
Lipidi Glucidi Tecnicamente solo le ultime tre Proteine ...tecnlab/Materiale Didattico/2015/Elementi...
55
Le macromolecole organiche •Lipidi •Glucidi •Proteine •Acidi nucleici Tecnicamente solo le ultime tre categorie sono costituite da macromolecole
Transcript of Lipidi Glucidi Tecnicamente solo le ultime tre Proteine ...tecnlab/Materiale Didattico/2015/Elementi...

Le macromolecole organiche
•Lipidi
•Glucidi
•Proteine
•Acidi nucleici
Tecnicamente
solo le ultime tre
categorie sono
costituite da
macromolecole

Le macromolecole, assieme, costituiscono la maggior
parte del contenuto secco degli organismi viventi

La caratteristica comune a tutte le macromolecole è di
essere costituite da strutture complesse (polimeri)
ottenute dall’assemblaggio di unità più piccole
(monomeri).
Così come con gli stessi mattoni si possono costruire
un numero enorme di case differenti, a partire dagli
stessi monomeri ciascun organismo può costruire le
proprie macromolecole
Di norma gli animali non ricavano le macromolecole
direttamente dagli esseri viventi di cui si cibano.
Durante la digestione, i polimeri vengono
‘scomposti’ in monomeri e assorbiti come tali.
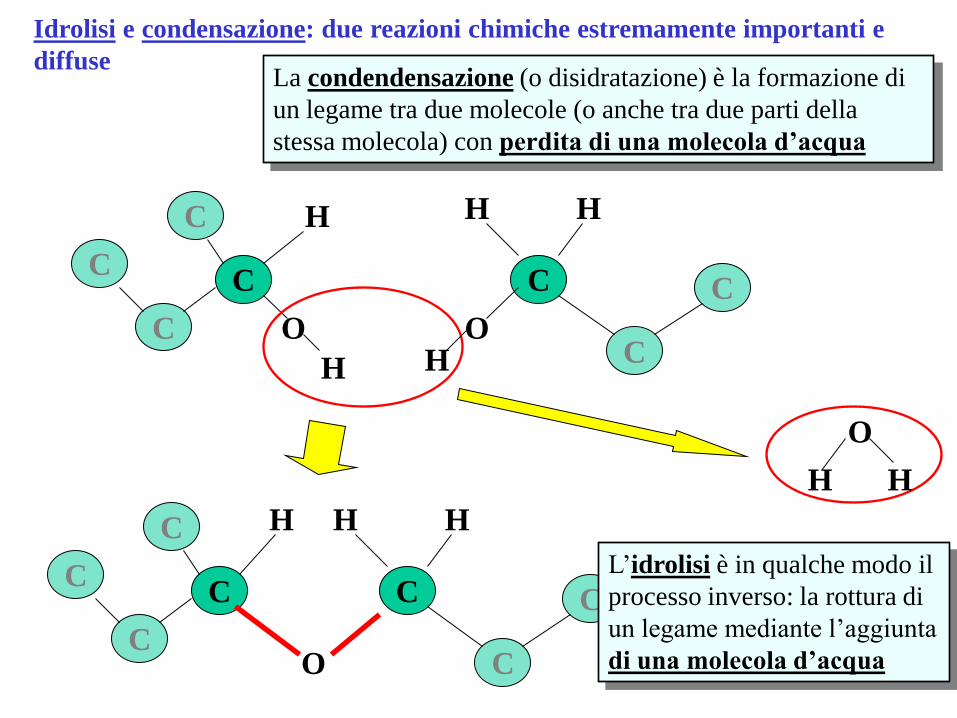
Idrolisi e condensazione: due reazioni chimiche estremamente importanti e
diffuse La condendensazione (o disidratazione) è la formazione di
un legame tra due molecole (o anche tra due parti della
stessa molecola) con perdita di una molecola d’acqua
C
C
C
O
H
H C
C
C C
H
O H
H
C
C
C
H C
C
C C
H
O
H
O
H H
L’idrolisi è in qualche modo il
processo inverso: la rottura di
un legame mediante l’aggiunta
di una molecola d’acqua

I lipidi sono un gruppo eterogeneo di sostanze. Essi sono
caratterizzati dalla loro insolubilità in acqua e dalla
affinità per i solventi apolari e per gli altri lipidi
• Trigliceridi
• Steroidi
• Fosfolipidi
• Carotenoidi
Funzione di deposito
Varie funzioni tra cui
quella di messaggeri
chimici (ormoni)
Sono la base costituente
delle membrane
biologiche
Fotopigmenti,
antiossidanti
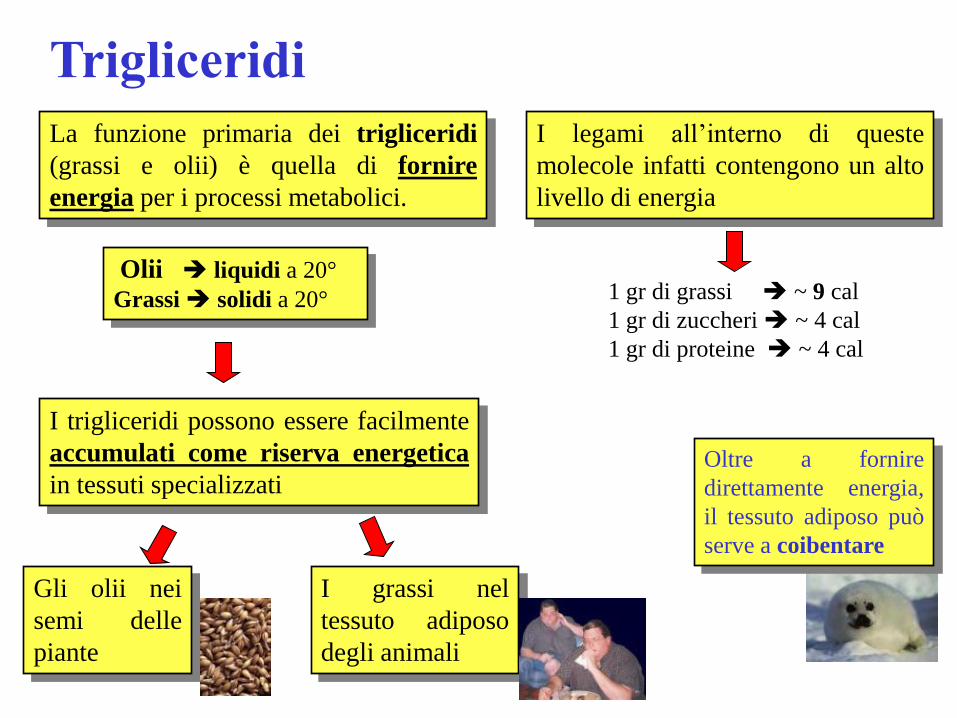
Trigliceridi
La funzione primaria dei trigliceridi
(grassi e olii) è quella di fornire
energia per i processi metabolici.
I legami all’interno di queste
molecole infatti contengono un alto
livello di energia
1 gr di grassi ~ 9 cal
1 gr di zuccheri ~ 4 cal
1 gr di proteine ~ 4 cal
Olii liquidi a 20°
Grassi solidi a 20°
I trigliceridi possono essere facilmente
accumulati come riserva energetica
in tessuti specializzati
Gli olii nei
semi delle
piante
I grassi nel
tessuto adiposo
degli animali
Oltre a fornire
direttamente energia,
il tessuto adiposo può
serve a coibentare
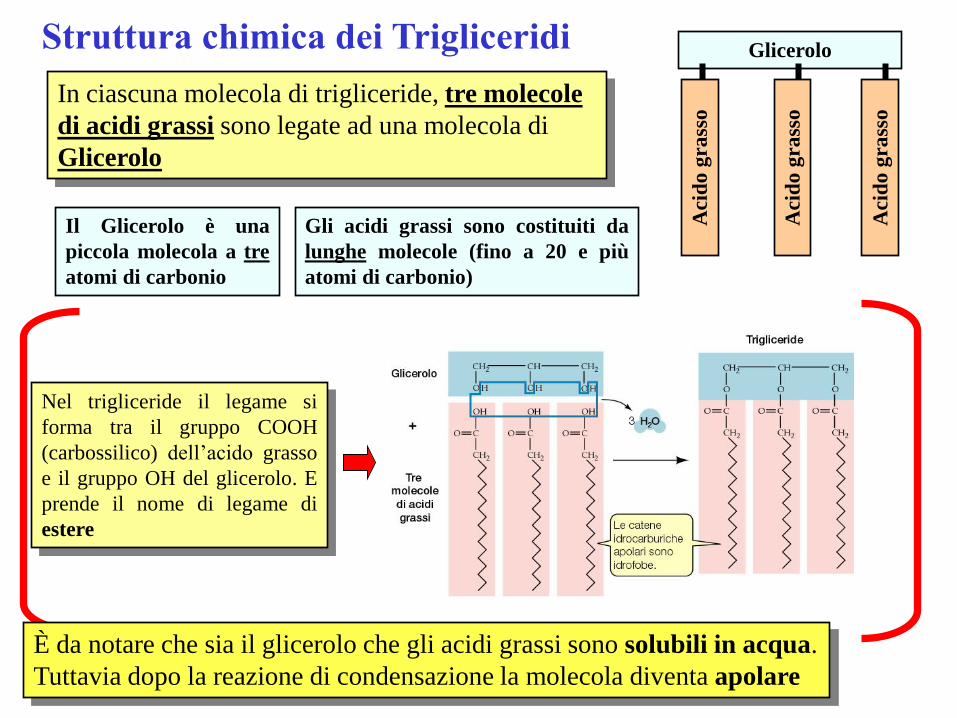
Struttura chimica dei Trigliceridi
In ciascuna molecola di trigliceride, tre molecole
di acidi grassi sono legate ad una molecola di
Glicerolo
Aci
do
gra
sso
Aci
do
gra
sso
Aci
do
gra
sso
Glicerolo
Il Glicerolo è una
piccola molecola a tre
atomi di carbonio
Gli acidi grassi sono costituiti da
lunghe molecole (fino a 20 e più
atomi di carbonio)
Nel trigliceride il legame si
forma tra il gruppo COOH
(carbossilico) dell’acido grasso
e il gruppo OH del glicerolo. E
prende il nome di legame di
estere
È da notare che sia il glicerolo che gli acidi grassi sono solubili in acqua.
Tuttavia dopo la reazione di condensazione la molecola diventa apolare
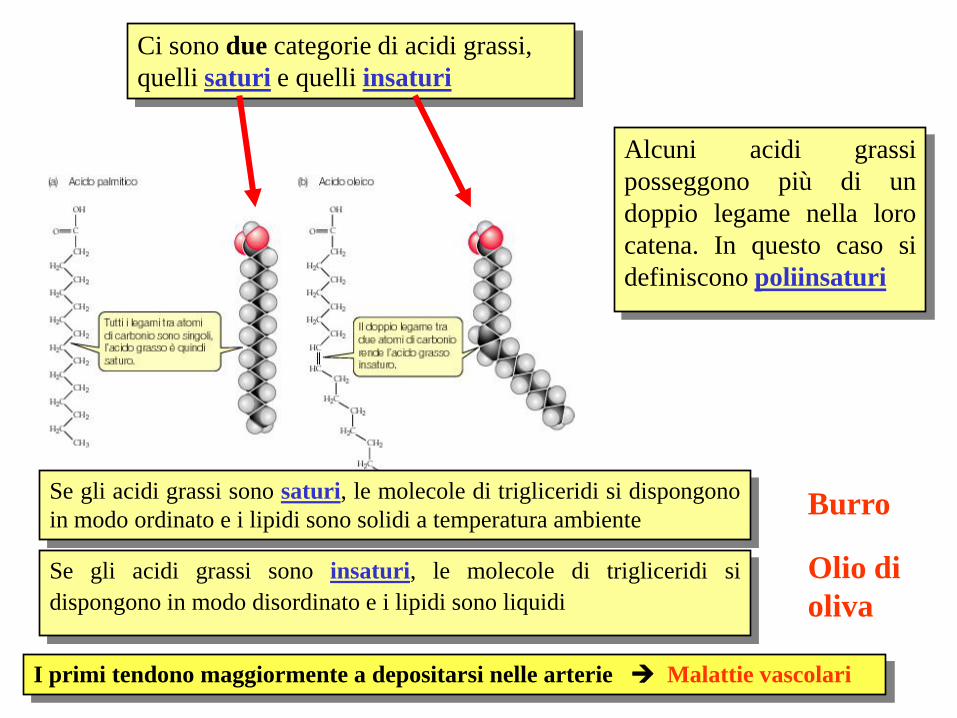
Ci sono due categorie di acidi grassi,
quelli saturi e quelli insaturi
Alcuni acidi grassi
posseggono più di un
doppio legame nella loro
catena. In questo caso si
definiscono poliinsaturi
Se gli acidi grassi sono saturi, le molecole di trigliceridi si dispongono
in modo ordinato e i lipidi sono solidi a temperatura ambiente
Se gli acidi grassi sono insaturi, le molecole di trigliceridi si
dispongono in modo disordinato e i lipidi sono liquidi
Burro
Olio di
oliva
I primi tendono maggiormente a depositarsi nelle arterie Malattie vascolari
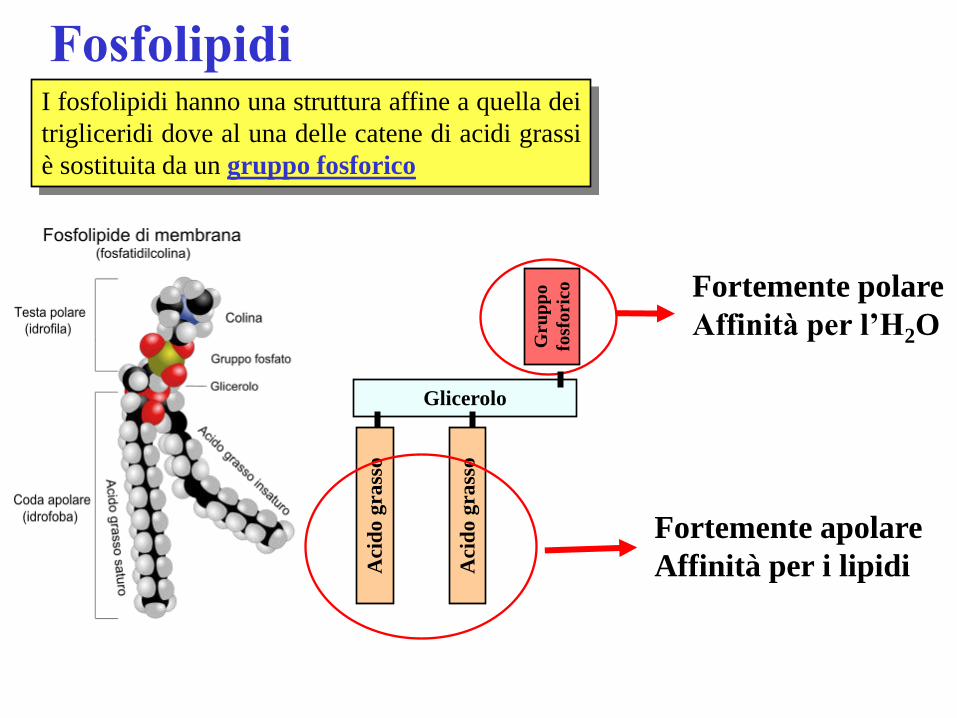
Fosfolipidi I fosfolipidi hanno una struttura affine a quella dei
trigliceridi dove al una delle catene di acidi grassi
è sostituita da un gruppo fosforico
Gru
pp
o
fosf
ori
co
Fortemente polare
Affinità per l’H2O
Aci
do
gra
sso
Aci
do
gra
sso
Glicerolo
Fortemente apolare
Affinità per i lipidi
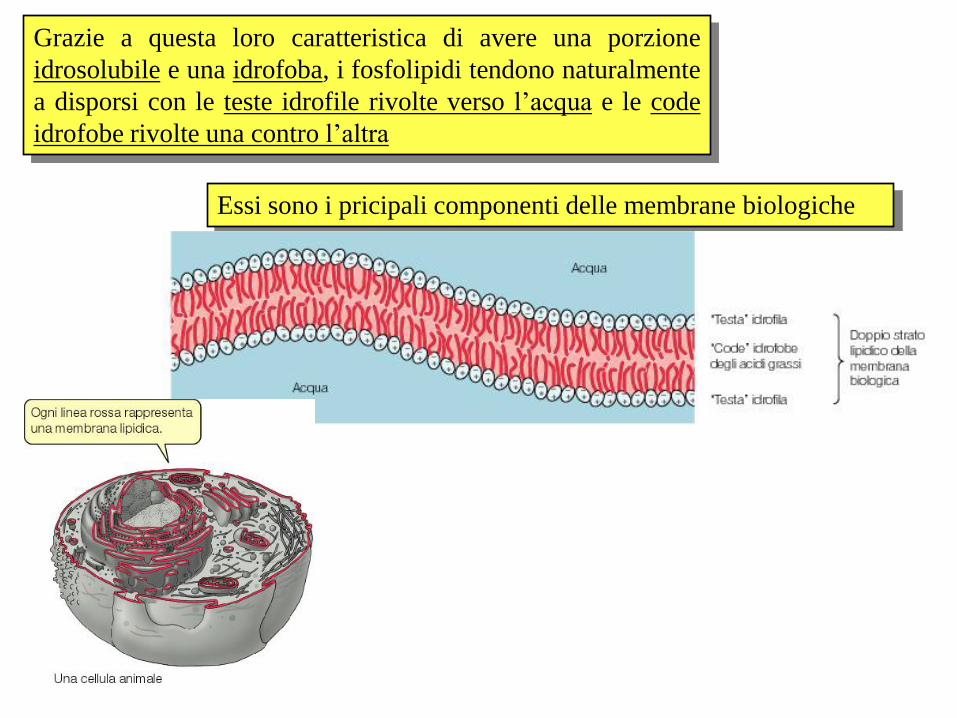
Grazie a questa loro caratteristica di avere una porzione
idrosolubile e una idrofoba, i fosfolipidi tendono naturalmente
a disporsi con le teste idrofile rivolte verso l’acqua e le code
idrofobe rivolte una contro l’altra
Essi sono i pricipali componenti delle membrane biologiche
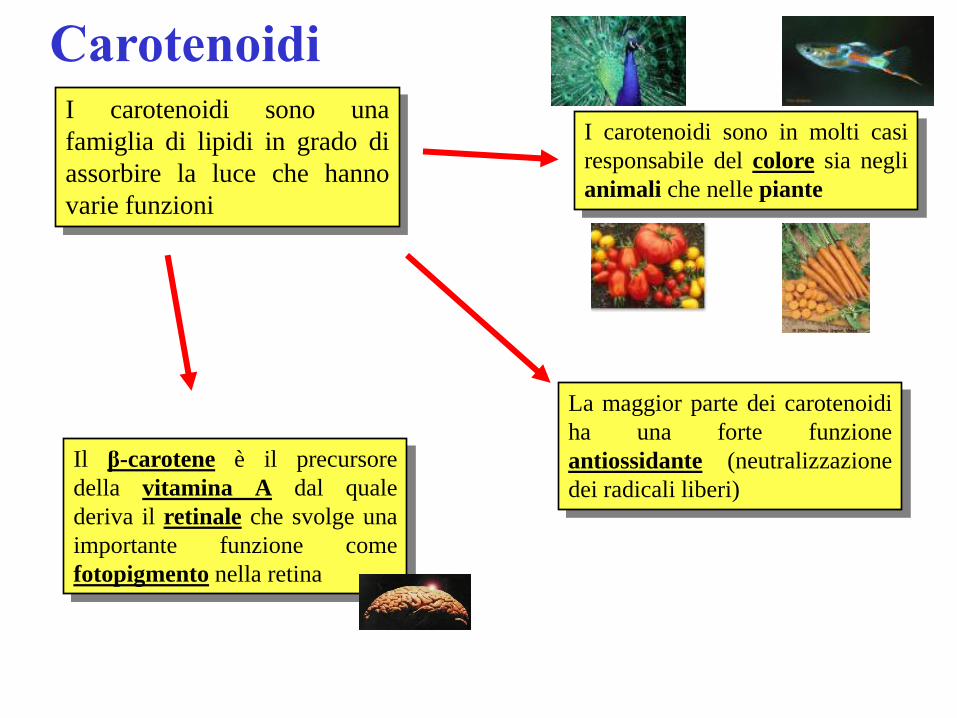
Carotenoidi I carotenoidi sono una
famiglia di lipidi in grado di
assorbire la luce che hanno
varie funzioni
Il β-carotene è il precursore
della vitamina A dal quale
deriva il retinale che svolge una
importante funzione come
fotopigmento nella retina
I carotenoidi sono in molti casi
responsabile del colore sia negli
animali che nelle piante
La maggior parte dei carotenoidi
ha una forte funzione
antiossidante (neutralizzazione
dei radicali liberi)

