FAGOCITOSI fagosoma precoce La cascata si€¦ · vita intracellulare fagosoma (o fagolisosoma)...
54
FAGOCITOSI fagosoma nascente fagosoma precoce fagosoma tardivo Fusione con lisosoma (fagolisosoma) morte batterica Ogni stadio ha marcatori specifici La cascata si completa in pochi minuti nella fagocitosi intervengono alcune frazioni del complemento
Transcript of FAGOCITOSI fagosoma precoce La cascata si€¦ · vita intracellulare fagosoma (o fagolisosoma)...

FAGOCITOSI
fagosomanascente
fagosomaprecoce
fagosomatardivo
Fusione con lisosoma
(fagolisosoma)morte
batterica
Ogni stadio ha marcatori specifici
La cascata si completa in pochi
minuti
nella fagocitosiintervengono alcune frazioni del
complemento

COMPLEMENTO (C’): 25 proteine plasmatiche 7 recettori diversi
riconoscono le componenti di C’Opsonizzazione batteri
lisi batteri
lisi cellule infettate (da virus)
Induzione dell’infiammazione
Chemiotassi dei leucociti
Normalità: OFF
ATTIVAZIONE
via classica(Complesso Ag/Ab)
via alternativasuperficie del patogeno
RICHIEDE ANTICORPI(RISPOSTA
ADATTATIVA)
via della lectinaMannano (patogeno) +
lectine (siero)In realtà la più
frequente..

Attivazione del complemento VIA ALTERNATIVA
C3Idrolisi spontanea
(instabile in fase acquosa)
C3aC3b
In presenza di un patogeno C3b si lega covalentemente alla superficie
C3bC3b legato si lega al “fattore B”B
Ba
Il fattore B legato a C3b viene tagliato dal fattore D (proteasi)
C3b+Bb = C3 convertasi della via alternativa
RUOLO CHIAVE NELL’INFIAMMAZIONE
C3bBb

C 7
C3
C3a
C3b
Complesso C3bBb3b:C5 convertasi
C3 convertasi
C5a
C5bC5b si lega e richiama
C6-C7-C8-C9 (polimerico)
Formando il MAC(membrane attack complex )
Che provoca la formazione di fori nelle membrane di cellule eucariotiche e batteri didermi
Una molecola di C3 o C5 convertasi può catalizzare il taglio di centinaia-migliaia di molecole bersaglio
C5b C 6 C 89
C 9C 9C 9

Opsonine:
Legano i microbi ai fagociti
C3b (C4b) ha un’azione OPSONIZZANTE
CON UN MECCANISMO A CERNIERA CHE COINVOLGE RECETTORI
sui batteri
e sui fagociti

C3b si lega in modo ASPECIFICO alle superfici batteriche
Il taglio espone un legame tioestere molto reattivo Che reagisce con gruppi OH
di cellule vicine
C3b OH
Se C3b si lega a cellule eucariotiche è distrutto dal fattore H (proteasi)
I batteri non hanno il fattore Hrestano opsonizzati
Sui fagociti ci sono recettori SPECIFICI che legano C3b, stimolando la fagocitosi dei batteri opsonizzati

NORMALITÀ
Fagocitosi lenta
RICHIAMO
chemiotassinecomplemento
Chemiotassine batteriche
ATTIVAZIONE
OPSONINEAnticorpi
Es. N-term rimosse dalle proteine
N-fMet-Leu-Phe: efficace a [10-11 M]

MEDIAZIONE DELL’INFIAMMAZIONE + RECLUTAMENTO DEI FAGOCITI
C5a-C3a-C4a
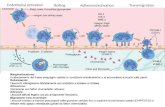
Chemiotassi positiva per i leucociti
VASODILATAZIONE
Espressione di molecole di adesione
selectine
integrine
DIAPEDESI

C5a-C3a-C4a si legano anche ai mastociti
ATTIVITA’ ANAFILOTOSSICA
Rilascio di istamina e NO
ne inducono la degranulazione
vasodilatazione
Cellule ricche di granuli citoplasmatici situate nel tessuto connettivo,
specialmente lungo i vasi sanguigni

MONOCITI
CELLULE A VITA LUNGA
DIFFERENZIABILI
ARRIVANO DOPO I PMN
RICHIAMO:
sostanze emesse dai PMN
ELIMINANO DETRITI E
BATTERI RESIDUI
ATTIVAZIONE
CITOCHINE:
I monociti possono stabilirsi nei tessuti
MACROFAGI
molecole proteiche prodotte da vari tipi cellulari
citochinecitochine

Attivazione dei macrofagi
TLR (Toll-Like-Receptors) Specifici per le sottopopolazioni di MAMP
Alcuni sulle membrane di molte cellule
Altri sulla membrana del fagosoma
Bersagli esterni Bersagli interni
TLR + bersaglio
NF-kB (nuclear Factor kB)
citochine

