
Biologia - libreriauniverso.it · Introduzione alla biologia 1 ... Reazione di formazione dei...
10
Biologia
Transcript of Biologia - libreriauniverso.it · Introduzione alla biologia 1 ... Reazione di formazione dei...

Biologia
01_AIMS_BIOLOGIA_01_001_026.indd 1 11/05/17 11:44

01_AIMS_BIOLOGIA_01_001_026.indd 2 11/05/17 11:44

1.1 GLI ELEMENTI E LE MOLECOLE FONDAMENTALI
Alcuni elementi sono talmente importanti per la sopravvivenza delle specie viventi sul nostro pianeta che vengono continuamente riciclati.
Tra questi l’elemento più diffuso è certamente il carbonio, dato che è alla base della composi-zione delle molecole organiche.
Esso viene rilasciato sotto forma di anidride carbonica, durante la fotosintesi.
L’azoto è un elemento altrettanto importante. Questo elemento è presente nell’atmosfera per circa il 78% e nel sottosuolo in forma di nitrati. Questi derivano dall’azoto atmosferico in seguito a precipitazione (pioggia).
Il fosforo è un altro elemento molto presente in natura e si ritrova in abbondanza sulle rocce sotto forma di fosfato, ossia prevalentemente legato all’ossigeno.
1.2 CARATTERISTICHE DEI VIVENTI
Vi sono una serie di caratteristiche che si ritro-vano in un essere vivente. Tra queste si ricordano:
y possibilità di riprodursi; y capacità di sviluppo e accrescimento; y continui processi evolutivi; y capacità di scambiare e trasformare l’energia;
y possesso di informazioni attraverso i geni; y capacità di rispondere alle esigenze nella
quotidianità; y formazione cellulare che garantisce il funzio-
namento dell’intero organismo.
1.3 BIOMOLECOLE
Le molecole biologiche, o biomolecole, sono la materia prima di cui sono costituiti tutti gli esseri viventi. Una cellula batterica contiene circa 5000 diversi tipi di biomolecole, mentre una cellula animale o vegetale ne contiene circa il doppio.
Si distinguono 4 classi diverse di biomole-cole: i carboidrati (o zuccheri), i lipidi (o grassi), le proteine (o protidi) e gli acidi nucleici.
1.4 CARBOIDRATI
I carboidrati, chiamati anche zuccheri o glucidi, sono la principale fonte energetica per la mag-gior parte degli esseri viventi. Sono detti carboi-drati in quanto contengono idrogeno e ossigeno combinati al carbonio nelle stesse proporzioni della molecola d’acqua.
I carboidrati si suddividono in 3 categorie (Figura 1.1):
y monosaccaridi; y disaccaridi; y polisaccaridi.
1Introduzione alla biologia
01_AIMS_BIOLOGIA_01_001_026.indd 3 11/05/17 11:44

Capitolo 1 • Introduzione alla biologia4
1.4.1 I monosaccaridi
I monosaccaridi rappresentano gli zuccheri più semplici, giacché non possono essere ulterior-mente demoliti per idrolisi; come tutti i carboi-drati, sono composti ternari polifunzionali, com-posti da carbonio, idrogeno e ossigeno.
Da un punto di vista chimico, possiamo dire che i monosaccaridi contengono molti gruppi
alcolici e un gruppo aldeidico (aldosi) o un gruppo chetonico (chetosi).
I monosaccaridi sono nominati aggiungendo al numero di atomi di carbonio presenti il suf-fisso -osio, per esempio uno zucchero semplice formato da tre atomi di carbonio si definisce triosio, invece uno formato da quattro atomi di carbonio si definisce tetrosio e così via.
Graficamente, gli zuccheri sono rappresen-tati attraverso strutture a catena aperta, defi-nite “proiezioni di Fischer”, dal chimico tedesco Hermann Emil Fischer. Esse raffigurano atomi di carbonio congiunti tra loro e ai propri sostituenti attraverso linee verticali e orizzontali. Tuttavia, in realtà, gli zuccheri esistono in catene chiuse, per cui si adottano anche le formule di Haworth, dal chimico inglese Walter Norman Haworth. Per esplicitare meglio questo concetto, pren-diamo in considerazione la molecola del glu-cosio (Figura 1.2).
ATTENZIONE: gli anomeri (a-D-Glucosio e b-D-Glucosio) hanno la medesima formula mole-colare (si definiscono stereoisomeri) e diversa posi-zione dei gruppi legati al carbonio anomerico (C-1).
