5 Apparato endocrino - psilab.unige.it · Apparato endocrino Ipofisi Ghiandole surrenali Epifisi...
79
Apparato endocrino
Transcript of 5 Apparato endocrino - psilab.unige.it · Apparato endocrino Ipofisi Ghiandole surrenali Epifisi...
Apparato endocrino

Il presente materiale didattico e ciascuna sua componente sono protetti dalle leggi sul copyright, sono qui proposti in forma aggregata per soli fini di studio e per uso personale.
Sono vietati forme e modi di diffusione, gratuite od onerose, diverse da quelle stabilite dal compilatore.
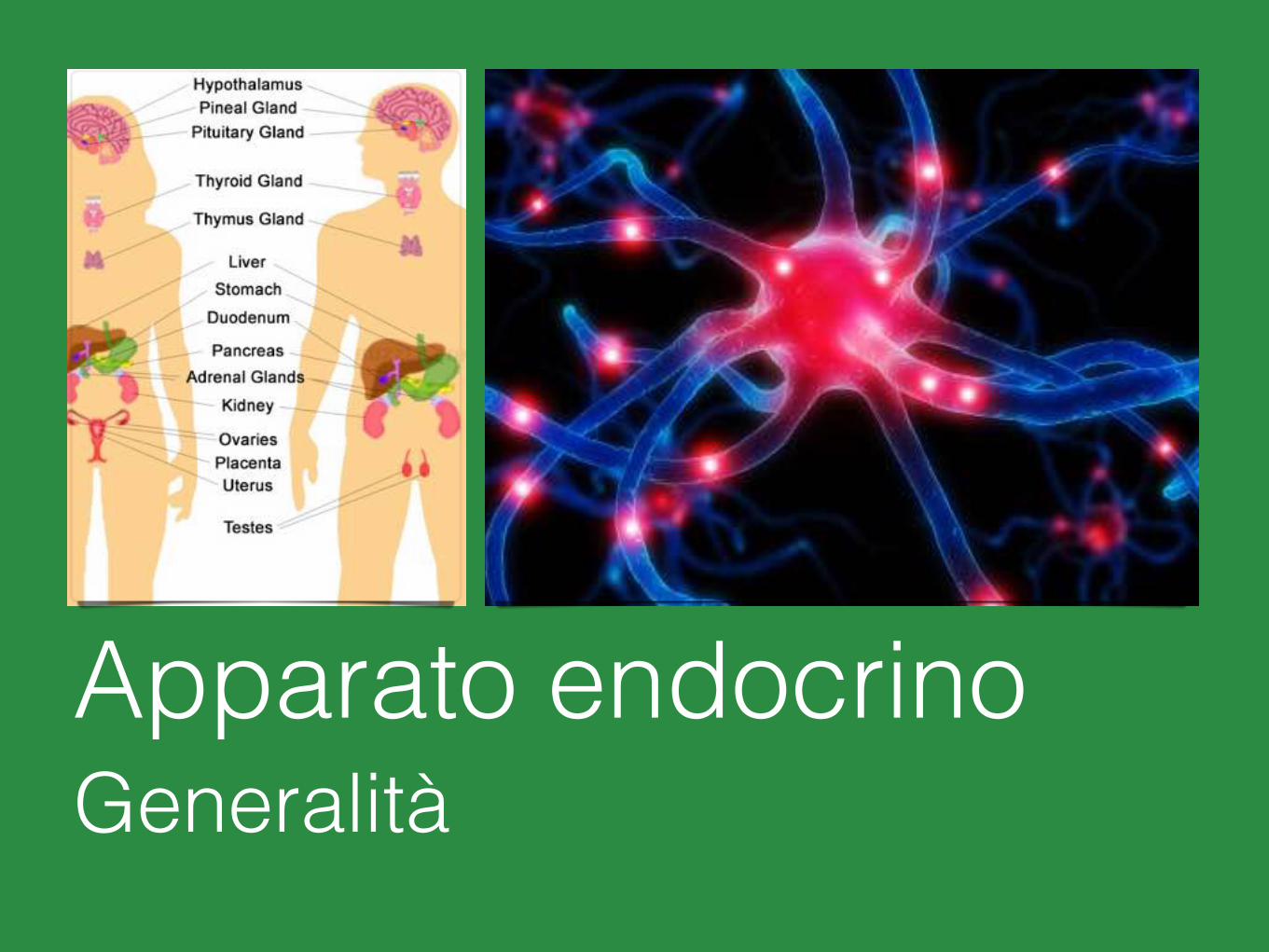
Apparato endocrinoGeneralità
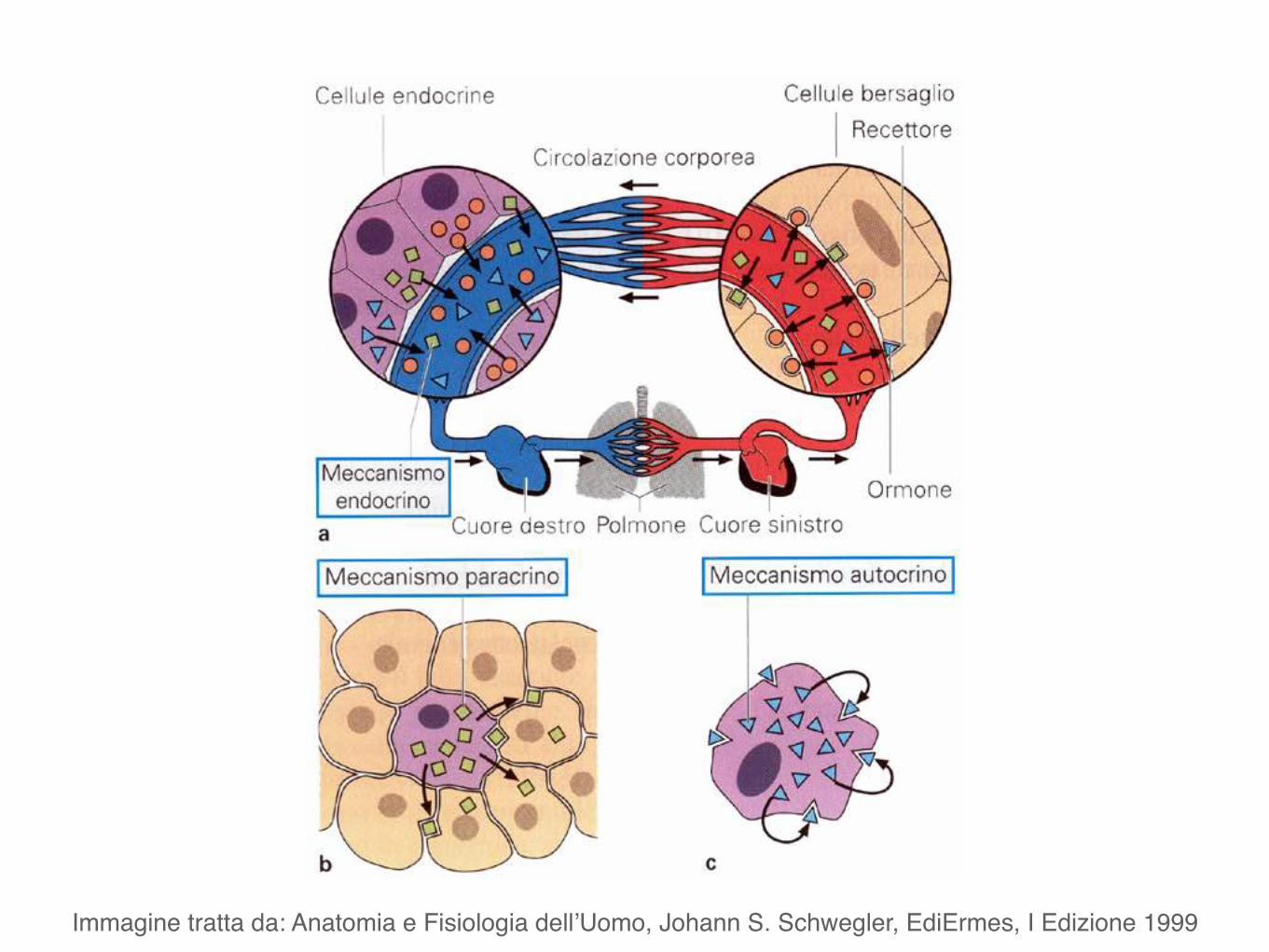
Immagine tratta da: Anatomia e Fisiologia dell’Uomo, Johann S. Schwegler, EdiErmes, I Edizione 1999

Tutti gli ormoni appartengono ad una delle seguenti classi di molecole:
Amine: derivano dagli aminoacidi tirosina e triptofano. Molte amine agiscono come neurotrasmettitori.
Ormoni peptidici: sono catene proteiche generalmente di 8-100 AA. Alcuni ormoni peptidici di dimensioni maggiori sono più o meno fortemente glicosilati (ormoni glicoproteici).
Ormoni steroidei: derivano dal colesterolo.
Derivati degli acidi grassi (prostaglandine e leucotrieni).
Struttura chimica degli ormoni
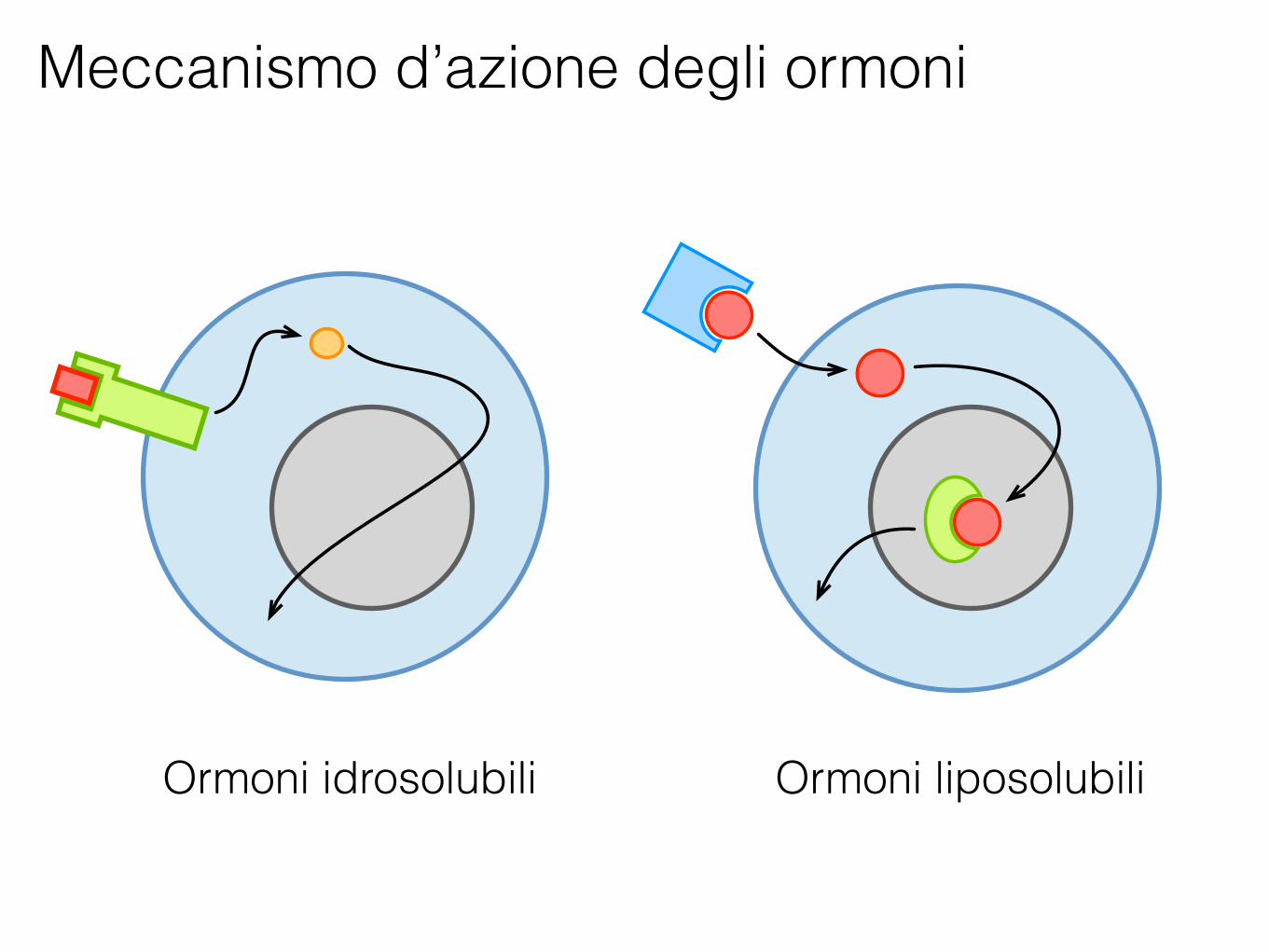
Meccanismo d’azione degli ormoni
Ormoni idrosolubili Ormoni liposolubili
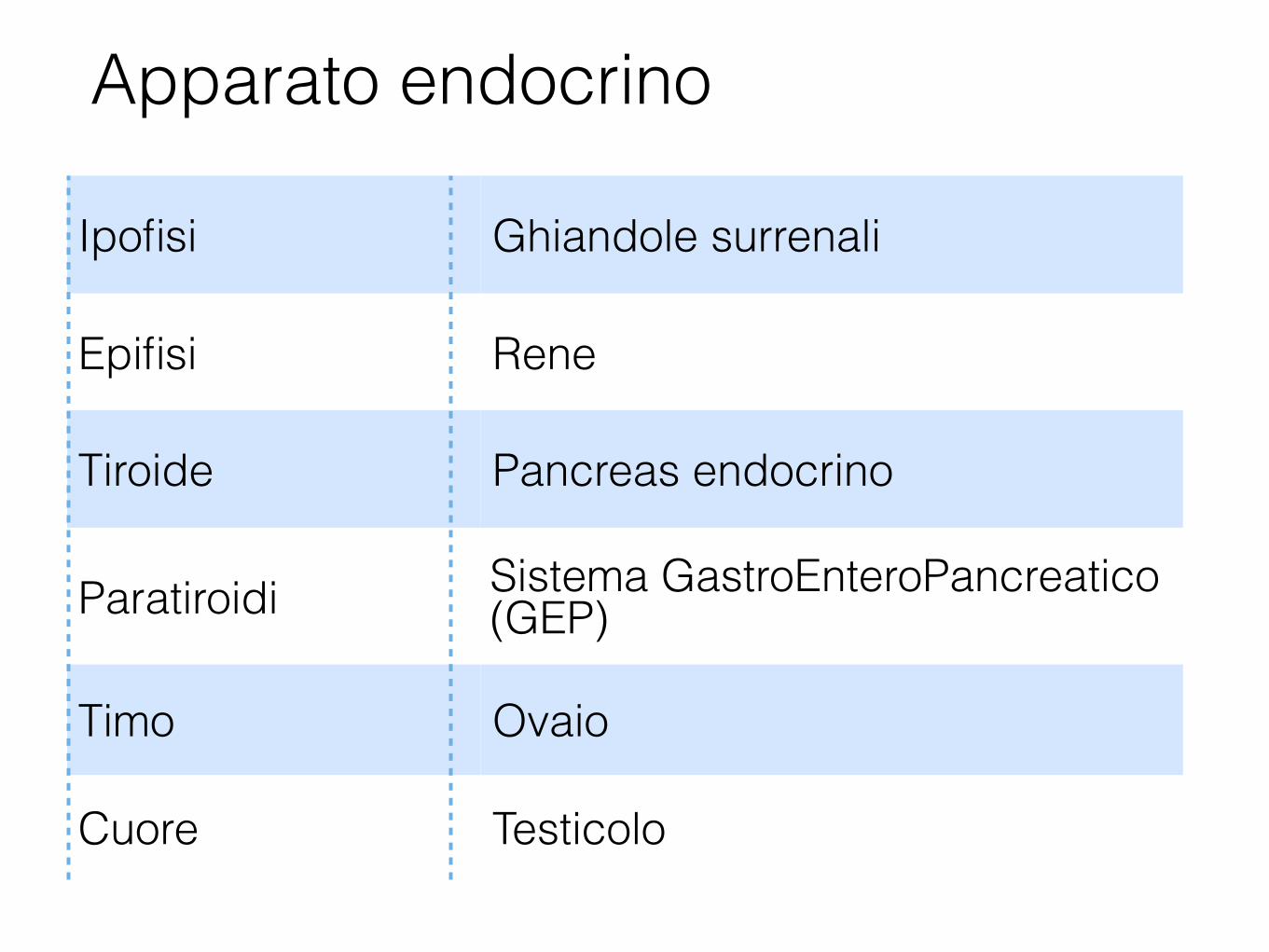
Apparato endocrino
Ipofisi Ghiandole surrenali
Epifisi Rene
Tiroide Pancreas endocrino
Paratiroidi Sistema GastroEnteroPancreatico (GEP)
Timo Ovaio
Cuore Testicolo
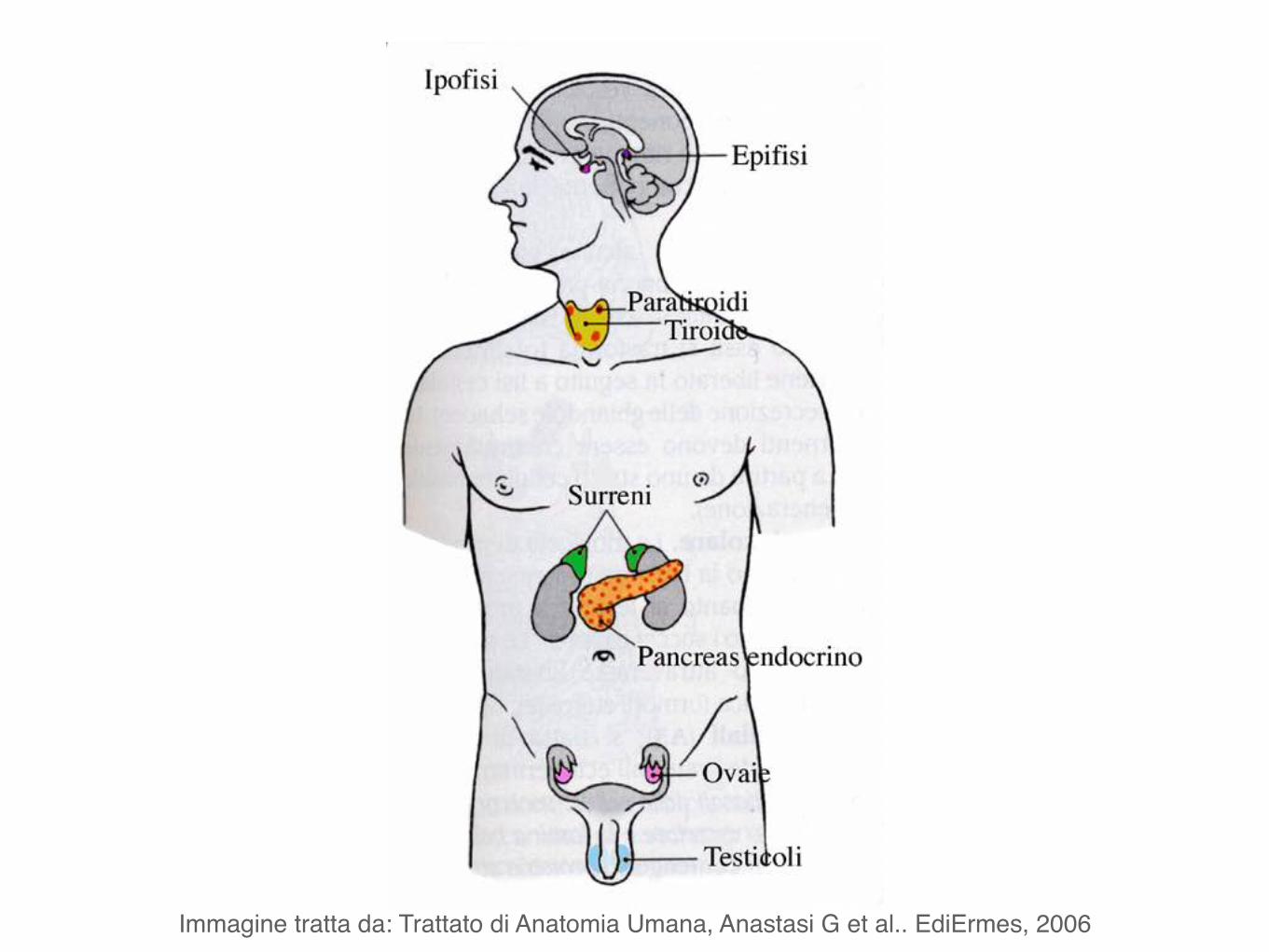
Immagine tratta da: Trattato di Anatomia Umana, Anastasi G et al.. EdiErmes, 2006
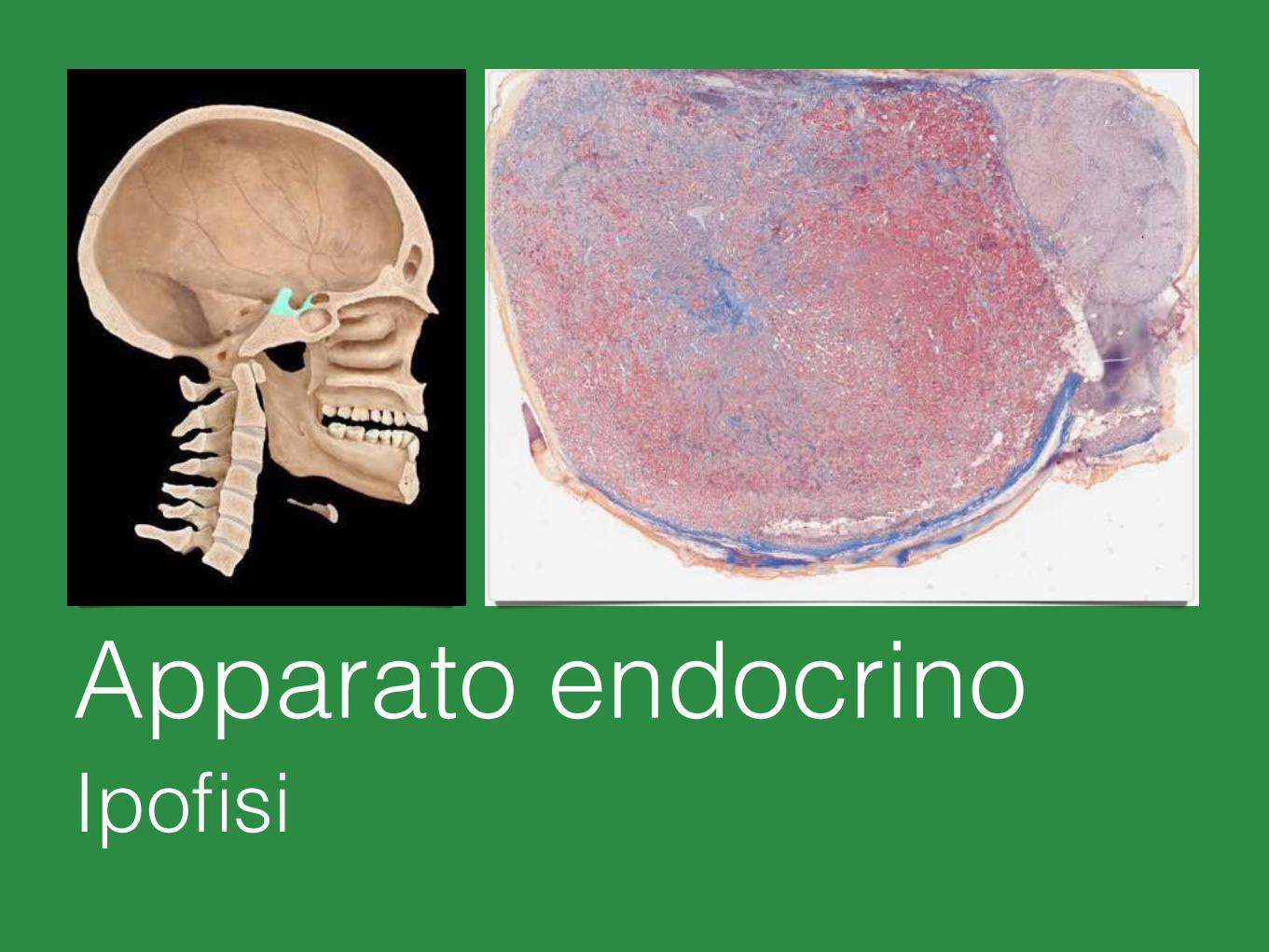
Apparato endocrinoIpofisi
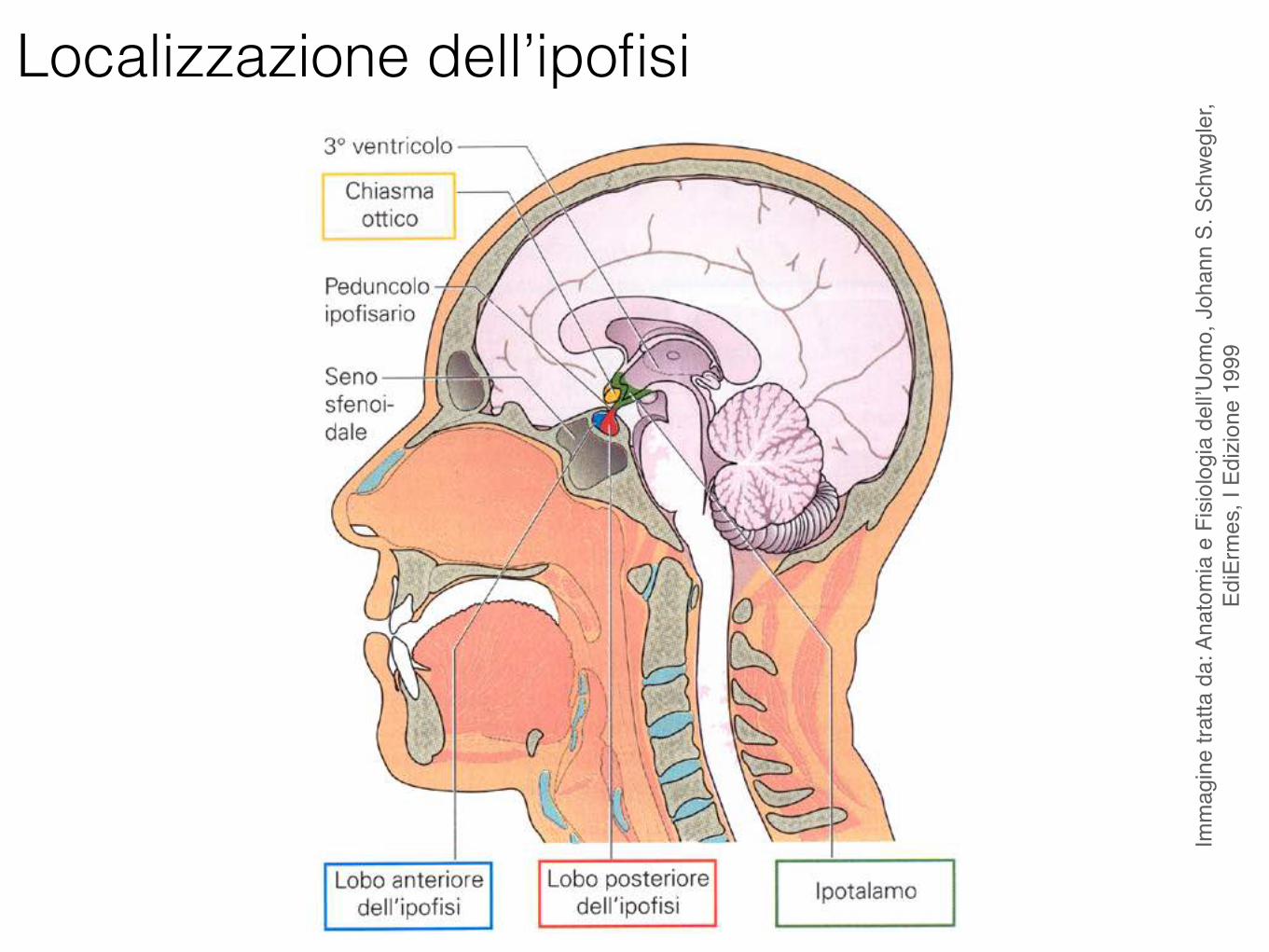
Localizzazione dell’ipofisi
Imm
agin
e tra
tta d
a: A
nato
mia
e F
isiol
ogia
del
l’Uom
o, J
ohan
n S.
Sch
wegl
er,
EdiE
rmes
, I E
dizio
ne 1
999