MACROFAGI: RUOLO CENTRALE NELLA RISPOSTA IMMUNITARIA
INTERAGISCONO CON ALTRE CELLULE ATTRAVERSO LE CITOCHINE- presentano l’antigene (APC)
Cellule delle mucose
INFIAMMAZIONE ACUTACELLULE DELL’IMMUNITA’
ADATTATIVA

REPRIMERE l’attivazionePoxvirus: proteina che mima quella adibita al controllo
dell’attivazione di C’
ELUSIONE DELLE DIFESE DELL’OSPITE
IMPEDIRE l’attivazione del complemento mimando i
tessuti dell’ospite
Es. Neisseria: acido sialico(comune sulle cellule eucariotiche) sul
lipooligosaccaride (LOS)

STRATI S
MASCHERAMENTO DEI RECETTORI PER IL COMPLEMENTO
Impedire la penetrazione del M.A.C
Es. Salmonella: rivestimenti esterni
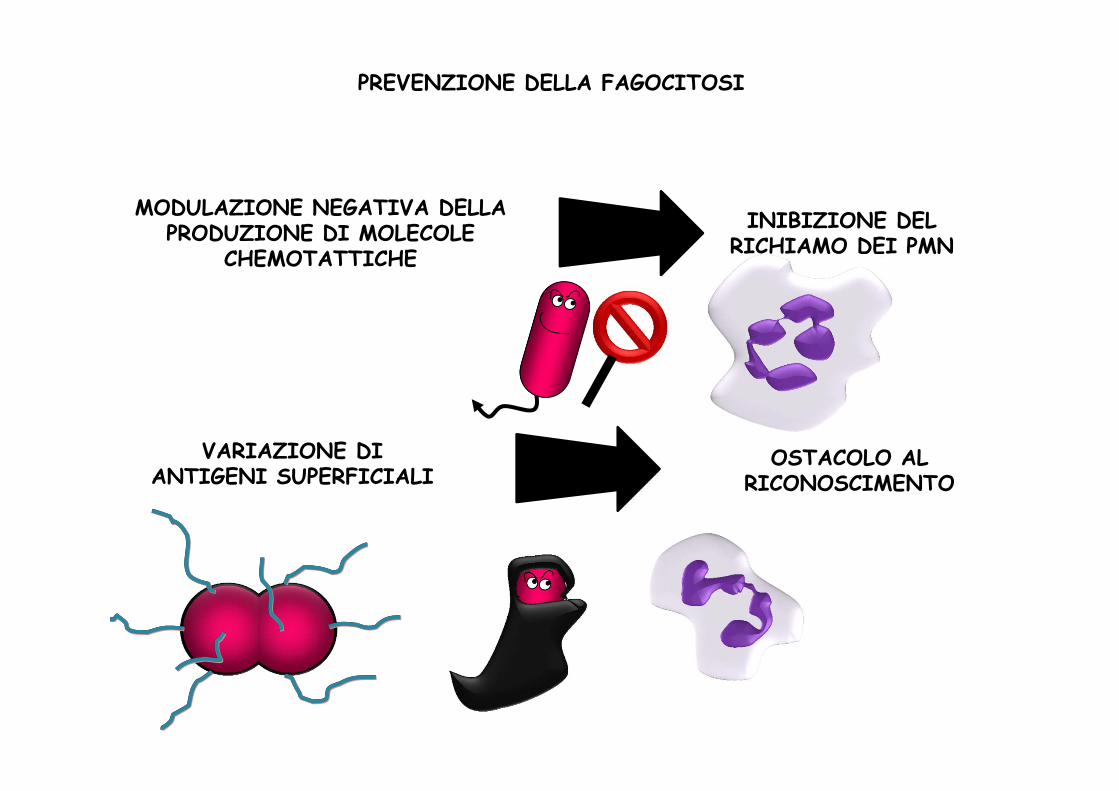
PREVENZIONE DELLA FAGOCITOSI
INIBIZIONE DEL RICHIAMO DEI PMN
MODULAZIONE NEGATIVA DELLA PRODUZIONE DI MOLECOLE
CHEMOTATTICHE
VARIAZIONE DIANTIGENI SUPERFICIALI
OSTACOLO AL RICONOSCIMENTO

PARALIZZARE IL CITOSCHELETRO DI ACTINA DEI FAGOCITI RESTANDO ALL’ESTERNO
OSTACOLO ALLA FAGOCITOSI
CIRCONDARSI DI UNA CAPSULA VISCOSA (BATTERI DIFFUSI
DAL SANGUE)

L’ESPLOSIONE DEI LISOSOMI NEL CITOPLASMA
microrganismi relativamente resistenti agli enzimi lisosomiali
UCCISIONE DEI FAGOCITI
Sono in grado di causare..