LATTE
MONOSACCARIDI
DISACCARIDI
POLISACCARIDI
Glucosio
Fruttosio
Galattosio
Saccarosio
Maltosio
Lattosio
Amido
Cellulosa
Glicogeno
MIELE
Figura 1.1 Struttura dei carboidrati.
Figura 1.2 Proiezioni di Fisher e di Haworth.
01_AIMS_BIOLOGIA_01_001_026.indd 4 11/05/17 11:44
Capitolo 1 • Introduzione alla biologia 5
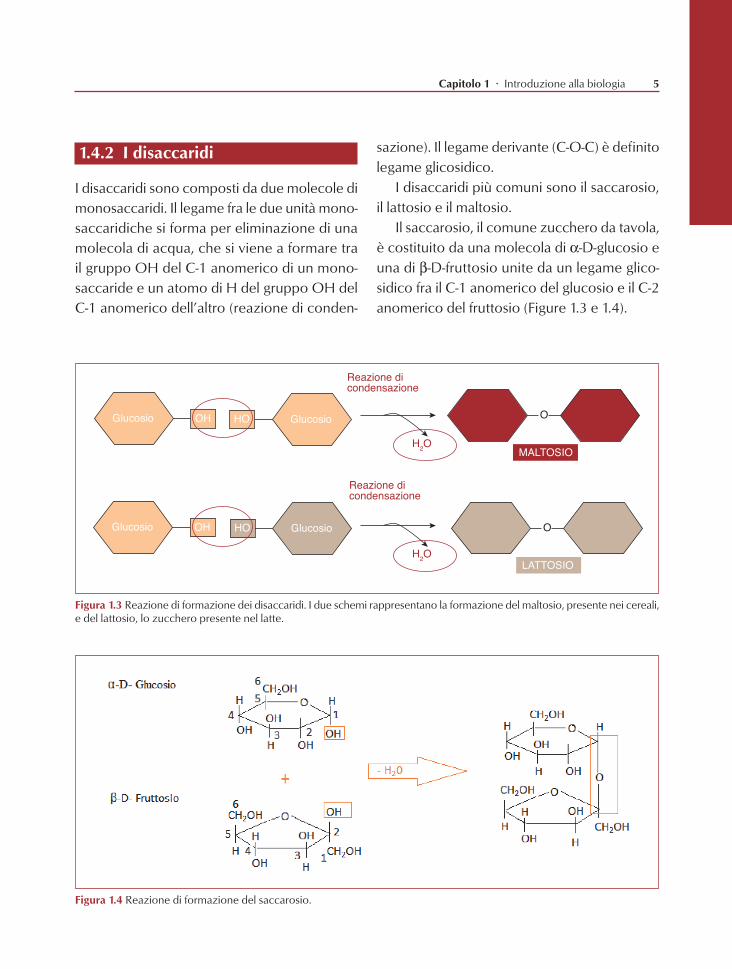
1.4.2 I disaccaridi
I disaccaridi sono composti da due molecole di monosaccaridi. Il legame fra le due unità mono-saccaridiche si forma per eliminazione di una molecola di acqua, che si viene a formare tra il gruppo OH del C-1 anomerico di un mono-saccaride e un atomo di H del gruppo OH del C-1 anomerico dell’altro (reazione di conden-
Reazione dicondensazione
Reazione dicondensazione
O
MALTOSIOH2O
H2O
O
LATTOSIO
GlucosioHOGlucosio OH
Glucosio OH GlucosioHO
Figura 1.3 Reazione di formazione dei disaccaridi. I due schemi rappresentano la formazione del maltosio, presente nei cereali, e del lattosio, lo zucchero presente nel latte.
Figura 1.4 Reazione di formazione del saccarosio.
sazione). Il legame derivante (C-O-C) è definito legame glicosidico.
I disaccaridi più comuni sono il saccarosio, il lattosio e il maltosio.
Il saccarosio, il comune zucchero da tavola, è costituito da una molecola di a-D-glucosio e una di b-D-fruttosio unite da un legame glico-sidico fra il C-1 anomerico del glucosio e il C-2 anomerico del fruttosio (Figure 1.3 e 1.4).
01_AIMS_BIOLOGIA_01_001_026.indd 5 11/05/17 11:44

Capitolo 1 • Introduzione alla biologia6
1.4.3 I polisaccaridi
I polisaccaridi sono i carboidrati più complessi e costituiscono una classe di polimeri, in cui l’unità ripetitiva, detta monomero, è il mono-saccaride.
