IMMUNITà ASPECIFICA - Axada Catania – Per … · Web viewCellule del kupffer nel fegato Cellule...
21
IMMUNITA’ ASPECIFICA I meccanismi dell’immunità naturale sono preesistenti al contatto con i microrganismi, e vengono rapidamente messi in moto dai microbi prima che si sviluppi una risposta immunitaria specifica verso di essi. Questo tipo di meccanismo immunitario è il sistema di difesa antimicrobica filogeneticamente più antico, e si ritrova in tutti gli organismi pluricellulari, compresi gli insetti e le piante, mentre l’immunità specifica compare per la prima volta nei vertebrati, e si sovrappone all’immunità innata al fine di potenziare le difese dell’ospite nei confronti dei patogeni esterni. L’immunità innata è la prima risposta dell’ospite in grado di prevenire le infezioni, ed in molti casi riesce ad eliminare i microbi. I meccanismi effettori dell’immunità innata sono spesso utilizzati per eliminare i microrganismi anche nell’ambito delle risposte specifiche. I linfociti B per esempio producono anticorpi, che per eliminare i microbi utilizzano due meccanismi dell’immunità naturale, i fagociti ed il sistema del complemento. L’immunità innata stimola le risposte specifiche e può influenzarne la natura, ottimizzando l’eliminazione dei diversi agenti patogeni. Alcune componenti del compartimento dell’immunità innata sono sempre vigili, anche prima che si
-
Upload
truonghanh -
Category
Documents
-
view
214 -
download
0
Transcript of IMMUNITà ASPECIFICA - Axada Catania – Per … · Web viewCellule del kupffer nel fegato Cellule...

IMMUNITA’ ASPECIFICA
I meccanismi dell’immunità naturale sono preesistenti al contatto con i microrganismi, e vengono rapidamente messi in moto dai microbi prima che si sviluppi una risposta immunitaria specifica verso di essi. Questo tipo di meccanismo immunitario è il sistema di difesa antimicrobica filogeneticamente più antico, e si ritrova in tutti gli organismi pluricellulari, compresi gli insetti e le piante, mentre l’immunità specifica compare per la prima volta nei vertebrati, e si sovrappone all’immunità innata al fine di potenziare le difese dell’ospite nei confronti dei patogeni esterni.L’immunità innata è la prima risposta dell’ospite in grado di prevenire le infezioni, ed in molti casi riesce ad eliminare i microbi.I meccanismi effettori dell’immunità innata sono spesso utilizzati per eliminare i microrganismi anche nell’ambito delle risposte specifiche. I linfociti B per esempio producono anticorpi, che per eliminare i microbi utilizzano due meccanismi dell’immunità naturale, i fagociti ed il sistema del complemento.L’immunità innata stimola le risposte specifiche e può influenzarne la natura, ottimizzando l’eliminazione dei diversi agenti patogeni.
Alcune componenti del compartimento dell’immunità innata sono sempre vigili, anche prima che si verifichi una infezione:come lo sono le barriere fisico-chimiche frapposte dalla cute e dalle superfici mucose. Altre componenti, quali i fagociti ed il sistema del complemento, sono invece inattive in condizioni normali, e si risvegliano prontamente in presenza di microrganismi.
L’immunità aspecifica comprende quattro tipi di barriere difensive:anatomica, fisiologica,endocitica/fagocitica ed infiammatoria.

