
Le origini dell’uomo moderno e la sua diffusione nel mondo...
15
1 Le origini dell’uomo moderno e la sua diffusione nel mondo viste attraverso l’identità linguistica Alessandra Colombo-846001 Lo studio dei resti fossili e l’analisi del DNA (in particolare DNA mitocondriale e cromosoma Y) in studi comparativi sulle attuali popolazioni umane portano a supporre un’origine dell’uomo moderno in Africa, ove egli visse con continuità, più a lungo che in altre regioni geografiche e dove mantenne popolazioni di relativa grande consistenza, sviluppando così alti livelli di diversità genetica intra-popolazione. La separazione in gruppi diversi si pensa sia cominciata attorno a 100.000 anni fa, quando successive migrazioni portarono l’uomo moderno alla conquista di nuovi territori, a discapito delle popolazioni di ominidi ivi presenti. Grazie all’identità materiale e linguistica delle varie popolazioni, al loro pool genetico, al contesto geo-climatico pleistocenico delle migrazioni e ai reperti archeologici è possibile ricostruire l’origine africana dell’uomo e risalire alle rotte migratorie seguite dall’uomo moderno nei suoi spostamenti trans-continentali. Introduzione Le origini dell’uomo moderno, hanno da sempre suscitato interesse presso la comunità scientifica. Se in principio la ricerca si avvaleva esclusivamente di discipline quali l’antropologia e l’archeologia, basate su uno studio di tipo morfologico, recentemente a esse si sono affiancati studi di tipo molecolare, geoarcheologico e geocronologico. La comunità scientifica si è scontrata per anni sull’interpretazione dei resti fossili, molti dei quali presentavano caratteristiche variabili a tal punto da non consentire un giudizio unanime sull’effettivo ruolo svolto nell’emersione dell’uomo anatomicamente moderno. Importanti risultati sono stati ottenuti dall’analisi di varie regioni di DNA di un ominide ormai estinto, ma a noi vicino filogeneticamente, Homo neanderthalensis, il cui genoma ha rivelato importanti novità ed è tuttora oggetto di studio. Al fine di comprendere le origini dell’uomo moderno è necessario delineare brevemente lo scenario dell’evoluzione umana a cominciare dalle epoche più antiche. Le prove molecolari dedotte da analisi tra i primati attuali e l’uomo, insieme ai dati paleontologici, convergono nel far risalire la divergenza evolutiva tra le linee che condurranno agli ominidi e alle moderne forme antropomorfe, a un intervallo di tempo compreso tra i 6 e i 7 Ma (Caramelli 2009). Tracce di bipedismo sono, infatti, presenti in resti fossili africani di 6 Ma, in una specie denominata Orrorin tugenensis, (Sawada et al.2002). La postura eretta sembra essere ormai acquisita 4.2 milioni di anni fa, con le prime forme del genere Australopithecus. Questi primi ominidi camminavano in posizione eretta, conservando tuttavia ottime capacità di arrampicamento sugli alberi, grazie alla lunghezza degli arti superiori maggiore degli inferiori. Di taglia ancora piccola, il succedersi di varie specie di questo genere ha dato origine alle prime forme di Homo. A partire da 2 milioni di anni fa si assiste ad un considerevole aumento delle dimensioni corporee e craniche lungo la linea evolutiva umana. La specie africana coinvolta in questo sviluppo è stata denominata Homo ergaster (o Homo erectus africano), caratterizzata da una lunghezza proporzionata degli arti, simile a quella odierna, un aumento di dimensioni del cervello e una riduzione della grandezza dei denti. Fino a quel momento l’evoluzione umana aveva riguardato solo il continente africano; poco sotto i 2 Ma invece, si ritrovano tracce di ominidi nelle regioni medio orientali e nella regione caucasica (Homo georgicus), risultato di una primissima migrazione dall’Africa. Popolazioni umane arcaiche si
-
Upload
phungkhanh -
Category
Documents
-
view
214 -
download
0
Transcript of Le origini dell’uomo moderno e la sua diffusione nel mondo...

1
Le origini dell’uomo moderno e la sua diffusione nel mondo viste attraverso l’identità linguistica
Alessandra Colombo-846001
Lo studio dei resti fossili e l’analisi del DNA (in particolare DNA mitocondriale e cromosoma Y) in studi comparativi sulle attuali popolazioni umane portano a supporre un’origine dell’uomo moderno in Africa, ove egli visse con continuità, più a lungo che in altre regioni geografiche e dove mantenne popolazioni di relativa grande consistenza, sviluppando così alti livelli di diversità genetica intra-popolazione. La separazione in gruppi diversi si pensa sia cominciata attorno a 100.000 anni fa, quando successive migrazioni portarono l’uomo moderno alla conquista di nuovi territori, a discapito delle popolazioni di ominidi ivi presenti. Grazie all’identità materiale e linguistica delle varie popolazioni, al loro pool genetico, al contesto geo-climatico pleistocenico delle migrazioni e ai reperti archeologici è possibile ricostruire l’origine africana dell’uomo e risalire alle rotte migratorie seguite dall’uomo moderno nei suoi spostamenti trans-continentali.
Introduzione
Le origini dell’uomo moderno, hanno da sempre suscitato interesse presso la comunità scientifica. Se in principio la ricerca si avvaleva esclusivamente di discipline quali l’antropologia e l’archeologia, basate su uno studio di tipo morfologico, recentemente a esse si sono affiancati studi di tipo molecolare, geoarcheologico e geocronologico. La comunità scientifica si è scontrata per anni sull’interpretazione dei resti fossili, molti dei quali presentavano caratteristiche variabili a tal punto da non consentire un giudizio unanime sull’effettivo ruolo svolto nell’emersione dell’uomo anatomicamente moderno. Importanti risultati sono stati ottenuti dall’analisi di varie regioni di DNA di un ominide ormai estinto, ma a noi vicino filogeneticamente, Homo neanderthalensis, il cui genoma ha rivelato importanti novità ed è tuttora oggetto di studio. Al fine di comprendere le origini dell’uomo moderno è necessario delineare brevemente lo scenario dell’evoluzione umana a cominciare dalle epoche più antiche. Le prove molecolari dedotte da analisi tra i primati attuali e l’uomo, insieme ai dati paleontologici, convergono nel far risalire la divergenza evolutiva tra le linee che condurranno agli ominidi e alle moderne forme antropomorfe, a un intervallo di tempo compreso
tra i 6 e i 7 Ma (Caramelli 2009). Tracce di bipedismo sono, infatti, presenti in resti fossili africani di 6 Ma, in una specie denominata Orrorin tugenensis, (Sawada et al.2002). La postura eretta sembra essere ormai acquisita 4.2 milioni di anni fa, con le prime forme del genere Australopithecus. Questi primi ominidi camminavano in posizione eretta, conservando tuttavia ottime capacità di arrampicamento sugli alberi, grazie alla lunghezza degli arti superiori maggiore degli inferiori. Di taglia ancora piccola, il succedersi di varie specie di questo genere ha dato origine alle prime forme di Homo. A partire da 2 milioni di anni fa si assiste ad un considerevole aumento delle dimensioni corporee e craniche lungo la linea evolutiva umana. La specie africana coinvolta in questo sviluppo è stata denominata Homo ergaster (o Homo erectus africano), caratterizzata da una lunghezza proporzionata degli arti, simile a quella odierna, un aumento di dimensioni del cervello e una riduzione della grandezza dei denti. Fino a quel momento l’evoluzione umana aveva riguardato solo il continente africano; poco sotto i 2 Ma invece, si ritrovano tracce di ominidi nelle regioni medio orientali e nella regione caucasica (Homo georgicus), risultato di una primissima migrazione dall’Africa. Popolazioni umane arcaiche si

2
diffondono anche in Asia (Homo erectus asiatico), dove sopravvivranno fino a circa 40.000 anni fa. Una particolare specie con ridottissime dimensioni corporee, tanto da essere considerata in un primo momento una forma patologica, è stata rinvenuta in un’isola indonesiana (Homo floresiensis). Essa rappresenterebbe una forma di nanismo insulare, un fenomeno che ha riguardato popolazioni molto arcaiche, precocemente arrivate in questi territori (probabilmente prima di 2 Ma). I discendenti di Homo ergaster o Homo erectus dei territori europei e africani sono stati classificati come appartenenti a varie specie: Homo antecessor e successivamente Homo heidelbergensis per le forme europee, Homo rhodesiensis per le forme africane. Un tempo venivano classificati generalmente come Homo sapiens “arcaico,” etichetta che ad oggi non ha più alcun significato (Caramelli 2009). A partire da 200.000 anni fa, si assiste in Europa, alla comparsa di caratteristiche morfologiche distintive di una particolare forma umana, Homo neanderthalensis, la quale è sopravvissuta fino a circa 28.000 anni fa. Nello stesso momento in Africa, si assiste alla comparsa delle prime forme di Homo con caratteri morfologici moderni, vedasi il cranio di Herto, datato a 160.000 anni fa (White et al.2003). La comparsa di Homo sapiens anatomicamente moderno, nel resto dei continenti, è attesa a partire da 70.000 anni fa, per le regioni asiatiche e da circa 40.000 anni fa per le regioni europee. Tuttavia, le prime forme extra-africane, che presentano una morfologia con parziali caratteristiche moderne, sono presenti in Medio Oriente poco meno di 100.000 anni fa. L’appellativo “anatomicamente moderno” è stato usato per identificare i reperti che presentano caratteristiche morfologico-scheletriche del tutto simili all’umanità attuale, e per distinguerli dai precedenti ominidi con caratteristiche più antiche. Il rapporto evolutivo intercorso tra gli individui moderni e i vari rappresentati di altri tipi umani, ancora presenti nelle prime fasi della loro diffusione, è tuttora incerto. Gli interrogativi che sono sorti si possono ricondurre a tre grandi questioni: � É avvenuto un cambiamento nel tempo
all’interno di una singola linea evolutiva, da
un’unica specie arcaica collocata in uno specifico continente, o la comparsa dell’uomo moderno ha interessato tutto il vecchio mondo?
