Alcuni dati sulla embriologia ed embriogenesi di « Olea Europaea » L.
23
This article was downloaded by: [York University Libraries] On: 19 October 2014, At: 18:42 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Giornale botanico italiano: Official Journal of the Societa Botanica Italiana Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/tplb19 Alcuni dati sulla embriologia ed embriogenesi di « Olea Europaea » L. Prof. Albina Messeri Published online: 14 Sep 2009. To cite this article: Prof. Albina Messeri (1950) Alcuni dati sulla embriologia ed embriogenesi di « Olea Europaea » L., Giornale botanico italiano: Official Journal of the Societa Botanica Italiana, 57:1-2, 149-169, DOI: 10.1080/11263505009430783 To link to this article: http://dx.doi.org/10.1080/11263505009430783 PLEASE SCROLL DOWN FOR ARTICLE Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) contained in the publications on our platform. However, Taylor & Francis, our agents, and our licensors make no representations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of the Content. Any opinions and views expressed in this publication are the opinions and views of the authors, and are not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon and should be independently verified with primary sources of information. Taylor and Francis shall not be liable for any losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoever or howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use of the Content. This article may be used for research, teaching, and private study purposes. Any substantial or systematic reproduction, redistribution, reselling, loan,
Transcript of Alcuni dati sulla embriologia ed embriogenesi di « Olea Europaea » L.

This article was downloaded by: [York University Libraries]On: 19 October 2014, At: 18:42Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH,UK
Giornale botanico italiano:Official Journal of the SocietaBotanica ItalianaPublication details, including instructions forauthors and subscription information:http://www.tandfonline.com/loi/tplb19
Alcuni dati sulla embriologiaed embriogenesi di « OleaEuropaea » L.Prof. Albina MesseriPublished online: 14 Sep 2009.
To cite this article: Prof. Albina Messeri (1950) Alcuni dati sulla embriologia edembriogenesi di « Olea Europaea » L., Giornale botanico italiano: Official Journal ofthe Societa Botanica Italiana, 57:1-2, 149-169, DOI: 10.1080/11263505009430783
To link to this article: http://dx.doi.org/10.1080/11263505009430783
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all theinformation (the “Content”) contained in the publications on our platform.However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness,or suitability for any purpose of the Content. Any opinions and viewsexpressed in this publication are the opinions and views of the authors, andare not the views of or endorsed by Taylor & Francis. The accuracy of theContent should not be relied upon and should be independently verified withprimary sources of information. Taylor and Francis shall not be liable for anylosses, actions, claims, proceedings, demands, costs, expenses, damages,and other liabilities whatsoever or howsoever caused arising directly orindirectly in connection with, in relation to or arising out of the use of theContent.
This article may be used for research, teaching, and private study purposes.Any substantial or systematic reproduction, redistribution, reselling, loan,

sub-licensing, systematic supply, or distribution in any form to anyone isexpressly forbidden. Terms & Conditions of access and use can be found athttp://www.tandfonline.com/page/terms-and-conditions
Dow
nloa
ded
by [
Yor
k U
nive
rsity
Lib
rari
es]
at 1
8:42
19
Oct
ober
201
4

Prof. ALBINA MESSERI
ALCUNI DATI SULLA EMBRIOLOGIA ED EMBRIOGENESI
DI (( OLEA EUROPAEA * L. (*)
(CON 40 FIGURE NEL TESTO)
prewntato 11 23 aprile 1949
La letteratura sull’olivo dal punto di vista embriologico ed einbrio- genetic0 t piuttosto scarsa, mentre abbondano dati generici sulla bio- logia di questa pianta. La memoria pib recente ed aggiornata sulla sistc- matica, distribuzionc geografica c coltivazione dell‘olivo t quella dcl CIFERRI (1942).
La diffcrenziazione delle gernme a fiore da quclle a legno nell’olivo si compie nci mesi inimediatamente precedenti la fioritura (VICODSKI DE PHILIPPIS 1937, 1938), e la fioritura di questa specie L stata oggetto di molti studi, fra i quali citerb quelli del CAMPBELL (1910, 1912 (I, 1912 by 1913) e quelli recenti del MORETTINI (1939, 1940 Q, 1940 b, 1949), che hanno portato alla conoxenza della autosterilith di molte razze di olivo e della tendenza verso la maxolinith, assai spiccata specialmente in certe razze meridionali (CAMPBELL 1910, 1912 a; PIROTTA e DE PERGOLA 1913 b). La abbondantissima fioritura L seguita dalla cascola, che si verifica in epoche pib o meno h e e sulle cui cause si discute molto. Della frazione di fiori che sfugge alla cascola, una partc si sviluppa in drupe normali, con un solo seme, per aborto precoce degli altri tre ovuli del pistillo, ed una parte, variabile da pianta a pianta e di anno in anno, si sviluppa in piccolissime olive partenocarpiche (FAVILLI 1943-45, 1947) dotate di tutte le caratteristiche delle drupe normali, per quello chc riguarda la costituzione del frutto, mentre all’interno dell’endocarpo si trovano tutti e quattro gli ovuli all0 stesso stadio di sviluppo, che hanno nel lor0 interno i resti cli un gametofito degenerante, senza traccia n i di zigoto n t di albume (MESSERI 1947). Sernbra quindi trattarsi di frutti parte- nocarpici, la cui presenza nell’olivo era del resto gih stata prospettata da PIROTTA e DE PERGOLA (1913 a). Sulla ernbriologia ed embriogenia del-
(*) Lavoro eseguito nell’Istituto Botanico dell‘universiti di Bari, diretto dalla --
Prof.ssa E. FRANCINI.
1491
. . [Nuovo Ciornale Botanico Itallano, n. s., vol. LVII , n. 1-2, 1950.
Dow
nloa
ded
by [
Yor
k U
nive
rsity
Lib
rari
es]
at 1
8:42
19
Oct
ober
201
4

150 MESSERI
I’olivo invece non si hanno dati organici in letteratura, ma solo notizie sui primi stadi di sviluppo del gametofito femminile (PIROTTA e DE PER- GOLA 1913 a) e pih recentemente dati pih numerosi ricavabili dalla memoria di ANDERSON (1931) sulla embriologia delle Celustraceae, Oleaceae e Apocinaceae. L’A. riferisce diverse notizie su Olea europaea, 0. chryso- phylla e 0. capensis; i suoi dati sono frammentari, ma costituiscono una preziosa informazione inquantocht si inquadrano con quelli di numerose altre Oleaceae studiate. A questi dati in maniera speciale mi riferirb nel corm di questo lavoro. Infine i dati pih recenti sulla cariologia del gencre Olea sono quelli di TAYLOR (1945).
La frammentarieth dei dati embriologici su questa pianta di cosi largo interesse, si spiegano forse colla difficoltl che essa offre in queste indagini dal punto di vista della tecnica istologica e citologica; queste dif- ficoltl sono state superate soltanto in parte da me, cosiccht sono riuscita a mettere insieme una serie di dati non del tutto conipleti. Le mie ri- cerche continuano tuttora, tuttavia i fatti finora accertati mi sembrano abbastanza numerosi e coordinati da decidermi a pubblicarli.
I dati che ho raccolto in questa nota si riferiscono in grande maggio- ranza ad una pianta della var. Ogliarola, che cresce nell’Orto Botanico dell’htituto di Bari; ho raccolto infatti, dal 1943, il materiale sempre dalla medesima pianta, per p t e r e avere un quadro pic facilmente inter- pretabile dal punto di vista biologico, senza indurvi variazioni dovute a differenza di razza, tanto piti possibili trattandosi di una pianta colti- vata. Ho anche utilizzato un poco del materiale raccolto dal compianto dott. ALDO BERCOVICH, gih assistente presso 1’Istituto Botanico di Bari.
I1 materiale offre una notevole difficolta alla fissazione e alla colora- zione. I fissativi che ho usato, con miglior rendimento, sono: Juell, Kar- petschenko, e soprattutto Delaunay. 11 Regaud all’acido picrico non ha dato resultati sodisfacenti. Le colorazioni usate sono state: ematossilina e safranina e, soprattutto, ematossilina ferrica.
Dcscrizione dell0 svolgimento delle diverse tappe dei jenorneni osservati. La mignolatura, come b noto, varia un poco nel tempo a seconda
dell’andamento della stagione; tuttavia, ai primi di Maggio, il mio olivo si trova circa agli inizi della fioritura, con la maggior parte dei fiori in piccoli bocci.
la settimana d i maggio. - I fiori pih giovani prelevati a questa data, ancora chiusi, presen_tano le antere abbastanza sviluppate, con una epidermide di uno strato di cellule ancora normalmente turgide con nucleo fortemente colorabile all’ematossilina ferrica e membrana spessa
Dow
nloa
ded
by [
Yor
k U
nive
rsity
Lib
rari
es]
at 1
8:42
19
Oct
ober
201
4

