Dipendente da contatto Autocrina Endocrina Paracrina 15-16... · De Leo -Fasano -Ginelli...
30
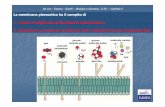
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Capitolo 5 Dipendente da contatto Segnali di posizione (migrazione dei leucociti) Paracrina Raggio breve (neurotrasmettitori) Autocrina Agisce sulla stessa cellula che ha prodotto il segnale (fattori di crescita) Endocrina Fattori rilasciati in circolazione (maggior parte degli ormoni) (eg, LH e FSH secreti dall’ipofisi, Agiscono sulle gonadi)
Transcript of Dipendente da contatto Autocrina Endocrina Paracrina 15-16... · De Leo -Fasano -Ginelli...
-
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Capitolo 5
Dipendente da contattoSegnali di posizione
(migrazione dei leucociti)
ParacrinaRaggio breve
(neurotrasmettitori)
AutocrinaAgisce sulla stessa cellula che
ha prodotto il segnale (fattori di crescita)
EndocrinaFattori rilasciati in circolazione (maggior parte degli ormoni)
(eg, LH e FSH secreti dall’ipofisi, Agiscono sulle gonadi)
-
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Capitolo 5
I recettori:
•hanno una elevata affinità di
legame con il ligando (consentendo una
risposta cellulare anche a basse
concentrazioni di ligando)
•Sono selettivamente specifici per il
loro ligando
•Sono legati reversibilmente con il
loro ligando tramite legami ad
idrogeno, Forze di van der Waals,
legami idrofobici
-
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Capitolo 5
-
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Capitolo 5
-
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Capitolo 5
-
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Capitolo 5
La fosforilazione è il meccanismo più comuneattraverso il quale le proteine vengono modificate lungo una cascata di segnali cellulari.
La fosforilazione avviene su aminoacidi che hanno un gruppo ossidrile disponibile per illegame con un gruppo fosfato (serina, treonina, tirosina).
La presenza di un gruppo fosfato introduce nella proteina una forte carica negativa con un elevato ingombro sterico. Questo causa unariorganizzazione dei legami tra aminoacidi, responsabili della struttura terziaria della proteina.
Cambia la struttura terziaria e si ha una transizione funzionale
È una modificazione reversibile. La defosforilazione è operata dalle fosfatasi
-
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Capitolo 5Ione Ca++ come secondo messaggero: Bassa concentrazione nel citoplasmaAccumulato in depositi intracellulari (RE)Lega una classe di proteine dette calmoduline (proteine regolatorie o subunitàregolatorie)
-
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Capitolo 5
Nelle miofibrille muscolariil Ca++ è immagazzinato nel ret sarcoplasmatico ed in seguito ad uno stimolo nervoso vi fuoriesce e lega una calmodulina detta troponina C che si associa ai filamenti di actina rendendoli capaci di interagire con la miosina
-
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Capitolo 5
Comunicazione tra cellule nervose (risposte elettriche)
Agiscono tramite l’azione di una classe di proteine ancorate ai lipidi (proteine G)
I recettori sono in grado di fosforilare (tirosina) i propri residui aminoacidici e quelli di altri enzimi
-
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Capitolo 5
Legano il GTPNella forma legata al GTP sono attiveHanno attività GTPasica (idrolizzano il GTP in GDP)Costituiscono una sorta di interruttori molecolari a tempo (idrolizzando il GTP ritornano allo stato inattivo, quindi si autoinattivano o autoregolano)
-
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Capitolo 5
interruttori molecolari a tempo
-
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Capitolo 5
Uno dei bersagli delle proteine G è l’adenilato ciclasi
-
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Capitolo 5
-
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Capitolo 5
PKA fosforila:
Glicogeno fosforilasi chinasi (attivandola)Glicogeno sintetasi (inibendola)Proteina fosfatasi I (inibendola)
La glicogeno fosforilasi chinasi fosforila la glicogeno fosforilasi attivandola: il glicogeno viene metabolizzato in glucosio.
La glicogeno sintetasi è responsabile della sintesi del glicogeno a partire dal glucosio
La proteina fosfatasi I defosforila la glicogeno sintetasi attivandola
Risultato finale dell’azione del glucagone (secreto dal pancreas in risposta ad una riduzione del glucosio ematico) sarà l’inibizione degli enzimi della glicogenolisi e liberazione di glucosio nel sangue
-
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Capitolo 5
Ormoni come LH, l’ormone adrenocorticotropo (ACTH) e l’ormone stimolante la tiroide (TSH) sono in grado di attivare PKA che trasloca nel nucleo e fosforila il fattore di trascrizione CREB.