Essi presentano catene molto lunghe, che possono essere lineari o ramificate.
Ne sono un esempio la cellulosa, l’amido e il glicogeno.
In generale, i polisaccaridi sono sostanze amorfe e insapore, che in acqua possono com-portarsi da colloidi (per esempio l’amido) o essere insolubili (come la cellulosa) in rela-zione alla loro composizione: numerosi gruppi ossidrili (polari) e lunghe catene carboniose (apolari).
I polisaccaridi si distinguono in:
y polisaccaridi di riserva, che comprendono l’amido, caratteristico delle cellule vegetali, e il glicogeno, presente nelle cellule animali;
y polisaccaridi strutturali, che servono come elementi di sostegno delle piante; il più dif-fuso è la cellulosa, costituente principale della parete delle cellule vegetali.
L’amido è presente in alimenti come le patate, il riso, il mais ecc. e viene utilizzato in diversi settori industriali come quello della carta, dei tessuti, degli adesivi e dei detersivi.
Negli ultimi anni sono stati impiegati anche materiali ottenuti dall’amido di mais, basti pen-sare ai sacchetti biodegradabili per la raccolta di rifiuti organici.
Da un punto di vista chimico, l’amido è for-mato da:
y amilosio, omopolisaccaride lineare formato da circa 300-600 molecole di glucosio unite
tra loro da legami glicosidici a-(1→4). Le cui catene lineari sono avvolte in una conforma-zione a elica;
y amilopectina, molecola ramificata formata da migliaia di molecole di glucosio, fino a 50000, unite da legami glicosidici a-(1→4). Ogni 25-30 molecole di glucosio è presente una ramificazione, anch’essa costituita da molecole di glucosio, unita alla catena prin-cipale da un legame glicosidico a-(1→6).
Quindi possiamo dire semplicemente che l’a-mido è un polisaccaride sintetizzato in natura a partire dal glucosio.
In acqua, per idrolisi, si scinde in maltosio (disaccaride) e glucosio. Per questo motivo rap-presenta per gli organismi vegetali una riserva di glucosio.
Il glicogeno è un polimero del glucosio, di costituzione molto simile a quella dell’amilo-pectina, e rappresenta una riserva di energia per gli animali, giacché da esso è possibile liberare molecole di glucosio mediante idrolisi. Infatti, quando nel circolo sanguigno il glucosio è pre-sente in una concentrazione eccessiva, questa viene ridotta formando glicogeno. Questo è poi immagazzinato nel fegato e nei muscoli e, quando il corpo ha bisogno di energia, viene degradato nuovamente in glucosio, che così rientra in circolo.
La cellulosa è un polisaccaride formato da lunghe e sottili catene parallele, unite fra loro mediante legami a idrogeno. Le unità mono-saccaridiche di cui è composta sono con-giunte mediante legami b(1→4) glucosidici. Quest’ultima caratteristica rende la cellulosa indigeribile da parte dell’uomo, non avendo quest’ultimo gli enzimi necessari a scindere questo tipo di legame.
01_AIMS_BIOLOGIA_01_001_026.indd 6 11/05/17 11:44

Capitolo 1 • Introduzione alla biologia 7
1.5 PROTEINE
Le proteine sono le macromolecole più abbon-danti e sono presenti in tutte le cellule e in tutti i compartimenti cellulari. In una cellula esistono migliaia di differenti proteine, diverse fra loro per dimensione e funzione biologica. Esse rappresen-tano lo strumento attraverso cui l’informazione genetica è espressa. Le proprietà e le attività svolte dalle proteine possono essere molteplici:
y catalitica: molte proteine sono enzimi, cioè sostanze la cui presenza assicura che siano realizzate numerose reazioni che, altrimenti, avverrebbero in tempi eccessivamente lunghi;
y regolatrice: gli ormoni ne sono un esempio; y difensiva: gli anticorpi fanno parte del sistema
immunologico e sono fondamentali nella difesa dell’organismo da microrganismi patogeni;
y strutturale: ne sono un esempio collagene, cheratina, elastina. Si tratta di un supporto meccanico.
Le proteine sono prodotte dalle cellule partendo da soli 20 amminoacidi.
Gli amminoacidi essenziali sono otto: valina, leucina, isoleucina, treonina, metionina, fenila-lanina, triptofano, lisina.
In generale, possiamo suddividere gli ammi-noacidi in tre gruppi: 1) amminoacidi essenziali, che l’organismo non è in grado di produrre e quindi sono introdotti attraverso l’alimentazione; 2) amminoacidi semiessenziali, la cui produzione è insufficiente in relazione al fabbisogno, per cui in determinate situazione risulta necessario inte-grarli con la dieta; 3) amminoacidi non essenziali, che sono normalmente sintetizzati e presenti nelle giuste quantità.
