Il fattore del sonno - Katawebdownload.kataweb.it/mediaweb/pdf/espresso/scienze/1976... · 2011. 9....
3
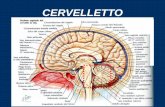
Il fattore del sonno si ottiene dal liquido cerebrospinale di capre non anestetizzate a cui siano state inserite due cannule di teflon. Le mem- brane che ricoprono le cavità che contengono il liquido cerebrospinale si possono attraversare senza rischi con le cannule; una volta tolti gli aghi, le membrane si g aldano da sole. Con il sistema indicato in questa illustrazione, per esempio, il liquido cerebrospinale normale può essere prelevato ininterrottamente dalla cavità cisternale del cer- vello della capra, attraverso la cannula posteriore, al ritmo di sei millilitri l'ora senza causare disagio all'animale. A sua volta, con la cannula anteriore tutto il sistema ventricolo-cisternale può essere riempito di liquido cerebrospinale artificiale. Nell'intervallo tra un espe- rimento e l'altro le cannule sono protette da un cappuccio metallico. 9//7 loMy. ) C-5 ‘) 2 /, \\\ CERVELLO / CERVELLETTO OSSO OCCIPITALE \ MIDOLLO SPINALE \\\\ \ \ \\ • \ . • \ \\,\ CANNULA INSERITA NEI VENTRICOLI , CANNULA COLLEGATA CON LA CISTERNA Il fattore del sonno Recentemente si è riusciti a isolare la sostanza che si accumula nel cervello durante la veglia prolungata e a trasferirla da un animale all'altro aumentando in quest'ultimo il normale periodo di sonno di John R. Pappenheimer tati positivi. Tuttavia tutti noi che abbia- mo preso parte agli esperimenti (Tracy B. Miller, Cecilie A. Goodrich, James Nicholl e io) avemmo la precisa sensa- zione che i gatti che avevano subito le trasfusioni di liquido da capre deprivate di sonno per quarantotto ore erano assai più assonnati in confronto agli stessi gat- ti ai quali era stato trasfuso liquido otte- nuto dalle capre che avevano dormito re- golarmente. In realtà i risultati furono così evidenti e potenzialmente di tale ri- levanza fisiologica che lasciammo da par- te altri progetti per dedicarci a tempo pieno allo studio sistematico del fenome- no Piéron. Il nostro interesse fu ulteriormente sti- molato quando sapemmo delle ricerche svolte nel laboratorio di Marcel Monnier a Basilea. Questi, insieme ai suoi colla- boratori, aveva appena scoperto che un fattore neuro-umorale collegato al sonno viene liberato nel sangue venoso del cer- vello quando determinate zone di una parte del cervello denominata talamo ventromediale vengono sottoposte a sti- molazioni elettriche. T l nostro primo obiettivo era quello di 1 stabilire un modo per valutare quanti- tativamente il fattore che produceva il sonno. Pensavamo che per isolare e de- purare il fattore attivo fossero necessarie centinaia di esperimenti. La prospettiva di verificare la durata del sonno in base a una analisi di migliaia di ore di regi- strazioni elettroencefalografiche non era molto attraente; sviluppammo perciò un semplice metodo di valutazione basato sulla attività notturna dei ratti. I ratti hanno un ciclo di sonno e di attività ben definito nelle ventiquattro ore: essi dor- mono di giorno circa il 65 per cento del tempo e di notte meno del 30 per cento del tempo. Mangiano e bevono soprat- tutto durante la notte. Il nostro progetto prevedeva l'infusio- ne nei ratti del presunto fattore sonno circa due ore prima dell'inizio del ciclo notturno; pensavamo che un tale fattore esogeno avrebbe potuto sopraffare lo sta- to naturale di veglia degli animali, facen- doli dormire di più e quindi essere meno attivi di notte. Costruimmo delle singole gabbie di at- tività per sedici ratti. Su ogni gabbia erano proiettati deboli raggi di luce rossa che convergevano su cellule fotocondut- trici. L'interruzione di un raggio lumino- so per due secondi metteva in moto un L 'idea che il sonno naturale venga causato da una sostanza secreta dal cervello durante la veglia, si accorda con l'esperienza di ogni giorno. Chiunque abbia osservato un bambino assonnato non avrà difficoltà a credere che alla base dell'intenso desiderio di dormire vi sia un qualche potente mecca- nismo chimico nel sistema nervoso cen- trale. Il commento «ho dormito come fossi drogato» esprime la sensazione in- tuitiva che una specie di «droga» natura- le abbia la funzione di produrre un son- no profondo e ristoratore. Così lago ammonisce Otello: «Non il papavero, né la mandragora né tutti gli sciroppi sonniferi del mondo saranno mai sufficienti a darti quel diSlce sonno che ieri era tuo.» È possibile che il cervello produca ogni mattina un naturale «sciroppo sonnife- ro» che ci fa dormire ogni notte? Più di sessanta anni fa lo psicofisiolo- go francese Henri Piéron effettuò espe- rimenti i quali sembravano dimostrare che durante una prolungata deprivazione di sonno il cervello produce una sostanza capace di indurre il sonno. Piéron e i suoi collaboratori praticarono delle tra- sfusioni di liquido cerebrospinale da cani che erano stati privati del sonno in cani normali e notarono che questi ultimi dor- mivano varie ore dopo la trasfusione. L'esperimento di Piéron, sebbene fos- se semplice, fu caratterizzato da difficol- tà tecniche e da confuse interpretazioni. Le trasfusioni implicavano un grande stress sia per gli animali donatori sia per i riceventi. I cani donatori erano tenuti svegli per dieci giorni o più, legati in posizione seduta di giorno e costretti a camminare continuamente durante la not- te. L'estrazione e la