� Se come sembra, l’espansione è avvenuta in Africa, gli individui di H. sapiens hanno sostituito le genti arcaiche che incontravano lungo il loro cammino, o ci sono stati incroci genetici prolifici?
� Inoltre, se questi incroci ci sono stati, in che percentuale hanno contribuito alla costituzione del pool genetico moderno? (Caramelli 2009)
L’origine dell’uomo moderno Quando e dove sorse la nostra specie? Negli ultimi decenni, numerosi studi di evoluzione molecolare hanno cercato di dare riposta a questo interrogativo. Sono state avanzate due principali ipotesi sull’evoluzione dell’uomo moderno, sulla base di studi condotti negli anni ‘80 e nel 2000 (Stringer and Wolpoff), le quali concordano sul fatto che Homo erectus sia il genere più vicino filogeneticamente a Homo sapiens, e che esso uscì dall’Africa e si diffuse nel resto del mondo circa 1.8 Ma (Ingman et al.2000). L’ipotesi delle origini africane recenti o Out of Africa (Vigilant et al.1991;Cann et al.1987) teorizza che l’uomo moderno si sia originato in Africa, in un periodo compreso tra i 100.000 e i 200.000 anni fa, e successivamente si sia spostato nel resto del mondo, andando a rimpiazzare gruppi di ominidi indigeni, con un piccolo o nullo rimescolamento genetico. Si esclude quindi qualsiasi evento di interbreeding (Stringer e Andrews 1988). L’ipotesi concorrente è il modello multiregionale. Essa propone che l’uomo moderno non si sia originato esclusivamente in Africa bensì da popolazioni di Homo erectus dislocate in varie zone del mondo, le quali furono soggette a interbreeding. A sostegno di tale ipotesi vi sono evidenze fossili di una continuità culturale e morfologica tra uomini arcaici e moderni al di fuori del continente africano (Wolpoff 1989). Tuttavia numerosi studi basati su resti fossili, analisi del DNA (mtDNA e del cromosoma Y),

3
studi geoarcheologici e geocronologici, e studi linguistici supportano l’ipotesi dell’origine africana di Homo sapiens (Out of Africa Hypothesis; Vigilant et al.1991; Cann et al. 1987; Horai et al.1995; Ruvolo et al.1993;Hedges 2000; Ingman et al.2000; Scozzari et al.2014; Muttoni et al.2010;Henn et al.2011; Atkinson 2011; Sforza 2010). Una nuova prospettiva verrebbe dal leaky model (Gibbons 2011) secondo il quale vi sarebbero stati almeno due eventi di interbreeding tra gli uomini arcaici e moderni nel corso della loro convivenza (figura 1). Vengono quindi rifiutate, almeno in parte, le ipotesi precedenti e viene avanzata l’ipotesi di una speciazione di Homo sapiens posteriore a tali eventi. I dati genomici suggeriscono solo un piccolo tasso di interbreeding, presumibilmente avvenuto ai margini, dove l’avanzata dell’uomo moderno incontrò gruppi arcaici, discendenti di H. erectus,
usciti dall’Africa 1.8 milioni di anni fa. Cosi come affermò John Relethford, un antropologo americano: “La strada seguita dall’evoluzione è piuttosto chiara: i nostri antenati si originarono in Africa, ma cosa successe dopo è più complesso” (Gibbons 2011).
Fig.1 I due modelli dell’origine dell’uomo moderno (a
sinistra) contestati da nuove rivelazioni basate su un antico
DNA, che suggerisce l’avvento di un limitato interbreeding
tra popolazioni arcaiche e moderne (Gibbons 2011).
Il sequenziamento dell’intero genoma nucleare di tre individui di sesso femminile di H. neanderthalensis, nel 2010, portò a nuove evidenze. Vissuti più di 38.000 anni fa, in Croazia, essi mostrano caratteri genetici comuni alle odierne popolazioni asiatiche ed europee, ma non a quelle africane. Si tratta di un 1-4% di DNA nucleare condiviso da queste popolazioni e che può essere ricondotto a H.neanderthalensis.
Secondo studi condotti da un team di scienziati guidati da Svante Pääbo, del Max Planck Institute di Lipsia, tale evidenza genetica si spiegherebbe con l’origine dell’uomo in Africa a cui seguirono eventi di incroci con Neanderthal in Medio Oriente o in Arabia, prima che avvenisse la dispersione dell’uomo moderno nel resto dell’Asia e in Europa, tra i 50 e gli 80 mila anni fa. Il ritrovamento di resti ossei nella Denisova Cave, sui Monti Altaj, nella Siberia del sud, portò alla scoperta di un secondo possibile evento di interbreedig. I resti ossei appartengono a un individuo facente parte di un gruppo di ominidi, i Denisoviani, staccatosi precocemente dai Neanderthal, datato a più di 50.000 anni fa. Comparando parti del genoma di Denisova con lo stesso segmento di DNA in 53 popolazioni umane odierne, il team trovò che i Denisoviani condividono da un 4% al 6% di DNA con popolazioni melanesiane, della Nuova Guinea e delle Bougainville Islands. Tali segmenti non sono mai stati trovati nei Neanderthal o in altre popolazioni viventi. Lo scenario più probabile prevede che i Neanderthal e le popolazioni di Denisova si separarono intorno a 200.000 anni fa. L’uomo moderno s’incrociò con i Neanderthal, quando lasciò il continente africano, intorno a 100.000 anni fa. Così si spiegano le tracce genetiche riscontrate nelle popolazioni asiatiche ed europee. Più tardi, un piccolo sottogruppo di H. sapiens, portatore di tracce genetiche conseguenti a interbreeding, si diresse verso est, in particolare verso la Melanesia e lì si incrociò con l’uomo di Denisova. Come risultato si ebbe che i Melanesiani ereditarono il DNA sia dai Neanderthal che dai Denisoviani: un 8% del loro DNA deriverebbe quindi da popolazioni arcaiche. Ne deriva in conclusione, che H. sapiens si mischiò con almeno due popolazioni arcaiche, in luoghi e tempi diversi (Gibbons 2011). Il dibattito è tuttora molto acceso e serviranno ulteriori evidenze per far luce sull’origine dell’uomo moderno.
Le analisi filogenetiche basate su studi del DNA Negli ultimi decenni, gli studi filogenetici basati su analisi del DNA mitocondriale e del

4
cromosoma Y hanno portato a delineare nuove prospettive. A causa delle sue caratteristiche, il tipo di DNA più usato fino ad oggi per studi di evoluzione umana e analisi filogenetiche, è il DNA mitocondriale (Caramelli 2009). L’utilizzo di geni mitocondriali presenta, infatti, alcuni vantaggi rispetto a quelli nucleari: si ha la disponibilità di un elevato numero di copie; un’apparente mancanza di ricombinazione genica (Olivio et al.1983); un elevato tasso di sostituzione genica, maggiore di quello dei geni nucleari (Brown et al.1979) e un’ereditarietà per linea materna (Giles et al.1979; Ingman et al.200). Il DNA posseduto all’interno di una specie è maggiormente diversificato, tanto più antiche sono le parentele tra gli individui. Lungo la genealogia, in questo caso materna, aumentano quindi le mutazioni e il numero di discendenti di un ipotetico antenato comune. Nel caso della trasmissione dl mtDNA, ciascuna donna può:
a) Non avere figli o avere solo maschi b) Avere solo figlie femmine c) Avere sia figlie femmine che maschi
I primi due casi porterebbero alla mancata trasmissione del materiale genetico mitocondriale alle successive generazioni e quindi all’estinzione di quel tipo. Ciò può succedere casualmente ad ogni generazione, e con il tempo, questa estinzione può anche riguardare tutti i tipi mitocondriali, tranne uno ovviamente. Se ciò è vero, esiste un’unica antenata per tutte le sequenze attuali, la quale non è stata l’unica femmina presente nella popolazione, ma l’unica che abbia beneficiato di condizioni riproduttive favorevoli (Looking for Eve Hypothesis) (Biondi e Rickards 2003;Caramelli 2009). Nel corso della storia recente vi è stata una vera e propria evoluzione degli studi genetici-molecolari. Negli anni ‘80, la maggior parte degli studi sulle molecole di mt dell’uomo veniva condotto attraverso RFLP analisi (Restriction-enzyme Fragment Lenght Polymorphysm), ricavando dati al fine di stimare il tasso evolutivo e temporizzare gli eventi evolutivi (Torroni et al.1998). Negli anni ‘90, si è passati all’utilizzo dei marcatori molecolari HVS-1 (Hyper Variable Segment), basandosi sul sequenziamento di mtDNA
confinato alle sole regioni di controllo (regioni non codificanti),le quali costituiscono meno del 7% del genoma mitocondriale complessivo (Ingman et al.2000). L’estrema variabilità del tasso di sostituzione dei siti e le conseguenti mutazioni parallele, causavano tuttavia delle difficoltà nella stima delle distanze genetiche e nelle ricostruzioni filogenetiche (Tamura e Nei 1993;Maddison et al.1992). Negli anni 2000 si è passati all’utilizzo del completo sequenziamento di mtDNA: da un 7% di DNA utilizzato con le HVS a un 100% di genoma mitocondriale in uso. L’incrementata mole di dati disponibili permette quindi una maggiore probabilità di risolvere la filogenesi (Ingman et al.2000). Con lo sviluppo delle moderne tecniche di biologia molecolare, come la Polymerase Chain Reaction (PCR), lo studio dell’origine dell’umanità fu affrontato anche dal punto di vista genetico e non più solamente paleontologico. Viene utilizzato sia materiale biologico d’individui attuali, sia DNA estratto da reperti antichi come le ossa. Tutti i lavori basati sull’analisi del DNA poggiano sul fatto che in alcune regioni di questa molecola, esiste un costante accumulo di mutazioni neutrali. Tramite l’utilizzo dell’orologio molecolare si può fornire così una stima del tempo di divergenza da un antenato comune di due o più sequenze o popolazioni (Biondi e Rickards 2003; Caramelli 2009). Un importante studio risalente al 2000 (Ingman et al.2000) segna la svolta nelle ricostruzioni filogenetiche: viene sequenziato l’intero genoma mitocondriale di 53 individui appartenenti a 14 phyla linguistici differenti. Si prende quindi in considerazione l’identità materiale delle comunità umane, in particolare la lingua. I dati ottenuti sono stati confrontati con quelli di studi paralleli (Kaessmann et al.1999), effettuati su determinate regioni di mtDNA, Xq13.3, negli stessi individui, portando ad una visione concorrente sull’evoluzione umana. Il risultato è la ricostruzione di un albero filogenetico (fig.2) con una buona robustezza in nodi, in termini di Bootstrap (un metodo di ricampionamento statistico con ripetizione).

