
La biochimica della resistenza alla malaria -...
6
Una coltura continua del parassita della malaria Plasmodium falcipa- rum può essere fatta crescere in questo dispositivo. I parassiti vengono coltivati in uno strato sottile di globuli rossi umani che ricopre il fondo del tubo orizzontale ed è sommerso dal terreno nutritivo. Il contenuto di ossigeno del terreno e le altre condizioni sperimentali possono essere - . variate per mezzo dei tubi verticali. Le cellule vengono rimosse per l'analisi al microscopio attraverso il tubo corto al centro. Fino a che non furono stabilite le condizioni per la coltura del parassita, nel 1976, le ricerche sulla malaria provocata dal P. falciparum, la forma più letale, dovevano venire eseguite con il sangue di volontari umani o di primati. Il destino dei parassiti della malaria viene controllato in strisci di globuli rossi su vetrini per microscopia. La colorazione (Giemsa) evidenzia i nuclei dei parassiti in viola scuro e il loro citoplasma in colore blu. La velocità di riproduzione viene misurata muliante conteggio dei numeri di parassiti per 100 globuli rossi. Questo striscio mostra i parassiti in vari stadi del loro ciclo vitale (si veda l'illustrazione in basso a pagina 85). ' evoluzione è causata dalla selezione naturale, che opera sulla variabi- lità genetica prodotta dalla muta- zione e dalla ricombinazione dei geni. I geni mutanti che conferiscono qualche vantaggio selettivo tendono a diventare più frequenti, mentre i mutanti svantag- giosi tendono a venire eliminati. Nelle popolazioni umane esistono pochi casi chiari di selezione a favore o contro geni specifici in risposta a forze di pressione selettiva. Gli esempi migliori sono costi- tuiti dalle malattie ereditarie: la selezione agisce contro i geni che provocano queste malattie, e agisce tanto più efficacemente quanto più le malattie sono gravi. Per questo motivo, le malattie eredita- rie mortali dovrebbero essere molto rare. Tuttavia alcune anomalie ereditarie dei globuli rossi, in particolare la talassemia e l'anemia falciforme, in alcune popolazio- ni sono presenti con frequenze sorpren- dentemente alte. Questa è forse una pro- va contro la teoria della selezione natura- le? Al contrario, i geni per l'anemia falci- forme e la talassemia dimostrano l'effica- cia della selezione come causa dell'evolu- zione. Gli stessi geni mutanti che provo- cano l'anomalia mortale dei globuli rossi negli individui omozigoti (che posseggo- no due geni anormali, uno da ciascuno dei genitori) proteggono gli individui etero- zigoti (che posseggono un gene anormale e uno normale) dagli effetti mortali della malaria, una malattia provocata da un parassita che infetta i globuli rossi. La protezione contro la malaria conserva le alte frequenze di questi geni per altro verso dannosi. L'efficacia della malaria come forza selettiva deriva dal potente effetto di que- sta malattia parassitaria sulla salute e sul- la capacità riproduttiva delle popolazioni umane. La malaria ha rappresentato sto- ricamente una delle cause più frequenti di morte. In Africa la malaria è tuttora en- demica: non colpisce la popolazione a ondate come le epidemie, ma piuttosto rappresenta una presenza costante che contribuisce a tassi di mortalità infantile che raggiungono il 50 per cento. Uccide circa il 10 per cento delle sue vittime di- rettamente e contribuisce alla morte delle altre diminuendo la capacità del sistema immunitario di combattere le altre infe- zioni. A causa della malaria un numero significativo di individui non vive abba- stanza a lungo da riprodursi: qualsiasi mutazione genetica che renda resistenti alla malaria deve perciò avere un forte vantaggio selettivo. Fu la coincidenza della distribuzione geografica dell'anemia falciforme con quella della malaria a suggerire la possibi- lità che il gene dell'anemia falciforme conferisse qualche tipo di resistenza. La dimostrazione clinica fu più difficile da ottenere, ma nel 1954 Anthony C. Alli- son dell'Università di Oxford dimostrò che i bambini eterozigoti per il gene del- l'anemia falciforme venivano colpiti mol- to più raramente dalla forma più grave di malaria rispetto ai bambini che non pos- sedevano questo gene. Tuttavia, poiché il parassita che induce la malaria non pote- va venire mantenuto in colture di labora- torio, non fu possibile dimostrare la resi- stenza a livello cellulare, né fu possibile stabilire il suo meccanismo biochimico. Recentemente noi abbiamo sfruttato un sistema di coltura in vitro di nuova conce- zione che ci ha permesso di studiare in che modo il gene dell'anemia falciforme e alcuni altri geni mutanti che alterano la funzionalità dei globuli rossi conferiscono la resistenza alla malaria. T globuli rossi, l'ambiente in cui il paras- -1 sita della malaria viene in contatto con le alterazioni funzionali codificate da questi geni, sono costituiti in gran parte di emoglobina, la proteina che cattura l'os- sigeno nei polmoni e lo cede ai tessuti. Le altre proteine del citoplasma dei globuli rossi sono enzimi metabolici. Alcuni di essi catalizzano la glicolisi, il processo in cui tramite passaggi successivi il glucosio viene degradato a lattato con formazione di due molecole di adenosintrifosfato (ATP). Altri catalizzano le reazioni di quella che è chiamata la via degli esosi monofosfati, che mantiene i coenzimi nicotinammide adenindinucleotide fosfa- to (NADP) e glutatione nella loro forma ridotta. L'ATP è il trasportatore univer- sale di energia per la cellula; il NADP ri- dotto (NADPH) e il glutatione ridotto servono a prevenire e a riparare i danni provocati dall'ossidazione. La membrana cellulare racchiude la cellula e ne control- la la forma e la deformabilità. Inoltre con- trolla i movimenti degli ioni verso l'inter- no e l'esterno della cellula; in particolare mantiene, consumando ATP, un'alta concentrazione interna di potassio contro la tendenza verso un equilibrio con l'am- biente esterno povero di potassio. Sulla faccia esterna della membrana i glicolipi- di e le glicoproteine caratterizzano in modo distintivo la superficie cellulare. É a livello della membrana cellulare che il parassita della malaria interagisce inizialmente con la cellula. Il parassita è un piccolo protozoo unicellulare del ge- nere Plasmodium, quattro specie del qua- le inducono la malaria nell'uomo; la for- ma più letale di malaria, che causa un milione di morti all'anno tra i bambini africani, è provocata dalla specie Plasmo- dium falciparum. Una forma specializzata del parassita viene iniettata nel sangue da una zanzara del genere Anopheles e migra nel fegato, dove si sviluppa e si divide a formare i merozoiti, la forma che infetta i globuli rossi. I merozoiti rientrano nel si- stema circolatorio e riconoscono e si le- gano alle membrane dei globuli rossi. A questo punto si attiva un meccanismo, tuttora non ben conosciuto, per cui il merozoite attraversa la membrana cellu- lare, che gli si richiude intorno. Così rac- chiuso in un vacuolo il parassita cresce, digerendo l'emoglobina per ottenere gli amminoacidi che gli servono per produrre le proprie proteine, e presumibilmente utilizzando il glucosio, l'ATP e i coenzimi ridotti del globulo rosso per il proprio La biochimica della resistenza alla malaria I geni di due malattie letali, l'anemia falciforme e la talassemia, sono favoriti dall'evoluzione perché proteggono contro la malaria. Il meccanismo di questa protezione ora può essere studiato in laboratorio di Milton J. Friedman e William Trager 82 83
Transcript of La biochimica della resistenza alla malaria -...
