B12. Metabolismo dei grassi - gsartor.org · Lipidi •Semplici –Sono molecole che non contengono...
82
1 Metabolismo dei grassi Metabolismo dei grassi Prof. Giorgio Sartor Copyright © 2001-2017 by Giorgio Sartor. All rights reserved. B12 - Versione 1.6.2 – Apr-17 B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi -2- Lipidi • Semplici – Sono molecole che non contengono legami esterei o amidici • Acidi grassi • Colesterolo • Complessi – Sono derivati di acidi grassi variamente esterificati o amidati. • Glicerofosfolipidi e sfingosidi • Trigliceridi
Transcript of B12. Metabolismo dei grassi - gsartor.org · Lipidi •Semplici –Sono molecole che non contengono...

1
Metabolismo dei grassiMetabolismo dei grassi
Prof. Giorgio Sartor
Copyright © 2001-2017 by Giorgio Sartor.
All rights reserved. B12 - Versione 1.6.2 – Apr-17
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 2 -
Lipidi
• Semplici– Sono molecole che non contengono legami
esterei o amidici• Acidi grassi• Colesterolo
• Complessi– Sono derivati di acidi grassi variamente
esterificati o amidati.• Glicerofosfolipidi e sfingosidi• Trigliceridi

2
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 3 -
Metabolismo dei grassi
1. Demolizione dei trigliceridi2. Catabolismo degli acidi grassi
3. Biosintesi – degli acidi grassi e colesterolo– dei lipidi complessi
4. Trasporto dei lipidi
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 4 -
I grassi (esteri del glicerolo)
CH3
H
H
OH
H O
H OCH3
O
O
O CH3C1
C2
C3
O
OCH3
OCH3
O
P
O
OO
ORNH2
OH
O
O
N+
OH
CH3 CH3
CH3
OH
OH
OH
OH
OH
OH
NH3
+OH
CH2OH
CH2
HOH
OH
1
2
3
N+
CH3CH3
CH3
OH
OCH3
O
P
O
OO
O
1
2
3
• Trigliceridi (TAG)
• Fosfolipidi
• Lisofosfolipidi

3
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 5 -
I grassi (esteri della sfingosina)
OHNH2
OH CH3
ONH
OH CH3
O
CH3
POO
O
N+
CH3CH3
CH3
• Sfingosina
• Ceramidi
• SfingomielinaOH
NH
OH CH3
O
CH3
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 6 -
Metabolismo dei grassi
1. Demolizione dei trigliceridi2. Catabolismo degli acidi grassi
3. Biosintesi – degli acidi grassi e colesterolo– dei lipidi complessi
4. Trasporto dei lipidi

4
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 7 -
Metabolismo dei grassi
• I grassi sono fra le principali fonti di energia metabolica
3602016Glucosio
11207016Glicogeno (muscolo)
192012016Glicogeno (fegato)
102000600017Proteine
5550001500037Grassi
Energia (kJ)g/70KgEnergia (kJ·mole-1)
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 8 -
Digestione dei grassi
CH3
H
H
OH
H O
H OCH3
O
O
O CH3C1
C2
C3
• A livello intestinale i grassi (triacilgliceroli, TAG) sono idrolizzati in C1 e C3 da lipasi pancreatiche mentre i C2 sono idrolizzati da lipasi intestinali

5
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 9 -
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 10 -
Lipasi• Catalizza l’idrolisi dei triacilgliceroli in posizione 1 e 3
formando 1,2-diacilgliceroli, 2-acilglicerolo e quindi glicerolo
CH3
H
H
OH
H O
H OCH3
O
O
O CH3
CH3
H
H
OH
H O
H OH
O
O CH3
CH3
O
O
CH3
H
H
OH
H OH
H OH
OHH
OH
HH
OH
H
OH
CH3
O
O
CH3
O
O
C1
C2
C3
H2O
H2O
H2O
6
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 11 -
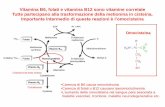
Lipolisi: meccanismo generale
R'
O
O
R"
R
Nu:
H
R'
O
O
R"
R
Nu+
H
R'
O
R
Nu O
R"H
R'
O
O O
R"H
R
Nu:H
H2O
H+
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 12 -
Triacilglicerolo lipasi EC 3.1.1.3 Ormone
(Adrenalina o glucagone)
Adenilatociclasi
(inattiva)
Adenilato ciclasi(attiva)
Fosfodiesterasi
AMP
Fosfatasi
Pi
Fosfatasi
Pi
ATP
ADPTriacilglicerololipasi
(b-inattiva)
Triacilglicerololipasi
(a-attiva)
P
P
Fosforilasi chinasi(b-inattiva)
Fosforilasi chinasi(a-attiva)
ATP
ADP
ATP cAMP + PPiProteinachinasi A(inattiva)
Proteinachinasi A(attiva)
Gα-GTP

7
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 13 -
Triacilglicerolo lipasi EC 3.1.1.3 Ormone
(Adrenalina o glucagone)
Adenilatociclasi
(inattiva)
Adenilato ciclasi(attiva)
Fosfodiesterasi
AMP
FosfatasiPi
Fosfatasi
Pi
ATP
ADPTriacilglicerololipasi
(b-inattiva)
Triacilglicerololipasi
(a-attiva)
P• Forma “a” attiva ed
indipendente da effettori allosterici
• Forma “b” meno attiva e dipendente da fattori allosterici
P
Fosforilasi chinasi(b-inattiva)
Fosforilasi chinasi(a-attiva)
ATP
ADP
ATP cAMP + PPiProteinachinasi A(inattiva)
Proteinachinasi A(attiva)
Gα-GTP
Insulina
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 14 -
Triacilglicerolo lipasi EC 3.1.1.3
• Triacilglicerolo lipasi da PseudomonasAeruginosacomplessata con un omologo di un trigliceride.
• Possiede un dominio legante Ca++

8
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 15 -
Detergenti
• L’accessibilità dei legami esterei dei TAG è favorita dalla presenza di sali di acili biliari che funzionano come emulsionanti:
OHOH
OH
CH3
CH3
CH3
CH3O
O
R
O
NH S
O
OO
R
O
NH
O
O
Acido colico (colato)
Acido taurocolico
Acido glicocolico
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 16 -
Fosfolipasi
N+
CH3CH3
CH3
O
OCH3
O
P
O
OO
O
O
CH31
2
3
PLA1
PLCPLD
PLA2

9
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 17 -
DESTINO DEI PRODOTTI
MECCANISMOFOSFOLIPASI
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 18 -
Fosfolipasi
• Fosfolipasi A1 EC 3.1.1.32• Fosfolipasi A2 EC 3.1.1.4• Fosfolipasi C EC 3.1.4.3 • Fosfolipasi D EC 3.1.4.4
• Lipasi EC 3.1.1.3
10
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 19 -
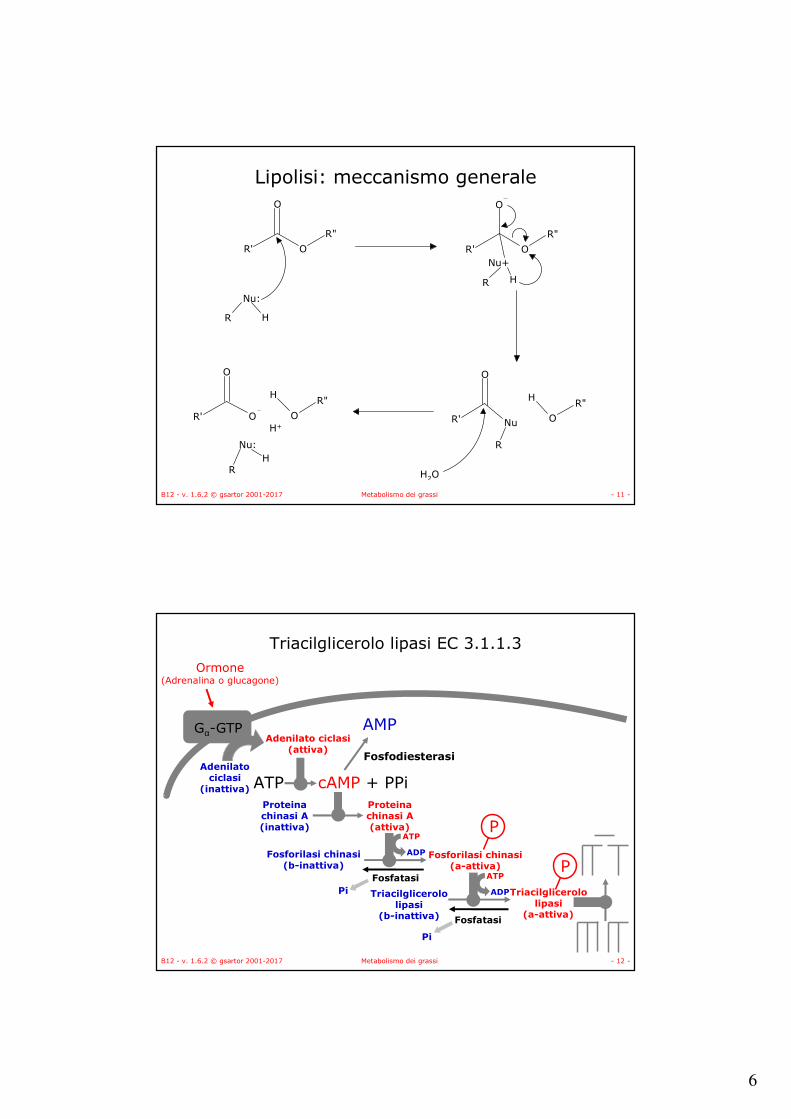
Fosfolipasi A1 (EC 3.1.1.32 )
N+
CH3CH3
CH3
O
OH
P
O
OO
O
CH3O
N+
CH3CH3
CH3
O
OCH3
O
P
O
OO
O
O
CH3
O
O
CH3
1
2
3
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 20 -
Fosfolipasi A1
1QD6
HIS
Ca++
ASP

11
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 21 -
Meccanismo (diade catalitica)
OO N
N
H
H O
H
H O
H
Ca++
O
O
R'
R"
OO N
N
H
H O
H
H
O
H
Ca++
O
O
R'
R"
OO N
N
H
H O
H
O
Ca++
O
R"
O
R'
H
Asp99
His48
Asp99
His48
Asp99
His48
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 22 -
Fosfolipasi A2 (EC EC 3.1.1.4)
N+
CH3CH3
CH3
OH
OCH3
O
P
O
OO
O
N+
CH3CH3
CH3
O
OCH3
O
P
O
OO
O
O
CH3
O
O
CH3
1
2
3

12
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 23 -
Fosfolipasi A2
• Il lisofosfolipide è un detergente potentissimo,• La fosfolipasi A2 è contenuta nel veleno di
serpenti (cobra), insetti (api), probabilmente anche in invertebrati marini (spugne), l’effetto è quello di lisare i globuli rossi attraverso l’effetto della lisofosfatidilcolina
• Nei mammiferi è secreta dal pancreas e una piccola quantità di lecitina viene secreta dal fegato, e quindi idrolizzata, per aiutare la solubilizzazione dei grassi.
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 24 -
Fosfolipasi A2
• Esistono almeno due famiglie di PLA2: –sPLA2: secreta, che permette l’idrolisi
dei fosfolipidi all’interfaccia, e possiede un dominio che si lega alla membrana (veleno delle api)
– cPLA2: citosolica, che viene utilizzata per la produzione di acidi grassi (arachidonato), inositolo fosfato come messaggeri intracellulari.

13
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 25 -
Fosfolipasi A2 (EC 3.1.1.4)
1S8G
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 26 -

14
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 27 -
Diade catalitica
ASP99
Ca++
HIS481FXF
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 28 -

15
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 29 -
Ipotetico modo d’azione di Fosfolipasi A2 su di una micella di fosfolipidi
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 30 -
FABS

16
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 31 -
FABS
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 32 -
Destino dei prodotti
• Glicerolo– Convertito in diidrossiacetonfosfato entra nella glicolisi– La conversione è catalizzata da:
• Glicerolo fosfato deidrogenasi • Diidrossiacetone chinasi
• Acidi grassi– Ossidazione
• Principalmente β-ossidazione– Produzione Acetil-CoA, NADH, FADH2
• Oppure ω-ossidazione– Processo aspecifico che porta alla produzione di composti
idrosolubili più facili da eliminare.

17
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 33 -
Glicerolo
• L’ossidazione del glicerolo a diidrossiacetone ècatalizzata dalla glicerolo fosfato deidrogenasi (EC 1.1.1.6)
1JQ5
OHOH
OH
NADH
O
OH
OHNAD+
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 34 -
Glicerolo
O
OH
OH PO
O
O
O
O
OH
ATPADP
• La fosforilazione del diidrossiacetone è catalizzata dalla diidrossiacetone chinasi (EC 2.7.1.29).
• Il diidrossiacetonfosfato cosìformatosi è convogliato nella glicolisi
1UOE DiidrossiacetoneOmologo ATP
Mg++

18
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 35 -
Glicerolo
O
OH
OH PO
O
O
O
O
OH
ATPADP
• La fosforilazione del diidrossiacetone è catalizzata dalla diidrossiacetone chinasi (EC 2.7.1.29).
• Il diidrossiacetonfosfato cosìformatosi è convogliato nella glicolisi
1UOEDiidrossiacetone Omologo ATP
Mg++
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 36 -
Lipidi
• Semplici– Sono molecole che non contengono legami
esterei o amidici• Acidi grassi• Colesterolo
• Complessi– Sono derivati di acidi grassi variamente
esterificati o amidati.• Glicerofosfolipidi e sfingosidi• Trigliceridi

19
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 37 -
Metabolismo dei grassi
1. Demolizione dei trigliceridi2. Catabolismo degli acidi grassi
3. Biosintesi – degli acidi grassi e colesterolo– dei lipidi complessi
4. Trasporto dei lipidi
β-ossidazione degli acidi grassi

20
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 39 -
Acidi grassi
• Acidi grassi saturi
• Acidi grassi insaturi
• Acidi grassi polinsaturi
O
O
CH31
23
4 67
89
1011
12 1415
1617
185 13
CH3O
O
12
34 6
78
9
5
10
1112 14
1516
17
18
13
CH3O
O
12
34 6
78
9
5
10
11
12
1415
1617
18
13
Acido stearico (C18)
Acido oleico (C18∆9)C18:1ω9
Acido linoleico (C18∆9,12)C18:2ω6
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 40 -
β-ossidazione acidi grassi
• Una volta che i TGA sono idrolizzati ad acidi grassi questi ultimi vengono demoliti fondamentalmente attraverso l’ossidazione,
• Il principale meccanismo è quello della β-ossidazione che distacca unità bicarboniose, sottoforma di Acetil-CoA, rompendo il legame tra il Cα e il Cβ,
O
O
CH3
H H
H H
O
O
CH3
H H
H H
S CH3
O
CoA
αβ
αβ

21
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 41 -
β-ossidazione acidi grassi
• A seconda del numero (n) di atomi di carbonio dell’acido grasso di producono n/2 molecole di Acetil-CoA.
• La reazione avviene nella matrice mitocondriale.
• Gli acidi grassi vengo attivati attraverso la formazione di un Acil-CoA.
• L’Acil-CoA viene trasportato all’interno della membrana interna mitocondriale attraverso il sistema carnitina/acil-carnitina.
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 42 -
Attivazione degli acidi grassi
• L’attivazione degli acidi grassi porta alla formazione di Acil-Coa
• Avviene nel citoplasma ad opera di una l’acil-CoA sintetasi(EC 6.2.1.3).
• Il processo è endoergonico(∆G = 31.4 kJ·mole-1)
• Viene reso spontaneo dalla razione di idrolisi di una molecola di ATP a AMP e PPi, il quale fornisce un surplus di energia convertendosi a 2Pi attraverso una pirofosfatasi, per un totale di -65.9 kJ·mole-1
di ATP impiegato.
CH3O
O
CH3S
OCoA
ATP
AMP + PPi
CoASH
2Pi
∆G = + 31.4 kJ mol-1 ∆G = - 32.3 kJ mol-1
∆G = - 33.6 kJ mol-1
1V25 1V26

22
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 43 -
Formazione di Acil-CoA
O
O CH3n
NO
N
N
OHOH
N
NH2
OP
O
OO
P
O
OO
P
O
OO
OP
O
OO
P
O
OO
OP
O
OHO
O
O
CH3 n
NO
N
N
OHOH
N
NH2
OP
O
O
CoA
S
H
O
O
CH3 n
NO
N
N
OHOH
N
NH2
OP
O
OS
CoA
O
CH3S
CoA
n
NO
N
N
OHOH
N
NH2
OP
O
OO
2
Aciladenosina
Intermedio tetraedrico
Acil-CoA
+
AMP
Acido grasso
ATP
+
PPi Pi
• Il meccanismo di reazione prevede la formazione di un derivato intermedio aciladenosina, legato all’enzima, che subisce l’attacco nucleofilo da parte dell’atomo di zolfo del CoA-SH.
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 44 -
Trasporto
• Il trasporto degli acilCoA nell’interno degli organelli subcellulari, dove avviene l’ossidazione degli acidi grassi, sfrutta il sistema carnitina/acilcarnitina.
O N+
OHO
CH3
CH 3
CH3
H

23
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 45 -
Carnitina• Il sistema carnitina/acilcarnitina è stato scoperto la
prima volta nei mitocondri. • Esso contiene carnitina acetiltransferasi (CAT), legata
strettamente alla faccia interna della membrana interna mitocondriale e quattro isoenzimi carnitina palmitoiltransferasi (CPT, CPT-IA nel fegato, CPT-IB nel muscolo e altre cellule, CPT-IC nel cervello e CPT-II).
• Gli enzimi CPT-I sono localizzati nella membrana esterna mitocondriale con le loro porzioni catalitica e regolatoria (inibita da malonilCoA) che si affacciano verso il lato citoplasmatico mentre CPT II è localizzata come la CAT.
• Tale varietà isoenzimatica è legata alla diversa affinitàper i diversi gruppi acilici:– CAT: C2-C10, CPTI: C6-C20– CPTII: C6-C18
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 46 -
Carnitina
• Anche i perossisomi contengono una CAT, – la carnitina octiltransferasi (CROT) che ha un’affinità
maggiore per C4-C16, è inibita da malonilCoA– ed un’altra acilcarnitina transferasi solubile non
inibita da malonilCoA.
• Anche altre membrane (reticolo endoplasmatico, sarcoplasmatico, membrana nucleare e membrana plasmatica) contengono carnitina aciltransferasi.