Approfondimento:
Radicali liberi, ossidazione e Carotenoidi I processi metabolici della cellula generano continuamente composti
dell’ossigeno (Radicali Liberi) caratterizzati dall’avere un numero spaiato di
elettroni e dalla forte tendenza a cedere O e ad ossidare le molecole
O2- anione superossido
H2O2 acqua ossigenata
OH- radicale idrossilico
I radicali liberi sono utilizzati per alcune
funzioni quali inattivare virus e batteri,
detossificare, attivare certi enzimi
Essi tuttavia hanno anche effetti dannosi sulla cellula:
- Reagiscono con il DNA provocando mutazioni (e tumori)
- Agiscono sugli acidi grassi dei fosfolipidi danneggiando le membrane
- Possono agire su certi enzimi di membrana (pompe ioniche) interferendo
con l’equilibrio idro-salino della cellula
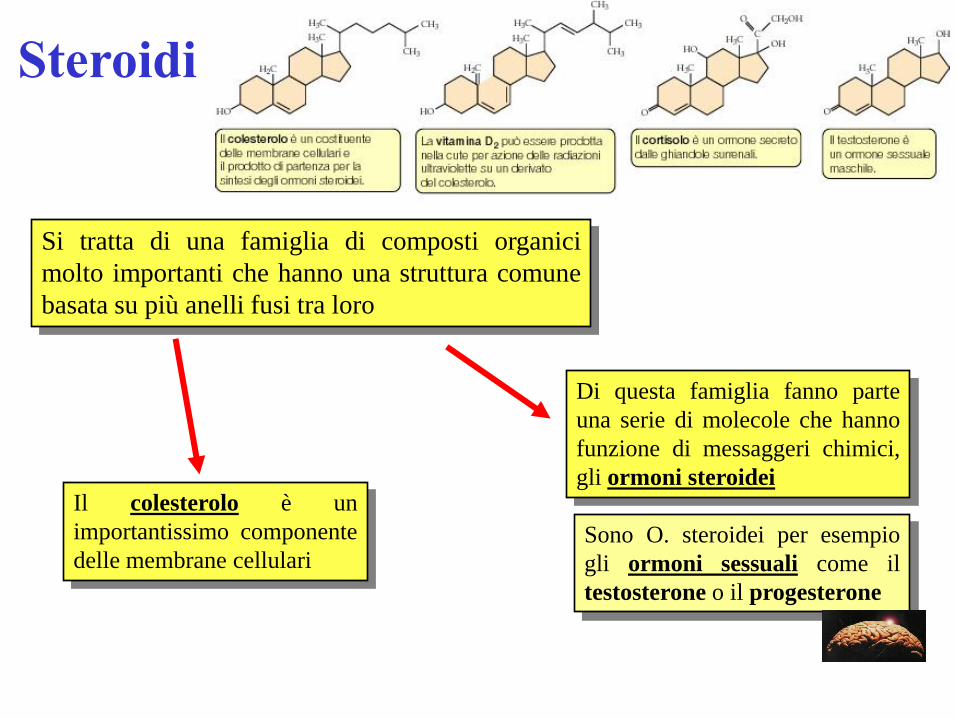
Steroidi
Si tratta di una famiglia di composti organici
molto importanti che hanno una struttura comune
basata su più anelli fusi tra loro
Il colesterolo è un
importantissimo componente
delle membrane cellulari
Di questa famiglia fanno parte
una serie di molecole che hanno
funzione di messaggeri chimici,
gli ormoni steroidei
Sono O. steroidei per esempio
gli ormoni sessuali come il
testosterone o il progesterone