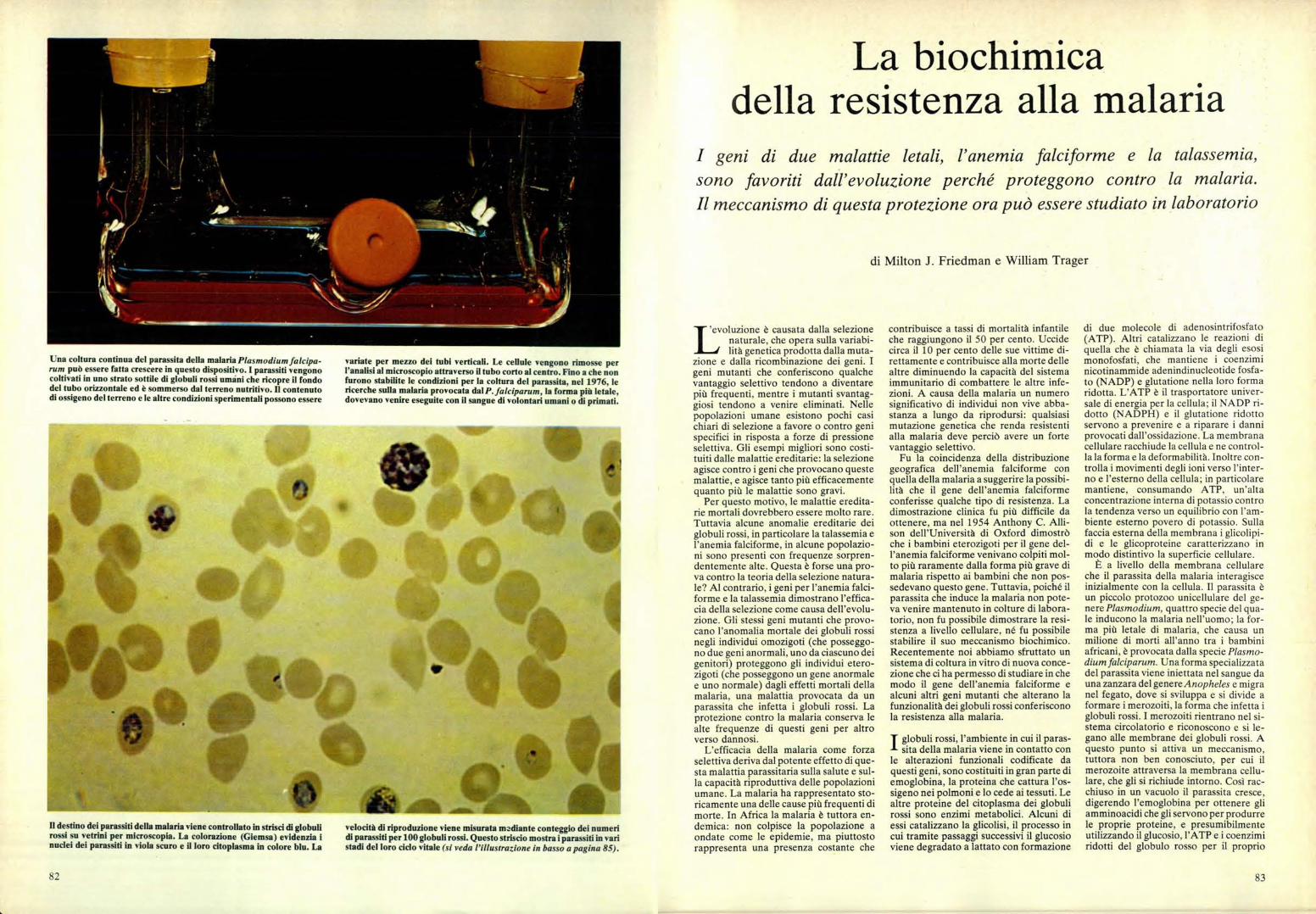
Una coltura continua del parassita della malaria Plasmodium falcipa-rum può essere fatta crescere in questo dispositivo. I parassiti vengonocoltivati in uno strato sottile di globuli rossi umani che ricopre il fondodel tubo orizzontale ed è sommerso dal terreno nutritivo. Il contenutodi ossigeno del terreno e le altre condizioni sperimentali possono essere
- .
variate per mezzo dei tubi verticali. Le cellule vengono rimosse perl'analisi al microscopio attraverso il tubo corto al centro. Fino a che nonfurono stabilite le condizioni per la coltura del parassita, nel 1976, lericerche sulla malaria provocata dal P. falciparum, la forma più letale,dovevano venire eseguite con il sangue di volontari umani o di primati.
Il destino dei parassiti della malaria viene controllato in strisci di globulirossi su vetrini per microscopia. La colorazione (Giemsa) evidenzia inuclei dei parassiti in viola scuro e il loro citoplasma in colore blu. La
velocità di riproduzione viene misurata muliante conteggio dei numeridi parassiti per 100 globuli rossi. Questo striscio mostra i parassiti in varistadi del loro ciclo vitale (si veda l'illustrazione in basso a pagina 85).
' evoluzione è causata dalla selezionenaturale, che opera sulla variabi-lità genetica prodotta dalla muta-
zione e dalla ricombinazione dei geni. Igeni mutanti che conferiscono qualchevantaggio selettivo tendono a diventarepiù frequenti, mentre i mutanti svantag-giosi tendono a venire eliminati. Nellepopolazioni umane esistono pochi casichiari di selezione a favore o contro genispecifici in risposta a forze di pressioneselettiva. Gli esempi migliori sono costi-tuiti dalle malattie ereditarie: la selezioneagisce contro i geni che provocano questemalattie, e agisce tanto più efficacementequanto più le malattie sono gravi.
Per questo motivo, le malattie eredita-rie mortali dovrebbero essere molto rare.Tuttavia alcune anomalie ereditarie deiglobuli rossi, in particolare la talassemia el'anemia falciforme, in alcune popolazio-ni sono presenti con frequenze sorpren-dentemente alte. Questa è forse una pro-va contro la teoria della selezione natura-le? Al contrario, i geni per l'anemia falci-forme e la talassemia dimostrano l'effica-cia della selezione come causa dell'evolu-zione. Gli stessi geni mutanti che provo-cano l'anomalia mortale dei globuli rossinegli individui omozigoti (che posseggo-no due geni anormali, uno da ciascuno deigenitori) proteggono gli individui etero-zigoti (che posseggono un gene anormalee uno normale) dagli effetti mortali dellamalaria, una malattia provocata da unparassita che infetta i globuli rossi. Laprotezione contro la malaria conserva lealte frequenze di questi geni per altroverso dannosi.
L'efficacia della malaria come forzaselettiva deriva dal potente effetto di que-sta malattia parassitaria sulla salute e sul-la capacità riproduttiva delle popolazioniumane. La malaria ha rappresentato sto-ricamente una delle cause più frequenti dimorte. In Africa la malaria è tuttora en-demica: non colpisce la popolazione aondate come le epidemie, ma piuttostorappresenta una presenza costante che
contribuisce a tassi di mortalità infantileche raggiungono il 50 per cento. Uccidecirca il 10 per cento delle sue vittime di-rettamente e contribuisce alla morte dellealtre diminuendo la capacità del sistemaimmunitario di combattere le altre infe-zioni. A causa della malaria un numerosignificativo di individui non vive abba-stanza a lungo da riprodursi: qualsiasimutazione genetica che renda resistentialla malaria deve perciò avere un fortevantaggio selettivo.