trasfusione del liqui- do cerebrospinale implicava una puntura della membrana atlanto-occipitale alla base del cervello, operazione che veniva compiuta senza anestesia; gli animali si dibattevano violentemente e spesso per- devano i sensi o rimanevano dopo la puntura parzialmente paralizzati. Gli e- sperimenti di Piéron avvenivano più di venticinque anni prima dell'introduzione dell'elettroencefalografia come strumen- to per l'analisi del sonno e i suoi risultati si basavano necessariamente sulla sem- plice osservazione del comportamento degli animali dopo le trasfusioni. Ciò nonostante, dopo molti esperimenti, Pié- ron giunse alla conclusione che mentre gli animali che ricevevano il liquido dai cani deprivati di sonno tendevano a dor- mire per varie ore, quelli che ricevevano il liquido di cani normali di solito rima- nevano svegli. Nel 1913 egli pubblicò una monografia sul sonno che compren- deva un ampio resoconto dei suoi espe- rimenti basati sulla trasfusione. L'esperi- mento di Piéron fu ripetuto nel 1939 da Jerome G. Schnedorf e A.C. Ivy della Nortwestern University. Essi riscontraro- no risultati positivi in nove delle venti trasfusioni. Il sonno nei cani riceventi aveva inizio circa quarantacinque minuti dopo la trasfusione, durava tra le due e le quattro ore e normalmente era accom- pagnato da un leggero aumento della temperatura corporea. Schnedorf e Ivy erano persuasi della realtà del fenomeno riscontrato da Piéron, ma si domanda- vano quali potessero essere i suoi rap- porti con il sonno normale a causa della variabilità della risposta, dell'aumento contemporaneo della temperatura corpo- rea e dell'eccezionale durata di depriva- zione di sonno necessaria per ottenere una risposta positiva. N essun ulteriore tentativo è stato com- piuto per analizzare il fenomeno Piéron fino a quando, nel 1965, i miei colleghi e io che lavoravamo presso il dipartimento di fisiologia della Harvard Medical School non prendemmo in esa- me il problema. In quel periodo stavamo studiando la composizione ionica del li- quido cerebrospinale in relazione al pro- blema del meccanismo del mantenimento del controllo della respirazione. A que- sto scopo avevamo messo a punto delle tecniche di perfusione delle cavità cere- brali di capre appositamente preparate ma non anestetizzate. La forma e le di- mensioni del cranio di questi animali rendono possibile l'introduzione di una cannula di teflon attraverso l'osso occipi- tale a livello della cavità cisternale, senza interferire con la libertà di movimento della testa e del collo (si veda l'illustrazio- ne nella pagina a fronte). In questo modo le membrane possono essere perforate in qualsiasi momento attraverso la cannula di teflon e il liquido cerebrospinale può essere estratto senza interruzione nella quantità di sei millilitri ogni ora senza danneggiare l'animale. Aggiungendo una seconda cannula, l'intero sistema ventri- colare e cisternale dell'animale non ane- stetizzato può essere perfuso di liquido ce- rebrospinale artificiale. Dopo l'estrazio- ne degli aghi, le membrane si saldano spontaneamente con scarso pericolo di infezione, purché durante l'operazione venga mantenuta una rigorosa asepsi. Tra un esperimento e l'altro i tubi ven- gono protetti da eventuali danni mecca- nici mediante un cappuccio metallico fis- sato alle corna dell'animale. Fatto questo, ci sembrò di aver creato condizioni favorevoli per riaffrontare lo studio del fenomeno Piéron senza arre- care il minimo disturbo agli animali. Par- timmo dal presupposto che, se il feno- meno era di fondamentale importanza nel meccanismo del sonno, esso doveva essere dimostrabile anche con depriva- zioni di sonno più brevi e che non dove- va essere limitato a una sola specie ani- male (ritenemmo cioè molto improbabile che ciascuna specie sintetizzasse un fat- tore diverso per una funzione così gene- ralizzata come il sonno). Con queste pre- messe, demmo inizio ad alcuni esperi- menti pilota con trasfusioni di liquido cisternale dalle nostre capre al sistema ventricolare di alcuni gatti. A questo sco- po preparammo alcuni gatti con tubi di teflon in maniera da poter perfonderli con liquidi lentamente e più volte senza alterare lo stato di coscienza normale dell'animale. Nei nostri primi esperimenti giudicam- mo i risultati dalla semplice osservazione comportamentale, proprio come aveva fatto Piéron cinquantacinque anni pri- ma. Non ci aspettavamo, in realtà, risul- 34 35
Transcript of Il fattore del sonno - Katawebdownload.kataweb.it/mediaweb/pdf/espresso/scienze/1976... · 2011. 9....
-
Il fattore del sonno si ottiene dal liquido cerebrospinale di capre nonanestetizzate a cui siano state inserite due cannule di teflon. Le mem-brane che ricoprono le cavità che contengono il liquido cerebrospinalesi possono attraversare senza rischi con le cannule; una volta tolti gliaghi, le membrane si galdano da sole. Con il sistema indicato inquesta illustrazione, per esempio, il liquido cerebrospinale normale
può essere prelevato ininterrottamente dalla cavità cisternale del cer-vello della capra, attraverso la cannula posteriore, al ritmo di seimillilitri l'ora senza causare disagio all'animale. A sua volta, con lacannula anteriore tutto il sistema ventricolo-cisternale può essereriempito di liquido cerebrospinale artificiale. Nell'intervallo tra un espe-rimento e l'altro le cannule sono protette da un cappuccio metallico.