I glucidi o carboidrati sono un gruppo eterogeneo di sostanze. Essi
hanno in comune la formula minima CH2O. Il nome carboidrati
deriva dal fatto che sono composti oltre che da C, da H e O nelle
proporzioni in cui si trovano nell’acqua
• Monosaccaridi
• Disaccaridi
• Polisaccaridi
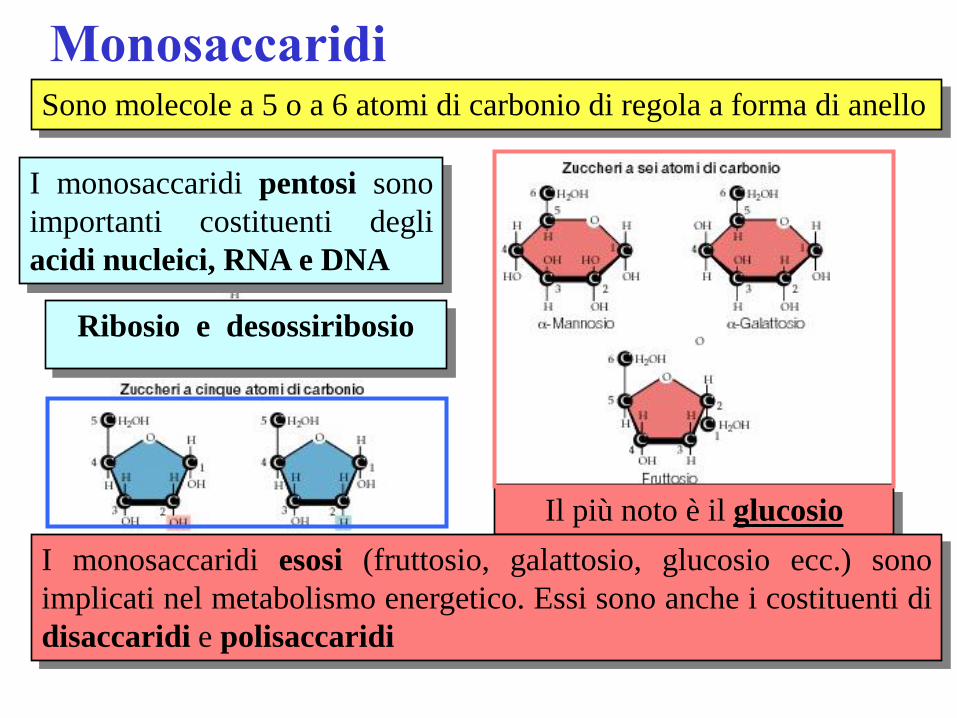
Monosaccaridi Sono molecole a 5 o a 6 atomi di carbonio di regola a forma di anello
I monosaccaridi pentosi sono
importanti costituenti degli
acidi nucleici, RNA e DNA
Ribosio e desossiribosio
Il più noto è il glucosio
I monosaccaridi esosi (fruttosio, galattosio, glucosio ecc.) sono
implicati nel metabolismo energetico. Essi sono anche i costituenti di
disaccaridi e polisaccaridi
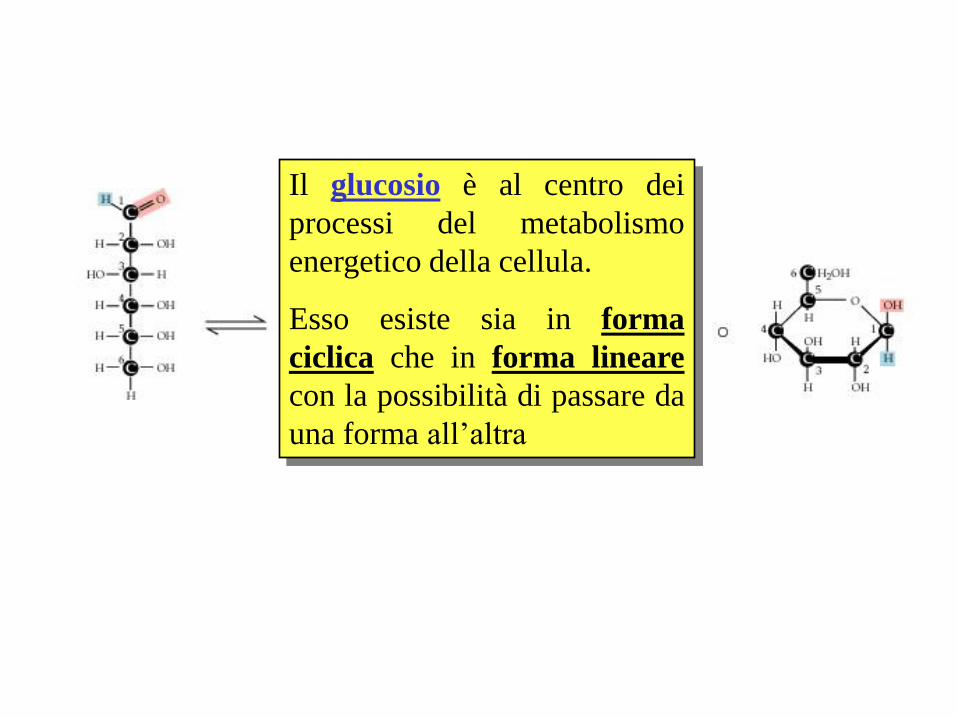
Il glucosio è al centro dei
processi del metabolismo
energetico della cellula.
Esso esiste sia in forma
ciclica che in forma lineare
con la possibilità di passare da
una forma all’altra
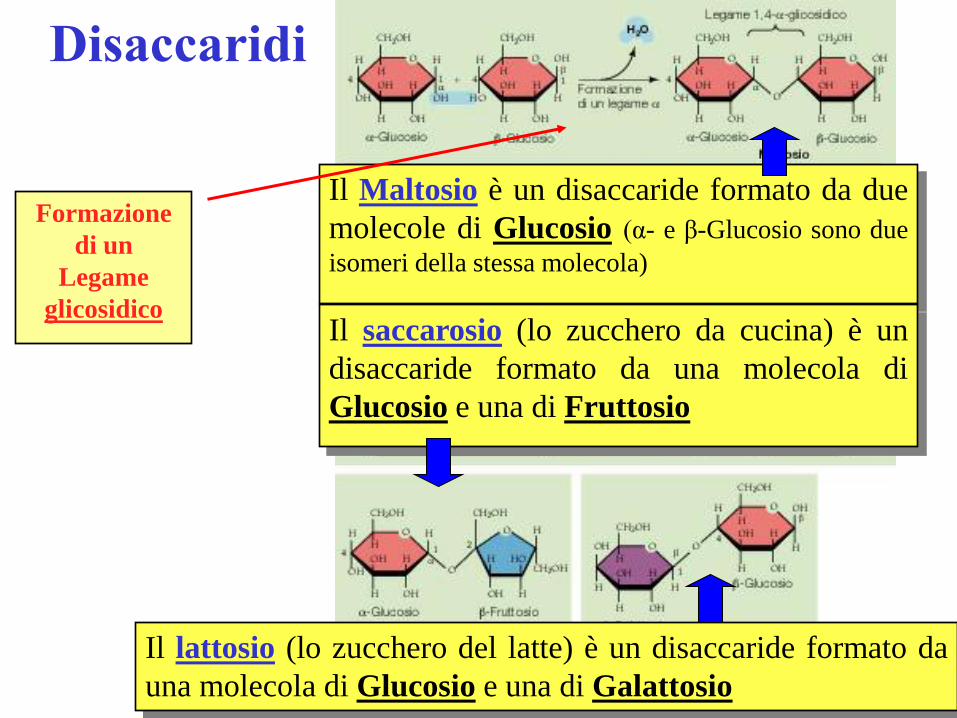
Disaccaridi
Il Maltosio è un disaccaride formato da due
molecole di Glucosio (α- e β-Glucosio sono due
isomeri della stessa molecola)
Il saccarosio (lo zucchero da cucina) è un
disaccaride formato da una molecola di
Glucosio e una di Fruttosio
Il lattosio (lo zucchero del latte) è un disaccaride formato da
una molecola di Glucosio e una di Galattosio
Formazione
di un
Legame
glicosidico
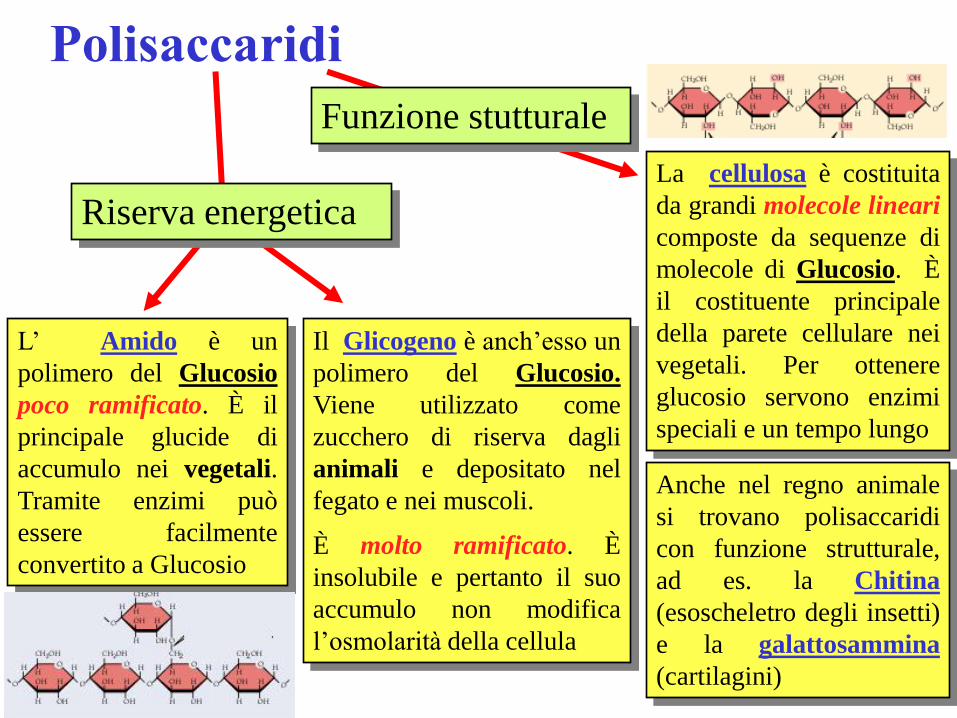
Polisaccaridi
Funzione stutturale
La cellulosa è costituita
da grandi molecole lineari
composte da sequenze di
molecole di Glucosio. È
il costituente principale
della parete cellulare nei
vegetali. Per ottenere
glucosio servono enzimi
speciali e un tempo lungo
Anche nel regno animale
si trovano polisaccaridi
con funzione strutturale,
ad es. la Chitina
(esoscheletro degli insetti)
e la galattosammina
(cartilagini)
Riserva energetica
L’ Amido è un
polimero del Glucosio
poco ramificato. È il
principale glucide di
accumulo nei vegetali.
Tramite enzimi può
essere facilmente
convertito a Glucosio
Il Glicogeno è anch’esso un
polimero del Glucosio.
Viene utilizzato come
zucchero di riserva dagli
animali e depositato nel
fegato e nei muscoli.
È molto ramificato. È
insolubile e pertanto il suo
accumulo non modifica
l’osmolarità della cellula

