AL DI FUORI DELLA CELLULA - giorgioventurini.net · tessuto e della diffusione dei nutrienti e...
31
1 AL DI FUORI DELLA CELLULA: •GLICOCALICE •MATRICE EXTRACELLULARE •LE ALTRE CELLULE A immediato contatto con la membrana plasmatica troviamo il GLICOCALICE : l’insieme delle componenti carboidratiche delle glicoproteine e dei glicolipidi, a cui si aggiunge materiale secreto dalle cellule. Il glicocalice si continua nella MATRICE EXTRACELLULARE, abbondante nei tessuti Connettivali.
Transcript of AL DI FUORI DELLA CELLULA - giorgioventurini.net · tessuto e della diffusione dei nutrienti e...

1
AL DI FUORI DELLA CELLULA:
•GLICOCALICE
•MATRICE EXTRACELLULARE
•LE ALTRE CELLULE
A immediato contatto con la membrana plasmatica troviamo il GLICOCALICE: l’insieme delle componenti
carboidratiche delle glicoproteine e dei glicolipidi, a cui si aggiunge materiale secreto dalle cellule.
Il glicocalice si continua nella MATRICE EXTRACELLULARE, abbondante nei tessuti
Connettivali.

2
Una cellula connettivale (cartilagine) in coltura insieme a globuli rossi: L’area priva di globuli rossi è occupata dalla matrice.
Le cellule connettivali producono una matrice extracellulare in cui restano immerse
TESSUTI CONNETTIVI: CELLULE IMMERSE IN ABBONDANTE SOSTANZA INTERCELLULARE
Posizione degli osteociti

3
Sezione di cartilagine:I condrociti sono contenuti nelle lacune cartilaginee, circondati dalla matrice che essi stessi secernono
I TESSUTI CONNETTIVI SONO FORMATI DA DIVERSI TIPI CELLULARI IMMERSI IN UNA SOSTANZA INTERCELLULARE,
COMPOSTA DA FIBRE E DA UNA COSIDDETTA MATRICE AMORFA.
I DIVERSI TESSUTI CONNETTIVI DIFFERISCONO PER I TIPI CELLULARI
PRESENTI E PER LE CARATTERISTICHE E LE QUANTITA’
DELLA SOSTANZA INTERCELLULARE PRESENTE

4
“LA MATRICE AMORFA” E’ FORMATA PRINCIPALMENTE DA PARTICOLARI
POLISACCARIDI, I GLICOSAMINOGLICANI (GAG), CHE POSSONO ASSOCIARSI A PROTEINE, PER
FORMARE I PROTEOGLICANI.
PROTEOGLICANI E GLICOSAMINOGLICANI INTERAGISCONO PER FORMARE COMPLESSI MOLECOLARI DI GRANDISSIME DIMENSIONI.
SONO INOLTRE PRESENTI SPECIFICHE GLICOPROTEINE, CON RUOLI FUNZIONALI DI
GRANDE IMPORTANZA
L’elevato numero di cariche negative (gruppi carbossile e gruppi solfato) permette alle molecole di attirare abbondanti cationi (soprattutto Na+)e quindi di attirare per osmosi e legare molte molecole di acqua, che è responsabile del turgore del tessuto e della diffusione dei nutrienti e dell’ossigeno
GLICOSAMINOGLICANI (GAG) = lunghe catene polisaccaridiche formate dalla ripetizione di dimeri
In genere, legati a proteine formano i proteoglicani.
-
--
A B
A B A B A B A B A B A B A B A B A B A B A B
MOLTI GAG SONO SOLFORATI

5
GLICOSAMINOGLICANI (GAG) = lunghe catene polisaccaridiche formate dalla ripetizione di dimeri
In genere, legati a proteine formano i proteoglicani
-
- -
GLICOSAMINOGLICANI
Dermatan-solfato
Acido Ialuronico
4 gruppi principali: Acido Ialuronico, Condroitin-Solfato e Dermatan-solfato, Cheratan-solfato, Eparan-solfato