9//7 loMy.) C-5
‘) 2/, \\\CERVELLO /
CERVELLETTO
OSSOOCCIPITALE
\MIDOLLO SPINALE
\\\\ \\ \\ • \
. • \
\\,\
CANNULA INSERITANEI VENTRICOLI
,
CANNULA COLLEGATACON LA CISTERNA
Il fattore del sonnoRecentemente si è riusciti a isolare la sostanza che si accumula nelcervello durante la veglia prolungata e a trasferirla da un animaleall'altro aumentando in quest'ultimo il normale periodo di sonno
di John R. Pappenheimer
tati positivi. Tuttavia tutti noi che abbia-mo preso parte agli esperimenti (TracyB. Miller, Cecilie A. Goodrich, JamesNicholl e io) avemmo la precisa sensa-zione che i gatti che avevano subito letrasfusioni di liquido da capre deprivatedi sonno per quarantotto ore erano assaipiù assonnati in confronto agli stessi gat-ti ai quali era stato trasfuso liquido otte-nuto dalle capre che avevano dormito re-golarmente. In realtà i risultati furonocosì evidenti e potenzialmente di tale ri-levanza fisiologica che lasciammo da par-te altri progetti per dedicarci a tempopieno allo studio sistematico del fenome-no Piéron.
Il nostro interesse fu ulteriormente sti-molato quando sapemmo delle ricerchesvolte nel laboratorio di Marcel Monniera Basilea. Questi, insieme ai suoi colla-
boratori, aveva appena scoperto che unfattore neuro-umorale collegato al sonnoviene liberato nel sangue venoso del cer-vello quando determinate zone di unaparte del cervello denominata talamoventromediale vengono sottoposte a sti-molazioni elettriche.
T l nostro primo obiettivo era quello di1 stabilire un modo per valutare quanti-tativamente il fattore che produceva ilsonno. Pensavamo che per isolare e de-purare il fattore attivo fossero necessariecentinaia di esperimenti. La prospettivadi verificare la durata del sonno in basea una analisi di migliaia di ore di regi-strazioni elettroencefalografiche non eramolto attraente; sviluppammo perciò unsemplice metodo di valutazione basatosulla attività notturna dei ratti. I ratti
hanno un ciclo di sonno e di attività bendefinito nelle ventiquattro ore: essi dor-mono di giorno circa il 65 per cento deltempo e di notte meno del 30 per centodel tempo. Mangiano e bevono soprat-tutto durante la notte.
Il nostro progetto prevedeva l'infusio-ne nei ratti del presunto fattore sonnocirca due ore prima dell'inizio del ciclonotturno; pensavamo che un tale fattoreesogeno avrebbe potuto sopraffare lo sta-to naturale di veglia degli animali, facen-doli dormire di più e quindi essere menoattivi di notte.
Costruimmo delle singole gabbie di at-tività per sedici ratti. Su ogni gabbiaerano proiettati deboli raggi di luce rossache convergevano su cellule fotocondut-trici. L'interruzione di un raggio lumino-so per due secondi metteva in moto un
L
'idea che il sonno naturale vengacausato da una sostanza secretadal cervello durante la veglia, si
accorda con l'esperienza di ogni giorno.Chiunque abbia osservato un bambinoassonnato non avrà difficoltà a credereche alla base dell'intenso desiderio didormire vi sia un qualche potente mecca-nismo chimico nel sistema nervoso cen-trale. Il commento «ho dormito comefossi drogato» esprime la sensazione in-tuitiva che una specie di «droga» natura-le abbia la funzione di produrre un son-no profondo e ristoratore. Così lagoammonisce Otello:«Non il papavero, né la mandragorané tutti gli sciroppi sonniferidel mondo saranno mai sufficientia darti quel diSlce sonnoche ieri era tuo.»
È possibile che il cervello produca ognimattina un naturale «sciroppo sonnife-ro» che ci fa dormire ogni notte?
Più di sessanta anni fa lo psicofisiolo-go francese Henri Piéron effettuò espe-rimenti i quali sembravano dimostrareche durante una prolungata deprivazionedi sonno il cervello produce una sostanzacapace di indurre il sonno. Piéron e isuoi collaboratori praticarono delle tra-sfusioni di liquido cerebrospinale da caniche erano stati privati del sonno in caninormali e notarono che questi ultimi dor-mivano varie ore dopo la trasfusione.