Fu la coincidenza della distribuzionegeografica dell'anemia falciforme conquella della malaria a suggerire la possibi-lità che il gene dell'anemia falciformeconferisse qualche tipo di resistenza. Ladimostrazione clinica fu più difficile daottenere, ma nel 1954 Anthony C. Alli-son dell'Università di Oxford dimostròche i bambini eterozigoti per il gene del-l'anemia falciforme venivano colpiti mol-to più raramente dalla forma più grave dimalaria rispetto ai bambini che non pos-sedevano questo gene. Tuttavia, poiché ilparassita che induce la malaria non pote-va venire mantenuto in colture di labora-torio, non fu possibile dimostrare la resi-stenza a livello cellulare, né fu possibilestabilire il suo meccanismo biochimico.Recentemente noi abbiamo sfruttato unsistema di coltura in vitro di nuova conce-zione che ci ha permesso di studiare in chemodo il gene dell'anemia falciforme ealcuni altri geni mutanti che alterano lafunzionalità dei globuli rossi conferisconola resistenza alla malaria.
T globuli rossi, l'ambiente in cui il paras--1 sita della malaria viene in contatto conle alterazioni funzionali codificate daquesti geni, sono costituiti in gran parte diemoglobina, la proteina che cattura l'os-sigeno nei polmoni e lo cede ai tessuti. Lealtre proteine del citoplasma dei globulirossi sono enzimi metabolici. Alcuni diessi catalizzano la glicolisi, il processo incui tramite passaggi successivi il glucosioviene degradato a lattato con formazione
di due molecole di adenosintrifosfato(ATP). Altri catalizzano le reazioni diquella che è chiamata la via degli esosimonofosfati, che mantiene i coenziminicotinammide adenindinucleotide fosfa-to (NADP) e glutatione nella loro formaridotta. L'ATP è il trasportatore univer-sale di energia per la cellula; il NADP ri-dotto (NADPH) e il glutatione ridottoservono a prevenire e a riparare i danniprovocati dall'ossidazione. La membranacellulare racchiude la cellula e ne control-la la forma e la deformabilità. Inoltre con-trolla i movimenti degli ioni verso l'inter-no e l'esterno della cellula; in particolaremantiene, consumando ATP, un'altaconcentrazione interna di potassio controla tendenza verso un equilibrio con l'am-biente esterno povero di potassio. Sullafaccia esterna della membrana i glicolipi-di e le glicoproteine caratterizzano inmodo distintivo la superficie cellulare.
É a livello della membrana cellulareche il parassita della malaria interagisceinizialmente con la cellula. Il parassita èun piccolo protozoo unicellulare del ge-nere Plasmodium, quattro specie del qua-le inducono la malaria nell'uomo; la for-ma più letale di malaria, che causa unmilione di morti all'anno tra i bambiniafricani, è provocata dalla specie Plasmo-dium falciparum. Una forma specializzatadel parassita viene iniettata nel sangue dauna zanzara del genere Anopheles e migranel fegato, dove si sviluppa e si divide aformare i merozoiti, la forma che infetta iglobuli rossi. I merozoiti rientrano nel si-stema circolatorio e riconoscono e si le-gano alle membrane dei globuli rossi. Aquesto punto si attiva un meccanismo,tuttora non ben conosciuto, per cui ilmerozoite attraversa la membrana cellu-lare, che gli si richiude intorno. Così rac-chiuso in un vacuolo il parassita cresce,digerendo l'emoglobina per ottenere gliamminoacidi che gli servono per produrrele proprie proteine, e presumibilmenteutilizzando il glucosio, l'ATP e i coenzimiridotti del globulo rosso per il proprio
La biochimicadella resistenza alla malaria
I geni di due malattie letali, l'anemia falciforme e la talassemia,sono favoriti dall'evoluzione perché proteggono contro la malaria.Il meccanismo di questa protezione ora può essere studiato in laboratorio
di Milton J. Friedman e William Trager
82 83

O O0,0O oo
4
-195 GRADICENTIGRADI
6®----TROFOZOITE
®C)® ®
e ®
PICCOLO PARASSITA-o-A• Q
MEROZOITI
2
3
5 AGGIUNTADI GLICEROLO
ESORBITOLO
8
0 5°2,
,
MALARIA
GENE PER L'ANEMIA FALCIFORME
Z GENE PER LA BETA-TALASSEMIA
Le distribuzioni dei geni per l'anemia falciforme e per la beta-talasse-mia coincidono con l'area dove la malaria provocata da P. falciparumera endemica prima del 1930 (in colore). Questa coincidenza geografi-
ca ha fatto pensare che la resistenza alla malaria potesse essere ilvantaggio evolutivo che tendeva a mantenere alta la frequenza dei geniresponsabili di malattie letali del sangue in certe popolazioni umane.
Il ciclo vitale del P. falciparum nei globuli rossi inizia con l'invasione diuna cellula da parte di un merozoite (1). Il parassita incorpora unagocciolina di citoplasma, così che in sezione appare come un anellosottile (2). La forma ad anello si nutre e cresce fino a diventare untrofozoite (3); sulla superficie del globulo rosso si formano sporgenzeche ne provocano l'adesione alle pareti dei vasi capillari. Lì il nucleo del
parassita si divide ripetutamente (4); ogni nucleo figlio acquista un po-co di citoplasma (5); il parassita si divide e dà origine a nuovi merozoiti,da 12 a 24, che provocano la rottura del globulo rosso ospite e dannoinizio a un nuovo ciclo (6). Alcuni trofozoiti si differenziano in gameto-citi maschili e femminili (7); questi gametociti vengono ingeriti dallezanzare e danno in tal modo inizio alla fase sessuale del ciclo vitale.
7
GAMETOCITA FEMMINILE GAMETOCITA MASCHILE
TROFOZOITE
GLOBULOROSSO1
2
COLTURA SINCRONA
Una coltura di P. falciparum si inizia inoculando una sospensione fresca di globuli rossi umani conglobuli rossi infetti prelevati da un paziente o da una coltura di riferimento (1). Le cellule vengonomantenute a temperatura corporea (su una piastra di terreno solido, come è mostrato in questodisegno, o nel tubo a U raffigurato a pagina 82) come uno strato sottile ricoperto di liquido nutritivo(2). La coltura può venire saggiata per misurare la moltiplicazione dei parassiti. Dopo tre o quattrogiorni le cellule contengono mescolanza di parassiti piccoli e grandi (3), che possono venire congelatie conservati per usi futuri (4), inoculati in nuove colture (5) o incubati in una soluzione di gelatinaper separare i trofozoiti (6); questi grossi parassiti possono venire coltivati da soli per produrremerozoiti (7) o mescolati con globuli rossi freschi per ottenere una coltura sincrona di parassiti (8).
metabolismo. Dopo un periodo di cresci-ta il nucleo del parassita si divide numero-se volte; in seguito ogni nucleo viene cir-condato da membrane insieme a parte delcitoplasma. In questo modo si formano da12 a 24 nuovi merozoiti, che provocano larottura del globulo rosso e invadono altrecellule.
La febbre periodica caratteristica dellamalaria è dovuta alla liberazione sincronadei merozoiti e delle tossine prodotte dalparassita in tutto il corpo. Nel caso di P.falciparum questa liberazione sincrona siverifica ogni 48 ore, il periodo del ciclo disviluppo dei parassiti nei globuli rossi. Lafebbre e la debilitazione che l'accompa-gna sono i sintomi principali della mala-ria. Nella malaria da P. falciparum, tutta-via, vi è un altro effetto più letale. Il glo-bulo rosso infetto sviluppa delle sporgen-ze sulla sua superficie che ne provocanol'adesione alle pareti dei vasi capillari,dove rimane fino alla maturazione delparassita. Quando un gran numero diglobuli rossi intasa in questo modo unorgano vitale come il cervello, può averluogo la morte del soggetto. L'indicazio-ne più chiara dell'effetto protettivo delgene per l'anemia falciforme è che pochiportatori del gene muoiono a causa dellecomplicazioni cerebrali nella malaria daP. falciparum.
