I linfociti T e il loro recettore -...
5
La «istruzione» di un linfocito T nel timo gli conferisce la capacità di distinguere le altre cellule dello stesso organismo da cellule estranee. Questa capacita è un elemento d'importanza critica nella specificità della risposta immunitaria. Il timocito, o linfocito T immaturo, che si nota nella microfotografia, appare ingrandito 20 900 diametri ed è unito a una cellula epiteliale che fa parte del rivestimento del timo (si veda lo schema a destra),I siti di legame scuri, presenti sulla superficie del timocito, sono le molecole dei recettori: esse probabilmente si legano a proteine che definiscono l'«io» e che sono esposte sulla cellula epiteliale del timo. Queste proteine sono codificate nel genoma da un segmento di DNA chiamato maggior complesso di istocompatibilità ( mHc ). Il legame può stimolare l'inizio della «istruzione », o maturazione, del timocito. I linfo- citi T maturi acquisiscono una scarsa affinità per le proteine del proprio io, codificate dal MHC, mentre hanno una affinità elevata per queste stesse proteine quando sono associate ad antigeni. I linfociti T innescano una risposta immunitaria solo contro le cellule infettate del proprio io, oppure contro cellule appartenenti a un individuo estraneo. La microfotografia è stata ottenuta da Andrew G. Farr dell'Università di Washington. CELLULA EPITELIALE DI TIMO RECETTORI DEL LINFOCITO T L e cellule del sistema immunitario per- mettono ai vertebrati di ricono- scere che materiali estranei, gli antigeni, hanno invaso il loro organismo. La risposta immunitaria che ne consegue è notevole per la sua specificità. Per esem- pio, un essere umano immunizzato me- diante vaccinazione contro il vaiolo può opporre resistenza a un'infezione provoca- ta da virus del vaiolo, ma non, per esem- pio, a un attacco di virus dell'influenza. Nell'ultimo decennio gli immunologi sono riusciti a riconoscere che il fattore più im- portante nella capacità del sistema immu- nitario di reagire specificamente ai virus è rappresentato da una classe di cellule di modeste dimensioni, i linfociti T(chiamati anche cellule T). I linfociti T, infatti, svol- gono un ruolo ausiliare essenziale nella ri- sposta che il sistema immunitario dà all'in- fezione batterica. Da lungo tempo è stata accettata l'idea che un'attività di questo tipo venga inne- scata da una molecola inclusa nella mem- brana dei linfociti Te chiamata «recettore dei linfociti T». Si presume che un antigene specifico si adatti e si leghi a questo recet- tore come una chiave si adatta alla giusta serratura, mettendo quindi in moto la serie complessa di eventi biochimici che costi- tuiscono la risposta immunitaria. Per tutta una serie di ragioni, l'isolamento del recet- tore dei linfociti T è risultato estremamen- te difficile e, fino a non molto tempo fa, è stato necessario dedurne le proprietà per via indiretta, senza essere guidati dalla co- noscenza della sua struttura, che oggi sta per essere messa a fuoco. Il chiarimento sulla struttura del recet- tore dei linfociti T porta con sé anche una migliore comprensione delle complesse in- terazioni che si instaurano tra linfociti T e altri elementi del sistema immunitario. In particolare, risulta sempre più evidente che il linfocito T è particolarmente adatto a fronteggiare le infezioni associate alle cellule dell'ospite, e non le infezioni che circolano liberamente nei liquidi corporei dell'ospite. Per svolgere questo compito, il recettore dei linfociti T deve non solo rico- noscere un antigene specifico, ma che an- che certe proteine di membrana della stes- sa cellula ospite. Un simile meccanismo di riconoscimento deve essere tenuto sotto ri- goroso controllo perché un linfocito T, se dovesse essere attivato soltanto dalle pro- teine dell'ospite, potrebbe facilmente rivol- gersi contro le cellule sane di quest'ultimo. La conseguente sensibilità del sistema di riconoscimento dei linfociti T è responsa- bile di molte affascinanti proprietà di que- sto sistema, importanti dal punto di vista medico. Per esempio, i linfociti T interver- ranno rapidamente nel rigetto di tessuto estraneo, chirurgicamente innestato o tra- piantato nel corpo. Pertanto le ricerche sul recettore dei linfociti T sono di notevole interesse anche per la chirurgia. V i sono, a dire il vero, due tipi di linfo- cito responsabili del riconoscimento di antigeni specifici e il secondo tipo è co- stituito dai linfociti B. I linfociti sia T sia B sono entrambi derivati dal midollo os- seo, ma i secondi subiscono un ulteriore sviluppo nel timo, una ghiandola che, nella specie umana, è situata immediatamente sotto la parte superiore dello sterno. Tutti e due i tipi circolano nel sangue e nella linfa e sono concentrati nei principali organi lin- fatici, che nei vertebrati superiori sono i linfonodi e la milza. Essi hanno vita lunga e, nella specie umana, possono sopravvi- vere anche molti anni senza dividersi. Pe- rò, come reazione a un antigene, si ingros- sano molto, si dividono rapidamente e se- cernono numerosi fattori proteici che con- tribuiscono alla eliminazione dell'organi- smo invasore o del materiale estraneo. È noto da tempo che la reazione iniziale di un linfocito B a un antigene è mediata da una proteina recettore, esposta sulla su- perficie della membrana cellulare. L'anti- gene si lega al recettore e il legame fa divi- dere e differenziare il linfocito B in un do- ne di plasmacellule. Queste secernono an- ticorpi, che presentano le medesime pro- prietà di legame nei riguardi dell'antigene che ha la molecola recettore inclusa nella membrana superficiale del linfocito B pa- rentale. In effetti, l'anticorpo è identico al recettore del linfocito B al quale l'antigene si è in origine legato, tranne che in esso non si riscontra l'estremità della catena amminoacidica che àncora la proteina re- cettore alla membrana del linfocito B. Sia il recettore dei linfociti B, sia gli anticorpi sono chiamati immunoglobuline. Dopo essere stati secreti nel sangue o nella linfa, gli anticorpi si legano all'anti- gene libero e lo contrassegnano in modo che sia poi distrutto da altri componenti del sistema immunitario. Questo quadro generale della «selezione» che un antigene fa di un linfocito B in base alla capacità di quest'ultimo di dare origine a un clone di cellule e di secernere anticorpi contro quel- l'antigene è detto «teoria della selezione clonale». La teoria è stata sviluppata negli anni sessanta da Sir Macfarlane Burnet del Walter and Eliza Hall Institute for Me- dical Research a Melbourne, da David W. Talmage, allora all'Università di Chicago, e da Niels Kaj Jerne, che lavorava in quel periodo per l'Organizzazione mondiale della sanità. Nel ruolo di recettori dei linfociti B, gli anticorpi sono presenti in piccole quantità. Però, quando un linfocito B viene stimola- to da un antigene, essi compaiono in solu- zione nel siero in notevoli quantità. Sono anche secreti a livelli elevati da certi tipi di tumore dei linfociti B come i plasmocito- mi. La pronta disponibilità di queste ele- vate concentrazioni di anticorpo solubile e il fatto che ogni molecola di anticorpo si possa legare a un antigene hanno permes- so di isolare facilmente l'anticorpo e di ac- quisire notevoli conoscenze sulla struttura del recettore dei linfociti B. Il recettore dei linfociti Tè stato, invece, molto più sfuggente. Il linfocito T, come il linfocito B, reagisce a un antigene dividen- dosi e dando origine a un clone; poi si dif- ferenzia in uno dei numerosi tipi di linfoci- to T, specifico per quell'antigene. I linfoci- ti T citotossici si legano all'antigene virale esposto sulla superficie di una cellula infet- ta e uccidono la cellula. I linfociti T sop- pressori inibiscono, invece, la risposta im- munitaria a un antigene, un certo tempo dopo che la risposta è stata messa in atto. I linfociti T coadiuvanti, infine, si legano all'antigene che è presente sulla superficie di un linfocito B. a sua volta già legato all'antigene. Ogni linfocito T coadiuvante libera quindi molecole ad azione ormono- simile, le linfochine, che permettono al lin- focito B di moltiplicarsi e di differenziarsi. Cosi, per liberare l'enorme potenziale di- struttivo di un linfocito B esiste un sistema a due chiavi: una (l'antigene libero) per il recettore dei linfociti B e l'altra (l'antigene sulla superficie del linfocito B) per il recet- tore dei linfociti T. I linfociti T non si differenziano mai in cellule che secernono anticorpi. Pertanto, diversamente dal recettore dei linfociti B, il loro recettore non è facilmente disponi- bile nelle quantità di composto chimico so- lubile e puro che sono necessarie per svol- gere un'analisi soddisfacente. Essendo gli anticorpi «costruiti» in maniera così raffi- nata ed essendo essi così efficienti nel rico- noscimento degli antigeni, per molti anni si è supposto che i linfociti T si basassero sulle stesse molecole dei linfociti B per le- garsi e per reagire all'antigene. M olti ricercatori hanno speso anni per esaminare le superfici dei linfociti T e i loro secreti alla ricerca di immunoglo- buline. Benché ricerche ad ampio raggio abbiano suggerito che i linfociti Tsoppres- sori possono legarsi all'antigene con mole- cole simili alle immunoglobuline, molti esperimenti hanno provato che, per la maggior parte, i linfociti T non sono asso- ciati a immunoglobuline. Non solo le im- munoglobuline non vengono secrete da questi linfociti, ma esse non si trovano neppure sulle loro membrane o nel loro citoplasma. Altri esperimenti dimostrano che i linfociti T non esprimono l'RNA mes- saggero trascritto dai geni per le immuno- globuline; inoltre quando, nei linfociti T, si esaminano i geni per le immunoglobuline si trova che essi non subiscono riassesta- menti come capita nei linfociti B. Questi risultati negativi si sono accu- mulati negli anni settanta e agli inizi degli anni ottanta. I risultati di esperimenti co- me quelli citati non sono però, di solito, molto persuasivi quando sono presi in con- siderazione singolarmente. Tuttavia, il nu- mero complessivo degli insuccessi ha fatto sì che gli esperti di immunologia moleco- lare cambiassero gradatamente il modo di impostare la ricerca. Se non era possibile trovare delle immunoglobuline associate ai linfociti T, occorreva cercare altrove, in queste cellule, il recettore per l'antigene. Alcuni anni prima, lo studio del trapian- to di tessuti estranei e del rigetto da parte dell'animale ospite aveva portato alle stes- se conclusioni gli esperti di immunologia cellulare: per esempio, le cellule prelevate da un ceppo di topi vanno incontro a un rapido rigetto quando sono iniettate in un topo appartenente a un diverso ceppo ge- netico. A partire dagli inizi degli anni tren- ta, Peter Gorer e altri ricercatori hanno dimostrato che un simile rigetto è provo- cato da molecole di antigene, presenti sulla superficie delle cellule di topo estranee. I linfociti T e il loro recettore Svolgono un compito chiave nel permettere all'organismo di lottare contro le infezioni virali, ma determinano anche il rigetto di un trapianto: base di questo comportamento è una molecola che è stata ormai identificata di Philippa Marrack e John Kappler 28 29
Transcript of I linfociti T e il loro recettore -...