L’ipofisi (ghiandola pituitaria) è lunga circa 1 cm, larga 1-1.5 cm e spessa 0.5 cm circa. Ha un peso di 0.4-1 g.
L’ipofisi comprende due porzioni principali:
• l’adenoipofisi (lobo anteriore) • la neuroipofisi (lobo posteriore)
Ipofisi

Adenoipofisi (lobo anteriore)
parte distale (lobo anteriore)
parte tuberale (infundibulare) parte intermedia
Neuroipofisi (lobo posteriore) è collegata al pavimento del diencefalo tramite il peduncolo ipofisario.
parte nervosa (lobo posteriore)
processo infundibolare eminenza mediana
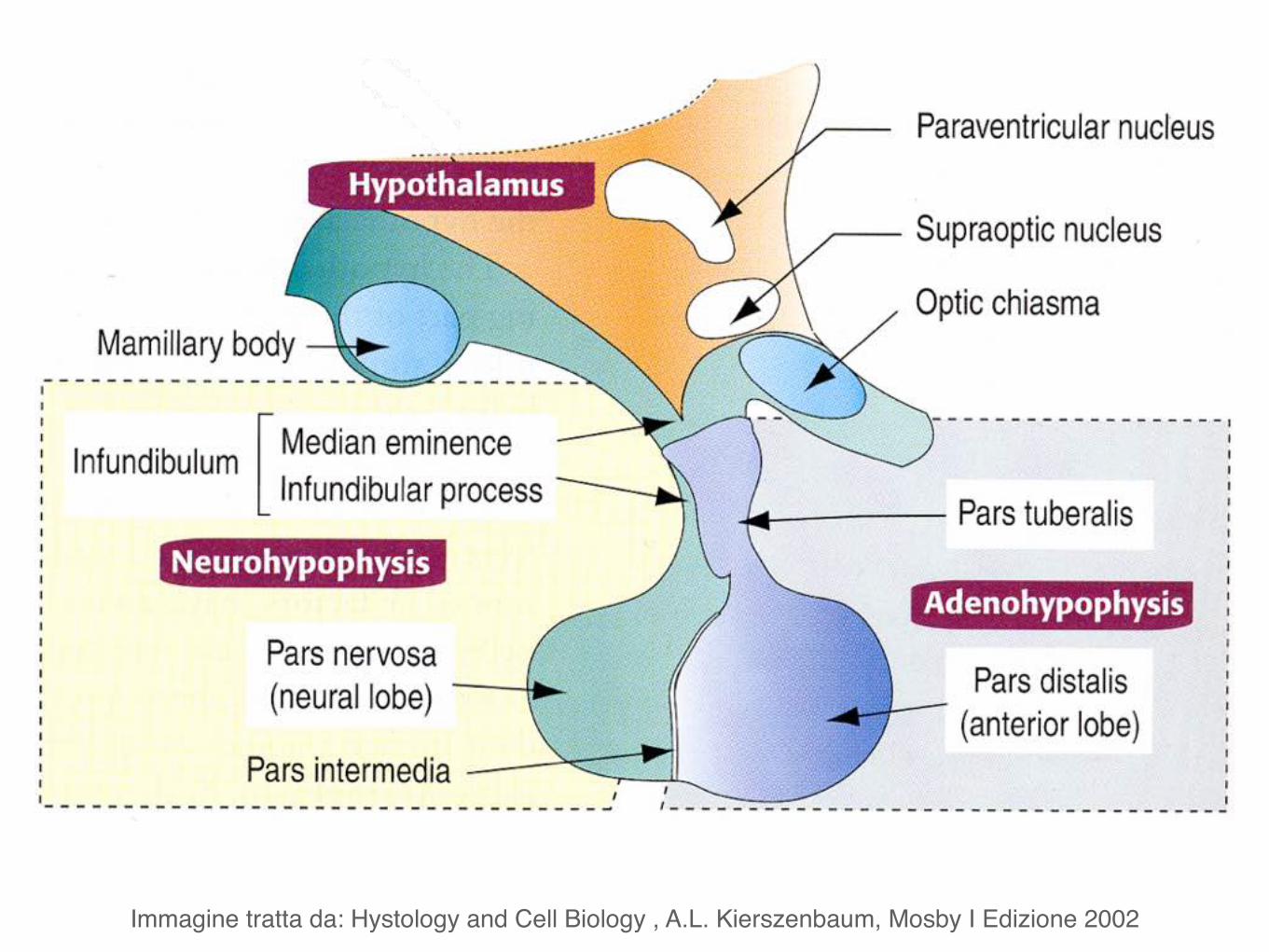
Immagine tratta da: Hystology and Cell Biology , A.L. Kierszenbaum, Mosby I Edizione 2002
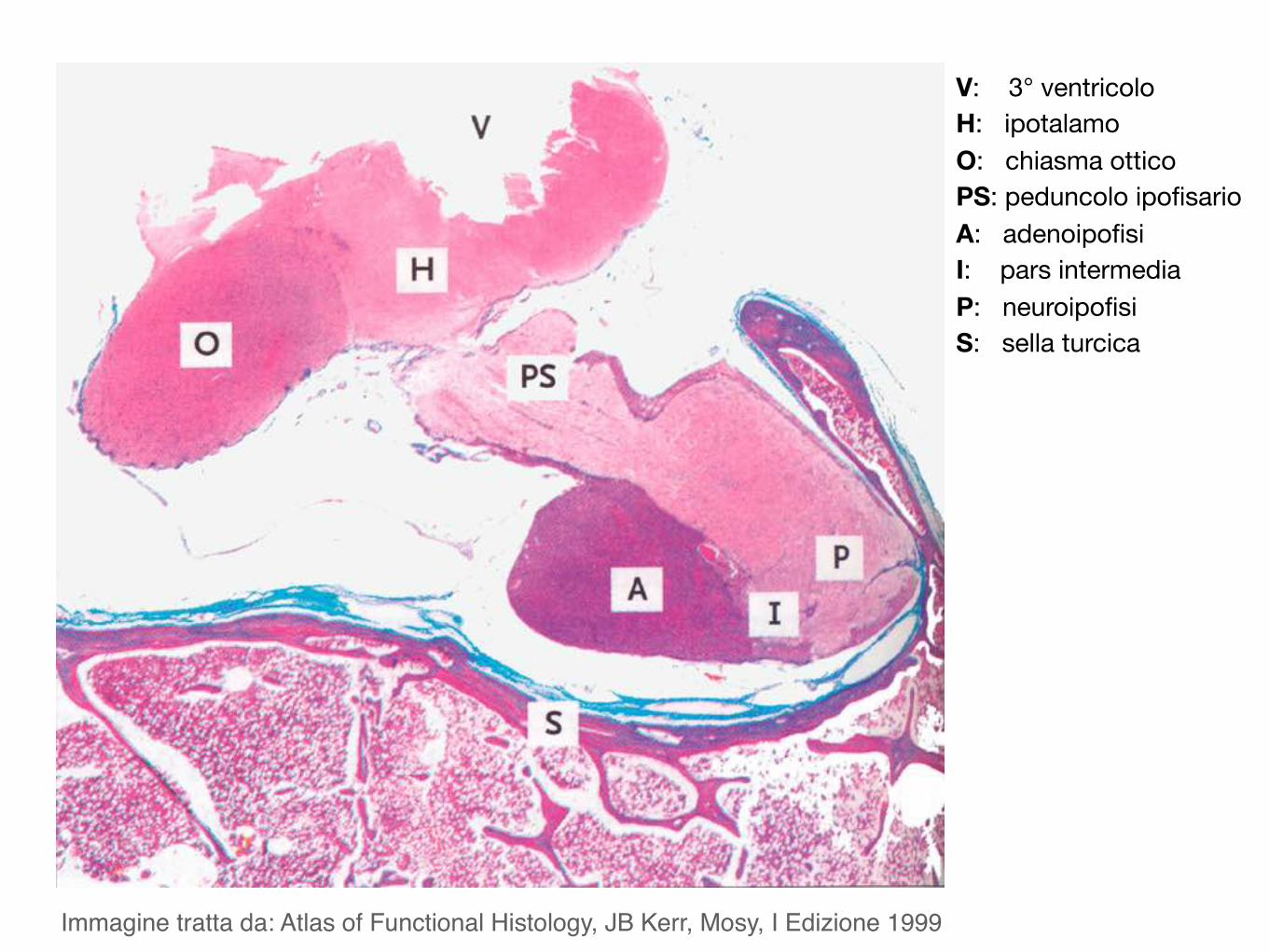
Immagine tratta da: Atlas of Functional Histology, JB Kerr, Mosy, I Edizione 1999
V: 3° ventricoloH: ipotalamoO: chiasma otticoPS: peduncolo ipofisarioA: adenoipofisiI: pars intermediaP: neuroipofisiS: sella turcica
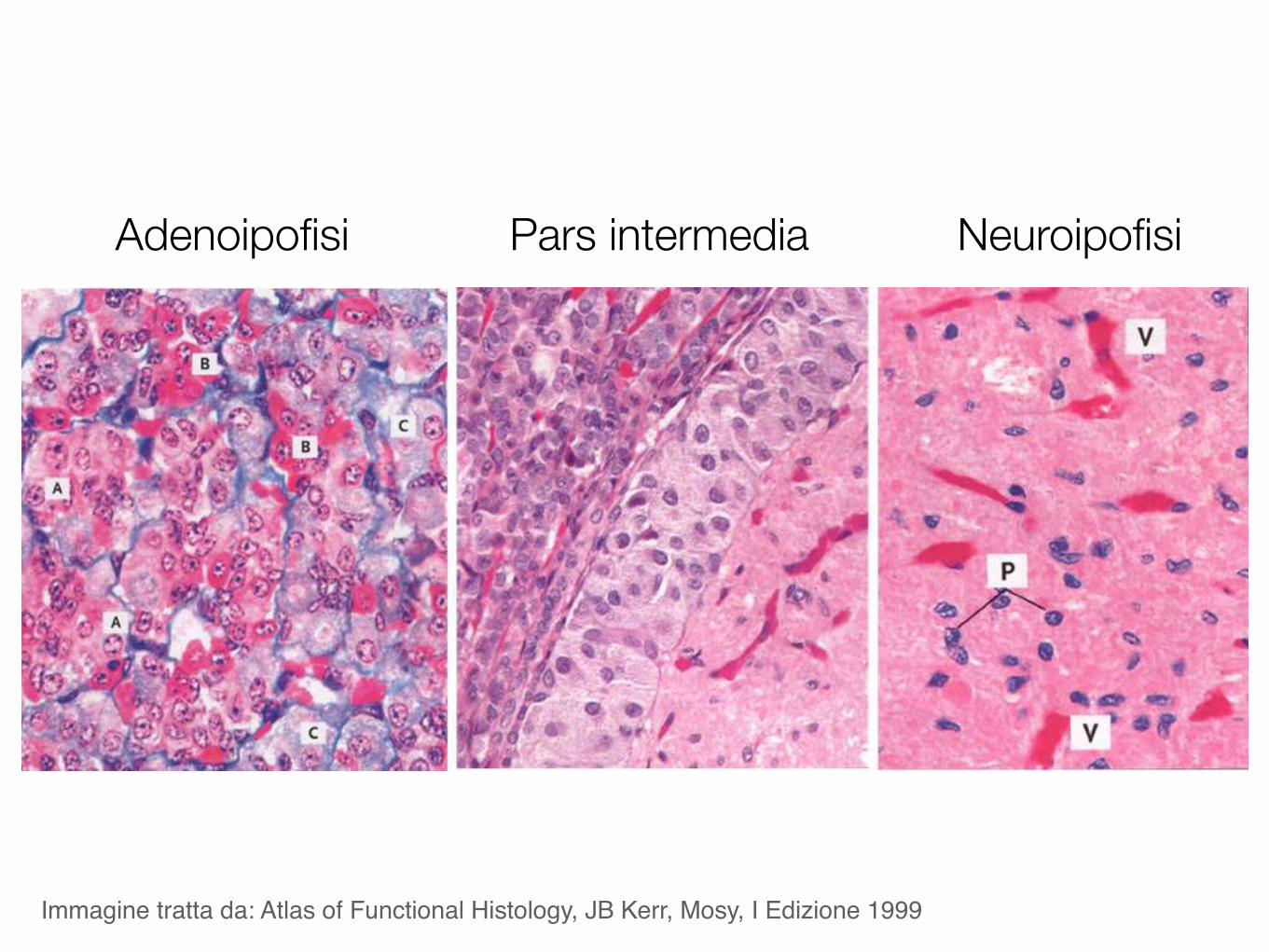
Adenoipofisi Pars intermedia Neuroipofisi
Immagine tratta da: Atlas of Functional Histology, JB Kerr, Mosy, I Edizione 1999
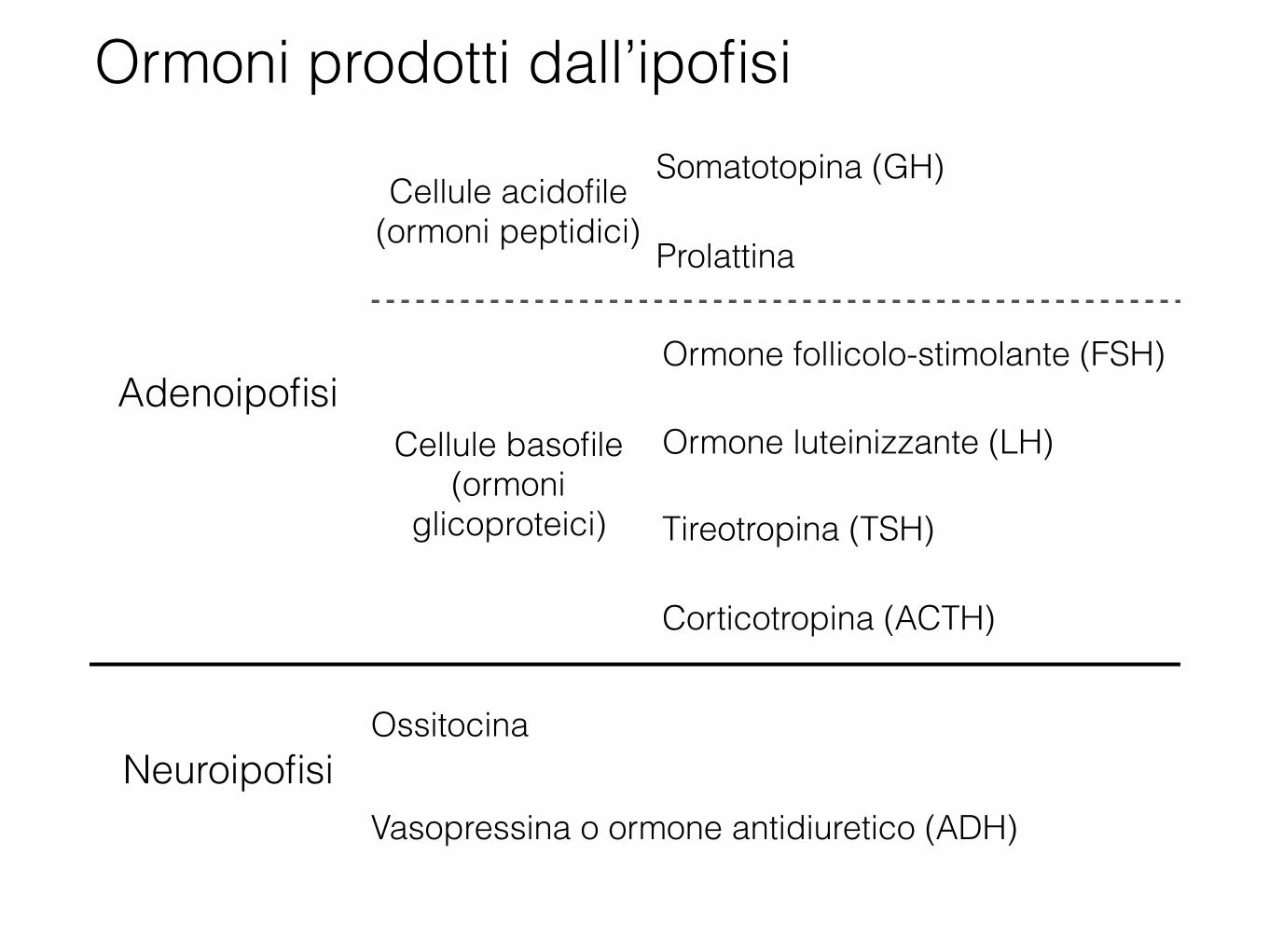
Adenoipofisi
Cellule acidofile (ormoni peptidici)
Somatotopina (GH)
Prolattina
Cellule basofile (ormoni
glicoproteici)
Ormone follicolo-stimolante (FSH)
Ormone luteinizzante (LH)
Tireotropina (TSH)
Corticotropina (ACTH)
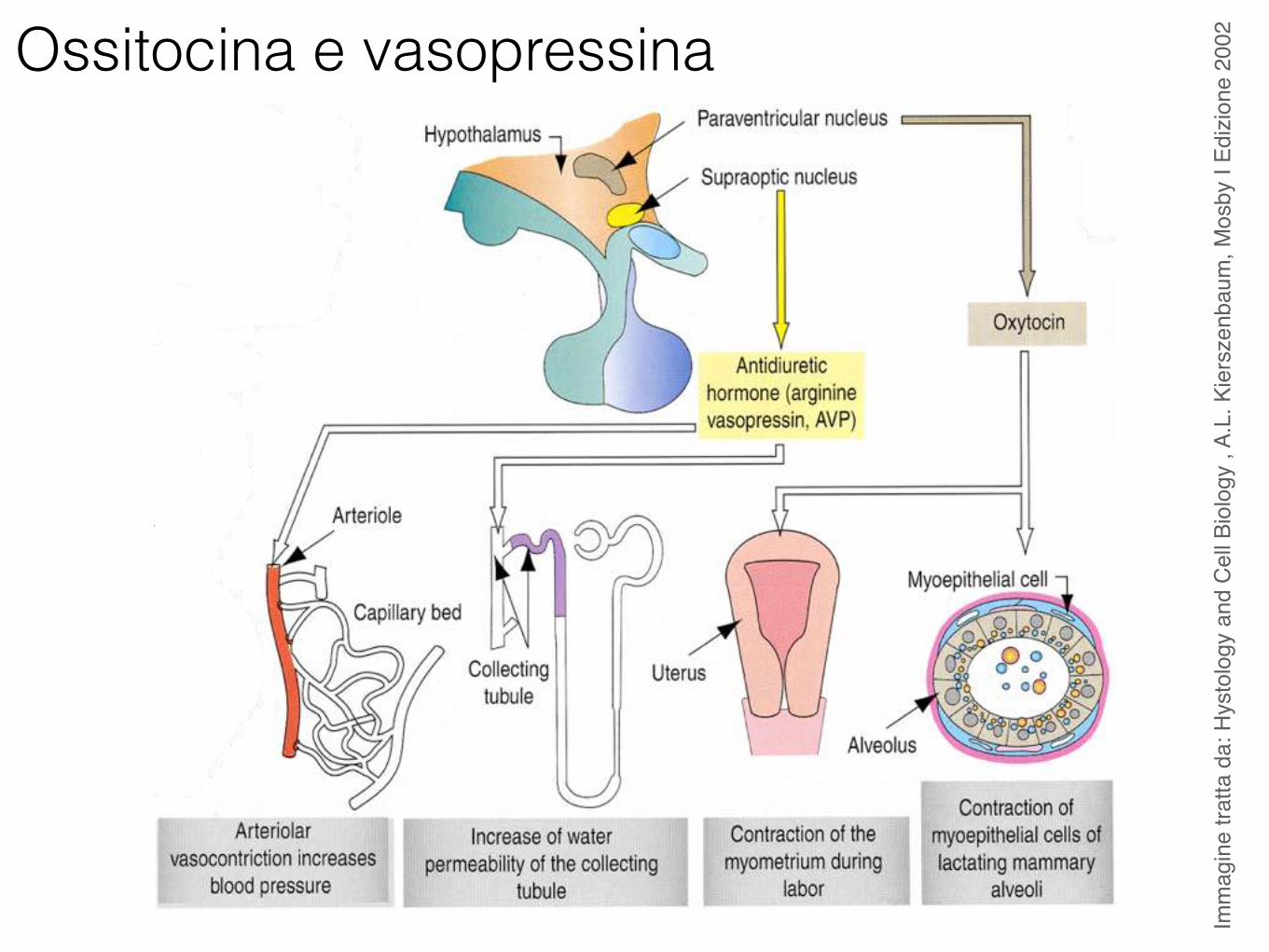
NeuroipofisiOssitocina
Vasopressina o ormone antidiuretico (ADH)
Ormoni prodotti dall’ipofisi

Neuroipofisi
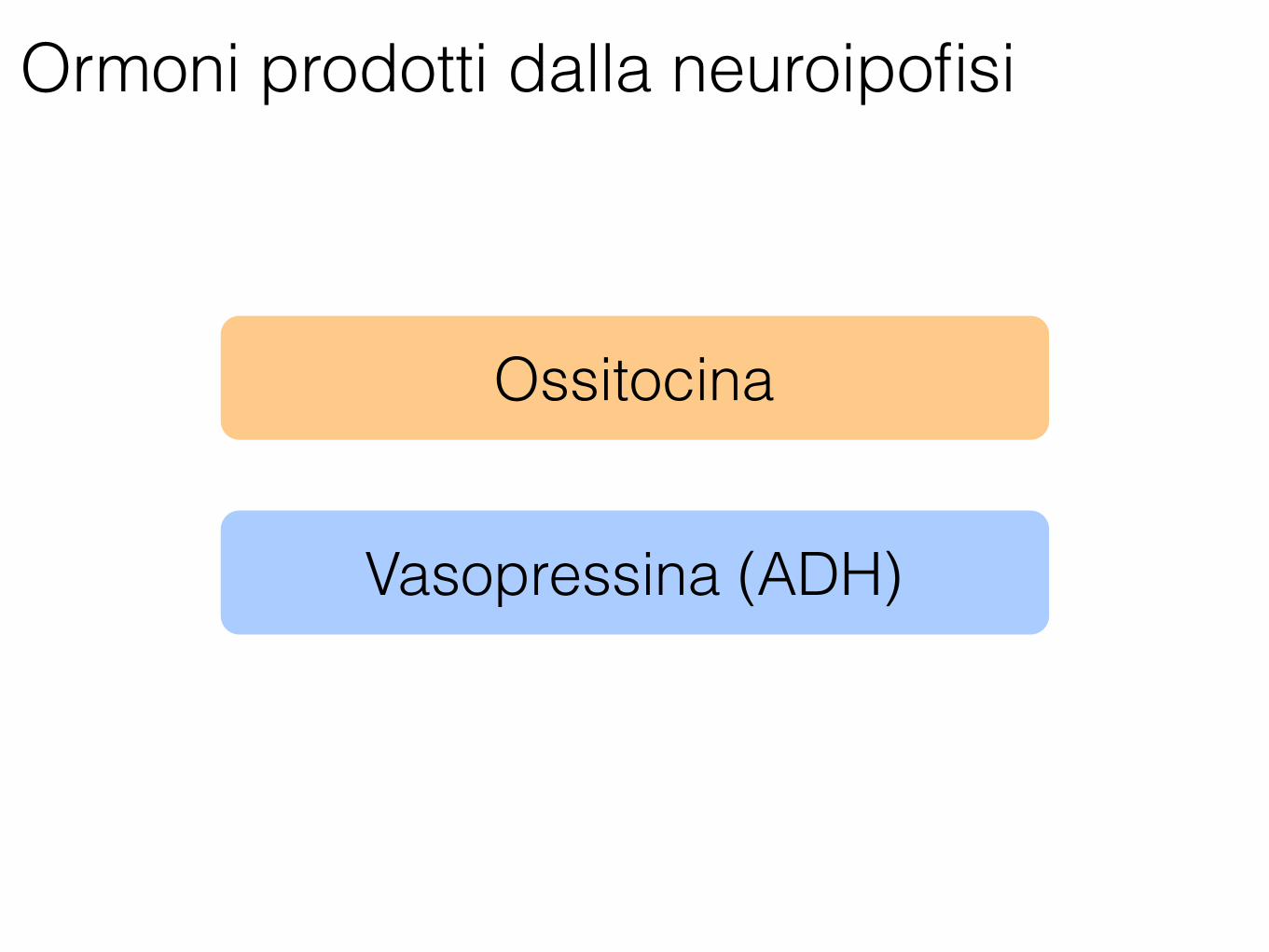
Ormoni prodotti dalla neuroipofisi
Ossitocina
Vasopressina (ADH)
Imm
agin
e tra
tta d
a: T
he H
uman
Bra
in, J
. Nol
te, M
osby
V E
dizio
ne 2
002
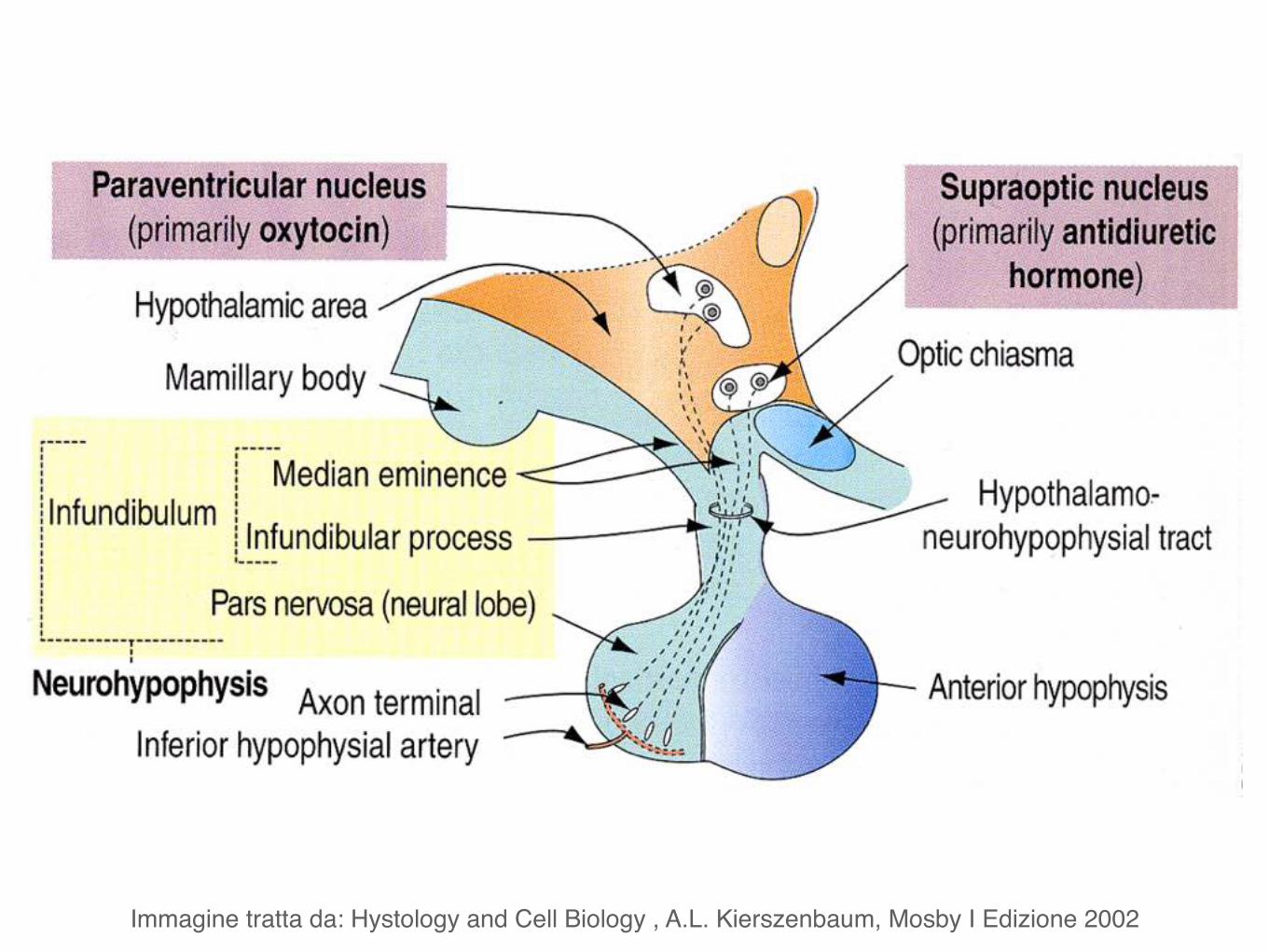
Immagine tratta da: Hystology and Cell Biology , A.L. Kierszenbaum, Mosby I Edizione 2002
Imm
agin
e tra
tta d
a: H
ysto
logy
and
Cel
l Bio
logy
, A.
L. K
iers
zenb
aum
, Mos
by I
Edizi
one
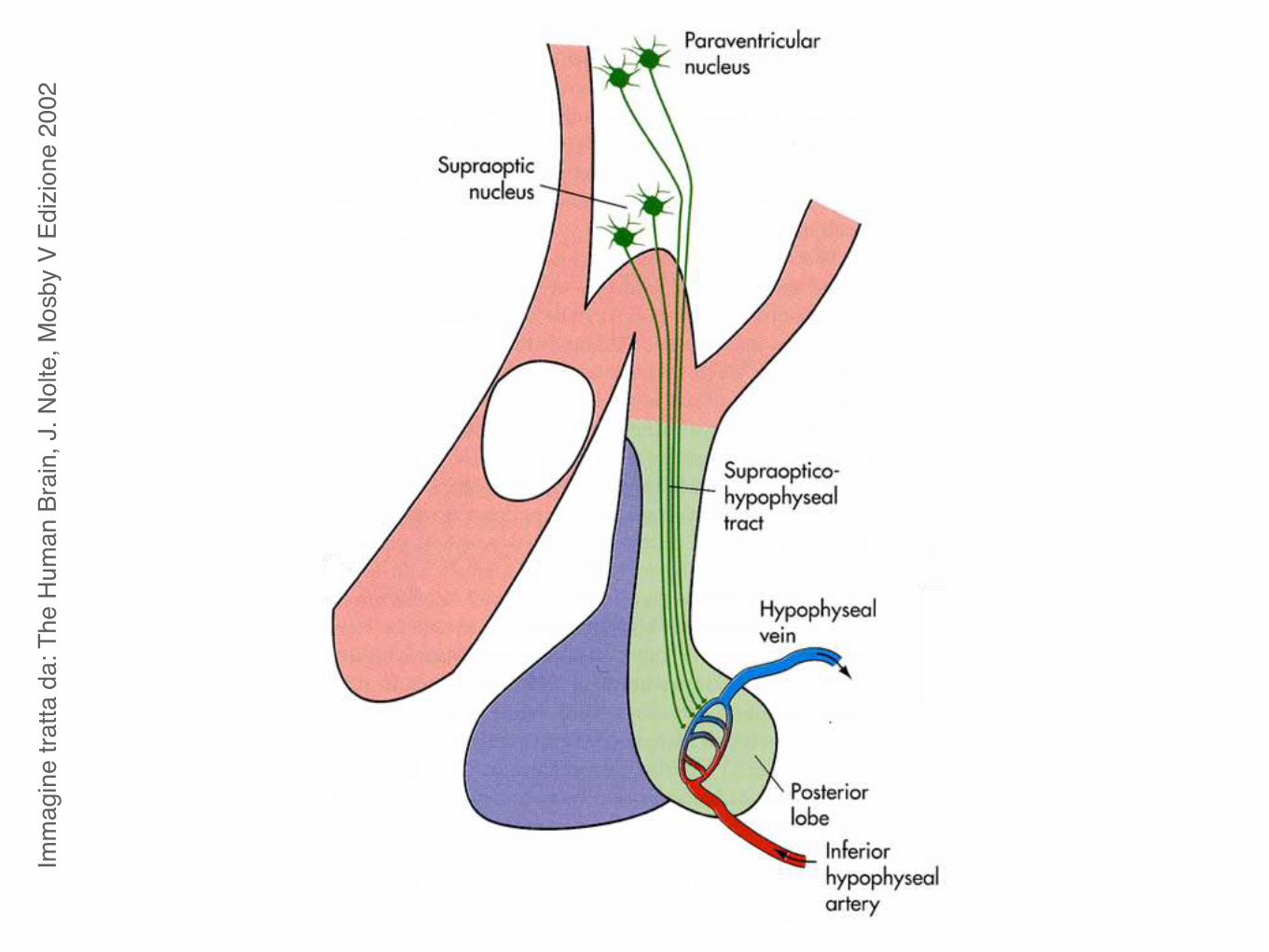
2002Ossitocina e vasopressina