T inus Pauling e i suoi colleghi riuscirono a definire una malattia molecolareper la prima volta quando dimostraronoche nell'anemia falciforme la molecola diemoglobina è alterata, e che gli individuiche ne sono affetti posseggono solo laforma alterata della molecola, chiamataemoglobina S, mentre alcuni membri del-le loro famiglie posseggono circa metàemoglobina S e metà emoglobina norma-le A. Studi sulle famiglie, pubblicati circanello stesso periodo, confermarono i ri-sultati di Pauling, dimostrando che l'ere-ditarietà dell'anemia falciforme potevaessere ascritta a un solo gene recessivo: lamalattia compare cioè solo nei membridella famiglia che sono omozigoti perquel gene.
L'emoglobina adulta è costituita da duecatene alfa e due catene beta; Vernon M.Ingram dell'Università di Cambridgedimostrò ben presto che nell'emoglobinaS solo la catena beta è anormale. L'ano-malia riguarda uno solo degli amminoaci-di che costituiscono la catena: un acidoglutammico sostituisce una valina. Gliindividui omozigoti per il gene dell'ane-mia falciforme (designati SS) possiedonosolo l'emoglobina S perché hanno en-trambi i geni per la catena beta mutati; gliindividui eterozigoti (AS), che sono por-tatori del carattere per l'anemia falcifor-me, hanno un solo gene mutato, e circa il40 per cento delle loro emoglobine sonodi tipo S. Se due portatori del carattereper l'anemia falciforme hanno quattro fi-gli, statisticamente uno su quattro saràaffetto da anemia falciforme, uno saràcompletamente normale e due sarannoportatori del carattere, in accordo con iprincipi della segregazione mendeliana.
I sintomi dell'anemia falciforme com-
84
85

INGRANDIMENTO DELLAMEMBRANA CELLULARE
C
1.3-DIFOSFO-GLICERATO
GLUTATIONERIDOTTO
NADP
NADPHGLUTATIONE
2.3-DIFOSFO-GLICERATO
LATTATO OSSIEMOGLOBINA DESOSSIEMOGLOBINA
GLUCOSIO
GLUCOSIO-6-FOSFATO
POTASSIO
i
>
SODI°
ADP
ATP
e
5ANEMIA
FALCIFORMEiPORTATOREDELL'ANEMIAFALCIFORME
Oujz
z O
LLJ
O Rj<cc
ccE 5
Un globulo rosso, la cellula più semplice dell'organismo, non è altro che citoplasma racchiuso dauna membrana. I glicolipidi e le glicoproteine sulla sua superficie (a) determinano le interazionidella cellula con il suo ambiente e con il parassita della malaria. Le proteine della membranacontrollano il trasporto di sostanze verso l'interno e l'esterno della cellula; in particolare, il po-tassio viene pompato verso l'interno e il sodio verso l'esterno (b). Questa reazione richiedeenergia, che viene fornita dall'ATP generato durante la glicolisi, il processo con cui il glucosioviene degradato a lattaio (c). La via degli esosi monofosfati (d) produce NADP ridotto e gluta-tione ridotto, i quali prevengono e riparano il danno ossidativo a livello della membrana cellulare.Il citoplasma è riempito per la maggior parte con la proteina emoglobina (e), che trasporta l'os-sigeno dai polmoni ai tessuti per mezzo di un processo che si basa su cambiamenti strutturali dellequattro catene di amminoacidi che la compongono e che è controllato in parte dal difosfoglicerato.
MADRE
c5' dPORTATORE
DELL'ANEMIA
NORMALEFALCIFORME
EMO-GLOBINA S
EMO-GLOBINA A
Questo diagramma rappresenta la trasmissione dei geni per l'emoglobina anormale S da parte digenitori entrambi eterozigoti per il gene anormale. Statisticamente, uno dei figli sarà omozigoteper il gene anormale (SS), cioè eredita due geni per l'emoglobina S ed è affetto da anemiafalciforme. Due figli erediteranno un gene per l'emoglobina S e un gene per l'emoglobina normaleA; essi sono eterozigoti (AS) come i loro genitori e sono «portatori» del carattere per l'anemiafalciforme. Un figlio sarà normale (AA). Le emoglobine vengono identificate per mezzo dell'elet-troforesi (in basso). I campioni su un supporto di acetato di cellulosa sono sottoposti a unacorrente elettrica. L'emoglobina S differisce dalla A per un solo amminoacido, che manca di unacarica negativa, e così l'emoglobina S migra verso il polo positivo meno rapidamente della A.
paiono quando i globuli rossi SS perdonoossigeno mentre circolano attraverso i tes-suti. Quando l'emoglobinaS è deossigena-ta, tende ad aggregarsi in lunghe fibresottili, che modificano la forma normale adisco dei globuli rossi in forme angolose,compresa la caratteristica forma a falce chedà il nome alla malattia. In una crisi dianemia falciforme alcune cellule deforma-te bloccano la circolazione locale del san-gue e impediscono l'afflusso di ossigeno.Quando la concentrazione di ossigeno di-minuisce, altri globuli rossi si deformano el'area di circolazione danneggiata si esten-de, causando intensi danni ai tessuti. Inmancanza di cure mediche sofisticate lasopravvivenza degli omozigoti per l'emo-globina S è molto bassa. Gli individuiportatori del carattere, invece, normal-mente non presentano alcun disturbo. Iloro globuli rossi AS contengono abba-stanza emoglobina normale e si defor-mano solo in condizioni estreme, come aelevate altitudini.
Ouattro anni fa, dopo mezzo secolo ditentativi di far crescere il parassita
della malaria in colture di laboratorio,uno di noi (Trager) fu in grado di definirele condizioni che permettono il manteni-mento di P. falciparum in colture conti-nue di globuli rossi umani in un mezzoartificiale simile a sangue. Questo sistemadi coltura viene attualmente utilizzato daalcuni ricercatori per sviluppare vaccinisperimentali contenenti materiale prove-nienti da vari stadi del ciclo cellulare delparassita, che vengono sperimentati suglianimali . Altri ricercatori stanno tentandodi purificare e analizzare gli agenti bio-chimici che hanno effetti particolari nelcorso di un'infezione. Il sistema ci ha an-che permesso di studiare nei dettagli leinterazioni del parassita della malaria coni globuli rossi mutanti ospiti.
Per capire come l'emoglobina dell'a-nemia falciforme protegge un portatoreeterozigote contro la malaria abbiamocoltivato il parassita della malaria in glo-buli rossi prelevati da donatori normali,da individui portatori del carattere perl'anemia falciforme e da pazienti affettida anemia falciforme. La coltivazioneavveniva nelle nostre condizioni stan-dard, in un'atmosfera con una concentra-zione di ossigeno del 17 per cento, checrea nel mezzo di coltura una tensione diossigeno simile a quella dei polmoni; l'e-moglobina era perciò completamente os-sigenata e i globuli rossi non si deformava-no. In queste condizioni i parassiti cresce-vano ugualmente bene in tutti e tre i tipi diglobuli rossi. Questo dimostra che non vi èalcuna alterazione sostanziale del metabo-lismo nei globuli rossi mutati e che la emo-globina S, come l'emoglobina A, può es-sere digerita dal plasmodio.