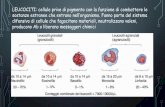
La «istruzione» di un linfocito T nel timo gli conferisce la capacità didistinguere le altre cellule dello stesso organismo da cellule estranee.Questa capacita è un elemento d'importanza critica nella specificità dellarisposta immunitaria. Il timocito, o linfocito T immaturo, che si notanella microfotografia, appare ingrandito 20 900 diametri ed è unito a unacellula epiteliale che fa parte del rivestimento del timo (si veda lo schemaa destra),I siti di legame scuri, presenti sulla superficie del timocito, sonole molecole dei recettori: esse probabilmente si legano a proteine chedefiniscono l'«io» e che sono esposte sulla cellula epiteliale del timo.Queste proteine sono codificate nel genoma da un segmento di DNAchiamato maggior complesso di istocompatibilità ( mHc ). Il legame puòstimolare l'inizio della «istruzione », o maturazione, del timocito. I linfo-citi T maturi acquisiscono una scarsa affinità per le proteine del proprioio, codificate dal MHC, mentre hanno una affinità elevata per queste stesseproteine quando sono associate ad antigeni. I linfociti T innescano unarisposta immunitaria solo contro le cellule infettate del proprio io, oppurecontro cellule appartenenti a un individuo estraneo. La microfotografiaè stata ottenuta da Andrew G. Farr dell'Università di Washington.
CELLULA EPITELIALE DI TIMO
RECETTORI DEL LINFOCITO T
L
e cellule del sistema immunitario per-mettono ai vertebrati di ricono-scere che materiali estranei, gli
antigeni, hanno invaso il loro organismo.La risposta immunitaria che ne consegueè notevole per la sua specificità. Per esem-pio, un essere umano immunizzato me-diante vaccinazione contro il vaiolo puòopporre resistenza a un'infezione provoca-ta da virus del vaiolo, ma non, per esem-pio, a un attacco di virus dell'influenza.Nell'ultimo decennio gli immunologi sonoriusciti a riconoscere che il fattore più im-portante nella capacità del sistema immu-nitario di reagire specificamente ai virus èrappresentato da una classe di cellule dimodeste dimensioni, i linfociti T(chiamatianche cellule T). I linfociti T, infatti, svol-gono un ruolo ausiliare essenziale nella ri-sposta che il sistema immunitario dà all'in-fezione batterica.
Da lungo tempo è stata accettata l'ideache un'attività di questo tipo venga inne-scata da una molecola inclusa nella mem-brana dei linfociti Te chiamata «recettoredei linfociti T». Si presume che un antigenespecifico si adatti e si leghi a questo recet-tore come una chiave si adatta alla giustaserratura, mettendo quindi in moto la seriecomplessa di eventi biochimici che costi-tuiscono la risposta immunitaria. Per tuttauna serie di ragioni, l'isolamento del recet-tore dei linfociti T è risultato estremamen-te difficile e, fino a non molto tempo fa, èstato necessario dedurne le proprietà pervia indiretta, senza essere guidati dalla co-noscenza della sua struttura, che oggi staper essere messa a fuoco.
Il chiarimento sulla struttura del recet-tore dei linfociti T porta con sé anche unamigliore comprensione delle complesse in-terazioni che si instaurano tra linfociti T ealtri elementi del sistema immunitario. Inparticolare, risulta sempre più evidenteche il linfocito T è particolarmente adattoa fronteggiare le infezioni associate allecellule dell'ospite, e non le infezioni checircolano liberamente nei liquidi corporeidell'ospite. Per svolgere questo compito, ilrecettore dei linfociti T deve non solo rico-
noscere un antigene specifico, ma che an-che certe proteine di membrana della stes-sa cellula ospite. Un simile meccanismo diriconoscimento deve essere tenuto sotto ri-goroso controllo perché un linfocito T, sedovesse essere attivato soltanto dalle pro-teine dell'ospite, potrebbe facilmente rivol-gersi contro le cellule sane di quest'ultimo.La conseguente sensibilità del sistema diriconoscimento dei linfociti T è responsa-bile di molte affascinanti proprietà di que-sto sistema, importanti dal punto di vistamedico. Per esempio, i linfociti T interver-ranno rapidamente nel rigetto di tessutoestraneo, chirurgicamente innestato o tra-piantato nel corpo. Pertanto le ricerche sulrecettore dei linfociti T sono di notevoleinteresse anche per la chirurgia.
Vi sono, a dire il vero, due tipi di linfo-cito responsabili del riconoscimento
di antigeni specifici e il secondo tipo è co-stituito dai linfociti B. I linfociti sia T siaB sono entrambi derivati dal midollo os-seo, ma i secondi subiscono un ulterioresviluppo nel timo, una ghiandola che, nellaspecie umana, è situata immediatamentesotto la parte superiore dello sterno. Tuttie due i tipi circolano nel sangue e nella linfae sono concentrati nei principali organi lin-fatici, che nei vertebrati superiori sono ilinfonodi e la milza. Essi hanno vita lungae, nella specie umana, possono sopravvi-vere anche molti anni senza dividersi. Pe-rò, come reazione a un antigene, si ingros-sano molto, si dividono rapidamente e se-cernono numerosi fattori proteici che con-tribuiscono alla eliminazione dell'organi-smo invasore o del materiale estraneo.
È noto da tempo che la reazione inizialedi un linfocito B a un antigene è mediatada una proteina recettore, esposta sulla su-perficie della membrana cellulare. L'anti-gene si lega al recettore e il legame fa divi-dere e differenziare il linfocito B in un do-ne di plasmacellule. Queste secernono an-ticorpi, che presentano le medesime pro-prietà di legame nei riguardi dell'antigeneche ha la molecola recettore inclusa nellamembrana superficiale del linfocito B pa-
rentale. In effetti, l'anticorpo è identico alrecettore del linfocito B al quale l'antigenesi è in origine legato, tranne che in essonon si riscontra l'estremità della catenaamminoacidica che àncora la proteina re-cettore alla membrana del linfocito B. Siail recettore dei linfociti B, sia gli anticorpisono chiamati immunoglobuline.
Dopo essere stati secreti nel sangue onella linfa, gli anticorpi si legano all'anti-gene libero e lo contrassegnano in modoche sia poi distrutto da altri componentidel sistema immunitario. Questo quadrogenerale della «selezione» che un antigenefa di un linfocito B in base alla capacità diquest'ultimo di dare origine a un clone dicellule e di secernere anticorpi contro quel-l'antigene è detto «teoria della selezioneclonale». La teoria è stata sviluppata neglianni sessanta da Sir Macfarlane Burnetdel Walter and Eliza Hall Institute for Me-dical Research a Melbourne, da David W.Talmage, allora all'Università di Chicago,e da Niels Kaj Jerne, che lavorava in quelperiodo per l'Organizzazione mondialedella sanità.
Nel ruolo di recettori dei linfociti B, glianticorpi sono presenti in piccole quantità.Però, quando un linfocito B viene stimola-to da un antigene, essi compaiono in solu-zione nel siero in notevoli quantità. Sonoanche secreti a livelli elevati da certi tipi ditumore dei linfociti B come i plasmocito-mi. La pronta disponibilità di queste ele-vate concentrazioni di anticorpo solubile eil fatto che ogni molecola di anticorpo sipossa legare a un antigene hanno permes-so di isolare facilmente l'anticorpo e di ac-quisire notevoli conoscenze sulla strutturadel recettore dei linfociti B.