MECCANISMO A CERNIERA (ZIPPER)
MECCANISMO A GRILLETTO (TRIGGER)
YERSINIA
SALMONELLA-SHIGELLA
MOLECOLE DI ADESIONE (INVASINE)
T3SS
ANCHE ENTRARE NELLE CELLULE E’ UN MODO DISFUGGIRE ALLA FAGOCITOSI

L’ESPLOSIONE DEI LISOSOMI NEL CITOPLASMA
MICRORGANISMI RELATIVAMENTE RESISTENTI AGLI ENZIMI LISOSOMIALI
UCCISIONE DEI FAGOCITI
SONO IN GRADO DI CAUSARE..
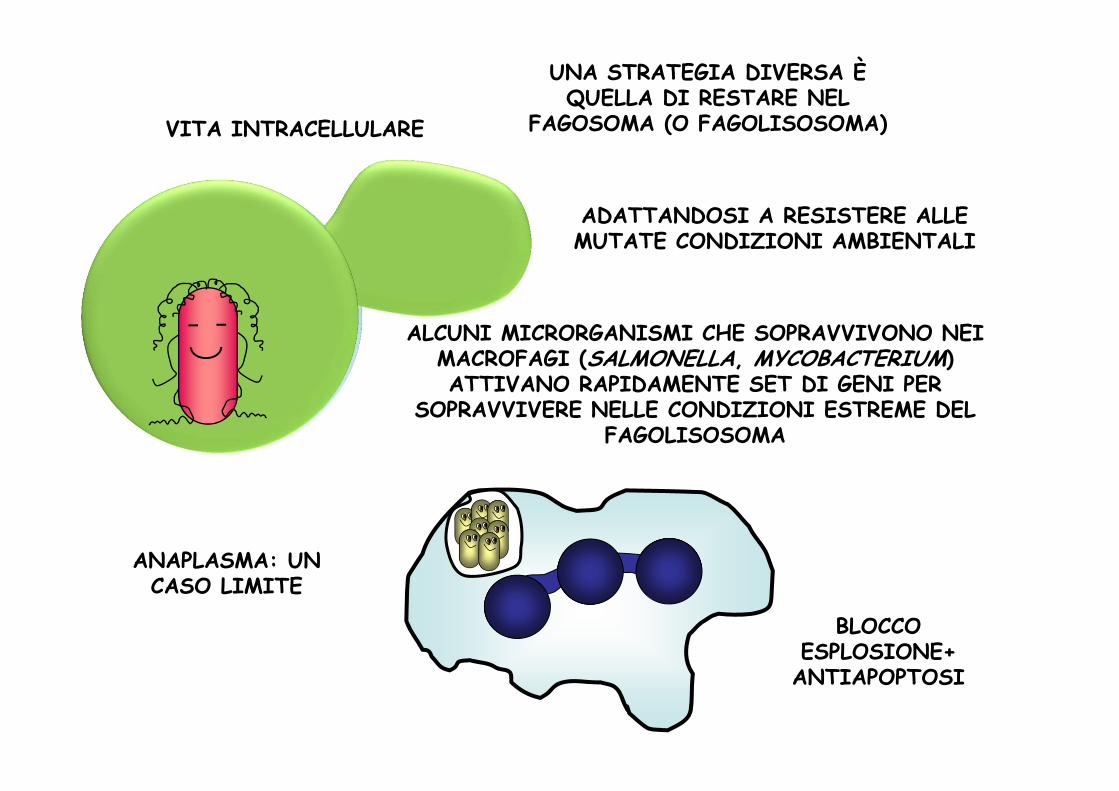
UNA STRATEGIA DIVERSA È QUELLA DI RESTARE NEL
FAGOSOMA (O FAGOLISOSOMA)VITA INTRACELLULARE