Barriere anatomicheLe barriere fisiche ed anatomiche che mirano a prevenire l’ingresso dei patogeni rappresentano la prima linea di difesa di un organismo nei confronti delle infezioni. La cute e la superficie delle mucose fanno parte di questa categoria in quanto rappresentano un’efficace barriera all’ingresso della maggior parte dei microrganismi. La cute è costituita da due strati: l’epidermide ed il derma. L’epidermide è formata da vari strati sovrapposti di cellule epiteliali ravvicinate. Lo strato più esterno è formato da cellule morte ed è ricco di una proteina impermeabile detta cheratina. Il derma è costituito da connettivo e contiene vasi sanguigni , follicoli piliferi , ghiandole sebacee e ghiandole sudoripare. Le ghiandole sebacee sono associate ai follicoli piliferi e producono una secrezione oleosa detta sebo. Il sebo è costituito da acido lattico ed acidi grassi che mantengono il pH della cute tra 3 e 5; questo pH inibisce la crescita della maggior parte di microrganismi. La cute intatta non solo previene l’ingresso della maggior parte dei patogeni, ma inibisce la crescita della maggior parte dei batteri a causa del suo basso pH. Soluzioni di continuo della cute, anche piccole, dovute a ferite o abrasioni rappresentano ovvie vie di infezione.La congiuntiva e gli apparati digerente, respiratorio e urogenitale non sono rivestiti da cute asciutta e protettiva , ma da membrane mucose costituite da uno strato esterno epiteliale e dal sottostante tessuto connettivo. Sebbene la maggior parte dei patogeni entrino nell’organismo legandosi alle mucose e superandole,svariati meccanismi di difesa non specifici cercano di impedire questo ingresso. Ad esempio la saliva, le lacrime e le secrezioni mucose hanno la funzione di lavare via potenziali invasori ed inoltre contengono sostanze ad azione antibatterica ed antivirale. Il muco, secreto da cellule epiteliali della mucosa intrappola i microrganismi estranei. Nel basso tratto respiratorio e nell’apparato gastrointestinale la mucosa è ricoperta da ciglia che si muovono in modo sincrono, al fine di espellere

i microrganismi intrappolati nel muco. Inoltre alcuni organismi non patogeni tendono a colonizzare le superfici mucose. Questa flora normale compete con i patogeni per i siti di attacco sulla superficie delle cellule epiteliali e per i nutrienti necessari alla loro sopravvivenza. L’adesione dei batteri alle mucose è dovuta all’interazione di protrusioni simili a peli sul batterio, dette fimbrie o pili, con certe glicoproteine o glicolipidi espressi solo dalle cellule epiteliali di certe mucose.
Barriere FisiolologicheLe barriere fisiologiche comprendono la temperatura, il pH, e vari fattori solubili. L’ acidità gastrica è una barriera fisiologica innata alle infezioni, in quanto ben pochi microrganismi ,una volta ingeriti, possono sopravvivere al basso pH dello stomaco.Molti fattori solubili contribuiscono all’ immunità non specifica. Tra questi si possono ricordare le proteine solubili lisozima, interferone e complemento. Il lisozima, un enzima idrolitico che si trova nelle secrezioni mucose e nelle lacrime, è capace di scindere lo strato peptidoglicanico della parete batterica. L’ interferone comprende una serie di proteine prodotte da cellule infettate da virus. Tra le loro funzioni vi è la capacità di legarsi alle cellule vicine e indurre uno stato di resistenza antivirale generalizzata.

Il complemento comprende un gruppo di più di venti proteine plasmatiche che circolano in forma inattiva ed una serie di proteine di membrana con funzione recettoriale. Vari meccanismi immunologici specifici e non specifici possono convertire le forme inattive delle proteine complementari in forme attive capaci di danneggiare le membrane degli organismi patogeni portando alla loro distruzione o facilitando la loro eliminazione. Tali meccanismi di attivazione fanno capo alle cosiddette via classica e via alternativa. Entrambe comportano l’attivazione della componente centrale C3 dalla quale dipende il realizzarsi delle diverse funzioni del sistema. La via classica è innescata in genere dal dominio funzionale di IgG e IgM legate ad antigeni presenti sulla superficie di vari microrganismi capaci di fissare il complemento. La via alternativa è attivata da molteplici sostanze quali aggreganti di IgA, le endotossine, i componenti di polisaccaridi di alcune capsule batteriche.I recettori di membrana riconoscono frammenti antigenici originati dai componenti plasmatici ed, interagendo, danno luogo ad attivazione cellulare.I componenti del complemento attivati contribuiscono ad una grande varietà di meccanismi difensivi: lisi cellulare (mediante l’inserimento delle componenti C5-C9 nello strato