Il recettore dei linfociti Tè stato, invece,molto più sfuggente. Il linfocito T, come illinfocito B, reagisce a un antigene dividen-dosi e dando origine a un clone; poi si dif-ferenzia in uno dei numerosi tipi di linfoci-to T, specifico per quell'antigene. I linfoci-ti T citotossici si legano all'antigene viraleesposto sulla superficie di una cellula infet-ta e uccidono la cellula. I linfociti T sop-pressori inibiscono, invece, la risposta im-
munitaria a un antigene, un certo tempodopo che la risposta è stata messa in atto.I linfociti T coadiuvanti, infine, si leganoall'antigene che è presente sulla superficiedi un linfocito B. a sua volta già legatoall'antigene. Ogni linfocito T coadiuvantelibera quindi molecole ad azione ormono-simile, le linfochine, che permettono al lin-focito B di moltiplicarsi e di differenziarsi.Cosi, per liberare l'enorme potenziale di-struttivo di un linfocito B esiste un sistemaa due chiavi: una (l'antigene libero) per ilrecettore dei linfociti B e l'altra (l'antigenesulla superficie del linfocito B) per il recet-tore dei linfociti T.
I linfociti T non si differenziano mai incellule che secernono anticorpi. Pertanto,diversamente dal recettore dei linfociti B,il loro recettore non è facilmente disponi-bile nelle quantità di composto chimico so-lubile e puro che sono necessarie per svol-gere un'analisi soddisfacente. Essendo glianticorpi «costruiti» in maniera così raffi-nata ed essendo essi così efficienti nel rico-noscimento degli antigeni, per molti anni
si è supposto che i linfociti T si basasserosulle stesse molecole dei linfociti B per le-garsi e per reagire all'antigene.
Molti ricercatori hanno speso anni peresaminare le superfici dei linfociti T
e i loro secreti alla ricerca di immunoglo-buline. Benché ricerche ad ampio raggioabbiano suggerito che i linfociti Tsoppres-sori possono legarsi all'antigene con mole-cole simili alle immunoglobuline, moltiesperimenti hanno provato che, per lamaggior parte, i linfociti T non sono asso-ciati a immunoglobuline. Non solo le im-munoglobuline non vengono secrete daquesti linfociti, ma esse non si trovanoneppure sulle loro membrane o nel lorocitoplasma. Altri esperimenti dimostranoche i linfociti T non esprimono l'RNA mes-saggero trascritto dai geni per le immuno-globuline; inoltre quando, nei linfociti T, siesaminano i geni per le immunoglobulinesi trova che essi non subiscono riassesta-menti come capita nei linfociti B.
Questi risultati negativi si sono accu-
mulati negli anni settanta e agli inizi deglianni ottanta. I risultati di esperimenti co-me quelli citati non sono però, di solito,molto persuasivi quando sono presi in con-siderazione singolarmente. Tuttavia, il nu-mero complessivo degli insuccessi ha fattosì che gli esperti di immunologia moleco-lare cambiassero gradatamente il modo diimpostare la ricerca. Se non era possibiletrovare delle immunoglobuline associateai linfociti T, occorreva cercare altrove, inqueste cellule, il recettore per l'antigene.
Alcuni anni prima, lo studio del trapian-to di tessuti estranei e del rigetto da partedell'animale ospite aveva portato alle stes-se conclusioni gli esperti di immunologiacellulare: per esempio, le cellule prelevateda un ceppo di topi vanno incontro a unrapido rigetto quando sono iniettate in untopo appartenente a un diverso ceppo ge-netico. A partire dagli inizi degli anni tren-ta, Peter Gorer e altri ricercatori hannodimostrato che un simile rigetto è provo-cato da molecole di antigene, presenti sullasuperficie delle cellule di topo estranee.
I linfociti T e il loro recettoreSvolgono un compito chiave nel permettere all'organismo di lottare controle infezioni virali, ma determinano anche il rigetto di un trapianto: basedi questo comportamento è una molecola che è stata ormai identificata
di Philippa Marrack e John Kappler
28 29

LINFOCITO TCITOTOSSICO
v MATURO
11 — —CELLULA MEMORIA T CELLULA MEMORIA T
CITOTOSSICA COADIUVANTE CELLULA MEMORIA B
20\
Iths—
ANTICORPI
ATTIVAZIONE DEI LINFOCITI T CITOTOSSICI
MACROFAGO '127 11.
1
ANTIGENE
117
PROTEINACODIFICATA LDAL MHC E DI CLASSE I
RECETTOREDI LINFOCITO T
SPECIFICO PER L'ANTIGENEE RISTRETTO ALLA CLASSE I
4a
ANTIGENE -TRASFORMATO
LINFOCITI TCITOTOSSICIIMMATURI
7a
CELLULA
8a
INTERLEUCHINE5a
INTERLEUCHINA-1
6a
1 b
2bPROTEINACODIFICATA DAL MHCE DI GLASSE II
'ROTEINAODIFICATA
)AL MHC
RECETTOREI LINFOCITO B
SPECIFICOPER
L'ANTIGENE
ANTIGENE
4 C
LINFOCITI B
\è/
LINFOCITI TCOADIUVANTI IMMATURI
X6b
. „
INTERLEUCHINEANTIGENE TRASFORMATO
8b
LINFOCITO TCOADIUVANTEMATURO
17
RECETTOREDI LINFOCITO T
SPECIFICO PER L'ANTIGENEE RISTRETTO ALLA CLASSE II
5h
13
ATTIVAZIONE DEI LINFOCITI T COADIUVANTI ATTIVAZIONE DEI LINFOCITI BPER PRODURRE ANTICORPI
MACROFAGOINFETTATO
PLASMACELLULA
Le proteine che caratterizzano ogni cel-lula come «propria» o come «estranea» so-no codificate da geni associati e localizzatiin una regione del DNA chiamata il mag-gior complesso di istocompatibilità (mHcdall'inglese major histocompatibility com-plex). Esse sono note come «proteine codi-ficate dal MHC», o semplicemente «protei-ne MHC». Una delle loro proprietà più sor-prendenti è l'estremo polimorfismo: vi so-no milioni di alleli, o varianti, dei geni delMHC, che codificano per ogni proteina;pertanto, la probabilità che due individuinon affini abbiano identiche proteine MHCè davvero scarsa.
Grazie alla ricerca di Gorer, è risultatosempre più chiaro che sia il rigetto dei tes-suti trapiantati sia l'esistenza di proteinecodificate dal mHC sono in stretta relazio-ne con la risposta immunitaria. Sir PeterMedawar e altri ricercatori hanno dimo-strato che responsabili del riconoscimentodi molecole antigeniche su un tessuto e-straneo trapiantato sono i linfociti; ricer-che successive hanno indicato che fonda-mentali nel rigetto di tessuti sono i linfoci-ti T, e non i linfociti B. Tuttavia, i trapiantinon sono esperimenti della natura, e gliimmunologi avevano quindi ancora dubbisul ruolo delle proteine codificate dal MHC.
Alcuni indizi sulle funzioni svolte dalleproteine MHC cominciarono a emergere al-la metà degli anni sessanta. Hugh O.McDevitt, allora al National Institute forMedical Research, in Inghilterra, assiemea Michael Sela del Weizmann Institute ofScience, in Israele, e Baruj Benacerraf, al-lora al National Institute of Allergy andInfectious Diseases, con i suoi collabora-tori hanno studiato la risposta di vari ceppidi animali agli antigeni e hanno trovatoche quella di un topo a certi antigeni sin-tetici dipende dal ceppo genetico a cui quel
topo appartiene. Per esempio, quando ilpolimero TGAL viene iniettato nei topi,quelli che possiedono alleli b nel loro MHCproducono anticorpi anti-TGAL, mentrequelli che possiedono alleli k no. È risulta-to così subito evidente che gli aneli in que-stione agiscono sulle funzioni dei linfoci-ti T e non su qualche altro tipo cellularedel sistema immunitario.
A questo punto alcuni immunologi han-no suggerito l'idea che il recettore dei lin-fociti T potesse essere codificato dal MHC.Se così fosse davvero, ci si aspetterebbeche differenti tipi di mLiC codifichino perdifferenti recettori dei linfociti T e quest'ul-timo punto spiegherebbe le differenze rile-vate da un ceppo di topi all'altro nella ri-sposta al TGAL. La teoria ha però dovutoessere scartata quando si è scoperto chealcune delle proteine codificate dal mFiC,che influiscono sul legame del TGAL, nonsono neppure espresse alla superficie deilinfociti T di topo. Pertanto, la fonte delledifferenze osservate non poteva essere so-lo il recettore dei linfociti T ed era inutilecontinuare a supporre che esso fosse codi-ficato dal MHC. Invece, gli esperimentihanno fornito una conferma a una questio-ne più sottile: le differenze rilevate nellarisposta immunitaria al TGAL (produzionedi anticorpi specifici) indicavano che leproteine codificate dal MHC influiscono sulmodo in cui quel polimero viene ricono-sciuto dal recettore del linfocito T. Questorisultato è stato il primo indizio che il pre-supposto per l'attività del recettore fosseun'interazione tra antigeni estranei e pro-teine codificate dal MHC.