1 EMBRIOLOGIA ED EMBRIOGENESI DI OLEA EUROPAEA L . ,151
ed un tessuto meccanico di uno strato di cellule a membrana sottile e cito- plasma molto vacuolizzato; fanno seguito il tessuto transitorio, formato da uno o due strati di cellule a carattere ancora meristematico, con nuclei grandi, citoplasma denso e pareti sottili, ed il tappeto, formato da uno strato di cellule non ancora molto allungate in serw radiale, multinu- cleate, con grandi nucleoli in ciascun nucleo.
L'interno della sacca pollinica L occupato dal tessuto sporigeno. A questo momento le cellule madri delle spore sono tutte in sinapsi. In molte antere si notano aree pih o meno grandi di cellule sporigene in degenerazione, sempre disposte in posizione periferica, subito sotto le cellule del tappeto.
Come t noto, i racemi di olivo sono composti di piccole infiorescenze a tre fiori, di cui il centrale, e pih lungamente pedicellato, t il pih precoce. Quando il fiore centrale L nello stadio ora descritto per quello che ri- guarda le antere, nei due laterali non si I: ancora delineata una distin- zione fra tessuto del tappeto e tessuto sporigeno; si distingue soltanio l'epidermide, if tessuto meccanico e il tessuto di transizione. Le future cellule del tappeto hanno per6 dimensioni maggiori e sono allungate in senso radiale in confront0 alle cdllule pih interne.
I1 gineceo in tutti i fiori L ancora molto poco progredito nel suo svi- luppo. Si vedono i quattro ovuli, abbozzati, nell'oyario; lo stilo e lo stigma sono molto lontani dal lor0 completo sviluppo. I n particolare, lo stigma si presenta come uno slargamento all'apice dello stilo, senza nessuna dif- ferenziazione di tessuto recettore.
Ho potuto inoltre cogliere pochi stadi intermedi fra quelli preceden- temente descritti e quelli trovati nella seconda settimana di Maggio. Si tratta di fiori con antere contenenti microspore gi21 libere (fig. 16) e con pistilli contenenti ovuli, che presentano cellule madri delle megaspore in sinapsi. A questo stadio il corpo ovulare t assai sviluppato, l'ovulo non ha ancora raggiunto la suanormale posizione anatropa e la nucella, che ancora non k rivestita dal tegumento, t contigua alle pareti dell'ovario (fig. 1). Contrariamente a quanto ha osservato ANDERSON (1931), non si accenna ancora alla formazione del canale micropilare.
La nucella appare di un solo strato all'apice e di pih strati verso la base (fig. 3). La cellula madre L di forma caratteristicamente rettangolare col grande nucleo in sinapsi spostato verso la met3 apicale di essa (fig. 2).
2a settimana di maggio. - I1 materiale che ho raccolto in questa data t abbondante, ma piuttosto omogeneo. Le antere hanno il polline gih formato. L'epidermide dell'antera va incontro a disfacimento e C O S ~
il tessu-to meccanico e quell0 di transizione. Pih individuabili sono le cel- lule del tappeto, che appaiono pih pallide che nelle antere descritte
Dow
nloa
ded
by [
Yor
k U
nive
rsity
Lib
rari
es]
at 1
8:42
19
Oct
ober
201
4

152 MESSERI
prima; tuttavia si distinguono bene in ogni cellula due o tre grandi nu- clei. Le cellule del tappeto sporgono verso la cavita della sacca pollinica e sono distinte le une dalle altre; si tratta di un tappeto di secrezione. I singoli granelli di polline sono rotondeggianti, senza una differenriarione fra endina ed esina e con un solo grande nucleo, nel citoplasma vacuoliz- zato. Qua e lh si notano delle zone di polline degenerante. I1 polline uni- nucleato era stat0 notato anche da’ANDERSoN (1931). 11 fiore in questo stadio i ancora con la corolla chiusa.
I1 pistillo i formato da un ovario diviso in due loggie, in ciaxuna delle quali trovano posto due ovuli anatropi. L’ovario si prosegue in uno stilo tozzo, che termina in alto con due stigmi papillosi. Sul tesuto stigmatic0 non si notano granelli di polline a questo stadio. .Del resto non c’i: polline fuori delle antere.
I quattro ovuli sono tutti delle stesse dimensioni, dello s t e m grado di vivacith e circa tutti e quattro nello s t e m stadio di sviluppo.Essi sono formati da un grande e s p e w tegumento, che forma il corpo dell’ovulo, alla radice del quale, completamente racchiusa da esso, sta la nucella, che contiene una diade. La nucella t formata da un solo strato di cellule all’apice, dove la cellula superiore della diade I: a contatto con l’epider- mide nucellare, ed invece i: pit3 spessa verso la base, dove, fra l’epider- mide nucellare e la cellula inferiore della diade, si frappongono una o due serie di piccole cellule schiacciate longitudinalmente. Questa dispo- sizione k stata osservata nel genere Olea anche da ANDERSON (1931).
presumibile che lo stadio di diade, cioi: di intercinesi fra la prima e la seconda divisione meiotica, sia, contrariamente -a1 comune, assai lungo nell’Olivo (figg. 4, 5).
Gih all0 stadio di diade si possono trovare gran parte delle cellule della nucella compresse e degenerate (fig. 4).
Le due cellule della diade non appaiono quasi mai ugualmente vi- vaci, ma la superiore ha il citoplasma pitl denso ed il nucleo molto grande picnotico, non mai tanto per6 da non mostrare abbastanza chiaramente lo stadio di profase (figg. 5, 6). La cellula inferiore della diade i: rnolto
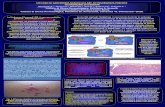
Fig. 1: 1-8 Maggio, ovulo con nucella nuda (x57). - Figg. 2 e 3: 1-8 Maggio, nucella con cellula rmdre delle megaswre ( x 800). -Fig. 4: 8 Maggio, diade ( x 800). -Fig. 5 a, b: 9 Maggio, nucelk contenente w diade con i nuclei delle due cellule in profase ( x 800). - Fig. 6: 9 Maggio, Cellula superim delh diade uniadeata e cellula inferiore binudeata (xs00). - Fig. 7: 9 Maggio, diade con cellula Mriore bioucleata ( ~ 8 0 0 ) . - Fig. 8: 8 Maggio, gametJto bioucleato non gennogliato ( ~ 8 0 0 ) . - Figg. 9 e 10: 1-8 Maggio, gametofiti bmucleati, germoglhi (~800). - Pig. 11: 8 Maggio, gametofito binucleato germogliante,caratteristicamente contorto nella nucelh degenerante (xs00). - Fig. 12: 18 Maggio, gVrmoQt0 binucleato poluiuato, in K-
cre&nento (XSOO). - Fig. 13: 18 Maggio, gxnetcrfito binudedlo sgusciilto fuori cialla nucclla, am i due nuclei Calvali ( ~ 8 0 0 ) . - Fig. 14: 18 Maggio, gametofito tetranucleato (XSOO). - Fig. 15: 18 Maggio, gme- tofilo tetranucleato, con nuclei micropilari picnotici ( ~ 8 0 0 ) . - Fig. 16: 7 Mageio, parete dell’antera e tetradi di microspore ( x 800).
Dow
nloa
ded
by [
Yor
k U
nive
rsity
Lib
rari
es]
at 1
8:42
19
Oct
ober
201
4