6
Modello di proteoglicano: le catene GAG si legano covalentemente ad una catena polipeptidica
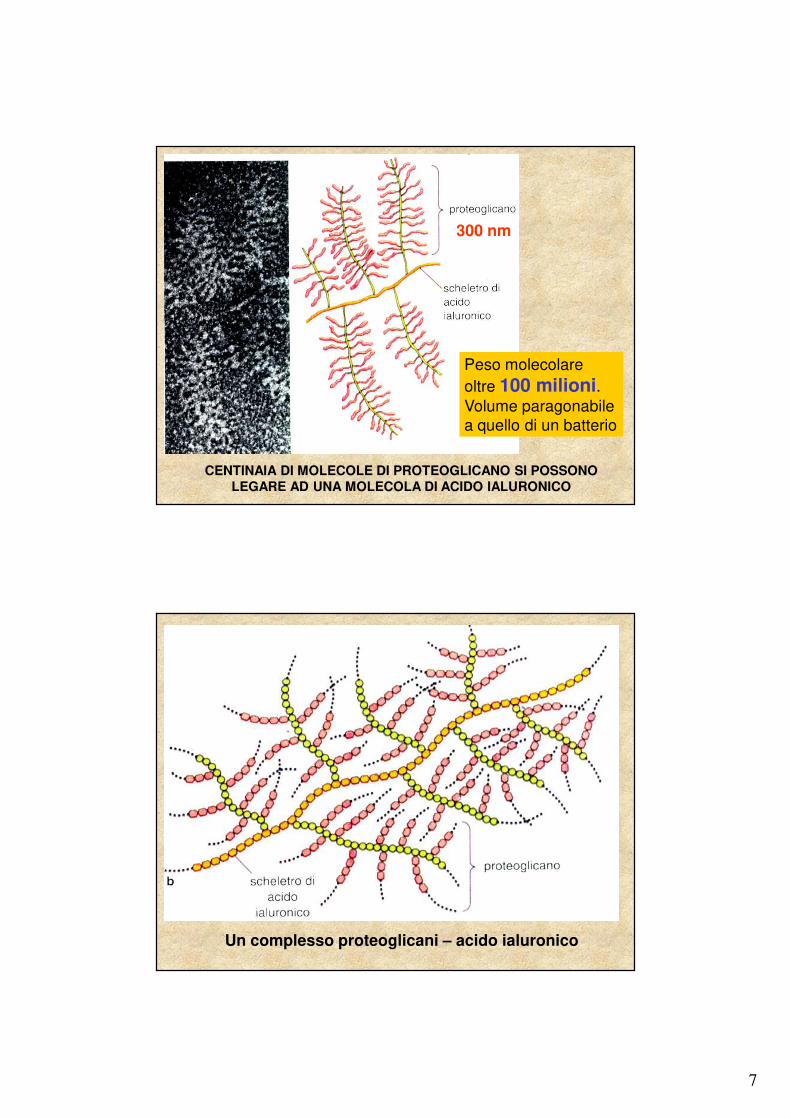
7
300 nm
Peso molecolare oltre 100 milioni.Volume paragonabile a quello di un batterio
CENTINAIA DI MOLECOLE DI PROTEOGLICANO SI POSSONO LEGARE AD UNA MOLECOLA DI ACIDO IALURONICO
Un complesso proteoglicani – acido ialuronico

8
Il grande numero di dimeri ripetuti e la scarsa flessibilità generano molecole di grande ingombro.
Le abbondanti cariche negative (COO- e SO3
-) richiamano cationi (soprattutto Na+) osmoticamente attivi che richiamano grandi quantità di acqua:
Il risultato è una pressione di rigonfiamento che genera il turgore e la resistenza alla compressione del tessuto
Batteri patogeni come lo stafilococco aureo secernono l’enzima ialuronidasi che, idrolizzando l’acido ialuronico, fluidifica i connettivi, facilitando la diffusione dei batteri.
L’acido ialuronico può anche essere presente in forma non associata a proteine, come polimero formato da migliaia di unità disaccaridiche ripetute. Ha proprietà lubrificanti e si riscontra in particolare nelle articolazioni