Come poter spiegare questa strana col-lezione di risultati? Le risposte sono
state date dalle ricerche pionieristiche diEthan M. Shevach e Alan S. Rosenthal del
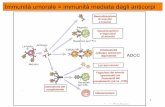
Esistono tre tipi principali di risposta del sistema immunitario, specifica nei riguardi dell'antigene:la risposta di un linfocito T citotossico (a sinistra), quella di un linfocito T coadiuvante (al centro)e quella di un linfocito B (a destra). L'antigene che penetra nel corpo (in alto) viene fagocitatoda un macrofago ( I a, 1b), trasformato in frammenti proteici più corti ( 2a, 2b) ed esposto in formaparzialmente digerita alla sua superficie (3a, 3b). In questa sede esso si lega a una delle due classidi proteine codificate dal MHC. Un linfocito T, il cui recettore si adatta al complesso proteina-an-tigene, si lega a quest'ultimo e, pertanto, è «selezionato» per la propagazione clonale dal complessoproteina-antigene (4a, 4b). Analogamente, l'antigene libero seleziona un linfocito B, il cui recettoresi adatta a esso e i due si legano (4c). I linfociti T destinati a diventare citotossici si leganoall'antigene associato con proteine codificate dal mHc e di classe I (.5a), mentre i futuri linfociti Tcoadiuvanti si legano all'antigene associato con proteine di classe!! (5b). Il legame con il linfoci-to T stimola la liberazione da parte del macrofago della interleuchina- 1, una molecola ormonosi-mile; il macrofago, a sua volta, stimola il linfocito T a dividersi e a differenziarsi (6a, 6b). Ladivisione cellulare prosegue fino a quando il linfocito T è stimolato da cellule che espongonoantigeni (7a, 7b). Un linfocito T maturo (8a, 8b) può allora svolgere la sua funzione. Se ècitotossico, può o legarsi a una cellula infetta che espone l'antigene ( 9) e ucciderla (10), oppurerimanere in circolo nel sangue o nella linfa come cellula memoria, la quale agisce più prontamentecon qualsiasi antigene dello stesso tipo che, in futuro, entrerà in contatto con essa (11). Ancheun linfocito T coadiuvante maturo può diventare una cellula memoria. La funzione del linfocito Tcoadiuvante consiste nello stimolare la proliferazione di linfociti B attivati. Un linfocito B, che hainglobato il suo antigene legato (12) e Io ha trasformato (13), presenta anche in superficie unframmento di antigene legato a una proteina codificata dal mHC e di classe 11(14). Il linfocito Tcoadiuvante maturo può legarsi a questo punto con il complesso antigene-proteina, presente sullinfocito B (15). Questo legame libera le interleuchine, che permettono al linfocito B di dividersie di differenziarsi ( 16); la divisione cellulare prosegue fintantoché il linfocito B viene stimolato dailinfociti T coadiuvanti ( 17). Le plasmacellule mature ( 18) liberano quindi i loro recettori, specificiper gli antigeni, come anticorpi e questi si legano agli antigeni liberi distruggendoli (19). Altrilinfociti T maturi, presenti nel clone, rimangono in circolo come cellule memoria (20). Lo schemainclude i risultati degli esperimenti descritti dagli autori in questo articolo assieme ai risultati dimolti altri esperimenti, ma certi particolari del modello sono ancora oggetto di vivaci controversie.
30 31

TOPO DI CEPPO K VIRUSDELLA CORIOMENINGITELINFOCITARIA
RECETTOREDEL LINFOCITO T
—411-
PROTEINA DI CEPPO B, VIRUSCODIFICATA DAL MHC DELLA CORIOMENINGITELINF CITARIA
LINFOCITO T CITOTOSSICOISOLATO,SPECIFICO PER IL VIRUSDELLA CORIOMENINGITELINFOCITARIA
TOPO DI CEPPO B
VIRUSDELLA CORIOMENINGITELINFOCITARIA
CELLULADI CEPPO BISOLATA
LA CELLULA BERSAGLIO
LA CELLULA BERSAGLIOE UCCISA
NON E UCCISA
CELLULA BERSAGLIO
1111
VIRUSDELLA CORIOMENINGITELINFOCITARIATRASFORMATO
LINFOCITCITOTOSSICO
PROTEINADI CEPPO B,
CODIFICATADAL MHC
LINFOCITOCITOTOSSICO
CELLULA BERSAGLIO
PROTEINADI CEPPO K,CODIFICATA DAL MHC
RECETTOREDEL LINFOCITO T
LINFOCITOCITOTOSSICO
LA CELLULA BERSAGLIONON E UCCISA
Un linfocito T reagisce a un antigene solo se legato a una proteinacodificata dal MHC, esposta su una cellula bersaglio. Linfociti T di topodi ceppo k, immunizzato contro il virus della coriomeningite linfocitaria,possono uccidere le cellule che espongono proteine di ceppo k, codificate
dal MHC, dopo che sono state infettate dal virus (al centro). Non ucci-dono, invece, le cellule infettate dal virus che non espongono le proteine(a destra) e neppure le cellule di topo che espongono le proteine diceppo k, codificate dal mnc, ma non sono infettate dal virus (a sinistra).
TOPO DI CEPPO K
M.
CELLULADI CEPPO KISOLATA-.....
W,.. PROTEINADI CEPPO K.
\l/ ' CODIFICATA DAL MHC
National Institute of Allergy and Infec-tious Diseases, di David H. Katz e Bena-cerraf della Harvard Medical School, diBernice Kindred dell'Università di Co-stanza nella Repubblica Federale Tede-sca, di Donald Shreffier della School ofMedicine dell'Università del Michigan e,in modo estremamente chiaro, dagli im-portanti esperimenti di Rolf Zinkernagel ePeter C. Doherty della Australian Natio-nal University.
Nel corso di un esperimento, Zinkerna-gel e Doherty hanno iniettato dosi subletalidel virus della cmt, (coriomeningite linfo-citaria) in topi e hanno isolato linfociti Tcitotossici, specifici per quel virus (si vedal'illustrazione di questa pagina), dagli ani-mali immunizzati. La funzione abituale diquesti linfociti consiste nel riconoscere gliantigeni virali sulle cellule infettate e neldistruggere queste ultime. La maggior par-te dei ceppi di topo produce anticorpi con-tro il virus della CML e, pertanto, vi eranoscarse probabilità che quel virus non riu-scisse a indurre in alcuni ceppi di topo unarisposta immunitaria simile a quella scate-nata dal TGAL. Malgrado questo fatto,Zinkernagel e Doherty hanno trovato conloro sorpresa che i linfociti Tdi topi appar-tenenti al ceppo k e immunizzati contro laCmL sono in grado di uccidere le celluleinfettate dal virus della cm". solo se esse
contengono almeno uno dei numerosi tipidi proteine codificate dal mFIC e caratteri-stiche del ceppo k. In altre parole, i linfo-citi T, letali per le cellule infettate che con-tenevano queste proteine, non potevanouccidere cellule di topi affini, infettati conlo stesso virus, ma dotati, per esempio, diproteine codificate dal MHC e caratteristi-che del ceppo d.
Questo esperimento e altri ancora han-no dimostrato che i linfociti T citotossicierano interessati non solo agli antigeni vi-rali delle cellule infettate, ma anche allesequenze amminoacidiche delle proteinecodificate dal MHC e presenti su quelle cel-lule. Altri ricercatori hanno dimostratopoco dopo che la stessa cosa avveniva coni linfociti T citotossici, che avevano unaspecificità per altri antigeni. Il fenomeno èoggi definito «restrizione da MHC». In ge-nerale, le proteine codificate dal MHC e ri-conosciute dai linfociti T citotossici appar-tengono a una singola classe strutturale, leproteine della classe I, che si trovano sullasuperficie di tutte le cellule nucleate del-l'organismo.
Noi e diversi altri ricercatori abbiamoquindi trovato che anche i linfociti T coa-diuvanti presentano questa restrizione daMHC; in generale, però, un linfocito T coa-diuvante riconosce una differente classe diproteine codificate dal MHC, le proteine di
classe II. L'espressione di queste proteineè notevolmente meno diffusa nell'organi-smo dell'espressione delle proteine dellaclasse I: negli esseri umani, le proteine del-la classe II si trovano soltanto nei linfoci-ti T e B, nei macrofagi e in certe celluleappartenenti ad altri tessuti.
Non appena è stata scoperta la restri-zione riguardo all'mxc, è risultato
chiaro che i fatti potevano essere spiegatida due tipi di teoria. La teoria del ricono-scimento associato suggerisce nella suaforma estrema che ogni linfocito T portarecettori di un unico tipo. Ciascuno di que-sti recettori si legherebbe in qualche modoa un complesso tra l'antigene e una parti-colare proteina codificata dal MHC, checompare sulla superficie della cellula cheespone quell'antigene. Per contro, la teoriadel duplice riconoscimento suggerisce nel-la sua forma estrema che ogni linfocito Tpresenta due tipi di recettore, uno dei qualisi legherebbe a un antigene specifico e l'al-tro a una proteina specifica, codificata dalMHC.