Proteine

Esistono decine di migliaia di diverse proteine. In ciascun essere vivente esse
svolgono funzioni fondamentali che nessun altro tipo di molecola potrebbe
svolgere
Nella cellula catalizzano migliaia di
differenti reazioni chimiche (Enzimi) Permettono il passaggio selettivo di
molecole attraverso la membrana
(Canali Ionici e Pompe) Trasportano specifiche molecole sia
nelle cellule che nei fluidi esterni
Attaccano selettivamente gli agenti
esterni (anticorpi del S.I.)
Fungono da messaggeri chimici
(Ormoni e neurotrasmettitori)
Hanno funzione strutturale sia
all’interno della cellula
(citoscheletro) che al di fuori (es
cheratine)
Regolano lo sviluppo e orchestrano il
funzionamento dell’intera cellula
Presiedono al movimento sia all’interno della cellula (cilia, trasporto
assoplasmatico) che a livello di sistema (es lavoro muscolare)
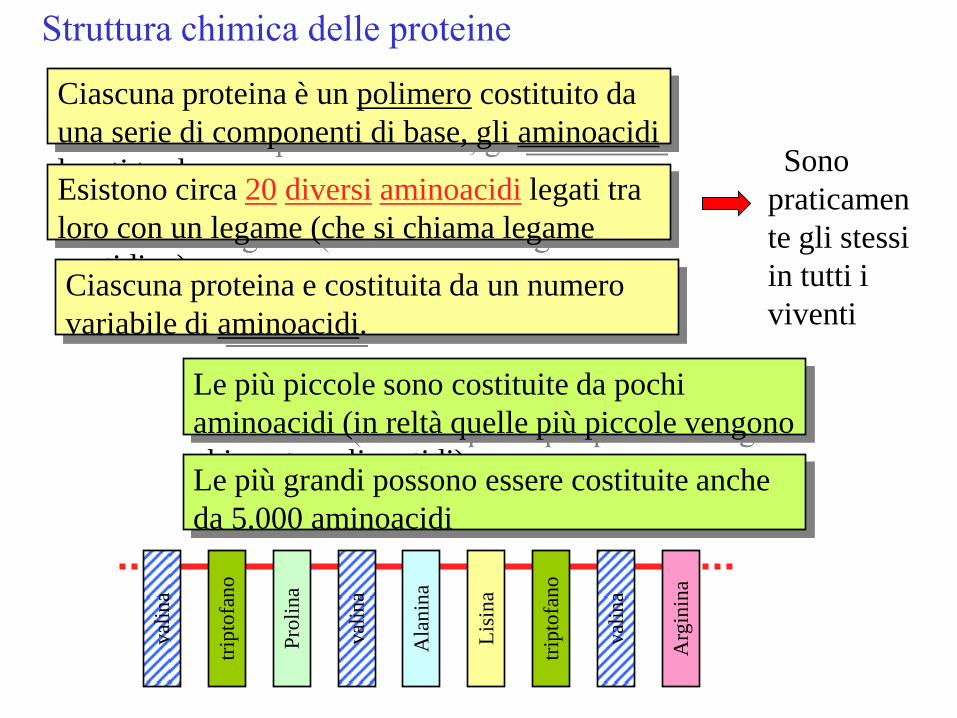
Struttura chimica delle proteine
Ciascuna proteina è un polimero costituito da
una serie di componenti di base, gli aminoacidi
legati tra loro Esistono circa 20 diversi aminoacidi legati tra
loro con un legame (che si chiama legame
peptidico) Ciascuna proteina e costituita da un numero
variabile di aminoacidi.
Le più piccole sono costituite da pochi
aminoacidi (in reltà quelle più piccole vengono
chiamate polipeptidi) Le più grandi possono essere costituite anche
da 5.000 aminoacidi
val
ina
Ala
nin
a
Pro
lin
a
Arg
inin
a
val
ina
Lis
ina
val
ina
trip
tofa
no
trip
tofa
no
Sono
praticamen
te gli stessi
in tutti i
viventi
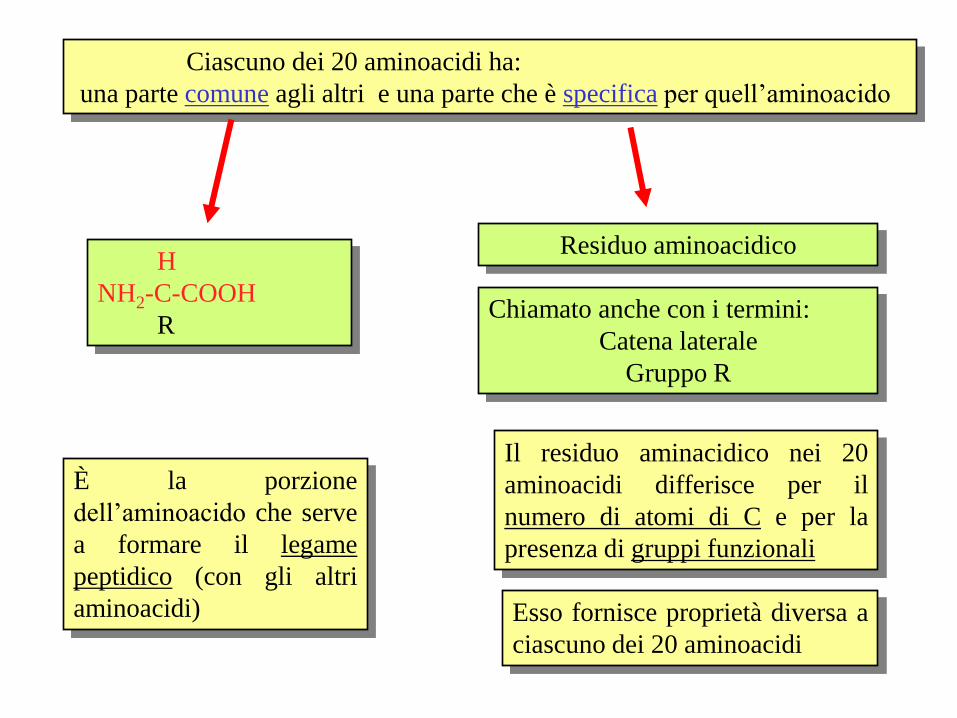
Ciascuno dei 20 aminoacidi ha:
una parte comune agli altri e una parte che è specifica per quell’aminoacido
H
NH2-C-COOH
R
È la porzione
dell’aminoacido che serve
a formare il legame
peptidico (con gli altri
aminoacidi)
Residuo aminoacidico
Chiamato anche con i termini:
Catena laterale
Gruppo R
Il residuo aminacidico nei 20
aminoacidi differisce per il
numero di atomi di C e per la
presenza di gruppi funzionali
Esso fornisce proprietà diversa a
ciascuno dei 20 aminoacidi


I venti aminoacidi differiscono tra loro in quanto:
• I residui sono carichi oppure elettricamente neutri
• I primi possono avere carica positiva o negativa
• Alcuni hanno residui idrofobici mentre altri hanno residui polari
• I residui hanno ingombri molto diversi
• Alcuni residui hanno gruppi funzionali peculiari
Questa differenza nelle caratteristiche dei Residui Aminoacidici
determina sia la struttura tridimensionale della proteina che le sue
funzionalità

Amino Acidi Idrofilici

Amino acidi idrofobici e amino acidi
“speciali”
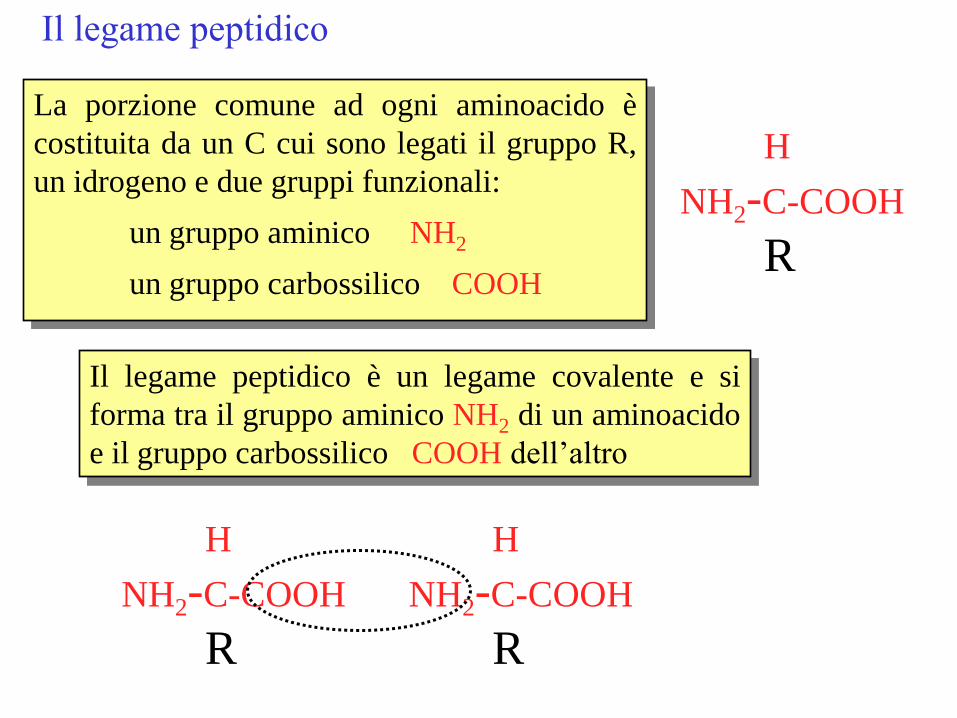
Il legame peptidico
H
NH2-C-COOH
R
La porzione comune ad ogni aminoacido è
costituita da un C cui sono legati il gruppo R,
un idrogeno e due gruppi funzionali:
un gruppo aminico NH2
un gruppo carbossilico COOH
Il legame peptidico è un legame covalente e si
forma tra il gruppo aminico NH2 di un aminoacido
e il gruppo carbossilico COOH dell’altro
H
NH2-C-COOH
R
H
NH2-C-COOH
R
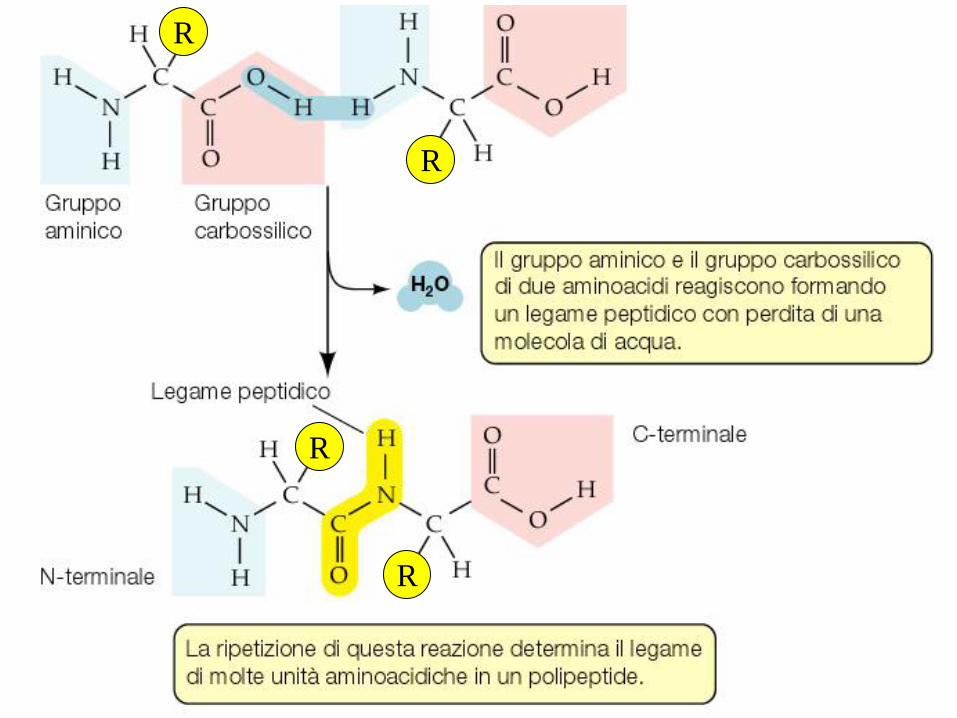
R
R
R
R
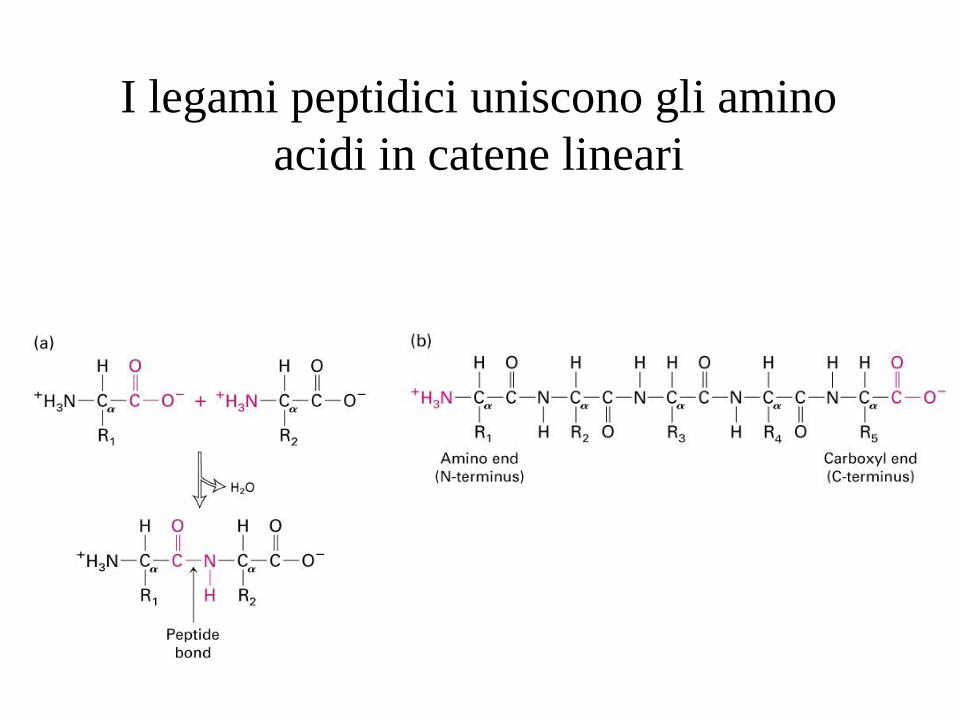
I legami peptidici uniscono gli amino
acidi in catene lineari