9
I proteoglicani regolano il traffico molecolare nel connettivo
Formando pori di dimensioni e densità di carica diversi possono funzionare come setacci molecolari (es. il proteoglicano perlecano, un eparan-solfato della lamina basale del glomerulo renale regola la filtrazione).
Proteoglicani possono legare proteine o altre molecole, determinandone una localizzazione precisa o concentrandole in una zona del tessuto.
LA COMPONENTE FIBROSA DELLA SOSTANZA INTERCELLULARE E’ FORMATA DA:
• FIBRE COLLAGENE E FIBRE RETICOLARI, COSTITUITE DALLA PROTEINA COLLAGENE
• FIBRE ELASTICHE, COSTITUITE DALLA PROTEINA ELASTINA

10
FIBRE COLLAGENE

11
L’ORGANIZZAZIONE DELLE FIBRE DI COLLAGENE
Tropocollagene (tripla elica)
La ricorrenza di residui di glicina ogni tre aminoacidi date le piccole dimensioni, permette di avvicinare le tre eliche per formare una struttura compatta
Il tropocollagene è formato da tre catene polipeptidiche di costituite da circa 300 ripetizioni della sequenza glicina-x-ydove x e y rappresentano in genere prolina e idrossiprolina. E’ anche comune la idrossilisina, cui si legano carboidrati. Le tre catene si avvolgono a formare una tripla elica

12
I gruppi OH dell’idrossiprolina e dell’idrossilisina e i residui di carboidrati sono importanti nella formazione di legami che permettono di formare le triple eliche e di assemblare le molecole di tropocollagene per formare le fibre.
Per idrossilare la prolina e la lisina è essenziale l’intervento dell’acido ascorbico (vitamina C)
LO SCORBUTO, grave malattia da carenza di vitamina C, porta alla impossibilità di formare fibre collagene stabili, a causa della mancata idrossilazione della prolina e della lisina
Gericault: Il naufragio del Medusa

13
“Alcuni persero le forze, e non potevano stare in piedi... Altri avevano la pelle cosparsa di macchie di sangue di un colore purpureo: ciò si estese poi alle loro caviglie, ai ginocchi, alle cosce, alle spalle, alle braccia e al collo. La loro bocca aveva un odore fetido e le loro gengive divennero così putrefatte da perdere brandelli di carne anche dalla radice dei denti, che pure cadevano” (1536).
La malattia può essere prevenuta o curata da una alimentazione ricca di vitamina C (acido ascorbico)

14
Le molecole di collagene si uniscono testa-coda, lasciando un breve interstizio. Tra una serie e quella parallela è presente uno sfalsamento di circa 70 nm (1/4 della lunghezza di una molecola). Di conseguenza il colorante che si accumula negli interstizi crea delle bande scure che ricorrono ogni 70 nm.