bilipidico, che porta all’alterazione delle funzioni della membrana cellulare), attivazione della fagocitosi, aumento della permeabilità vascolare, produzione di mediatori per la chemiotassi.
Barriere endocitiche e fagociticheUn altro importante meccanismo innato di difesa è l’ingestione di particelle extracellulari mediante la fagocitosi. La fagocitosi è un tipo di endocitosi, che è il termine generale per indicare la captazione di materiale dall’ambiente da parte di una cellula. La fagocitosi è l’ingestione di particelle tra cui anche microrganismi patogeni . Nella fagocitosi la membrana plasmatica si espande intorno alla particella estranea e forma grandi vescicole dette fagosomi. La fagocitosi è per lo più operata da cellule specializzate, come i monociti ed i neutrofili del sangue ed i macrofagi dei tessuti. La maggioranza dei tipi cellulari è invece capace di altre forme di endocitosi, quali l’endocitosi mediata dal recettore, in cui molecole extracellulari sono internalizzate dopo che si sono legate a specifici recettori cellulari, e la pinocitosi, attraverso la quale le cellule internalizzano dal terreno circostante materiale liquido e le molecole che esso contiene.
Barriere infiammatorieIl danno tissutale causato da una ferita o dall’invasione di un microrganismo patogeno induce una complessa sequenza di eventi noti nel loro insieme come risposta infiammatoria. I cinque segni cardinali dell’infiammazione sono:rubor,tumor,calor,dolor e functio laesa.
BARRIERE EPITELIALI: CUTE E MUCOSE

Le superfici epiteliali, se intatte, rappresentano una barriera fisica tra microbi e tessuti dell’ospite. Le principali superfici in cui l’ospite si interfaccia con l’ambiente esterno sono la cute, le mucose dell’apparato gastrointestinale e quelle dell’albero respiratorio: tutte e tre queste superfici sono protette da uno strato epiteliale continuo che previene l’ingresso dei microbi, e la perdita della sua integrità, predispone solitamente all’insorgenza di infezioni.Gli epiteli producono peptidi dotati di attività antibiotica naturale. Tra essi , meglio conosciute sono le cosiddette defensine, peptidi ricchi di cisteina e presenti nella cute di molte specie animali. Sono presenti in grande quantità nei granuli dei neutrofili, e rappresentano circa il 5% delle proteine in essi contenute. L’epitelio del tratto gastroenterico secerne peptidi dotati di potente attività antimicrobica, chiamati criptocidine, capaci di sterilizzare localmente il lume intestinale, ad esempio in corrispondenza delle cripte della parete. Gli epiteli producono inoltre numerose citochine attive nell’immunità innata, ad esempio i cheratinociti dell’epidermide producono IL1 e molte altre citochine.
CELLULE DELL’ IMMUNITA’ NATURALE
Cellule mononucleateIl sistema delle cellule mononucleate fagocitiche è costituito dai monociti circolanti del sangue e dai macrofagi tessutali Nel corso dell’ emopoiesi midollare i progenitori granulocito-macrofagici si differenziano in promonociti che lasciano il midollo ed entrano nel sangue, dove si differenziano ulteriormente divenendo monociti maturi.

Questi rimangono in circolo per circa otto ore, nel corso delle quali aumentano di volume, successivamente migrano nei tessuti e si trasformano nei diversi tipi di macrofagi tessutali.Il differenziamento del monocita in macrofago comporta vari cambiamenti nella cellula:il volume aumenta di 5-10 volte, gli organelli citoplasmatici aumentano di numero e di complessità, aumenta l’attività fagocitaria e la produzione di enzimi litici ed inizia la secrezione di vari fattori solubili.I macrofagi sono distribuiti in tutti i compartimenti dell’ organismo:alcuni, detti macrofagi stabili, sono residenti stabilmente in particolari tessuti, altri invece conservano la capacità di migrare e sono detti macrofagi liberi o nomadi. I macrofagi liberi si muovono nei tessuti con movimenti ameboidi, mentre i macrofagi stabili esercitano funzioni specializzate nei diversi tessuti e sono chiamati in modo diverso a seconda della sede in cui risiedono:
° Macrofagi alveolari nel polmone° Istiociti nei tessuti connettivi° Cellule del kupffer nel fegato° Cellule mesangiali nel rene° Cellule microgliali nel cervello° Osteoclasti nel tessuto osseo

I macrofagi sono normalmente in stato di quiescenza, ma nel corso della risposta immunitaria, possono essere attivati da vari stimoli.
La fagocitosi di un antigene particellare funziona da stimolo attivatorio iniziale, ma lo stato di attivazione può essere ulteriormente potenziato da citochine secrete dai linfociti T, dai mediatori dell’ infiammazione, e da prodotti della parete cellulare batterica. Uno degli attivatori più potenti dei macrofagi è l’interferone gamma (INFγ), secreto dai linfociti T attivati.