24
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 47 -
Trasporto nei mitocondri
• Il trasporto di Acil-Coaattraverso la membrana interna mitocondriale viene quindi mediato dalla carnitina attraverso tre sistemi enzimatici:A. Carnitina aciltransferasi I,
nel lato citoplasmatico,– CAT, CPT-IA nel fegato,
CPT-IB nel muscolo e altre cellule, CPT-IC nel cervello.
B. Carnitina acilcarnitinatraslocasi (CACT), nella membrana e
C. Carnitina aciltransferasi II, nel lato della matrice mitocondriale.
– CPT-II
Matrice
Spazio intermembrana
CPTI
CACT
Carnitina
Citoplasma
O-acilcarnitina
CPTII
Acil-CoA
O-acilcarnitina CoA-SH
Carnitina
Carnitina
CoA-SHAcil-CoA
+ +
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 48 -
Trasporto nei mitocondri
Matrice
Spazio intermembrana
CPTI
CACT
Carnitina
Citoplasma
O-acilcarnitina
CPTII
Acil-CoA
O-acilcarnitina CoA-SH
Carnitina
Carnitina
CoA-SHAcil-CoA
+ +Acetil-CoA Carnitina+
β-ossidazione
CAT
O-acetilcarnitina
CoA-SH
+
O-acetilcarnitina
O-acetilcarnitina
CACT
Carnitina
Carnitina

25
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 49 -
Trasporto nei perossisomi
• Nei perossisomi il trasporto di Acil-Coaattraverso la membrana viene mediato dalla carnitina attraverso:A. Carnitina aciltransferasi
I, nel lato citoplasmatico,
– CROT e solubile.
B. Carnitina acilcarnitinatraslocasi, nella membrana e
C. Carnitina aciltransferasiII, all’interno.
– CPT-II
Interno del perossisoma
CitoplasmaCROT
CACT
CPTII
Carnitina + Acil-CoA CoA-SH + O-acilcarnitina
Carnitina + Acil-CoA CoA-SH + O-acilcarnitina
CPTsolubile
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 50 -
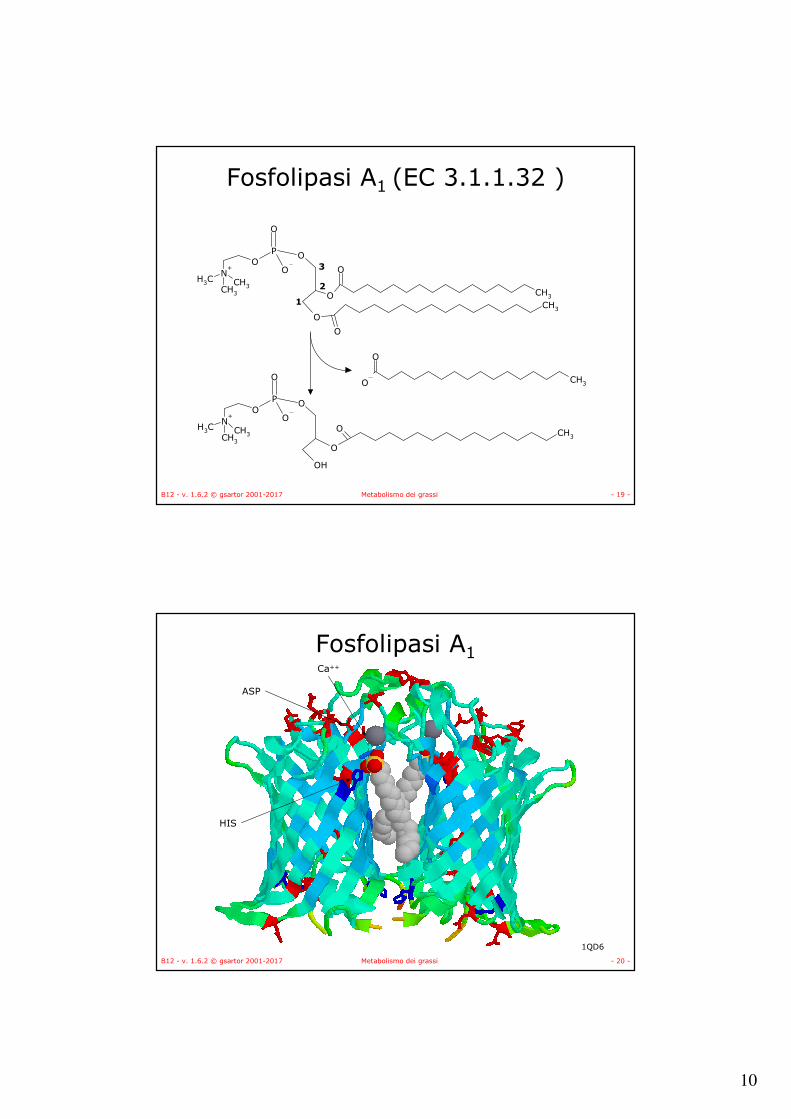
Carnitina aciltransferasi EC 2.3.1.7
S
O
CoACH3
n
O N+
OHO
CH3
CH3
CH3
SCoA H
O
CH3O
O N+
O
CH3
CH3
CH3
n
+
+
Carnitina Aciltransferasi II
(lato citoplasma)
Carnitina Aciltransferasi I
(lato matrice)

26
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 51 -
Carnitina aciltransferasi EC 2.3.1.7
Carnitina
CoA
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 52 -
Carnitina aciltransferasi EC 2.3.1.7
Carnitina

27
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 53 -
Carnitina aciltransferasi EC 2.3.1.7
CoA
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 54 -
Carnitina aciltransferasi EC 2.3.1.7
Carnitina
CoA
28
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 55 -
Carnitina aciltransferasi EC 2.3.1.7
Carnitina
CoA
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 56 -
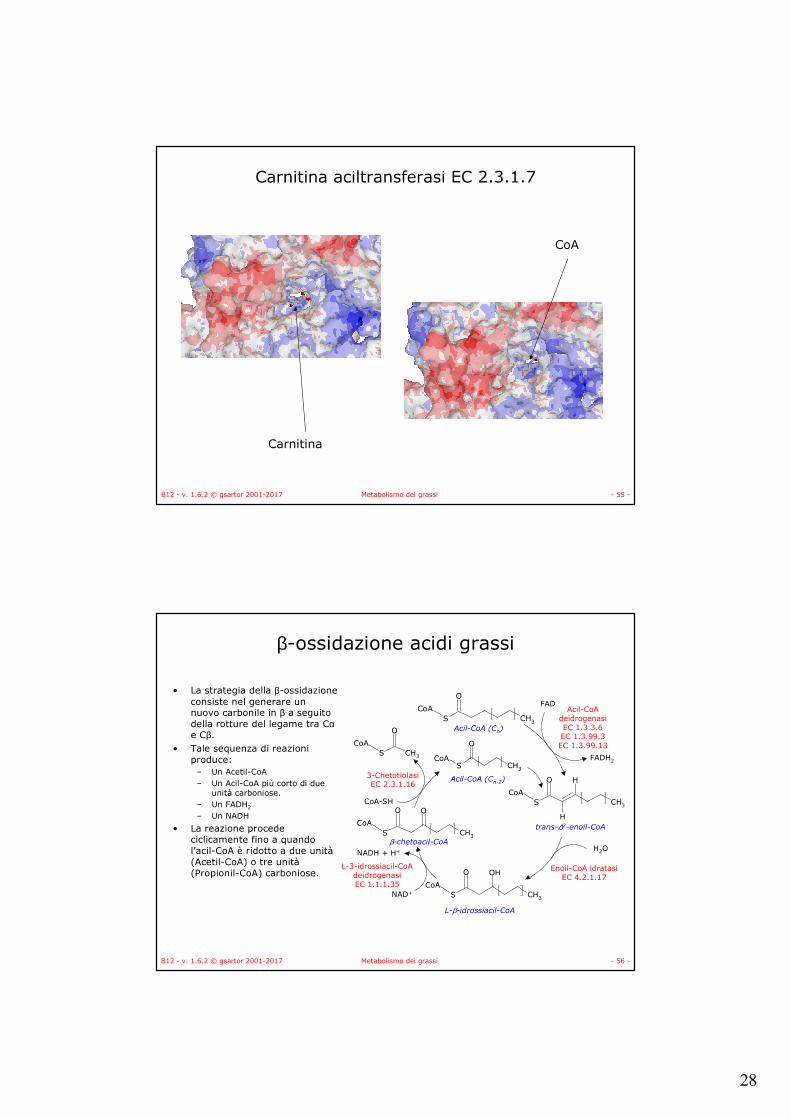
• La strategia della β-ossidazione consiste nel generare un nuovo carbonile in β a seguito della rotture del legame tra Cαe Cβ.
• Tale sequenza di reazioni produce:
– Un Acetil-CoA– Un Acil-CoA più corto di due
unità carboniose.– Un FADH2
– Un NADH
• La reazione procede ciclicamente fino a quando l’acil-CoA è ridotto a due unità(Acetil-CoA) o tre unità(Propionil-CoA) carboniose.
S
O
CoACH3
S
O
CoACH3
H
H
S
O
CoACH3
OH
S
O
CoACH3
O
S CH3
O
CoAS
O
CoACH3
FAD
FADH2
Acil-CoAdeidrogenasiEC 1.3.3.6EC 1.3.99.3EC 1.3.99.13
H2O
Enoil-CoA idratasiEC 4.2.1.17
NAD+
NADH + H+
L-3-idrossiacil-CoAdeidrogenasiEC 1.1.1.35
CoA-SH
3-ChetotiolasiEC 2.3.1.16
trans-∆2-enoil-CoA
L-β-idrossiacil-CoA
β-chetoacil-CoA
Acil-CoA (Cn)
Acil-CoA (Cn-2)
β-ossidazione acidi grassi

29
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 57 -
• La strategia della β-ossidazione consiste nel generare un nuovo carbonile in β a seguito della rotture del legame tra Cαe Cβ.
• Tale sequenza di reazioni produce:
– Un Acetil-CoA
– Un Acil-CoA più corto di due unità carboniose.
– Un FADH2
– Un NADH
• La reazione procede ciclicamente fino a quando l’acil-CoA è ridotto a due unità(Acetil-CoA) o tre unità(Propionil-CoA) carboniose.
S
O
CoACH3
S
O
CoACH3
H
H
S
O
CoACH3
OH
S
O
CoACH3
O
S CH3
O
CoAS
O
CoA
CH3
FAD
FADH2
Acil-CoAdeidrogenasiEC 1.3.3.6EC 1.3.99.3EC 1.3.99.13
H2O
Enoil-CoA idratasiEC 4.2.1.17
NAD+
NADH + H+
L-3-idrossiacil-CoAdeidrogenasiEC 1.1.1.35
CoA-SH
3-ChetotiolasiEC 2.3.1.16
trans-∆2-enoil-CoA
L-β-idrossiacil-CoA
β-chetoacil-CoA
Acil-CoA (Cn)
Acil-CoA (Cn-2)
β-ossidazione acidi grassi
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 58 -
• La strategia della β-ossidazione consiste nel generare un nuovo carbonile in β a seguito della rotture del legame tra Cαe Cβ.
• Tale sequenza di reazioni produce:
– Un Acetil-CoA– Un Acil-CoA più corto di due
unità carboniose.
– Un FADH2
– Un NADH
• La reazione procede ciclicamente fino a quando l’acil-CoA è ridotto a due unità(Acetil-CoA) o tre unità(Propionil-CoA) carboniose.
S
O
CoACH3
S
O
CoACH3
H
H
S
O
CoACH3
OH
S
O
CoACH3
O
S CH3
O
CoAS
O
CoACH3
FAD
FADH2
Acil-CoAdeidrogenasiEC 1.3.3.6EC 1.3.99.3EC 1.3.99.13
H2O
Enoil-CoA idratasiEC 4.2.1.17
NAD+
NADH + H+
L-3-idrossiacil-CoAdeidrogenasiEC 1.1.1.35
CoA-SH
3-ChetotiolasiEC 2.3.1.16
trans-∆2-enoil-CoA
L-β-idrossiacil-CoA
β-chetoacil-CoA
Acil-CoA (Cn)
Acil-CoA (Cn-2)
β-ossidazione acidi grassi

30
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 59 -
• La strategia della β-ossidazione consiste nel generare un nuovo carbonile in β a seguito della rotture del legame tra Cαe Cβ.
• Tale sequenza di reazioni produce:
– Un Acetil-CoA– Un Acil-CoA più corto di due
unità carboniose.– Un FADH2
– Un NADH
• La reazione procede ciclicamente fino a quando l’acil-CoA è ridotto a due unità(Acetil-CoA) o tre unità(Propionil-CoA) carboniose.
S
O
CoACH3
S
O
CoACH3
H
H
S
O
CoACH3
OH
S
O
CoACH3
O
S CH3
O
CoAS
O
CoACH3
FAD
FADH2
Acil-CoAdeidrogenasiEC 1.3.3.6EC 1.3.99.3EC 1.3.99.13
H2O
Enoil-CoA idratasiEC 4.2.1.17
NAD+
NADH + H+
L-3-idrossiacil-CoAdeidrogenasiEC 1.1.1.35
CoA-SH
3-ChetotiolasiEC 2.3.1.16
trans-∆2-enoil-CoA
L-β-idrossiacil-CoA
β-chetoacil-CoA
Acil-CoA (Cn)
Acil-CoA (Cn-2)
β-ossidazione acidi grassi
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 60 -
• La strategia della β-ossidazione consiste nel generare un nuovo carbonile in β a seguito della rotture del legame tra Cαe Cβ.
• Tale sequenza di reazioni produce:
– Un Acetil-CoA– Un Acil-CoA più corto di due
unità carboniose.– Un FADH2
– Un NADH
• La reazione procede ciclicamente fino a quando l’acil-CoA è ridotto a due unità(Acetil-CoA) o tre unità(Propionil-CoA) carboniose.
S
O
CoACH3
S
O
CoACH3
H
H
S
O
CoACH3
OH
S
O
CoACH3
O
S CH3
O
CoAS
O
CoACH3
FAD
FADH2
Acil-CoAdeidrogenasiEC 1.3.3.6EC 1.3.99.3EC 1.3.99.13
H2O
Enoil-CoA idratasiEC 4.2.1.17
NAD+
NADH + H+
L-3-idrossiacil-CoAdeidrogenasiEC 1.1.1.35
CoA-SH
3-ChetotiolasiEC 2.3.1.16
trans-∆2-enoil-CoA
L-β-idrossiacil-CoA
β-chetoacil-CoA
Acil-CoA (Cn)
Acil-CoA (Cn-2)
β-ossidazione acidi grassi

31
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 61 -
• La strategia della β-ossidazione consiste nel generare un nuovo carbonile in β a seguito della rotture del legame tra Cαe Cβ.
• Tale sequenza di reazioni produce:
– Un Acetil-CoA– Un Acil-CoA più corto di due
unità carboniose.– Un FADH2
– Un NADH
• La reazione procede ciclicamente fino a quando l’acil-CoA è ridotto a due unità (Acetil-CoA) o tre unità (Propionil-CoA) carboniose.
S
O
CoACH3
S
O
CoACH3
H
H
S
O
CoACH3
OH
S
O
CoACH3
O
S CH3
O
CoAS
O
CoACH3
FAD
FADH2
Acil-CoAdeidrogenasiEC 1.3.3.6EC 1.3.99.3EC 1.3.99.13
H2O
Enoil-CoA idratasiEC 4.2.1.17
NAD+
NADH + H+
L-3-idrossiacil-CoAdeidrogenasiEC 1.1.1.35
CoA-SH
3-ChetotiolasiEC 2.3.1.16
trans-∆2-enoil-CoA
L-β-idrossiacil-CoA
β-chetoacil-CoA
Acil-CoA (Cn)
Acil-CoA (Cn-2)
β-ossidazione acidi grassi
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 62 -
Destino dei prodotti
• I prodotti della β-ossidazione:– Acetil-CoA e corpi chetonici: entrano nel
ciclo di Krebs per produrre equivalenti riducenti (NADH e FADH2) che alimentano la fosforilazione ossidativa per la produzione di ATP.
– Un Acil-CoA più corto di due unitàcarboniose: rientra nel ciclo successivo di β-ossidazione.
– FADH2 e NADH che alimentano la fosforilazione ossidativa per la produzione di ATP.