ADATTANDOSI A RESISTERE ALLE MUTATE CONDIZIONI AMBIENTALI
ALCUNI MICRORGANISMI CHE SOPRAVVIVONO NEI MACROFAGI (SALMONELLA, MYCOBACTERIUM) ATTIVANO RAPIDAMENTE SET DI GENI PER
SOPRAVVIVERE NELLE CONDIZIONI ESTREME DEL FAGOLISOSOMA
BLOCCO ESPLOSIONE+ANTIAPOPTOSI
ANAPLASMA: UN CASO LIMITE
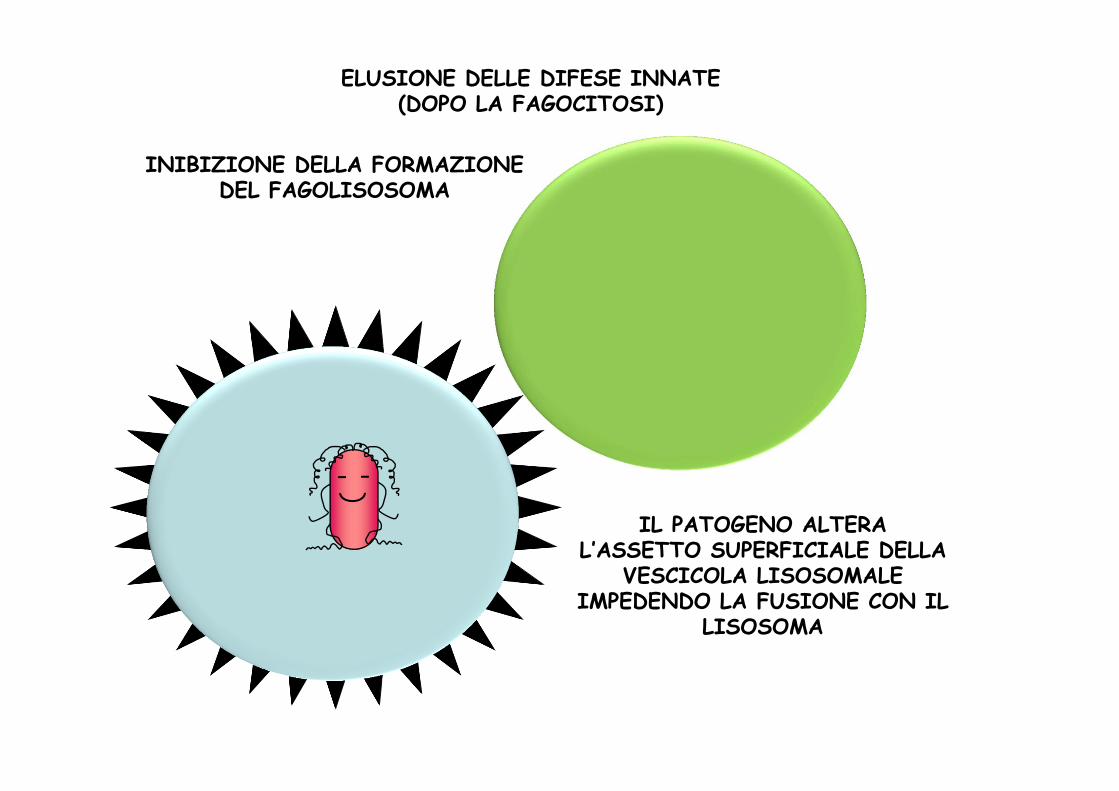
IL PATOGENO ALTERA L’ASSETTO SUPERFICIALE DELLA
VESCICOLA LISOSOMALE IMPEDENDO LA FUSIONE CON IL
LISOSOMA
INIBIZIONE DELLA FORMAZIONE DEL FAGOLISOSOMA
ELUSIONE DELLE DIFESE INNATE(DOPO LA FAGOCITOSI)