Rispetto ai macrofagi quiescenti, i macrofagi attivati hanno una maggiore attività fagocitica e microbicida, secernono una maggior quantità di mediatori dell’infiammazione ed hanno una maggiore capacità di attivare i linfociti T; inoltre secernono varie proteine con azione citotossica che contribuiscono all’eliminazione di un ampio spettro di patogeni, incluse cellule infettate da virus, cellule neoplastiche, e batteri intracellulari. Si può pertanto concludere che i macrofagi e i linfociti T interagiscono tra di loro in modo molto stretto durante la risposta immunitaria, facilitando l’uno l’azione dell’ altro.
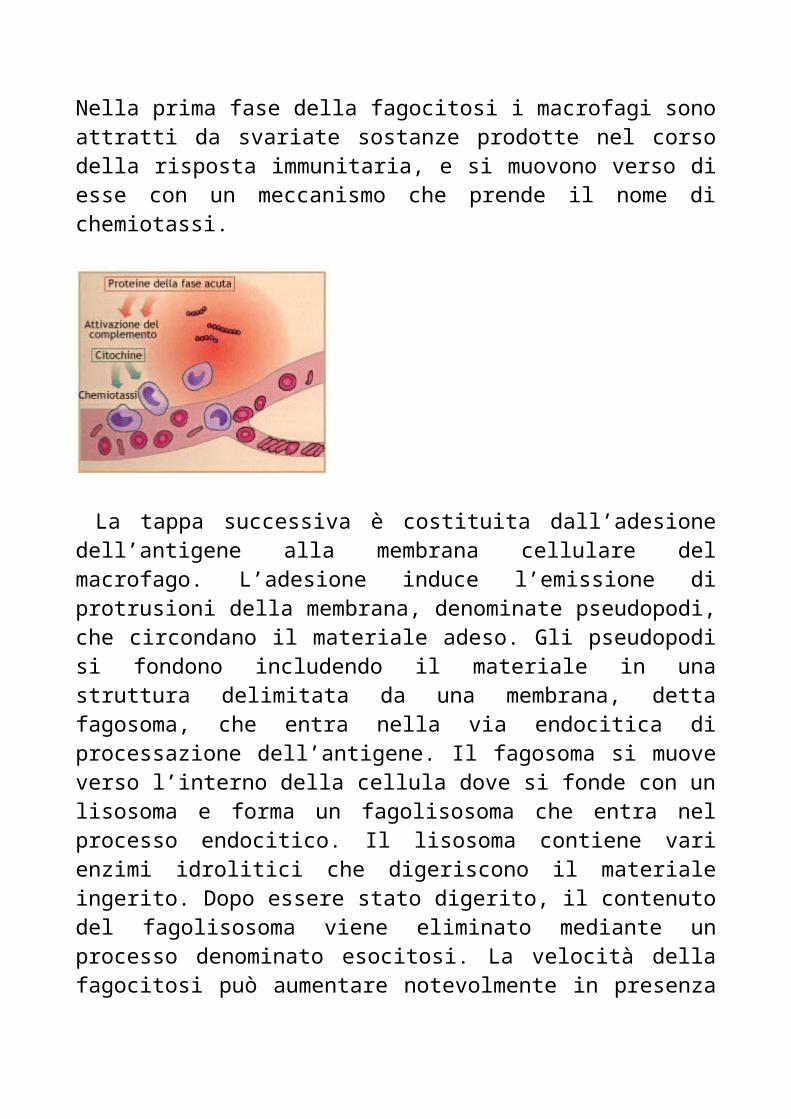
FagocitosiNella prima fase della fagocitosi i macrofagi sono attratti da svariate sostanze prodotte nel corso della risposta immunitaria, e si muovono verso di esse con un meccanismo che prende il nome di chemiotassi.