• Primaria: la sequenza lineare degli amino acidi
• Secondaria: l’organizzazione di parti di una catena polipeptidica
(esempio: l’ elica o il foglietto )
• Terziaria: la struttura tridimensionale completa di una catena
polipeptidica
• Quaternaria: l’associazione di due o più polipeptidi in una struttura
complessa multi-subunità
Quattro livelli di struttura determinano la
forma di una proteina
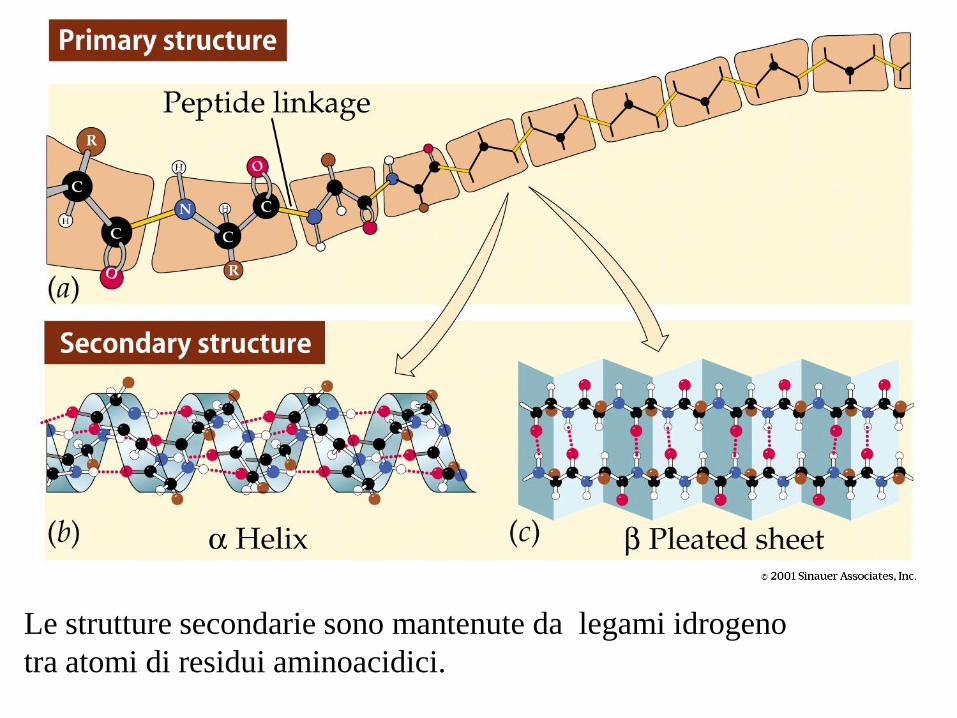
Le strutture secondarie sono mantenute da legami idrogeno
tra atomi di residui aminoacidici.
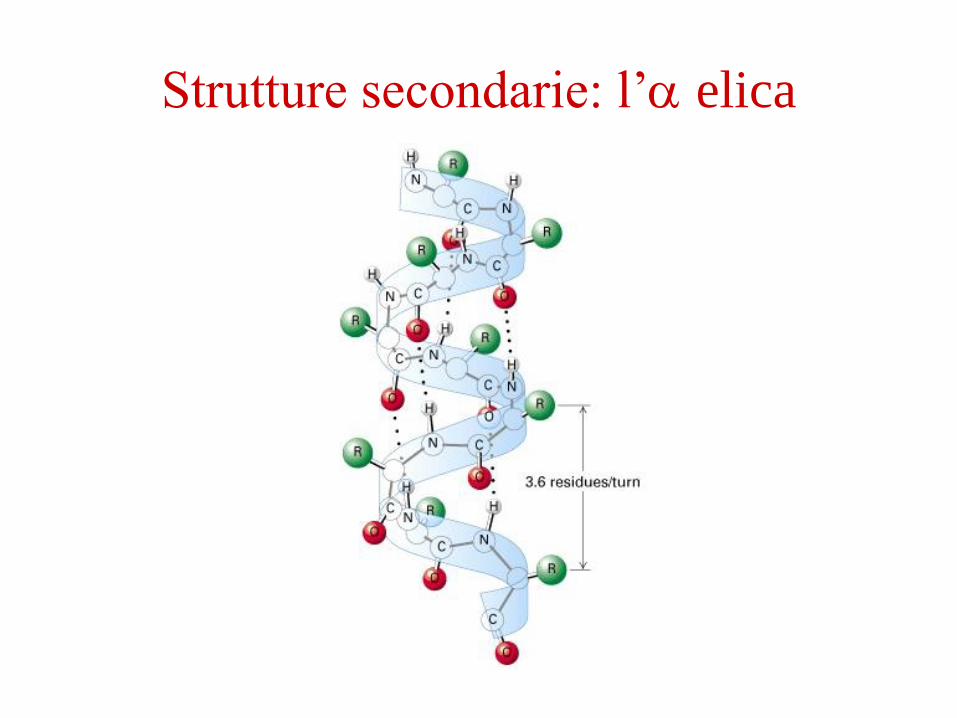
Strutture secondarie: l’ elica
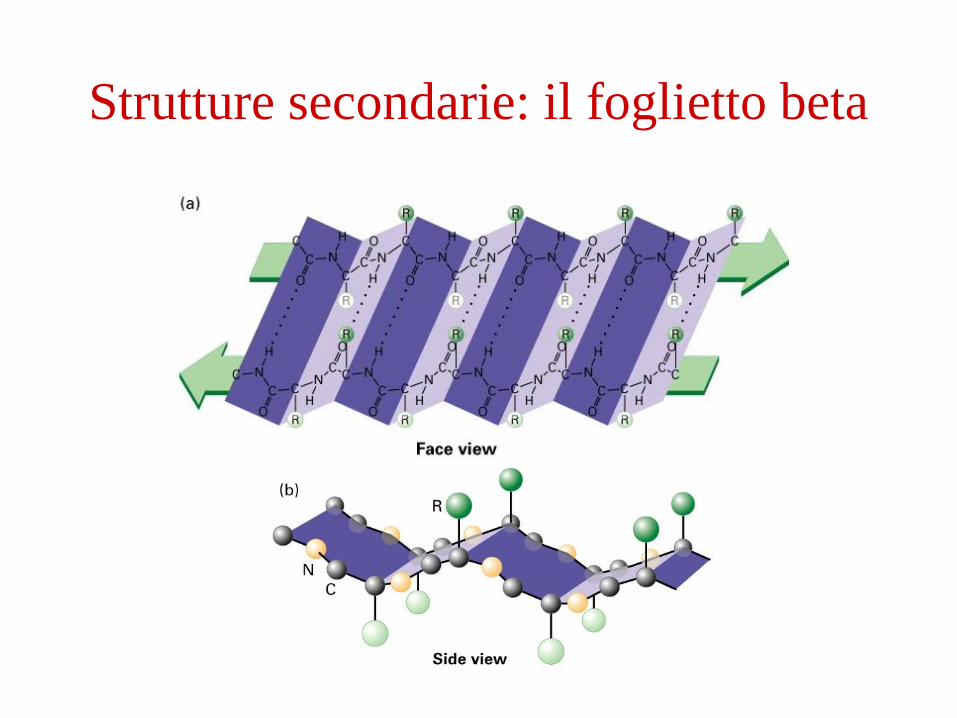
Strutture secondarie: il foglietto beta
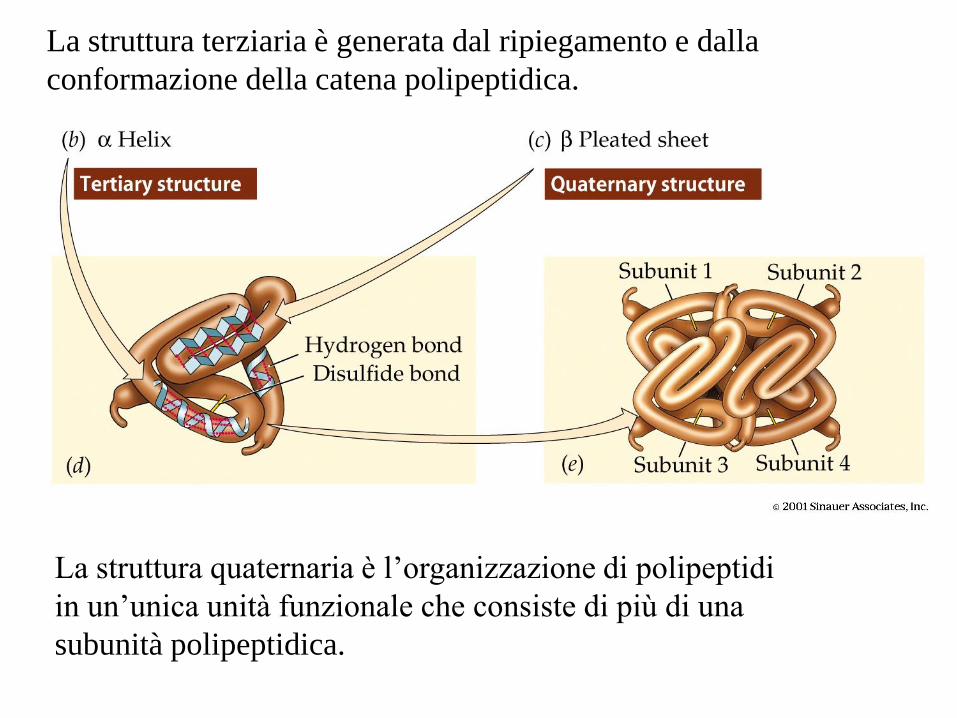
La struttura terziaria è generata dal ripiegamento e dalla
conformazione della catena polipeptidica.
La struttura quaternaria è l’organizzazione di polipeptidi
in un’unica unità funzionale che consiste di più di una
subunità polipeptidica.
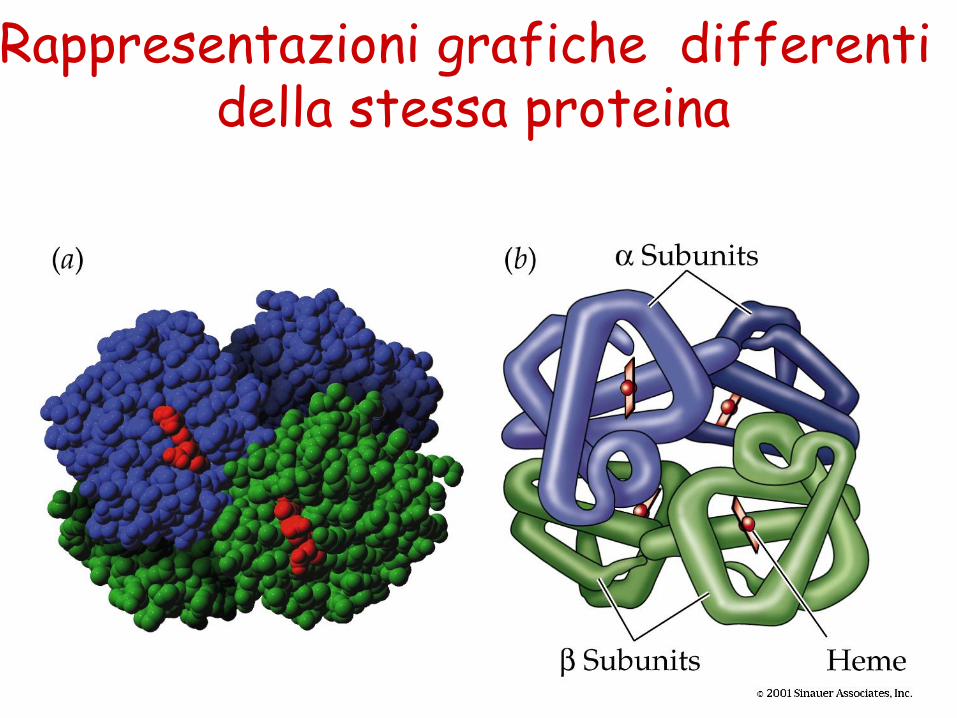
Rappresentazioni grafiche differenti della stessa proteina