5
Fig.2 Filogramma Neighbour-joining basato sul
sequenziamento completo di sequenze genomiche di mtDNA
(escludendo le zone D-loop). Dati ottenuti con PAUP*4.0
Beta (Sinauer Associates) e con 1.000 replicati Bootstrap, i
cui valori sono segnati in corrispondenza di ciascun nodo. Il
nodo marcato con * si riferisce a MRCA (Ingman et al.2000).
Si evidenzia, come il ramo più antico sia statisticamente significativo (NJ Bootstrap =100) e porti esclusivamente a linee africane, subsahariane. L’albero neighbour joining (NJ) si pone quindi come evidenza dell’origine africana dell’uomo moderno, secondo la teoria out of Africa, e pone gli Africani alla base dell’albero, in quanto clade più antico (Ingman et al.2000; Hedges 2000). I gruppi linguistici africani sono rappresentati in verde e in viola e si trovano alla base dell’albero. In giallo e in blu sono invece rappresentati i gruppi non africani, rispettivamente: asiatico ed europeo. Il nodo contrassegnato con un asterisco, con valore di Bootstrap pari a 98, è il punto da cui si dipartono i raggruppamenti monofiletici: asiatico, europeo ed africano. È il MRCA (Most Recent Common Ancestor), il più recente clade contenente sia
individui africani che non-africani (Ingman et al.2000). Interpretato letteralmente, l’albero indica come un particolare sottogruppo di Africani (contrassegnati in verde) sia più vicino filogeneticamente agli Europei e agli Asiatici rispetto che ad altri Africani (contrassegnati in viola).Vi è, infatti, un gruppo di 6 sequenze di Africani (sequenze dalla 33 a 38, in fig.2), che è geneticamente distante dagli altri Africani. Tale lignaggio rappresenterebbe i discendenti di una popolazione che diede evidentemente origine all’intero lignaggio dei non-Africani. Tuttavia la storia di un singolo gene o molecola può non sempre essere lo specchio di una popolazione; altri studi molecolari mostrano gli Africani come singolo gruppo (Nei e Roychoudhury 1993). È da sottolineare, infatti, come gli studi svolti unicamente su mtDNA riflettano unicamente la storia genetica in linea femminile (Ingman et al.2000). La maggioranza delle evidenze genetiche indica che ci furono solo all’incirca 10.000 individui che s’incrociarono per lungo tempo, prima della recente espansione dell’uomo moderno fuori dall’Africa (Harpending et al.1998). Una così piccola popolazione risulta incompatibile con il modello multiregionale, che richiederebbe un numero di individui ben maggiore per mantenere un flusso genetico tra i continenti (Hedges 2000). In conclusione, questi studi suggeriscono la presenza in passato di una popolazione fondatrice vissuta in Africa, portatrice del proprio set di alleli mitocondriali, e che le popolazioni africane hanno continuato a incrociarsi tra loro dopo l’esodo (Hedges 2000). L’orologio molecolare potrebbe rispondere in modo soddisfacente al quesito di dove venissero i nostri progenitori. Ma la specie più vicina al genere Homo, che sia H. erectus o qualsiasi altra specie, è sfortunatamente estinta. I fossili più antichi del moderno H. sapiens risalgono a 130.000 anni fa, tale data risulta essere quindi il limite superiore dell’origine della nostra specie (Lewin 1999). Studi su DNA fossili portano tuttavia a soglie inferiori. La separazione tra H. neandertalensis (una specie che non è necessariamente il nostro parente più prossimo) e

6
H. sapiens è stata stimata dall’orologio molecolare a 465.000 anni fa (Krings et al.2000). A fronte di ciò, la nostra specie probabilmente si originò tra i 130.000 e i 465.000 anni fa (Hedges 2000; Ingman et al.2000). Il tempo di divergenza tra uomo e scimpanzé viene stimato a 5 milioni di anni fa, basandosi su evidenze paleontologiche (Andrews 1992) e genetiche (Kuman e Hedges 1998), con un tasso di mutazione di 1.70*10-8 sostituzioni per sito per anno. L’età dell’antenato comune più recente per mtDNA, valutata sulla base della massima distanza tra due uomini (5.82*10-3 sostituzioni per sito tra Africani Mkamba e Sans) è stata datata a 171.500± 50.000 anni fa. Mentre, l’età del MRCA per il clade più giovane contenente sia sequenze di Africani che non-Africani, calcolata dalla distanza media di ciascun membro del clade rispetto al nodo condiviso (8.85*10-4 sostituzioni per sito), risulta essere di 52.000 ± 27.500 anni fa (Ingman et al.2000). Questa data potrebbe essere tuttavia troppo recente. Altri marcatori genetici, infatti, datano l’esodo dall’Africa intorno ai 100.000 anni fa (Nei e Roychoudhury 1993; Tishkoff et al.1996), data che ben concorda con le evidenze fossili e archeologiche di uomini moderni fuori dall’Africa (Ingman et al.2000). Recenti analisi del DNA (Green et al.2008) retrodaterebbero il tempo di divergenza tra H. neanderthalensis e il lignaggio umano a circa 800.000 anni fa (660.000 ± 140.000 anni). Questa divergenza potrebbe essere dovuta a popolazioni di ominidi che rimasero largamente isolate dalla riserva riproduttiva africana dopo la migrazione in Europa, intorno a 0.95-0.87 Ma, quando evolse in nuova specie. L’analisi è stata eseguita tramite la ricostruzione delle sequenze dell’intero genoma mitocondriale di un individuo di H. neanderthalensis di 38.000 anni fa, utilizzando 8.341 sequenze, identificate tra 4.8 Gb di DNA estratto da circa 0.3 g di ossa (Green et al.2008; Pennisi 2009). L’Africa è stata ormai confermata quale sorgente di tutti gli uomini moderni, ma la caratterizzazione delle variazioni genetiche e delle relazioni esistenti tra le varie popolazioni attraverso i continenti è tuttora enigmatica.
L’ équipe di Sarah Tishkoff (Tishkoff et al.2009) si è occupata di studiare 121 popolazioni africane, 4 popolazioni afro-americane e 60 non africane, con pattern di variazione di 1.327 microsatelliti nucleari e marcatori di inserzione/delezione. Sono stati identificati 14 cluster di popolazioni ancestrali in Africa in relazione all’etnia culturale, ovvero in base alla condivisione di proprietà linguistiche e/o culturali. Si osservano in molte popolazioni alti livelli d’incrocio avvenuti in passato: un riflesso delle migrazioni storiche trans-continentali compiute dagli antenati. I dati mostrano inoltre, l’evidenza di un’antichità condivisa tra le popolazioni di cacciatori-raccoglitori delle diverse zone geografiche. L’uomo moderno si originò in Africa c.a. 200.000 anni fa e si disperse nel resto del globo nei passati 100.000 anni (Campbell et al.2008). Gli uomini moderni sono vissuti con continuità in Africa, più a lungo che in qualsiasi altra regione geografica, e ivi hanno mantenuto una dimensione della popolazione effettiva relativamente ampia, con un conseguente alto livello di variabilità genetica intra-popolazione (Campbell et al.2008; Reed et al.2006). L’Africa si compone di più di 2.000 gruppi etnolinguistici differenti, rappresentanti circa un terzo delle lingue del mondo (www.ethnologue.com). Ad eccezione di qualche caso isolato, che non mostra chiare correlazioni con le altre lingue, esse sono state classificate in quattro grandi famiglie: lingua niger-kordofoniana (parlata in ampie regioni africane), Afroasiatica (parlata per lo più nel Sahara, NE- E Africa), nilo-sahariana (parlata per lo più in Sudan, Sahara e E-Africa) e khoesana (linguaggio contente dei suoni consonantici come click, parlata dai San, S-Africa e dagli Hadza e Sandawe, E-Africa) (Tishkoff et al.1996). Nonostante l’importanza della genetica di popolazione africana, la diversità genetica all’interno delle varie popolazioni africane non è ancora caratterizzata. A causa della considerevole variabilità ambientale, le popolazioni africane mostrano un ampio range di variazione linguistica, culturale e fenotipica (Campbell et al.2008; Reede et al.2006). A fronte di ciò, risulta difficile sia ricostruire la storia evolutiva umana che chiarire le relazioni tra le varie popolazioni (Tishkoff et al.2009).