13. E. D. Jarvis, C. Scharff, M. R. Grossman, J. A. Ramos,F. Nottebohm, Neuron 21, 775 (1998).
14. S. S. Burmeister, E. D. Jarvis, R. D. Fernald, PloS Biol. 3,e363 (2005).
15. L. Grosenick, T. S. Clement, R. D. Fernald, Nature 445,429 (2007).
16. K. J. O'Donovan, W. G. Tourtellotte, J. Milbrandt,J. M. Baraban, Trends Neurosci. 22, 167 (1999).
17. C. W. Whitfield, A.-M. Cziko, G. E. Robinson, Science 302,296 (2003).
18. C. M. Grozinger, N. Sharabash, C. W. Whitfield,G. E. Robinson, Proc. Natl. Acad. Sci. U.S.A. 100, 14519(2003).
19. S. A. Ament, M. Corona, H. S. Pollack, G. E. Robinson,Proc. Natl. Acad. Sci. U.S.A. 105, 4226 (2008).
20. M. E. Cummings et al., Proc. R. Soc. London Ser.B. Biol. Sci. 275, 393 (2008).
21. K. Replogle et al., BMC Genomics 9, 131 (2008).22. H. Liu, M. M. Wu, H. H. Zakon, Dev. Neurobiol. 67, 1289
(2007).23. C. Cirelli, C. M. Gutierrez, G. Tononi, Neuron 41, 35
(2004).
24. F. A. Champagne, D. D. Francis, A. Mar, M. J. Meaney,Physiol. Behav. 79, 359 (2003).
25. I. C. Weaver et al., Nat. Neurosci. 7, 847 (2004).26. D. L. Champagne et al., J. Neurosci. 28, 6037 (2008).27. D. W. Pfaff, Drive (MIT Press, Cambridge, MA, 1999).28. D. S. Falconer, T. F. C. Mackay, Introduction to
Quantitative Genetics (Longmans Green, Harlow Essex,UK, ed. 4, 1996).
29. A. Caspi, T. E. Moffitt, Nat. Rev. Neurosci. 7, 583 (2006).30. D. K. Lahiri, B. Maloney, Nat. Rev. Neurosci. 10.1038/
nrn2022-c1 (2006).31. A. Abbott, Nature 454, 154 (2008).32. J.Wang, K. G. Ross, L. Keller, PLoSGenet.4, e1000127 (2008).33. B. J. Dickson, Science 322, 904 (2008).34. D. A. Wheeler et al., Science 251, 1082 (1991).35. S. Fink, L. Excoffier, G. Heckel, Proc. Natl. Acad. Sci. U.S.A.
103, 10956 (2006).36. Y. Ben-Shahar, A. Robichon, M. B. Sokolowski, G. E.
Robinson, Science 296, 741 (2002).37. A. L. Toth, G. E. Robinson, Trends Genet. 23, 334 (2007).38. W. Enard et al., Nature 418, 869 (2002).39. W. Shu et al., Proc. Natl. Acad. Sci. U.S.A. 102, 9643 (2005).