Gli enzimi
In alcune reazioni chimiche il contenuto energetico dei prodotti della
reazione è complessivamente maggiore di quello dei reagenti.
Perché avvenga la reazione è necessario fornire energia
Queste reazioni si chiamano endoergoniche
In in altre reazioni chimiche il contenuto energetico dei prodotti della
reazione è minore di quello dei reagenti.
Quando la reazione chimica avviene si ha liberazione di energia
Queste reazioni si chiamano esoergoniche
Non è detto che perché una reazione è esoergonica essa
debba obbligatoriamente avvenire
Ad esempio la reazione che ha luogo durante la combustione di un
fiammifero è fortemente esoergonica ma perché avvenga occorre
inizialmente aggiungere energia per superare una barriera di attivazione
(sfregando la capocchia si produce calore)
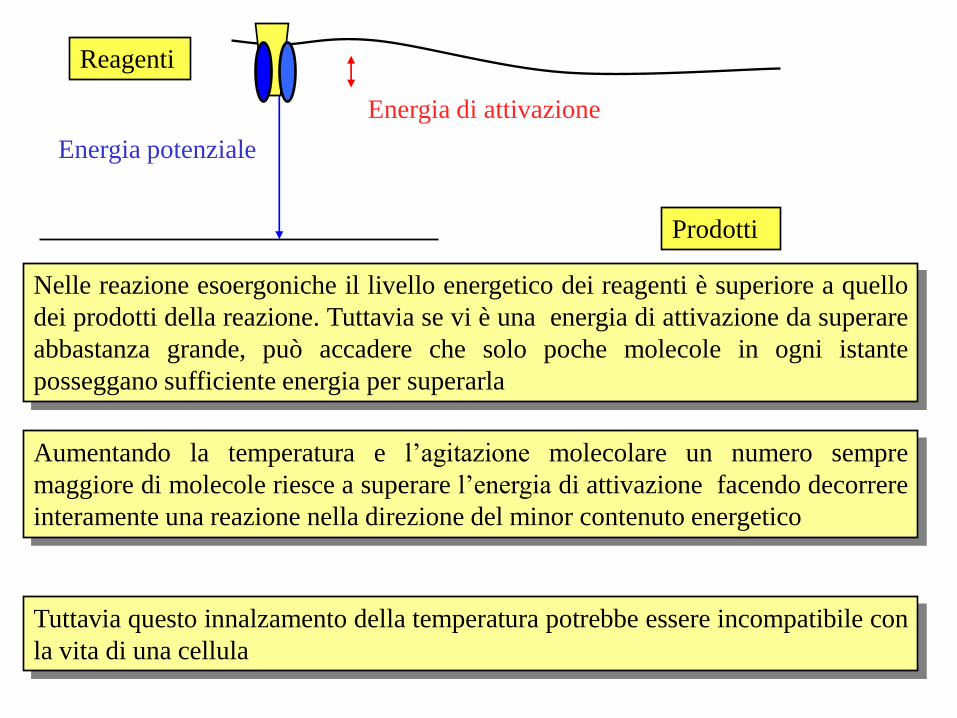
Energia potenziale
Energia di attivazione
Nelle reazione esoergoniche il livello energetico dei reagenti è superiore a quello
dei prodotti della reazione. Tuttavia se vi è una energia di attivazione da superare
abbastanza grande, può accadere che solo poche molecole in ogni istante
posseggano sufficiente energia per superarla
Reagenti
Prodotti
Aumentando la temperatura e l’agitazione molecolare un numero sempre
maggiore di molecole riesce a superare l’energia di attivazione facendo decorrere
interamente una reazione nella direzione del minor contenuto energetico
Tuttavia questo innalzamento della temperatura potrebbe essere incompatibile con
la vita di una cellula
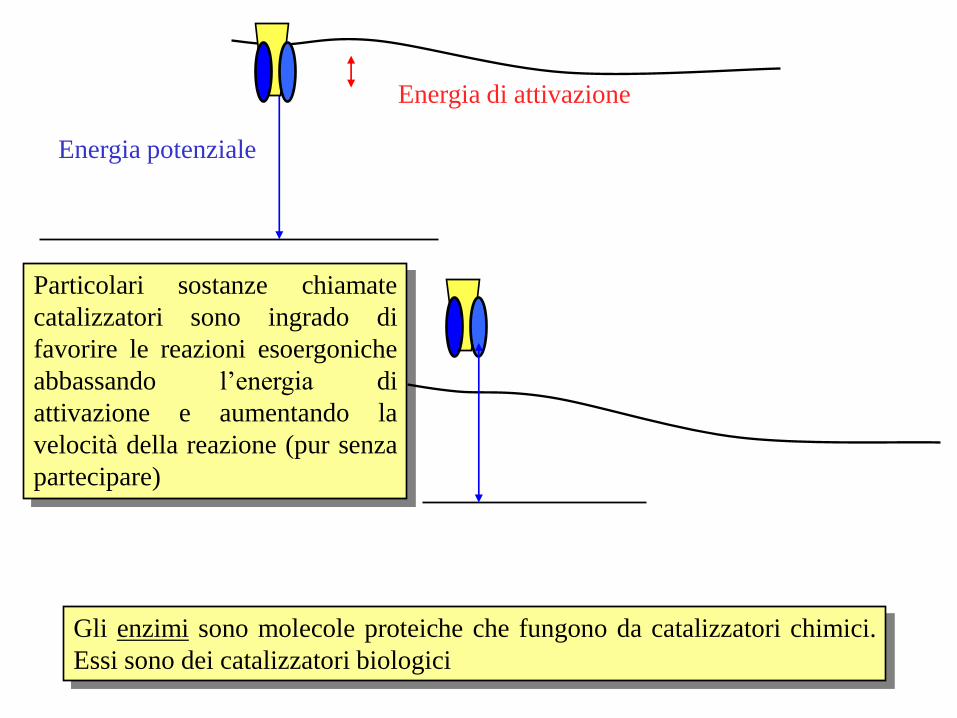
Gli enzimi sono molecole proteiche che fungono da catalizzatori chimici.
Essi sono dei catalizzatori biologici
Particolari sostanze chiamate
catalizzatori sono ingrado di
favorire le reazioni esoergoniche
abbassando l’energia di
attivazione e aumentando la
velocità della reazione (pur senza
partecipare)
Energia potenziale
Energia di attivazione

Gli enzimi aumentano la velocità delle reazioni chimiche abbassando l’energia
di attivazione
Grazie agli enzimi possono avvenire, a temperatura compatibili con la vita,
reazioni che altrimenti avverrebbero solo ad alte temperature
Grazie agli enzimi possono avvenire negli esseri viventi reazioni che altrimenti
avverrebbero con grande emissione di energia
Esistono migliaia di diversi tipi di enzimi ognuno specifico per una data reazione
chimica
Il nome della maggior parte degli enzimi è caratterizzato
dalla presenza del suffisso –asi (es. DNA-polimerasi)
Grazie agli enzimi possono avvenire anche reazioni endoergoniche mediante
l’accoppiamento di queste con reazioni esoergoniche
L’enzima agisce essenzialmente abbassando
l’energia di attivazione della reazione
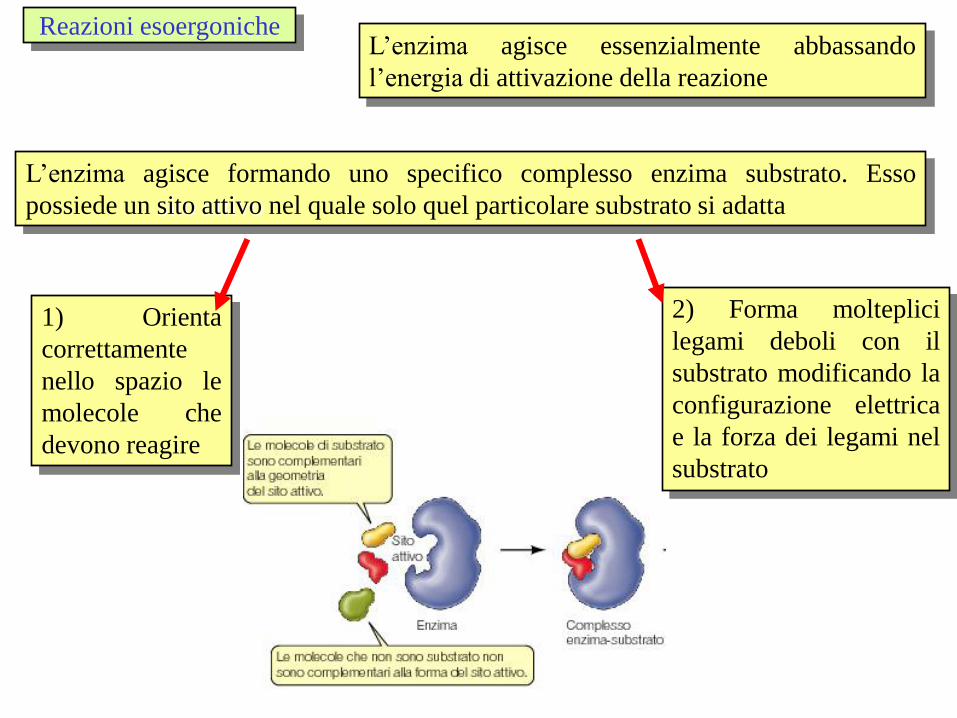
L’enzima agisce formando uno specifico complesso enzima substrato. Esso
possiede un sito attivo nel quale solo quel particolare substrato si adatta
1) Orienta
correttamente
nello spazio le
molecole che
devono reagire
2) Forma molteplici
legami deboli con il
substrato modificando la
configurazione elettrica
e la forza dei legami nel
substrato
Reazioni esoergoniche
Reazioni endoergoniche
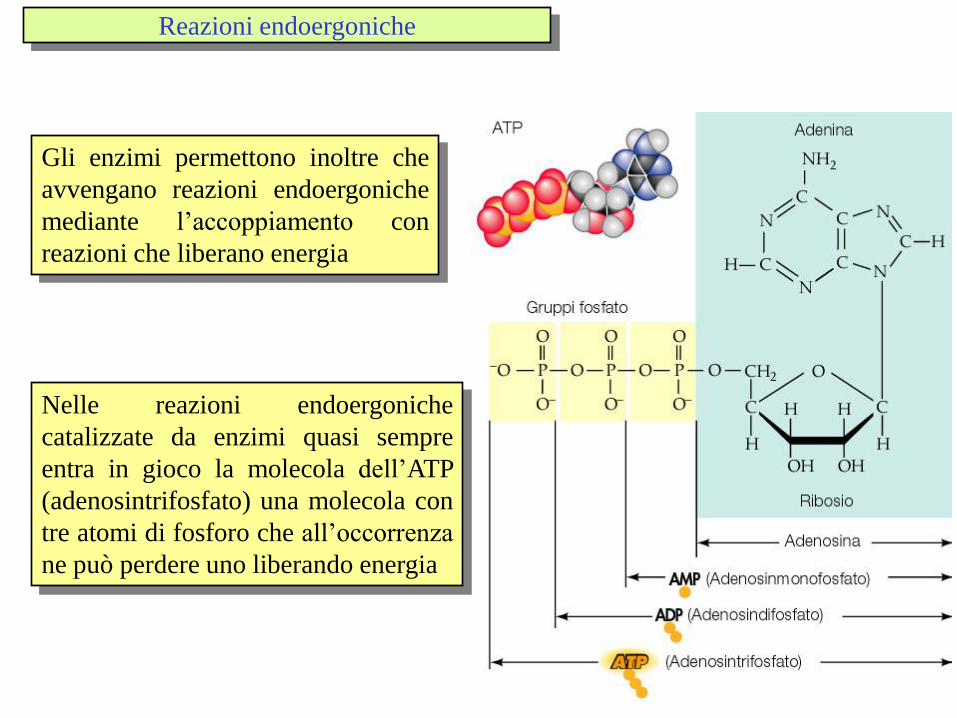
Nelle reazioni endoergoniche
catalizzate da enzimi quasi sempre
entra in gioco la molecola dell’ATP
(adenosintrifosfato) una molecola con
tre atomi di fosforo che all’occorrenza
ne può perdere uno liberando energia
Gli enzimi permettono inoltre che
avvengano reazioni endoergoniche
mediante l’accoppiamento con
reazioni che liberano energia
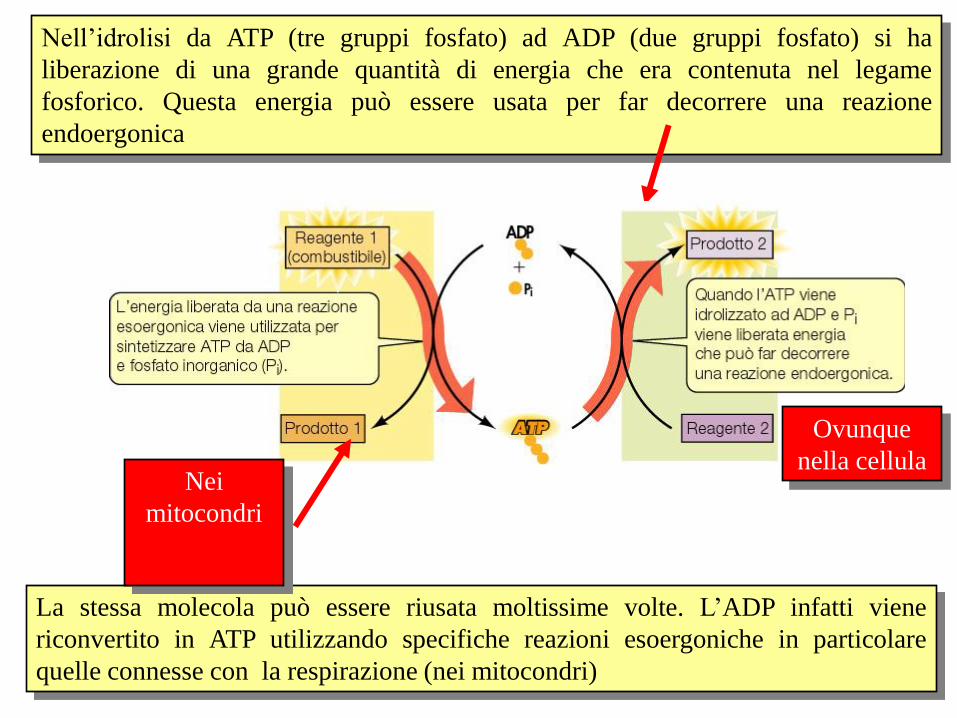
Nell’idrolisi da ATP (tre gruppi fosfato) ad ADP (due gruppi fosfato) si ha
liberazione di una grande quantità di energia che era contenuta nel legame
fosforico. Questa energia può essere usata per far decorrere una reazione
endoergonica
Ovunque
nella cellula
La stessa molecola può essere riusata moltissime volte. L’ADP infatti viene
riconvertito in ATP utilizzando specifiche reazioni esoergoniche in particolare
quelle connesse con la respirazione (nei mitocondri)
Nei
mitocondri