È possibile riconoscere una struttura comune a tutti gli amminoacidi: un gruppo carbossilico e
un gruppo amminico legati allo stesso atomo di carbonio (detto carbonio-a) e una catena late-rale (detta gruppo R) che influenza le proprietà dell’amminoacido stesso (Figura 1.5).
In un amminoacido possiamo distinguere due componenti: una acida (gruppo carbossi-lico) e una basica (gruppo amminico), per cui un amminoacido si comporta da “zwitterione” (o ione dipolare) (Figura 1.6).
In relazione all’intorno chimico, un amminoa-cido quindi può presentarsi o in forma anionica o in forma cationica o in forma zwitterionica. Inoltre, non bisogna dimenticare che al C-a è legato anche il gruppo R, per cui questo influen-zerà il comportamento dei gruppi carbossilico e amminico. Il valore di pH a cui una molecola ha una carica netta complessiva nulla è definito punto isoelettrico.
Le proteine sono polimeri di amminoacidi, in cui ciascuna unità amminoacidica è legata alla precedente e alla successiva mediante uno specifico legame covalente: legame peptidico. Dati due amminoacidi, il legame peptidico si forma mediante reazione di condensazione tra il gruppo amminico di un amminoacido e il gruppo carbossilico dell’altro.
Catena laterale
Gruppo amminico
Gruppo carbossilico
COOHR C
NH2
H
Figura 1.5 Gruppi comuni nelle strutture degli aminoacidi.
01_AIMS_BIOLOGIA_01_001_026.indd 7 11/05/17 11:44
Capitolo 1 • Introduzione alla biologia8
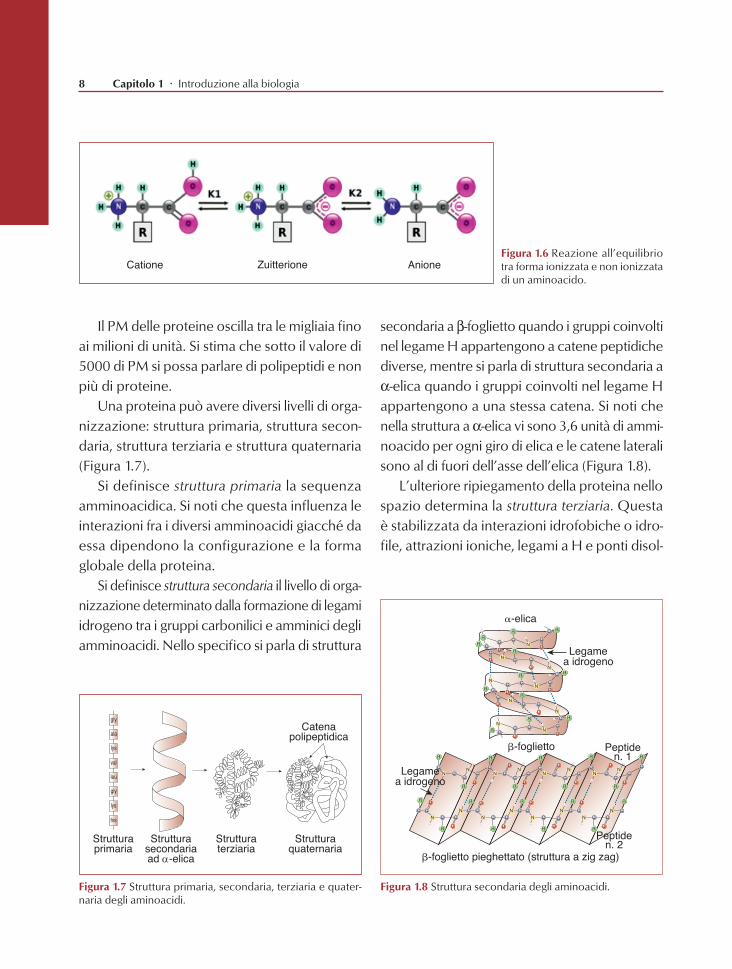
Il PM delle proteine oscilla tra le migliaia fino ai milioni di unità. Si stima che sotto il valore di 5000 di PM si possa parlare di polipeptidi e non più di proteine.
Una proteina può avere diversi livelli di orga-nizzazione: struttura primaria, struttura secon-daria, struttura terziaria e struttura quaternaria (Figura 1.7).