ALCUNI PATOGENI LISANO LA MEMBRANA DEL
FAGOSOMA
FUGA DAL FAGOSOMA
E SI MOLTIPLICANO NEL CITOPLASMA

-SHIGELLE IN MOTO-
SERVIZIO FOTOGRAFICO INSTITUT PASTEUR
ALCUNE SPECIE (LISTERIA, SHIGELLA, RICKETTSIA) POSSONO INTERVENIRE SUI FILAMENTI DI ACTINA DELL’OSPITE E FARSI
“SPINGERE” DA UNA CELLULA ALL’ALTRA

DIFESE ADATTATIVE
UMORALE (anticorpi)CELLULO-MEDIATA
ESPOSIZIONE ALL’AGENTE INFETTANTE
ANTICORPI: IMMUNOGLOBULINE RISPOSTA A MOLECOLE
ESTRANEE
VARIETA’ ENORME (RIARRANGIAMENTO GENICO)
CELLULE B SI SVILUPPANO NEL
MIDOLLOCELLULE T
SI SVILUPPANO NEL TIMO
RISPOSTA SPECIFICA

ANTIBODY GENERATOR
ANTIGEN
ANTIGENE: SOSTANZA NON RICONOSCIUTA COME “SELF”
INDUCE LA FORMAZIONE DI ANTICORPI
APTENE
“απτομαι” LEGARE STRINGERE
UN APTENE NON INDUCE LA FORMAZIONE DI ANTICORPI
MA E’ IN GRADO DI LEGARLI
“ANTIGENE INCOMPLETO”
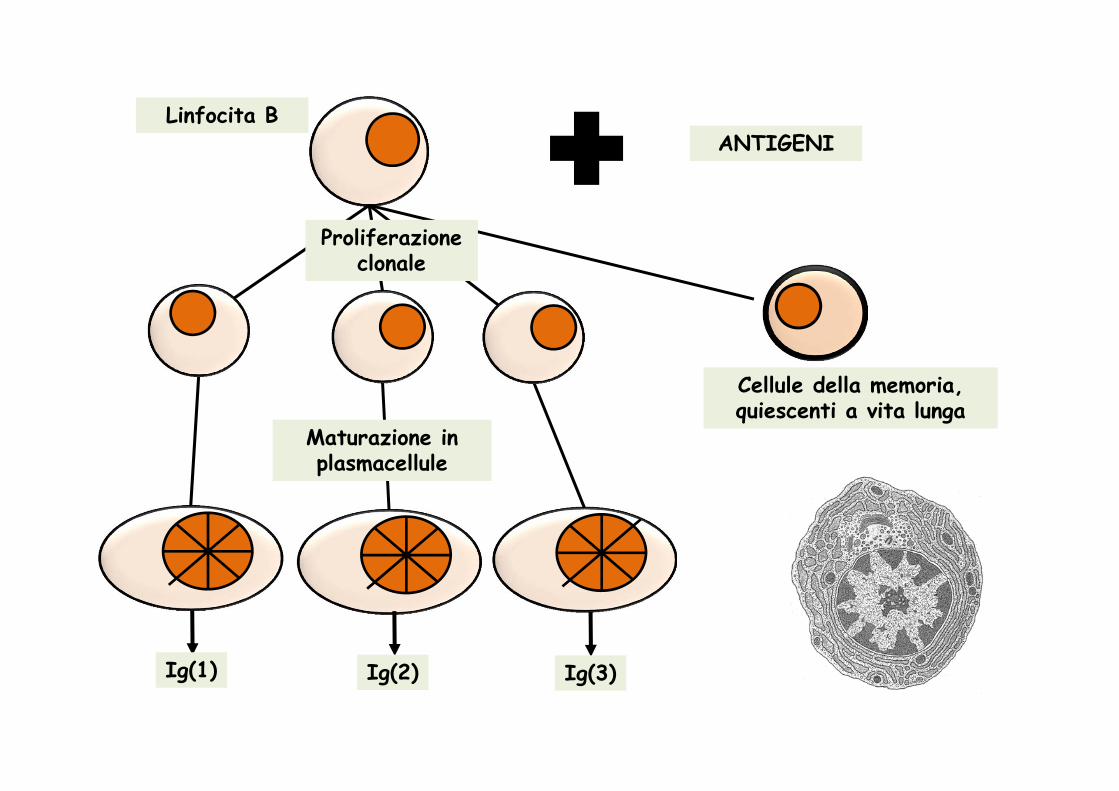
Proliferazione clonale
Maturazione in plasmacellule
Cellule della memoria, quiescenti a vita lunga
Ig(1) Ig(2) Ig(3)
Linfocita BANTIGENI

IMMUNITA’ CELLULO-MEDIATA
I PATOGENI INTRACELLULARI NON SONO RAGGIUNTI DA ANTICORPI, C’, FAGOCITI
INTERVENTO DEI LINFOCITI T
CD8CD4
HELPER KILLER
LE CELLULE SOMATICHE HANNO IL COMPLESSO MHC
MANTENGONO GLI ANTIGENI IN CONFORMAZIONE TALE DA
ESSERE RICONOSCIUTI

APC: ANTIGEN PRESENTING CELLES. UN MACROFAGO
ProliferazioneRisposta umorale Proliferazione
Risposta cellulo-mediata
1 2 3
4
1) TH interagisce con MHC-II
2) Reagisce a IL1
3) Produce IL2
4) Produce citochine che inducono la proliferazione dei linfociti B e T

UCCIDONO LE CELLULE CHE NON RICONOSCONO COME SELF
MENTRE LA RISPOSTA ADATTATIVA SI ESPLICA
DIFESE A PONTECellule natural killercellule T primitive
ES. CELLULE INFETTATE DA VIRUS/CELLULE TUMORALI
CON BASSI LIVELLI DIMHC-I
cellule B B1: sottopopolazione di B B B1
Producono “anticorpi naturali” (IgM)Che si mantengono nel siero senza
stimolazione antigenica Reagiscono con i carboidrati tipici dei
patogeni

Catena pesante (costante)
Catena pesante (variabile)
Catena leggera (costante)
Catena leggera (variabile)
Tenuti insieme da ponti disolfuro
Sono divisi in 5 classi
IgA, IgD, IgE, IgG, IgM
FUNZIONI
Neutralizzazione (es. tossine)opsonizzazione
Aggregazione allontanamento
Anticorpi: formati da diversi frammenti
Fc
Fab
Cerniera(hinge)