Dow
nloa
ded
by [
Yor
k U
nive
rsity
Lib
rari
es]
at 1
8:42
19
Oct
ober
201
4

154 ME SSER I
pili sviluppata in grandezza; io l’ho riscontrata sia con un solo nucleo grande in profase e il citoplasma un poco vacuolizzato (figg. 4,5), sia, piti spesso; con i due nuclei ravvicinati fra Ioro, nel centro della cellula (figg. 6,7, 8). La cellula superiore e quella inferiorc della diade general- mente non sono allineate l’una sull’altra, ma leggermente spostate la- teralmente l’una rispetto all’altra. Quando la cellula inferiore della diade ha gi2 diviso il suo nucleo, essa t pih marcatamente vacuojizzata special- mente nella sua parte superiore, in maniera che i nuclei appaiono spo- stati verso la regione calazalc della cellula.
Successivamente il gametofito germoglia, vacualizzandosi (fig. 9) e polarizzandosi ( f igg . 10, 11, 12). I1 gametofito binucleato L in un primo tempo contenuto dentro la nucella degenerante, ma presto sfonda le cellule della nucella e si fa strada nel canale micropilare (fig. 13) (cfr. PI- ROTTA e DE PERGOLA, 1913 a). L’ovulo nel frattempo non I: cresciuto di dimensioni; soltanto il gametofito binucleato si t enormemente accre- sciuto nella sua porzione .citoplasmatica micropilare, mentre i nuclei rimangono nell’ambito del citoplasma contenuto ancora nella nucella, o appena a1 di fuori.
Lo sgusciamcnto fuori della nucella mi sembra che avvenga un PO’
lateralmente a1 tetto di essa, perchi qualche volta ho potuto osservare che la cellula superiore della diade, che ha la sua normale sede sotto il tetto della nucella, era, a questo stadio, ancora perfettamente riconoscibile nclla sua sede, sotto le cellule della volta della nucella che mantencvano i consueti rapporti le une con le altre, mentre la nucella appariva rotta lateralmente ed un lungo pseudopodio del gametofito si prolungava attra- verso la rottura fino nella regione del micropilo. Raramente ho osservato nucelle con archesForio bicellulare dal quale si sono sviluppate due diadi, le cui cellule inferiori sono binucleate I1 gametofito binu- cleato germogliato si’ pub presentare con due aspetti e ciot o normal- mente polarizzato con un nucleo calazale ancora contenuto nel citoplasma endonucellare e uno micropilare, che t migrato in alto, al’ micropilo, nello pseudopodio citoplasmatico, che ha rotto la nucella, oppure non polarizzato, con ambedue i nuclei in posizione calazale. Dato che nei molti gametofiti maturi osservati l’apparato ovarico si trova sempre in posizione normalmente micropilare, penso che i gametofiti binucleati, che non si polarizzano, siano destinati a degenerare. Mi sono imbattuta in stadi di questo generc con i due nuclei normalmente vivaci (fig. 12) e con i nuclei picnotici (fig. 13).
Quindi per quanto riguarda il tip0 di costruzione del gametofito femminile, da tutto cib che ho sopra riportato, mi pare si possa con suffi- ciente sicurezza pronunziarsi per un tip0 di costruzione dimegasporiale.
Dow
nloa
ded
by [
Yor
k U
nive
rsity
Lib
rari
es]
at 1
8:42
19
Oct
ober
201
4

EMBRIOLOGIA ED EMBRIOGENESI DI OLEA EUROPAEA L. 155
Cib concorda con le osservazioni di ANDERSON (1931) su Olea europaea,. 0. chrysuphylla ed 0. capemis, mentre t noto che nelle altre Oleacee studiate prevale il tipo normale.
In Oka chrysophylla e 0. capensis sembra che la rottura della nu- cella e la fuoriuscita del gametofito non siano fenomeni ’cosi frequenti (ANDERSON 1931).
38 Settimana di Maggiu. - I fiori nel lor0 complesso non sono au- mentati di dimensioni. Essi si sono schiusi e le due grandi antere sono uscite dagli involucri. Le antere sono completamente mature, le sacche polliniche sono aperte. I1 tessuto transitorio b forrnato da grandi cellule a parete ispessita e le cellule del tappet0 sono state riassorbite o quasi; i granelli di polline sono maturi, rotondeggianti, con tre pori di germinazione, con esina ornata di una cuticola ondulata a cresta. Non si pub vedere a questo stadio se il polline b uni- o binucleato, perchb tutto il granello si colora fortemente di nero, senza lasciar vedere il contenuto. I1 pistillo ha uno stilo notevolmente pib lungo che nello stadio prece- dente. Le loggie ovariche contengono ciascuna due ovuli ugualmente sviluppati, circa delle dfmensioni di quelli dello stadio precedente.
A questo momento il gametofito si presenta o allo stadio binucleato (fig. 13) o a quello tetranucleato (figg. 14, 15); in ambedue.i casi perb i gametofiti hanno le medesime dimensioni, ciol: occupano tutto l’asse dell’ovulo, dalla sede della nucella, i cui resti sono sempre visibili, fino a1 micropilo, che in certi ovuli si presenta un PO’ divaricato.
In questo stadio il gametofito si trova a contatto della parete interna del tegumento, che presenta un tipico Mantelschicht, particolarmente accentuato nella regione equatoriale del sacco embrionale. Questo epi- telio interno t. stato dexritto da ANDERSON (1931) nel genere Olea e 1’A. sostiene che da questo strato dipende l’accrescimento intercalare dell’ovulo. Nel mio cam la differenziazione di questo tessuto appare nel suo aspetto pib tipico negli ovuli gih fecondati, ai primi stadi di dif- ferenziazione dell’albume, ed t a questo proposito che ne darb una de- scrizione piG dettagliata.
11 germogliamento del gametofito, e ciob il suo accrescimento soma- tico, si inizia all0 stadio di gametofito binucleato e c i d dal momento in cui si b compiuta la meiosi, dato il tip0 dimegasporiale di costruzione, e non sembra che una seconda crisi di accreximento importante in- tervenga fra lo stadiu binucleato ed il tetranucleato, perchb trovo i game- tofiti tetranucleati delle medesime dimensioni di quelli binucleati ger- rnogliati e polarizzati. Si possofio avere gametofiti tetranucleati con i due nuclei calazali vivaci e i due nuclei micropilari picnotici e con con- torno confuso (fig. 15), ma normalmente i gametufiti tetranucleati hanno
Dow
nloa
ded
by [
Yor
k U
nive
rsity
Lib
rari
es]
at 1
8:42
19
Oct
ober
201
4