15
PdR: peptidi di registro o telopeptidi

16
COLLAGENI
Fibrillari: Tipo I (>80%), II, III, V, XIAssociati a fibrille: Tipo IX, XIIFormano reticoli: Tipo IV (Lamina basale), VII
I collageni associati a fibrille non formano fibrille e, legandosi ai collageni fibrillari, probabilmente mediano le interazioni con altre proteine della matrice.
I collageni associati a fibrille presentano tratti flessibili, dovuti a regioni in cui manca l’avvolgimento a tripla elica
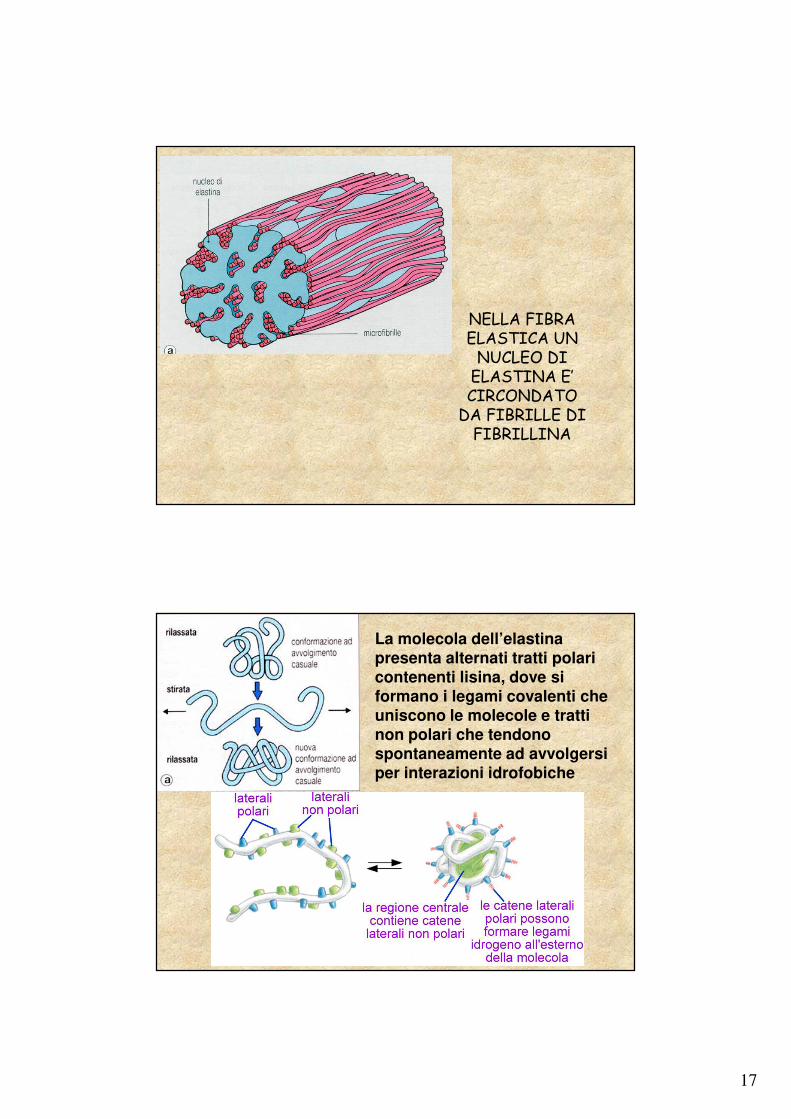
17
NELLA FIBRA ELASTICA UN
NUCLEO DI ELASTINA E’ CIRCONDATO
DA FIBRILLE DI FIBRILLINA
La molecola dell’elastina presenta alternati tratti polari contenenti lisina, dove si formano i legami covalenti che uniscono le molecole e tratti non polari che tendono spontaneamente ad avvolgersi per interazioni idrofobiche