Ogni tipo di teoria presenta vantaggi esvantaggi. La teoria del riconoscimentoassociato spiega l'osservazione in base al-la quale i linfociti T si legano solo rara-mente o all'antigene o alle proteine codifi-cate dal MHC senza legarsi contempora-
neamente a entrambe le molecole. La teo-ria spiega anche bene i risultati ottenuti daMcDevitt, Sela e Benacerraf sulle proprie-tà immunogene del TGAL nei topi. Se unsingolo recettore di linfocito T deve simul-taneamente legare l'antigene e la proteinacodificata dal MHC, l'incapacità dei topi diceppo k di reagire al TGAL potrebbe esseredovuta al fatto che l'antigene TGAL nonriesce a formare un complesso con la pro-teina codificata dal milc e di ceppo k. Dalcanto suo, la teoria del duplice riconosci-mento non presenta uno svantaggio che siriscontra nella teoria del riconoscimentoassociato: cioè essa non deve postulareun'interazione tra ogni forma di antigenee le forme invarianti della proteina codifi-cata dal MHC che si trovano in ogni singo-lo animale.
Benché l'ultima parola non sia stata an-cora detta, è probabilmente giusto affer-mare che i dati oggi disponibili sono infavore della teoria del riconoscimento as-sociato, con il suo modello di un unicorecettore del linfocito Tper l'antigene e perla proteina codificata dal MHC. Per esem-pio, abbiamo studiato le proprietà di linfo-citi T ibridi che esprimono recettori deri-vati da due differenti linfociti T parentali(si veda l'illustrazione a pagina 34). Ab-biamo dapprima isolato dei linfociti T coa-diuvanti, che riconoscono in maniera spe-cifica un antigene, l'ovalburnina di pollo,quando è associato a certe proteine di clas-se II, tipiche del ceppo k, e li abbiamo fusicon un tumore costituito da linfocili T, co-si da ottenere un ibridoma di linfociti T,cioè una cellula ibrida moltiplicatasi rapi-damente e che, in coltura, ha dato originein poco tempo a doni.
Per ottenere una cellula con due diffe-renti insiemi di recettori di linfociti T, ab-biamo fuso una delle cellule del nostro ibri-doma di linfociti Tcon un secondo insiemedi linfociti T, specifici per un certo antige-ne. Questo secondo insieme poteva reagiresoltanto all'antigene costituito dall'emo-cianina del mollusco Fissurella in presen-za di proteine di classe II e di ceppo f.
Abbiamo analizzato la risposta di linfo-citi T appena fusi agli antigeni ovalbuminadi pollo ed emocianina di Fissurella, asso-ciati a cellule che espongono gli antigeni eche appartengono o al ceppo k o al cep-pof. Se la teoria del duplice riconoscimen-to fosse giusta, i linfociti T appena fusidovrebbero presumibilmente portare quat-tro tipi di recettore, uno per ogni antigenee uno per ogni proteina codificata dal MHCe appartenente alla classe II. Pertanto essidovrebbero reagire a entrambi gli antigeniesposti su un linfocito di ceppo k e a en-trambi gli antigeni esposti su un linfocitodi ceppo f. Abbiamo trovato, invece, cheessi reagiscono agli antigeni solo comehanno fatto in precedenza i loro linfocitiparentali: all'antigene ovalbumina di pol-lo, associato con linfociti di ceppo k, e al-l'antigene emoglobina di Fissurella, asso-ciato con linfociti di ceppo f Non vi eraalcun cenno di reazione all'ovalbumina dipollo nei linfociti di ceppo f o all'emoglo-bina di Fissurella nei linfociti di ceppo k.
Se la teoria del riconoscimento associato è giusta, l'antigene e la proteina codi-
ficata dal MHC devono in qualche modoformare un complesso prima che un linfo-cito T possa legarsi a essi. I risultati didiversi esperimenti suggeriscono che tra leproteine codificate dal MHC e gli antigeniesista un qualche tipo di interazione; tut-tavia solo pochi esperimenti sono riuscitia rivelarla direttamente. Forse la dimo-strazione più sorprendente negli ultimitempi è quella offerta dalle ricerche di EmilR. Unanue e collaboratori della School ofMedicine della Washington University.
Unanue e Collaboratori hanno identifi-cato un piccolo antigene che può esserericonosciuto dai linfociti T quando è asso-ciato con proteine codificate dal MHC eappartenenti al ceppo k, ma non quando èassociato a proteine del ceppo d. Essi han-no posto uguali concentrazioni dell'antige-ne in soluzione su entrambi i lati di unamembrana semipermeabile e hanno ag-giunto da un solo lato proteine codificatedal MHC e appartenenti o all'uno o all'altroceppo. Dato che le molecole antigenichesono molto più piccole di entrambi i tipi diproteina, la membrana permetteva il liberopassaggio degli antigeni, ma confinava leproteine nel loro compartimento iniziale.
Unanue e il suo gruppo hanno trovatoche, quando le proteine codificate dal MHCe appartenenti al ceppo k venivano aggiun-te a un compartimento e la concentrazionedegli antigeni veniva poi lasciata equilibra-re, gli antigeni apparivano in maggiorequantità nel compartimento che contene-va le proteine del ceppo k. Le concentra-zioni dell'antigene nei due compartimentinon si modificavano quando, a un com-partimento, venivano aggiunte proteinecodificate dal MHC e appartenenti al cep-po d. Il risultato suggerisce che l'antigenesi leghi, almeno qualche volta, alla protei-na codificata dal mFiC assieme alla quale èriconosciuto, anche se il processo può av-venire con un'efficienza non elevata.
Anche dopo che erano state stabilitetutte le proprietà del recettore dei linfoci-ti T, citate sopra, l'identità della proteinaresponsabile dell'attività del recettore ri-maneva ancora avvolta dal mistero. Tut-tavia, fu chiaro da quel momento che ilrecettore dei linfociti T differisce per alme-no un aspetto importante dalla molecoladell'anticorpo. Indipendentemente dal fat-to che esso sia un'unica proteina distinta odue, perlomeno qualche sua componenteha una forte tendenza a riconoscere partedi una proteina codificata dal MHC. Lamolecola dell'anticorpo no.
Vi sono stati due importanti progressitecnologici che hanno permesso di comin-ciare a intravvedere la struttura del recet-tore dei linfociti T. Uno è stata la scopertaeffettuata agli inizi degli anni settanta daGeorge Kóhler e Cesar Milstein del Labo-ratory of Molecular Biology del MedicalResearch Council a Cambridge, i qualihanno trovato un modo per produrre ibri-domi di linfociti B, che possono secernerein vitro grandi quantità di tipi di anticorposelezionati. Questi anticorpi sono definiti
monoclonali e possono essere prodotti epurificati in enormi quantità. Il secondoprogresso è consistito nella elaborazionedi metodi grazie ai quali doni di linfociti To ibridomi di linfociti T con specificità perdeterminati antigeni e proteine codificatedal MHC potevano essere propagati in col-tura. Noi e numerosi altri ricercatori ab-biamo sfruttato entrambe queste conqui-ste per produrre anticorpi contro il recet-tore dei linfociti T e quindi per identificarele proprietà strutturali di questo recettore.
per produrre anticorpi contro il recetto-re dei linfociti T abbiamo innanzitutto
«costruito» un ibridoma di linfociti T cheportavano un recettore per il riconosci-mento dell'ovalbumina di pollo, antigeneassociato a una proteina di classe II, ap-partenente al ceppo d. I recettori presentiin questo tipo di ibridomi, quando sonolegati con il giusto antigene e la giusta pro-teina codificata dal MHC, danno una rapi-da risposta, secernendo linfochine. Gliibridomi sono utili dal punto di vista spe-rimentale perché la produzione di linfochi-ne può essere indotta e misurata rapida-mente in coltura.
Per ottenere un anticorpo di specificitànota si deve immunizzare un animale con-tro un antigene noto. Nel caso del recetto-re dei linfociti T la precisa identità dell'an-tigene non era, tuttavia, conosciuta. Lastrategia della nostra ricerca è consistitanell'immunizzare topi contro ibridomi dilinfociti T, nella speranza di indurre la pro-duzione di anticorpi che interferissero solocon le funzioni presumibilmente svolte dairecettori su quegli ibridomi. Abbiamo pen-sato che un anticorpo contro il recettore sisarebbe legato a quest'ultimo bloccandonela capacità di fissarsi all'ovalbumina dipollo e alla proteina di ceppo d, codificatadal MHC. Una diminuzione nella produzio-ne di linfochine indicherebbe il blocco.
Abbiamo immunizzato molti topi con-tro un numero elevato di ibridomi di linfo-citi T; quindi abbiamo prelevato il siero daogni topo, in tempi diversi a partire dal-l'immunizzazione, e lo abbiamo analizza-to in coltura allo scopo di valutare la suacapacità di bloccare la reazione degli ibri-domi di linfociti T alle proteine di ceppo de all'ovalbumina di pollo. Alla fine abbia-mo identificato molti topi che produceva-no l'anticorpo bloccante. L'antisiero pre-levato da questi animali aveva un'ulterioreproprietà incoraggiante: esso non blocca-va la risposta dei linfociti T quando ibri-domi di questi linfociti con differenti spe-cificità venivano posti, in coltura, a con-fronto con l'antigene e con la proteina co-dificata dal MHC, nei riguardi dei quali era-no specifici. Abbiamo ripetuto l'esperi-mento con altri ibridomi di linfociti T nelruolo dell'ibridoma originario. In ogni ca-so gli anticorpi bloccanti prodotti dai topierano efficaci solo contro ribridoma di lin-fociti T utilizzato per l'immunizzazione.