7
Le popolazioni africane e afroamericane, ad eccezione dei Dogon del Mali, presentano i più alti livelli di diversità genetica intra-popolazione (Ɵ=4Neų, dove il livello di diversità genetica è basato sulla variazione della lunghezza degli alleli microsatelliti; Ne è la dimensione della popolazione effettiva; e ų il tasso di mutazione dei microsatelliti). In aggiunta la variabilità genetica declina all’aumentare della distanza dall’Africa, in accordo con la teoria dell’effetto fondatore seriale e del modello di isolamento per distanza (IBD), risultante dalla migrazione degli uomini moderni dall’Africa verso il resto del mondo (Rosenberg et al.2002; Rosenberg et al.2006; Ramachandran et al 2005). All’interno dell’Africa, i San (Boscimani), popolazioni di cacciatori-raccoglitori che vivono attualmente in Africa meridionale, sono stati probabilmente una delle prime comunità a essersi separate dalla popolazione umana moderna ancestrale. Inoltre i San, i Khoi (Ottentotti) e alcuni gruppi di Pigmei dell’Africa centrale presentano la più grande diversità genetica tra le popolazioni africane. In accordo con l’ipotesi di un flusso genico bidirezionale, le popolazioni africane e del Medio Oriente condividono il maggior il numero di alleli, che risultano invece essere assenti nelle altre popolazioni. Mentre si osserva in Africa la maggior variabilità genetica intra-popolazione, sono le popolazioni dell’Oceania e i Nativi Americani a mostrare la maggior variabilità inter-popolazione, 8.36% e 4.59% rispettivamente (Thiskhoff et al.2009).
Il contesto geo-climatico delle migrazioni umane nel Pleistocene I cambiamenti climatici sono alla base delle modificazioni evolutive, qualora tali cambiamenti avvengano troppo repentinamente, le popolazioni non hanno il tempo sufficiente per adattarvisi, ed è probabile che si verifichino eventi di estinzione più che di speciazione. Nel periodo chiave per l’evoluzione di forme umane moderne, si registrarono ampie oscillazioni climatiche dovute alle glaciazioni del Pleistocene. Tra 100.000-200.000 anni fa, infatti, l’aridità causata dall’espansione dei ghiacciai in Europa e in tutto
l’emisfero boreale, costrinse le popolazioni africane a rifugiarsi in aree isolate e non ancora interessate da questi fenomeni. È quindi probabile che da qualcuna di queste popolazioni siano emersi i caratteri morfologici moderni, poiché i cambiamenti evolutivi possono presentarsi e fissarsi molto più velocemente se riguardano gruppi piccoli e isolati (Caramelli 2009). Una valutazione critica delle età magnetostratigrafiche e/o radiometriche (Muttoni et al.2010), rispetto ai resti fossili di ominidi e alle industrie litiche in siti chiave del Sud-Europa (Italia, Francia e Spagna), porta a pensare che la presenza dei primi ominidi in tale zona, sia da ricondursi a un intervallo di tempo tra, il Jaramillo subchron e il limite Brunhes-Matuyama (subchron C1r.1r,0.99-0.78 Ma). Questa finestra di tempo di c.a. 200.000 anni, coincide ampiamente con la transizione climatica globale del tardo Pleistocene Inferiore, all’interno del quale si osserva lo stadio isotopico marino (MIS) 22 (c.a.0.87Ma): il primo stadio freddo importante del Pleistocene. Lo studio condotto da Giovanni Muttoni e la sua équipe, nel 2010, suggerisce che l’aridificazione del Nord-Africa e dell’Est-Europa, particolarmente rigida durante il 22 MIS, provocò impulsi migratori di grossi erbivori, in particolar modo di elefanti, da queste regioni verso i rifugi del sud Europa, e gli uomini migrarono con loro. Si pensa quindi a una comune modalità di dispersione di uomini e elefanti, avvenuta nel tardo Pleistocene Inferiore, dai luoghi originari della savana africana verso il sud Europa, il buen retiro di elefanti e ominidi. La magnetostratigrafia è una tecnica frequentemente utilizzata nella datazione di sequenze stratigrafiche contenenti resti umani e artefatti. Il metodo consiste nel ricostruire, con appropriate tecniche di laboratorio, la sequenza degli eventi d’inversione della polarità magnetica terrestre, i quali vengono registrati nelle rocce grazie alla presenza di minerali ferromagnetici. Questi sedimenti, infatti, hanno la proprietà di fissare la direzione del campo magnetico terrestre al momento della loro deposizione sedimentaria.
Nel corso della storia della Terra il campo magnetico ha subito numerose inversioni, a lungo studiate e datate. La sequenza delle

8
magnetizzazioni delle rocce che si desidera datare viene confrontata con sequenze note delle inversioni del campo terrestre. Al pari di qualsiasi tecnica analitica, uno studio magnetostratigrafico deve rispettare alcuni criteri di qualità, in modo da ottenere risultati affidabili, riassunti recentemente da Langereis et.al. (2010). Gli studi magnetostratigrafici pionieri di Parés e Pérez-Gonzalez (1995) portarono a una prima robusta evidenza della presenza di ominidi nel nord della Spagna, a Gran Dolina (Carbonell et al.1995) fin dal Pleistocene Inferiore, nello specifico nella reversione del campo magnetico del tardo Matuyama chron, tra il Brunhes-Matuyama boundary (0.78 Ma) e probabilmente dopo il Jaramillo subchron (0.99 Ma). Un profilo magnetostratigrafico molto simile pone evidenze di ominidi in sedimenti a polarità inversa, appena al di sotto della transizione correlata al limite Brunhes-Matuyama, nel vicino sito di Sima del Elefante (Parés et al.2006). Recentemente dati cosmogenici sepolti di Sima del Elefante sembrano indicare un’occupazione di ominidi precedente, intorno a 1.1-1.2 Ma (Carbonell et al.2008), che dovrebbe collocarsi nel Matuyama, prima del Jaramillo. Nel bacino di Guadix-Baza nel sud della Spagna, è stato trovato il primo sito datato per la presenza di ominidi, risalente a un generico intervallo pre-Brunhes-Matuyama boundary (Oms et al.2000). Esso è stato nuovamente datato all’età pre-Jaramillo, con un intervallo di 1.2-1.25 Ma, utilizzando dati magnetostratigrafici di un profilo composito (Scott et al.2007). In questo quadro, un candidato per essere il più antico sito di occupazione in Europa potrebbe essere il sito di Pirro Nord, in Sud-Italia, dove sono state usate evidenze biostratigrafiche per datare l’occupazione umana tra 1.3-1.7 Ma (Arzarello et al.2007).Il momento preciso della prima colonizzazione del Sud-Europa da parte di ominidi è di difficile datazione, per le età controverse assegnate ai vari siti. Tale fatto è ben esemplificato nel caso dell’uomo di Ceprano, vissuto in centro-Italia, datato durante il limite Brunhes-Matuyama (Manzi 2004), fino a recenti studi che lo vedono invece di c.a.0.45 Ma (Muttoni et al.2009). Esso non si colloca più quindi, tra i siti candidati tra i più antichi del Sud-
Europa, bensì in epoca assai più recente. Spesso, associati ai siti di occupazione, si ritrovano dei manufatti litici, ascrivibili a due principali tecnologie: Oldowan e Acheulean. Il ritrovamento di tali artefatti può essere di aiuto nella datazione dei siti. Gli strumenti ricavati con la tecnica olduvaiana, risalgono a 2.6-2.5 Ma (Semaw et al.1997; Quade et al.2004) e furono prodotti in Africa da semplici strategie di riduzione litica, consistenti nella scheggiatura di pietre. La tecnologia acheuleana, utilizzata in Africa c.a.1.6-1.5 Ma (Asfaw et al.1992; Quade et al.2004), è più avanzata, e consiste nella rielaborazione della forma delle pietre fino a produrre strumenti bifacciali. L’esistenza della tecnologia olduvaiana in Europa, mentre in Africa persisteva quella acheuleana, può riflettere una deriva culturale, in condizioni di bassa densità demografica, che limitò il mantenimento e la trasmissione della tecnologia acheuleana, più sofisticata, in ambienti scarsamente popolati (Lycett e von-Cramon-Taubedel 2008).La datazione del sito di Pirro Nord è stata possibile, anche grazie al ritrovamento di artefatti ed ecofatti, con strumenti ascritti alla tecnologia olduvaiana e faune a mammiferi appartenenti alla Pirro Faunal Unit (FU) (Arzarello et al.2007). Tuttavia studi magnetostratigrafici svolti su sequenze lacustri di Leffe, in Nord-Italia (Muttoni et al.2007), con associazioni a mammiferi compatibili con la Pirro FU, mostrerebbero una datazione di c.a. 1 Ma, a cui si possono far risalire anche i manufatti litici di Pirro Nord. Dagli studi magnetostratigrafici di Kent et al.1971, si evince l’evidenza di un primo raffreddamento avvenuto intorno a 1.2 Ma, che si fece molto intenso tra il Jaramillo subchron e il Brunhes-Matuyama boundary (Chron C1r.1r. 0.99-0.78 Ma), suggerendo una corrispondenza con l’inizio della glaciazione alle medie latitudini, in Europa e Nord America. L’analisi del δ18O su foraminiferi planctonici di Berger et al.(1993) confermò un brusco cambiamento climatico globale avvenuto a partire da 40.000 anni fa, con ampie oscillazioni, tra MIS 23 e MIS 22, ed egli chiamò questo cambiamento, “climate revolution”.