32
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 63 -
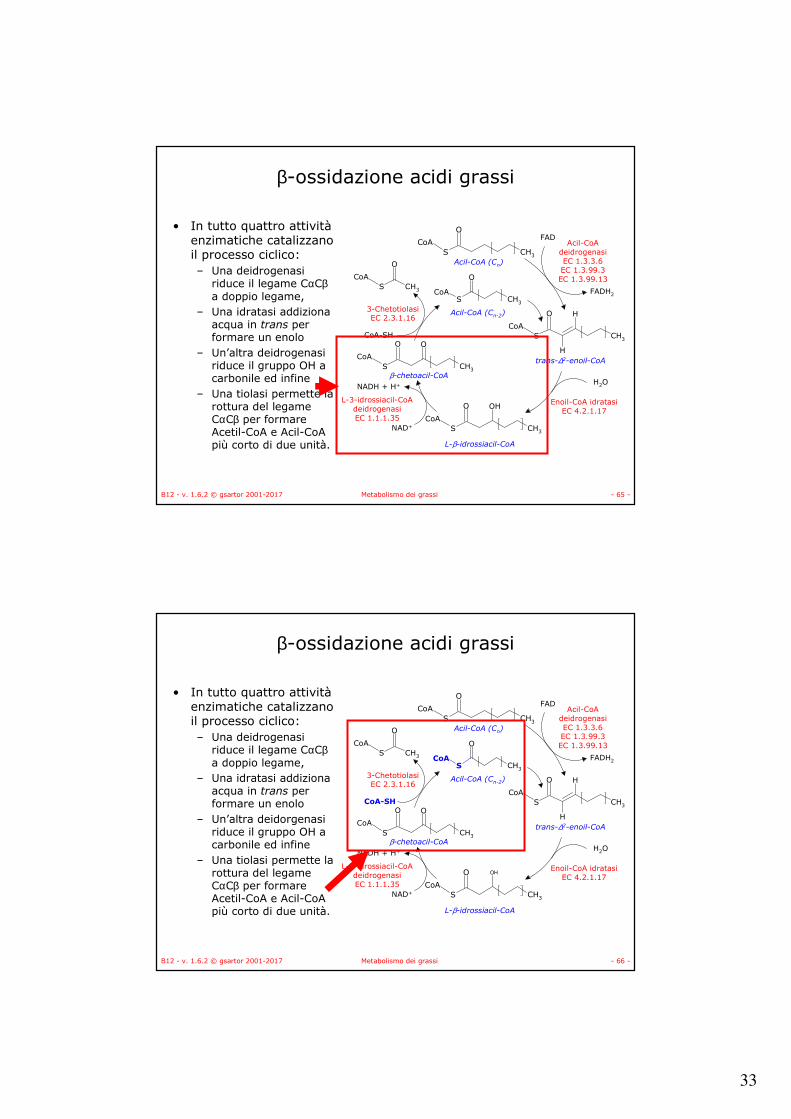
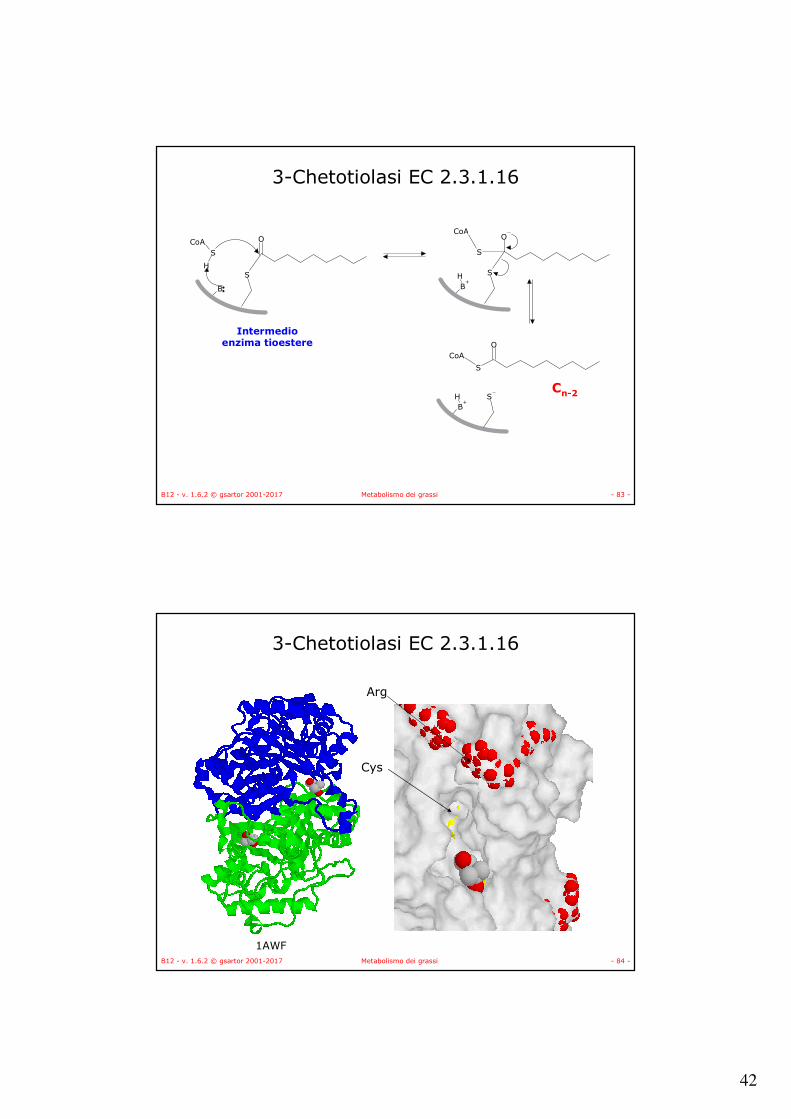
• In tutto quattro attivitàenzimatiche catalizzano il processo ciclico:– Una deidrogenasi
riduce il legame CαCβa doppio legame,
– Una idratasi addiziona acqua in trans per formare un enolo
– Un’altra deidrogenasi riduce il gruppo OH a carbonile ed infine
– Una tiolasi permette la rottura del legame CαCβ per formare Acetil-CoA e Acil-CoApiù corto di due unità.
S
O
CoACH3
S
O
CoACH3
H
H
S
O
CoACH3
OH
S
O
CoACH3
O
S CH3
O
CoAS
O
CoACH3
FAD
FADH2
Acil-CoAdeidrogenasiEC 1.3.3.6EC 1.3.99.3EC 1.3.99.13
H2O
Enoil-CoA idratasiEC 4.2.1.17
NAD+
NADH + H+
L-3-idrossiacil-CoAdeidrogenasiEC 1.1.1.35
CoA-SH
3-ChetotiolasiEC 2.3.1.16
trans-∆2-enoil-CoA
L-β-idrossiacil-CoA
β-chetoacil-CoA
Acil-CoA (Cn)
Acil-CoA (Cn-2)
β-ossidazione acidi grassi
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 64 -
• In tutto quattro attivitàenzimatiche catalizzano il processo ciclico:– Una deidrogenasi
riduce il legame CαCβa doppio legame,
– Una idratasi addiziona acqua in trans per formare un enolo
– Un’altra deidrogenasi riduce il gruppo OH a carbonile ed infine
– Una tiolasi permette la rottura del legame CαCβ per formare Acetil-CoA e Acil-CoApiù corto di due unità.
S
O
CoACH3
S
O
CoACH3
H
H
S
O
CoACH3
OH
S
O
CoACH3
O
S CH3
O
CoAS
O
CoACH3
FAD
FADH2
Acil-CoAdeidrogenasiEC 1.3.3.6EC 1.3.99.3EC 1.3.99.13
H2O
Enoil-CoA idratasiEC 4.2.1.17
NAD+
NADH + H+
L-3-idrossiacil-CoAdeidrogenasiEC 1.1.1.35
CoA-SH
3-ChetotiolasiEC 2.3.1.16
trans-∆2-enoil-CoA
L-β-idrossiacil-CoA
β-chetoacil-CoA
Acil-CoA (Cn)
Acil-CoA (Cn-2)
β-ossidazione acidi grassi
33
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 65 -
• In tutto quattro attivitàenzimatiche catalizzano il processo ciclico:– Una deidrogenasi
riduce il legame CαCβa doppio legame,
– Una idratasi addiziona acqua in trans per formare un enolo
– Un’altra deidrogenasi riduce il gruppo OH a carbonile ed infine
– Una tiolasi permette la rottura del legame CαCβ per formare Acetil-CoA e Acil-CoApiù corto di due unità.
S
O
CoACH3
S
O
CoACH3
H
H
S
O
CoACH3
OH
S
O
CoACH3
O
S CH3
O
CoAS
O
CoACH3
FAD
FADH2
Acil-CoAdeidrogenasiEC 1.3.3.6EC 1.3.99.3EC 1.3.99.13
H2O
Enoil-CoA idratasiEC 4.2.1.17
NAD+
NADH + H+
L-3-idrossiacil-CoAdeidrogenasiEC 1.1.1.35
CoA-SH
3-ChetotiolasiEC 2.3.1.16
trans-∆2-enoil-CoA
L-β-idrossiacil-CoA
β-chetoacil-CoA
Acil-CoA (Cn)
Acil-CoA (Cn-2)
β-ossidazione acidi grassi
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 66 -
• In tutto quattro attivitàenzimatiche catalizzano il processo ciclico:– Una deidrogenasi
riduce il legame CαCβa doppio legame,
– Una idratasi addiziona acqua in trans per formare un enolo
– Un’altra deidorgenasiriduce il gruppo OH a carbonile ed infine
– Una tiolasi permette la rottura del legame CαCβ per formare Acetil-CoA e Acil-CoApiù corto di due unità.
S
O
CoACH3
S
O
CoACH3
H
H
S
O
CoACH3
OH
S
O
CoACH3
O
S CH3
O
CoAS
O
CoACH3
FAD
FADH2
Acil-CoAdeidrogenasiEC 1.3.3.6EC 1.3.99.3EC 1.3.99.13
H2O
Enoil-CoA idratasiEC 4.2.1.17
NAD+
NADH + H+
L-3-idrossiacil-CoAdeidrogenasiEC 1.1.1.35
CoA-SH
3-ChetotiolasiEC 2.3.1.16
trans-∆2-enoil-CoA
L-β-idrossiacil-CoA
β-chetoacil-CoA
Acil-CoA (Cn)
Acil-CoA (Cn-2)
β-ossidazione acidi grassi

34
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 67 -
• In tutto quattro attivitàenzimatiche catalizzano il processo ciclico:– Una deidrogenasi
riduce il legame CαCβa doppio legame,
– Una idratasi addiziona acqua in trans per formare un enolo
– Un’altra deidorgenasiriduce il gruppo OH a carbonile ed infine
– Una tiolasi permette la rottura del legame CαCβ per formare Acetil-CoA e Acil-CoApiù corto di due unità.
S
O
CoACH3
S
O
CoACH3
H
H
S
O
CoACH3
OH
S
O
CoACH3
O
S CH3
O
CoAS
O
CoACH3
FAD
FADH2
Acil-CoAdeidrogenasiEC 1.3.3.6EC 1.3.99.3EC 1.3.99.13
H2O
Enoil-CoA idratasiEC 4.2.1.17
NAD+
NADH + H+
L-3-idrossiacil-CoAdeidrogenasiEC 1.1.1.35
CoA-SH
3-ChetotiolasiEC 2.3.1.16
trans-∆2-enoil-CoA
L-β-idrossiacil-CoA
β-chetoacil-CoA
Acil-CoA (Cn)
Acil-CoA (Cn-2)
β-ossidazione acidi grassi
E il ciclo si ripete
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 68 -
• In tutto quattro attivitàenzimatiche catalizzano il processo ciclico:– Una deidrogenasi
riduce il legame CαCβa doppio legame,
– Una idratasi addiziona acqua in trans per formare un enolo
– Un’altra deidrogenasi riduce il gruppo OH a carbonile ed infine
– Una tiolasi permette la rottura del legame CαCβ per formare Acetil-CoA e Acil-CoApiù corto di due unità.
S
O
CoACH3
S
O
CoACH3
H
H
S
O
CoACH3
OH
S
O
CoACH3
O
S CH3
O
CoAS
O
CoACH3
FAD
FADH2
Acil-CoAdeidrogenasiEC 1.3.3.6EC 1.3.99.3EC 1.3.99.13
H2O
Enoil-CoA idratasiEC 4.2.1.17
NAD+
NADH + H+
L-3-idrossiacil-CoAdeidrogenasiEC 1.1.1.35
CoA-SH
3-ChetotiolasiEC 2.3.1.16
trans-∆2-enoil-CoA
L-β-idrossiacil-CoA
β-chetoacil-CoA
Acil-CoA (Cn)
Acil-CoA (Cn-2)
β-ossidazione acidi grassi
35
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 69 -
S
O
CoACH3
S
O
CoA
CH3
H
H
S
O
CoACH3
OH
S
O
CoACH3
O
S CH3
O
CoAS
O
CoACH3
FAD
FADH2
H2O
Enoil-CoA idratasiEC 4.2.1.17
NAD+
NADH + H+
L-3-idrossiacil-CoAdeidrogenasiEC 1.1.1.35
CoA-SH
3-ChetotiolasiEC 2.3.1.16
trans-∆2-enoil-CoA
L-β-idrossiacil-CoA
β-chetoacil-CoA
Acil-CoA (Cn)
Acil-CoA (Cn-2)
Acil-CoAdeidrogenasi
EC 1.3.3.6EC 1.3.99.3
EC 1.3.99.13
Acil-CoA deidrogenasiEC 1.3.3.6 - EC 1.3.99.3 - EC 1.3.99.13
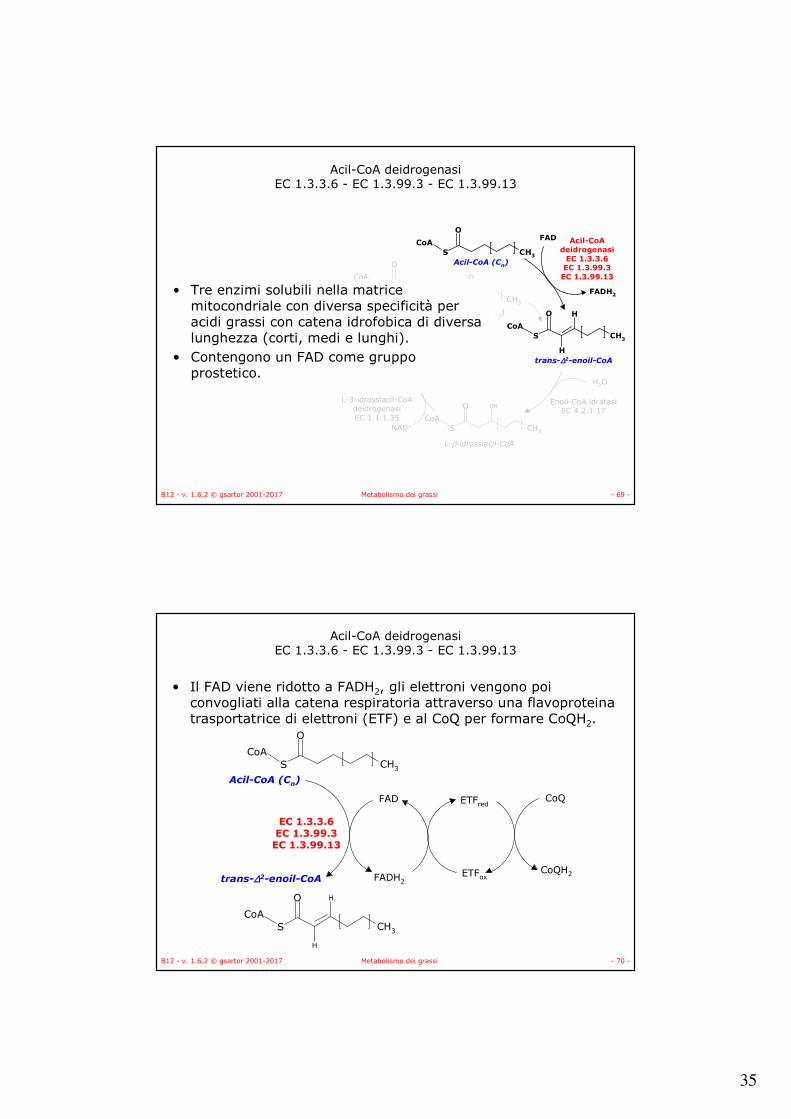
• Tre enzimi solubili nella matrice mitocondriale con diversa specificità per acidi grassi con catena idrofobica di diversa lunghezza (corti, medi e lunghi).
• Contengono un FAD come gruppo prostetico.
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 70 -
S
O
CoACH3
S
O
CoACH3
H
H
FAD
FADH2trans-∆2-enoil-CoA
Acil-CoA (Cn)
ETFox
ETFredCoQ
CoQH2
EC 1.3.3.6EC 1.3.99.3
EC 1.3.99.13
Acil-CoA deidrogenasiEC 1.3.3.6 - EC 1.3.99.3 - EC 1.3.99.13
• Il FAD viene ridotto a FADH2, gli elettroni vengono poi convogliati alla catena respiratoria attraverso una flavoproteina trasportatrice di elettroni (ETF) e al CoQ per formare CoQH2.
36
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 71 -
Acil-CoA deidrogenasiEC 1.3.3.6 - EC 1.3.99.3 - EC 1.3.99.13
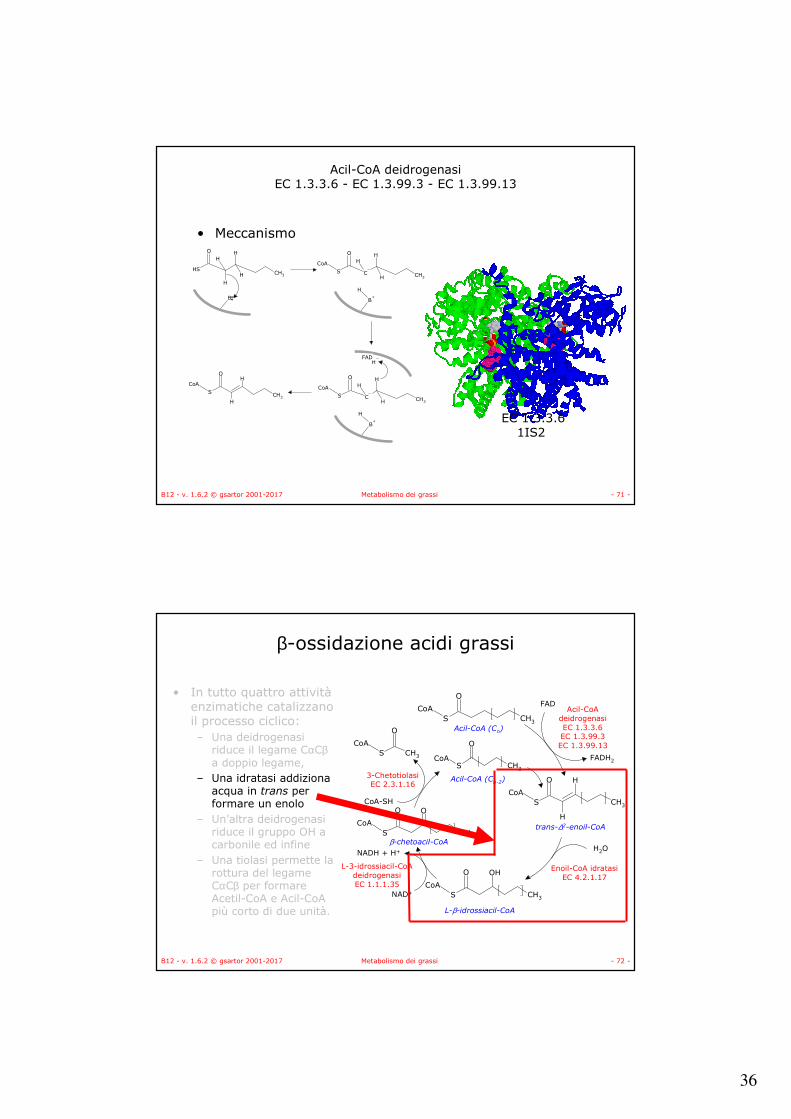
• Meccanismo
SH
O
CH3
H
H
H
H
B
S C
O
CoA
CH3
H
H
H
B+
H
S C
O
CoA
CH3
H
H
H
B+
H
H
S
O
CoA
CH3H
H
FAD
EC 1.3.3.61IS2
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 72 -
• In tutto quattro attivitàenzimatiche catalizzano il processo ciclico:– Una deidrogenasi
riduce il legame CαCβa doppio legame,
– Una idratasi addiziona acqua in trans per formare un enolo
– Un’altra deidrogenasi riduce il gruppo OH a carbonile ed infine
– Una tiolasi permette la rottura del legame CαCβ per formare Acetil-CoA e Acil-CoApiù corto di due unità.
S
O
CoACH3
S
O
CoACH3
H
H
S
O
CoACH3
OH
S
O
CoACH3
O
S CH3
O
CoAS
O
CoACH3
FAD
FADH2
Acil-CoAdeidrogenasiEC 1.3.3.6EC 1.3.99.3EC 1.3.99.13
H2O
Enoil-CoA idratasiEC 4.2.1.17
NAD+
NADH + H+
L-3-idrossiacil-CoAdeidrogenasiEC 1.1.1.35
CoA-SH
3-ChetotiolasiEC 2.3.1.16
trans-∆2-enoil-CoA
L-β-idrossiacil-CoA
β-chetoacil-CoA
Acil-CoA (Cn)
Acil-CoA (Cn-2)
β-ossidazione acidi grassi