La tappa successiva è costituita dall’adesione dell’antigene alla membrana cellulare del macrofago. L’adesione induce l’emissione di protrusioni della membrana, denominate pseudopodi, che circondano il materiale adeso. Gli pseudopodi si fondono includendo il materiale in una struttura delimitata da una membrana, detta fagosoma, che entra nella via endocitica di processazione dell’antigene. Il fagosoma si muove verso l’interno della cellula dove si fonde con un lisosoma e forma un fagolisosoma che entra nel processo endocitico. Il lisosoma contiene vari enzimi idrolitici che digeriscono il materiale ingerito. Dopo essere stato digerito, il contenuto del fagolisosoma viene eliminato mediante un processo denominato esocitosi. La velocità della fagocitosi può aumentare notevolmente in presenza di opsonine, molecole che si legano sia all’antigene che al macrofago.
Attività antimicrobica e citotossicaI macrofagi attivati producono varie sostanze con attività antimicrobica e citotossica, responsabili della distruzione intracellulare dei microrganismi fagocitati.Meccanismi di uccisione ossigeno-dipendenti:I fagociti attivati producono numerosi metaboliti intermedi reattivi dell’ossigeno, e metaboliti intermedi reattivi dell’azoto, dotati di potente azione microbicida. Durante la fagocitosi nei macrofagi si verifica un processo noto come burst respiratorio, che produce l’attivazione di una ossidasi di membrana che catalizza la reazione di riduzione
dell’ossigeno ad anione superossido, un prodotto intermedio reattivo dell’ossigeno estremamente tossico per i microrganismi fagocitati.L’anione superossido genera altri potenti agenti ossidanti, tra i quali si ricordano i radicali idrossilici, l’ossigeno singoletto ed il perossido di idrogeno. Quando il lisosoma si fonde con il fagosoma, la mieloperossidasi agisce sul perossido di idrogeno e produce agenti ossidanti più durevoli ed estremamente tossici come l’ipoclorito. I macrofagi attivati con il lipopolisaccaride della parete cellulare batterica in presenza di una citochina prodotta dai linfociti T, INF γ, vengono indotti ad esprimere elevati livelli di ossido nitrico sintetasi (NOS), che ossida la L- arginina formando citrullina e un radicale reattivo, l’ossido nitrico.Meccanismi d’azione indipendenti dall’ossigeno:I macrofagi attivati contengono il lisozima ed enzimi idrolitici la cui azione degradativa non coinvolge l’ossigeno. Essi contengono inoltre un gruppo di peptidi con azione antimicrobica e citotossica detti defensine (vedi sopra).Patogeni resistenti:Alcuni microrganismi riescono a sopravvivere ed a moltiplicarsi all’interno del macrofago; questi microbi intracellulari comprendono la Salmonella typhimurium, ed il Micobacterium tubercolosis. Alcuni di questi impediscono la fusione del lisosoma col fagosoma e proliferano nel citoplasma del macrofago infettato. I macrofagi secernono fattoriI macrofagi attivati secernono numerose importanti proteine che svolgono un ruolo centrale nello sviluppo della risposta immunitaria. Tra queste sono comprese l’interleuchina1, il TNFα e l’interleuchina6, citochine in grado di promuovere il processo infiammatorio. Ognuna di queste ha un’azione propria, ad esempio l’interleuchina1 attiva i linfociti, mentre tutte e tre agiscono sul centro ipotalamico della termoregolazione, responsabile della risposta febbrile. I macrofagi attivati secernono vari altri fattori coinvolti nella risposta immunitaria. Questi
comprendono un gruppo di proteine sieriche denominate complemento; inoltre gli enzimi idrolitici contenuti nei lisosomi possono anche essere secreti dal macrofago attivato. Esso è in grado di produrre anche fattori solubili come il fattore di necrosi tumorale. Il sistema del complemento consiste di una famiglia di proteine plasmatiche, che fungono da ponte tra il riconoscimento dei microbi e lo sviluppo dell’infiammazione e delle funzioni effettrici.
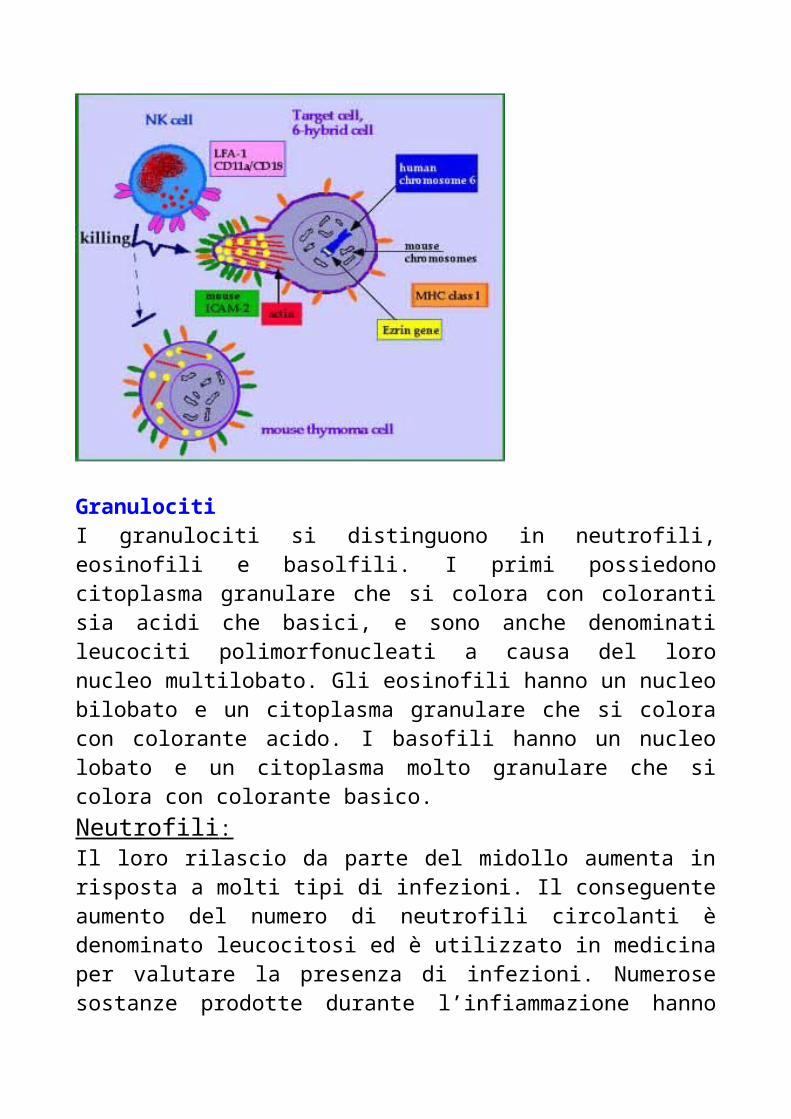
Cellule N KUn gruppo ristretto di linfociti del sangue periferico, detti cellule Null, non esprimono le molecole di membrana caratteristiche dei linfociti T e B. Queste cellule non esprimono neppure i recettori per l’antigene dei linfociti T e B, e sono pertanto prive di due attributi fondamentali dell’immunità: la specificità e la memoria. Le cellule null comprendono una popolazione linfocitaria dotata di una funzione ben definita, i linfociti natural killer, cellule di aspetto granulare che nell’uomo rappresentano il 5-10 % dei linfociti di sangue periferico.