40. S. Haesler et al., PLoS Biol. 5, e321 (2007).41. M. Groszer et al., Curr. Biol. 18, 354 (2008).42. J. Sebat et al., Science 316, 445 (2007), published
online 14 March 2007; 10.1126/science.1138659.43. J. D. Jensen, A. Wong, C. F. Aquadro, Trends Genet. 23,
568 (2007).44. J. C. Hall, J. Neurogenet. 17, 1 (2003).45. We thank J. Desjardins, A. Fernald, K. A. Hughes,
D. B. Kelley, K. Maruska, C. Olin, M. B. Sokolowski,L. J. Stubbs, members of the Clayton and Robinsonlaboratories, and two anonymous reviewers for reviewsof this manuscript and C. Harrell for graphical assistance.Research by the authors cited here was supported bythe following grants: NIH R01 NS051820 and NS045264(D.F.C.); NIH NS34950 Javits Award and UnitedStates–Israel Binational Science Foundation 200596(R.D.F.); and NIH R01 GM073644, NSF Frontiers inBiological Research EF04-25852, U.S. Department ofAgriculture AG2003-35302-13490, and a BurroughsWellcome Fund Innovation Award (G.E.R.).
10.1126/science.1159277
REVIEW
Oxytocin, Vasopressin, and theNeurogenetics of SocialityZoe R. Donaldson1 and Larry J. Young1,2*
There is growing evidence that the neuropeptides oxytocin and vasopressin modulate complexsocial behavior and social cognition. These ancient neuropeptides display a marked conservation ingene structure and expression, yet diversity in the genetic regulation of their receptors seems tounderlie natural variation in social behavior, both between and within species. Human studies arebeginning to explore the roles of these neuropeptides in social cognition and behavior and suggestthat variation in the genes encoding their receptors may contribute to variation in human socialbehavior by altering brain function. Understanding the neurobiology and neurogenetics of socialcognition and behavior has important implications, both clinically and for society.
Social interactions affect every aspect ofour lives, from wooing a mate and caringfor our children to determining our suc-
cess in the workplace. Abnormal manifestationsof social behavior, such as the pathologicaltrusting associated with Williams-Beuren Syn-drome (1), social withdrawal in depression, anddecreased social cognition in autism, profoundlyaffect the lives of those who suffer from thesedisorders. Neuroscientists once considered socialbehavior to be too hopelessly complex to under-stand at a mechanistic level, but advances in ani-mal models of social cognition and bonding, aswell as application of new technologies in humanresearch have demonstrated that the molecularbasis of social behavior is not beyond the realmof our understanding. There appears to be markedconservation in the molecular mechanisms regulat-
ing social behavior across diverse species, includ-ing our own.
Interacting with other neurotransmitter systemswithin specific neural circuits, neuropeptides haveemerged as central players in the regulation of socialcognition and behavior. Neuropeptides may act asneurotransmitters, if released within synapses, or asneurohormones, activating receptors distant fromthe site of release, which provides evolutionaryflexibility to their actions (2). Within vertebrates, amajority of work relating neuropeptides to socialbehavior has focused on members of the oxytocin/vasopressin family. Homologs of oxytocin andvasopressin existed at least 700 million years agoand have been identified in such diverse organismsas hydra, worms, insects, and vertebrates. Amongthese distant taxa, oxytocin- and vasopressin-relatedpeptides play a general role in the modulation ofsocial and reproductive behaviors. In contrast tothis apparent conservation in function, the spe-cific behaviors affected by these neuropeptidesare notably species-specific.
Only recently have scientists begun to dissectthe roles of oxytocin, vasopressin, and their re-
lated receptors in human social behavior. Whereashuman social behavior is more nuanced and com-plex than the behaviors typically assayed in otheranimals, this complexity has created unique op-portunities to design finely honed tasks that haverevealed a potential role for these peptides in per-sonality, trust, altruism, social bonding, and ourability to infer the emotional state of others. Here,we review the evidence of evolutionary conserva-tionwithin the vasopressin/oxytocin peptide fam-ily, briefly discuss the role of these peptides andtheir respective receptors in modulating socialbehavior and bonding, and provide a synthesisof recent advances implicating the oxytocin andvasopressin systems in human trust, cooperation,and social behavior.
Conservation of Neuropeptide SystemsRegulating Social BehaviorThe mammalian oxytocin and vasopressin non-apeptides, so called for their nine–amino acidcomposition, differ from each other at only twoamino acid positions (Fig. 1). Oxytocin, vasopres-sin, and their respective nonmammalian vertebratelineages are thought to have arisen from a gene-duplication event before vertebrate divergence.Within these lineages, peptides vary by a singleamino acid, and their genes are found near eachother on the same chromosome. Invertebrates, withfew exceptions, have only one oxytocin/vasopressinhomolog, whereas vertebrates have two (3, 4).
In mammals, oxytocin and vasopressin are pro-duced primarily within hypothalamic brain regionsand then shuttled to the pituitary for peripheralrelease or projected to various brain regions.Notably,just as oxytocin andvasopressin are expressedwithinthe hypothalamus of mammals, their homologs areexpressed within similar neurosecretory brainregions of organisms as diverse as worms and fish.A characterization of annepressin (the homolog ofoxytocin/vasopressin in segmented worms) andvasotocin (vasopressin’s counterpart in bony fish)revealed conserved neural expression of these genes
1Center for Behavioral Neuroscience, Yerkes Regional PrimateResearch Center, Emory University, Atlanta, GA 30322, USA.2Department of Psychiatry and Behavioral Sciences, EmoryUniversity, Atlanta, GA 30329, USA.
*To whom correspondence should be addressed. E-mail:[email protected]
7 NOVEMBER 2008 VOL 322 SCIENCE www.sciencemag.org900
Genetics of Behavior
CORRECTED 13 MARCH 2009; SEE LAST PAGE
Neuron
Review
The Challenge of Translationin Social Neuroscience: A Review of Oxytocin,Vasopressin, and Affiliative Behavior
Thomas R. Insel1,*1National Institute of Mental Health, National Institutes of Health, Bethesda, MD 20892, USA*Correspondence: [email protected] 10.1016/j.neuron.2010.03.005
Social neuroscience is rapidly exploring the complex territory between perception and action where recog-nition, value, and meaning are instantiated. This review follows the trail of research on oxytocin and vaso-pressin as an exemplar of one path for exploring the ‘‘dark matter’’ of social neuroscience. Studies acrossvertebrate species suggest that these neuropeptides are important for social cognition, with gender- andsteroid-dependent effects. Comparative research in voles yields amodel based on interspecies and intraspe-cies variation of the geography of oxytocin receptors and vasopressin V1a receptors in the forebrain. Highlyaffiliative species have receptors in brain circuits related to reward or reinforcement. The neuroanatomicaldistribution of these receptorsmay be guided by variations in the regulatory regions of their respective genes.This review describes the promises and problems of extrapolating these findings to human social cognition,with specific reference to the social deficits of autism.
Social neuroscience has come a long way in a short time.Two decades ago, a gap existed between behavioral neurosci-ence, systems neuroscience, behavioral ecology, and socialpsychology. Today, the field of social neuroscience fills thisgap with abundance: social neuroscience now has its own jour-nals, textbooks, societies, and, according to PubMed, nearly3000 research papers (as of February 22, 2010). Much of thisstunning growth has been driven by human neuroimagingstudies seeking the neural correlates of psychological pro-cesses, from face perception to social preferences. Socialneuroscience has a different foundation in animal studies, builton molecular and cellular approaches as well as the tools ofsystems neuroscience. In fact, the history of animal studies ofsocial perception and social behavior with classical lesion andneurophysiological techniques extends back several decadesin the venerable literatures of neuroethology and behavioralneuroendocrinology. This review will follow a single thread ofsocial neuroscience spun from this older animal research,recently woven into human studies and now suggesting potentialtreatments of human disorders of social behavior, such asautism.
Social Neuroscience in 2010Most of social neuroscience can be separated into studies ofeither receptive or expressive processes. Receptive studies,which emerged from neuroethology, focus on sensory process-ing. From the elegant work on pheromone receptors in mice(Dulac and Torello, 2003) to the careful mapping of face cells inhuman and nonhuman primates (Kanwisher, 2006), this arm ofsocial neuroscience has described the neural geography and,in some cases, the cellular landscape by which sensory informa-tion is initially encoded as social. A fundamental insight fromreceptive studies is that, in most vertebrates, the brain employsspecific receptors or cortical regions for processing social
information, whether that information is from pheromonal/olfac-tory, audio-vocal, somatosensory, or visual cues. That is, socialinformation is not simply complex multisensory perception; it isperceived and encoded in unique ways in the brain.Expressive studies, long the domain of behavioral neurosci-
ence and behavioral neuroendocrinology, focus on socialinteractions: communication, reproductive behavior (especiallyparental care and sex), agonistic actions (aggression and preda-tion), and affiliative behaviors (including social play). In verte-brates (and many invertebrates), nearly all of these behaviorsare influenced by gonadal steroids (estrogens and androgens),acting via their nuclear receptors. The mapping of gonadalsteroid receptor expression in the brain helped identify keyregions for social behavior (McEwen et al., 1979; Pfaff andKeiner, 1973; Pfaff and McEwen, 1983). Additionally, steroidreceptors are transcription factors, and by identifying the genesregulated by steroid receptors, a molecular basis for socialbehavior could be proposed (Warnmark et al., 2003).Between the receptive and expressive arms sits the great dark
matter of social neuroscience. (Figure 1) What happens betweenthe stage when a percept is encoded as ‘‘social’’ to the stagewhen a ‘‘go’’ signal is given for initiating social behavior? Howdoes the brain distinguish prey from predator, juvenile fromadult, novel from familiar, kin from unrelated conspecific? Whatare the neural mechanisms that facilitate or inhibit social interac-tion? These are the questions that have been more difficult toanswer.Recently, researchers have begun to address these questions
with human neuroimaging studies seeking to map variousaspects of higher-order processing of social information andeven develop computational principles for the neural basis ofsocial cognition (Behrens et al., 2009). Human neuroimagingstudies can describe the cortical patterns, but mechanisticstudies are still mostly the domain of animal research. In fact,
768 Neuron 65, March 25, 2010 ª2010 Elsevier Inc.
Neuron
Review
The Challenge of Translationin Social Neuroscience: A Review of Oxytocin,Vasopressin, and Affiliative Behavior
Thomas R. Insel1,*1National Institute of Mental Health, National Institutes of Health, Bethesda, MD 20892, USA*Correspondence: [email protected] 10.1016/j.neuron.2010.03.005
Social neuroscience is rapidly exploring the complex territory between perception and action where recog-nition, value, and meaning are instantiated. This review follows the trail of research on oxytocin and vaso-pressin as an exemplar of one path for exploring the ‘‘dark matter’’ of social neuroscience. Studies acrossvertebrate species suggest that these neuropeptides are important for social cognition, with gender- andsteroid-dependent effects. Comparative research in voles yields amodel based on interspecies and intraspe-cies variation of the geography of oxytocin receptors and vasopressin V1a receptors in the forebrain. Highlyaffiliative species have receptors in brain circuits related to reward or reinforcement. The neuroanatomicaldistribution of these receptorsmay be guided by variations in the regulatory regions of their respective genes.This review describes the promises and problems of extrapolating these findings to human social cognition,with specific reference to the social deficits of autism.
Social neuroscience has come a long way in a short time.Two decades ago, a gap existed between behavioral neurosci-ence, systems neuroscience, behavioral ecology, and socialpsychology. Today, the field of social neuroscience fills thisgap with abundance: social neuroscience now has its own jour-nals, textbooks, societies, and, according to PubMed, nearly3000 research papers (as of February 22, 2010). Much of thisstunning growth has been driven by human neuroimagingstudies seeking the neural correlates of psychological pro-cesses, from face perception to social preferences. Socialneuroscience has a different foundation in animal studies, builton molecular and cellular approaches as well as the tools ofsystems neuroscience. In fact, the history of animal studies ofsocial perception and social behavior with classical lesion andneurophysiological techniques extends back several decadesin the venerable literatures of neuroethology and behavioralneuroendocrinology. This review will follow a single thread ofsocial neuroscience spun from this older animal research,recently woven into human studies and now suggesting potentialtreatments of human disorders of social behavior, such asautism.
Social Neuroscience in 2010Most of social neuroscience can be separated into studies ofeither receptive or expressive processes. Receptive studies,which emerged from neuroethology, focus on sensory process-ing. From the elegant work on pheromone receptors in mice(Dulac and Torello, 2003) to the careful mapping of face cells inhuman and nonhuman primates (Kanwisher, 2006), this arm ofsocial neuroscience has described the neural geography and,in some cases, the cellular landscape by which sensory informa-tion is initially encoded as social. A fundamental insight fromreceptive studies is that, in most vertebrates, the brain employsspecific receptors or cortical regions for processing social
information, whether that information is from pheromonal/olfac-tory, audio-vocal, somatosensory, or visual cues. That is, socialinformation is not simply complex multisensory perception; it isperceived and encoded in unique ways in the brain.Expressive studies, long the domain of behavioral neurosci-
ence and behavioral neuroendocrinology, focus on socialinteractions: communication, reproductive behavior (especiallyparental care and sex), agonistic actions (aggression and preda-tion), and affiliative behaviors (including social play). In verte-brates (and many invertebrates), nearly all of these behaviorsare influenced by gonadal steroids (estrogens and androgens),acting via their nuclear receptors. The mapping of gonadalsteroid receptor expression in the brain helped identify keyregions for social behavior (McEwen et al., 1979; Pfaff andKeiner, 1973; Pfaff and McEwen, 1983). Additionally, steroidreceptors are transcription factors, and by identifying the genesregulated by steroid receptors, a molecular basis for socialbehavior could be proposed (Warnmark et al., 2003).Between the receptive and expressive arms sits the great dark
matter of social neuroscience. (Figure 1) What happens betweenthe stage when a percept is encoded as ‘‘social’’ to the stagewhen a ‘‘go’’ signal is given for initiating social behavior? Howdoes the brain distinguish prey from predator, juvenile fromadult, novel from familiar, kin from unrelated conspecific? Whatare the neural mechanisms that facilitate or inhibit social interac-tion? These are the questions that have been more difficult toanswer.Recently, researchers have begun to address these questions
with human neuroimaging studies seeking to map variousaspects of higher-order processing of social information andeven develop computational principles for the neural basis ofsocial cognition (Behrens et al., 2009). Human neuroimagingstudies can describe the cortical patterns, but mechanisticstudies are still mostly the domain of animal research. In fact,
768 Neuron 65, March 25, 2010 ª2010 Elsevier Inc.
13. E. D. Jarvis, C. Scharff, M. R. Grossman, J. A. Ramos,F. Nottebohm, Neuron 21, 775 (1998).
14. S. S. Burmeister, E. D. Jarvis, R. D. Fernald, PloS Biol. 3,e363 (2005).
15. L. Grosenick, T. S. Clement, R. D. Fernald, Nature 445,429 (2007).
16. K. J. O'Donovan, W. G. Tourtellotte, J. Milbrandt,J. M. Baraban, Trends Neurosci. 22, 167 (1999).
17. C. W. Whitfield, A.-M. Cziko, G. E. Robinson, Science 302,296 (2003).
18. C. M. Grozinger, N. Sharabash, C. W. Whitfield,G. E. Robinson, Proc. Natl. Acad. Sci. U.S.A. 100, 14519(2003).
19. S. A. Ament, M. Corona, H. S. Pollack, G. E. Robinson,Proc. Natl. Acad. Sci. U.S.A. 105, 4226 (2008).
20. M. E. Cummings et al., Proc. R. Soc. London Ser.B. Biol. Sci. 275, 393 (2008).
21. K. Replogle et al., BMC Genomics 9, 131 (2008).22. H. Liu, M. M. Wu, H. H. Zakon, Dev. Neurobiol. 67, 1289
(2007).23. C. Cirelli, C. M. Gutierrez, G. Tononi, Neuron 41, 35
(2004).
24. F. A. Champagne, D. D. Francis, A. Mar, M. J. Meaney,Physiol. Behav. 79, 359 (2003).
25. I. C. Weaver et al., Nat. Neurosci. 7, 847 (2004).26. D. L. Champagne et al., J. Neurosci. 28, 6037 (2008).27. D. W. Pfaff, Drive (MIT Press, Cambridge, MA, 1999).28. D. S. Falconer, T. F. C. Mackay, Introduction to
Quantitative Genetics (Longmans Green, Harlow Essex,UK, ed. 4, 1996).
29. A. Caspi, T. E. Moffitt, Nat. Rev. Neurosci. 7, 583 (2006).30. D. K. Lahiri, B. Maloney, Nat. Rev. Neurosci. 10.1038/
nrn2022-c1 (2006).31. A. Abbott, Nature 454, 154 (2008).32. J.Wang, K. G. Ross, L. Keller, PLoSGenet.4, e1000127 (2008).33. B. J. Dickson, Science 322, 904 (2008).34. D. A. Wheeler et al., Science 251, 1082 (1991).35. S. Fink, L. Excoffier, G. Heckel, Proc. Natl. Acad. Sci. U.S.A.
103, 10956 (2006).36. Y. Ben-Shahar, A. Robichon, M. B. Sokolowski, G. E.
Robinson, Science 296, 741 (2002).37. A. L. Toth, G. E. Robinson, Trends Genet. 23, 334 (2007).38. W. Enard et al., Nature 418, 869 (2002).39. W. Shu et al., Proc. Natl. Acad. Sci. U.S.A. 102, 9643 (2005).
40. S. Haesler et al., PLoS Biol. 5, e321 (2007).41. M. Groszer et al., Curr. Biol. 18, 354 (2008).42. J. Sebat et al., Science 316, 445 (2007), published
online 14 March 2007; 10.1126/science.1138659.43. J. D. Jensen, A. Wong, C. F. Aquadro, Trends Genet. 23,
568 (2007).44. J. C. Hall, J. Neurogenet. 17, 1 (2003).45. We thank J. Desjardins, A. Fernald, K. A. Hughes,
D. B. Kelley, K. Maruska, C. Olin, M. B. Sokolowski,L. J. Stubbs, members of the Clayton and Robinsonlaboratories, and two anonymous reviewers for reviewsof this manuscript and C. Harrell for graphical assistance.Research by the authors cited here was supported bythe following grants: NIH R01 NS051820 and NS045264(D.F.C.); NIH NS34950 Javits Award and UnitedStates–Israel Binational Science Foundation 200596(R.D.F.); and NIH R01 GM073644, NSF Frontiers inBiological Research EF04-25852, U.S. Department ofAgriculture AG2003-35302-13490, and a BurroughsWellcome Fund Innovation Award (G.E.R.).
10.1126/science.1159277
REVIEW
Oxytocin, Vasopressin, and theNeurogenetics of SocialityZoe R. Donaldson1 and Larry J. Young1,2*
There is growing evidence that the neuropeptides oxytocin and vasopressin modulate complexsocial behavior and social cognition. These ancient neuropeptides display a marked conservation ingene structure and expression, yet diversity in the genetic regulation of their receptors seems tounderlie natural variation in social behavior, both between and within species. Human studies arebeginning to explore the roles of these neuropeptides in social cognition and behavior and suggestthat variation in the genes encoding their receptors may contribute to variation in human socialbehavior by altering brain function. Understanding the neurobiology and neurogenetics of socialcognition and behavior has important implications, both clinically and for society.
Social interactions affect every aspect ofour lives, from wooing a mate and caringfor our children to determining our suc-
cess in the workplace. Abnormal manifestationsof social behavior, such as the pathologicaltrusting associated with Williams-Beuren Syn-drome (1), social withdrawal in depression, anddecreased social cognition in autism, profoundlyaffect the lives of those who suffer from thesedisorders. Neuroscientists once considered socialbehavior to be too hopelessly complex to under-stand at a mechanistic level, but advances in ani-mal models of social cognition and bonding, aswell as application of new technologies in humanresearch have demonstrated that the molecularbasis of social behavior is not beyond the realmof our understanding. There appears to be markedconservation in the molecular mechanisms regulat-
ing social behavior across diverse species, includ-ing our own.
Interacting with other neurotransmitter systemswithin specific neural circuits, neuropeptides haveemerged as central players in the regulation of socialcognition and behavior. Neuropeptides may act asneurotransmitters, if released within synapses, or asneurohormones, activating receptors distant fromthe site of release, which provides evolutionaryflexibility to their actions (2). Within vertebrates, amajority of work relating neuropeptides to socialbehavior has focused on members of the oxytocin/vasopressin family. Homologs of oxytocin andvasopressin existed at least 700 million years agoand have been identified in such diverse organismsas hydra, worms, insects, and vertebrates. Amongthese distant taxa, oxytocin- and vasopressin-relatedpeptides play a general role in the modulation ofsocial and reproductive behaviors. In contrast tothis apparent conservation in function, the spe-cific behaviors affected by these neuropeptidesare notably species-specific.
Only recently have scientists begun to dissectthe roles of oxytocin, vasopressin, and their re-
lated receptors in human social behavior. Whereashuman social behavior is more nuanced and com-plex than the behaviors typically assayed in otheranimals, this complexity has created unique op-portunities to design finely honed tasks that haverevealed a potential role for these peptides in per-sonality, trust, altruism, social bonding, and ourability to infer the emotional state of others. Here,we review the evidence of evolutionary conserva-tionwithin the vasopressin/oxytocin peptide fam-ily, briefly discuss the role of these peptides andtheir respective receptors in modulating socialbehavior and bonding, and provide a synthesisof recent advances implicating the oxytocin andvasopressin systems in human trust, cooperation,and social behavior.