37
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 73 -
S
O
CoACH3
S
O
CoA
CH3
H
H
S
O
CoACH
3
OH
S
O
CoACH3
O
S CH3
O
CoAS
O
CoACH3
FAD
FADH2
Acil-CoAdeidrogenasiEC 1.3.3.6EC 1.3.99.3EC 1.3.99.13
H2O
Enoil-CoA idratasiEC 4.2.1.17EC 4.2.1.74
NAD+
NADH + H+
L-3-idrossiacil-CoAdeidrogenasiEC 1.1.1.35
CoA-SH
3-ChetotiolasiEC 2.3.1.16
trans-∆2-enoil-CoA
L-β-idrossiacil-CoA
β-chetoacil-CoA
Acil-CoA (Cn)
Acil-CoA (Cn-2)
β-ossidazione acidi grassi
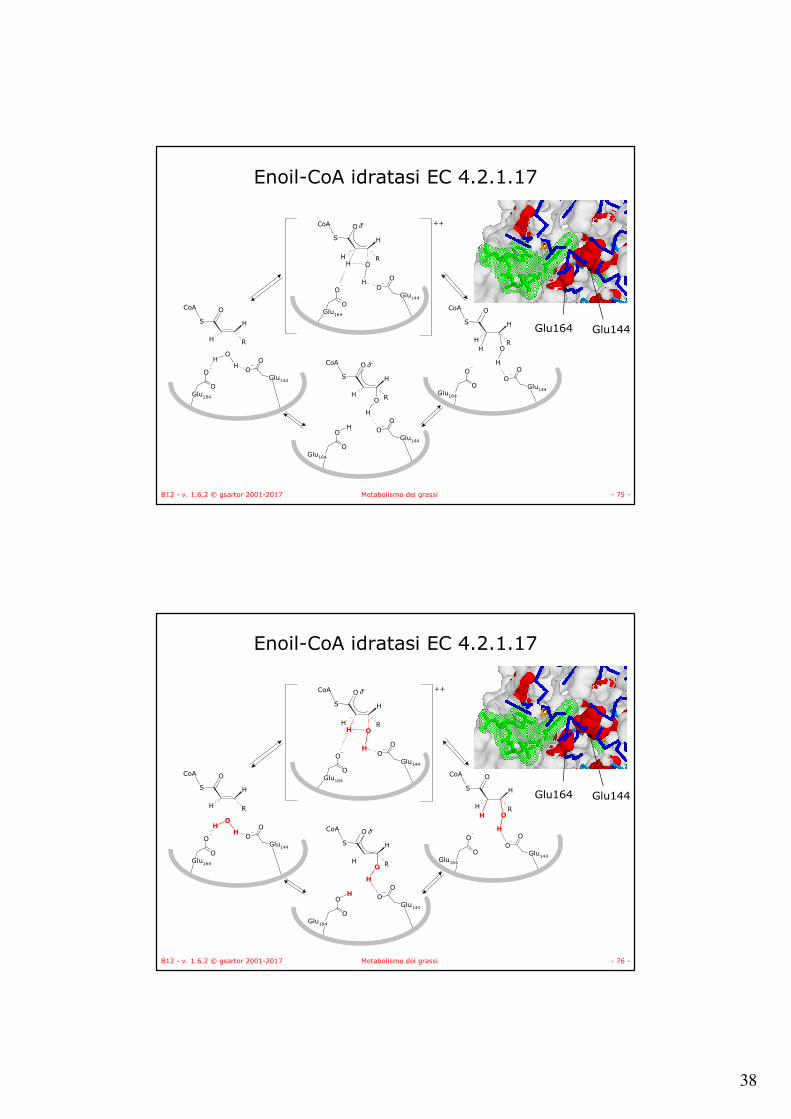
• Idratazione stereo specifica.• L’enoil-CoA idratasi catalizza l’idratazione
stereospecifica al doppio legame transproducendo L-idrossiacil-CoA.
• Esamero. • Due enzimi:
• EC 4.2.1.17: maggiore affinità per enoil-CoA a corta catena.
• EC 4.2.1.74: maggiore affinità per enoil-CoA a lunga catena.
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 74 -
Enoil-CoA idratasi EC 4.2.1.17
Esanoil-CoA
1MJ3
38
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 75 -
Enoil-CoA idratasi EC 4.2.1.17
S
OCoA
RH
H
Glu164
O
O
HO
H
Glu144
OO
Glu164
O
O
H O
H
Glu144
OO
S
OCoA
RH
H
Glu164
O
OS
OCoA
RH
H
O
H
Glu144
OO
Glu164
O
OH
O
H
Glu144
OO
H
S
OCoA
RH
H
δ− ++
δ−
Glu164 Glu144
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 76 -
Enoil-CoA idratasi EC 4.2.1.17
S
OCoA
RH
H
Glu164
O
O
HO
H
Glu144
OO
Glu164
O
O
H O
H
Glu144
OO
S
OCoA
RH
H
Glu164
O
OS
OCoA
RH
H
O
H
Glu144
OO
Glu164
O
OH
O
H
Glu144
OO
H
S
OCoA
RH
H
δ− ++
δ−
Glu164 Glu144

39
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 77 -
• In tutto quattro attivitàenzimatiche catalizzano il processo ciclico:– Una deidrogenasi
riduce il legame CαCβa doppio legame,
– Una idratasi addiziona acqua in trans per formare un enolo
– Un’altra deidrogenasi riduce il gruppo OH a carbonile ed infine
– Una tiolasi permette la rottura del legame CαCβ per formare Acetil-CoA e Acil-CoApiù corto di due unità.
S
O
CoACH3
S
O
CoACH3
H
H
S
O
CoACH3
OH
S
O
CoACH3
O
S CH3
O
CoAS
O
CoACH3
FAD
FADH2
Acil-CoAdeidrogenasiEC 1.3.3.6EC 1.3.99.3EC 1.3.99.13
H2O
Enoil-CoA idratasiEC 4.2.1.17
NAD+
NADH + H+
L-3-idrossiacil-CoAdeidrogenasiEC 1.1.1.35
CoA-SH
3-ChetotiolasiEC 2.3.1.16
trans-∆2-enoil-CoA
L-β-idrossiacil-CoA
β-chetoacil-CoA
Acil-CoA (Cn)
Acil-CoA (Cn-2)
β-ossidazione acidi grassi
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 78 -
S
O
CoACH3
S
O
CoACH3
H
H
S
O
CoA
CH3
OH
S
O
CoA
CH3
O
S CH3
O
CoAS
O
CoACH3
FAD
FADH2
Acil-CoAdeidrogenasiEC 1.3.3.6EC 1.3.99.3EC 1.3.99.13
H2O
Enoil-CoA idratasiEC 4.2.1.17
NAD+
NADH + H+
L-3-idrossiacil-CoAdeidrogenasiEC 1.1.1.35
CoA-SH
3-ChetotiolasiEC 2.3.1.16
trans-∆2-enoil-CoA
L-β-idrossiacil-CoA
β-chetoacil-CoA
Acil-CoA (Cn)
Acil-CoA (Cn-2)
β-ossidazione acidi grassi
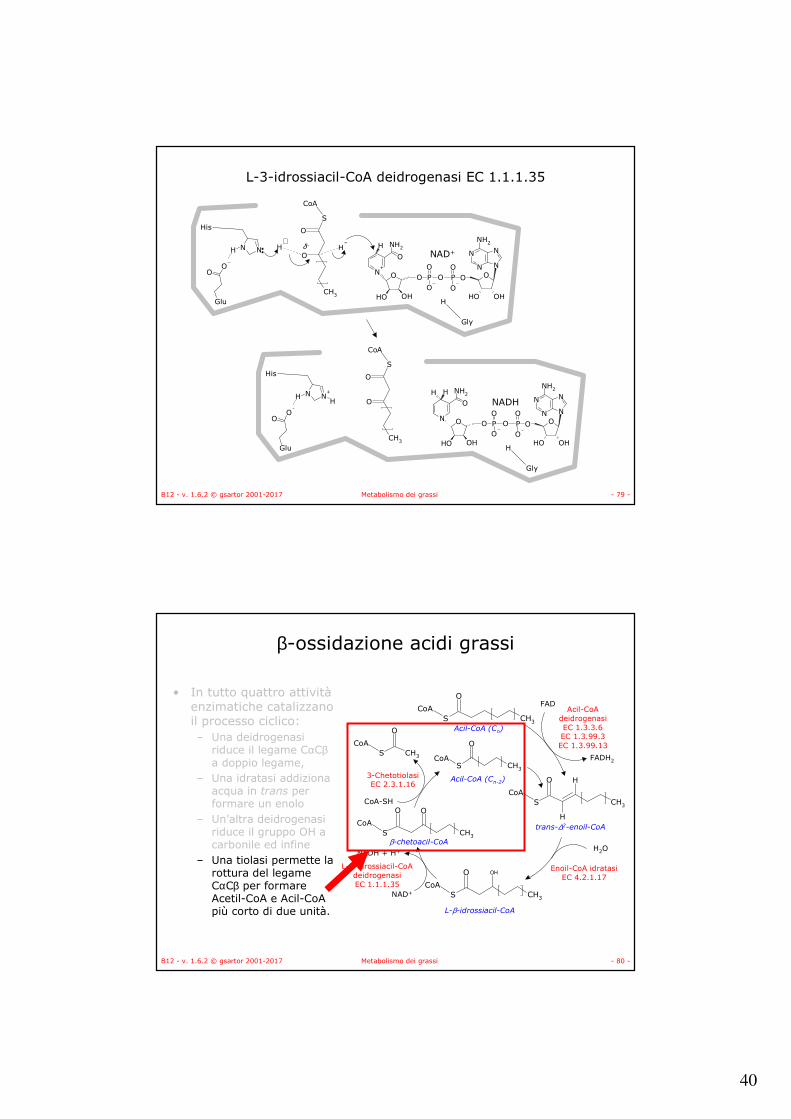
• L’enzima L-3-idrossiacil-CoA deidrogenasi catalizza l’ossidazione di L-β-idrossiacil-CoA.
• È un enzima specifico per l’isomero L.• Il NADH prodotto entra nella catena respiratoria a livello del
complesso I.
40
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 79 -
L-3-idrossiacil-CoA deidrogenasi EC 1.1.1.35
S
O
CoA
CH3
O
H H
Glu
OO
N N H
His
NNO
N
OH
N
NH2
OH
OPOO
OPOO
OO
OH
N+
OH
NH2
O
H
Gly
H
S
O
CoA
CH3
O
Glu
OO
N N+
H
His
HNN
O
N
OH
N
NH2
OH
OPOO
OPOO
OO
OH
N
OH
NH2
O
HH
Gly
H
+δ− -
NAD+
NADH
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 80 -
• In tutto quattro attivitàenzimatiche catalizzano il processo ciclico:– Una deidrogenasi
riduce il legame CαCβa doppio legame,
– Una idratasi addiziona acqua in trans per formare un enolo
– Un’altra deidrogenasi riduce il gruppo OH a carbonile ed infine
– Una tiolasi permette la rottura del legame CαCβ per formare Acetil-CoA e Acil-CoApiù corto di due unità.
S
O
CoACH3
S
O
CoACH3
H
H
S
O
CoACH3
OH
S
O
CoACH3
O
S CH3
O
CoAS
O
CoACH3
FAD
FADH2
Acil-CoAdeidrogenasiEC 1.3.3.6EC 1.3.99.3EC 1.3.99.13
H2O
Enoil-CoA idratasiEC 4.2.1.17
NAD+
NADH + H+
L-3-idrossiacil-CoAdeidrogenasiEC 1.1.1.35
CoA-SH
3-ChetotiolasiEC 2.3.1.16
trans-∆2-enoil-CoA
L-β-idrossiacil-CoA
β-chetoacil-CoA
Acil-CoA (Cn)
Acil-CoA (Cn-2)
β-ossidazione acidi grassi

41
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 81 -
S
O
CoACH3
S
O
CoACH3
H
H
S
O
CoACH3
OH
S
O
CoA
CH3
O
S CH3
O
CoAS
O
CoA
CH3
FAD
FADH2
Acil-CoAdeidrogenasiEC 1.3.3.6EC 1.3.99.3EC 1.3.99.13
H2O
Enoil-CoA idratasiEC 4.2.1.17
NAD+
NADH + H+
L-3-idrossiacil-CoAdeidrogenasiEC 1.1.1.35
CoA-SH
3-ChetotiolasiEC 2.3.1.16
trans-∆2-enoil-CoA
L-β-idrossiacil-CoA
β-chetoacil-CoA
Acil-CoA (Cn)
Acil-CoA (Cn-2)
β-ossidazione acidi grassi
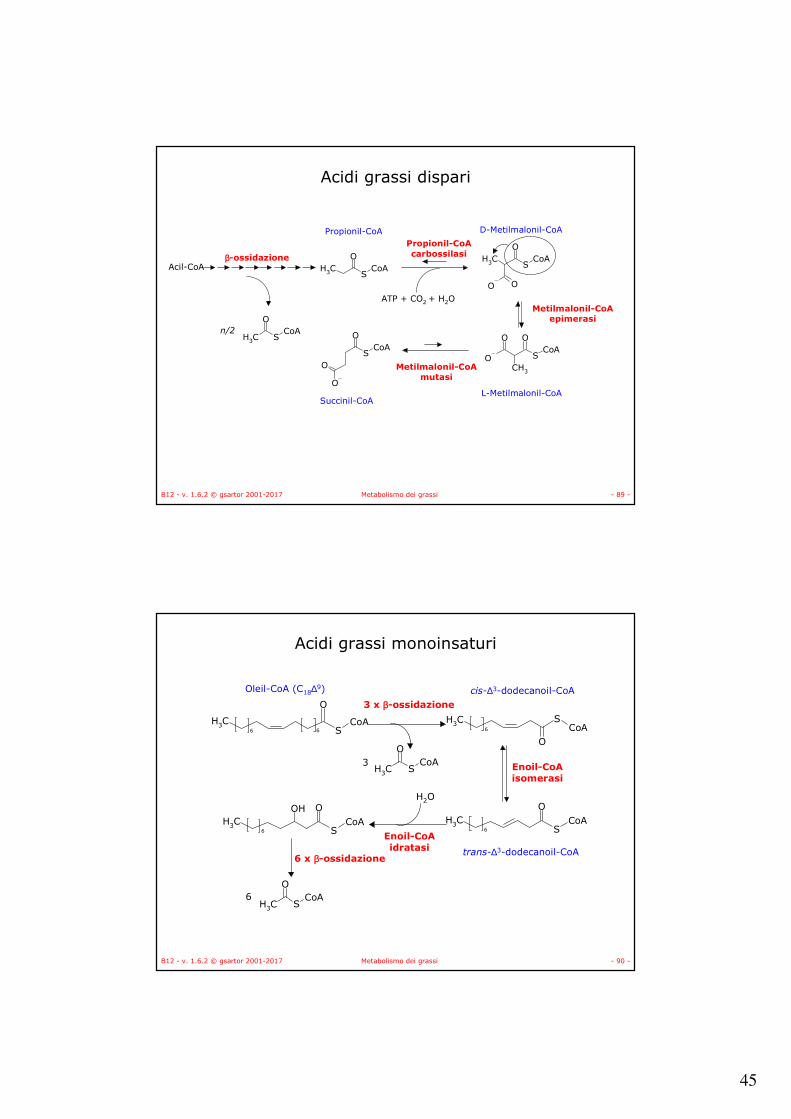
• La chetotiolasi catalizza la scissione del β-chetoacil-CoA.• Si produce un Acil-CoA di due atomi di carbonio più corto.• Il ciclo ricomincia.
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 82 -
3-Chetotiolasi EC 2.3.1.16
S
O
CoA
O
S
S
O
CoA
O
B+
H
S
B+
H
S
O
B+
H
S
O
CoACH2-
S
O
CoACH2
S
O
CoACH3S
O
B
Cn
Intermedioenzima tioestere
C2
42
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 83 -
3-Chetotiolasi EC 2.3.1.16
S
O
B
CoAS
H
B+
H
CoA
S
S
O
B+
H
CoA
S
O
S
Intermedioenzima tioestere
Cn-2
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 84 -
3-Chetotiolasi EC 2.3.1.16
1AWF
Cys
Arg