Le cellule N K furono descritte per la prima volta nel 1976, quando si evidenziò che alcune cellule erano in grado di uccidere un ampio spettro di cellule tumorali in assenza di una pregressa immunizzazione nei confronti del tumore. Successivamente si osservò che queste cellule svolgono un ruolo importante nella difesa nei confronti delle cellule tumorali e cellule infettate da alcuni virus. In alcuni casi la cellula N K utilizza i propri recettori per riconoscere situazioni anomale, quali una ridotta espressione di MHC di classe1 o la presenza di antigeni non canonici espressi da queste cellule. Inoltre esse riescono a riconoscere una potenziale cellula bersaglio dalla presenza di anticorpi sulla stessa loro superficie, diretti contro cellule tumorali o infettate da virus.
GranulocitiI granulociti si distinguono in neutrofili, eosinofili e basolfili. I primi possiedono citoplasma granulare che si colora con coloranti sia acidi che basici, e sono anche denominati leucociti polimorfonucleati a causa del loro nucleo multilobato. Gli eosinofili hanno un nucleo bilobato e un

citoplasma granulare che si colora con colorante acido. I basofili hanno un nucleo lobato e un citoplasma molto granulare che si colora con colorante basico. Neutrofili : Il loro rilascio da parte del midollo aumenta in risposta a molti tipi di infezioni. Il conseguente aumento del numero di neutrofili circolanti è denominato leucocitosi ed è utilizzato in medicina per valutare la presenza di infezioni. Numerose sostanze prodotte durante l’infiammazione hanno azione chemiotattica sui neutrofili e promuovono il loro accumulo nel sito del danno. Tra queste si possono ricordare alcune componenti del complemento, fattori della cascata coagulativa e prodotti secreti dai linfociti T.
I neutrofili sono fagociti attivi, come i macrofagi, e la loro fagocitosi è simile a quella decritta per il macrofago, eccetto per il fatto che il neutrofilo possiede due tipi di granuli ripieni di sostanze battericide, quelli primari e quelli secondari. I granuli primari sono più grandi e densi e rappresentano una varietà di lisosoma: contengono perossidasi, lisozima, e vari enzimi.I granuli secondari sono più piccoli e contengono collagenasi, lattoferrina e lisozima. La capacità dei