Si definisce struttura primaria la sequenza amminoacidica. Si noti che questa influenza le interazioni fra i diversi amminoacidi giacché da essa dipendono la configurazione e la forma globale della proteina.
Si definisce struttura secondaria il livello di orga-nizzazione determinato dalla formazione di legami idrogeno tra i gruppi carbonilici e amminici degli amminoacidi. Nello specifico si parla di struttura
secondaria a b-foglietto quando i gruppi coinvolti nel legame H appartengono a catene peptidiche diverse, mentre si parla di struttura secondaria a a-elica quando i gruppi coinvolti nel legame H appartengono a una stessa catena. Si noti che nella struttura a a-elica vi sono 3,6 unità di ammi-noacido per ogni giro di elica e le catene laterali sono al di fuori dell’asse dell’elica (Figura 1.8).
L’ulteriore ripiegamento della proteina nello spazio determina la struttura terziaria. Questa è stabilizzata da interazioni idrofobiche o idro-file, attrazioni ioniche, legami a H e ponti disol-
Figura 1.6 Reazione all’equilibrio tra forma ionizzata e non ionizzata di un aminoacido.
Strutturaprimaria
Strutturasecondariaad α-elica
Strutturaterziaria
Strutturaquaternaria
gly
ala
lys
val
leu
gly
lys
his
Catenapolipeptidica
Figura 1.7 Struttura primaria, secondaria, terziaria e quater-naria degli aminoacidi.
�-foglietto pieghettato (struttura a zig zag)
R
R
R
R
R
R
R
R
R
R
R
R
R
R R
R
R
N
NN
N
N
N
N
N
N
N
N
N
N
N
ND
D
D
DN
D
D
D
D
D
D
D
D
D
D
DR
CC
C
CCC C
CC
C CC
CC
CC C
CC
C C
C C
CC
CC
C CC C
C
CC
Peptiden. 2
Peptiden. 1
Legamea idrogeno
�-elica
Legamea idrogeno
RN
R
C
R
D
C
NC
C
RR
N
DCC
N
DC
DC
D
RN CC
DCC N
RND
C
R
ND
CCD
R
C C
RR
NC C
C
D
RC
�-foglietto
Figura 1.8 Struttura secondaria degli aminoacidi.
Catione Zuitterione Anione
01_AIMS_BIOLOGIA_01_001_026.indd 8 11/05/17 11:44

Capitolo 1 • Introduzione alla biologia 9
furo. Inoltre, le strutture terziarie possono essere anche distinte in globulari o fibrose.
L’aggregazione di due o più catene polipep-tidiche, ognuna con struttura terziaria, mediante interazioni deboli o forti determina la struttura quaternaria. Ne sono esempio l’emoglobina, pro-teina globulare a struttura quaternaria, e il col-lagene, proteina fibrosa a struttura quaternaria.
1.6 PEPTIDI E PROTEINE
I peptidi sono molecole a basso PM, costituiti generalmente da meno di 50 residui amminoa-cidici, rigorosamente codificati dal codice gene-tico. A differenza delle proteine, i peptidi non presentano una configurazione tridimensionale ben definita.
Le tecniche di purificazione adottate per lo studio di una miscela di peptidi sono:
y cromatografia a scambio ionico, in cui il mec-canismo di separazione è basato sulla diversa affinità dei diversi soluti (ionici) nei confronti dei gruppi attivi della resina;
y cromatografia di esclusione molecolare, basata sulla diversa dimensione dei peptidi. La fase stazionaria, ovvero la resina, è formata da gra-nuli di gel contenenti pori di dimensioni note:
– dialisi, in cui una soluzione concentrata è separata dal solvente mediante una mem-brana semipermeabile: in questo modo solo i peptidi più piccoli potranno diffon-dere attraverso i pori della membrana;
– elettroforesi, consistente nella migrazione di ioni in un campo elettrico, in cui di norma si adotta il gel di poliacrilammide come supporto. Questa tecnica permette di separare le proteine in base alle dimen-sioni e alla carica.
1.7 LA DENATURAZIONE PROTEICA
Una proteina si definisce denaturata quando subisce una modifica strutturale importante che influisce sulla sua funzione. Le condizioni che inducono questo cambiamento possono essere:
y termiche, ossia estremo caldo o freddo, raggi UV o agitazione magnetica;
y chimiche, ossia utilizzo di sostanze chimiche quali per esempiogli acidi.