43
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 85 -
Stechiometria• Per ogni ciclo si producono:
– NADH che equivale a 2.5 moli di ATP prodotte nella catena respiratoria.
– FADH2 che equivale a 1.5 moli di ATP prodotte nella catena respiratoria.
• Per la degradazione dell’acido palmitico (C16):CH3-(CH2)14-CO-S-CoA + 7FAD + 7NAD+ + 7H2O + 7CoASH →
→ 8CH3-CO-S-CoA + 7FADH2 + 7NADH + 7H+
7FADH2 + 10.5Pi + 10.5ADP + 3.5O2 →7FAD + 17.5H2O + 10.5ATP 320 kJ·mol-1
7NADH + 7H+ + 17.5Pi + 17.5ADP + 3.5O2 →7NAD+ + 24.5H2O + 17.5ATP 534 kJ·mol-1
8CH3-CO-S-CoA + 16O2 + 80Pi + 80ADP →→ 8 CoA + 88H2O + 16CO2 + 80ATP 2440 kJ·mol-1
CH3-(CH2)14-CO-S-CoA + 108Pi +108ADP +23O2 →→ 108ATP + 16 CO2 + 130H2O + CoASH 3294 kJ·mol-1
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 86 -
Stechiometria• Per ogni ciclo si producono:
– NADH che equivale a 2.5 moli di ATP prodotte nella catena respiratoria.
– FADH2 che equivale a 1.5 moli di ATP prodotte nella catena respiratoria.
• Per la degradazione dell’acido palmitico (C16):CH3-(CH2)14-CO-S-CoA + 7FAD + 7NAD+ + 7H2O + 7CoASH →
→ 8CH3-CO-S-CoA + 7FADH2 + 7NADH + 7H+
7FADH2 + 10.5Pi + 10.5ADP + 3.5O2 →7FAD + 17.5H2O + 10.5ATP 320 kJ·mol-1
7NADH + 7H+ + 17.5Pi + 17.5ADP + 3.5O2 →7NAD+ + 24.5H2O + 17.5ATP 534 kJ·mol-1
8CH3-CO-S-CoA + 16O2 + 80Pi + 80ADP →→ 8 CoA + 88H2O + 16CO2 + 80ATP 2440 kJ·mol-1
CH3-(CH2)14-CO-S-CoA + 108Pi +108ADP +23O2 →→ 108ATP + 16 CO2 + 130H2O + CoASH 3294 kJ·mol-1

44
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 87 -
Rendimento
• Combustione dell’acido palmitico:CH3-(CH2)14-COOH + O2 → CO2 + H2O
∆G°’ = -9790 kJ·mol-1
• Ossidazione dell’acido palmitico e respirazione cellulare: CH3-(CH2)14-COOH + O2 → CO2 + H2O
(108 – 2)·ATP = ∆G°’ = -3233 kJ·mol-1
Efficienza = 3233/9790 = 33%
• Produzione di 130 H2O metabolica per mole di acido palmitico che viene β-ossidato.
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 88 -
Pari e dispari
• Dalla β-ossidazione di acidi grassi con atomi si carbonio pari (n) si formano n/2 acetil-CoA attraverso (n/2 – 1) cicli di reazioni.
• Dalla β-ossidazione di acidi grassi con atomi si carbonio dispari (n) si formano n/2-1 acetil-CoA + 1 propionil-CoA da (n/2-1) cicli di reazioni.
45
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 89 -
Acidi grassi dispari
CH3
O
SCoA
CH3
O
SCoA
O O
O
SCoA
CH3
O
O
O
SCoA
O
O
CH3
O
SCoA
Propionil-CoAPropionil-CoAcarbossilasi
ATP + CO2 + H2O
D-Metilmalonil-CoA
Metilmalonil-CoAepimerasi
L-Metilmalonil-CoA
Metilmalonil-CoAmutasi
Succinil-CoA
Acil-CoAβ-ossidazione
n/2
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 90 -
Acidi grassi monoinsaturi
S
O
CoACH36 6
SCoA
CH3
O6
SCoA
OCH3
6SCoA
OCH3
OH
6
CH3
O
SCoA
CH3
O
SCoA
Oleil-CoA (C18∆9)
3 x β-ossidazione
Enoil-CoAisomerasi
cis-∆3-dodecanoil-CoA
trans-∆3-dodecanoil-CoA
H2O
3
6
6 x β-ossidazione
Enoil-CoAidratasi

46
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 91 -
Acidi grassi polinsaturi
S
O
CoACH34 6
CH3
O
SCoA
CH3
O
SCoA
S
O
CoACH3
4
S
O
CoACH34S
CoAO
CH34
SCoA
OCH3
4S
CoA
O
CH34
Linoeil-CoA (C18∆9∆12)
3 x β-ossidazione
Enoil-CoAisomerasi
cis-∆3-cis-∆3-dodecanoil-CoA
H2O
3
Enoil-CoAidratasi trans-∆3-cis-∆6-dodecanoil-CoA
β-ossidazione
cis-∆4-decanoil-CoA
NADPH + H+ NADP+
Dienoil-CoAreduttasitrans-∆2-cis-∆4-decanoil-CoA
trans-∆3-decanoil-CoA
H2O
Enoil-CoAidratasi
4 x β-ossidazione
5
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 92 -
Perossisomi
• La β-ossidazione avviene in molte strutture subcellulari, in particolare nei perossisomi.
• Sono deputati alla formazione di H2O2 che viene utilizzata come sistema di difesa da batteri, virus, ecc.
• L’accettore di elettroni della acil-CoA ossidasi è il FAD:
FAD → FADH2
FADH2 + O2 → FAD + H2O2
2 H2O2 → H2O + O2

47
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 93 -
ω-ossidazione
• Nel reticolo endoplasmatico può avvenire la ω-ossidazione.
• Catalizzata da enzimi che appartengono alla classe delle ossidasi miste (CytP450, CYP)
• Provocano una idrossilazione e quindi una ossidazione.• È un processo aspecifico che converte molecole lipofile in
prodotti più idrosolubili più facili da eliminare.• È un processo che detossifica le cellule da molecole lipofile.
O
O
CH 3n
O
O
n
OHO
O
n
O
O
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 94 -
Struttura del CYP (1PO5)
AA basici
48
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 95 -
Struttura del CYP (1PO5)
AA basiciCys436
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 96 -
Struttura del CYP (1PO5)
AA basiciCys436
Cavità

49
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 97 -
Ciclo del CYP
Fe3+H2O
Fe3+RH
Fe2+RH
[Fe2+O2RH]-1
FeO3+RH
Fe3+ROH
[Fe2+O2RH]-2[Fe2+OOHRH]-1
ROH RH
H+
e-
e-
H+
H2O
H2O
H2O
(substrato)(prodotto)
O2
NAD(P)H-citocromoP450 reduttasi
citocromo b5
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 98 -
Meccanismo ciclico del CYP
O
OH
O
H
N
NN
N
Fe4+
O2-
S
N
NN
NFe
3+
S
N
NN
N
HOO
Fe3+
S
N
NN
NFe
2+
S
OHR
N
NN
NFe
2+
O
S
N
NN
NFe
3+
OHH
S
N
NN
NFe
3+
S
N
NN
N
O O
Fe3+
S
N
NN
N
O O
Fe3+
S
-
N
NN
NFe
2+
OO
S
H
ROH RH
RH
H2O2
H+
e-
H+
O2
H2O
I
II
III
e-
IVV
RH
-
RH
+.
H+
H2O
H2O
VI
VII
RH
RH
2-RH
RH
A

50
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 99 -
Meccanismo ciclico del CYP
O
OH
O
H
N
NN
N
Fe4+
O2-
S
N
NN
NFe
3+
S
N
NN
N
HOO
Fe3+
S
N
NN
NFe
2+
S
OHR
N
NN
NFe
2+
O
S
N
NN
NFe
3+
OHH
S
N
NN
NFe
3+
S
N
NN
N
O O
Fe3+
S
N
NN
N
O O
Fe3+
S
-
N
NN
NFe
2+
OO
S
H
ROH RH
RH
H2O2
H+
e-
H+
O2
H2O
I
II
III
e-
IVV
RH
-
RH
+.
H+
H2O
H2O
VI
VII
RH
RH
2-RH
RH
A
I. P450 acquo Fe3+
(basso spin)Lega RH
II. P450 canfora Fe3+
(alto spin)entra 1e-, riduzione a Fe2+
III. P450 canfora Fe2+
Lega O2
IV. P450 con O2legato, equivalente a Fe3+-O2
-
entra 1e-, riduzione a O2
2-
V. P450 perossido
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 100 -
Meccanismo ciclico del CYP
O
OH H
N
NN
N
Fe4+
O2-
S
N
NN
NFe
3+
S
N
NN
N
HOO
Fe3+
S
N
NN
NFe
2+
S
OHR
N
NN
NFe
2+
O
S
N
NN
NFe
3+
OHH
S
N
NN
NFe
3+
S
N
NN
N
O O
Fe3+
S
N
NN
N
O O
Fe3+
S
-
N
NN
NFe
2+
OO
S
H
ROH RH
RH
H2O2
H+
e-
H+
O2
H2O
I
II
III
e-
IVV
RH
-
RH
+.
H+
H2O
H2O
VI
VII
RH
RH
2-RH
RH
A
Entra 1H+
VI. P450 idroperossidoEntra 1H+ esce H2O
VII. P450 Fe4+ O2-
catione radicale sulla proteinasi forma ROH
A. L’idroperossido VI si può formare per reazione di IIcon H2O2

51
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 101 -
Meccanismo
• L’ossigeno è legato non ad angolo retto.• Il legame dell’ossigeno allontana il ligando
(RH) solo dopo che I due atomi di ossigeno si sono ridotti il ligando si riavvicina. Ciò previene la formazione di ROS.
• Gli elettroni per la riduzione dell’ossigeno so forniti da una proteina Fe-S (P450 batterica o mitcondriale) o da una NADPH-citocromo P450 ossidoreduttasi FAD/FMN dipendente (microsomi).
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 102 -
Meccanismo generale del CYP
NADPH + H+
NADP+CYTP450Reduttasi
FMNH2/FADH2
CYTP450Reduttasi FMN/FAD
CYP Fe2+
CYP Fe3+
RH
ROH
2H+ + O2
H2O

52
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 103 -
Metabolismo dei grassi
1. Demolizione dei trigliceridi2. Catabolismo degli acidi grassi
3. Biosintesi – degli acidi grassi e colesterolo– dei lipidi complessi
4. Trasporto dei lipidi
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 104 -
Biosintesi degli acidi grassi
• La sintesi degli acidi grassi segue un percorso diverso rispetto al catabolismo:– Le catene di acidi grassi sono costruite per addizione
di unità di due atomi di carbonio derivate dal acetil-CoA.
– Le unità di acetato sono attivate dalla formazione di malonil-CoA.
– Gli intermedi della biosintesi sono legati a SH di proteine (proteine trasportatrici di acili, ACP) e non a CoA-SH.
– La sintesi avviene nel citoplasma mentre la degradazione è mitocondriale.

53
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 105 -
Biosintesi degli acidi grassi
• Negli animali la sintesi è catalizzata da una serie di enzimi componenti del complesso acido grasso sintasi (EC 2.3.1.85 EC 2.3.1.86) mentre nelle piante e nei batteri gli enzimi sono separati
• La biosintesi usa come sistema redox il NADP+/NADPH (la degradazione usa il sistema NAD+/NADH).
• L’addizione dell’unità C2 è alimentata dal ∆G negativo della decarbossilazione del malonil-CoA.
• L’allungamento è ripetuto fino alla formazione di C16(Acido palmitico).
• Altri enzimi catalizzano la ramificazione e l’insaturazione.
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 106 -
Strategia
CH3
O
SCoA
O
SCoA
O
O
CH3
O
S ACP
SO ACP
CH3 S
O
ACP
O
CH3
O
SCoA
O
O
O
O (CH2)14
CH3
Acetil-CoA Malonil-CoA
Acetil-ACP
H2O
NADPH
NADP+
NADP+
NADPH
CO2
CO2
CO2
INVERSO DELLA β-OSSIDAZIONE
Dopo sette cicli
6
EC 2.3.1.85EC 2.3.1.86

54
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 107 -
Energia
• La spinta è il potere riducente del NADPH
Acetil-CoA + 7 Malonil-CoA + 14 NADPH + 14H+ →→ Palmitoil-CoA + 7 HCO3
- + 7 CoA-SH + 14 NADP+
Formazione di esteri
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 108 -
Quindi:• Per produrre acidi grassi, nel citoplasma servono:
– Acetil-CoA– Malonil-CoA
• (Acetil-CoA + CO2 → Malonil-CoA)
– Equivalenti riducenti come NADPH
55
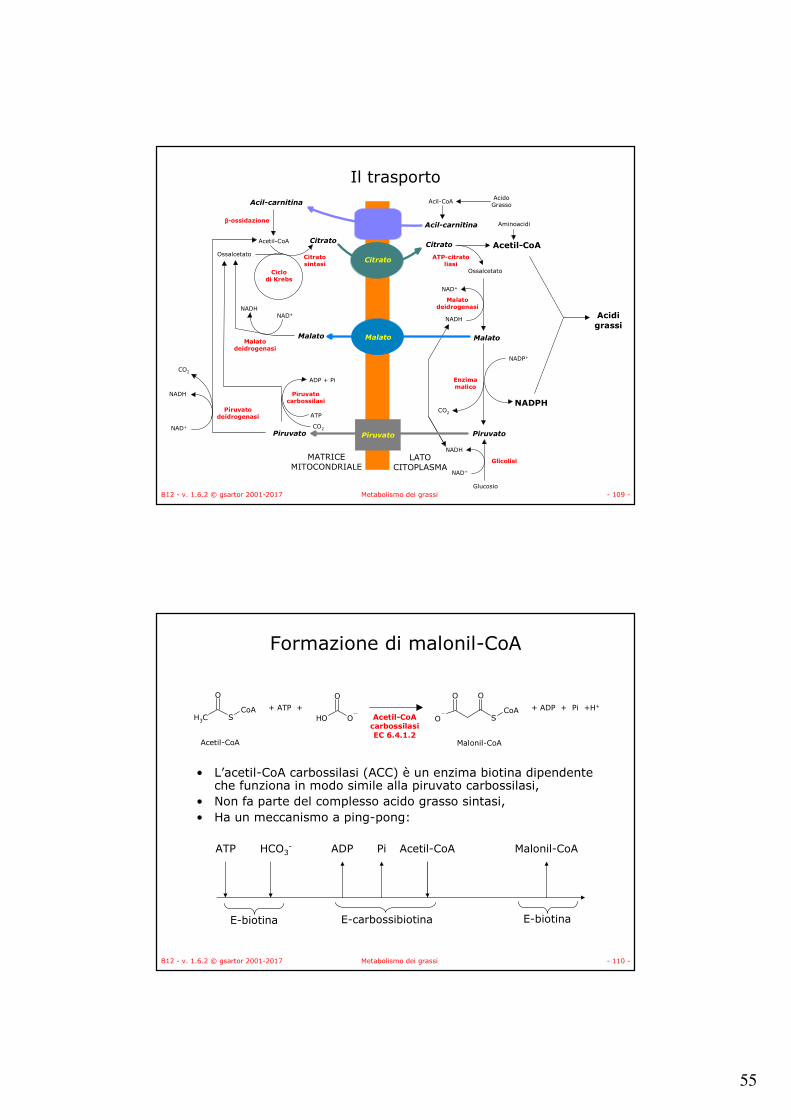
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 109 -
Il trasporto
Piruvato
Glucosio
Glicolisi
Enzimamalico
Malato
Malatodeidrogenasi
Ossalcetato
Citrato
Acil-carnitina
AcidoGrassoAcil-CoA
Aminoacidi
Acetil-CoA
ATP-citratoliasi
Acidigrassi
Ciclodi Krebs
Citrato
Ossalcetato
Malatodeidrogenasi
Citratosintasi
Acetil-CoA
Acil-carnitina
β-ossidazione
Piruvato
Malato
Piruvatocarbossilasi
Piruvatodeidrogenasi
NAD+NADH
NADP+
NADPH
ATP
ADP + Pi
CO2NAD+
NADH
CO2
NAD+
NADH
NAD+
NADH
CO2
MATRICEMITOCONDRIALE
LATOCITOPLASMA
Citrato
Malato
Piruvato
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 110 -
Formazione di malonil-CoA
CH3
O
SCoA
O
OOH
O
SCoA
O
O+ ATP +
Acetil-CoAcarbossilasiEC 6.4.1.2
Acetil-CoA Malonil-CoA
+ ADP + Pi +H+
• L’acetil-CoA carbossilasi (ACC) è un enzima biotina dipendente che funziona in modo simile alla piruvato carbossilasi,
• Non fa parte del complesso acido grasso sintasi,• Ha un meccanismo a ping-pong:
E-biotina
ATP HCO3-
E-carbossibiotina
ADP Pi Acetil-CoA
E-biotina
Malonil-CoA