neutrofili di uccidere i microrganismi ingeriti è molto superiore a quella dei macrofagi, a causa del loro burst respiratorio più potente. Eosinofili, basofili e mastcellule : Gli eosinofili come i neutrofili sono cellule mobili e fagocitarie che migrano dal sangue ai tessuti; pur essendo però la loro attività fagocitaria meno intensa. I basofili invece sono privi di attività fagocitaria e svolgono la loro funzione liberando le sostanze farmacologicamente attive, contenute nei loro granuli. Essi svolgono un ruolo centrale in alcune risposte allergiche.I precursori delle mastcellule si formano nel midollo osseo nel corso dell’emopoiesi, sono rilasciati nel sangue come precursori e si differenziano solo quando entrano nei tessuti. Le mastcellule si trovano in numerosi tessuti tra i quali la cute, il tessuto connettivo e il tessuto epiteliale mucoso dei tratti respiratorio, genito-urinario e digestivo. Essi contengono istamina e altre sostanze farmacologicamente attive importanti nello sviluppo dell’allergia.
Cellule dendriticheLe cellule dendritiche devono il loro nome alla fitta rete di lunghi processi della membrana che si dipartono dal soma cellulare, simili ai dendriti delle cellule nervose. Esse processano e presentano l’antigene ai linfociti T-helper. In base alla loro localizzazione vengono suddivise in:Cellule del Langerhans che si trovano nell’epidermide e nelle mucoseCellule dendritiche interstiziali che popolano la maggior parte degli organiCellule dendritiche interdigitate che si trovano in regioni ricche di linfociti T degli organi linfatici secondari e nella regione midollare del timoCellule dendritiche circolanti che raggruppano quelle presenti nel sangue che costituiscono lo 0,1% dei leucociti presenti e quelle presenti nella linfa.

Queste cellule sono cellule presentanti l’antigene molto più potenti dei macrofagi o dei linfociti B che necessitano entrambi di un’attivazione preventiva. Dopo aver fagocitato o endocitato l’antigene nei tessuti queste cellule migrano nel sangue e nella linfa raggiungendo così vari organi linfatici, dove presentano l’antigene ai linfociti T.
Tessuto linfatico associato alle mucoseLe membrane mucose che ricoprono i tratti digestivo, respiratorio e urogenitale sono la principale sede d’ingresso nell’organismo dei microrganismi patogeni. La difesa di queste superfici estremamente vulnerabili è assicurata da un tessuto linfatico organizzato denominato tessuto linfatico associato alle mucose (malt). Esso comprende aspetti diversi che vanno dagli infiltrati linfatici presenti nei villi intestinali, alle strutture organizzate presenti nelle tonsille, nell’appendice, e nelle placche di Peyer. L’importanza del malt nella difesa dell’organismo è provata dalla sua ricca popolazione di plasmacellule anticorposecernenti, il cui numero è di gran lunga superiore a quello delle plasmacellule totali presenti nella milza, nei linfonodi e nel midollo osseo.
Tessuto linfatico associato alla cuteCome sappiamo la pelle rappresenta una barriera anatomica importante verso l’ambiente esterno, grazie anche alla sua estesa superficie. Lo strato esterno della pelle, l’epidermide è costituito dai cheratinociti, i quali producono un gran numero di citochine coinvolte nelle infiammazioni locali, inoltre essi possono essere indotti a produrre MHC di classe 2, e possono comportarsi da cellule presentanti l’antigene. Nella matrice posta tra le cellule epidermiche si trovano le cellule di Langerhans. L’epidermide contiene infine i linfociti intraepidermici simili a quelli presenti nel malt; si pensa che l’unica funzione di
questo tipo cellulare sia di combattere gli antigeni che entrano attraverso la pelle.