Per verificare l'effetto della deformazionesulla crescita del parassita aggiungemmoun piccolo numero di globuli rossi infetta-ti a colture di globuli rossi AS e SS in unaatmosfera di ossigeno al 17 per cento.Durante le 48 ore successive, il periodo diun ciclo di crescita, tutti i parassiti abban-donarono i globuli rossi normali in cui
PADRE
C51
86
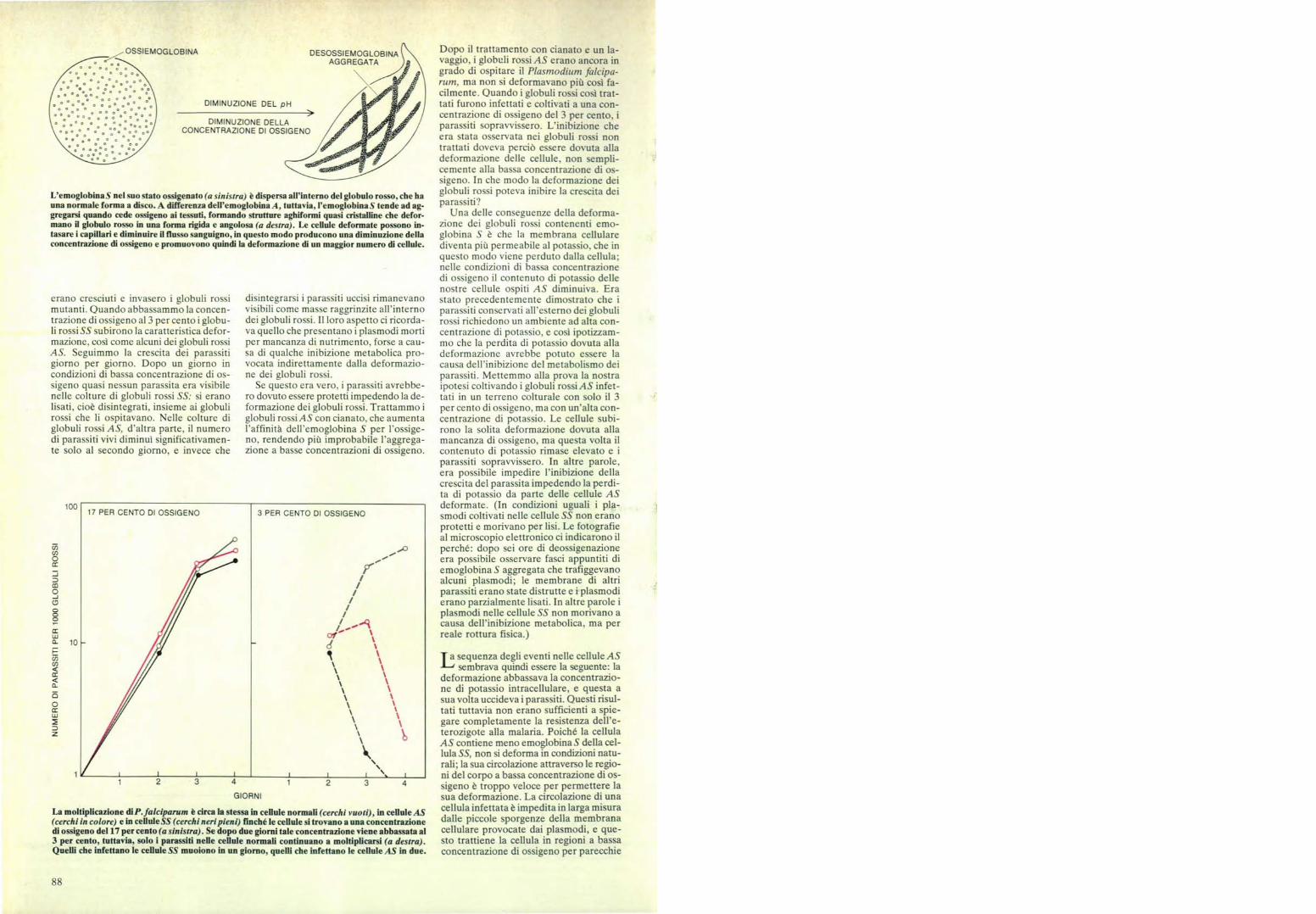
OSSIEMOGLOBINA
o ° 0 o 0° .° o . oo °. o o o .° 030
0 0 0 0 °o. ° 0 oo o o
o 00 0 o o o o 0°° °. o oo o o o . o °o 00.0
o o 00 0 ° o ° ° o o
K:: 0 o ...°0°0o o o oo 0 0. oo o
o
DESOSSIEMOGLOBINAAGGREGATA
DIMINUZIONE DEL pH
DIMINUZIONE DELLACONCENTRAZIONE DI OSSIGENO
L'emoglobina 5 nel suo stato ossigenato (a sinistra) è dispersa all'interno del globulo rosso, che hauna normale forma a disco. A differenza dell'emoglobina A, tuttavia, l'emoglobina S tende ad ag-gregarsi quando cede ossigeno ai tessuti, formando strutture aghiformi quasi cristalline che defor-mano il globulo rosso in una forma rigida e angolosa (a destra). Le cellule deformate possono in-tasare i capillari e diminuire il flusso sanguigno, in questo modo producono una diminuzione dellaconcentrazione di ossigeno e promuovono quindi la deformazione di un maggior numero di cellule.
17 PER CENTO DI OSSIGENO 3 PER CENTO DI OSSIGENO
/ofci
•
2
3
4
2
3
4
GIORNI
La moltiplicazione di P. falciparum è circa la stessa in cellule normali (cerchi vuoti), in cellule AS(cerchi in colore) e in cellule SS (cerchi neri pieni) finché le cellule si trovano a una concentrazionedi ossigeno del 17 per cento (a sinistra). Se dopo due giorni tale concentrazione viene abbassata al3 per cento, tuttavia, solo i parassiti nelle cellule normali continuano a moltiplicarsi (a destra).Quelli che infettano le cellule SS muoiono in un giorno, quelli che infettano le cellule AS in due.
100
10
Dopo il trattamento con cianato e un la-vaggio, i globuli rossi AS erano ancora ingrado di ospitare il Plasmodium falcipa-rum, ma non si deformavano più così fa-cilmente. Quando i globuli rossi così trat-tati furono infettati e coltivati a una con-centrazione di ossigeno del 3 per cento, iparassiti sopravvissero. L'inibizione cheera stata osservata nei globuli rossi nontrattati doveva perciò essere dovuta alladeformazione delle cellule, non sempli-cemente alla bassa concentrazione di os-sigeno. In che modo la deformazione deiglobuli rossi poteva inibire la crescita deiparassiti?
Una delle conseguenze della deforma-zione dei globuli rossi contenenti emo-globina S è che la membrana cellularediventa più permeabile al potassio, che inquesto modo viene perduto dalla cellula;nelle condizioni di bassa concentrazionedi ossigeno il contenuto di potassio dellenostre cellule ospiti AS diminuiva. Erastato precedentemente dimostrato che iparassiti conservati all'esterno dei globulirossi richiedono un ambiente ad alta con-centrazione di potassio, e così ipotizzam-mo che la perdita di potassio dovuta alladeformazione avrebbe potuto essere lacausa dell'inibizione del metabolismo deiparassiti. Mettemmo alla prova la nostraipotesi coltivando i globuli rossi AS infet-tati in un terreno colturale con solo il 3per cento di ossigeno, ma con un'alta con-centrazione di potassio. Le cellule subi-rono la solita deformazione dovuta allamancanza di ossigeno, ma questa volta ilcontenuto di potassio rimase elevato e iparassiti sopravvissero. In altre parole,era possibile impedire l'inibizione dellacrescita del parassita impedendo la perdi-ta di potassio da parte delle cellule ASdeformate. (In condizioni uguali i pla-smodi coltivati nelle cellule SS non eranoprotetti e morivano per lisi. Le fotografieal microscopio elettronico ci indicarono ilperché: dopo sei ore di deossigenazioneera possibile osservare fasci appuntiti diemoglobina S aggregata che trafiggevanoalcuni plasmodi; le membrane di altriparassiti erano state distrutte e i plasmodierano parzialmente lisati. In altre parole iplasmodi nelle cellule SS non morivano acausa dell'inibizione metabolica, ma perreale rottura fisica.)