56
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 111 -
Acetil-CoA carbossilasi (EC 6.4.1.2)
• Ha un peso molecolare da 4000 a 8000 kD è formata da monomeri di 230 kD
• Ogni subunità contiene le attività per:– Il trasporto della carbossibiotina, – La biotina carbossilasi– La transcarbossilasi– I siti di regolazione allosterica
• La forma polimerica è attiva, i singoli monomeri non sono attivi.
• L’equilibrio:Monomeri inattivi ↔ polimero attivo
• È regolato dal prodotto (palmitoil-CoA) che lo sposta verso i monomeri, mentre il citrato lo sposta verso il polimero.
• Gli effetti allosterici sono regolati dalla fosforilazione.
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 112 -
Acetil-CoA carbossilasi (EC 6.4.1.2)
• Possiede numerosi siti di fosforilazione,• È fosforilata da una chinasi e defosforilata da una fosfatasi.• La ACC defosforilata è attivata da bassa concentrazione di
citrato ed inibita da alta concentrazione di (grasso) acil-CoA, al contrario la ACC fosforilata attivata da alta concentrazionedi citrato e inibita da bassa concentrazione di (grasso) acil-CoA.
1 1200 2345
P
P P P P P P
23 25 29 76 77 95
Protein chinasi calmodulinadipendente
Caseinachinasi II?
Protein chinasi cAMP dipendenteProtein chinasi C
Protein chinasi AMP dipendente Protein chinasi C
Protein chinasi cAMP dipendente
AMP-protein chinasi

57
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 113 -
Acetil-CoA carbossilasi (EC 6.4.1.2)
• È un enzima a biotina
• La biotina si lega ad una lisina nel sito attivo dell’enzima formando un lungo braccio flessibile ad una estremità del quale vi è il sito di carbossilazione.
NH NH
S
CH3CH3
O
OH
O
NH NH
S
CH3CH3
O
O
N
O
NHH
Sito di carbossilazione
Lisina
Legameε-peptidico
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 114 -
Acetil-CoA carbossilasi (EC 6.4.1.2)
• Il lungo braccio flessibile permette il movimento della biotina tra il sito di carbossilazione e il sito di decarbossilazione e formazione del malonil-CoA.
• La carbossilazione avviene ad opera di carbossifosfato che si forma nel sito di carbossilazione per reazione di ATP e bicarbonato.
NHNH
SCH3
CH3
O
O
N O
NH
H
OO
OP
O
OO
NNH
SCH3
CH3
O
O
NO
NH
H
O
O
NNH
S
CH3
CH3
O
O
N
O
NH
H
O
O
OP
O
OO
Carbossifosfato
HCO3ATP
ADP
Biotina
Carbossibiotina

58
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 115 -
Acetil-CoA carbossilasi (EC 6.4.1.2)
• La decarbossilazione e formazione di malonil-CoAavviene nel secondo sito della Acetil-CoA carbossilasi dove si lega acetil-CoA per formare malonil-CoA.
NNH
S
CH3
CH3
O
O
N
O
NH
H
O
O
NHNH
S
CH3
CH3
O
O
N
O
NH
H
S
OCH3
CoA
O O
S
O
CoA
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 116 -
Formazione di Acetil-ACP
CH3
O
SCoA
O
SCoA
O
O
CH3
O
S ACP
SO ACP
CH3 S
O
ACP
O
CH3
O
SCoA
O
O
O
O (CH2)14
CH3
Acetil-CoA Malonil-CoA
Acetil-ACP
H2O
NADPH
NADP+
NADP+
NADPH
CO2
CO2
CO2
INVERSO DELLA β-OSSIDAZIONE
Dopo sette cicli
6
• La reazione è catalizzata da [ACP]acetiltransferasi(EC 2.3.1.38),
• Per il malonil-CoA invece vi è una [ACP]maloniltransferasi(EC 2.3.1.39)

59
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 117 -
Acyl Carrier Protein (ACP)
• Il legame avviene attraverso la Ser terminale che lega il gruppo prostetico:
ACPOP
O
OO
CH3
CH3OH
HO
N
H
O
N
H
SH
Fosfopanteina
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 118 -
Strategia
CH3
O
SCoA
O
SCoA
O
O
CH3
O
S ACP
SO ACP
CH3
S
O
ACP
O
CH3
O
SCoA
O
O
S
O (CH2)14
CH3
ACP
S
O
ACP
OH
CH3
H
S
O
ACP
CH3
H
H
O
O (CH2)14
CH3
Acetil-CoA Malonil-CoA
Acetil-ACP
H2O
NADPH
NADP+
NADP+
NADPH
CO2
CO2
CO2
INVERSO DELLA β-OSSIDAZIONE
Dopo sette cicli
6
Acido grasso sintasiEC 2.3.1.85EC 2.3.1.86
Acetoacetil-ACP
β-chetoacil [ACP]reduttasi
EC 1.1.1.100
β-idrossibutirril[ACP] deidratasi
EC 4.2.1.58
Enoil-[ACP] reduttasiNADPH EC 1.3.1.10
NADH EC 1.3.1.9
D-idrossibutirril-ACP
(nella β-ossidazione
L-idrossienoil-CoA)Crotonil-ACP
Butirril-ACP
β-chetoacil-ACPsintetasi
EC 2.3.1.416 Malonil-CoA
Palmitoil-ACP
Acil-ACP-idrolasiEC 3.1.2.14
Palmitato
60
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 119 -
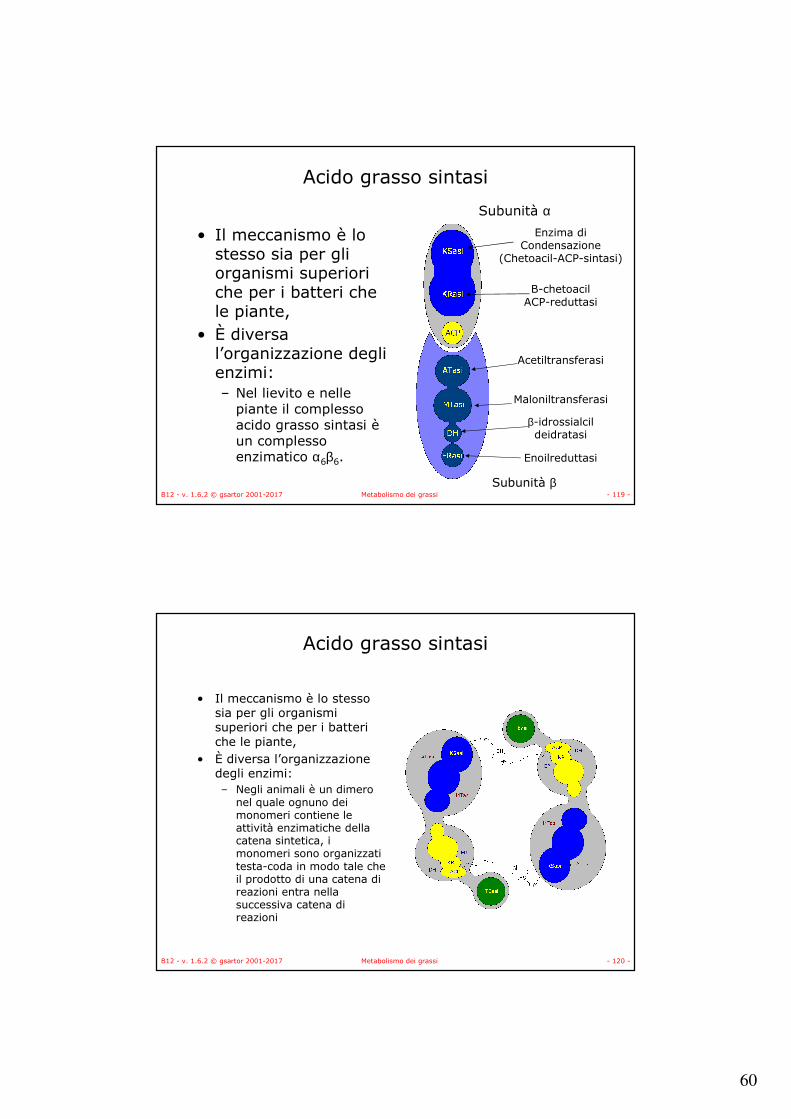
Acido grasso sintasi
• Il meccanismo è lo stesso sia per gli organismi superiori che per i batteri che le piante,
• È diversa l’organizzazione degli enzimi:– Nel lievito e nelle
piante il complesso acido grasso sintasi èun complesso enzimatico α6β6.
Subunità α
Subunità β
Enzima diCondensazione
(Chetoacil-ACP-sintasi)
B-chetoacilACP-reduttasi
Acetiltransferasi
Maloniltransferasi
β-idrossialcildeidratasi
Enoilreduttasi
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 120 -
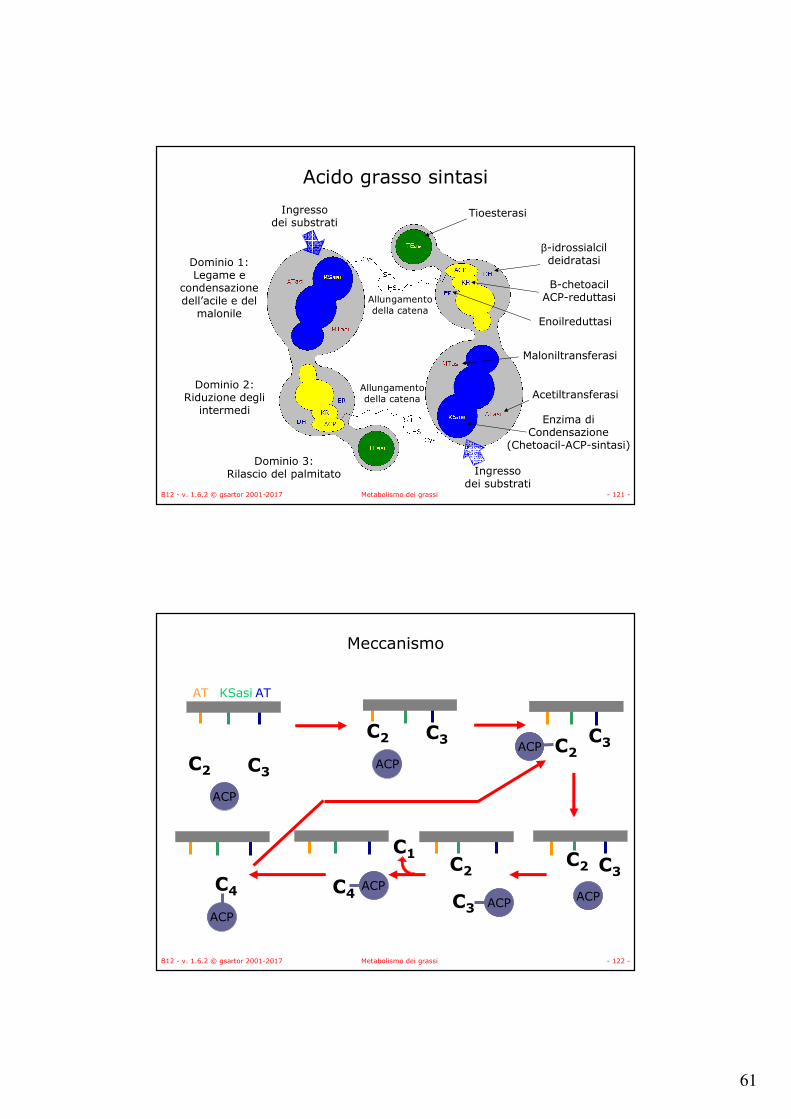
Acido grasso sintasi
• Il meccanismo è lo stesso sia per gli organismi superiori che per i batteri che le piante,
• È diversa l’organizzazione degli enzimi:– Negli animali è un dimero
nel quale ognuno dei monomeri contiene le attività enzimatiche della catena sintetica, i monomeri sono organizzati testa-coda in modo tale che il prodotto di una catena di reazioni entra nella successiva catena di reazioni
61
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 121 -
Acido grasso sintasi
Ingressodei substrati
Ingressodei substrati
Dominio 1:Legame e
condensazionedell’acile e del
malonile
Dominio 2:Riduzione degli
intermedi
Dominio 3:Rilascio del palmitato
Allungamentodella catena
Allungamentodella catena
Enzima diCondensazione
(Chetoacil-ACP-sintasi)
B-chetoacilACP-reduttasi
Acetiltransferasi
Maloniltransferasi
β-idrossialcildeidratasi
Enoilreduttasi
Tioesterasi
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 122 -
Meccanismo
C2 C3
C1
C4
C2 C3 C2C3
C2C2 C3
C3
C4
ACP
ACPACP
ACPACPACP
ACP
AT ATKSasi

62
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 123 -
Meccanismo
AT
O
H
KSasi
S
H
MT
O
H
CH3 S
O
CoAS
O
CoA
O O
ACP
S
H
KSasi
S
H
AT
O
HO
O
O
MT
O
CH3
O
SACP
KSasi
S
H
CH3
O
AT
OO
O
O
MT
O
ACP
S H
AT
O
HO
O
O
MT
O
CH3
O
S
KSasi
ACP
S H
AT
O
H
S
O
O
O
ACP
CH3
O
S
KSasi MT
O
H
AT
O
H H
S
KSasi MT
O
H
S
O
ACP
CH3
AT
O
H
S
O
ACP
O
CH3
S
KSasi
H
MT
O
H
Acetil-CoA
Malonil-CoA
Acetil transferasi Malonil transferasi
3-chetoacil-ACP sintasi
CO2H2O
NADPHNADP+NADP+NADPH
C2 C3
C1
C4
C2 C3 C2C3
C2C2 C3
C3
C4
ACP
ACPACP
ACPACPACP
ACP
AT ATKSasi
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 124 -
Meccanismo
AT
O
H
KSasi
S
H
MT
O
H
CH3 S
O
CoAS
O
CoA
O O
ACP
S
H
KSasi
S
H
AT
O
HO
O
O
MT
O
CH3
O
SACP
KSasi
S
H
CH3
O
AT
OO
O
O
MT
O
ACP
S H
AT
O
HO
O
O
MT
O
CH3
O
S
KSasi
ACP
S H
AT
O
H
S
O
O
O
ACP
CH3
O
S
KSasi MT
O
H
AT
O
H H
S
KSasi MT
O
H
S
O
ACP
CH3
AT
O
H
S
O
ACP
O
CH3
S
KSasi
H
MT
O
H
Acetil-CoA
Malonil-CoA
Acetil transferasi Malonil transferasi
3-chetoacil-ACP sintasi
CO2H2O
NADPHNADP+NADP+NADPH

63
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 125 -
Allungamento della catena
• L’allungamento della catena procede come inverso della β-ossidazione dell’acile attraverso legame con CoA.
• A differenza della β-ossidazione l’enoil-CoA reduttasi usa il NADPH invece del FADH2.
RS
O
CoA
CH3 S
O
CoA HS
CoA
SCoAR
O O
SCoAR
OH O
H
SCoAR
H O
H
SCoAR
O
NADH + H+ NAD+
NADPH + H+NAPD+
Acil-CoA (Cn) β-chetoacil-CoA
L-β-idrossiacil-CoA
α−β-trans-enoil-CoAAcil-CoA (Cn+2)
Tiolasi L-β-idrossiacil-CoAdeidrogenasi
Enoil-CoAidratasi
Enoil-CoAreduttasi
H2O
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 126 -

64
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 127 -
Mono insaturazione
• La singola insaturazione viene inserita nella catena acilica nel processo di allungamento della catena del Acil-ACP.
• Nell’E. Coli agisce una β-idrossidecanoil-ACP idratasi che elimina una molecola d’acqua dal D-b-idrossidecanoil-ACPintermedio.
• Negli eucarioti la reazione avviene attraverso un’ossidazione del prodotto finale (stearoil-CoA, C18) ad opera di O2 nel reticolo endoplasmatico.
O2
S
OCoA
7CH3
7CH3 S
OCoA
16
Stearoil-CoA (C18)
1/2 H2O
Oleil-CoA (C18∆9)
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 128 -
Mono insaturazione
• La singola insaturazione viene inserita nella catena acilica nel processo di allungamento della catena del Acil-ACP.
• Nell’E. Coli agisce una β-idrossidecanoil-ACP idratasi che elimina una molecola d’acqua dal D-b-idrossidecanoil-ACPintermedio.
• Negli eucarioti la reazione avviene attraverso un’ossidazione del prodotto finale (stearoil-CoA, C18) ad opera di O2 nel reticolo endoplasmatico.
S
OCoA
7CH3
7
CH3 S
OCoA
16
O2
Stearoil-CoA (C18)
Oleil-CoA (C18∆9)
H2O+Desaturasi
(Fe+++)
Desaturasi(Fe++)
2 Cyt b5
(Fe++)
2 Cyt b5
(Fe+++)
Cyt b5
reduttasi(Fe++)
Cyt b5
reduttasi(Fe+++)
NADH + H+
NAD+ + 1/2
2H+
+ 2H+

65
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 129 -
Poli insaturazione
• La gestione degli acidi grassi polinsaturi (PUFA) è diversa nei diversi organismi, quindi i processi sono diversi a seconda dell’organismo.
• Nei mammiferi:
O
O
O
SCoA
O
SCoA
O
SCoA
O
SCoA
O
O
Acil-CoAsintetasi
Linoleato (C18-∆9, 12)
Linoleil-CoA (C18-∆9, 12-CoA)
C18-∆6, 9, 12-CoA
CoA-SH + ATP
AMP + PPi
Malonil-CoA
CO2 + CoASH
C20-∆8, 11, 14-CoA
2H+
Arachidonoil-CoA (C20-∆5, 8, 11, 14-CoA)
Arachidonato (C20-∆5, 8, 11, 14)
H2O
CoASH
Desaturazione
Desaturazione
2H+
Allungamento
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 130 -
Controllo e regolazione della biosintesi
• L’acetil-CoA ha un ruolo centrale della regolazione del metabolismo degli acidi grassi e dei glucidi.
• L’acetil-CoA carbossilasi è regolata allostericamente dal citrato (attivatore) e dagli acil-CoA (inibitore).
• Il malonil-CoA agisce invece come inibitore del trasporto di Acil-CoA all’interno dei mitocondri a livello della formazione dell’acil-carnitina.
• Vi è poi un azione di controllo a livello di interazione tra gli organi mediata dagli ormoni attraverso le cascate enzimatiche attivate dal cAMP.