9
Fig.3 Scenario paleogeografico dell’Europa mostrante le possibili rotte migratorie seguite dai grandi erbivori (in particolar modo
dagli elefanti) e dagli uomini durante il tardo Pleistocene Inferiore, c.a. 0.87 Ma (MIS 22). Sono illustrati nella mappa
esclusivamente i siti che mostrano prove della presenza di ominidi ed erbivori. Vengono inoltre rappresentate le varie zone
climatiche-geografiche, caratteristiche del periodo pleistocenico analizzato (Muttoni et al.2010).
Il cambiamento climatico globale avvenne tra MIS 36 (c.a. 1.2 Ma) e MIS 13 (0.54-0.46),e all’interno di questa transizione, MIS 22 (c.a. 0.87 Ma) rappresenta la prima grande glaciazione continentale pleistocenica dell’emisfero boreale (Shackleton e Opdyke 1976; Berger et al.1993;Shackleton 1995; Lourens et al.2004). Tale transizione climatica avvenne nel Chron C1r.1r, la stessa finestra temporale calcolata per la presenza delle prime popolazioni del Sud-Europa. È possibile che la globale transizione climatica nel MIS 22, del tardo Pleistocene Inferiore, abbia causato un critico cambiamento climatico in Africa, la patria degli ominidi, così da spingerli a migrare dall’Europa all’Europa del sud. In aggiunta, un’aridificazione progressiva in Africa e nell’est Europa, durante il tardo Pleistocene Inferiore, particolarmente nel MIS 22, portò alla ricerca di migliori rifugi nel sud Europa, da parte di mammiferi, compreso l’uomo (Muttoni et al.2010).
Vi sono molti argomenti e dati che considerano il diffondersi in Europa di emigranti asiatici e africani come fattore scatenante del più importante turnover faunistico del tardo Pleistocene Inferiore (fauna medio galeriana in Italia; fauna cromeriana NE Europa; fauna
triaspoliana E, Europa), intercorso durante il MIS 22, c.a. 0.87Ma (Head e Gibbard 2005). Vedasi la migrazione di Elephas antiquus dall’Africa, e di Mammuthus trogontherii dall’Asia. Tale cospicua migrazione deve avere presumibilmente lasciato delle tracce nei record geologici e archeologici africani e asiatici. La Danakil depression nella zona nord-est africana, è una probabile area di tappa della migrazione. Vi sono stati trovati, infatti, resti di ominidi associati ad ambienti ricchi d’acqua, risalenti al Jaramillo-Matuyama boundary (0.99 Ma) (Martinez-Navarro et al.2004;Albianelli e Napoleone 2004). Vi sono varie ipotesi sulle possibili rotte migratorie intraprese da uomini e animali in fuga dalla terra di origine, dedotta situarsi nell’est Africa (probabilmente nella depressione Danakil), verso il Sud-Europa (fig.3). La via più verosimile intrapresa dai grandi erbivori e dagli ominidi è attraverso la valle del Nilo, verso le rive del Mediterraneo del sud. Dal corridoio del Sinai andarono fino al Levante, e da qui la migrazione di ominidi e erbivori sarebbe proseguita verso l’Europa dell’est attraverso il Bosforo, esposto a fasi alterne nel Pleistocene o, in alternativa, attraverso il condotto del paleo-Eufrate verso la Transcaucasia e da qui verso le coste del Mar Nero. Entrambe queste rotte (Bosforo e Eufrate)

10
sono sostenute da ritrovamenti di strumenti del Paleolitico Inferiore attraverso la Turchia (Khun 2002), e da evidenze in Israele del Pleistocene Inferiore (e.g. Gesher Benot, Ya’aqov, Evron, e ‘Ubeidiya). Successivamente, dall’est Europa o dalle coste del Mar Nero, proseguirono verso ovest attraverso il corridoio del Danubio. Raggiunsero il Nord-Italia, in particolare la valle del Po, la quale rimase largamente esposta, sopra il livello del mare, per la prima volta nel Pleistocene, nel MIS 22. Da qui, essi andarono a colonizzare il Sud-Italia, e attraverso le Alpi Marittime e i Pirenei raggiunsero anche Francia e Spagna (Fig.3). Vengono invece escluse le vie migratorie ipotizzate attraverso lo Stretto di Gibilterra e il Canale di Sicilia, entrambe sommerse dal mare in quel periodo. Lo stretto di Gibilterra è stato aperto sull’Atlantico sino alla fine del Miocene, e il livello variabile del mare pleistocenico non interessò in modo sostanziale i c.a. 5 km di profondità del canale. In accordo con Straus (2001), i dati geocronologici e archeologici indicano che solo alla fine del Paleolitico (c.a.11 ka) si ebbe un possibile attraversamento umano dello stretto. Il Canale di Sicilia rimase sommerso anche durante la massima bassa marea del Pleistocene, come dimostra la presenza di mammiferi endemici, indicanti come la Sicilia si comportò come un arcipelago durante il Pleistocene (Villa 2001; Marra 2005). Inoltre, Villa (2001) mostrò come la colonizzazione dell’Italia nel Pleistocene Medio iniziò da nord, e non dalla via a sud, sulla rotta nord Africa-Sicilia. Si esclude anche la possibilità di un passaggio verso sud attraverso l’Europa centrale, percorrendo le aree alpine del nord, vista la rigidità del clima in prossimità della coltre di ghiaccio della Fennoscandia durante MIS 22, 0.87 Ma (Muttoni et al.2010).
Uno studio italiano “ridisegna” l’origine di Homo sapiens Un team di ricercatori coordinati dai genetisti della Sapienza, Fulvio Cruciani e Rosaria Scozzari, ha condotto uno studio sulla distribuzione geografica e sulla filogenesi del cromosoma Y umano, identificando varianti
genetiche che hanno permesso di ridisegnare nuovi scenari relativi all’origine e alle migrazioni di Homo sapiens (Scozzari et al.2014). Infatti, i risultati dello studio spingono a retrodatare le tappe migratorie del Sapiens e avvalorano l’ipotesi che i primi passi della nostra specie siano avvenuti in Africa centro-occidentale, e non nell’area orientale del continente, come finora ritenuto dalla comunità scientifica. Inoltre la diversità genetica del cromosoma Y osservata oggi al di fuori del continente africano, sembrerebbe essersi originata nella penisola arabica, che si viene a configurare non più solo quale corridoio di transito temporaneo di H. sapiens, ma una regione in cui la nostra specie ha sostato per molto tempo. Il lavoro si è avvalso di una tecnica innovativa (next-generation sequencing) per sequenziare a elevata profondità (50X), circa 1.5 milioni di basi del cromosoma Y umano, in 68 individui maschi non imparentati, rappresentativi delle linee di discendenza paterne più antiche e selezionati mediante uno screening di migliaia di campioni. Sono state così individuate 2.368 varianti geniche, di cui l’80% identificate per la prima volta, in questo lavoro. I dati hanno evidenziato un drastico cambiamento nelle linee genetiche (fig.4): i nodi basali risultano essere molto più antichi (intorno a 196 mila anni) di quanto finora ritenuto, e molte delle principali tappe migratorie affrontate da H. sapiens sono state retrodatate, dimostrando la sua esistenza in Africa centro-occidentale già 200 mila anni fa. Il ridotto numero di rami presenti nella filogenesi dall’origine fino a 115 mila anni fa suggerisce l’esistenza di un piccolo numero (circa 10) di “padri fondatori” africani della nostra specie; questo numero risulta essere molto minore di quello osservato nell’albero del DNA mitocondriale (che fornisce informazioni sulle linee di discendenza materne). Inoltre, solo uno di questi “padri fondatori” avrebbe dato origine a tutte le linee di discendenza maschile che si trovano oggi fuori dall’Africa. La distribuzione geografica delle varianti identificate, (fig.5), ha così riaperto un campo di ricerca in cui sembravano esserci risposte acclarate rispetto all’origine in Africa orientale di H. sapiens.