CREB attivato è in grado di legarsi al promotore di alcuni geni attivando la trascrizione
Le funzioni principali di questi ormoni consistono nel determinare l’aumento della sintesi di progesterone (LH), la secrezione di cortisolo (ACTH) e la sintesi dell’ormone tiroideo (TSH)
-
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Capitolo 5
Il recettore attiva una proteina Gq che a sua volta attiva la fosfolipasi Cβ (proteina periferica) che genera l’inositolo trifosfato (IP3) ed il diacilglicerolo (DAG). Il IP3 si lega al canale del Ca++ aprendolo (diff passiva) aumentando la concentrazione del Ca nel citosol. Le subunità della PKC sono calmoduline. La PKC liberata dalle calmoduline èlibera di muoversi nel citosol, si posizionerà sul lato interno della membrana e verràattivata dal DAG
-
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Capitolo 5
-
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Capitolo 5
AMPLIFICAZIONE DEL SEGNALEA partire da poche molecole di ligando e poche molecole di recettore attivato si determina ad ogni passaggio un aumento di molecole segnale che attivano un gran numero di enzimi e quindi una risposta cellulare efficiente
Ogni molecola di recettore può reclutare numerose proteine G in successione (fin
quando è occupato dal ligando)
La proteina G attiva l’adenilato ciclasi con un rapporto stechiometrico 1:1
Adenilato ciclasi e fosfolipasi C producono grandi quantità di secondi messaggeri: per
ogni molecole di adenilato ciclasi si hanno migliaia di molecole di cAMP; l’apertura del
canale del Ca++ causa il rilascio di quantità millimolari di Ca++ nel citosol.
Le proteine chinasi PKA e PKC possono fosforilare diversi bersagli amplificando
ancora il bersaglio.
L’amplificazione del segnale è molto utile per consentire alla cellula repentine
modifiche del metabolismo.
-
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Capitolo 5
Proteine transmembrana monopasso (in genere omodimeri)Hanno attività tirosin-chinasicaCostituiscono molti recettori per fattori di crescita, differenziamento e per l’insulina
L’attivazione del recettore da parte del ligando porta all’autofosforilazione del recettore stesso. Si formano dei dimeri recettoriali: le regioni chinasiche dei due monomeri sono vicine e si possono fosforilare a vicenda.Il ligando favorisce la dimerizzazione
-
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Capitolo 5
I residui fosforilati dei recettori sono riconosciuti da proteine che possiedono domini in grado di legare le tirosine fosforilate (SH2).
La queste, le più note sono quelle della via Ras/MAPK
-
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Capitolo 5
Il recettore fosforilato viene legato dalla proteine adattatrice Grb2 che a sua volta lega
SOS.
L’assemblaggio di queste due proteine porta all’attivazione di RAS.
RAS è proteina G monomerica.
SOS induce il rilascio di GDP ed il legame con GTP con RAS necessario per l’attivazione
-
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Capitolo 5
RAS attiva una chinasi (RAF) che a sua volta attiva a cascata due chinasi: MAPKK(MEK) e MAPK (ERK). ERK attivata trasloca nel nucleo dove fosforila i suoi bersagli(fattori di trascrizione). In questo modo l’attività del recettore modifica l’espressione genicaQuesto tipo di risposta è amplificativa
-
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Capitolo 5
Recettore nicotinico dell’acetilcolina: è un canale per il sodio. Il legame dell’acetilcolina apre il canale e permette l’ingresso del Na. In questo modo sidepolarizza la membrana e si aprono altri canali del Na in maniera tale che il segnale elettrico si propaghi sino ad i tubuli T della m plasmatica delle cellule muscolari.Queste strutture sono in contatto con il reticolo sarcoplasmatico. L’arrivo dell’onda elettrica induce il rilascio di Ca++ dal ret. Sarcoplasmatico: il primo evento che porta alla contrazione muscolare.
-
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Capitolo 5
La risposta recettoriale non è
temporalmente indefinita.I segnali intracellulari sono transitori e si esauriscono in pochi minuti (anche se gli effetti possono durare parecchie ore).Autospegnimento delle proteine GPresenza di enzimi in grado di spegnere i segnali: cAMP fosfodiesterasi e fosfatasi.Una cascata di segnali attivatori è sempre associata ad una di segnali inibitori. La risposta ad uno stimolo è il risultato del dosaggio tra le due cascate di segnali.
Desensibilizzazione dei recettori dopo uno stimolo prolungato o troppo intenso
-
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Capitolo 5
GRK fosforila la porzione citoplasmatica del recettore beta-adrenergico impedendo il suo legame con la proteina G poiché viene legato dall’arrestina
-
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Capitolo 5
-
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Capitolo 5
-
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Capitolo 5
-
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Capitolo 5
-
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Capitolo 5











![(Ipofisi 2013 [modalità compatibilità]) - autistici.org - presentazioni... · Cosa fanno? • Alterano l ... nelle gonadi un’azione paracrina locale L’Inibina B è il marcatore](https://static.fdocumenti.com/doc/165x107/5c6580fb09d3f2826e8cc042/ipofisi-2013-modalita-compatibilita-presentazioni-cosa-fanno-.jpg)