66
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 131 -
Controllo e regolazione della biosintesi
Piruvato
Glucosio
Glicolisi
Enzimamalico
Malato
Malatodeidrogenasi
Ossalacetato
Citrato
Acil-carnitina
AcidoGrassoAcil-CoA
Aminoacidi
Acetil-CoA
ATP-citratoliasi
Acil-CoA
Ciclodi Krebs
Citrato
Ossalacetato
Malatodeidrogenasi
Citratosintasi
Acetil-CoA
Acil-carnitina
β-ossidazione
Piruvato
Malato
Piruvatocarbossilasi
Piruvatodeidrogenasi
NAD+NADH
NADP+
NADPH
ATP
ADP + Pi
CO2NAD+
NADH
CO2
NAD+
NADH
NAD+
NADH
CO2
MATRICEMITOCONDRIALE
LATOCITOPLASMA
Malonil-CoAACC
Citrato
Malato
Piruvato
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 132 -
Acetil-CoACarbossilasi
(attiva a basso citrato)
Controllo e regolazione della biosintesi
Ormone(Adrenalina o glucagone)
Adenilatociclasi
(inattiva)
Adenilato ciclasi(attiva)
Fosfodiesterasi
AMP
FosfatasiPi
Fosfatasi
Pi
ATP
ADP Acetil-CoACarbossilasi(attiva ad
alto citrato)
P
P
Fosforilasi chinasi(b-inattiva)
Fosforilasi chinasi(a-attiva)
ATP
ADP
ATP cAMP + PPiProteinachinasi A(inattiva)
Proteinachinasi A(attiva)
Gα-GTP

67
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 133 -
Metabolismo dei grassi
• Demolizione dei trigliceridi• Catabolismo degli acidi grassi• Biosintesi
–degli acidi grassi e colesterolo–dei lipidi complessi
• Trasporto dei lipidi
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 134 -
Lipidi
• Semplici– Sono molecole che non contengono legami
esterei o amidici• Agidi grassi• Colesterolo
• Complessi– Sono derivati di acidi grassi variamente
esterificati o amidati.• Glicerofosfolipidi e sfingosidi• Trigliceridi

68
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 135 -
Colesterolo• Il colesterolo appartiene alla
famiglia degli steroidi,
• È un componente fondamentale delle membrane biologiche,
• È il precursore nella sintesi degli acidi biliari (acido colico, acido taurocolico, acido glicocolico) e della sintesi degli ormoni steroidei (testosterone, estradiolo, progesterone) e della vitamina D3.
• Viene sintetizzato prevalentemente nelle cellule epatiche a partire da acetil-CoA.
CH3
CH3
CH3
OH
CH3
CH3
H
H
H
H
CH3
CH3
OH
CH3
CH3
CH3H
HH
H
H
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 136 -
Biosintesi del colesterolo
• Viene sintetizzato nelle cellule epatiche a partire da acetil-CoA per formare 3-R-mevalonato.
CH3 S
O
CoA
CH3 S
O
CoA HS
CoA
SCoACH3
O O
SCoA
OH O
CH3
O
O
CH3 S
O
CoA HS
CoA
SCoA
OH OH
CH3
O
O
HOH
OH H
CH3
O
O
H
HS
CoA
NADPH + H+
NAPD+
Acetil-CoA (Cn) Acetoacetil-CoA
3-idrossi-3-metilglutaril-CoA
HMG-CoA
3-R-mevalonato
Tiolasi HMG sintasi
HMG-CoA reduttasi I riduzione
Intermedio legatoall'enzima
NADPH + H+NAPD+
II riduzione
69
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 137 -
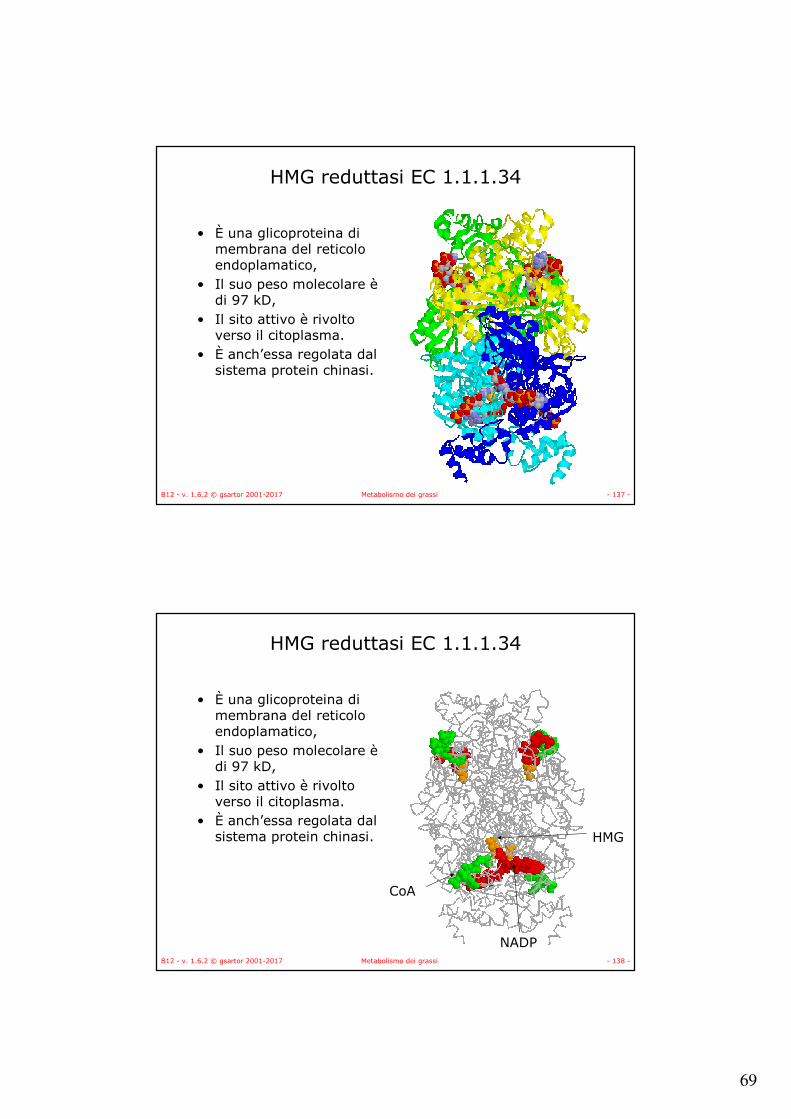
HMG reduttasi EC 1.1.1.34
• È una glicoproteina di membrana del reticolo endoplamatico,
• Il suo peso molecolare èdi 97 kD,
• Il sito attivo è rivolto verso il citoplasma.
• È anch’essa regolata dal sistema protein chinasi.
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 138 -
HMG reduttasi EC 1.1.1.34
• È una glicoproteina di membrana del reticolo endoplamatico,
• Il suo peso molecolare èdi 97 kD,
• Il sito attivo è rivolto verso il citoplasma.
• È anch’essa regolata dal sistema protein chinasi. HMG
CoA
NADP

70
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 139 -
HMG reduttasi EC 1.1.1.34
• Viene inibita dalle statine, usate come farmaci per ridurre elevati livelli di colesterolo.
Mevastatina
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 140 -
HMG-CoA reduttasichinasi (inattiva)
Controllo e regolazione della biosintesi del colesteroloOrmone
(Adrenalina o glucagone)
Adenilatociclasi
(inattiva)
Adenilato ciclasi(attiva)
Fosfodiesterasi
AMP
FosfatasispecificaPi
PATP
ADP
ATP cAMP + PPi
HMG-CoA reduttasichinasi chinasi (attiva)
Gα-GTP
HMG-CoA reduttasichinasi chinasi (inattiva)
HMG-CoA reduttasichinasi (attiva)
ATP
ADP
HMG-CoAreduttasi(attiva)
PHMG-CoAreduttasi(inattiva)
FosfatasispecificaPi
3-R-mevalonato
HMG-CoA
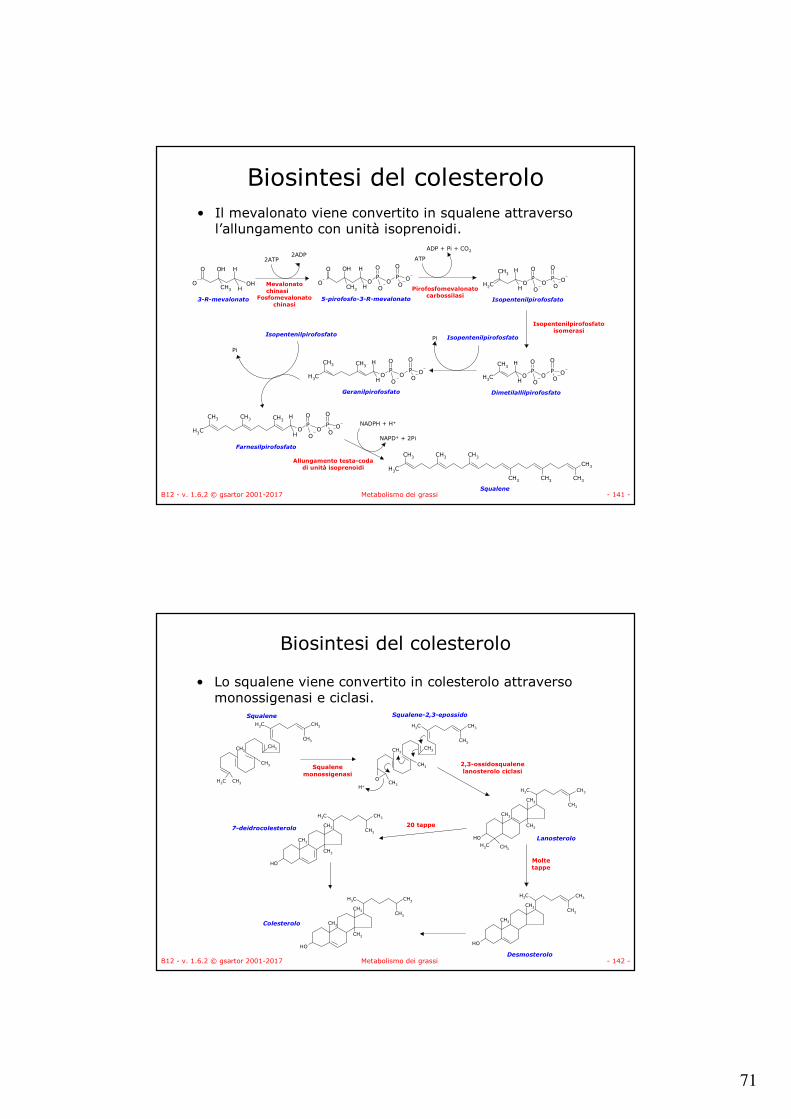
71
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 141 -
Biosintesi del colesterolo• Il mevalonato viene convertito in squalene attraverso
l’allungamento con unità isoprenoidi.
OH
OH H
CH3
O
O
H
P
O
OOO
OH H
CH3
O
O
H
P
O
OO P
O
OOOCH2
HCH3
H
P
O
OO
P
O
OOOCH3
HCH3
H
P
O
OOCH3
CH3
P
O
OOO
HCH3
H
P
O
OO
CH3
CH3 CH3
P
O
OOO
HCH3
H
P
O
OO
CH3
CH3
CH3CH3CH3
CH3CH3CH3
3-R-mevalonato
Mevalonatochinasi
5-pirofosfo-3-R-mevalonato
2ATP2ADP
ATP
ADP + Pi + CO2
IsopentenilpirofosfatoFosfomevalonatochinasi
Pirofosfomevalonatocarbossilasi
Dimetilallilpirofosfato
Isopentenilpirofosfatoisomerasi
Pi
Geranilpirofosfato
Pi
IsopentenilpirofosfatoIsopentenilpirofosfato
Farnesilpirofosfato
NADPH + H+
NAPD+ + 2Pi
Allungamento testa-codadi unità isoprenoidi
Squalene
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 142 -
Biosintesi del colesterolo
• Lo squalene viene convertito in colesterolo attraverso monossigenasi e ciclasi.
CH3CH3
CH3
CH3
CH3
CH3
CH3
CH3
CH3
CH3
CH3
CH3
CH3
CH3
CH3
O
CH3
CH3
CH3
CH3
CH3
CH3
OHCH3
CH3
CH3
CH3
CH3
CH3
CH3
OH
CH3
CH3
CH3CH3
CH3
OH
CH3
CH3
CH3
CH3
CH3
CH3
OH
CH3
Squalenemonossigenasi
Squalene Squalene-2,3-epossido
H+
Lanosterolo
2,3-ossidosqualenelanosterolo ciclasi
7-deidrocolesterolo
Desmosterolo
20 tappe
Moltetappe
Colesterolo
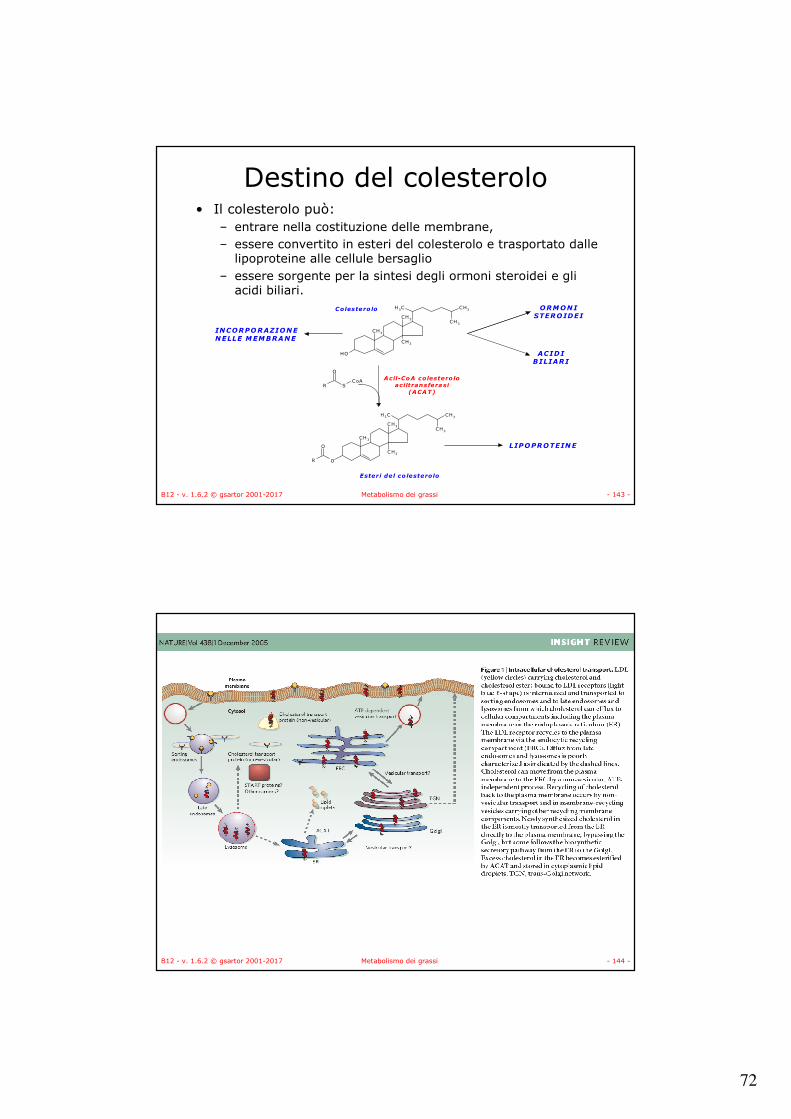
72
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 143 -
Destino del colesterolo• Il colesterolo può:
– entrare nella costituzione delle membrane,– essere convertito in esteri del colesterolo e trasportato dalle
lipoproteine alle cellule bersaglio– essere sorgente per la sintesi degli ormoni steroidei e gli
acidi biliari.
CH 3
CH3
CH 3
CH 3
CH 3
OH
CH 3
SCoA
O
R
CH 3
CH3
CH3
CH3
CH 3
O
C H 3
O
R
Colestero lo
INCORPO RAZIONE
NELLE M EM BRANE
Acil-CoA colestero loaciltransferasi
(ACA T)
ORM ONISTEROIDEI
Esteri del co lesterolo
LIPOPROTEIN E
ACIDIBILIARI
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 144 -