In altre parole l’enegia contenuta nei cibi che mangiamo viene utilizzata per
produrre ATP (tramite la respirazione nei mitocondri). L’ATP in seguito viene
utilizzato quale molecola di scambio per tutti i processi della cellula che richiedono
energia.
• Metabolismo -Reazioni chimiche endoergoniche
• Movimento (es muscolo, movimento ciliare)
• Pompe di membrana (per mantenere la concentrazione di soluti)
• Mantenimento della temperatura corporea
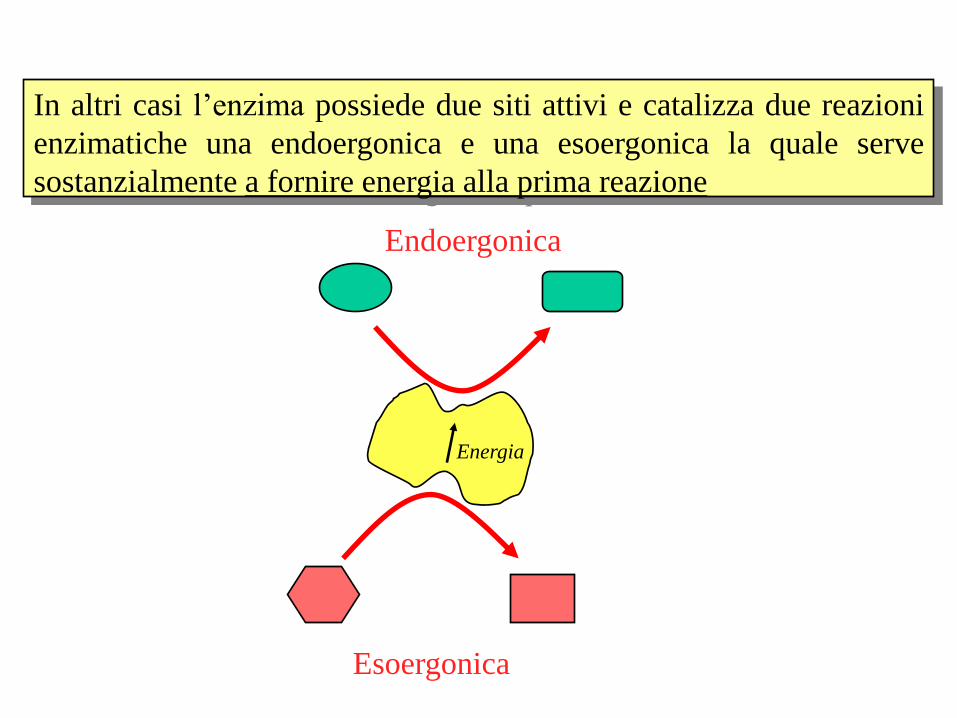
In altri casi l’enzima possiede due siti attivi e catalizza due reazioni
enzimatiche una endoergonica e una esoergonica la quale serve
sostanzialmente a fornire energia alla prima reazione
Endoergonica
Esoergonica
Energia
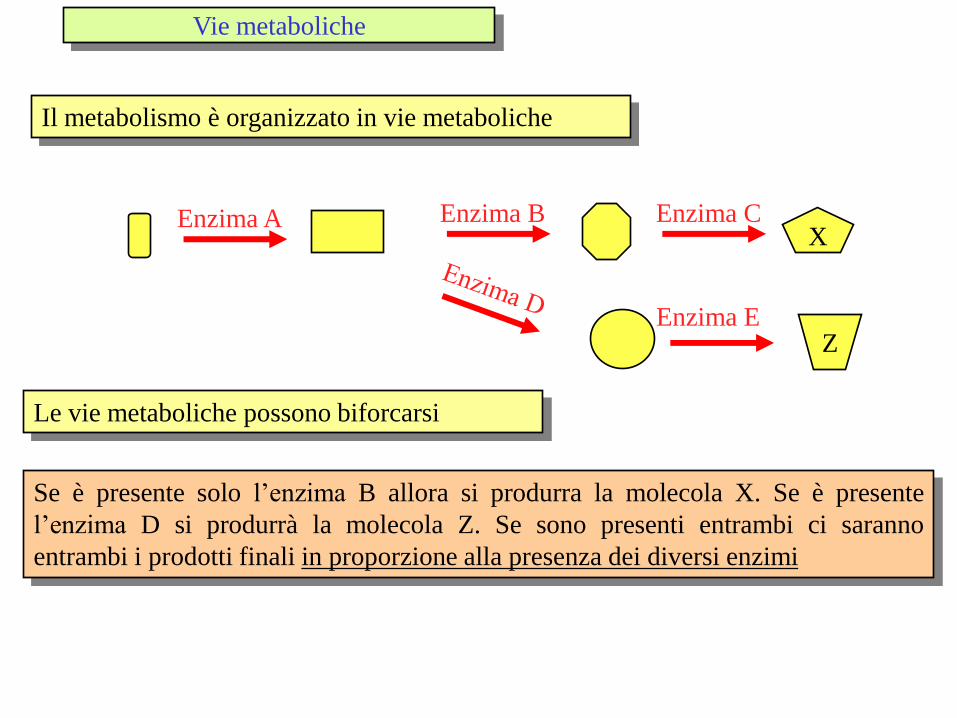
Il metabolismo è organizzato in vie metaboliche
X Enzima A Enzima B Enzima C
Z Enzima E
Le vie metaboliche possono biforcarsi
Se è presente solo l’enzima B allora si produrra la molecola X. Se è presente
l’enzima D si produrrà la molecola Z. Se sono presenti entrambi ci saranno
entrambi i prodotti finali in proporzione alla presenza dei diversi enzimi
Vie metaboliche
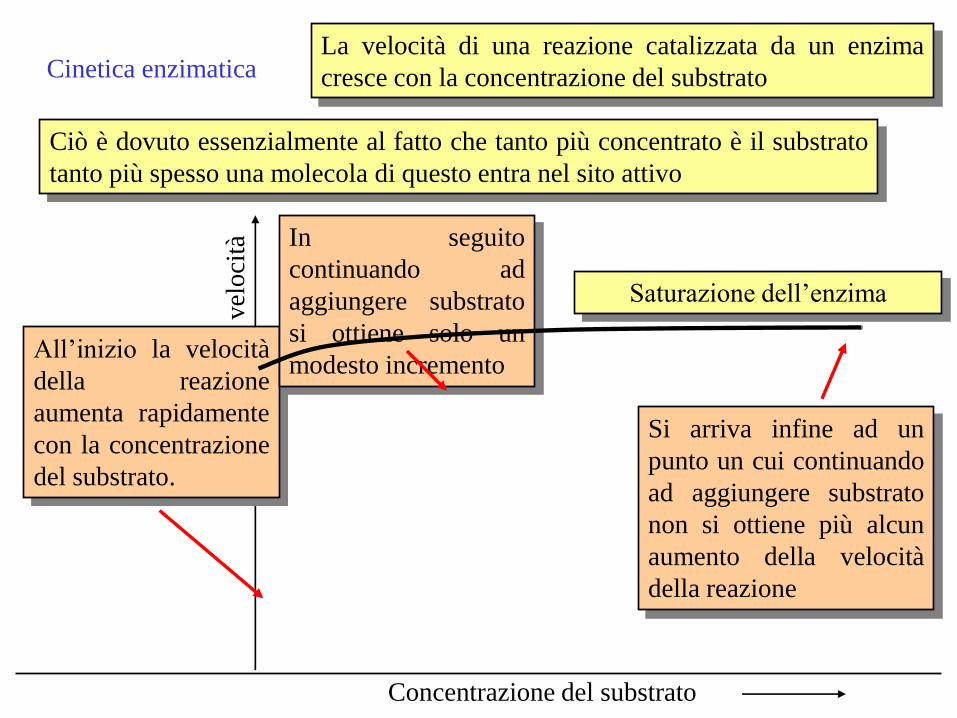
Cinetica enzimatica La velocità di una reazione catalizzata da un enzima
cresce con la concentrazione del substrato
Ciò è dovuto essenzialmente al fatto che tanto più concentrato è il substrato
tanto più spesso una molecola di questo entra nel sito attivo
All’inizio la velocità
della reazione
aumenta rapidamente
con la concentrazione
del substrato.
In seguito
continuando ad
aggiungere substrato
si ottiene solo un
modesto incremento
Si arriva infine ad un
punto un cui continuando
ad aggiungere substrato
non si ottiene più alcun
aumento della velocità
della reazione
Saturazione dell’enzima
Concentrazione del substrato
vel
oci
tà

Date queste caratteristiche della cinetica enzimatica, quando il substrato è
presente in eccesso saranno la concentrazione dell’enzima e la sua affinità
con il substrato (in molti enzimi questa può essere variata) a determinare il
decorrere di una data reazione