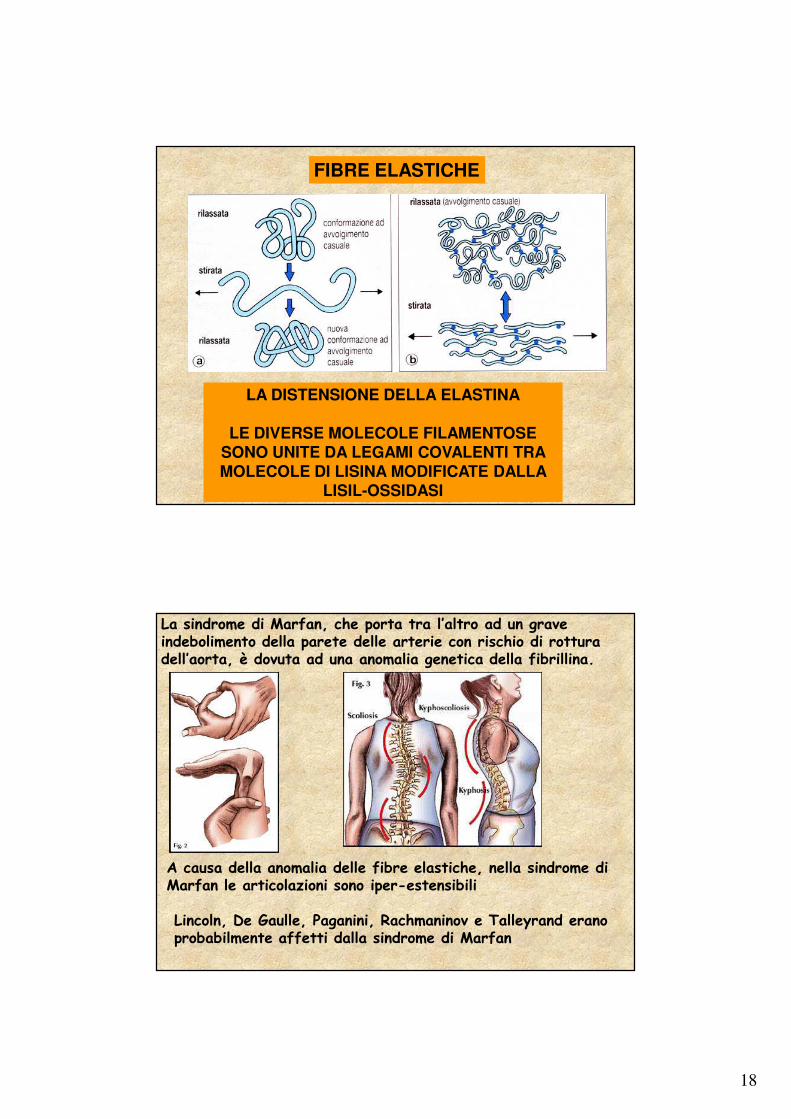
18
LA DISTENSIONE DELLA ELASTINA
LE DIVERSE MOLECOLE FILAMENTOSE SONO UNITE DA LEGAMI COVALENTI TRA MOLECOLE DI LISINA MODIFICATE DALLA
LISIL-OSSIDASI
FIBRE ELASTICHE
La sindrome di Marfan, che porta tra l’altro ad un grave indebolimento della parete delle arterie con rischio di rottura dell’aorta, è dovuta ad una anomalia genetica della fibrillina.
A causa della anomalia delle fibre elastiche, nella sindrome di Marfan le articolazioni sono iper-estensibili
Lincoln, De Gaulle, Paganini, Rachmaninov e Talleyrand erano probabilmente affetti dalla sindrome di Marfan