IgD, E, G monomeriche
IgD: recettore per Agsulle cellule B non esposte all’antigene
IgE: implicate nei fenomeni allergici allergene
istamina
Mast-Zellen
IgG: principali circolanti, legano C’
Via classica
4 forme (IgG, IgG2, IgG3, IgG4)
Passano il filtro placentare (immunità passiva al feto)
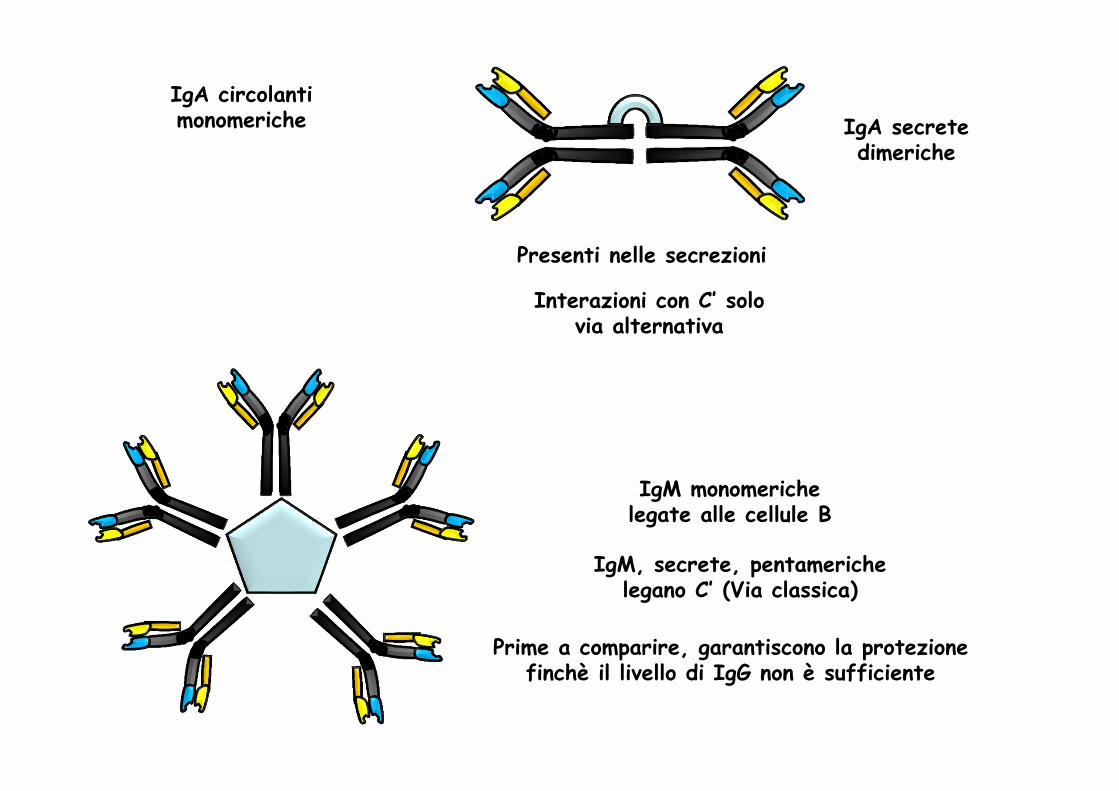
IgA circolanti monomeriche
IgM, secrete, pentamerichelegano C’ (Via classica)
IgM monomerichelegate alle cellule B
Prime a comparire, garantiscono la protezione finchè il livello di IgG non è sufficiente
IgA secretedimeriche
Presenti nelle secrezioni
Interazioni con C’ solo via alternativa

VIA CLASSICA
ComplessoC1qr2s2
C1q cambia conformazione
C1q Legame con superficie patogeno o anticorpi
Attiva C1r (proteasi)
Attiva C1s (proteasi)
C4C2
C4aC4bC2a C2b
ComplessoC4bC2bC3 convertasi
MBP
VIA DELLA LECTINA
M
Mannan Binding Protein MBP(lectina)MBL-associated serine proteaseMASP1 e MASP 2 simili a C1r, C1s
Attivazione di MASP1/Masp2