11
Fig.4-5 Maximum parsimony tree (fig.4) ottenuto da 2.386 varianti geniche. Il numero di mutazioni per ciascun ramo è mostrato al
di sopra di essi. Le date stimate sono riportate in box, affianco a ciascun nodo (in alto date ottenute con metodo Beast, in basso con
metodo rho. I colori usati rispecchiano la corrente nomenclatura (Karafet et al. 2008; Scozzari et al. 2012) usata per i maggiori
aplogruppi. In fig.5 si mostra la distribuzione degli aplogruppi MSY nel loro attuale home-range nel continente africano (Scozzari et
al.2014).
Lo studio rivoluziona anche le tempistiche dell’uscita dal continente africano della popolazione di H. sapiens che ha dato origine al popolamento del pianeta, in un intervallo tra 145-64 mila anni fa. Una delle implicazioni principali emerse è che l’occupazione della penisola arabica possa essere avvenuta più di 100 mila anni fa, e non 60 mila anni fa, e che non sia stata un’escursione temporanea, come precedentemente ipotizzato. Sembra, piuttosto, sia stato l’evento che ha dato origine, in tempi successivi, a tutta la diversità genetica del cromosoma Y osservata oggi al di fuori del continente africano. Gli scenari proposti aprono la possibilità ad un lungo gap temporale in cui la popolazione di uomini esistente nella penisola arabica sia rimasta geneticamente isolata prima di espandersi nuovamente nel resto del mondo (Scozzari et al.2014). Nuovi scenari si aprono quindi sull’evoluzione dell’uomo moderno.
La diversità fonemica: evidenza dell’origine africana dell’uomo moderno La migrazione out of Africa è consistita in una catena molto ramificata di successive migrazioni in cui tribù o porzioni di tribù (clan, bande di caccia) hanno colonizzato nuovi territori vergini aventi potenziali risorse (acqua, selvaggina, percorsi di migrazione delle mandrie, pesce, frutti, tuberi, semi) adeguate alla dimensione dei gruppi e alla loro prevedibile crescita numerica. La tribù
di cacciatori-raccoglitori era un gruppo sostanzialmente endogamico che parlava una sola lingua ed era orientativamente composto da un migliaio di individui. Abitava un territorio abbastanza vasto da consentire il sostentamento della tribù con un’economia di caccia e raccolta, ma non a sufficienza da provocare una disgregazione dell’unità linguistica o un impedimento della comunicazione sociale all’interno del gruppo. La distanza fra tribù vicine era probabilmente di diverse decine di chilometri, anche 100-200 km, per un tempo di percorrenza di 4/8 giorni di marcia; anche le spedizioni di caccia potevano richiedere spostamenti di giorni, così come i trasferimenti tra i campi d’inverno e quelli d’estate, se le condizioni climatiche lo richiedevano. Al termine della migrazione out of Africa, si ebbe l’inizio dell’era agropastorale, e la popolazione complessiva si accrebbe di un migliaio di volte (era diventata 1-15 milioni): gli esseri umani moderni si erano ormai installati in tutte le regioni del mondo. La densità di popolazione sul territorio, pur vicina ai limiti di sostenibilità, era comunque molto bassa, compresa probabilmente tra 0,013 e 0,2 abitanti/km2. La migrazione, cioè il processo di gemmazione di nuove colonie dalle tribù esistenti, ripetuto per circa 50.000 anni, portò all’esistenza di un numero di tribù stimabile in 5-10 mila. L’era agropastorale vide diffusioni demiche multiple, con forte differenziamento genetico dovuto a una selezione naturale differenziale, diversa a seconda
12
delle aree (significativo il fatto che oggi siano documentate nel mondo circa 7.000 lingue). Le fasi della migrazione si possono riassumere nei seguenti momenti. In principio, un clan di famiglie tra loro imparentate o comunque legate, costituito da diverse decine di individui che parlano la stessa lingua e condividono la stessa cultura, si separò dalla tribù e si spostò in un territorio vergine conveniente. Le favorevoli condizioni ambientali favorirono la crescita demografica di generazione in generazione fino a ad approssimarsi ai limiti di sostenibilità del territorio. Questo è il momento più probabile affinché il processo ricominci, con la separazione di una nuova colonia che va alla ricerca di un nuovo territorio. Reiterati episodi di separazione di gruppi dalle tribù di origine conducono al costituirsi di popolazioni, la cui diversità genetica è minore rispetto a quella della popolazione fondatrice: solo una porzione delle varianti geniche presenti nella popolazione originante è presente nel gruppo che si separa. Questo fenomeno di deriva genetica, l’effetto fondatore seriale, provoca una progressiva perdita di diversità genetica nelle popolazioni che via via si costituiscono, e un progressivo aumento di diversità inter-popolazioni, dovuto all’accumulo indipendente di mutazioni, non solo biologiche ma anche culturali. Le distanze geografiche e genetiche tra coppie di popolazioni sono inoltre significativamente correlate, secondo un modello d’isolamento dovuto alla distanza (Sforza 2010).
Più complesso e non facilmente quantificabile, è il quadro dell’espansione delle lingue e delle correlazioni tra diversità linguistiche e genetiche a diversa scala. Eventi demografici legati alla storia delle popolazioni hanno certamente avuto effetti sia sulla diversità genetica sia su quella linguistica. Inoltre, la creazione di barriere linguistiche può aver portato ad un rafforzamento dell’isolamento genetico tra gruppi che parlavano lingue diverse. La ricostruzione del passato evolutivo delle lingue e la loro classificazione in famiglie mediante metodi linguistici è sempre stata accompagnata da accese discussioni e controversie, e non sembra in grado di spingere la sua analisi oltre 7.000 anni fa. Il Dottor Quentin Atkinson, PhD, Senior Lecturer nel Dipartimento
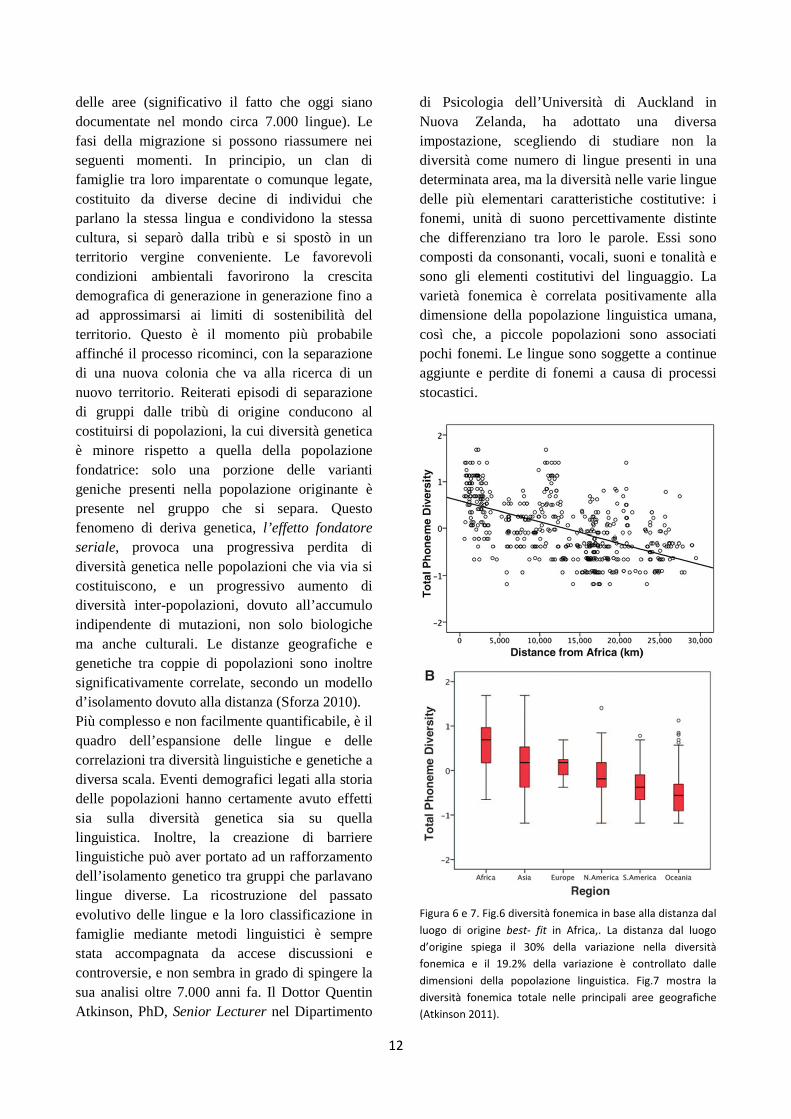
di Psicologia dell’Università di Auckland in Nuova Zelanda, ha adottato una diversa impostazione, scegliendo di studiare non la diversità come numero di lingue presenti in una determinata area, ma la diversità nelle varie lingue delle più elementari caratteristiche costitutive: i fonemi, unità di suono percettivamente distinte che differenziano tra loro le parole. Essi sono composti da consonanti, vocali, suoni e tonalità e sono gli elementi costitutivi del linguaggio. La varietà fonemica è correlata positivamente alla dimensione della popolazione linguistica umana, così che, a piccole popolazioni sono associati pochi fonemi. Le lingue sono soggette a continue aggiunte e perdite di fonemi a causa di processi stocastici.
Figura 6 e 7. Fig.6 diversità fonemica in base alla distanza dal
luogo di origine best- fit in Africa,. La distanza dal luogo
d’origine spiega il 30% della variazione nella diversità
fonemica e il 19.2% della variazione è controllato dalle
dimensioni della popolazione linguistica. Fig.7 mostra la
diversità fonemica totale nelle principali aree geografiche
(Atkinson 2011).