19
Nel connettivo troviamo •Fibroblasti (condrociti, osteociti)•Adipociti•Mastociti
•Cellule di derivazione emopoietica:
•Macrofagi•Globuli bianchi migranti dal sangue
I fibroblasti producono e secernono la matrice extracellulare

20
Immagine di un MASTOCITO al microscopio elettronico. I numerosi granuli scuri rappresentano vescicole secretorie ripiene di istamina e di eparina. La superficie cellulare, ricca di estroflessioni, testimonia la attività ameboide
SVILUPPO DI UN ADIPOCITO.I grassi, in nero nell’immagine, si accumulano all’inizio come piccole gocce, che poi si uniscono fino a formare una unica grande goccia lipidica che schiaccia in periferia nucleo e citoplasma
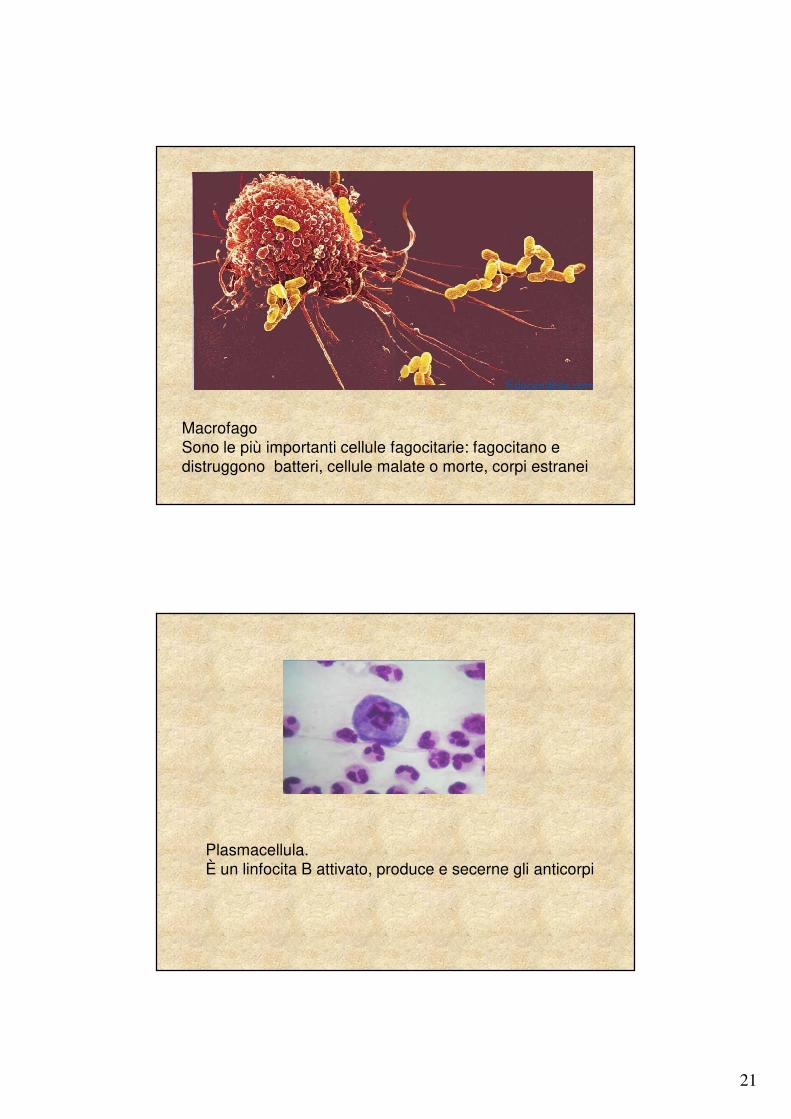
21
MacrofagoSono le più importanti cellule fagocitarie: fagocitano e distruggono batteri, cellule malate o morte, corpi estranei
Plasmacellula. È un linfocita B attivato, produce e secerne gli anticorpi

22
I complessi di proteoglicano –acido ialuronico si connettono alle fibre collagene creando una rete complessa, cui a loro volta possono legarsi le cellule
Le cellule si ancorano alla matrice extracellulare direttamente tramite i proteoglicani, le cui catene proteiche possono essere integrali di membrana o legate ai fosfolipidi con legami covalenti.
Le cellule possono anche legarsi a proteoglicani e collagene tramite specifici recettori o tramite proteine adesive come la fibronectina o la laminina

23
LEGAME A EPARINA FIBRINA
LEGAME A COLLAGENE
LEGAME A FIBRINA
LEGAME A CELLULE
LEGAME A EPARINA E ACIDO
IALURONICO
Nel connettivo sono presenti importanti GLICOPROTEINE, come ad esempio la FIBRONECTINA, capace di legare cellule, fibre connettivali e proteoglicani
LEGAME A FIBRINA
FIBRONECTINA