-- 156 MESSERI
i 4 nuclei vivaci (fig. 14). Raramente si trovano dei fiori che hanno le antere normalmente sviluppate col polline maturo e nei quali il pistillo i: pi6 piccolo del normale; esso consta di un piccolo ovario con due log- gie e quattro piccolissimi ovuli, in alcuni dei quali si osserva una cellula pili grande nel centro; la nucella non b differenziata dal corpo dell’ovulo, 1’0vu10 non b ancora anatropo, cii, che complica l’osservazione. Le log- gie dell’ovario sono chiuse perchb lo stilo t gii formato e assai lungo; manca del tutto il tessuto recettore all’apice deilo stilo, che non 6 affatto slargato nelle caratteristiche papille stigmatiche.
Nei fiori normalmente sviluppati a questo stadio non sembra che il polline abbia ancora iniziato la germinazione sugli stigmi.
Fino a questo punto dell0 sviluppo le pareti dell’ovario sono formate da un tessuto omogeneo c non c’t nessun inizio di differenziazione di un endocarpo. 4 S e t t i n h a di Maggio. - I fiori di quest’epoca hanno gii comple-
tamcnte spollinato; in essi la corolla t appassita e con essa gli stami, che sono epicorollini. Tutto il pistilo non si L molto accresciuto dallo stadio precedentc, lo stilo si t allungato, lo stigma i piti differenziato dallo stilo, sotto forma di un corpo convesso apicale, ed i tutto pieno di granelli pollinici aderenti e germinanti, Si vedono i tubetti pollinici spor- gere dai pori germinativi. ‘ Gli ovuli sono un poco piti sviluppati rispetto a quelli che conte- nevano gametofiti all0 stadio binucleato e tetranucleato, tutti e quattro della medesima grandezza. Due di essi, uno per loggia, sono perfetta- mente anatropi, con l’asse calazo-micropilare perfettamente diritto e col micropilo beante proprio in posizione micropilare (fig. 1 7 4 . G h altri due sono leggermente campilotropi, con l’asse ovulare piegato un poco in fuori, cosiccht il niicropilo appare spostato, un poco lontano dalla base dello stigma (fig. 17 b). A questo stadio in tutti e quattro gli ovuli di un pistillo c’L il gametofito maturo.
A propsito della posiz’one che gli ovuli occupano nelle loggie ova- riche, ANDERSON (1931) pubblica una figura di una sezione longitudinale di un ovario di OIea europaea, con due ovuli, dei quali uno perfettamente anatropo, con l’asse corrispondente a quello della loggia, e l’altro col mi- cropilo spostato rispetto a1 prinio e col cavo del sacco embrionale un PO’ curvo, proprio come ho trovato io nel mio materiale.
Lo spazio che occupa il gametofito maturo in un ovulo non i molto superiore a quello occupato dal gametofito binucleato germogliato e da quello tetranucleato. Non sono mai riuscita a osservare le antipodi an- Cora vivaci che devono essere effimere, cosiccht non posso stabilire con esattezza il tipo di sviluppo del gametofito; in qualche cam per6 ho po- tuto ricostruire alla calaza i resti di tre cellule.
9
Dow
nloa
ded
by [
Yor
k U
nive
rsity
Lib
rari
es]
at 1
8:42
19
Oct
ober
201
4

Dow
nloa
ded
by [
Yor
k U
nive
rsity
Lib
rari
es]
at 1
8:42
19
Oct
ober
201
4

Si tratterebbc quindi del tipo Scilla, chc del resto c’l: in altre Olcacee (cfr. ANDERSON 1931, e BERCOVICH per PhilIyrea (I)) . Le sinergidi sono caratteristicamente appuntitc, molto spcsso cosi fortemente colorabili all’e- matossilina ferrica, che appaiono come una massa nera (fig. 18 a, 18 b, 20 Q 21 b) ; in materiale fissato in Delaunay csse appaiono meno deformate e meno fortementc colorate (figg. 26, 27, 29a). Presentano un grande vacuolo nella parte infcriore del loro corpo, regolarissimo; a1 margine superiore di questo vacuolo si affaccia i l nuclco, piuttosto piccolo, con un nucleolo. La partc superiore della ccllula, caratteristicamente appuntita, b occupata da un protoplasma denso, perforato da piccoli vacuoli che formano una rete densa; pih che come \._acuoli, essi appaiono come corpi figurati vacuolizzati. Quest0 speciale apparato vacuolare tiene il posto dell’apparato filamentoso che sta appunto nella porzione pili micropilare di molte sinergidi. La cellula uovo si trova sotto una sinergide (fig. 27) spostata molto verso il micropilo; essa quasi si impegna nel micropilo (figg. 18a, 32 c). H o dctto gii che fino dallo stadio binucleato germogliato il citoplasma si insinua come un tappo nel micropilo preparando cosi la sede per la ccllula uovo. La cellula uovo b molto diafana in confront0 alle sinergidi, pii! o meno tipicamcntc piriforme (figg. 25, 28, 29 b), con un grande vacuolo verso il micropilo ed il nucleo spostato verso la calaza (figg. 21 a, 29b, 3 2 ~ ) ; Oltrc a cellulc uovo con questa fisionomia nor- male, ne ho trovate di atipichc sia nella vacuolizzazione, in quanto o non sono vacuolizzate o hanno vacuolo calazale, sia nelle dimensioni, che al- cune volte sono minuscole (figg. 20b, 22). Prevale pert, l’aspetto nor- male (figg. 25, 27).
I nuclei polari sono scmpre vivaci, grandi, mai fusi, sempre ravvi- cinati fra loro e giacciono circa a meta della lunghezza del gametofito, un po’ spostati verso la ragione calazale (figg. 24, 25, 33).
In diversi casi mi pare di avere osservato la fecondazionc in atto. Cosi nella fig. 23 ho riprodotto una cellula uovo a citoplasma denso, senza la caratteristica vacuolizzazione, alla quale aderisce una struttura che interpreterei come un tubetto pollico.
In altri casi una delle sinergidi giace sopra un corpo che put, essere interpretato come il tubetto pollinico (figg. 21 b, 22, 31); i vacuoli delle sinergidi sono ripieni di corpi irregolari molto colorabili in ematossi- lina ferrica, la cellula uovo non senibra ancora fecondata, per la sua ca- ratteristica vacuoliwzione (fig. 22). Non ho mai trovato gametofiti in cui i nuclei polari fossero fusi; essi sono semprc ravvicinati e nella posi- zione che ho descritto sopra (figg. 24, 33). In questo stadio spesso il -_
(I) Dalla tesi di laurea, intdita (Univ. di Pisa, 25-XJ939).
Dow
nloa
ded
by [
Yor
k U
nive
rsity
Lib
rari
es]
at 1
8:42
19
Oct
ober
201
4