L'esperimento di Piéron, sebbene fos-se semplice, fu caratterizzato da difficol-tà tecniche e da confuse interpretazioni.Le trasfusioni implicavano un grandestress sia per gli animali donatori sia peri riceventi. I cani donatori erano tenutisvegli per dieci giorni o più, legati inposizione seduta di giorno e costretti acamminare continuamente durante la not-te. L'estrazione e la trasfusione del liqui-do cerebrospinale implicava una punturadella membrana atlanto-occipitale allabase del cervello, operazione che venivacompiuta senza anestesia; gli animali sidibattevano violentemente e spesso per-devano i sensi o rimanevano dopo lapuntura parzialmente paralizzati. Gli e-sperimenti di Piéron avvenivano più di
venticinque anni prima dell'introduzionedell'elettroencefalografia come strumen-to per l'analisi del sonno e i suoi risultatisi basavano necessariamente sulla sem-plice osservazione del comportamentodegli animali dopo le trasfusioni. Ciònonostante, dopo molti esperimenti, Pié-ron giunse alla conclusione che mentregli animali che ricevevano il liquido daicani deprivati di sonno tendevano a dor-mire per varie ore, quelli che ricevevanoil liquido di cani normali di solito rima-nevano svegli. Nel 1913 egli pubblicòuna monografia sul sonno che compren-deva un ampio resoconto dei suoi espe-rimenti basati sulla trasfusione. L'esperi-mento di Piéron fu ripetuto nel 1939 daJerome G. Schnedorf e A.C. Ivy dellaNortwestern University. Essi riscontraro-no risultati positivi in nove delle ventitrasfusioni. Il sonno nei cani riceventiaveva inizio circa quarantacinque minutidopo la trasfusione, durava tra le due ele quattro ore e normalmente era accom-pagnato da un leggero aumento dellatemperatura corporea. Schnedorf e Ivyerano persuasi della realtà del fenomenoriscontrato da Piéron, ma si domanda-vano quali potessero essere i suoi rap-porti con il sonno normale a causa dellavariabilità della risposta, dell'aumentocontemporaneo della temperatura corpo-rea e dell'eccezionale durata di depriva-zione di sonno necessaria per ottenereuna risposta positiva.
Nessun ulteriore tentativo è stato com-piuto per analizzare il fenomeno
Piéron fino a quando, nel 1965, i mieicolleghi e io che lavoravamo presso ildipartimento di fisiologia della HarvardMedical School non prendemmo in esa-me il problema. In quel periodo stavamostudiando la composizione ionica del li-quido cerebrospinale in relazione al pro-blema del meccanismo del mantenimentodel controllo della respirazione. A que-sto scopo avevamo messo a punto delletecniche di perfusione delle cavità cere-brali di capre appositamente preparatema non anestetizzate. La forma e le di-mensioni del cranio di questi animali
rendono possibile l'introduzione di unacannula di teflon attraverso l'osso occipi-tale a livello della cavità cisternale, senzainterferire con la libertà di movimentodella testa e del collo (si veda l'illustrazio-ne nella pagina a fronte). In questo modole membrane possono essere perforate inqualsiasi momento attraverso la cannuladi teflon e il liquido cerebrospinale puòessere estratto senza interruzione nellaquantità di sei millilitri ogni ora senzadanneggiare l'animale. Aggiungendo unaseconda cannula, l'intero sistema ventri-colare e cisternale dell'animale non ane-stetizzato può essere perfuso di liquido ce-rebrospinale artificiale. Dopo l'estrazio-ne degli aghi, le membrane si saldanospontaneamente con scarso pericolo diinfezione, purché durante l'operazionevenga mantenuta una rigorosa asepsi.Tra un esperimento e l'altro i tubi ven-gono protetti da eventuali danni mecca-nici mediante un cappuccio metallico fis-sato alle corna dell'animale.
Fatto questo, ci sembrò di aver creatocondizioni favorevoli per riaffrontare lostudio del fenomeno Piéron senza arre-care il minimo disturbo agli animali. Par-timmo dal presupposto che, se il feno-meno era di fondamentale importanzanel meccanismo del sonno, esso dovevaessere dimostrabile anche con depriva-zioni di sonno più brevi e che non dove-va essere limitato a una sola specie ani-male (ritenemmo cioè molto improbabileche ciascuna specie sintetizzasse un fat-tore diverso per una funzione così gene-ralizzata come il sonno). Con queste pre-messe, demmo inizio ad alcuni esperi-menti pilota con trasfusioni di liquidocisternale dalle nostre capre al sistemaventricolare di alcuni gatti. A questo sco-po preparammo alcuni gatti con tubi diteflon in maniera da poter perfonderlicon liquidi lentamente e più volte senzaalterare lo stato di coscienza normaledell'animale.
Nei nostri primi esperimenti giudicam-mo i risultati dalla semplice osservazionecomportamentale, proprio come avevafatto Piéron cinquantacinque anni pri-ma. Non ci aspettavamo, in realtà, risul-
34
35
-
•=k-r.:=wr"---.A.w"-Air.y"L-M==~"--EREE='—'
m-- — .,—.-
o-- -=-
i11 1
i% il
11111111111
ii 911111 E IIII JUIlliiiiiiiii II • •fill E IEREMMIIIIMIMMI•••
••••• ‘,, II M-,:c IMRE • •..1••••••••••11111 111 1•••• ••n•IIITFPT—-- .. fill I •MIEMMIZE f IMEIMINNI 1111~~....
'.;-.Eimmi• mem ,', ',
' , •."'.