Conservation of Neuropeptide SystemsRegulating Social BehaviorThe mammalian oxytocin and vasopressin non-apeptides, so called for their nine–amino acidcomposition, differ from each other at only twoamino acid positions (Fig. 1). Oxytocin, vasopres-sin, and their respective nonmammalian vertebratelineages are thought to have arisen from a gene-duplication event before vertebrate divergence.Within these lineages, peptides vary by a singleamino acid, and their genes are found near eachother on the same chromosome. Invertebrates, withfew exceptions, have only one oxytocin/vasopressinhomolog, whereas vertebrates have two (3, 4).
In mammals, oxytocin and vasopressin are pro-duced primarily within hypothalamic brain regionsand then shuttled to the pituitary for peripheralrelease or projected to various brain regions.Notably,just as oxytocin andvasopressin are expressedwithinthe hypothalamus of mammals, their homologs areexpressed within similar neurosecretory brainregions of organisms as diverse as worms and fish.A characterization of annepressin (the homolog ofoxytocin/vasopressin in segmented worms) andvasotocin (vasopressin’s counterpart in bony fish)revealed conserved neural expression of these genes
1Center for Behavioral Neuroscience, Yerkes Regional PrimateResearch Center, Emory University, Atlanta, GA 30322, USA.2Department of Psychiatry and Behavioral Sciences, EmoryUniversity, Atlanta, GA 30329, USA.
*To whom correspondence should be addressed. E-mail:[email protected]
7 NOVEMBER 2008 VOL 322 SCIENCE www.sciencemag.org900
Genetics of Behavior
CORRECTED 13 MARCH 2009; SEE LAST PAGE
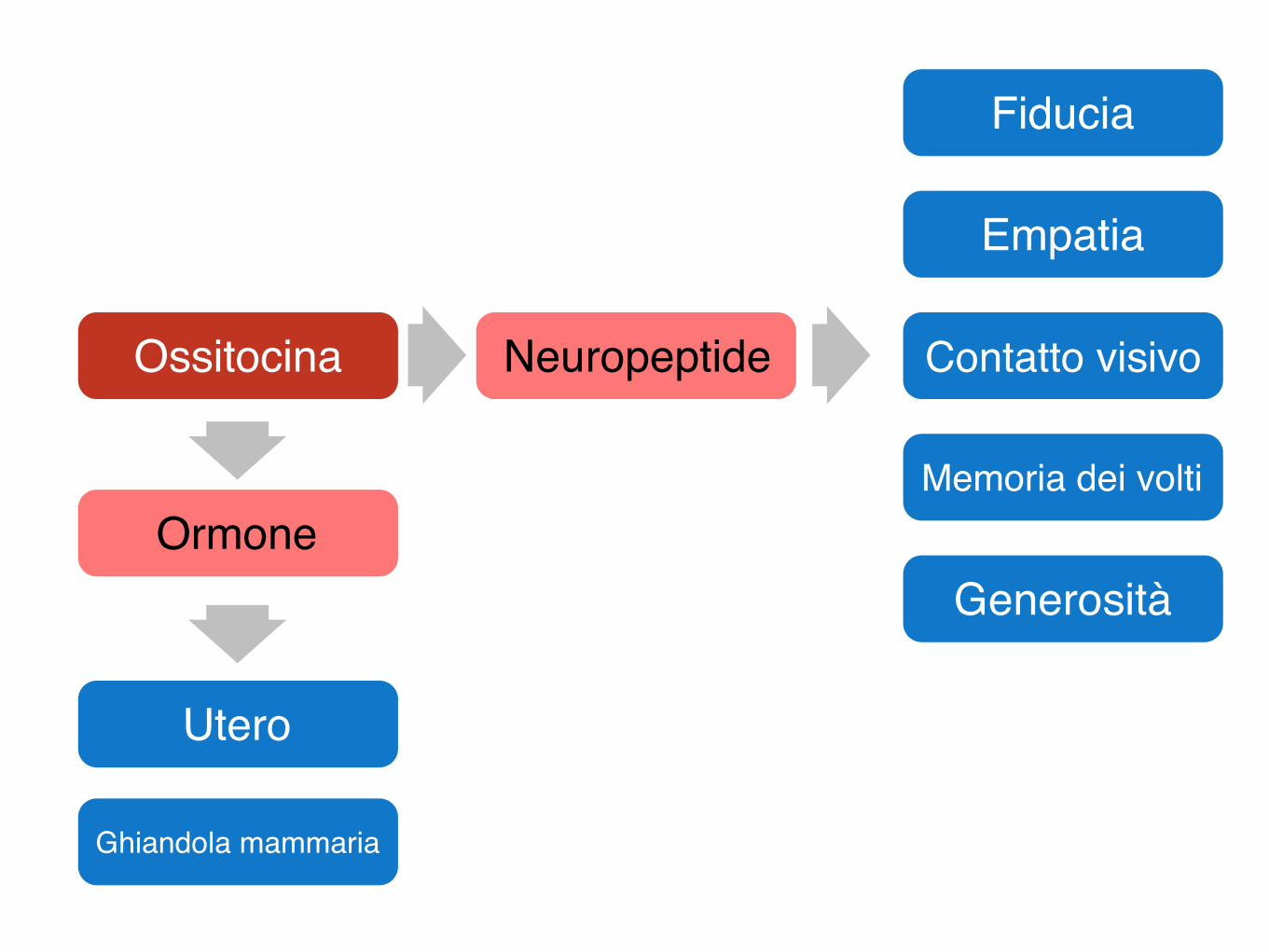
Ossitocina
Utero
Ghiandola mammaria
Fiducia
Empatia
Contatto visivo
Memoria dei volti
GenerositàOrmone
Neuropeptide

Adenoipofisi
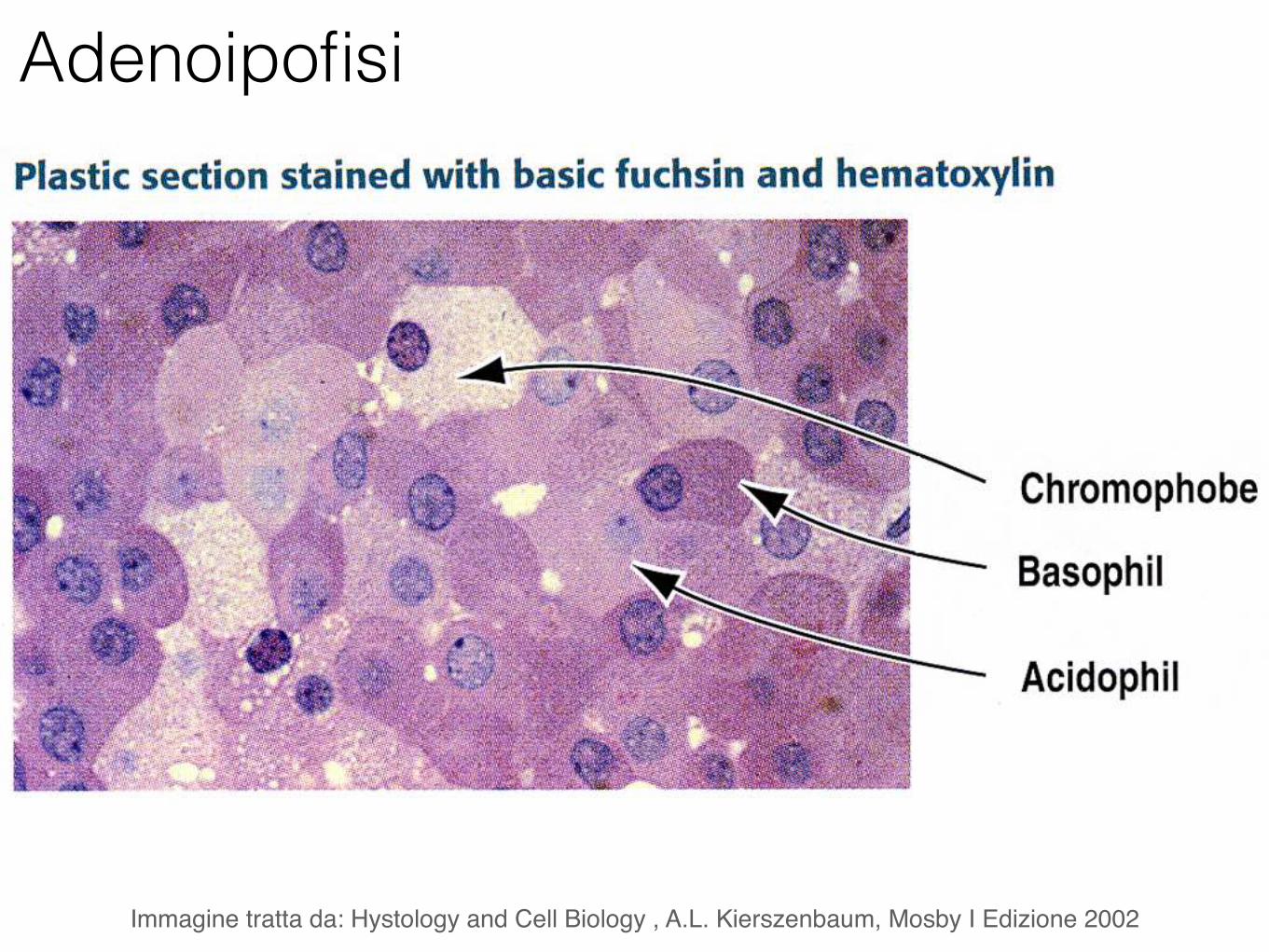
Adenoipofisi
Immagine tratta da: Hystology and Cell Biology , A.L. Kierszenbaum, Mosby I Edizione 2002
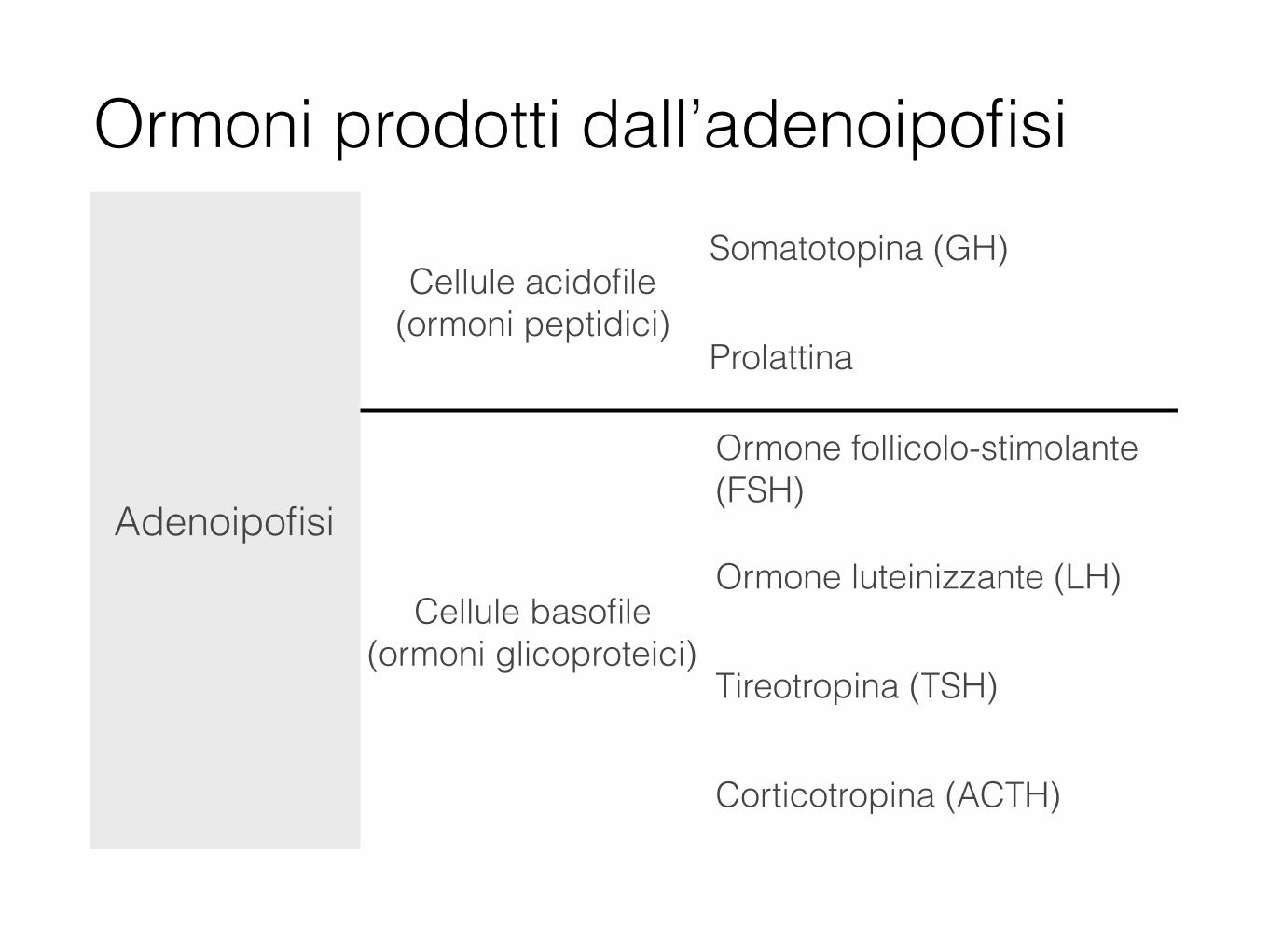
Adenoipofisi
Cellule acidofile (ormoni peptidici)
Somatotopina (GH)
Prolattina
Cellule basofile (ormoni glicoproteici)
Ormone follicolo-stimolante (FSH)
Ormone luteinizzante (LH)
Tireotropina (TSH)
Corticotropina (ACTH)
Ormoni prodotti dall’adenoipofisi
Imm
agin
e tra
tta d
a: T
he H
uman
Bra
in, J
. Nol
te, M
osby
V E
dizio
ne 2
002
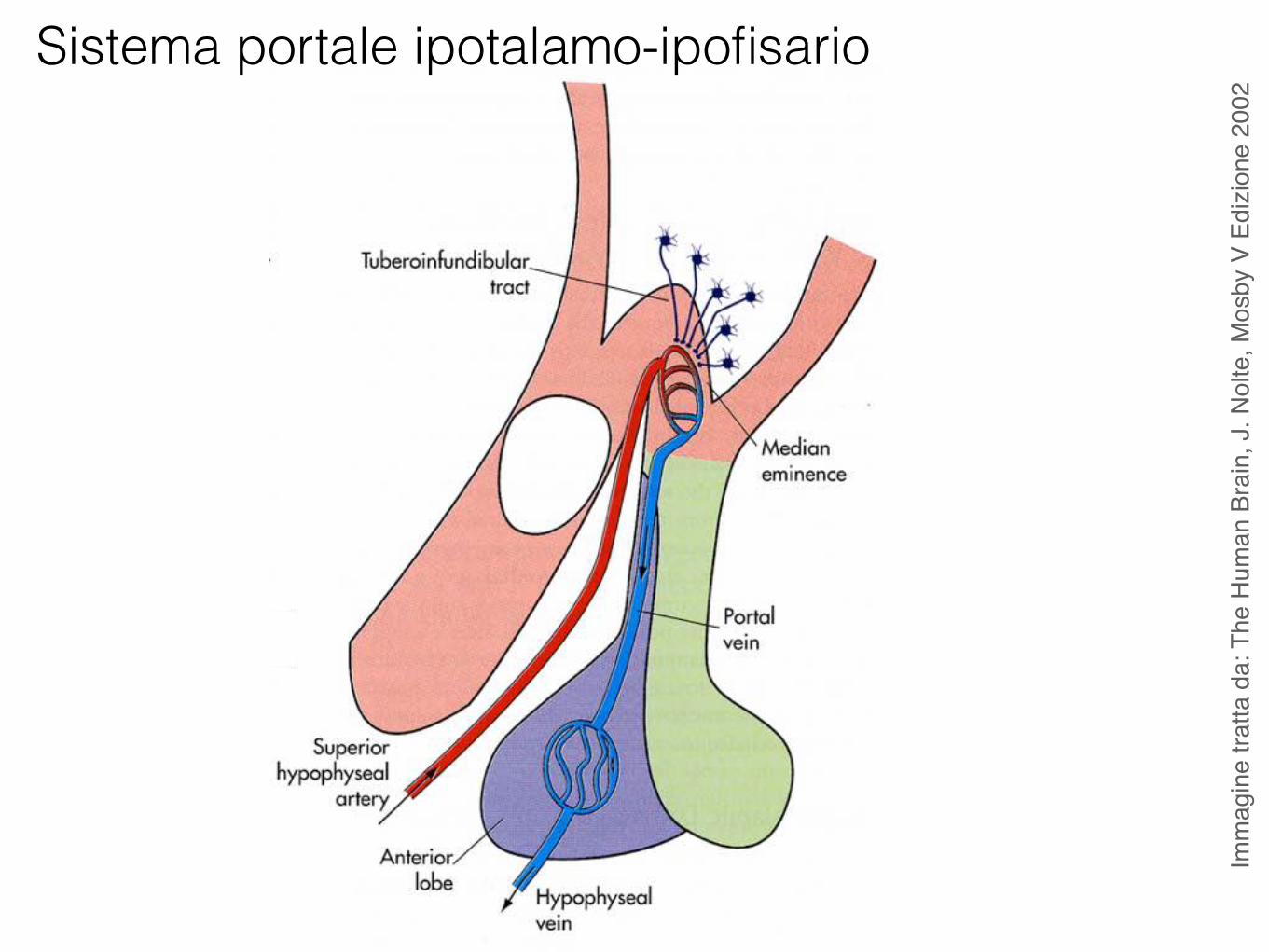
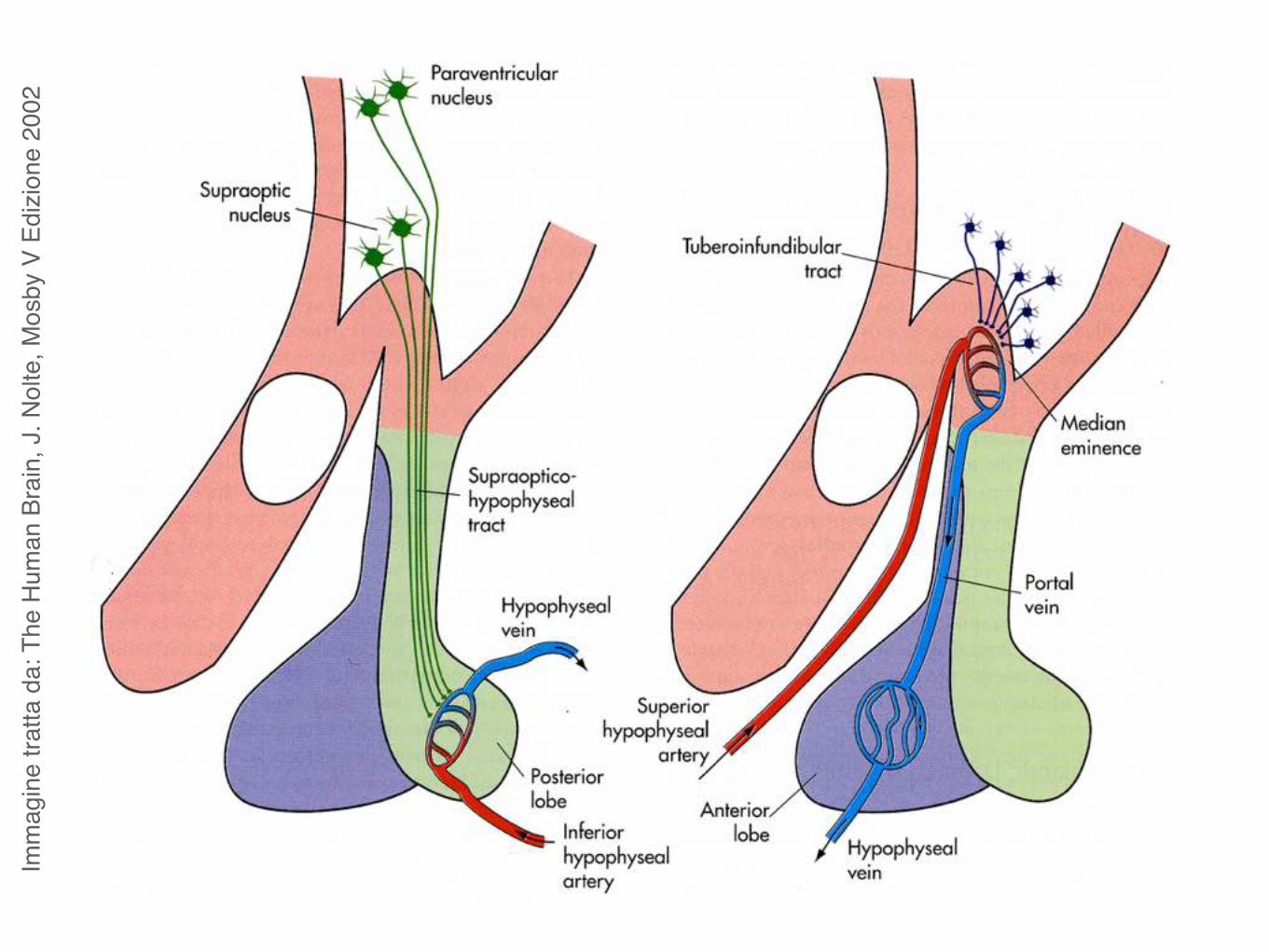
Sistema portale ipotalamo-ipofisario
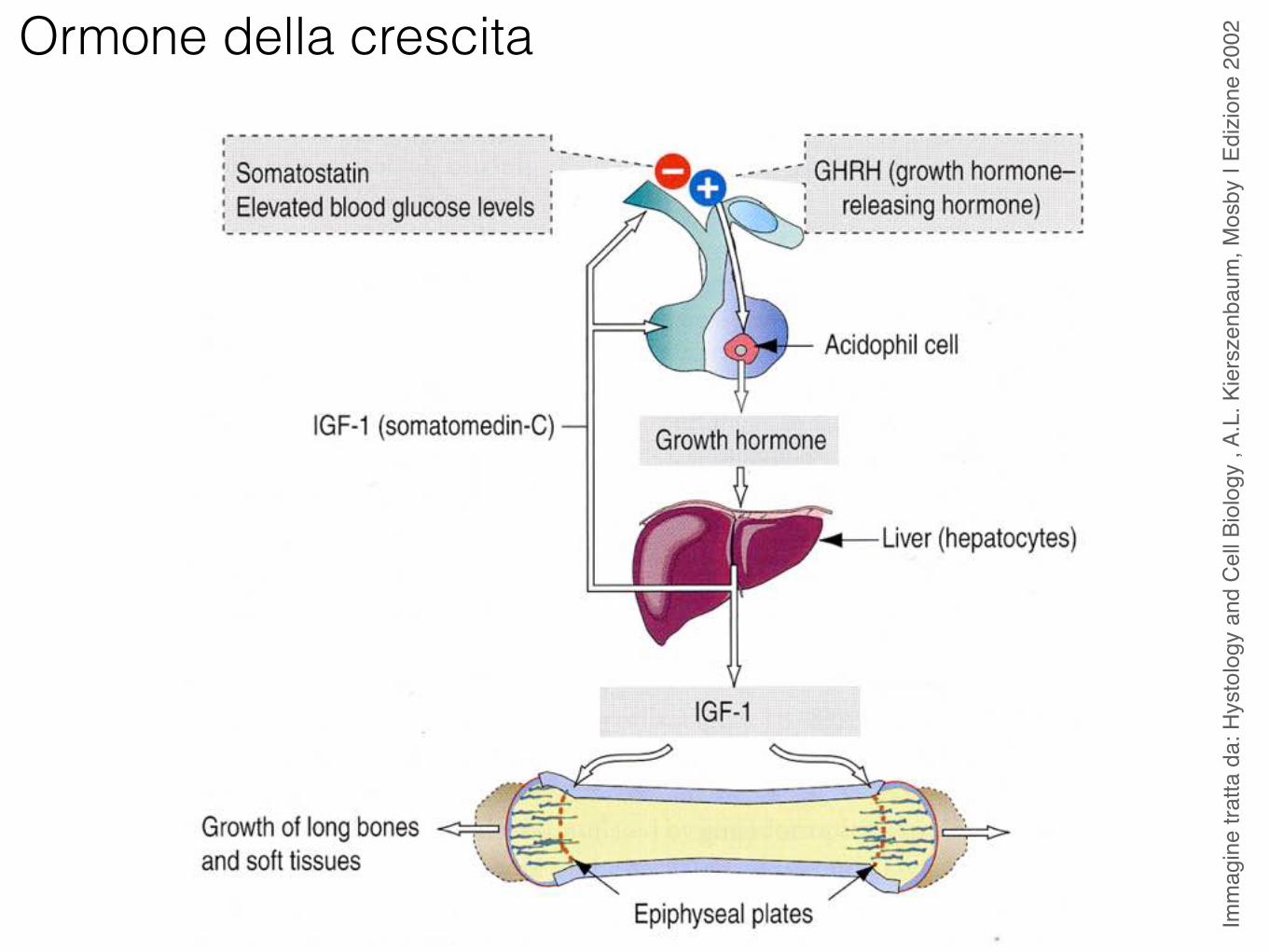
Ormone della crescita
Imm
agin
e tra
tta d
a: H
ysto
logy
and
Cel
l Bio
logy
, A.