EMBRIOLOGIA ED EMBRIOGENESI DI OLEA EUROPAEA L. 159
canale micropilare si h molto raccorciato e l’ovulo t divenuto quasi beante per la formazione di on grande micropilo. Le sinergidi e specialmente la cellula uovo si affacciano a questa apertura (figg. 25, 30). Impegnato nel micropilo c’k anche il citoplasma della cellula proendospermatica. Alle volte invece il canale micropilare si mantiene allungato (fig. 32 a, b, c).
Nei numerosi pistilli che ho esaminato in questo stadio di sviluppo, che dura circa dal 25 a1 29 di Maggio, non ho mai riscontrato chc fra i quattro ovuli di un pistillo ce ne fosse uno preferito. Sono circa tutti delle medesime dimensioni e si presentano nel medesimo stadio di sviluppo. L’antagonismo fra gli ovuli per il raggiungimento della monospermia t un fenomeno certamente posteriore all0 stadio di niaturith del gameto- fit0 femminile. E non c’t niente che lasci supporre, all’atto della feconda- zione, la preferenza di un solo ovulo di un pistillo.
Nei pochi fiori che a questo stadio hanno ancora le antere in posto, queste sono giA aperte; gran parte del polline b uscito, quello ancora in posto, completamente maturo, presenta le ernie dell’entina, che spor- gono attraverso ai tre pori germinativi. Le estroflessioni dell’entina fanno capo, nell’interno del granello, a tre semilune chiare.
A questo stadio cominciano a comparire nelle pareti dell’ovario delle cellule sclerenchimatiche sparse che preludono alla formazione dell’en- docarpo legnoso. Esse compaiono nella parte profonda della parete del- l’ovario, internamente alla zona dove corrono i fasci conduttori. Sono scarsissime, distanziate e poco progredite nella differenziazione. La lor0 parete I: ispessita e poligonale ed il protoplasma degenerante L ancora visibile all‘interno di esse.
Sono assai rari i fiori con gametofito maturo che non presentano pol- line germinante sullo stigma.
Dalla la settimana di Giugno alla l a settimana di Agosto. - In mate- riale del 5 giugno la fecondazione h avvenuta e l’ovulo preferito t giA notevolmente cresciuto a carico degli altri tre. I tre ovuli regrediti sono di dimensioni diverse fra loro. I n quelli meno compressi si osserva an- Cora il cavo del sacco embrionale, con i resti del gametofito (fig. 36). In alcuni questi resti sono rappresentati soltanto da una massa citopla- smatica, nella quale non si distinguono piti i nuclei o comunque corpi figurati, in altri invece si distinguono ancora bene l’apparato ovarico ed i due nuclei polari ravvicinati, ma non fusi. Non ho mai trovato i nuclei polari fusi negli ovuli non preferiti; essi si trovano all0 stadio in cui li ho descritti alla fine di maggio, o comunque sono degenerati in quell0 stadio. Sembrerebbe ci& che essi non fossero stati fecondati.
ANDERSON (1931) dice a questo proposito che si ha ~robabdmente in Olea la fecondazione di un solo ovulo, mentre tutti e quattro arrivano
Dow
nloa
ded
by [
Yor
k U
nive
rsity
Lib
rari
es]
at 1
8:42
19
Oct
ober
201
4

160 MESSERI
a completo sviluppo e sostiene che anche in Fraxinus, contrariamente a quanto dice SOMMER (in ANDERSON 1931), la monospermia non di- pende dall’aborto piti o meno precoce di tre ovuli su quattro, ma dalla mancata fecondazione di tre su quattro ovuli, tutti ugualmente pronti, all0 stadio maturo. Le mie osservazioni confermano pienamente che tutti e quattro gli ovuli di un pistillo arrivano all0 stadio maturo, ma non ho purtroppo dati sufficienti per pronunziarmi in quanti dei quattro ovuli avvenga la fecondazione.
L’ovulo preferito presenta la cellula uovo fecondata, trasformata in uno zigoto allungato a forma di pelo, che giace proprio nella regione del micropilo (figg. 34 a, 35 a), ed un abbondante albume cellulare (figg. 34 a,
Le cellule dell’albume sono, fino dagli stadi piti precoci, eterogenee, fatto che si niette bene in evidenza con fissazione in Delaunay (fig. 37 a, b). In particolare quelle che circondano lo zigoto hanno citoplasma denso con caratteristiche goccioline sferiche sospese e nuclei grandi a forma ame- b i d e , particolarniente colorabili. Le cellule lontane, in particolare quelle calazali, hanno grandi dimensioni, citoplasma abbondantemente vacuo- lizzato, privo o pverissimo di quelle goccie che caratterizzano le cellule micropilari, e nuclei piti piccoli e pid pallidi (figg. 37 a , b, e 38 a, b).
Gli stadi intcrmedi fra quelli descritti alla fine di maggio e i presenti, ciot fra la maturith, la fecondazione e lo sviluppo dell’albunie nell’ovulo preferito, devono essere fugacissimi, dato che non ho mai potuto trovare niente di intermedio che mi aiutasse a comprendere il meccanismo che regola lo sviluppo dell‘ovulo preferito a danno degli altri tre. Non ho mai trovato un caso di due ovuli preferiti. Sembra assai probabile per6 che nei gametofiti degli ovuli destinati a regredire non si arrivi mai alla fecondazione data la costanza con cui trovo, nei resti di essi, i due nu- clei polari ravvicinati e non fusi, uguali fra lor0 nelle dimensioni e nel numero dei nucleoli, tanto da non poter far supporre che uno solo dei nuclei sia stato fecondato. L’albume sembra esere cellulare fino dai suoi primi stadi di sviluppo ed il suo metabolismo in confront0 a quello del gametofito prima della fecondazione si manifesta sempre con la com- parsa delle goccioline. l?i evidente anche fino dai primi stadi del suo svi- luppo un dimorfismo fra albume micropilare che si sviluppa nella sfera embrionale e quello calazale. Le goccioline sono caratteristiche dell’al- bume micropilare, il quale si introflette nel micropilo a circondare anche a1 di sopra lo zigoto (fig. 38a).
L‘albume di Olea europaea t quindi assai diverso da quello osservato in 0. capensis (ANDERSON 1931), con cellule tetranucleate diverse di forma nelle due zone superiore ed inferiore del gametofito.
b, 35a, b, c, 37 a, b, 38 a, b, 39).
Dow
nloa
ded
by [
Yor
k U
nive
rsity
Lib
rari
es]
at 1
8:42
19
Oct
ober
201
4

EMBRIOLOGIA ED EMBRIOGENESI DI OLEA EUROPAEA L. 161 __ .----
35a i j I
I
!
I I
I I !
Fig. 34 a, b: 5 Giugno, a zigoto germogliante (z), ciroondaro da UM celluh dell'dbume, e ccllulc dcll'e- pitelio dcl tegurncnco; b cellulc dcll'albume a circa a mctl della uvich del s1cm embrionale c cellule del te- gummto (XSOO). - Fig. 35 a . b , c: 5 Giugno, zigoto filifcrrne (z) e ccllule dell'albume: a, a contatto con lo zi- goto; b, c ira a mtta &lL o v i t l dcll'ovulo; c, nelh regione o l a u l e ( ~ 8 0 0 ) . - Fig. 36: 8 Giugno, due ovuli della medwitna loggia, con prevalenu di un ovulo aull'alrro ( ~ 5 7 ) .
Dow
nloa
ded
by [
Yor
k U
nive
rsity
Lib
rari
es]
at 1
8:42
19
Oct
ober
201
4