13
Quentin Atkinson, esperto nell’applicazione di metodi matematici alla linguistica, ha analizzato un campione di 504 lingue (il 7% delle circa 7.000 lingue totali, oggi documentate nel mondo) utilizzando il World Atlas of Language Structures (WALS). Dallo studio (Atkinson 2011) risulta che il numero di fonemi più alto si trova in lingue parlate in Africa, e decresce all’aumentare della distanza dal continente africano (fig.6 e 7). Il maggior numero di fonemi si trova, infatti, nelle lingue Khoisan, parlate dai Boscimani e dagli Ottentotti, in Africa meridionale. Esse presentano più di 100 fonemi, fino a 141 secondo il catalogo UPSID, incluse consonanti “clic” (particolari schiocchi della lingua contro il palato o i denti), consonanti stridenti e faringalizzate. Il più piccolo numero di fonemi (calcolato in una decina) si riscontra invece in lingue parlate nelle Hawaii, o in altre isole tropicali del Pacifico e nell’America meridionale: regioni situate ai margini estremi, temporali e spaziali, della migrazione dall’Africa. Per confronto, la lingua italiana presenta 30 fonemi mentre l’inglese, circa 45. Tenendo sotto controllo alcune variabili, quali la varianza di dimensioni delle popolazioni e altri fattori confondenti, Atkinson ha modellato il quadro della diversità fonemica che sarebbe potuta conseguire a diversi potenziali punti di origine dell’espansione del linguaggio. In accordo con un declino della diversità genetica e fenotipica umana all’aumentare della distanza dall’Africa, secondo l’effetto fondatore seriale, nel corso delle migrazioni umane, viene proposto un andamento analogo per lo sviluppo della cultura umana e del linguaggio. Il modello più adatto a spiegare il quadro della diversità fonemica odierna pone l’origine del linguaggio in Africa centro meridionale. Fuori dall’Africa, la maggiore diversità fonemica si riscontra in lingue sviluppate nel sud est asiatico, in accordo con una maggiore diversità genetica ivi osservata (Fig.7). Questo fatto suggerisce che le popolazioni del sud est asiatico siano cresciute molto rapidamente dopo l’uscita dall’Africa dei loro antenati. Nelle Americhe si osserva una diminuzione della diversità fonemica correlata alla distanza delle popolazioni dallo stretto di Bering, confermando l’ipotesi che i primi americani siano venuti
dall’Asia attraversando il ponte di terre emerse che univa la Siberia all’Alaska, e da lì si siano espansi fino alla punta dell’America meridionale (Atkinson 2011). Lo studio linguistico di Atkinson (2011) ben si accorda con le evidenze precedentemente esposte di un origine dell’uomo anatomicamente moderno avvenuta in Africa, ciò nonostante è ancora in discussione la circoscrizione dell’esatta area d’origine della migrazione di Homo sapiens, saranno quindi necessarie nuove evidenze per appurarla.
Bibliografia e sitografia • Albianelli, A., Napoleone, G., (2004). Magnetostratigraphy of
thw Homo-Bearing Pleistocene Dandiero Basin (Danakil
depression, Eritrea). Rivista Italiana di Paleontologia e
Stratigrafia 110, 35–44.
• Andrews, P. (1992), Evolution and environment in the
Hominoidea. Nature 360, 641±646.
• Arzarello, M., Marcolini, F., Pavia, G., Pavia, M., Petronio, C.,
Petrucci, M., Rook, L., Sardella, R., (2007). Evidence of earliest
human occurrence in Europe: the site of Pirro Nord (Southern
Italy). Naturwissenschaften 94, 107–112.
• Asfaw, B., Beyene, Y., Suwa, G., Walker, R.C., White, T.D.,
WoldeGabriel, G., Yemane, T., (1992). The earliest Acheulean
from Konso–Gardula. Nature 360, 732–735.
• Atkinson Q.D. (2011), Phonemic Diversity Supports a Serial
Founder Effect Model of Language Expansion from Africa,
Science, 346-349.
• Berger, W.H., Bickert, T., Schmidt, H., Wefer, G.,
(1993).Quaternary oxygen isotope record of pelagic
foraminifers: Site 806, Ontong Java Plateau. Proceedings ODP:
Scientific Results, vol. 130, pp. 381–395.
• Biondi G., Rickards O., (2003), Uomini per caso.Miti, fossili e
molecole nella nostra storia evolutiva, Editori Riuniti.
• Brown, W. M., George, M. Jr & Wilson, A. C., (1979). Rapid
evolution of animal mitochondrial DNA. Proc. Natl Acad. Sci.
USA 76, 1967±1971.
• Campbell MC, Tishkoff SA. (2008), Annu. Rev. Genomics Hum.
Genet;9:403.
• Cann, R. L., Stoneking, M. & Wilson, A. C. (1987),Mitochondrial
DNA and human evolution. Nature 325, 31±36.
• Caramelli D., (2009), Antropologia molecolare, manuale di
base, Firenze University Press, cap. 11 Evoluzione umana e
popolamento.
• Carbonell, E., Bermudez de Castro, J.M., Arsuaga, J.L., Diez,
J.C., Rosas, A., Cuenca-Bescos, G., Sala, R., Mosquera, M.,
Rodriguez, X.P., (1995). Lower Pleistocene hominids and
artifacts from Atapuerca-TD6 (Spain). Science 269, 826–829.
• Carbonell, E., Bermúdez de Castro, J.M., Parés, J.M., Pérez-
González, A., Cuenca-Bescós, G., Ollé, A., Mosquera, M.,
Huguet, R., van der Made, J., Rosas, A., Sala, R., Vallverdú, J.,
García, N., Granger, D.E., Martinón-Torres, M., Rodríguez, X.P.,
Stock, G.M., Vergès, J.M., Allué, E., Burjachs, F., Cáceres, I.,
Canals, A., Benito, A., Díez, C., Lozano, M.,Mateos, A., Navazo,
M., Rodríguez, J., Rosell, J., Arsuaga, J.L., (2008). The first
hominin of Europe. Nature 452, 465–470.

14
• Ethnologue. (www.ethnologue.com)
• Gibbons A., (2011), A New View Of the Birth of Homo sapiens,
Science, Vol. 331 no. 6016 pp. 392-394.
• Giles, R. E., Blanc, H., Cann, H. M. & Wallace, D. C., (1980).
Maternal inheritance of human mitochondrial DNA. Proc. Natl
Acad. Sci. USA 77, 6715±6719.
• Green R.E., Malaspinas A.S., Krause j., Briggs A.W., Johnson
P.L.f., Uhler C., Meyer M., Good J.M., Maricic T., Stenzel U.,
Prϋfer K., Siebauer M., Burbano H., Ronan M.,. Rothberg J.M.,
Egholm M., Rudan P., Brajkovic´ D. , Kucan Z., Gusiic´ I.
,Wikstrӧm M., Laakkonen L., Kelso J., Slatkin M., Pӓӓbo
S.,(2008), A Complete Neandertal Mitochondrial Genome
Sequence Determined by High-Throughput Sequencing, Cell
134, 416–426.
• Harpending H. C., Batzer M. A., Gurven M., Jorde L.B., Rogers
A. R., and Sherry S. T. (1998), Genetic traces of ancient
demography, Proceedings of the National Academy of Science,
USA, 95,1961-1967.
• Head, M.J., Gibbard, P.L., (2005). Early–Middle Pleistocene
transitions: an overview and recommendation for the defining
boundary. In: Head, M.J., Gibbard, P.L. (Eds.), Early–Middle
Pleistocene Transitions: The Land–Ocean Evidence: Special
Publications, vol. 247. Geological Society, London, pp. 1–18.
• Hedges, S. B. (2000). A start for population genomics. Nature
408:652-653.
• Henn B.M., Gignoux C.R., Jobin M., Granka M.L., Macpherson
J.M., Kidd J.M., Rodriguéz-Botigué L., Ramachandran S., Hon L.,
Brisbin A., Lin A., Underhill P., Comas D., Kidd K.K., Norman
P.J., Parham P., Bustamante C., Mountain J.L., Feldman M.W.,
(2011), Hunter-gatherer genomic diversity suggests a southern
African origin for modern humans. Proc Natl Acad Sci
USA.;108:5154-5162.
• Horai, S., Hayasaka, K., Kondo, R., Tsugane, K. & Takahata, N.,
(1995), Recent African origin of modern humans revealed by
complete sequences of hominoid mitochondrial DNAs. Proc.
Natl Acad. Sci. USA 92, 532±536.
• Ingman, M., Kaessmann, H., Pääbo, S. & Gyllensten, U. (2000),
Nature 408, 708–713.
• Kaessmann, H., Heissig, F., von Haeseler, A. & Paabo, S. (1999),
DNA sequence variation in a non-coding region of low
recombination on the human X chromosome. Nature Genet.
22, 78±81.
• Karafet TM, Mendez FL, Meilerman MB, Underhill PA, Zegura
SL, Hammer MF. (2008). New binary polymorphisms reshape
and increase resolution of the human Y chromosomal
haplogroup tree. Genome Res 18: 830–838.
• Kent, D.V., Opdyke, N.D., Ewing, M., (1971). Climate change in
the North Pacific using icerafted detritus as a climatic
indicator. Geological Society of America Bulletin 82,2741–
2754.