T a sequenza degli eventi nelle cellule AS1--d sembrava quindi essere la seguente: ladeformazione abbassava la concentrazio-ne di potassio intracellulare, e questa asua volta uccideva i parassiti. Questi risul-tati tuttavia non erano sufficienti a spie-gare completamente la resistenza dell'e-terozigote alla malaria. Poiché la cellulaAS contiene meno emoglobina S della cel-lula SS, non si deforma in condizioni natu-rali; la sua circolazione attraverso le regio-ni del corpo a bassa concentrazione di os-sigeno è troppo veloce per permettere lasua deformazione. La circolazione di unacellula infettata è impedita in larga misuradalle piccole sporgenze della membranacellulare provocate dai plasmodi, e que-sto trattiene la cellula in regioni a bassaconcentrazione di ossigeno per parecchie
erano cresciuti e invasero i globuli rossimutanti. Quando abbassammo la concen-trazione di ossigeno al 3 per cento i globu-li rossi SS subirono la caratteristica defor-mazione, così come alcuni dei globuli rossiAS. Seguimmo la crescita dei parassitigiorno per giorno. Dopo un giorno incondizioni di bassa concentrazione di os-sigeno quasi nessun parassita era visibilenelle colture di globuli rossi SS: si eranolisati, cioè disintegrati, insieme ai globulirossi che li ospitavano. Nelle colture diglobuli rossi AS, d'altra parte, il numerodi parassiti vivi diminuì significativamen-te solo al secondo giorno, e invece che
disinte grarsi i parassiti uccisi rimanevanovisibili come masse raggrinzite all'internodei globuli rossi. Il loro aspetto ci ricorda-va quello che presentano i plasmodi mortiper mancanza di nutrimento, forse a cau-sa di qualche inibizione metabolica pro-vocata indirettamente dalla deformazio-ne dei globuli rossi.
Se questo era vero, i parassiti avrebbe-ro dovuto essere protetti impedendo la de-formazione dei globuli rossi. Trattammo iglobuli rossi AS con cianato, che aumental'affinità dell'emoglobina S per l'ossige-no, rendendo più improbabile l'aggrega-zione a basse concentrazioni di ossigeno.
88
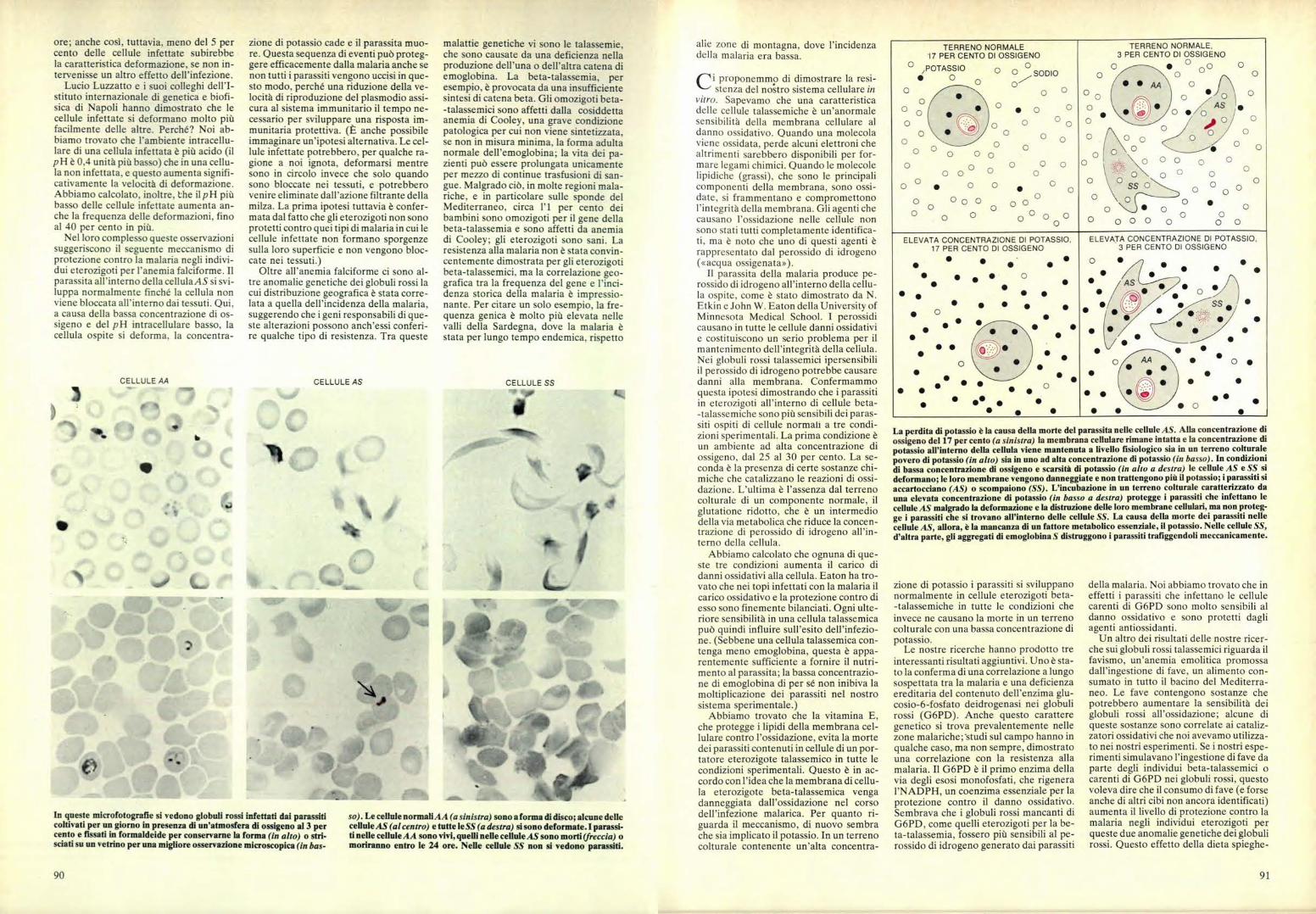
TERRENO NORMALE17 PER CENTO DI OSSIGENO
O OPOTASSIO•1
OO O SODIO
O0
• O °O
O 0
oo• O
o o
oo 00 o o
oo oo o o
o o o o o00 o o
oo • o o •o• ooo o o o o oo o o
o o n oo o - o -
TERRENO NORMALE,3 PER CENTO DI OSSIGENO
OO • O O
OO O O
• • AA OO o • i - O0O / AS •
o •o • / o o
di.o o o
o oo----o o
o 00 o o
o o o oo o O osS o\ oo oo o o •
o • o , ooo 000 o - o 0
ELEVATA CONCENTRAZIONE DI POTASSIO17 PER CENTO DI OSSIGENO
• • • • ••• • • O•• •••
e•
O• •
• • e • ••
• • • • • c ---,
• •......
•
• ••
O((,._3u.)'-
•• / • I• • 7/ )• -__________.--- ••• • • 0• • • •
• •• • • •• • •
ELEVATA CONCENTRAZIONE DI POTASSIO,3 PER CENTO DI OSSIGENO
O • • •• • ------ .