Risposta primaria Risposta secondaria
IgM
IgG
Ig-totali
Le percentuali relative di IgM e IgG sono diverse
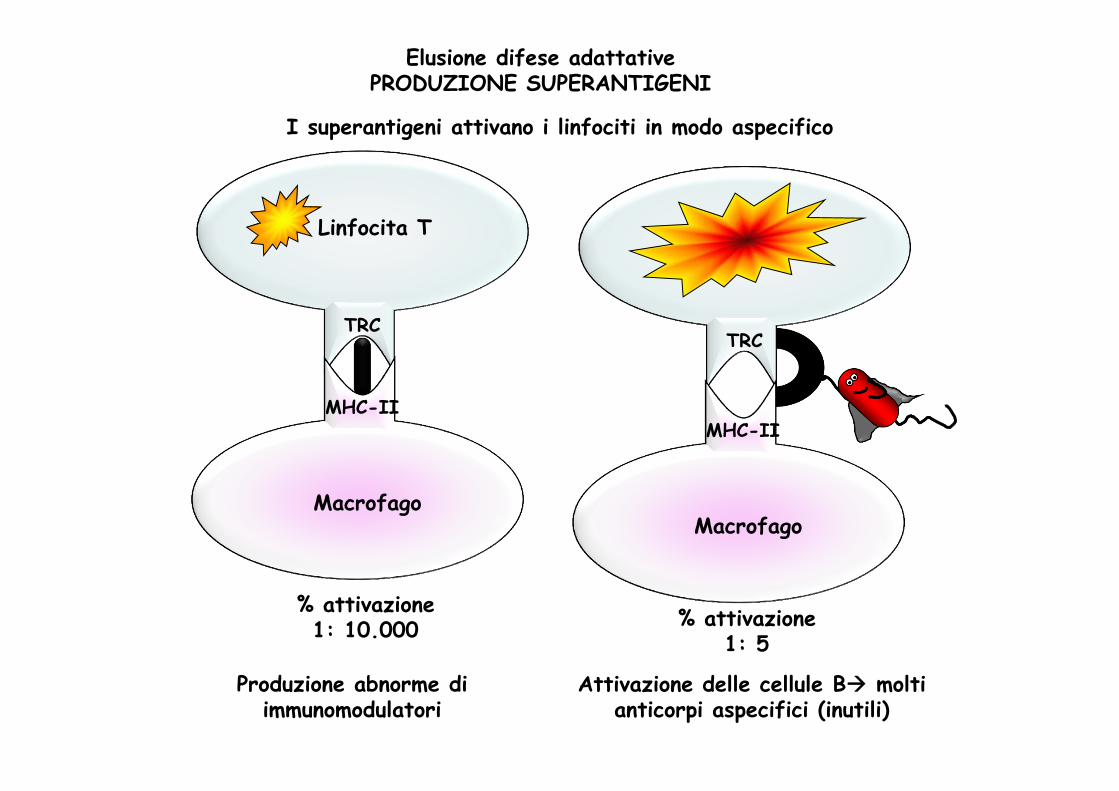
% attivazione1: 5
% attivazione1: 10.000
Linfocita T
TRC
Macrofago
MHC-II
I superantigeni attivano i linfociti in modo aspecifico
Linfocita T
TRC
Macrofago
MHC-II
Elusione difese adattativePRODUZIONE SUPERANTIGENI
Attivazione delle cellule B molti anticorpi aspecifici (inutili)
Produzione abnorme di immunomodulatori

Elusione difese adattativeInattivazione immunoglobuline
Alcuni patogeni mucosali (es. Neisseria) producono IgA proteasi

Es: SalmonellaPhoP/PhoQ sopravvivenza
intracellulare/interferenza con la presentazione degli antigeni batterici
Elusione difese adattativeBlocco della presentazione dell’antigene
Es: LegionellaAttraverso T4SS, blocca la sintesi di
MHC-I e MHC-II assenza di presentazione

TOSSINE
ESOTOSSINE
CITOLISINE
TOSSINE A,B
TOSSINE SUPERANTIGENE
ENDOTOSSINA
FRAZIONE LIPIDICA della membrana
esterna
BATTERI ALL’ATTACCO: LE TOSSINE
LE TOSSINE CAUSANO DIRETTAMENTE DANNO:

TOSSINE PROTEICHE (ESOTOSSINE)proteine solubili prodotte da batteri vivi durante la fase
esponenziale di crescita sono i veleni più potenti per l’uomo
Le specie batteriche che producono esotossine causano in genere malattie in cui la tossina ha un ruolo importante
Letalità delle tossine proteiche in confronto a quelle di alcuni veleni
TossinaDose
Tossica (mg/Kg)
ospite (X) Rispetto alla dose tossica di:
stricnina endotossine Veleno di serpente
botulinica 0,8 x 10-8 topo 3 x 106 3 x 107 3 x 105
tetanica 4,0 x 10-8 topo 1 x 106 1 x 107 1 x 105
NeurotossinaShigella 2,3 x 10-6 coniglio 1 x 106 1 x 107 1 x 105
difterica 6,0 x 10-5 cavia 2 x 103 2 x 104 2 x 102