• Krings, M., Geisert, H., Schmitz, R.W., Krainitzki, H. & Pääbo, S.
(2000), Proc. Natl Acad. Sci. USA 96, 5581–5585.
• Kuhn, S.L., (2002). Paleolithic Archeology in Turkey.
Evolutionary Anthropology 11,198–210.
• Kumar, S. & Hedges, S. B. (1998), A molecular timescale for
vertebrate evolution. Nature 392, 917±920.
• Langereis, C.G., Krijgsman, W., Muttoni, G., Menning, M.,
(2010). Magnetostratigraphy—concepts, definitions, and
applications. Newsletter on Stratigraphy 43 (3), 207–233.
• Lewin, R. (1999), Human Evolution, Science, Malden, MA.
• Lycett, S.J., von Cramon-Taubadel, N., (2008). Acheulean
variability and hominin dispersals: a model-bound approach.
Journal of Archaeological Science 35 (3), 553–562.
• Maddison, D. R., Ruvolo, M. & Swofford, D. L. (1992).
Geographic origins of human mitochondrial DNA: phylogenetic
evidence from control region sequences. Syst. Biol. 41,
111±124.
• Manzi, G., (2004). Human evolution at the Matuyama–Brunhes
boundary. Evolutionary Anthropology 13, 11–24.
• Marra, A.C., (2005). Pleistocene mammals of Mediterranean
islands. Quaternary International 129, 5–14.
• Martinez-Navarro, B., Rook, L., Segid, A., Yosief, D., Ferreti,
M.P., Shoshani, J., Tecle, T.M., Libsekal, Y., (2004). The large
fossil mammals from Buia (Eritrea): systematics, biochronology
and paleoenvironments. Rivista Italiana di Paleontologia e
Stratigrafia110, 61–88.
• Muttoni, G., Ravazzi, C., Breda, M., Laj, C., Kissel, C., Mazaud,
A., Pini, R., Garzanti, E., (2007). Magnetostratigraphy of the
Leffe lacustrine succession (Southern Alps, Italy): evidence for
an intensification of glacial activity in the Alps at Marine
Isotope Stage 22 (0.87 Ma). Quaternary Research 67, 161–173.
• Muttoni, G., Scardia, G., Kent, D. V., (2010). Human migration
into Europe during the late Early Pleistocene climate
transition. Palaeogeography, Palaeoclimatology,
Palaeoecology; 296,79–93.
• Muttoni, G., Scardia, G., Kent, D.V., Swisher, C.C., Manzi, G.,
(2009). Pleistocene magnetochronology of early hominin sites
at Ceprano and Fontana Ranuccio, Italy. Earth and Planetary
Science Letters 286, 255–268.
• Nei, M. & Roychoudhury, A. K. (1993). Mol. Biol. Evol. 10, 927–
943.
• Olivio, P. D., Van de Walle, M. J., Laipis, P. J. & Hauswirth, W.
W., (1983), Nucleotide sequence evidence for rapid genotypic
shifts in the bovine mitochondrial DNA D-loop. Nature 306,
400
• Oms, O., Parés, J.M., Martínez-Navarro, B., Agustí, J., Toro, I.,
Martínez-Fernández, G., Turq, A., (2000). Early human
occupation of Western Europe: paleomagnetic dates for two
paleolithic sites in Spain. Proceedings of the National Academy
of Sciences 97, 10666–10670.
• Parés, J.M., Pérez-Gonzalez, A., (1995). Paleomagnetic age for
homind fossils at Atapuerca Archaeological site, Spain. Science
269, 830–832.
• Parés, J.M., Pérez-Gonzalez, A., Rosas, A., Benito, A., Bermudez
de Castro, J.M., Carbonell, E., Huguet, R., (2006). Matuyama-
age lithic tools from the Sima del Elefante site, Atapuerca
(northern Spain). Journal of Human Evolution 50, 163–169.
• Quade, J., Levin, N., Semaw, S., Stout, D., Renne, P., Rogers,
M.J., Simpson, S., (2004). Paleoenvironments of the earliest
stone toolmakers, Gona, Ethiopia. Geological Society of
America Bulletin 116, 1529–1544.
• Ramachandran S, Deshpande O., Roseman C.C., Feldman
M.W., Rosenberg N.A., Cavalli-Sforza L.L.,(2005), Support from
the relationship of genetic and geographic distance in human
populations for a serial founder effect originating in Africa,
Proc. Natl. Acad. Sci. U.S.A;102:15942.
• Reed FA, Tishkoff SA. (2006), Current Opinion in Geneics &
Development;16:597.
• Rosenberg N.A., Pritchard J.K., Weber J.L., Cann H.M., Kidd
K.K., Zhivotovsky L.A., Feldman M.W. (2002), Genetic structure
of human populations, Science;298:2381.

15
• Rosenberg N.A., Mahajan S., Gonzalez-Quevedo C., Blum M.G.,
Nino-Rosales L., Ninis V., Das P., Hegde M., Molinari L., Zapata
G., Weber J.L., Belmont J.W., Patel P. (2006), Low Levels of
Genetic Divergence across Geographically and Linguistically
Diverse Populations from India PLoS Genet;2:e215.
• Ruvolo, M., Zehr S., Von Dornum M., Pan D., Chang B., Lin J.,
(1993), Mitochondrial COII sequences and modern human
origins. Mol. Biol. Evol. 10,1115±1135.
• Sawada, D., Piburn, M., Judson, E., Turley, J., Falconer, K.,
Benford, R. & Bloom, I., (2002). Measuring reform practices in
science and mathematics classrooms: The Reformed Teaching
Observation Protocol. School Science and Mathematics,
102(6), 245-253.
• Scott, G.R., Gibert, L., Gibert, J., (2007). Magnetostratigraphy
of the Orce region (Baza Basin), SE Spain: New chronologies for
Early Pleistocene faunas and hominid occupation sites.
Quaternary Science Reviews 26, 415–435.
• Scozzari R, Massaia A, D’Atanasio E, Myres NM, Perego UA,
Trombetta B, Cruciani F. (2012). Molecular dissection of the
basal clades in the human Y chromosome phylogenetic tree.
PLoS ONE 7: e49170.
• Scozzari R, Massaia A, Trombetta B, Bellusci G, Myres NM,
Novelletto A, Cruciani F. (2014), An unbiased resource of
novel SNP markers provides a new chronology for the human
Y chromosome and reveals a deep phylogenetic structure in
Africa. Genome Research; 24:535-544.
• Semaw, S., Renne, P., Harris, J.W.K., Feibel, C.S., Bernor, R.L.,
Fesseha, N., Mowbray, K., (1997). 2.5-million-year-old stone
tools from Gona, Ethiopia. Nature 385, 333–336.
• Sforza L.L.C. L’evoluzione della cultura, (2010), Codice Edizioni,
Torino, Capitolo 12, L’espansione out of Africa, Capitolo 13,
L’effetto fondatore seriale.
• Shackleton, N.J., (1995). New data on the evolution of Pliocene
climate variability. In:Vrba, E., Denton, G.H., Partridge, T.C.,
Burckle, L.H. (Eds.), Palaeoclimate and evolution, with
emphasis on human origins. Yale University Press, New
Haven,pp. 242–248.
• Shackleton, N.J., Opdyke, N.D., (1976). Oxygen-isotope and
paleomagnetic stratigraphy of Pacific core V28-239: Late
Pliocene to latest Pleistocene. Geological Society of America
Memoir 145, 449–464.
• Stringer, C. B. & Andrews, P. (1998), Science 239, 1263–1268.
• Tamura, K. & Nei, M., (1993). Estimation of the number of
nucleotide substitutions in the control region of mitochondrial
DNA in humans and chimpanzees. Mol. Biol. Evol. 10, 512±526.
• Tishkoff, S. A., Dietzsch E., Speed W., Pakstis A.J., KiddJ.R.,
Cheung K., Bonné-Tamir B., Santachiara-Benerecetti A.S.,
Moral P., Krings M.,Pääbo S., Watson E, Risch N., Jenkins T.,
Kidd K.K. (1996). Global Patterns of Linkage Disequilibrium at
the CD4 Locus and Modern Human Origins, Science 271, 1380–
1387.
• Tishkoff S.A., Reed F.A., Friedlaender F.R., Ehret C, Ranciaro A.,
Froment A, Hirbo J.B., Awomoyi A.A., Bodo J.M., Doumbo O.,
(2009). The genetic structure and history of Africans and
African Americans. Science 324: 1035–1044.
• Torroni, A., Bandelt H.J., D’Urbano L., Lahermo P., Moral P.,
Sellitto D., Rengo C., Forster P., Savontaus M.L., Bonné-
TamirB., Scozzari R., (1998), mtDNA analysis reveals a major
late Paleolithic population expansion from southwestern to
northeastern Europe. Am. J. Hum. Genet. 62, 1137±1152.
• Vigilant, L., Stoneking, M., Harpending, H., Hawkes, K.
&Wilson, A. C. (1991), African populations and the evolution of
human mitochondrial DNA. Science 253, 1503±1507.
• Villa, P., (2001). Early Italy and the colonization of Western
Europe. Quaternary International 75, 113–130.
• Wolpoff, M. H. (1989), in The Human Revolution: Behavioral
and Biological Perspectives on the Origins of Modern Humans
(eds Mellars, P. & Stringer, C.) 62–108 Princeton Univ. Press.


