L. K
iers
zenb
aum
, Mos
by I
Edizi
one
2002
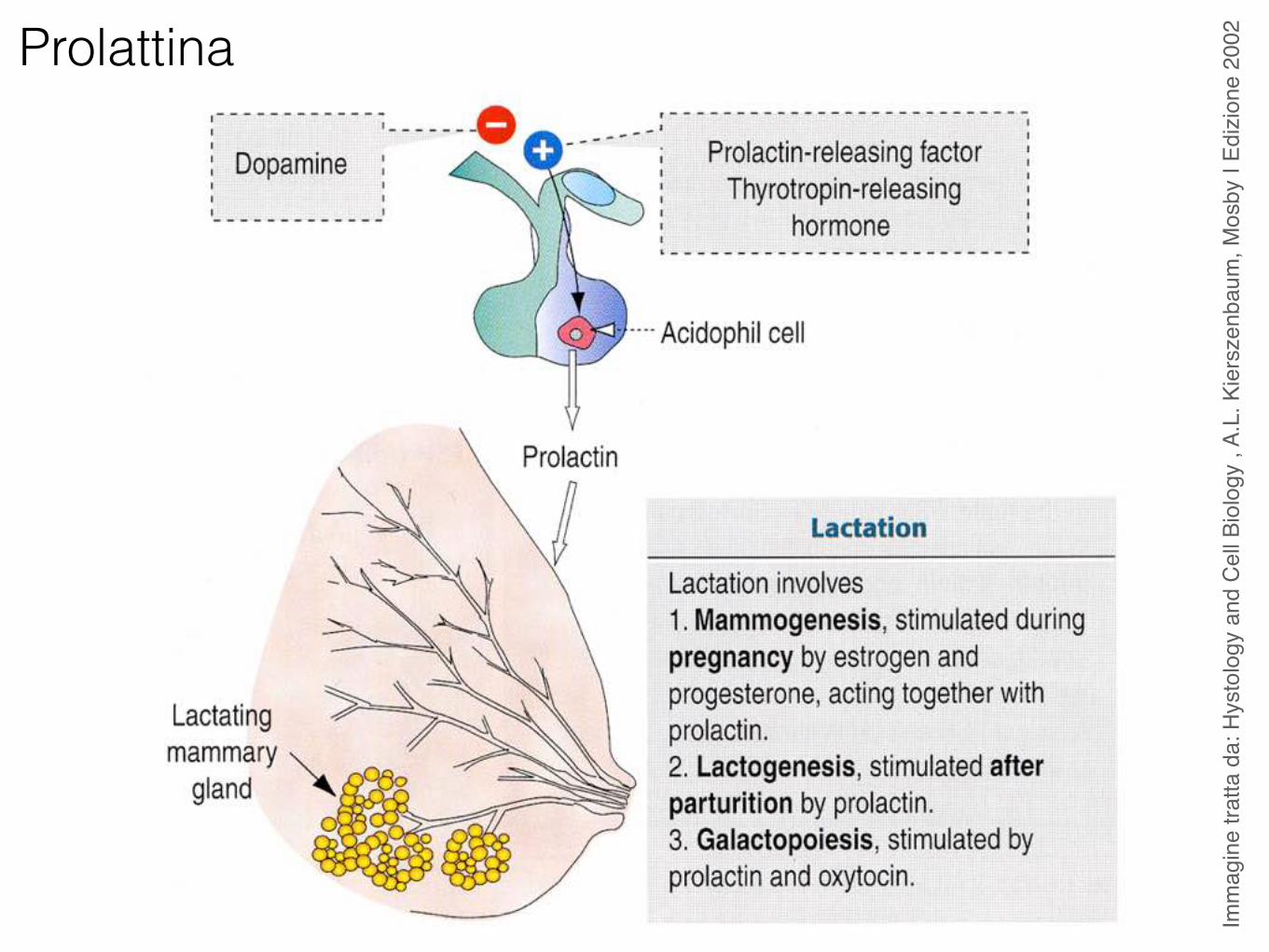
Prolattina
Imm
agin
e tra
tta d
a: H
ysto
logy
and
Cel
l Bio
logy
, A.
L. K
iers
zenb
aum
, Mos
by I
Edizi
one
2002
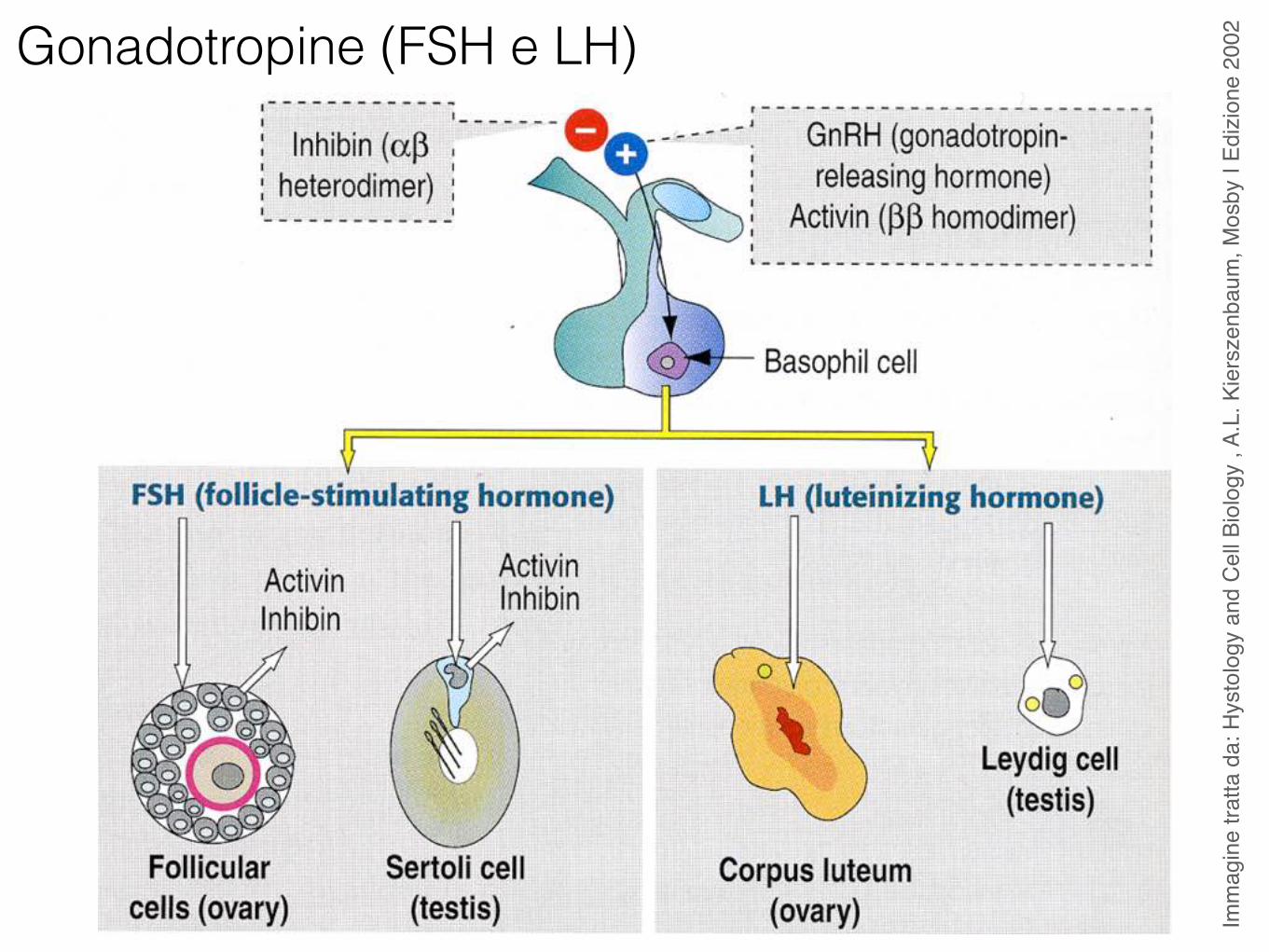
Gonadotropine (FSH e LH)
Imm
agin
e tra
tta d
a: H
ysto
logy
and
Cel
l Bio
logy
, A.
L. K
iers
zenb
aum
, Mos
by I
Edizi
one
2002
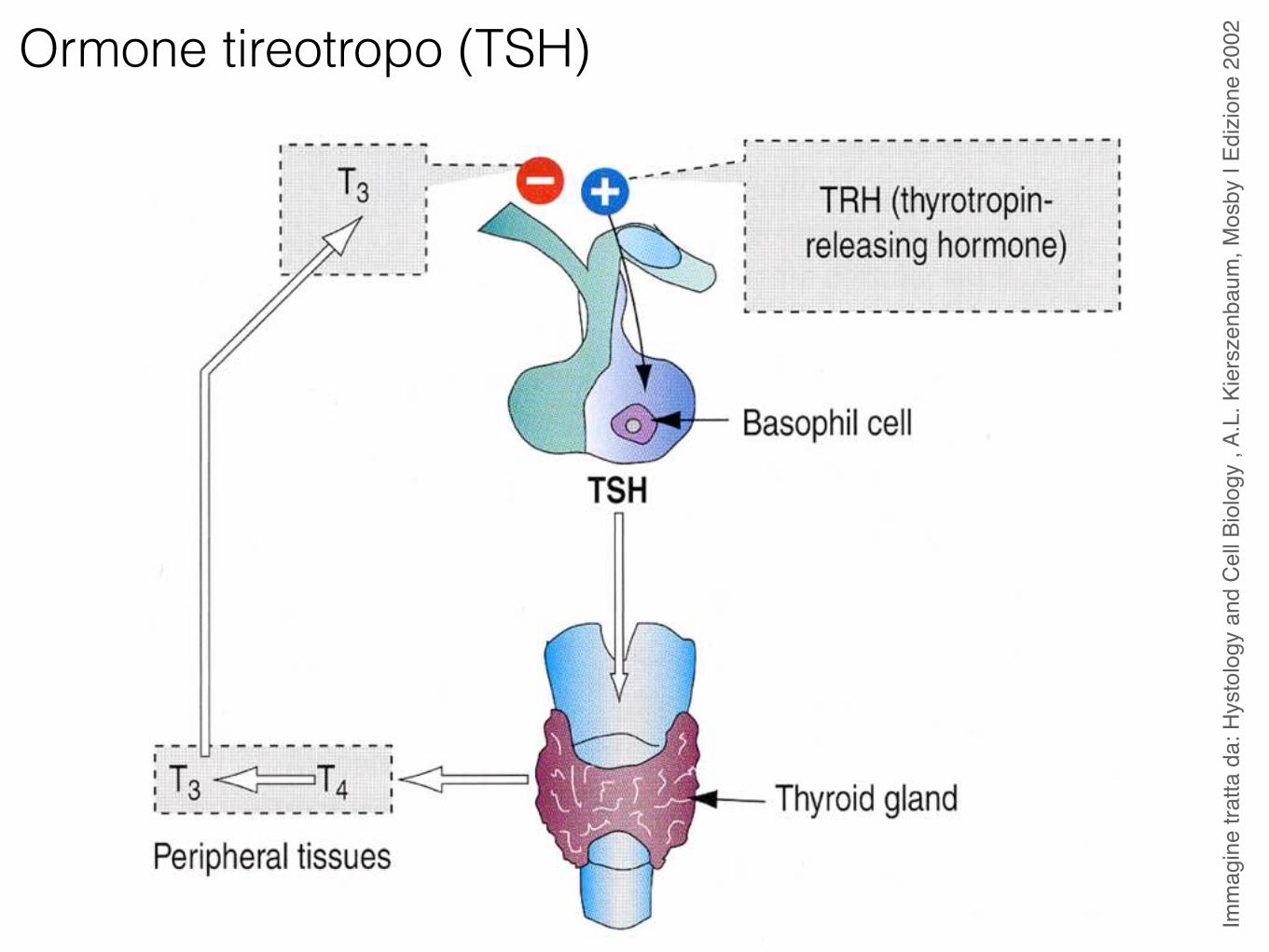
Ormone tireotropo (TSH)
Imm
agin
e tra
tta d
a: H
ysto
logy
and
Cel
l Bio
logy
, A.
L. K
iers
zenb
aum
, Mos
by I
Edizi
one
2002
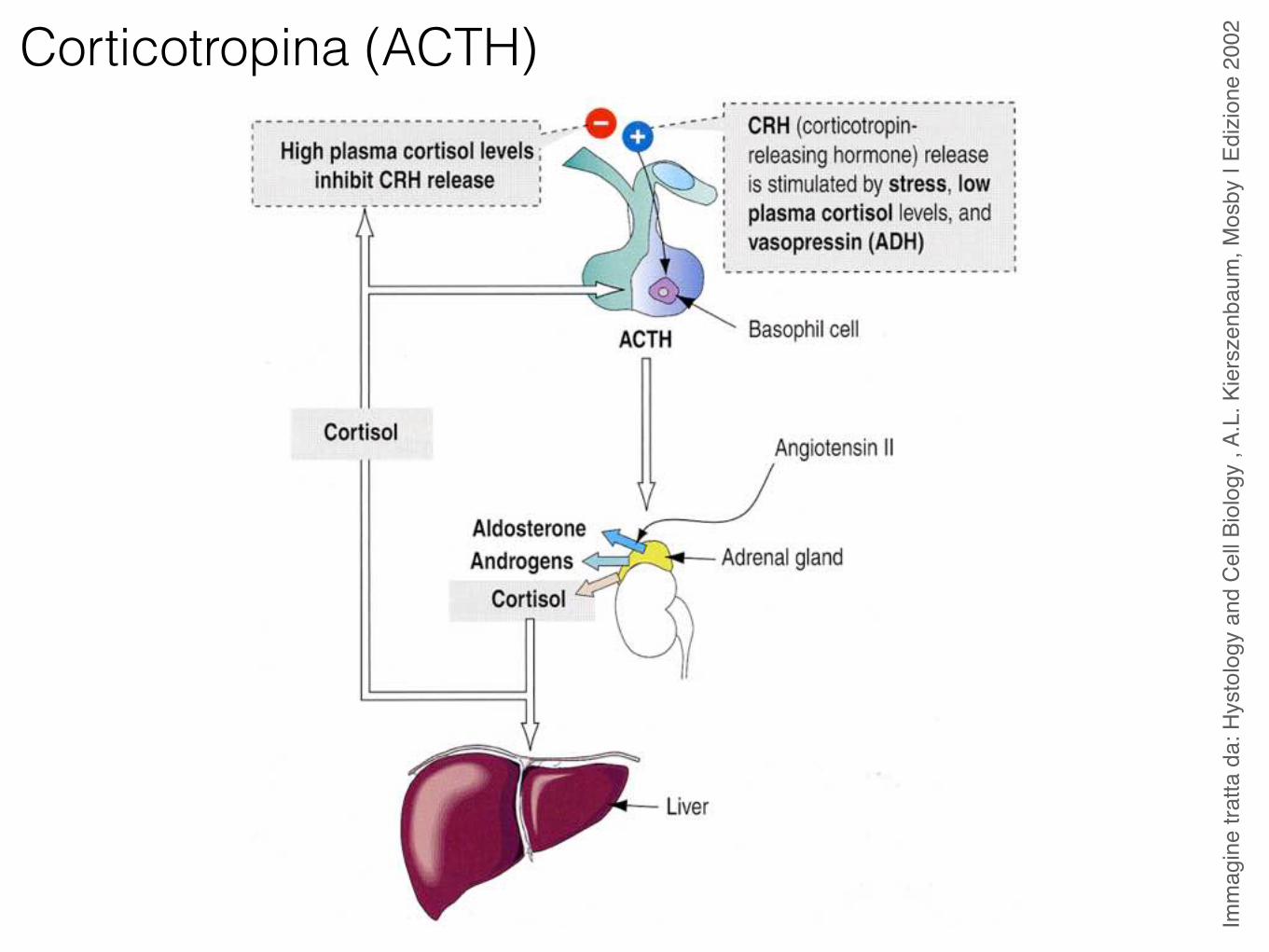
Corticotropina (ACTH)
Imm
agin
e tra
tta d
a: H
ysto
logy
and
Cel
l Bio
logy
, A.