24
La fibronectina, collegando proteoglicani, fibre e cellule, regola le posizioni delle componenti del connettivo.Le cellule si legano grazie a specifici recettori: le INTEGRINE
Le integrine sono anche collegate con il citoscheletro (microfilamenti di actina). In questo modo l’ancoraggio delle cellule alla matrice risulta meccanicamente resistente.
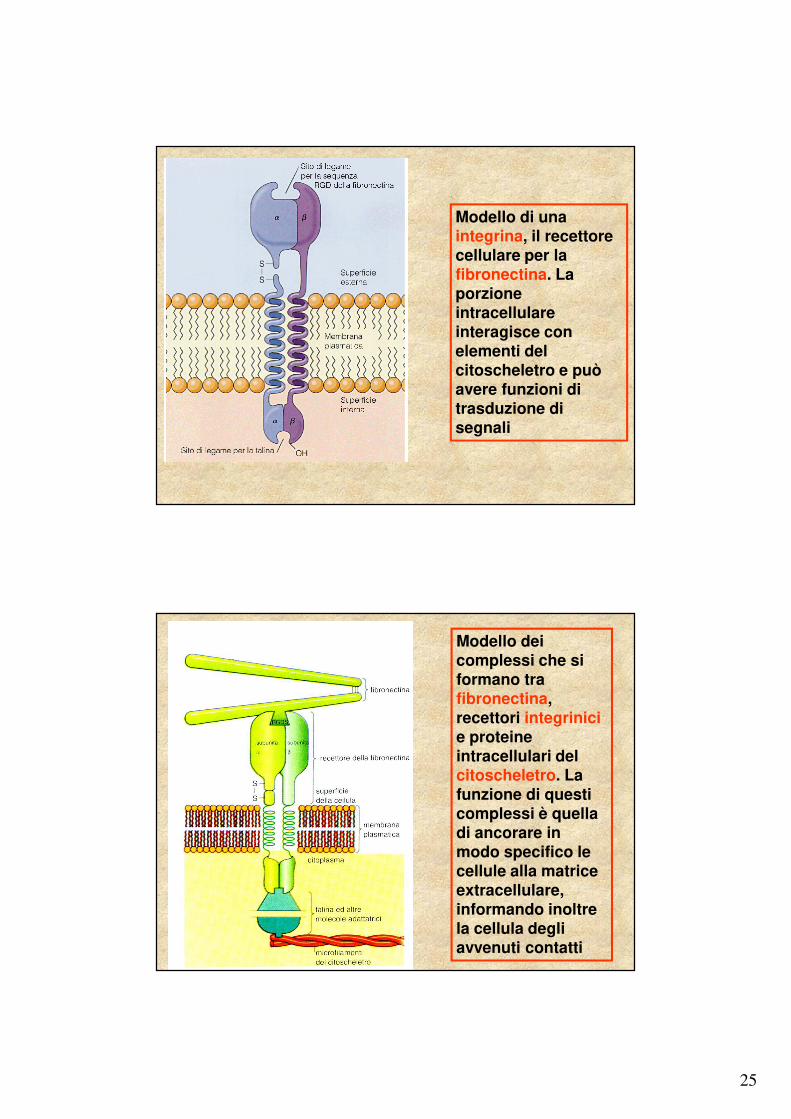
25
Modello di una integrina, il recettore cellulare per la fibronectina. La porzione intracellulare interagisce con elementi del citoscheletro e può avere funzioni di trasduzione di segnali
Modello dei complessi che si formano tra fibronectina, recettori integrinicie proteine intracellulari del citoscheletro. La funzione di questi complessi è quella di ancorare in modo specifico le cellule alla matrice extracellulare, informando inoltre la cellula degli avvenuti contatti
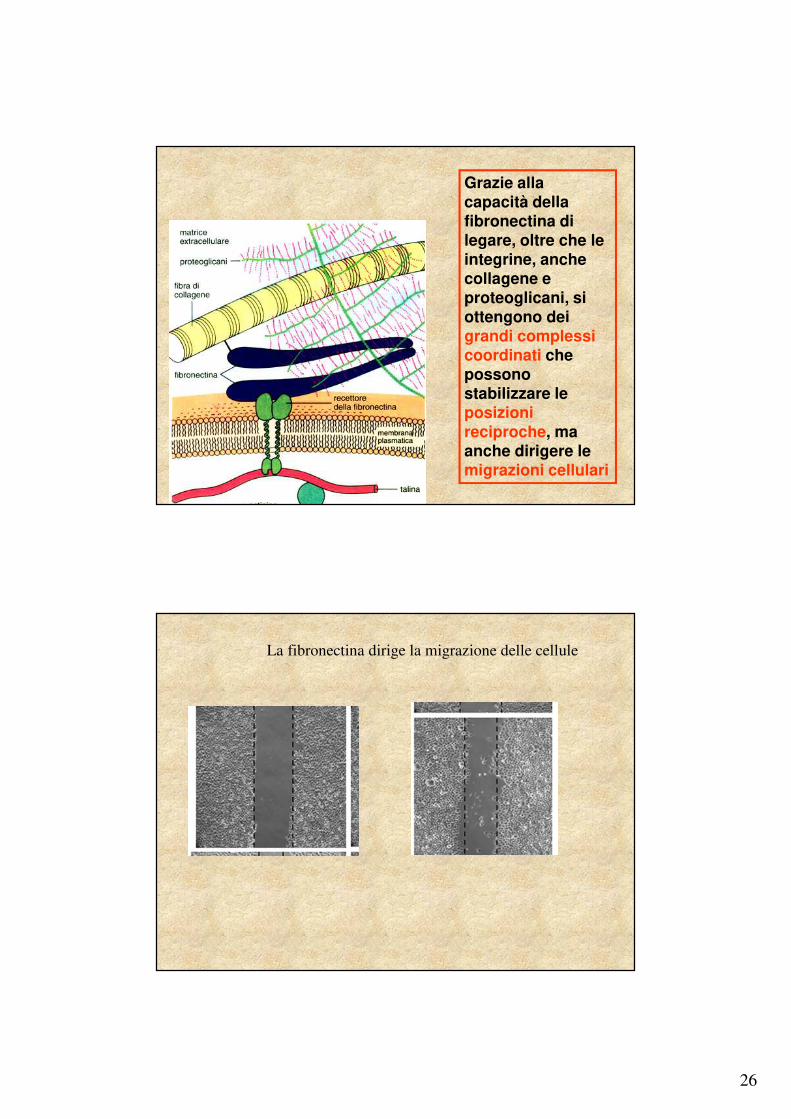
26
Grazie alla capacità della fibronectina di legare, oltre che le integrine, anche collagene e proteoglicani, si ottengono dei grandi complessi coordinati che possono stabilizzare le posizioni reciproche, ma anche dirigere le migrazioni cellulari
La fibronectina dirige la migrazione delle cellule