I ''')
ME 1111.MWilli
.. , -
' ' '' ///',' . ,„'
'"
'i
M••••1I•Ellammessmla
•••••NEMEREMil
1111 li ii1111111 11111
wourilfr// ,, ,, ,fl'O ' o misME ' IM•••• ~mia Nomi
••• imissommmimis mia
•i
•!111111111
Il II
ili
ME ,,••
111•111•••••111~111111~••• • MI
•11~111• ' •MaiMETIniffilh.1-1
Il I III I Id I.a IIIIMMMEMMIIMI EMailiellE
111i , iIalli
Elleq 114ME IIMIMMIE
M ••••••••••n:.-~1.~
M MIERI 5' 'Palw
Il--------
Ligillinal~~wwwww~EZIE!:~misudill
LiildiLlid~oh
-------~-- —p—
MnERE
-- I EE1E--___Twea
DALLE ORE 18 ALLE ORE 24
LIQUIDOCEREBROSPINALE
ARTIFICIALE
LIQUIDOCEREBROSPINALE
PRELEVATO DACAPRE NORMALI
LIQUIDOCEREBROSPINALE
PRELEVATO DACAPRE DEPRIVATE
DI SONNO
LIQUIDOCEREBROSPINALE
ARTIFICIALE
LIQUIDOCEREBROSPINALE
PRELEVATO DACAPRE NORMALI
LIQUIDOCEREBROSPINALE
PRELEVATO DACAPRE DEPRIVATE
DI SONNO
0,1 0,2 0,3 0.4 0.5 0.6 0,7 0.8 0,9RAPPORTO FRA L'ATTIVITÀ NOTTURNA DURANTE L'ESPERIMENTO
E L'ATTIVITÀ NELLA NOTTE PRECEDENTE
DALLE ORE 18 ALLE ORE 24
Il fattore del sonno siene iniettalo lentamente nei ventricoli cerebralidi un animale da esperimenti) come il ratto, attraverso una cannula.senza recare alcun disturbo per l'animale normalmente cosciente.L'azione del fattore viene misurata controllando l'attività notturnadel ratto di laboratorio tenuto in una gabbia appositamente costruitacome quella che appare nel disegno. La gabbia, che fa parte di un
blocco di sedici appartenenti al laboratorio dell'autore presso la facoltàdi medicina della iian arti iiiteriI, altrasersata da deboli raggi diluce rossa tocalizzati sii cellule foloconduttrici. Quando il ratto si!mune, l'interruzione per due secondi di uno dei raggi di luce mette inazione un contatore situato in una stanza contigua e permette la rego-lare registrazione (kW:Utilità del ratto per tutta la durata delle 24 ore.
RATTO A
RATTO B
1—ht
Mentre si misurava l'attività notturna dei ratti venivano effettuatecontemporaneamente registrazioni elettroencefalografiche dei poten-ziali cerebrali: questo per ottenere una prova più specifica dell'effettosonno che segue all'infusione di liquido cerebrospinale prelevato dacapre deprivate di sonno. Gli elettroencefalogrammi qui rappresentati,ottenuti simultaneamente da due diversi ratti, sono caratterizzali dacicli episodici di sonno a onda lenta (bande colorate). Gli episodi
ognuno dei quali dura alcuni minuti, si distinguono per cambiamentiabbastanza rilevanti delle tensioni nelle basse frequenze. La registra-zione dell'attività notturna dei due ratti si può vedere nel grafico alcentro. Una deviazione verso l'alto del pennino (che segnala l'interru-zione di un raggio di luce) si riferisce al ratto A così come unadeviazione verso il basso si riferisce al ratto B. Come dimostra questasequenza di registrazioni, al sonno si alternano brevi periodi di veglia.
DALLE ORE 18 ALLE ORE 24 DALLE ORE 18 ALLE ORE 24
DALLE ORE 18 ALLE ORE 24 DALLE ORE 18 ALLE ORE 24
contatore nella camera accanto. L'insie-me dei sedici contatori veniva fotografa-to automaticamente ogni sei ore per ot-tenere una registrazione della attività diogni ratto nelle ventiquattro ore. La stan-za del ratto era ad aria condizionata e atemperatura controllata ed era mantenu-ta a un ciclo di dodici ore di luce e dodicidi oscurità.
Abbiamo sviluppato delle semplici tec-niche per impiantare i tubi-guida ventri-colari nei ratti e abbiamo notato che glianimali sopportavano ripetute infusioniintraventricolari a un ritmo che ci con-sentiva di introdurre un millilitro di li-quido ogni trenta minuti.
Presto ci rendemmo conto che era ne-cessario lavorare solo con ratti maschiper evitare le grandi variazioni nella atti-vità notturna, connesse nelle femminecon i quattro giorni del ciclo sessuale.C'erano anche grandi variazioni d'attivi-tà tra i diversi ratti maschi, ma l'attivitànotturna d'ogni singolo ratto era quasicostante da una notte all'altra. Ogni rat-to poteva quindi essere utilizzato comecontrollo di se stesso: la media della suaattività notturna prima dell'infusione ser-viva come indicazione di base per unconfronto con l'attività che veniva misu-rata durante le notti in cui veniva affet-tuata l'infusione.
T risultati della prima serie di esperi--1- menti sul liquido cerebrospinale inte-ro, sterilizzato, preso da capre normali eda capre sottoposte a quarantotto ore dideprivazione di sonno, hanno dimostra-to che l'attività notturna dei ratti usatiper l'esperimento si riduceva al 63 percento circa del normale, durante le primesei ore dopo l'infusione di 0,1 millilitri diliquido cerebrospinale estratto da capredeprivate di sonno. Inoltre abbiamo tro-vato che questa riduzione era significati-vamente maggiore di quella conseguentel'infusione di soluzione salina sterile, odi liquido estratto da capre normali. Lariduzione dell'attività notturna in seguitoall'infusione di liquidi normali venne at-tribuita al fatto che i liquidi venivanoinfusi durante le due ore precedenti ilciclo d'oscurità, cioè in un periodo in cuigli animali normalmente dormono. Que-sti sono quindi privati di quasi due ore disonno e tendono a recuperarlo più tardidurante la notte.