• • •• / AS --", • /i)• ,i • ,--'
0/ SS• I • •
0/
••e / • • •
• •
• AA
•
••
• • • O
••• •
•
• • •• O
• • •
La perdita di potassio è la causa della morte del parassita nelle cellule AS. Alla concentrazione diossigeno del 17 per cento (a sinistra) la membrana cellulare rimane intatta e la concentrazione dipotassio all'interno della cellula viene mantenuta a livello fisiologico sia in un terreno colturalepovero di potassio (in alto) sia in uno ad alta concentrazione di potassio (in basso). In condizionidi bassa concentrazione di ossigeno e scarsità di potassio (in alto a destra) le cellule AS e SS sideformano; le loro membrane vengono danneggiate e non trattengono più il potassio; i parassiti siaccartocciano (AS) o scompaiono (SS). L'incubazione in un terreno colturale caratterizzato dauna elevata concentrazione di potassio (in basso a destra) protegge i parassiti che infettano lecellule AS malgrado la deformazione e la distruzione delle loro membrane cellulari, ma non proteg-ge i parassiti che si trovano all'interno delle cellule SS. La causa della morte dei parassiti nellecellule AS, allora, è la mancanza di un fattore metabolico essenziale, il potassio. Nelle cellule SS,d'altra parte, gli aggregati di emoglobina S distruggono i parassiti trafiggendoli meccanicamente.
CELLULE AA CELLULE AS CELLULE SS
In queste microfotografie si vedono globuli rossi infettati dai parassiticoltivati per un giorno in presenza di un'atmosfera di ossigeno al 3 percento e fissati in formaldeide per conservarne la forma (in alto) o stri-sciati su un vetrino per una migliore osservazione microscopica (in bas-
so). Le cellule normaliAA (a sinistra) sono a forma di disco; alcune dellecellule AS (al centro) e tutte le SS (a destra) si sono deformate. I parassi-ti nelle cellule AA sono vivi, quelli nelle celluleAS sono morti (freccia) omoriranno entro le 24 ore. Nelle cellule SS non si vedono parassiti.
ore; anche così. tuttavia. meno del 5 percento delle cellule infettate subirebbela caratteristica deformazione, se non in-tervenisse un altro effetto dell'infezione.
Lucio Luzzatto e i suoi colleghi dell'I-stituto internazionale di genetica e biofi-sica di Napoli hanno dimostrato che lecellule infettate si deformano molto piùfacilmente delle altre. Perché? Noi ab-biamo trovato che l'ambiente intracellu-lare di una cellula infettata è più acido (ilpH è 0,4 unità più basso) che in una cellu-la non infettata, e questo aumenta signifi-cativamente la velocità di deformazione.Abbiamo calcolato, inoltre, the il p H piùbasso delle cellule infettate aumenta an-che la frequenza delle deformazioni, finoal 40 per cento in più.
Nel loro complesso queste osservazionisuggeriscono il seguente meccanismo diprotezione contro la malaria negli indivi-dui eterozigoti per l'anemia falciforme. Ilparassita all'interno della cellulaAS si svi-luppa normalmente finché la cellula nonviene bloccata all'interno dai tessuti. Qui,a causa della bassa concentrazione di os-sigeno e del pH intracellulare basso, lacellula ospite si deforma, la concentra-
zione di potassio cade e il parassita muo-re. Questa sequenza di eventi può proteg-gere efficacemente dalla malaria anche senon tutti i parassiti vengono uccisi in que-sto modo, perché una riduzione della ve-locità di riproduzione del plasmodio assi-cura al sistema immunitario il tempo ne-cessario per sviluppare una risposta im-munitaria protettiva. (E anche possibileimmaginare un'ipotesi alternativa. Le cel-lule infettate potrebbero, per qualche ra-gione a noi ignota, deformarsi mentresono in circolo invece che solo quandosono bloccate nei tessuti, e potrebberovenire eliminate dall'azione filtrante dellamilza. La prima ipotesi tuttavia è confer-mata dal fatto che gli eterozigoti non sonoprotetti contro quei tipi di malaria in cui lecellule infettate non formano sporgenzesulla loro superficie e non vengono bloc-cate nei tessuti.)
Oltre all'anemia falciforme ci sono al-tre anomalie genetiche dei globuli rossi lacui distribuzione geografica è stata corre-lata a quella dell'incidenza della malaria.suggerendo che i geni responsabili di que-ste alterazioni possono anch'essi conferi-re qualche tipo di resistenza. Tra queste
malattie genetiche vi sono le talassemie,che sono causate da una deficienza nellaproduzione dell'una o dell'altra catena diemoglobina. La beta-talassemia, peresempio. è provocata da una insufficientesintesi di catena beta. Gli omozigoti beta--talasse miei sono affetti dalla cosiddettaanemia di Cooley, una grave condizionepatologica per cui non viene sintetizzata,se non in misura minima, la forma adultanormale dell'emoglobina; la vita dei pa-zienti può essere prolungata unicamenteper mezzo di continue trasfusioni di san-gue. Malgrado ciò, in molte regioni mala-riche, e in particolare sulle sponde delMediterraneo, circa l'l per cento deibambini sono omozigoti per il gene dellabeta-talassemia e sono affetti da anemiadi Cooley; gli eterozigoti sono sani. Laresistenza alla malaria non è stata convin-centemente dimostrata per gli eterozigotibeta-talassemici, ma la correlazione geo-grafica tra la frequenza del gene e l'inci-denza storica della malaria è impressio-nante. Per citare un solo esempio, la fre-quenza genica è molto più elevata nellevalli della Sardegna, dove la malaria èstata per lungo tempo endemica, rispetto
alle zone di montagna, dove l'incidenzadella malaria era bassa.
Ci proponemmo di dimostrare la resi-stenza del nostro sistema cellulare in
vitro. Sapevamo che una caratteristicadelle cellule talassemiche è un'anormalesensibilità della membrana cellulare aldanno ossidativo. Quando una molecolaviene ossidata, perde alcuni elettroni chealtrimenti sarebbero disponibili per for-mare legami chimici. Quando le molecolelipidiche (grassi), che sono le principalicomponenti della membrana, sono ossi-date, si frammentano e compromettonol'integrità della membrana. Gli agenti checausano l'ossidazione nelle cellule nonsono stati tutti completamente identifica-ti, ma è noto che uno di questi agenti èrappresentato dal perossido di idrogeno(«acqua ossigenata»).
Il parassita della malaria produce pe-rossido di idrogeno all'interno della cellu-la ospite, come è stato dimostrato da N.Etkin e John W. Eaton della University ofMinnesota Medica! School. I perossidicausano in tutte le cellule danni ossidativie costituiscono un serio problema per ilmantenimento dell'integrità della cellula.Nei globuli rossi talassemici ipersensibiliil perossido di idrogeno potrebbe causaredanni alla membrana. Confermammoquesta ipotesi dimostrando che i parassitiin eterozigoti all'interno di cellule beta--talassemiche sono più sensibili dei paras-siti ospiti di cellule normali a tre condi-zioni sperimentali. La prima condizione èun ambiente ad alta concentrazione diossigeno, dal 25 al 30 per cento. La se-conda è la presenza di certe sostanze chi-miche che catalizzano le reazioni di ossi-dazione. L'ultima è l'assenza dal terrenocolturale di un componente normale, ilglutatione ridotto, che è un intermediodella via metabolica che riduce la concen-trazione di perossido di idrogeno all'in-terno della cellula.
Abbiamo calcolato che ognuna di que-ste tre condizioni aumenta il carico didanni ossidativi alla cellula. Eaton ha tro-vato che nei topi infettati con la malaria ilcarico ossidativo e la protezione contro diesso sono finemente bilanciati. Ogni ulte-riore sensibilità in una cellula talassemicapuò quindi influire sull'esito dell'infezio-ne. (Sebbene una cellula talassemica con-tenga meno emoglobina, questa è appa-rentemente sufficiente a fornire il nutri-mento al parassita; la bassa concentrazio-ne di emoglobina di per sé non inibiva lamoltiplicazione dei parassiti nel nostrosistema sperimentale.)