27
• LE CELLULE DEGLI EPITELI DI RIVESTIMENTO (EPIDERMIDE, EPITELI INTESTINALI E DELL’APPARATO RESPIRATORO), • LE CELLULE MUSCOLARI, • LE CELLULE ADIPOSE, • LE CELLULE ENDOTELIALI DEI VASI SANGUIGNI,• LE CELLULE DI SCHWANN DELLA MIELINA
ADERISCONO AD UN SOTTILE STRATO DI SOSTANZA EXTRACELLULARE DETTO MEMBRANA O LAMINA BASALE
• La lamina basale separa gli epiteli dal sottostante connettivo.

28
RETE DI COLLAGENE DI TIPO IV NELLA MEMBRANA BASALE
La LAMININA, una glicoproteina della membrana basale, presenta siti di legame per cellule, proteoglicani e collagene. E’ responsabile del legame delle cellule epiteliali alla membrana basale.
Recettori di membrana

29
Immagine della laminina al microscopio elettronico
RETE COLLAGENE DI TIPO IV E LAMININA NELLA MEMBRANA BASALE

30
Un modello della membrana basale
IN UNA CAMERA DI COLTURA LE CELLULE MIGRANO LUNGO UNA STRISCIA RICOPERTA DI FIBRONECTINA (EVIDENZIATA DALLE LINEE BIANCHE). LE CELLULE CHE LASCIANO LA TRACCIA DI FIBRONECTINA CAMBIANO MORFOLOGIA E PERDONO LA CAPACITÀ DI MIGRARE

31
“SENTIERI” DI LAMININA O DI FIBRONECTINA GUIDANO LE MIGRAZIONI DELLE CELLULE DURANTE LO SVILUPPO