Infusioni simulate ottenevano lo stessorisultato. Tuttavia la differenza tra glieffetti dell'infusione di liquido cerebro-spinale normale e quelli di liquido cere-brospinale estratto da capre deprivate disonno era statisticamente molto rilevan-te. La nostra valutazione basata sull'atti-vità notturna è utile perché permette diselezionare una grande varietà di cam-pioni, ma, naturalmente, non è specificaper il sonno.
Una dimostrazione più rigorosa del-l'effetto sonno richiede o una registra-zione continua, oppure la media di molteregistrazioni elettroencefalografiche.noto che il sonno, come è indicato dal-l'elettroencefalogramma, è caratterizzatoda cicli episodici di durata variabile tra
L'attività notturna dei ratti si riduce approssimativamente al 63 per cento del livello normale,durante la prima metà della notte che fa seguito alla infusione intraventricolare d'un decimo dimillilitro di liquido cerebrospinale prelevato da capre deprivate di sonno (istogrammi in alto).La riduzione dell'attività che segue l'infusione dei due liquidi di controllo (liquido cerebrospina-le artificiale e liquido cerebrospinale preso da capre normali) dipende dal fatto che tutte le infu-sioni sono state praticate due ore prima del ciclo notturno, quando i ratti sono di norma addor-mentati: essi vengono perciò deprivati di sonno e tendono a compensare il deficit nella prima partedella notte. Durante la seconda metà della notte i livelli di attività dei ratti sperimentali mostra-no un recupero quasi completo raggiungendo il normale livello d'attività (istogrammi in basso).
LIQUIDOCEREBROSPINALE
PRELEVATO DACAPRE NORMALI
LIQUIDOCEREBROSPINALE
PRELEVATO DACAPRE DEPRIVATE
DI SONNO
0 0,1 0,2 0,3 0,4 0,5 0 6 0 7 0 8 0 9 1 0 0,1 0,2 0,3 0,4 0,5 0,6 0,7 0 8 0,9RAPPORTO FRA L'ATTIVITÀ NOTTURNA DURANTE L'ESPERIMENTO
E L'ATTIVITA NELLA NOTTE PRECEDENTE
LIQUIDOCEREBROSPINALE
PRELEVATO DACAPRE NORMALI
LIQUIDOCEREBROSPINALE
PRELEVATO DACAPRE DEPRIVATE
DI SONNO
-20 o 20 40 60 -20 o 20 40 60VARIAZIONI NELLA DURATA DEL SONNO A ONDA LENTA (PER CENTO)
Misurazioni simultanee dell'altisità notturna dei ratti e della durata dei loro episodi di sonno aonda lenta sono state ottenute dopo infusioni intraventricolari di liquido cerebrospinale presoda capre deprivate di sonno e da capre di controllo. L'evidente correlazione tra la riduzionedell'attività notturna e l'aumento della durata del sonno a onda lenta associato con l'infusionedi liquido proveniente da capre deprivate di sonno, ha dimostrato che una sostanza capace diindurre il sonno si accumula nel sistema cerebrospinale durante uno stato di veglia prolungata.
36 37
-
500r
o
500_
t4i0MM9
500~
O
500
500r
O
500
Gli elettroencefalogrammi di coniglio dopo infusione di fattore del sonno concentrato (graficoin alto) e dopo 24 ore di deprivazione di sonno (grafico al centro) vengono qui rappresentatiinsieme all'elettroencefalogramma normale dello stesso coniglio (grafico in basso). Ognuna diqueste registrazioni di un minuto rappresenta la media di 20 registrazioni fatte nelle ore che prece-dono o seguono la registrazione qui illustrata. 1 primi due grafici indicano la grande ampiezza delleonde lente che sono caratteristiche sia del sonno causato da infusione del fattore, sia del sonnoprofondo che segue la deprivazione di sonno. Presumibilmente quest'ultimo è prodotto da unaccumularsi di fattore sonno naturale durante un periodo di veglia. Le tensioni medie dei tre e-sempi qui mostrati sono 140 microvolt (in alto), 138 microvolt (al centro), 91 microvolt (in basso).
novanta minuti circa nell'uomo e pochiminuti nei piccoli animali da laboratorio.
Nei ratti adulti e nei conigli ogni ciclodi sonno è essenzialmente costituito da«sonno a onda lenta», è cioè caratteriz-zato da un tracciato elettroencefalografi-co con differenze di potenziale piuttostoampie e basse frequenze.
Facendo esperimenti su ratti a cui era-no stati applicati degli elettrodi (oltre aitubi collegati con i ventricoli) abbiamopotuto dimostrare che la riduzione del-l'attività notturna in seguito all'infusio-ne del liquido cerebrospinale estratto dacapre deprivate del sonno era propriolegata a un aumento della durata delsonno a onda lenta, quale viene misuratomediante registrazioni elettroencefalo-grafiche continue (si veda la figura inbasso a pagina 36). Questi risultati cihanno largamente confermato il fenome-no Piéron. Abbiamo così dimostrato,inoltre, che erano sufficienti un giorno odue di privazione di sonno perché la so-stanza che lo provoca fosse presente nel
o100
80
o
uJooUio_— 60
Ui
o
O
ozz 40OUi
oO
Ui
o2
< 20
liquido cerebrospinale e che questa so-stanza non è specifica della specie.