L. K
iers
zenb
aum
, Mos
by I
Edizi
one
2002
Imm
agin
e tra
tta d
a: T
he H
uman
Bra
in, J
. Nol
te, M
osby
V E
dizio
ne 2
002
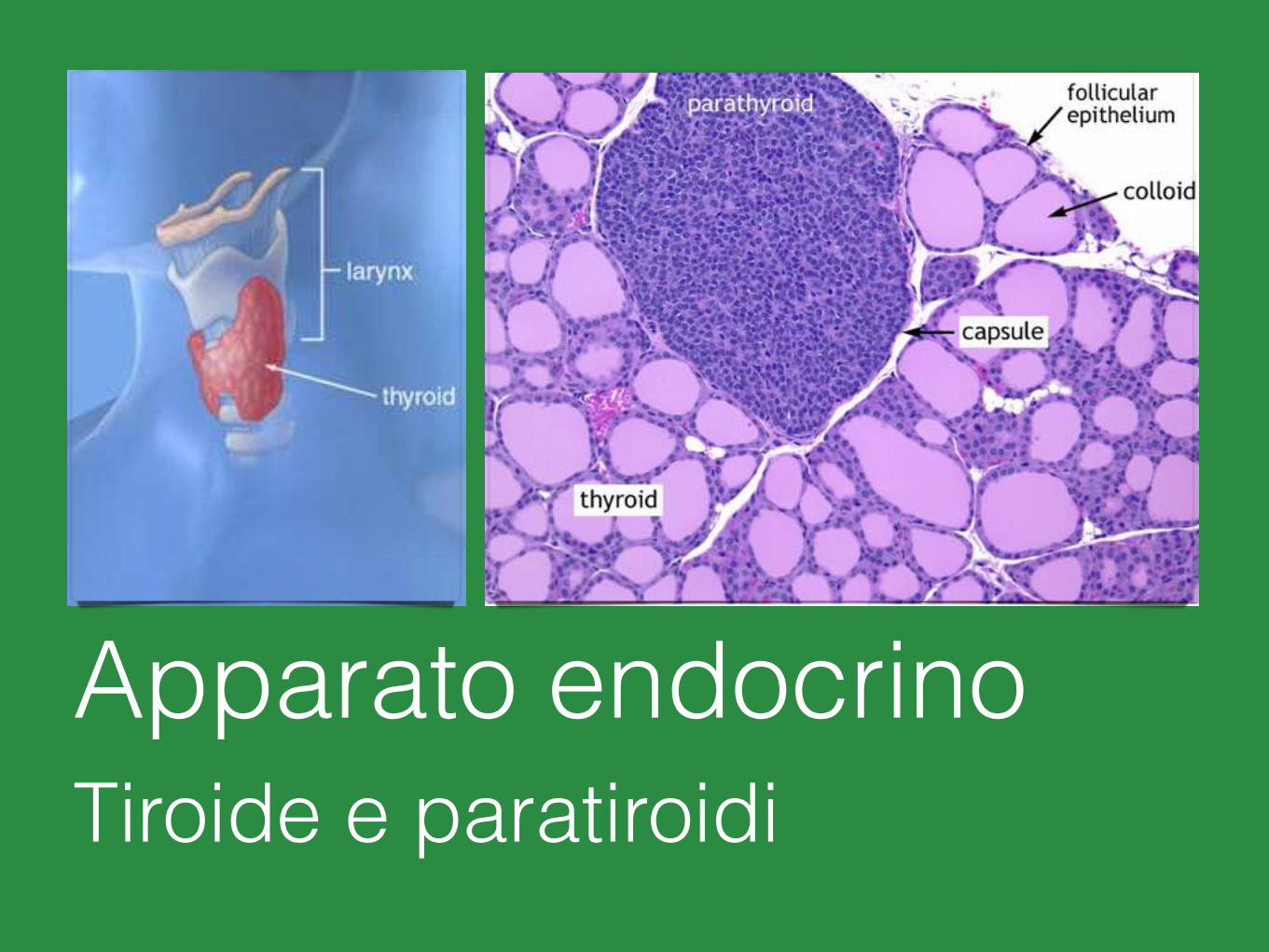
Apparato endocrinoTiroide e paratiroidi
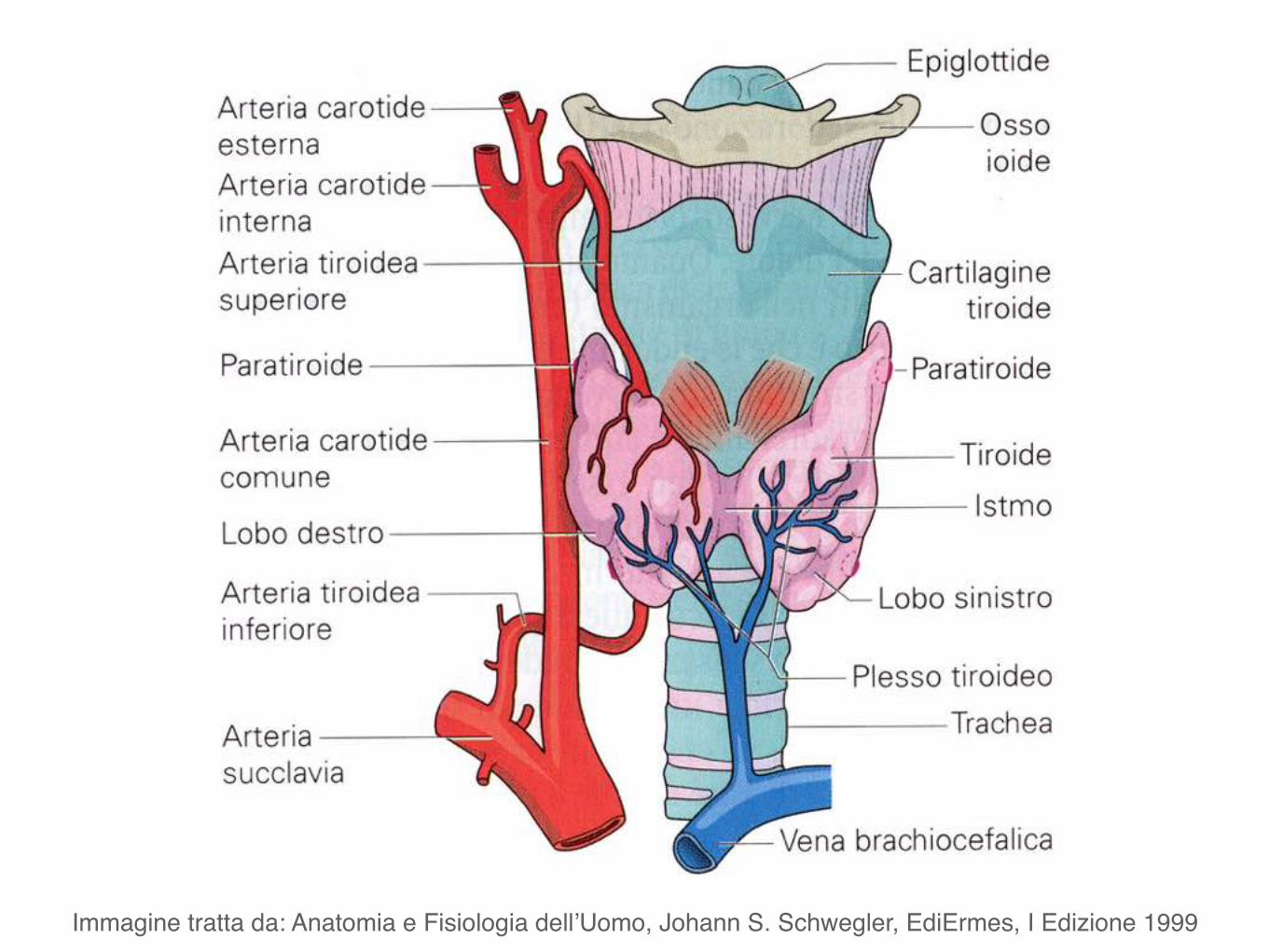
Immagine tratta da: Anatomia e Fisiologia dell’Uomo, Johann S. Schwegler, EdiErmes, I Edizione 1999
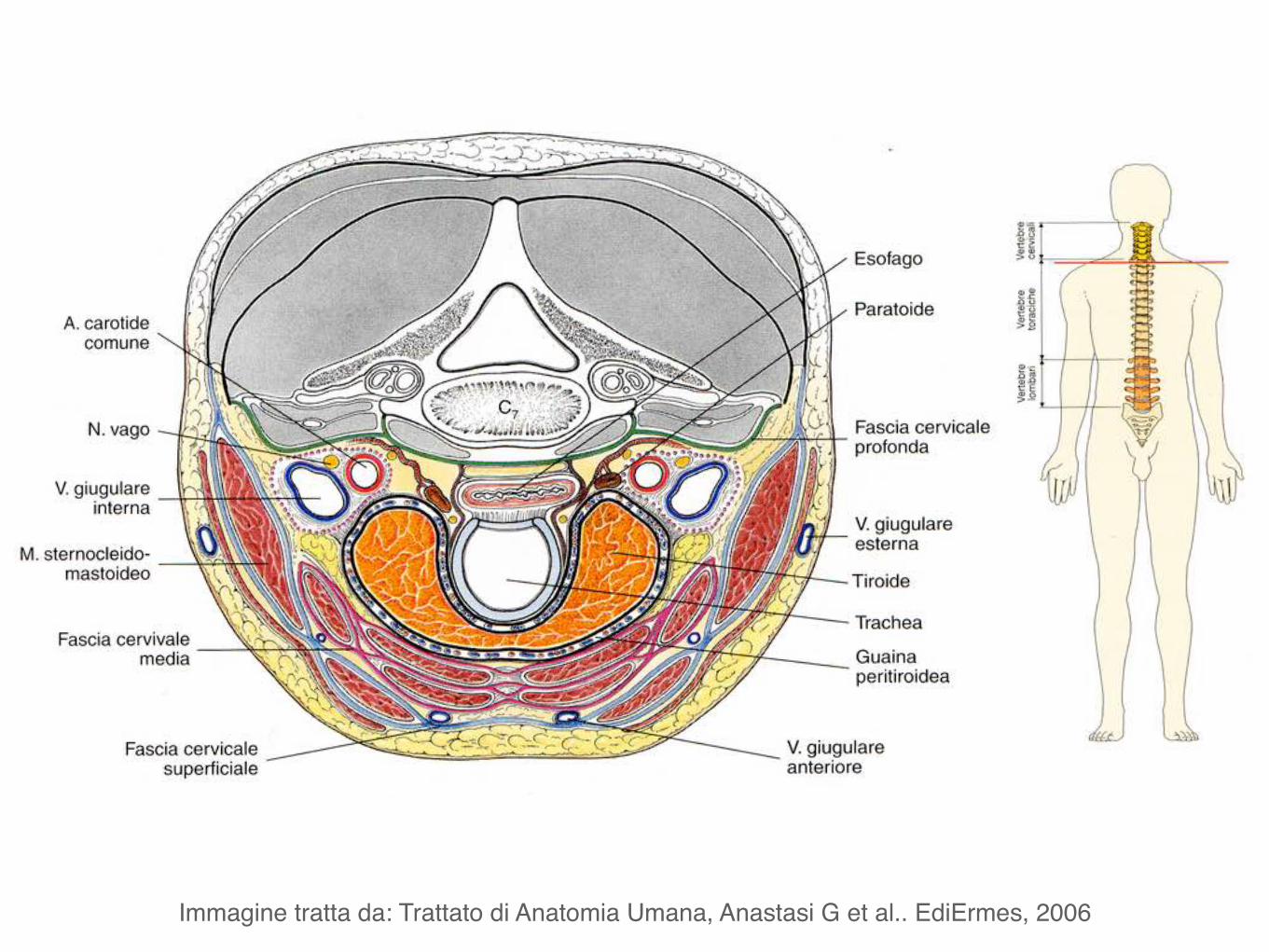
Immagine tratta da: Trattato di Anatomia Umana, Anastasi G et al.. EdiErmes, 2006

Microscopicamente i due lobi della tiroide sono costituiti da follicoli.
I follicoli, in numero di parecchie migliaia nell’uomo, sono di dimensione variabile e sono rivestiti da un singolo strato di cellule appiattite, cuboidi o colonnari, i tireociti.
Nel tessuto connettivo tra i follicoli si trovano le cellule parafollicolari o cellule C.
Struttura della tiroide
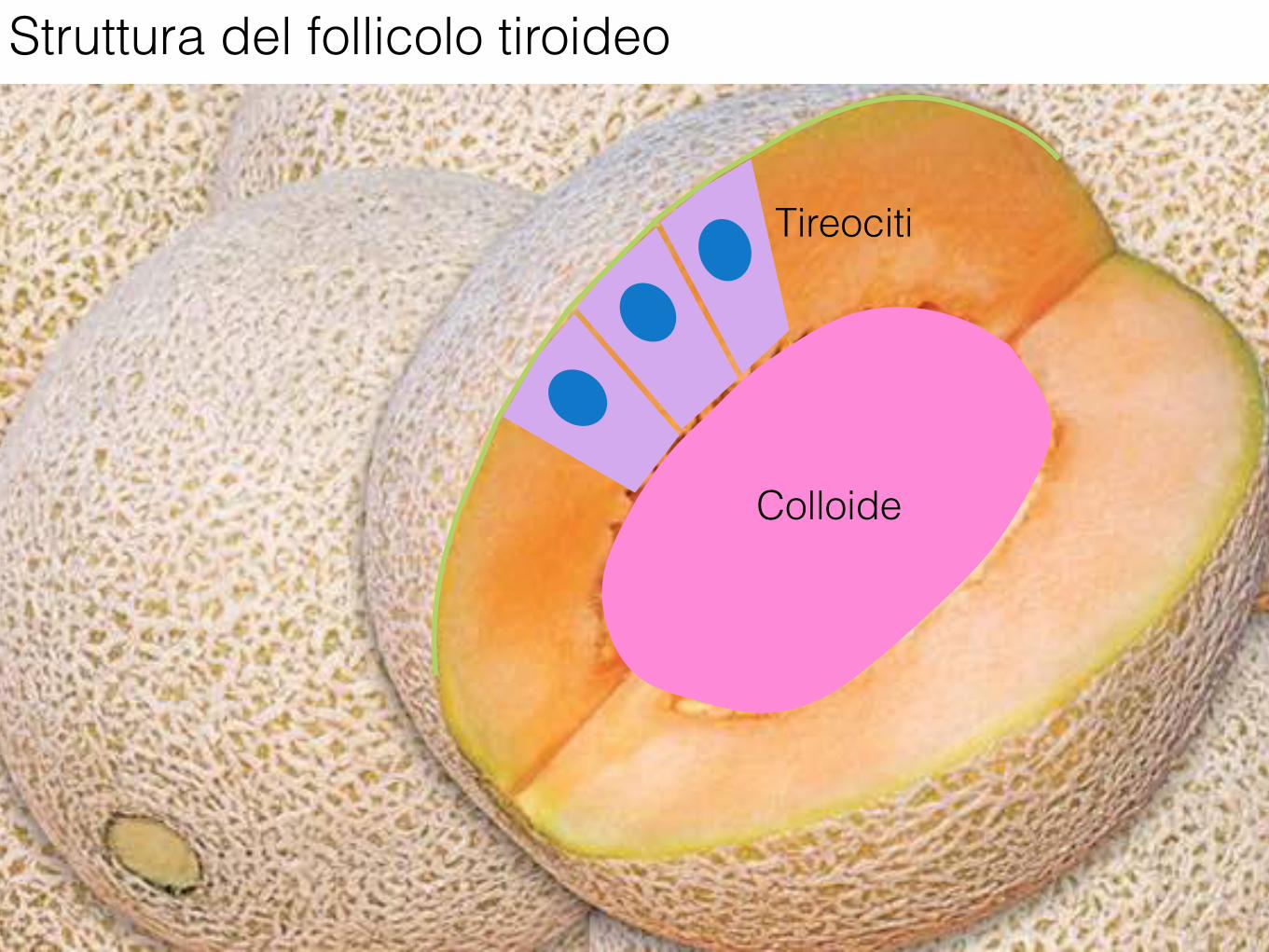
Struttura del follicolo tiroideo
Tireociti
Colloide

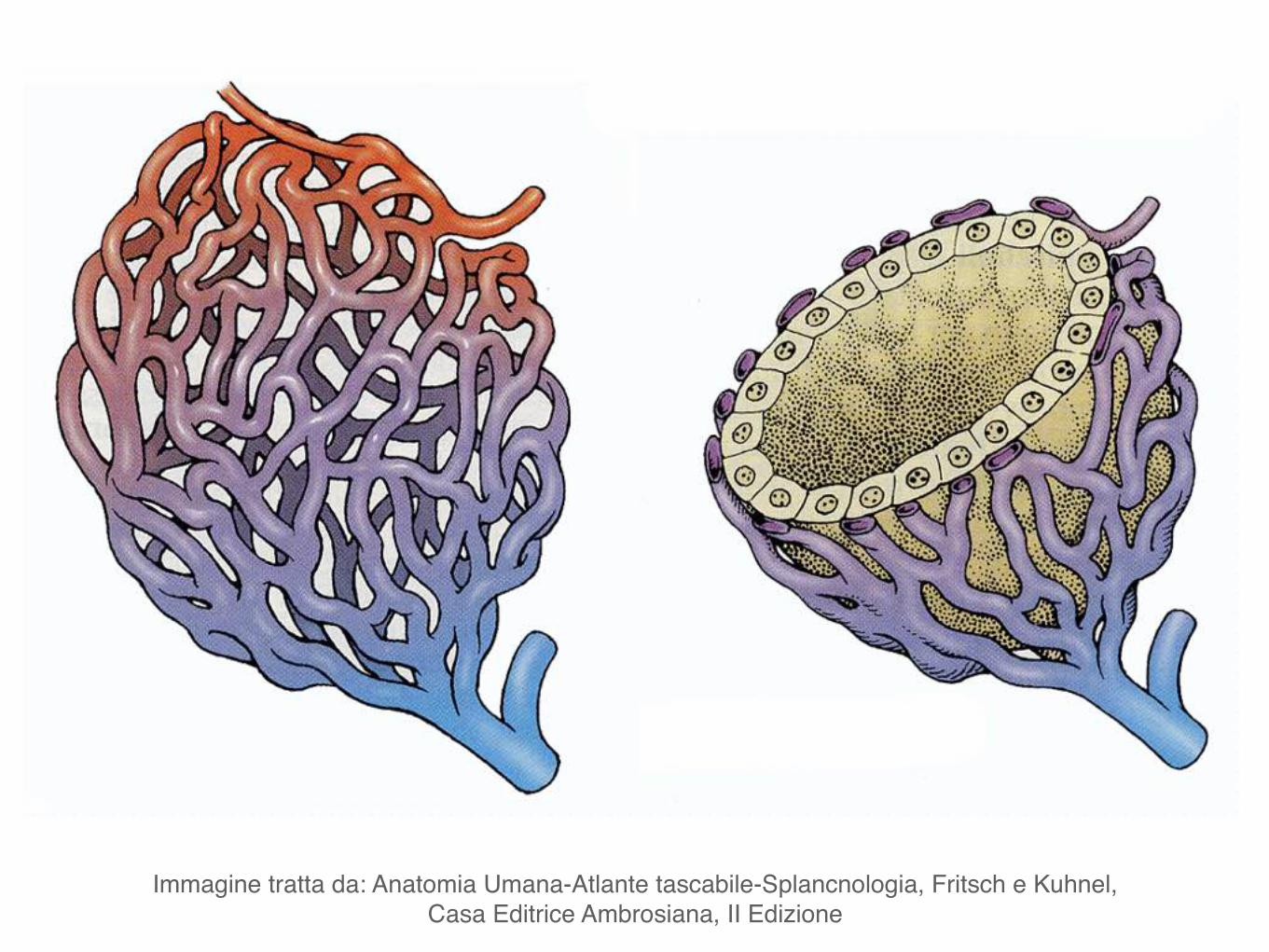
Immagine tratta da: Anatomia Umana-Atlante tascabile-Splancnologia, Fritsch e Kuhnel, Casa Editrice Ambrosiana, II Edizione

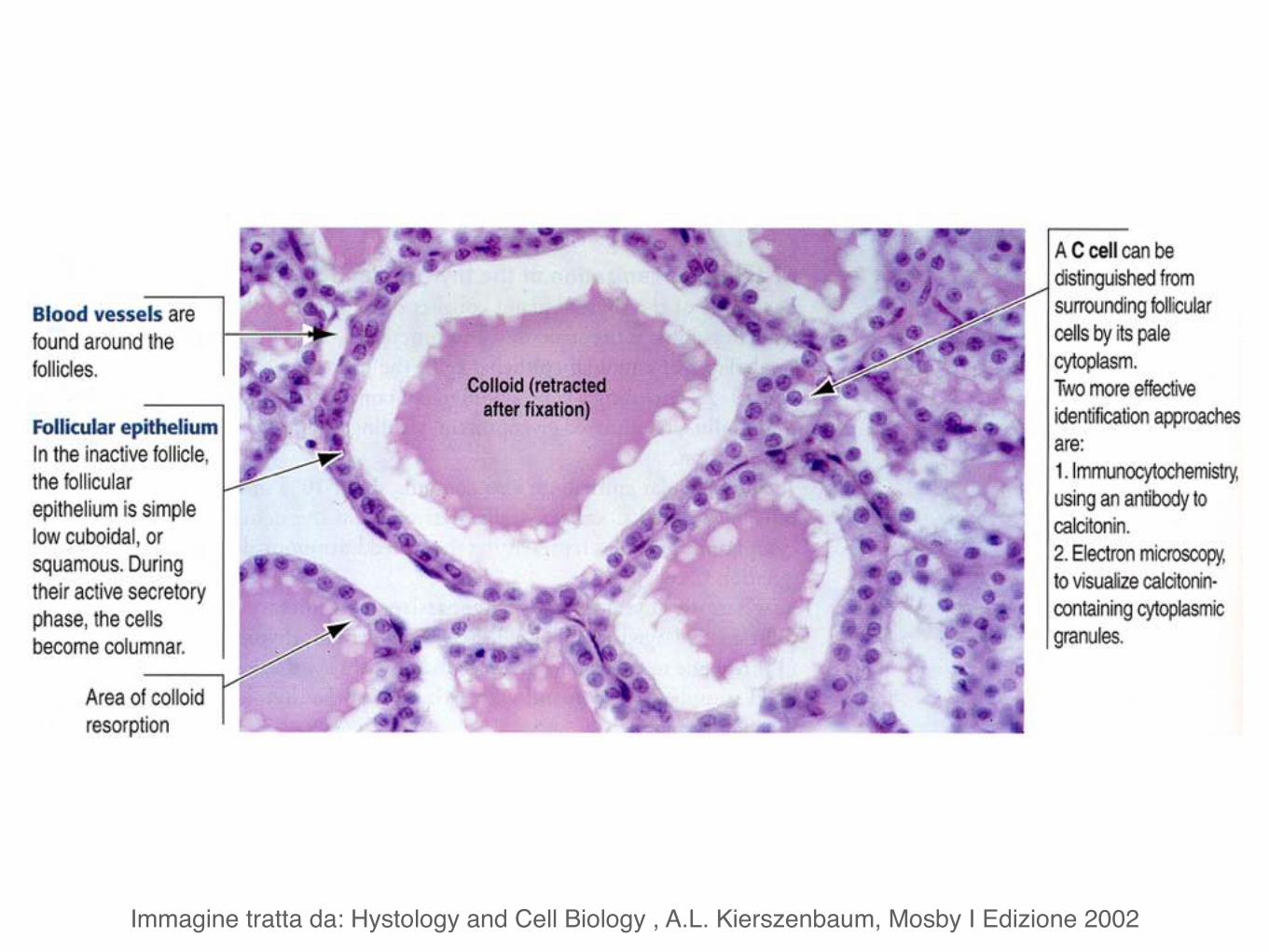
Immagine tratta da: Hystology and Cell Biology , A.L. Kierszenbaum, Mosby I Edizione 2002