162 MESSERI
Questo stadio si protrae per tutta la prima metl del mese di Giugno. In ovari fissati in questo period0 infatti troviamo sempre lo zigoto sotto forma di un’unica cellula allungata come un pelo, che si insinua fra le cellule dell’albume, col suo unico grande nucleo in posizione apicale ed un grande vacuolo, che occupa quasi tutto lo spazio retro- stante. Esso pub essere lungo fino ad attraversare quasi tutto il micro- pilo (fig. 38a) . Una volta sola in materiale del 17 Giugno, ho osservato un embrione sempre a forma di pelo, ma pluricellulare, sviluppato in lunghezza fino a raggiungere il cavo del gametofito. La sua moltiplica- zione fino alla formazione di un cordone pluricellulare, i posteriore a1 suo allungamento come cellula unica. Quando lo zigoto ha raggiunto questa posizione, esso si lascia dietro tutta la regione del canale micro- pilare, occupata da cellule dell’albume, che, come avevo gia detto, si erano spinte fin 18 a fasciare lo zigoto (fig. 39). Queste cellule gia supe- rate dallo Zigoto appaiono degeneranti, picnotiche, ben diverse da quelle che si trovano nella regione apicale dello zigoto. Evidentemente queste cellule micropilari hanno gia assolto il loro compito di dirigere ed ali- mentare l’accrcximento dello zigoto fino a raggiungere la regione del- l’albume, nella quale si dovri piG tardi sviluppare l’embrione.
A questo stadio l’albume e lo zigoto, come gia il gametofito dallo stadio binucleato germogliato in poi, si trovano a diretto contatto.con le cellule della parete interna dell’unico tegumento. Restano ancora visibili alla cataza i resti delle cellule della nucella, attraverso alle quali sguscip fuori il gametofito binucleato. Le cellule della parete interna del tegumento hanno l‘aspetto di un epitelio secretore, assai diverso nelle varie regioni dell’ovulo. Esse sono appiattite, a xarsa vitahi, spew semi-riassorbite nella regione micropilare dove arriva lo zigoto col suo accrescimento apicale (fig. 34 a), mentre sono allungate radialmente a citoplasma molto denso, nucleo grande, cariche di inclusi, nella regione circa equatoriale dell‘ovulo, occupata dalle cellule dell’albume a maggiori dimensioni (fig. 34 b). L’ovulo evidentemente cresce di mole per accre- scimento intercalare soprattutto di questa regione (cfr. ANDERSON 1931).
La differenziazione dell’endocarpo, sotto forma di cellule pietrose nel parenchima dell‘ovario interno alla corona dei fasci conduttori, non k progredita in confronto a quanto si osservava alla fine di Maggio. Si tratta sempre di rare cellule pietrose. I1 parenchima di questa regione k scompaginato in confronto a quanto si osserva allo stadio di gametofito maturo. Anche ANDERSON (1931) ha osservato che l’endocarpo si forma dopo l’antesi.
Lo stadio di proembrione allungato, piliforme, t in generale molto lungo. I primi stadi di sviluppo dell’embrione vero e proprio li ho tro-
Dow
nloa
ded
by [
Yor
k U
nive
rsity
Lib
rari
es]
at 1
8:42
19
Oct
ober
201
4

EMBRIOLOGIA ED EMBRIOGENESI DI OLEA EUROPAEA L. 163
Fig. 37 a, b: 8 Giugno, zigoto unictllulut piliforrne (L) CiKw.da10 da cellule dcll'albumr: a -to alto Zi- 8010, b nel ccntm c allz alu (xe00). - Fig. 38 a, b: 10 Luglio, a ziuoto unioclluLn allulylto (2). cilmpduo da celluk dcll'albume, b album nellr regionc m c d h dell'ovulo (xs00). - Fig. 39: 17 G i w . mrpo embrio- rule pluricdluhre, Blifonm. circhdrto dall'albume dimotfo (x%oo).
Dow
nloa
ded
by [
Yor
k U
nive
rsity
Lib
rari
es]
at 1
8:42
19
Oct
ober
201
4

164 MESSERI
vati alla fine di giugno (verso il 28), ma con una certa frequenza soltanto nel luglio e perfino in agosto. L'embrione si forma per moltiplicazionc della cellula apicale del proembrione, quando questo ha raggiunto una determinata posizione, sempre uguale in tutti i casi osservati, e precisa- mente subito sotto la base del canale micropilare, in posizione leg- germente laterale a questo. Non ho mai trovato una testa embrionale in posizione assiale rispetto a1 micropilo; quando lo zigoto esce dal collo
Fig. 400, b: 28 Giugno, inizio della differenziarionc dell'cmbrimc (~800).
micropilare ~ S S O cresce lateralmente. Gli stanno aderenti grandissime cellule dell'albume, il cui nucleo appare spesso in profase.
Le prime moltiplicazioni portano alla formazione di una specie di morula, in cui non si distingue un dermatogeno da un territorio interno e che ha una specie di sospensore che si insinua dentro il micropilo fatto da una serie di. cellule con le medesime caratteristiche di quelle della morula (figg. 40 a, b). L'embrione si mantiene sotto forma di morula fino ai primi di Agosto, c i d per piii di un mese di tempo.
CONCLUSIONI
Le osservazioni riportate nel testo mi permettono di trarre le se- guenti conclusioni.
Gli ovari di olivo sono biloculari e ciascuna loggia t fornita di duc ovuli ugualmente sviluppati ed orientati (ovuli anatropi), dall'inizio della loro differenziazione nel carpello fino all0 stadio di gametofito maturo e all'atto della fecondazione. Ciascun ovulo t formato da un grande unico
Dow
nloa
ded
by [
Yor
k U
nive
rsity
Lib
rari
es]
at 1
8:42
19
Oct
ober
201
4

EMBRIOLOGIA ED EMBRIOGENESI DI OLEA EUROPAEA L. 165
tegumento, che racchiude nel suo interno una piccola nucella. Una si- mile costituzione dell’ovulo t caratteristica di tutto l’ordine delle Ligu- strales, di cui 1’0livo fa parte.
Nella prima settimana di maggio (nell’olivo che ho seguito) si ha la inegasporogenesi.
La divisione riduzionale della cellula madre deve esere fuggevo- lissima. La diade I: racchiusa in una nucella formata di un solo strato di cellule. Dalla cellula inferiore della diade si sviluppa direttamente il gametofito binucleato, che t quindi di costruzione dimegasporiale. L‘ac- crescimento somatic0 pi6 notevole si ha allo stadio di gametofito binu- cleato. Questo infatti sfonda la nucella, i cui resti restano a lungo a fa- sciare la porzione calazale del gametofito, e si spinge nel canale micro- pilare, divaricandolo fino a formare la cavita del sacco embrionale, che si mantiene circa delle medesime dimensioni fino alla fecondazione. Anche gli ovuli non crescono molto dallo stadio di diade a quello di game- tofito maturo. Sono state osscrvate delle anomalie nella polarizzazionc del gametofito binucleato. Dal gametofito binucleato si passa a1 tetra- nucleato, il quale conserva circa le medesime dimensioni del binucleato germogliato, ma manifesta con maggiore evidenza la differenza di co- stituzione del citoplasma e dei nuclei delle due porzioni micropilare c. calazale del gametofito, che giA si notavano nel gametofito binucleato. I1 citoplasma micropilare appare assai piti denso del calazale ed i nuclei cosi carichi di colore e cosi difficili a differenziarsi, che sembrano spesso picnotici. Questo stadio i l’unico rappresentato nel materiale fissato a meta di Maggio.
L’ultima divisione porta a1 gametofito maturo circa nell’ultima set- timana di Maggio. Nel gametofito maturo ho potuto osservare una volta sola i resti di tre antipodi; l’apparato ovarico invece i normalmente vi- vace in tutti e quattro gli ovuli di ciascun ovario, formato da sinergidi con uno speciale apparato micropilare, vacuolizzato, e da una cellula uovo di forma normale.
La fecondazione avviene, nel mio olivo, nell’ultima settimana di Maggio. Ho potuto vedcre in atto l’entrata del budello pollinico in un certo numero di gametofiti e probabilmente la fusione del nucleo del- l’oosfera con un nucleo del tubetto pollinico, ma non ho mai osservato la fecondazione del nucleo secondario, anzi non sono mai riuscita ad osser- vare la fusione dei due nuclei polari che si mostrano sempre ravvicinati, ma non fusi, nella parte equatoriale del gametofito.
Nella prima settimana di Giugno si i gii stabilita una netta diffe- renza fra i quattro ovuli dell’oyario. Uno t molto cresciuto e presenta lo zigoto filiforme, unicellulare, che si insinua fra le cellule dcll‘albume,
Dow
nloa
ded
by [
Yor
k U
nive
rsity
Lib
rari
es]
at 1
8:42
19
Oct
ober
201
4