Avendo ottenuto tali risultati, ci siamo uniti a Manfred L. Karnovsky del
Dipartimento di chimica biologica dellaHarvard Medical School per svolgere unprogramma di ricerca sull'identificazionedel fattore che produce sonno. Questoprogramma è durato cinque anni ed è an-cora in corso. Sono stati comunque com-piuti considerevoli progressi nella purifi-cazione del fattore sonno dal liquido ce-rebrospinale, e più recentemente dall'in-tero cervello di animali deprivati di son-no. Per poter disporre di sufficiente ma-teriale per iniziare tali esperimenti, ab-biamo tenuto in laboratorio un gruppodi venti capre provviste in permanenza ditubi. Non potevamo però praticare pun-ture nel cervello delle capre più d'unavolta ogni tre settimane se non volevamocorrere il rischio che si formassero delleaderenze delle membrane cisternali.
Sulla base di tale programma di lavoro
e di alcuni errori inevitabili, sono statinecessari quasi tre anni per raccogliere isei litri di liquido necessari per mettere apunto le procedure di purificazione. Illavoro è stato costantemente ostacolatodal fatto che le procedure di fraziona-mento standard introducevano dei fatto-ri di contaminazione che interferivanocon la valutazione del sonno naturale,quando venivano introdotti direttamentenei ventricoli cerebrali. Inoltre, alcunidei normali componenti il liquido cere-brospinale possono causare reazioni dieccitazione, qualora vengano immessinella cavità ventricolare in alta concen-trazione. Alla fine, comunque, siamo riu-sciti a ottenere un grado considerevole dipurificazione e concentrazione del fatto-re sonno da grandi quantità di liquidocerebrospinale.
I soluti con un peso molecolare supe-riore a circa 500 sono stati eliminati fil-trando il liquido attraverso una serie disetacci molecolari ultrafini. Soluti conun peso molecolare inferiore a 350 sonostati eliminati trattando il liquido contecniche di gel-filtrazione. Abbiamo no-tato che una soluzione concentrata di li-quido cerebrospinale contenente solo so-luti con un peso molecolare compresotra 350 e 500 riduceva l'attività notturnadei ratti del 50 per cento rispetto allivello normale per le dodici ore dopol'infusione nei ventricoli del cervello deiratti; nei conigli la durata normale delsonno era aumentata del 50 per centocirca durante le sei ore dopo l'infusioneintraventricolare.
L'effetto sonno delle soluzioni concen-trate poteva essere annullato per incuba-zione con un enzima proteolitico, e que-sto dimostra che la molecola del fattoresonno deve contenere almeno un legamepeptidico. La frazione attiva contenevagruppi amminici primari che reagivanoai test di fluorescenza standard nella ma-niera caratteristica dei peptidi. È possibi-le, comunque, che i peptidi fossero pre-senti come impurità nella frazione attivae sarebbe prematuro concludere che ilfattore sonno è un peptide semplice.
La quantità di fattore sonno nel liqui-do cerebrospinale è evidentemente moltoridotta. In realtà, la maggior parte delmateriale che abbiamo raccolto con tan-ta fatica da litri di liquido cerebrospinaleè stata consumata per imparare comefare a purificare e a concentrare il fatto-re. Dal momento che molte sostanze tro-vate nel liquido cerebrospinale sono pre-senti in concentrazione molto più altanel cervello, abbiamo tentato di estrarreil fattore sonno dai tessuti cerebrali dicapre deprivate di sonno, servendoci dialcune delle procedure di frazionamentousate in precedenza per il liquido cere-brospinale.
Le estrazioni iniziali dall'intero ence-falo, dal tronco cerebrale e dalla cortec-cia cerebrale venivano effettuate con tec-niche di cui è nota l'efficacia per l'estra-zione di ormoni composti da pochi am-minoacidi. Gli estratti cerebrali venivanopoi sottoposti alle stesse tecniche di puri-ficazione usate per il liquido cerebrospi-
nale. Si constatò che erano attive nel pro-muovere il sonno le stesse frazioni checontenevano il fattore del sonno nel li-quido cerebrospinale. Quando abbiamosperimentato l'effetto di questo fattore inconigli in cui erano stati infusi estratti pu-rificati di tessuti cerebrali di capre depri-vate di sonno, constatammo che l'attivi-tà espressa in grammi di tessuto cerebra-le appariva maggiore negli estratti prepa-rati dal tronco cerebrale che in quellipreparati dalla corteccia cerebrale, ma lavalutazione non era abbastanza precisadal punto di vista quantitativo da per-mettere di esserne certi.
IT I sonno supplementare prodotto da
un'infusione del nostro fattore non èininterrotto; si tratta, piuttosto, di unaumento della durata e della frequenzadegli episodi di sonno. Gli animali daesperimento sono facilmente disturbatidai rumori e si svegliano a intervalli permangiare o per bere. Abbiamo notatoche l'ampiezza del tracciato a onda lentadurante gli episodi di sonno era note-volmente maggiore dell'ampiezza osser-vata normalmente. Questo si poteva fa-cilmente verificare su registrazioni elet-troencefalografiche standard, ma per a-nalizzare l'effetto quantitativamente ab-biamo fatto passare i segnali in uscitadell'elettroencefalografo attraverso filtrielettrici correttivi e integratori per otte-nere la tensione media integrata. Le inte-grazioni venivano registrate su nastro o-gni due minuti. Abbiamo trovato che neiconigli l'ampiezza del tracciato nell'arcodelle basse frequenze durante il sonno aonda lenta provocato dal fattore sonnoera superiore del 50 per cento all'am-piezza misurata nel sonno normale. Laampiezza, per quel che riguarda le altrefrequenze, era immutata. Evidentementela sostanza che provoca il sonno ha uneffetto specifico sugli eventi che determi-nano le tensioni a onda lenta.