73
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 145 -
Metabolismo dei grassi
1. Demolizione dei trigliceridi2. Catabolismo degli acidi grassi
3. Biosintesi – degli acidi grassi e colesterolo– dei lipidi complessi
4. Trasporto dei lipidi
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 146 -
Lipidi
• Semplici– Sono molecole che non contengono legami
esterei o amidici• Agidi grassi• Colesterolo
• Complessi– Sono derivati di acidi grassi variamente
esterificati o amidati.• Glicerofosfolipidi e sfingosidi• Trigliceridi
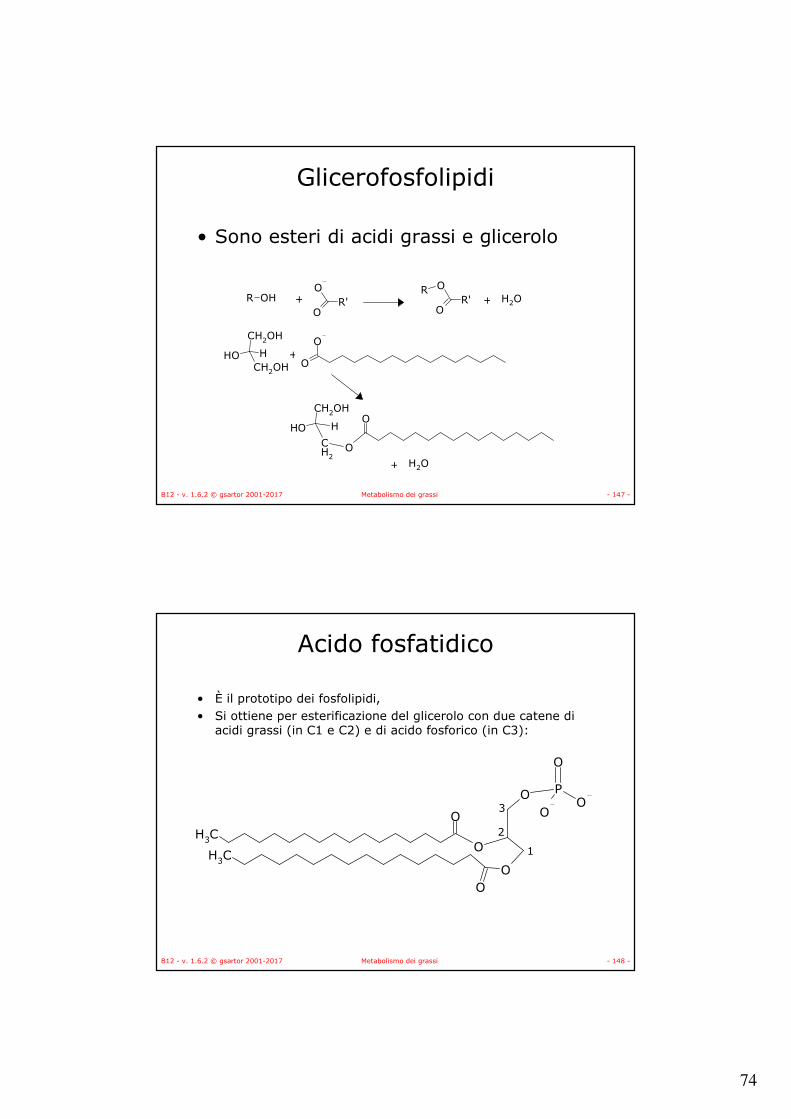
74
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 147 -
Glicerofosfolipidi
• Sono esteri di acidi grassi e glicerolo
O
O
CH2OH
CH2OHOH H
R OH R'O
OR'
O
O
R
CH2OH
CH2
OH H
O
O
H2O++
+
H2O+
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 148 -
Acido fosfatidico
• È il prototipo dei fosfolipidi,• Si ottiene per esterificazione del glicerolo con due catene di
acidi grassi (in C1 e C2) e di acido fosforico (in C3):
O
OCH3
OCH3
O
P
O
OO
O
1
2
3

75
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 149 -
Biosintesi dell’acido fosfatidico
• L’acido fosfatidico viene sintetizzato a partire da glicerolo-3-P e acil-ACP.
• Negli eucarioti la sorgente è il diidrossiacetonfosfato
OH
OH
OH
OH
O
O
P
O
OO
P
O
OO
O
OH
OH
O
O
O
P
O
OO
O
R
O
R
SCoA
H SCoA
O
O
OH
P
O
OO
O
R
O
R
SCoA
H SCoA
O
R
SACP
H SACP
O
R
SCoA
H SCoA
O
R
SACP
H SACP
O
R
O
O
O
R
O
POOO
o
o
o
o
NADH + H+ NAD+
NADPH + H+ NAPD+
ATPADP
GliceroloGlicerolo-3-fosfatoDiidrossiacetonfosfato
1-acil-diidrossiacetonfosfato
1-acil-glicerolo-3-fosfato
Acido fosfatidico
Glicerolochinasi
Glicerolo-3-fosfato
deidrogenasi
Diidrossiaceton
fosfatoaciltransferasi
Glicerolo-3-fosfato
aciltransferasi
Acil-diidrossiaceton
fosfato reduttasi
1-acilglicerolo
aciltransferasi
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 150 -
Fosfolipidi
• Nei fosfolipidi la funzione acida libera del gruppo fosfato è esterificata con un alcool:
– Colina
– Serina
– Etanolamina
– Glicerolo
– Inositolo
O
OCH3
OCH3
O
P
O
OO
ORNH2
OH
O
O
N+
OH
CH3 CH3
CH3
OH
OH
OH
OH
OH
OH
NH3
+OH
CH2OH
CH2
HOH
OH
1
2
3
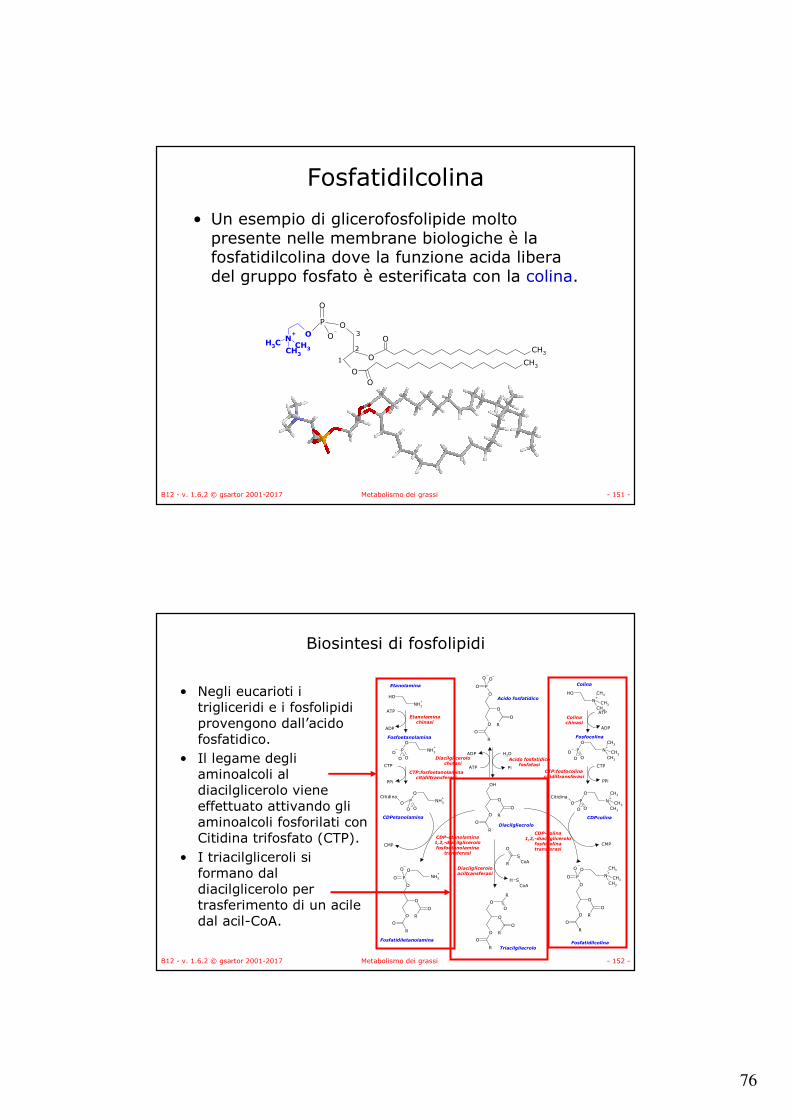
76
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 151 -
Fosfatidilcolina
• Un esempio di glicerofosfolipide molto presente nelle membrane biologiche è la fosfatidilcolina dove la funzione acida libera del gruppo fosfato è esterificata con la colina.
N+ O
CH3CH3
CH3
O
OCH3
OCH3
O
P
O
OO
1
2
3
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 152 -
Biosintesi di fosfolipidi
• Negli eucarioti i trigliceridi e i fosfolipidi provengono dall’acido fosfatidico.
• Il legame degli aminoalcoli al diacilglicerolo viene effettuato attivando gli aminoalcoli fosforilati con Citidina trifosfato (CTP).
• I triacilgliceroli si formano dal diacilglicerolo per trasferimento di un aciledal acil-CoA.
O
O
O
O
R
O
R
P
OO
O
O
OH
O
O
R
O
R
O
O
O
O
R
O
R
O
R
O
O
O
O
R
O
R
P
OO
O N+CH3
CH3CH3
O
O
O
O
R
O
R
P
OO
O NH3
+
OH
N+CH3
CH3CH3
O
N+CH3
CH3CH3
P
OOO
O
N+CH3
CH3CH3
P
OOO
Citidina
OH
NH3
+
O
NH3
+P
OOO
O
NH3
+P
OOO
Citidina
O
R
SCoA
H SCoA
CMP
CTP
PPi
ATP
ADP
CMP
CTP
PPi
ATP
ADP
ATP
ADP
Pi
H2O
Acido fosfatidico
Diacilgliecrolo
Triacilgliecrolo
FosfatidiletanolaminaFosfatidilcolina
Colina
Fosfocolina
CDPcolina
Etanolamina
Fosfoetanolamina
CDPetanolamina
Diacilglicerolochinasi
Acido fosfatidicofosfatasi
Colinachinasi
CTP:fosfocolinacitidiltransferasi
CTP:fosfoetanolaminacitidiltransferasi
Etanolaminachinasi
CDP-etanolamina1,2,-diacilglicerolofosfoetanolamina
transferasi
CDP-colina1,2,-diacilglicerolo
fosfocolinatransferasi
Diacilgliceroloaciltransferasi

77
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 153 -
Biosintesi di fosfatidilserina
• Nei mammiferi la sintesi di fosfatidilserina avviene attraverso la scambio con etanolamina.
O
O
O
O
R
O
R
P
OO
O NH3
+
NH3+
OH
O
ONH3+
OH
O
O
NH3+
OHNH3+
OH
O
O
O
O
R
O
R
P
OO
ONH3
+
O
O
Enzima di scambiodelle basi
(reticolo endoplasmatico)
Fosfatidiletanolamina
Fosfatidilserina
CO2
Fosfatidilserinadecarbossilasi(mitocondri)
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 154 -
Metabolismo dei grassi
1. Demolizione dei trigliceridi2. Catabolismo degli acidi grassi
3. Biosintesi – degli acidi grassi e colesterolo– dei lipidi complessi
4. Trasporto dei lipidi

78
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 155 -
Complessi lipoproteici
• I lipidi circolano negli organismi sottoforma di complessi lipoproteici:
• Gli acidi grassi sono coniugati con l’albumina• I triacilgliceroli, i fosfolipidi, il colesterolo ed i suoi esteri
sono trasportati come lipoproteine.
100-500< 0.950Chilomicroni
30-800.950-1.006VLDL (Very Low Density Lipoprotein)
25-501.006-1.019IDL (Intermediate Density Lipoprotein)
18-281.019-1.063LDL (Low Density Lipoprotein)
5-151.063-1.210HDL (High Density Lipoprotein)
Diametro (nm)
Densità(g/ml)
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 156 -
Complessi lipoproteici
• I lipidi circolano negli organismi sottoforma di complessi lipoproteici:
• Gli acidi grassi sono coniugati con l’albumina• I triacilgliceroli, i fosfolipidi, il colesterolo ed i suoi esteri
sono trasportati come lipoproteine.
Colesterolo Fosfolipidi
Chilomicroni
VLDL (Very Low Density Lipoprotein)
IDL (Intermediate Density Lipoprotein)
LDL (Low Density Lipoprotein)
HDL (High Density Lipoprotein)
TriacilgliceroliProteine

79
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 157 -
Complessi lipoproteici
• Le lipoproteine vengono sintetizzate nel fegato e nell’intestino (chilomicroni).
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 158 -
Complessi lipoproteici
• Le lipoproteine vengono sintetizzate nel fegato e nell’intestino (chilomicroni).

80
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 159 -
Complessi lipoproteici
Nei chilomicroni, VLDL, IDL, HDL3-634.100E
Nelle HDL8-1032.500D
Nei chilomicroni, VLDL, IDL, HDL8-158.800C-3
Nei chilomicroni, VLDL, IDL, HDL3-88.800C-2
Nei chilomicroni, VLDL, IDL, HDL4-77.000C-1
Proteina principale nelle LDL80-100500.000B-100
Solo nei chilomicroni<5240.000B-48
Si trova come dimero nelle HDL30-508.700A-2
Proteina principale nelle HDL90-12028.000A-1
DistribuzioneConcentrazione
nel plasma (mg/100ml)
PMApoproteina
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 160 -
Complessi lipoproteici• La sintesi delle
apoproteine, dei fosfolipidi, del triacilglicerolo, del colesterolo e dei suoi esteri avviene nel reticolo endoplasmatico,
• L’assemblaggio dei componenti nelle prelipoproteineavviene nel RE e queste vengono trasferite nel Golgi.
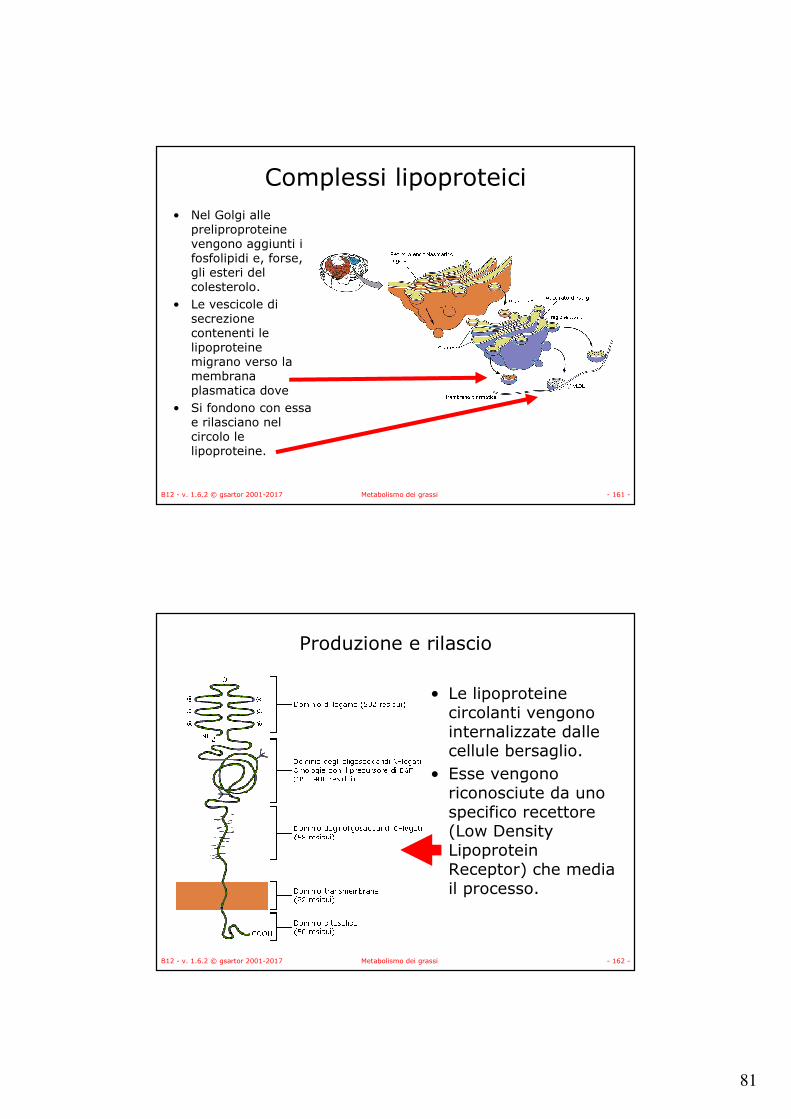
81
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 161 -
Complessi lipoproteici• Nel Golgi alle
preliproproteinevengono aggiunti i fosfolipidi e, forse, gli esteri del colesterolo.
• Le vescicole di secrezione contenenti le lipoproteine migrano verso la membrana plasmatica dove
• Si fondono con essa e rilasciano nel circolo le lipoproteine.
B12 - v. 1.6.2 © gsartor 2001-2017 Metabolismo dei grassi - 162 -
Produzione e rilascio
• Le lipoproteine circolanti vengono internalizzate dalle cellule bersaglio.
• Esse vengono riconosciute da uno specifico recettore (Low Density LipoproteinReceptor) che media il processo.

82
Crediti e autorizzazioni all’utilizzo• Questo materiale è stato assemblato da informazioni raccolte dai seguenti testi di Biochimica:
– CHAMPE Pamela , HARVEY Richard , FERRIER Denise R. LE BASI DELLA BIOCHIMICA [ISBN 978-8808-17030-9] – Zanichelli
– NELSON David L. , COX Michael M. I PRINCIPI DI BIOCHIMICA DI LEHNINGER - Zanichelli – GARRETT Reginald H., GRISHAM Charles M. BIOCHIMICA con aspetti molecolari della Biologia
cellulare - Zanichelli– VOET Donald , VOET Judith G , PRATT Charlotte W FONDAMENTI DI BIOCHIMICA [ISBN 978-
8808-06879-8] - Zanichelli
• E dalla consultazione di svariate risorse in rete, tra le quali:– Kegg: Kyoto Encyclopedia of Genes and Genomes http://www.genome.ad.jp/kegg/– Brenda: http://www.brenda.uni-koeln.de/– Protein Data Bank: http://www.rcsb.org/pdb/– Rensselaer Polytechnic Institute:
http://www.rpi.edu/dept/bcbp/molbiochem/MBWeb/mb1/MB1index.html
• Il materiale è stato inoltre rivisto e corretto dalla Prof. Giancarla Orlandini dell’Università di Parma alla quale va il mio sentito ringraziamento.
Questo ed altro materiale può essere reperito a partire da: http://www. gsartor.org/pro
• Il materiale di questa presentazione è di libero uso per didattica e ricerca e può essere usato senza limitazione, purché venga riconosciuto l’autore usando questa frase:
Materiale ottenuto dal Prof. Giorgio Sartor
Università di Bologna
Giorgio SartorUfficiale: [email protected]: [email protected]
Aggiornato il 11/04/2017 10:15:50