166 MESSERI
giP abbondantemente divise; gli altri tre sono gii compressi dal primo e presentano resti degeneranti dell’apparato ovarico, dove non t possi- bile distinguere le cellule l’una dall’altra, e i due nuclei polari non fusi, ancora delle dimensioni ed aspetto che avevano nel gametofito maturo, probabilmente non fecondati. Nell’ovulo preferito, invece, l’albome non solo t gih abbondantemente diviso, ma presenta un caratteristico di- morfismo, con cellule a citoplasma denso, ripieno di goccioline, nella regione micropilare, alle quali fanno rixontro cellule a citoplasma po- vero e vacuolizzato nella regione calazale.
Lo zigoto si mantiene indiviso, crescendo in lunghezza dentro il micropilo, come un pelo, per tutta la prima met2 di Giugno ed anche oltre, dopodicht si divide per dar luogo ad un corpo proembrionale, pluricellulare, filiforme, che si insinua sempre pili profondamente fra le cellule dell’albume. Nel fraitempo le cellule dell’albume, che il corpo proembrionale crescendo si laxia dietro verso l’apertura -micropilare, degenerano. Esso si porta cosi alla zona in cui il canale micropilare si apre internamente nella cavitP del sacco, gii completamente ripieno di cellule dell’albume. Li, in una regione caratteristica, e ciot aderente alle pareti della cavith, lungo le quali il corpo proembrionale striscia nel suo accrescimento, si forma il primo abbozzo embrionale. I1 pili precoce da me trovato t del 18 Giugno, il pili tardivo del 7 agosto. Questo abbozzo 2 formato da un corpo pluricellulare subsferico. La differenziazione del- l’embrione non t ancora cominciata nella prima decade di agosto.
A questo proposito desidero ricordare che anche in Probascidea Jussieui e P. Zutea lo zigoto si allunga fino a portare la sua testa in una zona caratteristica del sacco embrionale e solo dopo avere raggiunta que- sta sede, lo zigoto da orig‘ine all’embrione (MARTINI, 1939).
EIBLIOGRAFIA
ANDERSON A., 1931. - Studien iiber die Embryologie der Familien Celastraceae, Olea- ceae w d Apocynaceae. Lunds Universitets Arsskrift. N. F. Avd. 2. Bd. 27. Nr. 7. Kungl. Fysiografiska Sallskapets Handligar. N. F. 42. Nr. 7 (pp. 41-74) - 1-112.
CAMPBELL C., 1910. - Osservazioni e ricerche sull’Olivo chiamato u maschio )I. Bull.
-, 1912 a. - Sull’olivo a Dekkcir u del sud tunisino e salla impollinatione artificiale de& olivi praticata dagli arabi di certe oasi africane. Nuovo Giorn. Bot. It., n. s.,
-, 1912 b. - Un caso di partenocarpia nell’olivo. Nuovo Giorn. Bot. It., n. s., 19,
-, 1913. - Questioni e ricerche sulk biologla fiorale defl’olivo. Annali di Botanica,
SW. Bot., It., 5-12.
19, 73-85.
86-89.
11, 209-227.
Dow
nloa
ded
by [
Yor
k U
nive
rsity
Lib
rari
es]
at 1
8:42
19
Oct
ober
201
4

EMBRIOLOGIA ED EMBRIOGENESI DI OLEA EUROPAEA L. 167
Cisaap~e~~ C., 1921. - Studi sull'olivo in terra di Bari. Pubblicaz. delh s t a . Agraria Sperim. in Bari, 4, 1-54.
C ~ R R I R., 1942. - Reunti progressi dq l i studi botanico-ugrari sull'divo. Atti Con- vegno Studi olivicoli, 49-98.
FAVILLI R., 1943-45. - Rapporti fra la prescnza di semi vani e lo sviluppo delle drupe nell'divo. Annali Fac. Agr. Univ. di Pisa, n. s., 6, 1-14.
-, 1947. - Rictrchc sdl'aborto del seme nell'olivo. Annali Fac. Agr. Univ. di Pisa, n. s., 8, 118-125.
MARTINI E., 1939. - Riccrche embriologiche sulle Martyniaceae. Nuovo Giorn. Bot. It., n. s., 46, 197-226.
-mu A., 1947. - Osservazioni morfdogiche sulle Q olive passerine a. Nuovo Giorn. Bot. It., n. s., 54, 314315.
MORB~~NI A., 1939. - Ricerche sulla biologia fiorale dell'olivo. Nuovo Giorn. Bot. It., n. s., 46, 1-70.
-, 1940 a. - L'agente vettore del polline dell'olivo. Nuovo Giorn. Bot. It., n. s., 43, 264-270.
-, 1940 6. - Prim0 contributo allo studio della cascola dei $on e d a fnrtti dell'olivo. Atti R. Acc. Georg., ser. 6, 3, 33-52. - e BAGAOLI E., 1949. - L'antosterilit6 dell'olivastra Seggianese n del M. Amiata. Annali Speriment. Agraria, n. s., 7, 493-507.
PJROITA R., 1919. - Osservazioni s u l fiore dell'olivo. Atti R. Acc. dci Lincei, ser. V, 28, 20 sem., fasc. 1. - e DE hcou D., 1913, a. - Partenocarpia nell'olivo. Bull. Soc. Bot. It., 122-124. - -, 1913 b. - Sull'olivo maschio. Nota preventiva. Bull. Soc. Bot. It., !24-125.
TAYLOR H., 1945. - Cyfo-taxonomy and phylogeny of the Oleaceae. Brittonia, 5, 337-
VIGODSKI DE PHILIPPIS A,, 1937. - L'epoca della differenziazione delle gemme fiorali
-, 1938. - Ulteriori osservazioni sull'epoca di differenziazione delle gemme jiorali
.
365.
nell'olivo. Nuovo Giorn. Bot. It., n. s., 44, 484-487.
nell'divo. Nuovo Giorn. Bot. It., n. s., 45, 599-600.
SUMMARY
In this paper a first set of observations on the ernbriology and embriogeny of the Olive tree has been collected.
The material for this study comes all from a single specimen which grows in the Botanical Garden of Bari. Difficulties have been met in fixing, staining and cutting, therefore my observations are far from being complete. This researche is still going on.
The pystil of the Olive tree is bilocular with two ovules in each locule. Meiotic division must be fugitive as I have observed it only rarely. Diade stage is longer. The lower cell of the diade divides soon giving a binucleated cell, which develops into a dimegasporial binucleatcd gametophyte. The upper diade cell never divides, but degenerates rather late. The nucella has a very low position into the single in- tegument of theovule and it is very thin, formed by one layer of cells at its top and by two or three layers laterally.
The binucleated gametophyte breaks down the nuceltus and grows up filling the micropilar channel almost completely. At this stage the gametophyte almost reaches the size of the ripe stage. The binucleated gametophyte, through the tetra:
Dow
nloa
ded
by [
Yor
k U
nive
rsity
Lib
rari
es]
at 1
8:42
19
Oct
ober
201
4