La specificità degli antisieri ottenuti perun particolare ibridoma di linfociti T ci hafatto ritenere che eravamo davvero riuscitia produrre anticorpi contro un particolare
32 33

TEORIA DEL RICONOSCIMENTO ASSOCIATO
RECETTORE PERLA PROTEINADI CEPPO F E PERL'EMOCIANINADI FISSURELLA
CELLULA CHEESPONE L'ANTIGENE
PROTEINA DI CEPPO F,CODIFICATA DAL MHC
LINFOCITO
EMOCIANINAjae
FISSURELLA/r
LINFOCITO
, T IBRIDO
OVALBUMINA_ __5#4DI POLLO
PROTEINA DI CEPPO K.CODIFICATA DAL MHC
RECETTORE PERLA PROTEINADI CEPPO K E PERL'OVALBUMINADI POLLO
--"\gamil(CEPPO K
/Li 4t%N- ADI POLLO
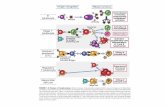
Due teorie tra loro incompatibili possono però spiegare il riconoscimento sia dell'antigene sia delleproteine codificate dal mHc da parte dei linfociti T. Secondo la teoria del riconoscimento associato,ogni linfocito T porta un recettore, che può legarsi a un complesso di un antigene con una proteinacodificata dal ADiC (in alto a sinistra). Secondo la teoria del duplice riconoscimento, invece, ognilinfocito T ha due recettori, uno per l'antigene e uno per la proteina codificata dal mliC (in bassoa sinistra). Per comprovare le due teorie, gli autori hanno «costruito» un linfocito T ibrido fon-dendo due linfociti T normali. Un linfocito T parentale è stato isolato da animali con proteine diceppo f, codificate dal MHC, dopo che erano stati immunizzati contro l'emocianina di Fissurella.Un secondo linfocito T parentale è stato isolato, invece, da animali di ceppo k, immunizzati control'ovalbumina di pollo. Secondo una versione estrema della prima teoria, il linfocito ibrido dovrebbericonoscere l'emocianina di Fissurella solo su cellule di ceppo! e l'ovalbumina di pollo solo sucellule di ceppo k (in alto a destra). Secondo una versione estrema della seconda teoria, l'ibridodovrebbe riconoscere, invece, i due antigeni sulle cellule di entrambi i ceppi (in basso a destra). Illinfocito ibrido si è comportato proprio conformemente a quanto sostiene la prima teoria.
(CEPPO F
OVAL-BU MINADI POLLO
mi CEPPO K
EMO-CIANINADI FISSU-RELLA j
CEPPO F
EMO-CIAN I NADI FISSU-RELLA
LCEPPO K
OVAL-BUMINA
POLLO
CEPPO F
TCI ANI NADI
/su.RELLA
TEORIA DEL DUPLICE RICONOSCIMENTO
RECETTOREPER LA PROTEINA?2DI CEPPO F
RECETTOREPER L'ENOCIANINADI FISSURELLA
,
i / flo4. LINFOCITO i
T IBRIDO
\ -----11k jRECETTOREPER L'OVALBUMINADI POLLO
1(14 /
PER LA PROTEIN*RECETTORE
DI CEPPO K
CATENE LEGGERE
REGIONE COSTANTE
REGIONE VARIABILE
REGIONE DI CONGIUNZIONE
REGIONE RESPONSABILE DELLA DIVERSITÀ
MICROGLOBULINA BETA-2
CATENE PESANTICATENA ALFA
CATENA ALFA I I CATENA BETA
CATENA BETA
LEGAMEDISOLFURO
PROTEINA CODIFICATADAL MHC E DI CLASSE II
PROTEINA CODIFICATADAL MHC E DI CLASSE I
1MUN
CATENA ALFA
MICROGLOBULINABETA-2
RECETTORE DI LINFOCITO T
n nn aCITOPLASMA
IMMUNOGLOBULINA M
[1 CELLULAREMEMBRANA
Le strutture molecolari della proteina di classe I e di quella di classe II,codificate dal MHC, e inoltre del recettore del linfocito T edi una immu-noglobulina o anticorpo, sono simili; esse hanno anche in comune alcunesequenze di amminoacidi e sono caratterizzate da anse costituite, all'in-terno di ogni catena, da circa 70 amminoacidi; gli atomi di zolfo, a ogniestremità delle anse, sono uniti da legami covalenti. Le proteine di clas-se I, nei vertebrati superiori, sono espresse alla superficie di ogni cellulanucleata in associazione con la beta-2-microglobulina, una proteina non
codificata dal mitc. Le proteine di classe II sono espresse, invece, solosulla superficie di cellule selezionate, come i linfociti B. Nel recettore dellinfocito T e nella immunoglobulina sono state identificate una regionevariabile, una di congiunzione e una responsabile della diversità, ognunacodificata da una sequenza di DNA, selezionata a caso in mezzo a parec-chie sequenze presenti nel genoma. Queste regioni danno origine a unasostanziale variabilità combinatoriale nelle molecole espresse da differen-ti cellule. Gli schemi, molto semplificati, non sono disegnati in scala.
recettore dei linfociti T. Questo recettore èl'unica struttura sulla superficie dell'ibri-doma di linfociti T, che presumibilmentesarebbe bloccato da un anticorpo in unmodo cosi specifico, perché soltanto essovarierebbe da un clone di linfociti T a unaltro. Cionondimeno il nostro compitonon era ancora finito. Dato che solo pic-cole quantità di anticorpi potevano venireisolate da ogni topo, per «immortalare» lecellule che secernevano questi anticorpiabbiamo fatto ricorso al metodo di Kóhlere Milstein.
Kathryn Haskins e Janice White del no-stro laboratorio hanno estratto plasmacel-lule B da uno dei topi immunizzati controun ibridoma di linfociti T, specifici perl'antigene ovalbumina di pollo. Le plasma-cellule sono state poi fuse con cellule tu-morali in modo da ottenere ibribomi diplasmacellule che sarebbero stati facil-mente coltivati in vitro. Questi ibridomihanno secreto anticorpi, i quali sono statiselezionati in base alla capacità di blocca-re il riconoscimento dell'ovalbumina dipollo e della proteina di classe II e di cep-po d da parte dell'ibridoma di linfociti T,specifici per l'ovalbumina di pollo. Un soloibridoma di plasmacellule ha mostratoquesta proprietà. Analoghi anticorpi sonostati ottenuti all'incirca nello stesso perio-do da James P. Allison, allora al C an-cer Center dell'Università del Texas aSmithville, negli Stati Uniti, e da Stefan C.W. Meuer e Ellis Reinherz della HarvardMedical School.
Grazie soprattutto alla riuscita propa-gazione degli anticorpi per i recettori
specifici dei linfociti T è stato possibilecostruire un'immagine di questi recettoriproteici. Il solido legame tra recettore eanticorpo ha fatto sì che i ricercatori po-tessero purificare il recettore in grandiquantità, sufficienti a mettere in luce le sueproprietà molecolari di base. Cosa che for-se può sorprendere, esiste a quanto pareuna stretta rassomiglianza strutturale trala molecola dell'anticorpo e il recettore deilinfociti T. Entrambi sono costituiti da duecatene polipeptidiche codificate da geniben distinti nel DNA e tenuti uniti da fortilegami covalenti che collegano due atomidi zolfo (si veda l'illustrazione della pagi-na a fronte). Negli anticorpi le due catenedifferiscono per la dimensione e sono chia-mate catena pesante e catena leggera; cia-scuna include una sequenza di amminoa-cidi relativamente costante in un singoloanimale, perfino per anticorpi che si lega-no ad antigeni distinti. Ogni catena ha pe-rò anche un lungo tratto di amminoacidiche varia in misura considerevole negli an-ticorpi con diversa specificità antigenica.
La variabilità di un anticorpo all'inter-no di un animale deriva dal genoma di que-st'ultimo. L'idea di fondo è che ogni cate-na polipeptidica dell'anticorpo consta ditre o quattro regioni, ciascuna delle qualipuò essere codificata da uno o dall'altro didiversi segmenti di DNA, selezionati a ca-so. La variabilità combinatoria che insor-ge nella «costruzione» dell'anticorpo porta
a un numero elevato di anticorpi ben di-stinti tra loro.
Per esempio, una catena pesante constadi quattro regioni, cioè di quattro sequenzeamminoacidiche: la regione costante, unaregione di congiunzione, una regione re-sponsabile della diversità e la regione va-riabile. La regione costante è codificata daun solo segmento di DNA, mentre il seg-mento di DNA che codifica per la regionedi congiunzione può essere selezionato acaso tra quattro segmenti distinti. Analo-gamente la regione responsabile della di-versità è codificata da un segmento di DNA
selezionato tra più di dieci e la regione va-riabile è codificata da un segmento di DNA
selezionato tra più di 100. Il numero disequenze amminoacidiche che possonoaver origine dalle varie combinazioni è,pertanto, superiore a 4 x 10 x 100, o4000. Anche la catena leggera consta diregioni codificate da differenti combina-zioni di DNA e una ulteriore variabilità traeorigine dal fatto che i punti di contatto trala regione di congiunzione, quella respon-sabile della diversità e la regione variabilesono imprecisi. Secondo Susumu Tonega-wa del Massachusetts Institute of Techno-logy (mur), il numero di molecole di anti-corpo diverse può raggiungere il miliardo(si veda l'articolo Le molecole del sistema
immunitario di Susumu Tonegawa in (<LeScienze» n. 208, dicembre 1985).