1.8 ENZIMI
Gli enzimi sono proteine capaci di catalizzare le reazioni chimiche che avvengono all’interno degli organismi viventi. Le modalità con cui ope-rano possono essere molteplici, in linea generale possiamo descrivere il processo catalitico come la formazione del complesso enzima-substrato, così espressa:
E + S ↔ ES ↔ EP ↔ E + P
Il substrato (S) si lega all’enzima (E) a livello del sito catalitico e forma il complesso enzima-sub-strato (ES), attraverso il quale il substrato viene trasformato in prodotto (EP). Il prodotto così formato viene rilasciato dall’enzima (P + E) che è così disponibile a interagire con un’altra mole-cola di substrato. In questo processo, l’enzima né viene consumato né si altera.
La reazione catalitica di un enzima può assol-vere a due funzioni: aumentare la velocità di reazione (che può essere a seconda dei casi anche 1014 volte superiore) e/o permettere una reazione che normalmente non avverrebbe. Il principio alla base dell’aumento della velocità di reazione è l’abbassamento dell’energia di atti-
01_AIMS_BIOLOGIA_01_001_026.indd 9 11/05/17 11:44

Capitolo 1 • Introduzione alla biologia10
vazione della reazione, come si può notare nel grafico (Figura 1.9).
Si noti che molti enzimi per poter funzionare richiedono un cofattore, che può essere uno ione metallico o una molecola organica più complessa (in tal caso si parla di coenzimi). Per esempio le vitamine possono essere precursori di coenzimi o fungere esse stesse da coenzimi.
1.8.1 Enzimi regolatori
La sopravvivenza cellulare dipende dalla gestione delle risorse. Affinché questa sia ottimale esi-stono delle proteine regolatrici il cui compito è per l’appunto la minimizzazione degli “sprechi” attraverso l’aumento o la diminuzione dell’atti-vità dei sistemi enzimatici a cui sono relazionate, in risposta a determinati segnali. Essi trovano attività in numerosi processi fisiologici, come per esempiola digestione, le attività ormonali, la coagulazione del sangue ecc.
L’attività di un enzima può essere regolata mediante diversi meccanismi, che possiamo rias-sumere in tre tipologie: regolazione allosterica, regolazione covalente e regolazione mediata da proteine controllo.
La regolazione allosterica dipende dal legame reversibile con uno specifico ligando (effettore allosterico), che regola l’intensità dell’attività dell’enzima, per cui è anche definito modula-tore. Esso può essere positivo, se attiva l’attività enzimatica, o negativo, se invece la inibisce.
La regolazione covalente consiste in modifi-cazioni covalenti, cioè rottura o formazione di nuovi legami, catalizzate da enzimi che conver-tono la loro forma attiva in inattiva o viceversa. Si tratta di una regolazione più lenta di quella allosterica poiché richiede alcuni minuti.
Per quanto riguarda la regolazione mediata da proteine di controllo, prendiamo per esempiola calmodulina, una proteina in grado di percepire i livelli di calcio intracellulare. Infatti, quando questa ha i propri siti occupati dagli ioni calcio, può legarsi a specifici enzimi determinandone l’attivazione.
1.9 LIPIDI
I lipidi sono macromolecole polari, per cui risul-tano essere solubili in acqua e insolubili in sol-venti apolari. Da un punto di vista funzionale, si distinguono in:
y lipidi di deposito, che hanno funzione ener-getica e protettiva e sono rappresentati prin-cipalmente da trigliceridi;
y lipidi strutturali, che sono i costituenti fonda-mentali delle membrane (fosfolipidi, glicoli-pidi e colesterolo).
Convenzionalmente, tuttavia, vengono classifi-cati in base alla presenza o assenza di acidi grassi nella molecola. Pertanto possiamo suddividerli in:
y lipidi semplici, quando non contengono mole-cole di acidi grassi legate covalentemente (terpeni e steroidi);
Coordinata di reazione
Ener
gia
liber
a
Ea
Ea
Reazionecatalizzata
∆G
Prodotti
Reazionenoncatalizzata
Reagenti
Non c’è differenza nell’energia liberain gioco tra reazionicatalizzate e non catalizzate
Una reazione catalizzatapossiede energiadi attivazione minore
Una reazione non catalizzata possiede energia di attivazionemaggiore di quelladella stessa reazione catalizzata
Figura 1.9 Confronto dell’energia di attivazione in presenza e in assenza dell’enzima.
01_AIMS_BIOLOGIA_01_001_026.indd 10 11/05/17 11:44


