Le tossine proteiche sono fortemente antigeniche
ANTITOSSINE
TOSSOIDI
ANATOSSINE
anticorpi specifici che possono bloccarne l’attività
Esotossine che hanno perso l’attività tossica per invecchiamento; restano antigeni vaccini
Tossoidi ottenuti con trattamenti artificiali (vaccini antitetanico e antidifterico)

subunità A: responsabile dell’attività biologica
BA
B
MOLTE TOSSINE HANNO UNA STRUTTURA A,B
La componente biologica della tossina resta inattiva finchè non si libera dalla tossina nativa
A
subunità B: lega il recettore,e trasporta “A” nella cellula

A/B: tossina sintetizzata in un singolo polipeptide, dominio A e il dominio B coesistono e sono separati da una proteasi
Sito di taglio
COOH
NH2
S S
A+B tossina sintetizzata e secreta in subunità indipendenti che interagiscono direttamente sul bersaglio
Uno dei componenti della tossina dell’antrace si lega alla membrana della cellula e lega uno degli altri due componenti
LF
PA63
PA63
EF

A-B : le subunità sono sintetizzate separatamente ma si associano prima del legame al loro bersaglio
A
“A-5B” indica che il dominio di legame è formato da 5 subunità identiche
Es. enterotossina di V. choleraee altre enterotossine
La maggior parte delle tossine di tipo A/B è esportata attraverso il sistema di secrezione di tipo II

Tossine che agiscono nella cellula:
la subunità B si lega al recettore e forma un poro attraverso cui la subunità A entra nel citoplasma
(es. Colera)
INGRESSO DIRETTO
INGRESSOINDIRETTO
la tossina nativa entra nella cellula in un endosoma. Il pH si abbassa causando la
separazione di A e B; la subunità B provoca il rilascio di A nel citoplasma attraverso la
membrana dell’endosoma (es. Antrace)
AA

Ogni esotossina ha un meccanismo d’azione caratteristico
La tossina tetanica e la tossina botulinica hanno struttura simile e agiscono entrambe sul tessuto nervoso
Ma provocano effetti opposti
TETANO BOTULISMO
Blocco dei neuroni inibitori
Paralisi spastica
Blocco della placca motrice
Paralisi flaccida

DIFTERITE
ADP-RIBOSILAZIONE
DI EF2(fattore di
allungamento)
Blocco della sintesi proteica
Morte cellulare
COLERA
ATTIVAZIONE DELL’ADENIL-
CICLASI
Perdita massiva di elettroliti e acqua
Altre esotossine

Per essere efficaci le tossine secrete devono attraversare la membrana
Attraverso la membrana interna: -SEC (sistema generale di traslocazione)-TAT /twin arginine traslocase
Esterno
Periplasma

SEC-dipendentiT2SS, T5SS (T4SS)
SEC-indipendentiT1SS T3SS (T4SS)
Traslocazione attraverso la membrana esterna (didermi)
Traslocano proteine che sono state portate al periplasma da SEC
Traslocano proteine direttamente senza l’intervento di SEC

Le specie che modificano il citoscheletrodelle cellule dell’ospite
Shigella, Salmonella e EPEC (uno dei virotipi di E. coli)
Possiedono in genere uno o più sistemi di secrezione di tipo III (TTSS)
Citoscheletro

Salmonella: invade attivamente gli enterociti
dirigendo gli effetti sul citoscheletro dell’ospite dall’interno di un vacuolo legato alla membrana
EPEC/EHEC: restano all’esterno della cellula ospite
ma intimamente connessi alla superficie cellulare

Salmonella, Shigella E e. coli EPEC utilizzano T3SS per riarrangiare il citoscheletro di actina
Shigella: entra nelle cellule M
si replica nel citoplasma
diffonde da cellula a cellula polimerizzando l’actina dell’ospite

CagA dà due tipi di segnale alla cellula: provoca la formazione di un piedistallo per H. pylori, ed il rilascio di citochine pro-infiammatorie
Stimoli“Growth-factor-like”
Trascrizione IL-8
PolimerizzazioneActina
CagA P
Formazione del piedistallo
Un esempio di tossina secreta attraverso un T4SS è CagA di Helicobacter pylori

E. coli 6-ac. Salmonella
7-ac.Bacteroides
5-ac.Bacteroides ha una forma PENTA-acilata che non induce risposte infiammatorie e sembra antagonizzi le forme tossiche
EVASIONE DALLE DIFESE ADATTATIVE
Modificazione antigenica Es. catene LPS meno acilate, sfuggono ai TLR

In alcuni patogeni enterici i sistemi di secrezione di tipo III traslocano
nell’enterocita le proteine responsabili delle alterazioni del citoscheletro