Nel recettore dei linfociti T, le due cate-ne sono chiamate catena alfa e catena be-ta. Nel topo esse hanno entrambe un pesomolecolare di circa 43 000 unità di massaatomica (dalton); nella specie umana la ca-tena alfa pesa circa 50 000 unità di massaatomica e la catena beta circa 39 000.Confrontando le catene alfa e le catenebeta estratte da differenti cloni di linfoci-ti T, abbiamo trovato che su ogni catena,di clone in clone, compaiono certe sequen-ze fisse di amminoacidi. Altre sequenze,invece, possono variare.
Con la scoperta delle proteine che co-stituiscono il recettore dei linfociti T, sonoentrate in gioco le tecniche della biologiamolecolare. Queste tecniche permettono airicercatori di analizzare i geni che codifi-cano per una data proteina e un'analisi diquesto tipo può chiarire la struttura di unaproteina molto più rapidamente di quantopossano fare i metodi biochimici. Inoltre,dati i ruoli analoghi dei recettori dei linfo-citi T e delle immunoglobuline nel sistemaimmunitario e le somiglianze strutturaliche stanno emergendo tra loro, è sembratoprobabile che i segmenti di DNA che codi-ficano per il recettore dei linfociti T subis-sero un riassestamento prima che le pro-
teme che costituiscono il recettore si espri-messero sulla superficie del linfocito T,proprio come accade per i segmenti diDNA che codificano per il recettore dei lin-fociti B. Si è scatenata quindi subito unagara per trovare i geni responsabili del re-cettore dei linfociti T.
primi attendibili candidati al ruolo digeni sono stati comunicati simultanea-
mente da due gruppi: Stephen M. Hedrick,Mark M. Davis e collaboratori dei Natio-nal Institutes of Health, dell'Universitàdella California a San Diego e della Stan-ford University, e Tak W. Mak e collabo-ratori dell'Ontario C ancer Institute. En-trambi i gruppi, il primo lavorando su genidi topo e il secondo su geni umani, pensa-vano che le proteine del recettore dei linfo-citi T sarebbero state trovate soltanto neilinfociti T e non, per esempio,) nei linfoci-ti B. Per confermare questa loro supposi-zione, essi hanno adottato tecniche speri-mentali raffinate e sono stati presto in gra-do di identificare i geni espressi solo neilinfociti B o solo nei linfociti T, ma non inentrambi. Sono stati identificati dapprimai geni che codificano per la catena beta delrecettore e, nell'anno successivo, sono sta-ti isolati anche quelli per la catena alfa.Dall'analisi di questi geni si. sono potuti
34 35

L'effetto del timo sul linfocito T è stato dimostrato in un esperimento diRolf Zinkernagel, allora alla Scripps Clinic and Research Foundation.Topi ibridi figli di genitori di ceppo a e di ceppo b sono serviti comecontrolli. Essi sono stati immunizzati contro il virus della coriomeningitelinfocitaria (1) e hanno poi fornito linfociti T citotossici, specifici per ilvirus (2). Questi linfociti sono stati in grado di uccidere le cellule infettatedal virus e provenienti da topi sia di ceppo a sia di ceppo b (3). Per
determinare il ruolo del timo, quest'organo è stato asportato da un altrotopo ibrido e tutti i linfociti T préesistenti, assieme ad altre cellule linfoidi,sono stati distrutti irradiando l'animale (4). Nuove cellule staminali delmidollo osseo, da cui normalmente si sviluppano i linfociti T, sono stateallora ottenute da un altro topo ibrido (5) e il timo irradiato di un topodi ceppo b è stato trapiantato nell'ibrido timectomizzato (6). Le cellulestaminali ibride sono state fatte sviluppare nel topo ibrido con timo
TOPO DI CEPPO A TOPO DI CEPPO
VIRUS DELLACORIOMENINGITELINFOCITARIA
TOPO IBRIDO
VIRUS DELLA LINFOCITI T CITOTOSSICI IBRIDICORIOMENINGITELINFOCITARIA
2
3RECETTORE SPECIFICOPER IL CEPPO BRECETTORE SPECIFICO
PROTEINAPER IL CEPPO ADI CEPPO 8,A,'
PROTEINA Di CEPPO A, CODIFICATA I— CODIFICATA DAL MHC DAL MHC
CELLULA MORTA DI CEPPO A CELLULA MORTA DI CEPPO B
LINFOCITO TCITOTOSSICO IBRIDO,
SVILUPPATOSIIN TIMO DICEPPO B
10
CELLULA DI CEPPO A,VIVA E INFETTATA
RECETTORESPECIFICOPER ILCEPPO B...,
••
CELLULA MORTADI CEPPO B
CELLULE 01 ., 5STAMiNALi e•-•
IBRIDE
TOPO IBRIDO TOPO DI CEPPO
TIMO DI CEPPO BIRRADIATO
VIRUS DELLACORiOMENINGITE
8 LINFOCITARIA
conoscere numerosi particolari sulla strut-tura del recettore dei linfociti T.
Sia la catena alfa, sia la catena beta delrecettore di questi linfociti hanno la regio-ne variabile, la regione costante e la regio-ne di congiunzione. Inoltre, i ricercatorisopra citati hanno confermato la presenzadi una regione responsabile della diversitànella catena beta e probabilmente di una
e analoga anche nella catena alfa.L sequenze amminoacidiche di ciascunadi queste regioni sono simili alle sequenzeanaloghe presenti nelle immunoglobuline,ma non sono affatto identiche.
Davis e collaboratori da un lato e LeroyHood e collaboratori del California Insti-tute of Technology dall'altro hanno presoin esame l'organizzazione delle sequenzedi DNA che codificano per la catena betadel recettore e hanno trovato finora 12segmenti di DNA che codificano per altret-tante regioni di congiunzione, due segmen-ti che. codificano per regioni responsabilidella diversità (ognuno di questi segmentipuò essere trascritto secondo l'uno o l'al-tro di tre sistemi di lettura) e circa 20 seg-menti che codificano per regioni variabili.Il numero delle possibili combinazioni diamminoacidi nella catena beta è, pertanto,almeno 12 x 2 x 3 x 20, cioè 1440. Maanche questo numero è troppo piccolo:una stima realistica della variabilità deveanche tenere conto delle mutazioni e diuna congiunzione inesatta, che possonoinfluire sul recettore dei linfociti T, propriocome influiscono sulla molecola di anti-corpo. La variabilità nella catena alfa puòanche essere superiore: benché non sianostate identificate finora regioni responsabi-
li della diversità, sembrano esservi più re-gioni di congiunzione e regioni variabili diquante ve ne siano per la catena beta; forseanche 100 per ogni tipo. Le catene alfa ebeta possono dunque combinarsi per for-mare differenti tipi di recettore dei linfoci-ti T, nell'ordine di 10 milioni, il che è suf-ficiente a giustificare il ben noto repertoriodi linfociti T presente in un animale.
T e nuove informazioni acquisite sulla1-d struttura del recettore dei linfociti Thanno permesso di riproporre in terminiben più precisi molte delle questioni ri-guardanti il sistema immunitario. Vi sonoessenzialmente tre tipi di riconoscimentoche sono stati osservati e per i quali si vor-rebbe ora dare una spiegazione a livellostrutturale. In primo luogo, il linfocito Tnon reagisce alle proteine del proprio «io»,codificate dal MHC. In secondo luogo, illinfocito T reagisce quando si trova simul-taneamente in presenza di un antigene e diuna proteina del proprio io, codificata dalMHC, ma generalmente non quando si tro-va in presenza di un antigene associato auna proteina codificata dal MHC, ma ap-partenente a un altro individuo. In terzoluogo, il linfocito T può reagire a una pro-teina di un altro individuo, codificata dalMHC, se in assenza dell'antigene. È questol'effetto che spiega il rigetto di tessuti inne-stati o trapiantati. Associata a queste os-servazioni vi è una questione inerente losviluppo: in che modo i precursori dei lin-fociti T si differenziano nel timo in celluleche hanno queste proprietà?