- 168 MESSERI
nucleated stage, develops into the ripe gametophyte. The 4 ovules of each pystil are equally developed until1 the ripe stage, and keep about the size they had reached at the binucleated stage.
T h e stages just described take place about'from the first week of May (diade stage) to the last week of May (ripe gametophyte). It has never been possible to ob- serve the whole ripe gametophyte, the calazal part degenerating soon. So, we cannot establish the type of development. Probably it is of Scilla type.
The ovaric apparatus is formed by two sinergides and one egg cell of normal size and form. The two polar nuclei are allways closely approached.
Fertilization occurs in the last week of May. Pollen tubes have been observed in several gametophytes and it could seem very likely that fertilization of the egg might occur in all the four ovules of each ovarium.
The first week of June the ovules are already unequally grown in each ovary. Three of them remain of the size reached before fertilisation, while the fourth one has grown rapidly, crushing the other three against the walls of the ovary. In the grown up ovule we can observe the zygote in the micropilar region and the albu- men, formed by completely divided cells (cellular albumen), surrounding the ferti- lized egg, and prolonging into the embryosac cavity. T h e other three ovules show the rest of the ovaric apparatus where it is impossible to distinguish whether the egg has been fertilized or not and the two polar nuclei, which did not fuse, being only closely approached. It is very likely that they have not been fertilized, in fact they are both of the same size and neither of them shows two nucleoli, to make it possible to suppose that fertilisation took place before they fuse.
Owing to the embryo development the following observations have been made: The zygote, during the first half of June, is represented by one cell, thread-like, gro- wing amidst the albumen cells, with an apical nucleus. The albumen cells, amongst which the zygote grows, are different in form and content by those far from it. Their content is characterized by plenty of droplets scattered in the cytoplasm, which are lacking in the albumen cells far from the zygote. At the end of June the zygote divides into a thread body, which is formed by several cells disposed in one row, which runs through the albumen cells from the micropilar region inside, toward the embriosac cavity, already filled by the albumen tissue. The albumen cells of the micropilar region, in the meanwhile, degenerate, collapsing in a brownish mass. At the same time the apical cells of the proembryonic body divides into a roundish plu- ricellular body, which represents the first embryonic initial.
During the month of July, unicellular zygotes, threadlike pluricellular bodies without, or with the embrionic head already formed, may be found.
During the mounth of August the embrionic head has always formed, but the embryo is far from being built up.
EXPLANATION OF FIGURES
Fig. 1 : May 8th, ovule with nacke nucella ( x 57). - Fig. 2 and 3: May 8th, nu- cella and megaspore mother cell ( x 800). - Fig. 4: May 8th, diade ( x 800). - Fig. 5 a, b: May 9 th, nucella containing a diade, the two cells show the nucleus in prophase ( x 800). - Fig. 6: May 9th, diade, upper cell with one nucleus, lower one with two nuclei ( x 800). - Fig. 7: May 9th, diade, lower cell binucleated ( x 800). - Fig. 8: May 8th, binucleated gamethopyte, not yet developed ( x 800). - Fig. 9 and 10: May lrst-8th, developing binucleated gametophytes ( x 800). - Fig. 11:
Dow
nloa
ded
by [
Yor
k U
nive
rsity
Lib
rari
es]
at 1
8:42
19
Oct
ober
201
4

EMBRIOLOGIA ED EMBRIOGENESI DI OLEA EUROPAEA L. 169
May 8th, developing binucleated gametophyte, characteristically rolled inside the degenerating nucella ( X 800). - Fig. 12: May 18th, polarized binucleated gametophyte ( x 800). - Fig. 13: May 18th, binucleated gametophyte, escaped off the nucella with the two nuclei still in the calaza ( ~ 8 0 0 ) . - Fig. 14: May 18th, tetranucleated gametophyte ( x 800). - Fig. 15: May 18th, tetranucleated gametophyte with pic- notic micropilar nuclei ( x 800). - Fig. 16: May 7th, anther walls and pollente trads ( x 800). - Fig. 17 a, b: May 25th, longitudinal section of a pystil, with 4 ovules equally developed ( x 11). - Fig. 18 a, b: May 25th, synergids and egg cell of one of the ovule of the fig. 17b ( ~ 8 0 0 ) . - Fig. 19: May 25th, sinergid of the other ovule of the fig. 17b ( x 800). - Fig. 20 a, b.: May 25th, sinergids and egg cell of the ovule a of the fig. 17 a ( x 800). - Fig. 21 a, b: May 25, sinergids, egg cell and portion of the pollen tube of the ovule b of the fig. 17a ( x 800). - Fig. 22:-May 25th, sinergid, egg cell and pollen tube ( x 800). - Fig. 23: May 25th, egg cell adhering to'a pollen tube ( x 800). Fig. 24: May 25th, polar nuclei closely approached each other ( x 800). - Fig. 25: May 27th, ripe gametophyte (x310). - Fig. 26: May 27th, ovaric appa- ratus with sinergids. - Fig. 27: May 27th, ovaric apparatus ( ~ 8 0 0 ) . - Fig. 28: May 27th, egg' cell ( x 800). - Fig. 29a, b: May 27th, sinergids with vacuolar appa- ratus at their apices and egg cell ( x 800). - Fig. 30: May 27th, two ovules of the same locule with ripe gametophytes ( x 800). - Fig. 31 : May 29th, degenerating sinergid and pollen tube ( ~ 8 0 0 ) . - Fig. 32 a, b, c: May 28th, sinergids and egg cell, their position in relation to the micropile ( x 800). - Fig. 33: May 27th, polar nuclei closely near ( x 800). - Fig. 34 a, b: June 5th, a growing zygote, surrounded by an albumen cell, epithelium cells of the inner wall of the ovule cavity; b albumen cells about at the equator of the embrionic sac and cells of the integument of the ovule ( X 800). - Fig. 35 a, b, c: June Sth, thread like zygote (z) and albumen cells: a at the zygote level; b about at the equator of the ovule cavity, c in the calazal region (x800). - Fig. 36: June 8, two ovules of the same locule, one prevaling on the other ( x 57). Fig. 37 a, b: June 8, threadlike, unicellular zygote (2) surrounded by albumen cells: a at the zygote level, b at the equator and at the calaza ( ~ 8 0 0 ) . - Fig. 38 a, b: July 10, prolonged, unicellular zygote (z), surrounded by the albumen cells, b albumen cells in the median region of the ovule ( x 800). - Fig. 39: June 171h, pluricellular proembrionic body, threadlike, surrounded by the albumen cells which are dimor- phic ( x 800). - Fig. 40 a, b: June 28th, beginning of the differentiation of the em- bryo ( ~ 8 0 0 ) .
RIASSUNTO
L'A. studia l'embriologia e la embriogenesi d i Olea europaea L. inquadrandone le'varie fasi nel tempo. La costruzione del gametofito femminile b di tip0 dimega- sporiale, il gametofito maturo 6 probabilmente 8-nucleato. Tut t i e 4 gli ovuli d i u n pistillo raggiungono lo stadio maturo, e la concorrenza fra loro, che porta alla mono- spermia, si stabilisce soltanto a1 momento della fecondazione. Non si k potuto sta- bilire se un solo ovulo viene fecondato e quindi i: ancora oscuro il meccasnismo che porta alla monospermia. La formazione dell'embrione b preceduta da un lungo pe- ricdo (pic d i un mese) durante il quak lo zigoto i: quiescente, quindi si sviluppa in un corpo proembrionale. L'albume i: cellulare ed ha uno svilupp0 molt0 anticipato sull'embrione; esso i: spiccatarnente polimorfo nelle varie zone del sacco embrionale.
Dow
nloa
ded
by [
Yor
k U
nive
rsity
Lib
rari
es]
at 1
8:42
19
Oct
ober
201
4