Se un fattore sonno esogeno, cioè nonnaturale, estratto da tessuti cerebrali dianimali deprivati di sonno poteva pro-durre sonno supplementare con tensionia onda lenta di ampiezza abnorme, sem-brava corretto supporre che, deprivandodel sonno gli animali da esperimento, a-vremmo ottenuto tracciati simili per ac-cumulo del fattore sonno endogeno onaturale. Gli esperimenti dimostraronoche è proprio così. L'ampiezza degli elet-troencefalogrammi a bassa frequenza delsonno a onda lenta dopo 24 ore di depri-vazione di sonno era indistinguibile daquella degli elettroencefalogrammi regi-strati nello stesso coniglio dopo infusio-ne di fattore sonno esogeno. Il sonno aonda lenta che seguiva la deprivazione disonno, differiva comunque da quello suc-cessivo all'infusione del fattore sonnoper la modalità d'insorgenza. Nel primocaso il sonno a onda lenta di grandeampiezza comincia quasi subito; nel se-condo, cioè nel sonno supplementareprodotto da un fattore esogeno, non rag-giunge il massimo che dopo due o tre oredall'infusione. Il ritardo nell'inizio delsonno dopo l'infusione del liquido nei
ventricoli del cervello può essere attri-buito al periodo di tempo necessario per-ché il fattore sonno esogeno si diffondadal sistema ventricolare al suo sito diattività nel tessuto cerebrale. La sostanzaendogena, invece, può accumularsi den-tro il cervello in vicinanza del suo sitoattivo.
T l successo nell'estrazione e purificazio-1 ne del fattore sonno dai tessuti cere-brali degli animali deprivati di sonno co-stituisce un importante progresso perchéapre la via all'acquisizione di grandi quan-tità di materiale per dare inizio a stu-di sistematici sulla sua struttura chimicae la funzione fisiologica. La nostra inda-gine sui tessuti cerebrali come materialeutile a questo scopo ha coinciso con glistudi condotti indipendentemente da KojiUkizono e dai suoi collaboratori dell'U-niversità di Tokio. Questi autori hannoestratto e purificato una sostanza cheprovoca il sonno dal tronco cerebrale di1000 ratti deprivati di sonno. La sostan-za estratta a Tokio sembra avere alcuneproprietà in comune con la nostra. Essaè presente nelle stesse frazioni purificate,e ha effetti similari sull'attività notturnae sulla durata del sonno dei ratti. Fino aquesto punto nessuno dei due gruppi èriuscito a ottenere una sostanza abba-stanza purificata da permettere studistrutturali definitivi, ma per raggiungerequesto fine si continua a lavorare sia aTokio sia a Boston.
La quantità di sostanza attiva che si
può estrarre dal cervello di animali de-privati di sonno è estremamente ridotta:un milionesimo di grammo, circa, perogni 100 grammi di tessuto cerebrale. Èquindi evidente che le fonti naturali nonsono in grado di fornire una quantitàsufficiente di questa sostanza allo statopuro per poter effettuare sistematici stu-di fisiologici e biochimici sulla modalitàd'azione di tale fattore. Perciò i nostrisforzi sono al momento diretti a indivi-duare la struttura chimica della sostanzaallo scopo di riuscire a produrla sinteti-camente.
Se si potrà produrre sinteticamente ilfattore sonno molti problemi di grandeimportanza e interesse potranno essereaffrontati sperimentalmente. Qual è, peresempio, il rapporto fra il fattore sonnoe la teoria del «neurotrasmettitore» diM. Jouvet? Quale relazione c'è fra ilfattore sonno e le modificazioni biochi-miche che avvengono durante il sonno?Non si potrebbe marcare il fattore sonnocon un indicatore radioattivo per indivi-duarne l'origine e il luogo di azione alivello cellulare? Sarebbe possibile alte-rare la molecola in modo tale da facili-tarne l'ingresso nel cervello dopo som-ministrazione orale o endovenosa apren-do così la strada a possibili applicazionicliniche? Se avessimo a disposizione scor-te sufficienti di sostanza allo stato puropotremmo tentare di avvicinarci alla so-luzione di uno dei più misteriosi problemidella fisiologia contemporanea, cioè lafunzione e il meccanismo del sonno.
ORE DOPO L'INFUSIONE34
7
2 3 4 5
7
ORE DOPO LA DEPRIVAZIONE DI SONNO
L'aumento del sonno a onda lenta è stato osservato anche nei conigli in seguito all'infusioneintraventricolare di fattore del sonno concentrato, prelevato dal cervello di capre e pecoredeprivate di sonno (in colore). L'aumento del sonno a onda lenta associato a tali infusioni è quiparagonalo all'aumento che si verifica negli stessi conigli senza l'infusione, ma dopo 24 ore dideprivazione di sonno (in nero). Il ritardo di due o tre ore nell'inizio del sonno che seguel'infusione indica che il fattore del sonno esogeno deve diffondersi lentamente fino al sito attivonel cervello, mentre il fattore sonno formatosi durante il periodo di deprivazione si trova nellesue immediate adiacenze e lo raggiunge senza difficoltà. Le sbarre verticali indicano la pro-babilità statistica d'errore nei confronti di tredici diversi test eseguiti su cinque differenti conigli.
38
39
Page 1Page 2Page 3