La spiegazione più lineare della tolle-ranza immunologica è che i doni di linfo-
citi T che reagiscono alle proteine del pro-prio io, codificate dal mHC, sono eliminatinel timo in un modo o nell'altro. Non vi èancora una descrizione precisa di comeuna simile selezione potrebbe aver luogo.Si potrebbe suggerire che, a un certo sta-dio dello sviluppo, i linfociti T muoiono sei loro recettori, che hanno una specificitàper un determinato antigene, si legano auna qualsiasi molecola espressa dalle cel-lule dell'organismo a cui essi appartengo-no. Questa spiegazione è nota come «teo-ria dell'aborto clonale». Essa sostiene che,allorché i linfociti T si sviluppano, tuttiquelli che reagiscono in maniera specificaalle proteine del proprio io, codificate dalmHc, oppure ad altri antigeni del proprioio, verrebbero uccisi perché continuamen-te bombardati da quelle molecole. Mori-rebbero anche i linfociti T che reagisconoin modo specifico alle proteine del proprioio, codificate dal MHC e associate a un an-tigene estraneo, se dovessero legarsi a si-mili complessi, mentre in un animale noncolpito da un antigene di questo tipo essi sisvilupperebbero fino a raggiungere la pie-na maturità. Potrebbero così sopravviverefino a quando un antigene estraneo cheinvadesse l'organismo scatenasse la lororeazione.
La risposta del linfocito T a una protei-na codificata dal MHC e associata a unantigene fa sorgere delle perplessità. RolfZinkernagel e collaboratori e Michael J.Bevan, allora al MIT, hanno riconosciutoinnanzitutto le dimensioni del problemanel corso di uno studio da essi intrapresosulla «istruzione» dei linfociti Tnel timo (siveda l'illustrazione qui sotto). Essi hanno
incrociato topi appartenenti ai ceppi a e b,generando cosi una progenie ibrida checonteneva nel proprio organismo proteinecodificate dal MHC e appartenenti a en-trambi i ceppi. Il timo di ogni topo ibridoè stato quindi asportato e l'animale è statosuccessivamente irradiato allo scopo di di-struggere tutti i suoi linfociti B e T. Unnuovo timo è stato quindi trapiantato inesso da un topo di ceppo b e nuove cellulestaminali del midollo osseo di un altro ani-male ibrido dello stesso tipo sono state pa-rimenti trapiantate.
In un topo ibrido normale le cellule sta-minali ibride si svilupperebbero in linfoci-ti T maturi in grado di reagire collettiva-mente all'antigene, associato o con unaproteina codificata dal MHC e appartenen-te al ceppo a, o con una proteina codifica-ta dal MHC e appartenente al ceppo b. Ne-gli animali da laboratorio, tuttavia, sullecellule non linfatiche del timo erano pre-senti soltanto proteine di ceppo b, anchese, nel resto del corpo, si trovavano, inve-ce, proteine caratteristiche di entrambi iceppi. È stato trovato con sorpresa che ilinfociti T maturi di questi animali poteva-no reagire all'antigene soltanto se questoera associato a una proteina appartenenteal ceppo b e codificata dal MHC, e non seera associato, invece, a una proteina diceppo a. Così, allorché i linfociti T si svi-luppavano nel timo di animali appartenen-ti al ceppo b, i loro recettori venivano ap-parentemente selezionati per riconosceresoltanto quegli antigeni che erano associa-ti alle proteine codificate dal MHC, presentiin quell'organo. A quanto pare i linfociti Tnon riescono a riconoscere gli antigeni as-
di ceppo b (7). Quando questo topo è statoimmunizzato contro il virus della coriomenin-gite linfocitaria (8), i linfociti T isolati da essonon sono stati in grado di uccidere le celluleinfettate dal virus e derivate da topi di ceppo a(9), anche se erano in grado di uccidere le stes-se cellule appartenenti ai topi di ceppo b (10).
sociati a proteine codificate dal mHC edestranee al timo nel quale sono ospitati.
ono state avanzate molte ipotesi perspiegare questo inatteso risultato, ma
nessuna è del tutto soddisfacente. Forse,perché i linfociti T possano maturare e di-ventare funzionanti, i loro recettori devo-no legarsi debolmente alle proteine delproprio io, codificate dal MHC e presentinel timo. Successivamente quei linfociti T,i cui recettori hanno la massima affinitàper le proteine del proprio io e codificatedal mHC, potrebbero andare incontro a unaborto clonale; i linfociti T che rimango-no, con una affinità scarsa ma ancora po-sitiva per le proteine del proprio io e codi-ficate dal mHc, verrebbero allora riversatiin circolo. Quando, però, un antigene silega a una di queste proteine, l'affinità delrecettore presente sui linfociti T circolantiper quell'antigene potrebbe aumentare inmaniera sostanziale.
Che dire dell'intensa reazione dei linfo-citi T a un tessuto estraneo trapiantato?La spiegazione che trova maggior favoreè che a un linfocito T una proteina estra-nea, codificata dal MHC, sembra chimica-mente la stessa cosa del complesso tra unaproteina del proprio io, codificata dalMHC, e un antigene. Questa spiegazionetiene conto di parecchie osservazioni. Peresempio, il recettore del linfocito T che silega alla proteina del proprio io, codificatadal MHC, sembra essere lo stesso recettoreche anche si lega alla proteina estranea.Inoltre, questo recettore sembra avere unapredisposizione a legarsi all'antigene inpresenza o della proteina di classe Io dellaproteina di classe II, ma non di entrambe,indipendentemente dal fatto che la protei-na codificata dal MHC sia o no estranea.
Nell'ultimo anno si sono svolte nume-rose ricerche per trovare un fondamento aqueste predisposizioni del recettore. Fino-ra, però, non sono state osservate differen-ze ovvie tra i linfociti T ristretti all'una oall'altra classe di proteine codificate dalMHC. Nel 1984, Tonegawa e collaboratorihanno trovato un terzo tipo di gene, il genegamma, che subisce una riassestamentoda parte dei linfociti T. Le proprietà dellasequenza di questo gene fanno pensare chela proteina che il gene codifica non facciaparte del recettore del linfocito T, così co-me questo recettore viene correntementeinteso. Cionondimeno la proteina si espri-me solo nei linfociti T citotossici e cosìcontribuisce, forse, in qualche modo nonsospettato alla specificità, nei riguardi delmHc, della cellula che la contiene.
Se esiste una somiglianza sul piano chi-mico tra una proteina codificata da unMHC estraneo e un complesso costituito dauna proteina codificata dal mHc del pro-prio io e un antigene, si potrebbe pensareche l'antigene che fa parte del complessosia piccolo. Rispetto a un antigene grosso,un antigene piccolo si adatterebbe anchepiù facilmente alla fenditura, nel recettoredel linfocito T, che funge da legame. Re-centi ricerche condotte da numerosi grup-pi, tra cui quelli di Emil Unanue e collabo-
ratori e di Ronald H. Schwartz e collabo-ratori del National Institute of Allergy andInfectious Diseases e il nostro, in collabo-razione con Howard M. Grey del NationalJewish Center for Immunology and Respi-ratory Medicine, hanno confermato chel'antigene legato è piccolo.
Per esempio, abbiamo trovato che i lin-fociti T reagiscono a cellule vive, tenute inincubazione assieme a un antigene, mentrenon rispondono allo stesso antigene quan-do esso viene aggiunto alle cellule dopoche sono state fissate con una sostanzachimica. Quando alle cellule fissate conuna sostanza chimica si aggiunge solo unpiccolo frammento dell'antigene, il linfoci-to T reagisce di nuovo. La ricerca confer-ma che le cellule che espongono l'antigene,e alle quali può legarsi un linfocito T, han-no già trasformato, o digerito, in qualchemodo l'antigene.
Dobbiamo sollevare un'ultima obiezio-ne. L'esistenza dei linfociti B dimo-
stra chiaramente che si può sviluppare unsistema in cui un recettore riconosca unantigene nativo e libero e si leghi a esso.Questo legame finisce per portare a un'e-spansione e a un differenziamento clonalidei linfociti B, il che dà origine ad anticorpiefficaci nel distruggere l'antigene. Perché,allora, i linfociti T hanno messo in atto unsistema così elaborato per riconoscerel'antigene solo in associazione con le pro-teine prodotte dal mHc?
Si può dare a questa domanda una ri-sposta teleologica: il linfocito T è pro-grammato solo per reagire con un antigenefissato su una cellula bersaglio, e non conun antigene libero. Per esempio, un linfo-cito T cnotossico è programmato per uc-cidere cellule infettate da un virus e, per-tanto, inibisce la crescita e la diffusione delvirus. Un linfocito del genere non può uc-cidere una particella virale libera e, pertan-to, le sue risorse potrebbero venir sperpe-rate se non vi fosse il mezzo per dirigere lasua attenzione su un virus che si sviluppaall'interno di una cellula nucleata. Così ilsistema immunitario ha programmato ilrecettore del linfocito T in modo che possalegarsi a un antigene virale solo quandopuò.legarsi anche a una proteina codifica-ta dal MHC del proprio io e appartenentealla classe I, proteina che è presente sullasuperficie di ogni cellula nucleata.
Il sistema immunitario, ancor più astu-tamente, ha progettato dei recettori sui lin-fociti T coadiuvanti, i quali sono destinatia interagire, in primo luogo, con i linfoci-ti B e con altre cellule del sistema immuni-tario. Questi recettori si legano all'antige-ne solo quando esso è associato con pro-teine codificate dal MHC del proprio io eappartenenti alla classe I, proteine chevengono espresse solo alla superficie deilinfociti B e delle altre cellule immunitarie.Il sistema immunitario si è così evoluto inmodo che entrambe le classi delle proteinecodificate dal mHc fungano da segnale peri linfociti T, guidando questi ultimi versol'antigene esattamente nei punti dove la lo-ro azione possa essere efficace.
36
37