Regolazione della funzione enzimatica
L’azione degli enzimi è regolata da una gran quantità i fattori legati
ai bisogni della cellula e alla disponibilità dei composti.
Ci sono essenzialmente due modi per regolare l’attività di una determinata via
metabolica
1) Si può controllare la produzione di un dato enzima aumentandola
o diminuendola tramite un’azione sui meccanismi di regolazione del
gene
1) Si può agire direttamente sulla funzionalità di un dato enzima
agendo su uno dei suoi siti attivi
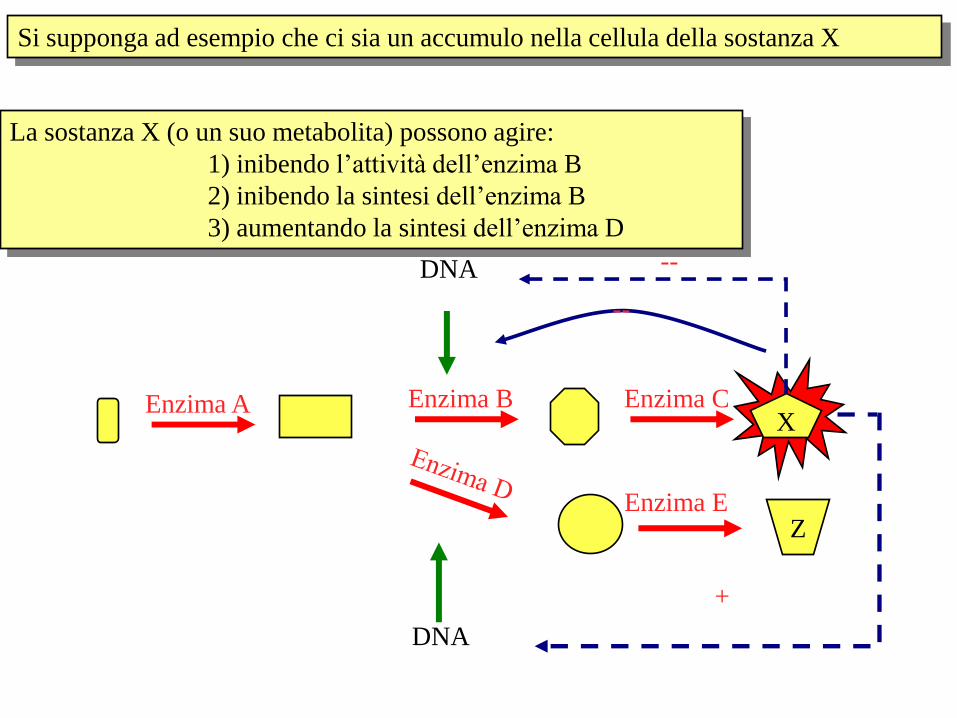
X Enzima A Enzima B Enzima C
Z Enzima E
Si supponga ad esempio che ci sia un accumulo nella cellula della sostanza X
La sostanza X (o un suo metabolita) possono agire:
1) inibendo l’attività dell’enzima B
2) inibendo la sintesi dell’enzima B
3) aumentando la sintesi dell’enzima D
DNA
+
--
DNA --
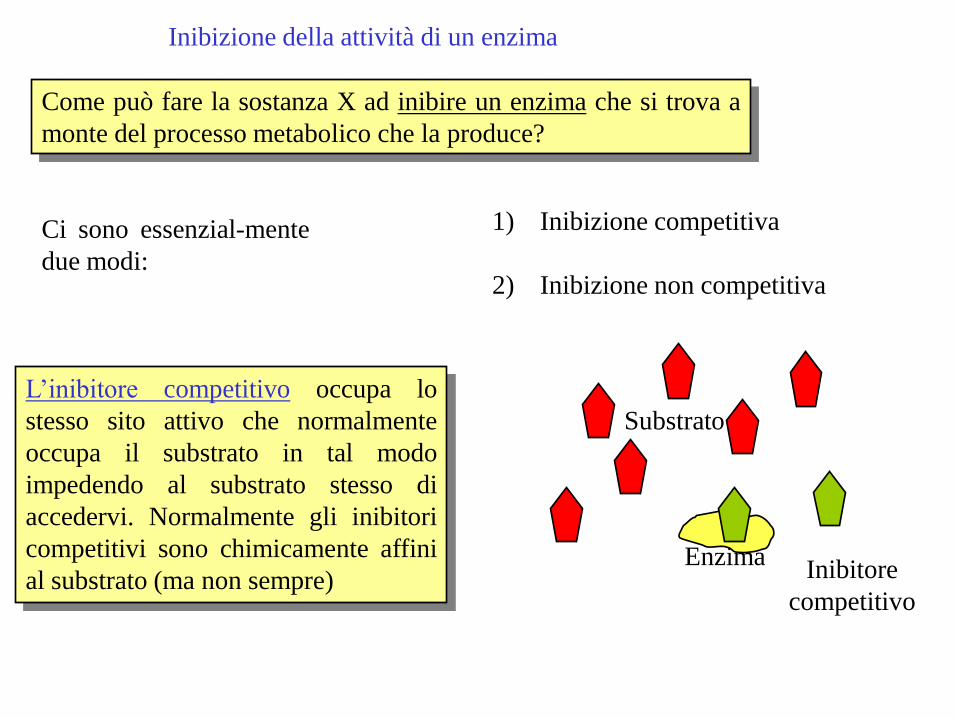
Inibizione della attività di un enzima
Come può fare la sostanza X ad inibire un enzima che si trova a
monte del processo metabolico che la produce?
Ci sono essenzial-mente
due modi:
1) Inibizione competitiva
2) Inibizione non competitiva
L’inibitore competitivo occupa lo
stesso sito attivo che normalmente
occupa il substrato in tal modo
impedendo al substrato stesso di
accedervi. Normalmente gli inibitori
competitivi sono chimicamente affini
al substrato (ma non sempre) Enzima
Inibitore
competitivo
Substrato
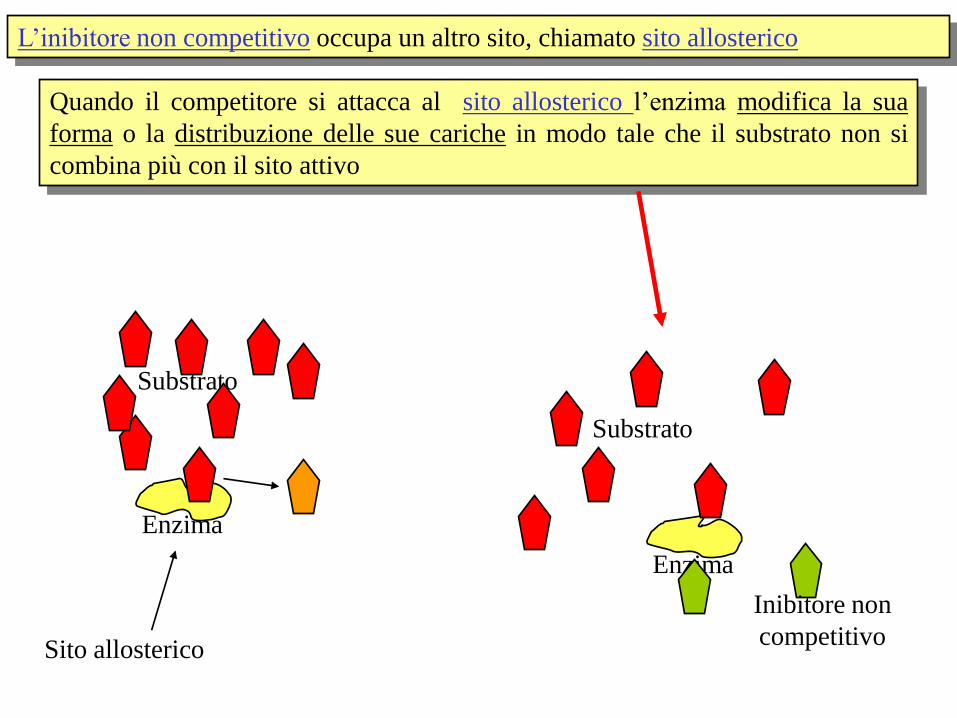
L’inibitore non competitivo occupa un altro sito, chiamato sito allosterico
Enzima
Inibitore non
competitivo
Substrato
Enzima
Substrato
Sito allosterico
Quando il competitore si attacca al sito allosterico l’enzima modifica la sua
forma o la distribuzione delle sue cariche in modo tale che il substrato non si
combina più con il sito attivo

Molte tossine e veleni sono inibitori enzimatici (sia
competitivi che non)
Gli inibitori naturali (il cui scopo è regolare il metabolismo) si legano
reversibilmente ai siti dell’enzima. Al contrario veleni e tossine
spesso si legano irreversibilmente bloccando ogni ulteriore attività
enzimatica

Isozimi
Gli isozimi o isoenzimi sono forme leggermente diverse dello stesso
enzima (possono avere uno o più aminoacidi di differenza)
Uno dei vantaggi degli eterozigoti * è la presenza di isozimi
Si supponga ad esempio che l’enzima A1 funzioni in modo ottimale a 20 gradi
di temperatura mentre la forma A2 lavori meglio a 30 gradi
Gli individui che possiedono
A1 saranno favoriti a 20 gradi
mentre quelli che hanno A2
saranno favoriti a 30 gradi
Gli individui hanno una
mistura di A1 e A2
potranno vivere in un
ambito di temperature più
ampio
* Ogni individuo possiede due copie dello stesso gene che possono
essere identiche (omozigoti) o diverse tra loro (eterozigoti)
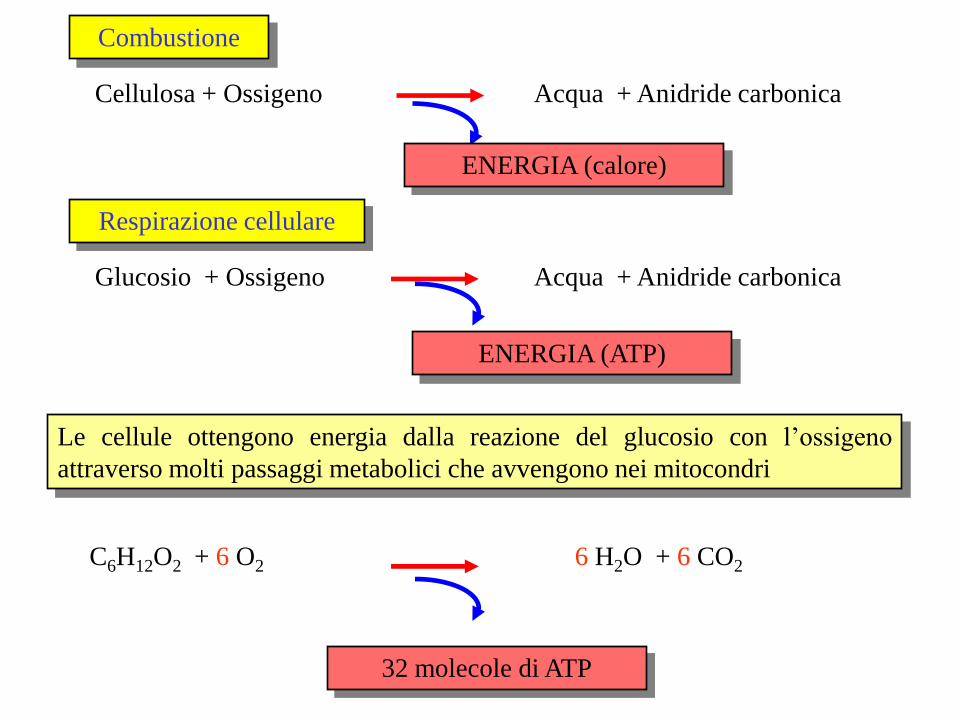
Cellulosa + Ossigeno Acqua + Anidride carbonica
Combustione
Glucosio + Ossigeno Acqua + Anidride carbonica
Respirazione cellulare
ENERGIA (calore)
ENERGIA (ATP)
C6H12O2 + 6 O2
6 H2O + 6 CO2
32 molecole di ATP
Le cellule ottengono energia dalla reazione del glucosio con l’ossigeno
attraverso molti passaggi metabolici che avvengono nei mitocondri
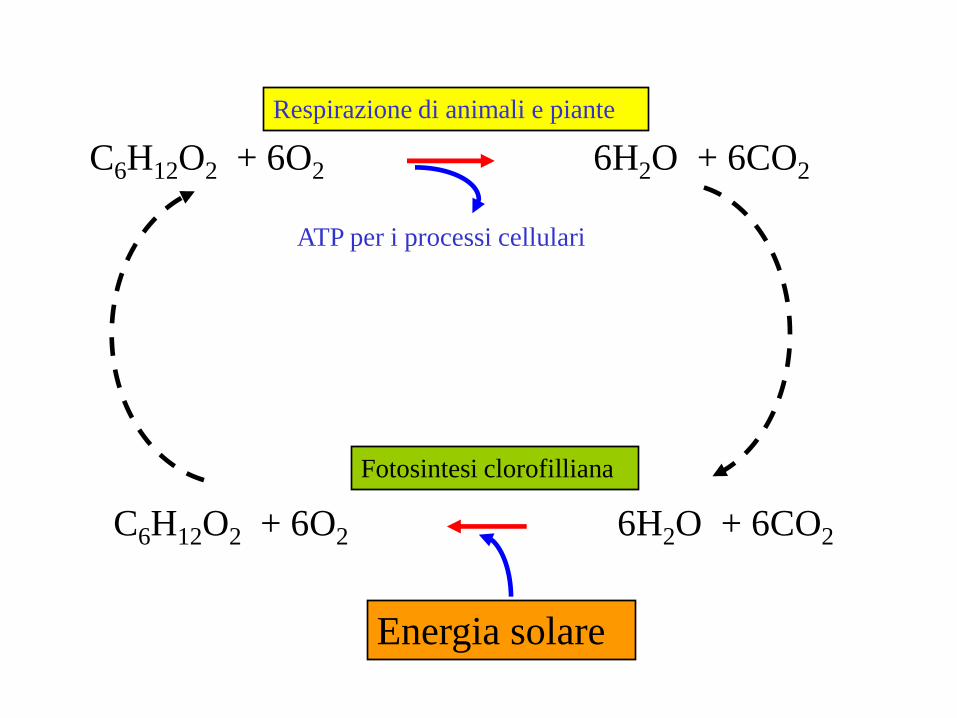
C6H12O2 + 6O2 6H2O + 6CO2
Respirazione di animali e piante
ATP per i processi cellulari
C6H12O2 + 6O2 6H2O + 6CO2
Energia solare
Fotosintesi clorofilliana


