Abbiamo trovato che la vitamina E,che protegge i lipidi della membrana cel-lulare contro l'ossidazione, evita la mortedei parassiti contenuti in cellule di un por-tatore eterozigote talassemico in tutte lecondizioni sperimentali. Questo è in ac-cordo con l'idea che la membrana di cellu-la eterozigote beta-talassemica vengadanneggiata dall'ossidazione nel corsodell'infezione malarica. Per quanto ri-guarda il meccanismo, di nuovo sembrache sia implicato il potassio. In un terrenocolturale contenente un'alta concentra-
zione di potassio i parassiti si sviluppanonormalmente in cellule eterozigoti beta--talassemiche in tutte le condizioni cheinvece ne causano la morte in un terrenocolturale con una bassa concentrazione dipotassio.
Le nostre ricerche hanno prodotto treinteressanti risultati aggiuntivi. Uno è sta-to la conferma di una correlazione a lungosospettata tra la malaria e una deficienzaereditaria del contenuto dell'enzima glu-cosio-6-fosfato deidrogenasi nei globulirossi (G6PD). Anche questo caratteregenetico si trova prevalentemente nellezone malariche ; 'studi sul campo hanno inqualche caso, ma non sempre, dimostratouna correlazione con la resistenza allamalaria. Il G6PD è il primo enzima dellavia degli esosi monofosfati, che rigeneral'NADPH, un coenzima essenziale per laprotezione contro il danno ossidativo.Sembrava che i globuli rossi mancanti diG6PD, come quelli eterozigoti per la be-ta-talassemia, fossero più sensibili al pe-rossido di idrogeno generato dai parassiti
della malaria. Noi abbiamo trovato che ineffetti i parassiti che infettano le cellulecarenti di G6PD sono molto sensibili aldanno ossidativo e sono protetti dagliagenti antiossidanti.
Un altro dei risultati delle nostre ricer-che sui globuli rossi talassemici riguarda ilfavismo, un'anemia emolitica promossadall'ingestione di fave, un alimento con-sumato in tutto il bacino del Mediterra-neo. Le fave contengono sostanze chepotrebbero aumentare la sensibilità deiglobuli rossi all'ossidazione; alcune diqueste sostanze sono correlate ai cataliz-zatori ossidativi che noi avevamo utilizza-to nei nostri esperimenti. Se i nostri espe-rimenti simulavano l'ingestione di fave daparte degli individui beta-talassemici ocarenti di G6PD nei globuli rossi, questovoleva dire che il consumo di fave (e forseanche di altri cibi non ancora identificati)aumenta il livello di protezione contro lamalaria negli individui eterozigoti perqueste due anomalie genetiche dei globulirossi. Questo effetto della dieta spieghe-
90
91

La sequenza di eventi qui raffigurata potrebbe proteggere gli eterozigoti AS contro la malaria.Una cellula infettata dal parassita è caratterizzata dalla presenza di sporgenze sulla sua superficie eda un pH intracellulare basso. Una cellula non infettata AS trascorrerà un breve periodo di deos-sigenazione in un capillare senza deformarsi (1); una cellula infettata, d'altra parte, rimarrà bloc-cata abbastanza a lungo da permettere alla bassa ossigenazione e al pH basso di provocare la de-formazione (2). La deformazione causa la perdita di potassio attraverso la membrana cellulare (3).Privato di potassio, il parassita muore (4). La morte di una frazione dei parassiti può concedereall'organismo dell'eterozigote il tempo sufficiente per lo sviluppo di una risposta immunitaria.
rebbe anche i risultati inconcludenti deglistudi sulla resistenza alla malaria di questie terozigoti.
Il terzo risultato accessorio riguarda ineonati. Durante i primi mesi di vita i
neonati sono quasi completamente pro-tetti contro la malaria. Geoffrey Pasvol,R. J. M. Wilson e D. J. Weatherall del-l'Università di Oxford hanno recente-mente dimostrato che l'emoglobina fetale
(che consiste di catene alfa e gamma, per-siste per un certo tempo dopo la nascita esi trova anche nei globuli rossi di alcuniadulti) può contribuire a questa protezio-ne; anche i globuli rossi di individui adultierano resistenti all'infezione da parte deiplasmodi se contenevano emoglobinafetale. Abbiamo dimostrato che i parassitiche si trovano all'interno dei globuli rossiche contengono emoglobina fetale sonomolto sensibili al danno ossidativo, comequelli che infettano i globuli rossi talas-semici. In ogni caso è improbabile che lamalaria sia stata una forza selettiva nel-l'evoluzione dell'emoglobina fetale. Laproteina fetale ha una più alta affinità perl'ossigeno dell'emoglobina adulta, e ciòmigliora il rifornimento di ossigeno al fetoin sviluppo: questa è stata probabilmentela forza selettiva che ne ha determinatol'evoluzione. La sensibilità agli ossidantidei globuli rossi fetali e la conseguenteresistenza alla malaria sono probabilmen-te effetti secondari di questo adattamentoalle condizioni dello sviluppo del feto.
Per riassumere, l'evoluzione ha prodot-to la selezione di globuli rossi mutanti
che funzionano sufficientemente bene incondizioni normali, ma si danneggianoquando sono infettati dal Plasmodiumfalciparum, e questo danno uccide il pa-rassita. In altre parole, questi globuli rossisono marginalmente vitali e l'infezione lirende non vitali e incapaci di ospitare ilparassita intracellulare. L'alterazione ge-netica nel caso delle cellule AS è una ten-denza appena contenuta a deformarsi,che è stimolata dalla presenza del parassi-ta. L'alterazione dei globuli rossi eterozi-goti talassemici consiste in un aumentodella sensibilità dei lipidi della membranacellulare nei confronti del perossido di.idrogeno generato dal parassita. In en-trambi i casi l'effeto finale è la perdita dipotassio che inibisce il metabolismo delparassita.
La storia evolutiva di un gene può esse-re molto complessa. Quando una nuovamutazione casuale si manifesta, non èpossibile prevedere il suo effetto o il suopotenziale evolutivo. Sicuramente non sisarebbe potuto prevedere che certi geniavrebbero causato malattie letali negliomozigoti e protetto gli eterozigoti dallamalaria. Il contrasto tra le due opposteforze selettive della malaria da una partee dell'anemia falciforme e della beta-ta-lassemia dall'altra parte ha prodotto unpolimorfismo bilanciato: una situazionein cui il vantaggio dell'eterozigote combi-nato con lo svantaggio dell'omozigotemantiene il gene mutante a un livello difrequenza nella popolazione basso maconsistente.
I meccanismi della vita e della morte sifondano sulla chimica e sulla biologiamolecolare; possono essere riprodotti ecompresi, almeno in parte, in laboratorio.I loro effetti finali sulla specie umana nonsi decidono, tuttavia, né in laboratorio nénelle singole cellule degli esseri umanima, lentamente e imprevedibilmente, nel-la lotta evolutiva per la vita combattutasulla scala dei continenti e dei millenni.
92















![la malaria in italia [Sola lettura] - Scuola Leonardo€¦ · La malaria in Italia meccanismi di trasmissione della malattia è statofondamentaleperl’Europael’Italia. • Un programma](https://static.fdocumenti.com/doc/165x107/604f506483935a5c5477eb8f/la-malaria-in-italia-sola-lettura-scuola-leonardo-la-malaria-in-italia-meccanismi.jpg)