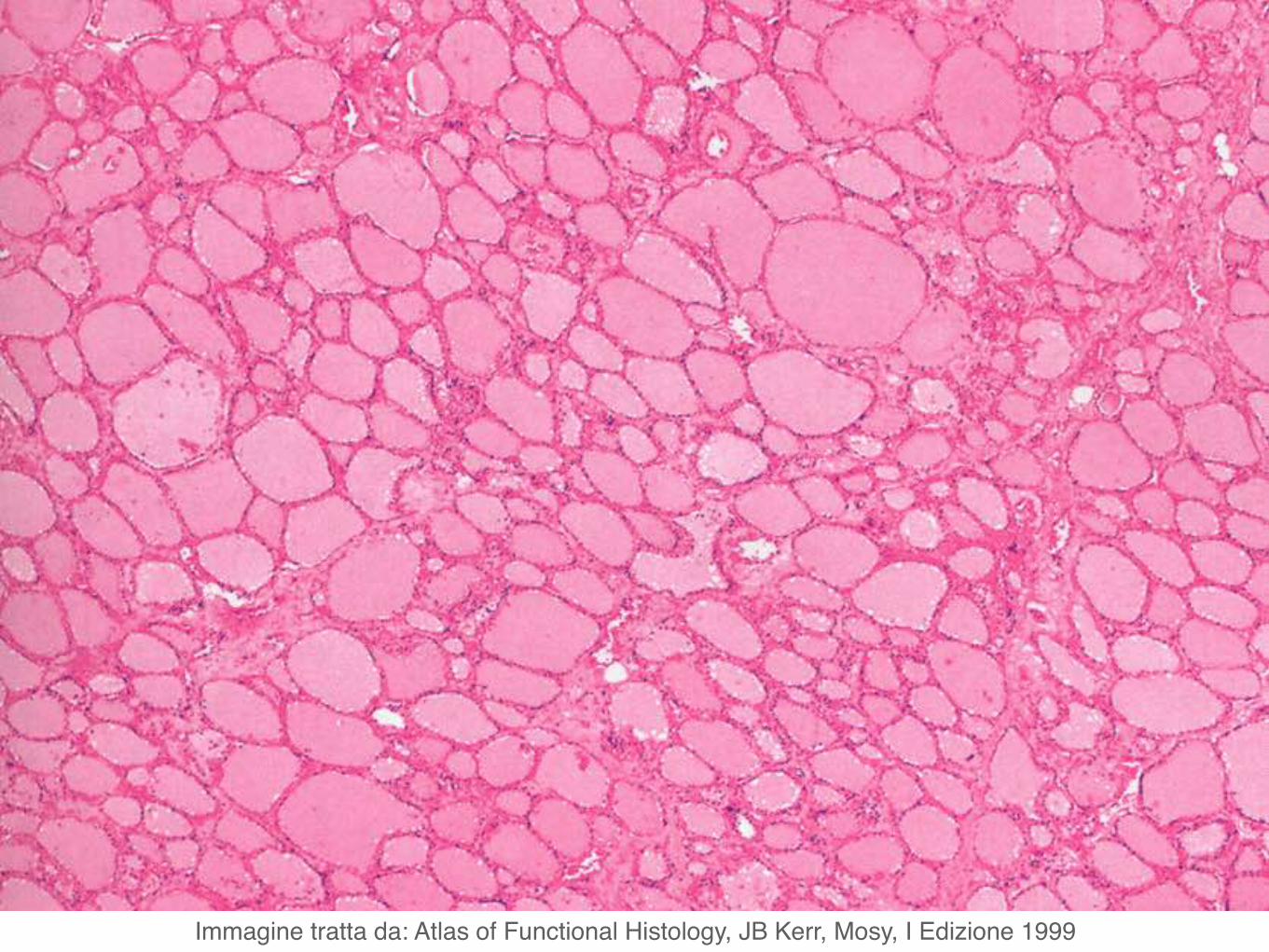
Immagine tratta da: Atlas of Functional Histology, JB Kerr, Mosy, I Edizione 1999
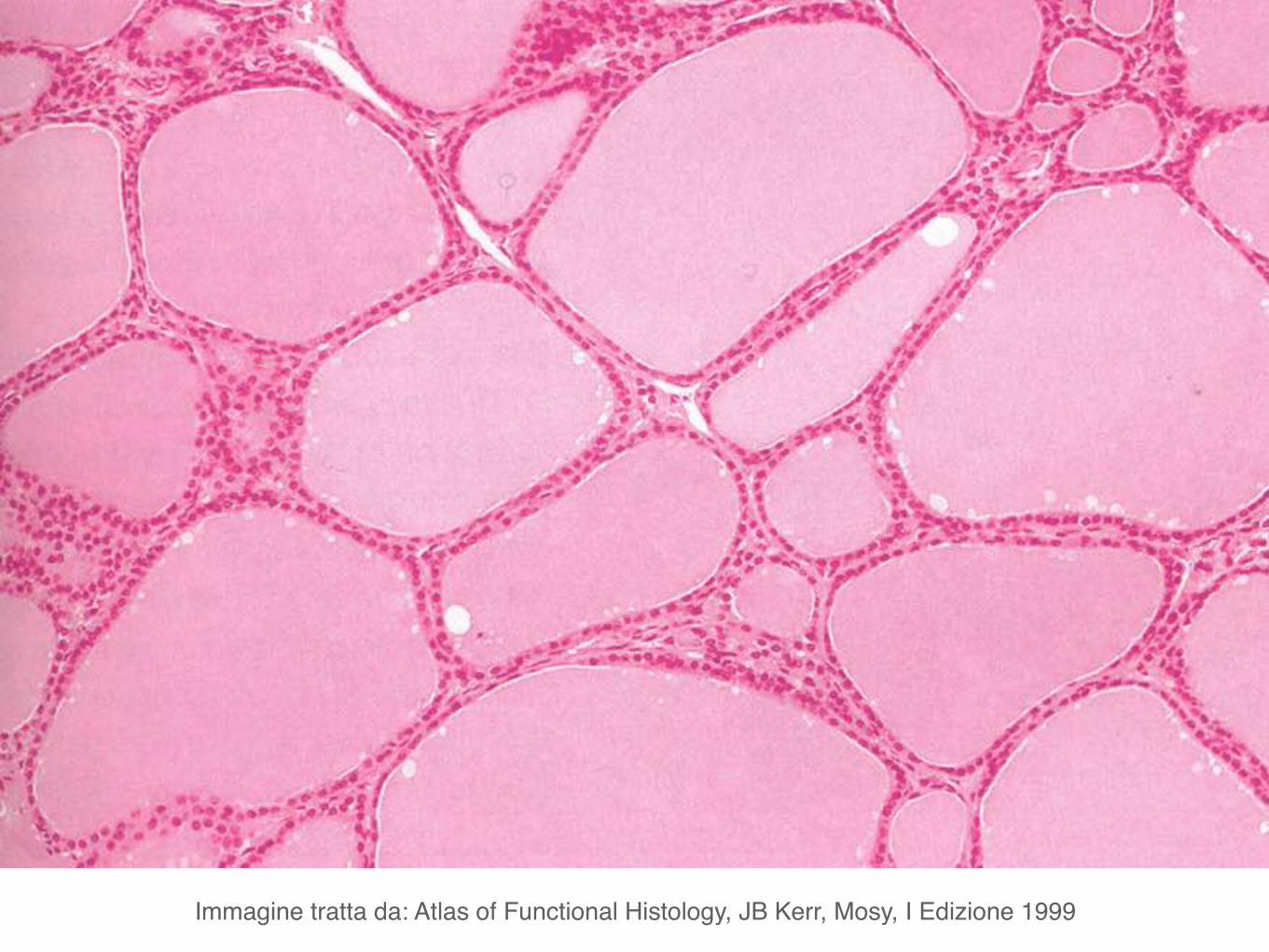
Immagine tratta da: Atlas of Functional Histology, JB Kerr, Mosy, I Edizione 1999

Le cellule follicolari della tiroide producono la proteina tireoglobulina che viene poi convertita in T3 (tiroxina) e T4. Inoltre concentrano iodio prelevandolo dal sangue.
Le cellule parafollicolari o cellule C producono calcitonina cha ha un effetto ipocalcemizzante. Antagonista della calcitonina é il paratormone, prodotto dalle paratiroidi.
Ormoni prodotti dalla tiroide

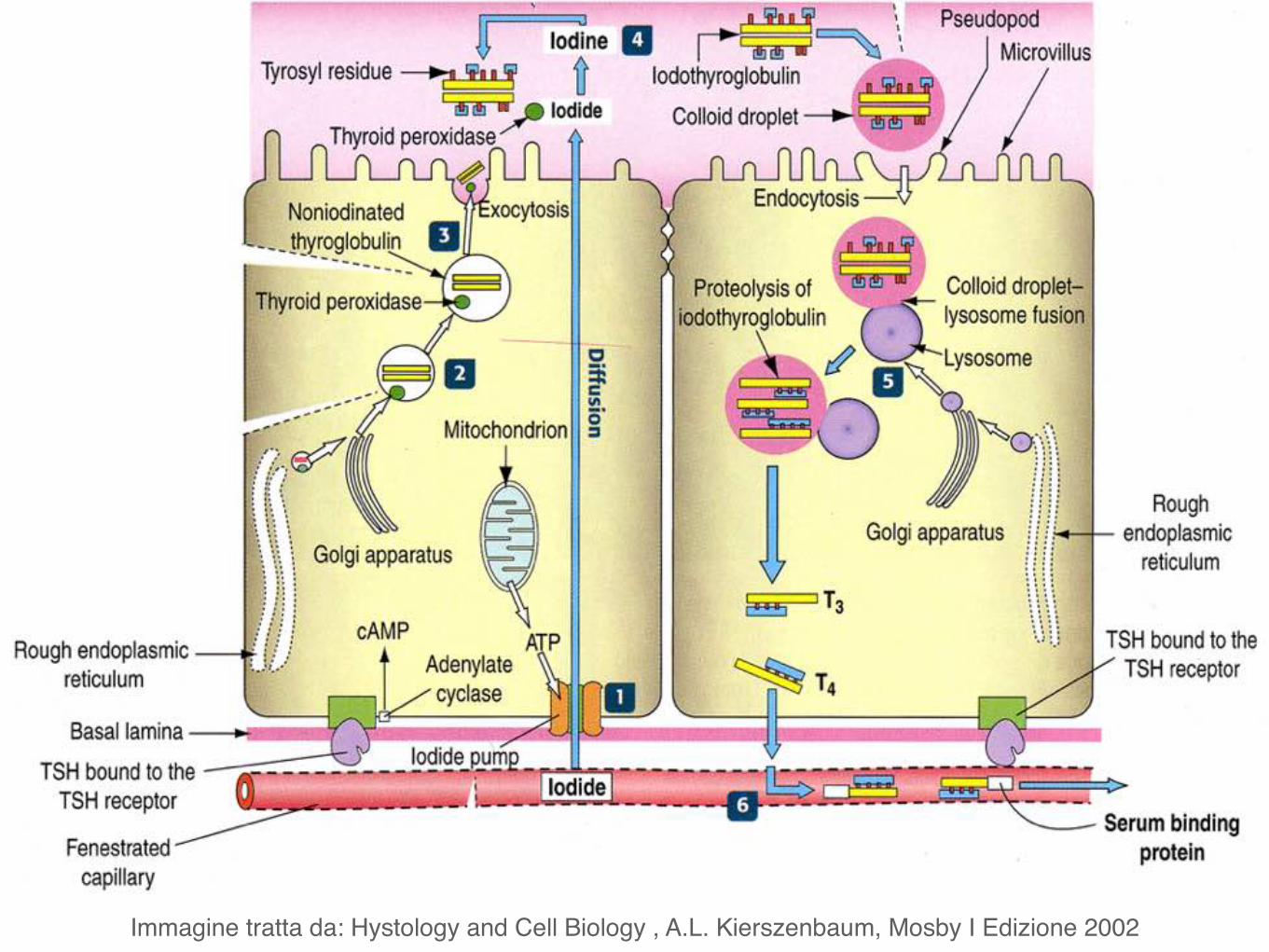
Immagine tratta da: Hystology and Cell Biology , A.L. Kierszenbaum, Mosby I Edizione 2002
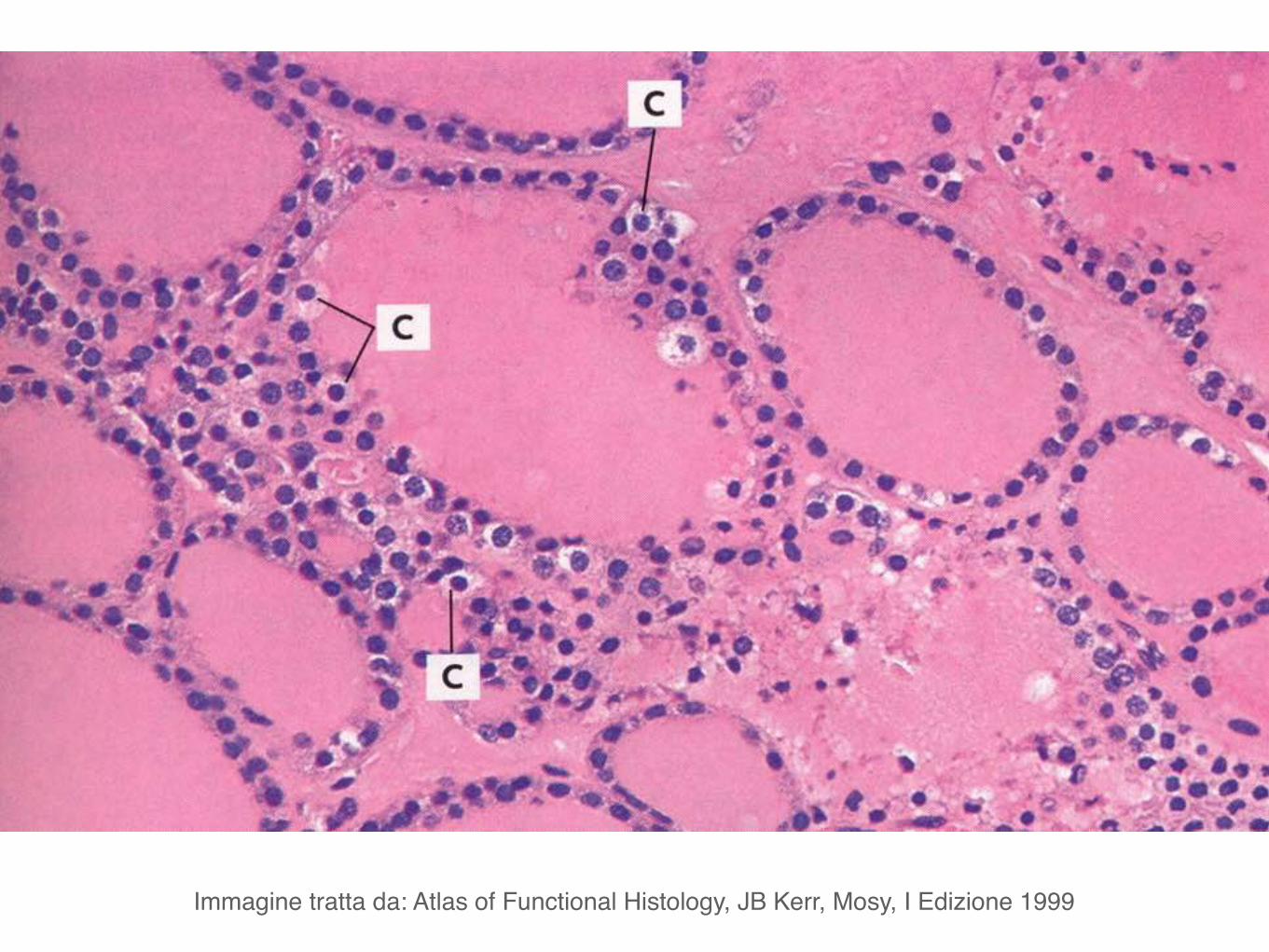
Immagine tratta da: Atlas of Functional Histology, JB Kerr, Mosy, I Edizione 1999

Paratiroidi

Hanno forma per lo più ovale e dimensioni variabili (peso tra 0,2 e 0,5 g).
Sono poste a contatto con la faccia posteriore dei lobi laterali della tiroide.
Generalmente sono in numero di quattro, due superiori e due inferiori.
Paratiroidi
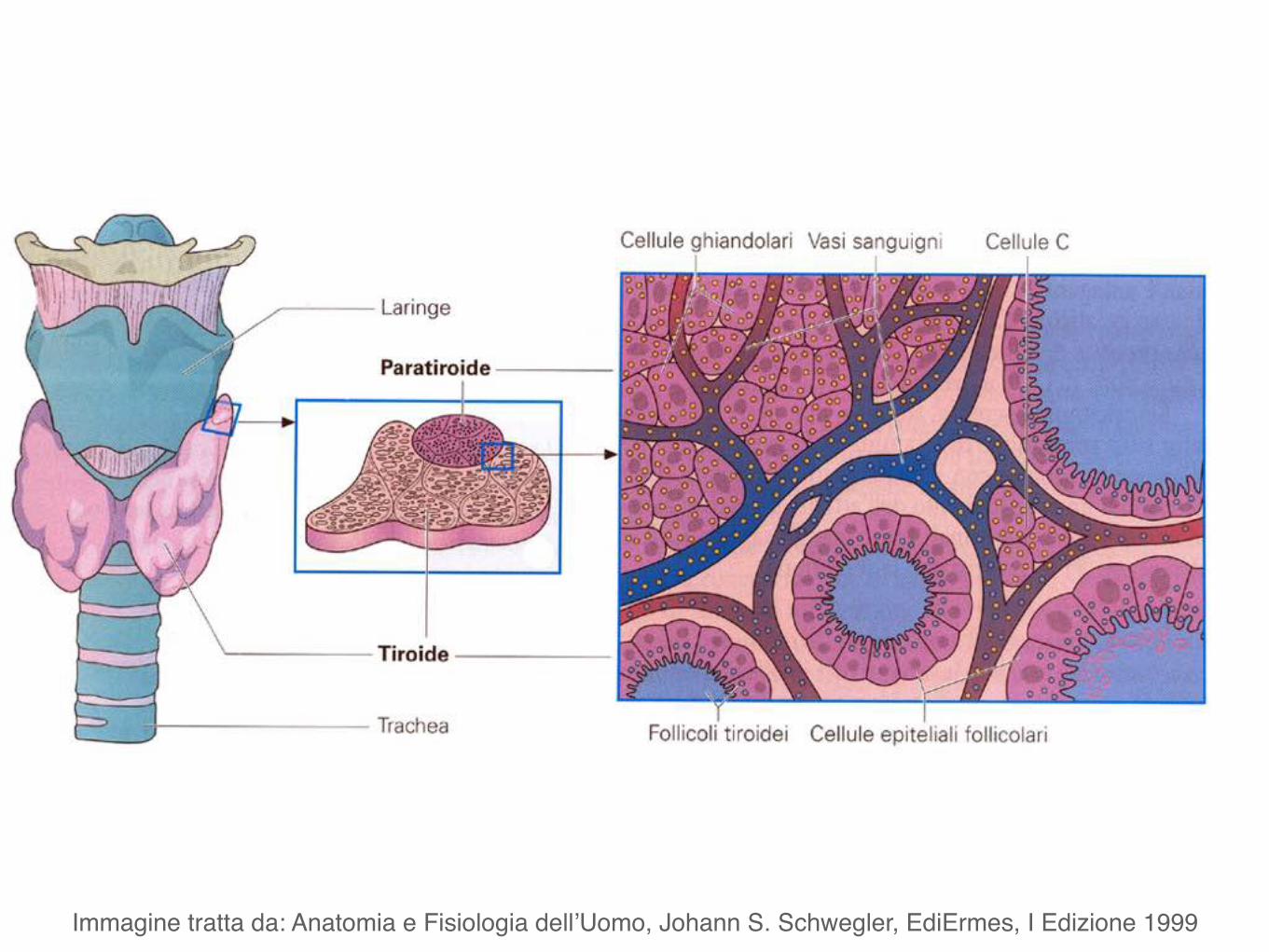
Immagine tratta da: Anatomia e Fisiologia dell’Uomo, Johann S. Schwegler, EdiErmes, I Edizione 1999
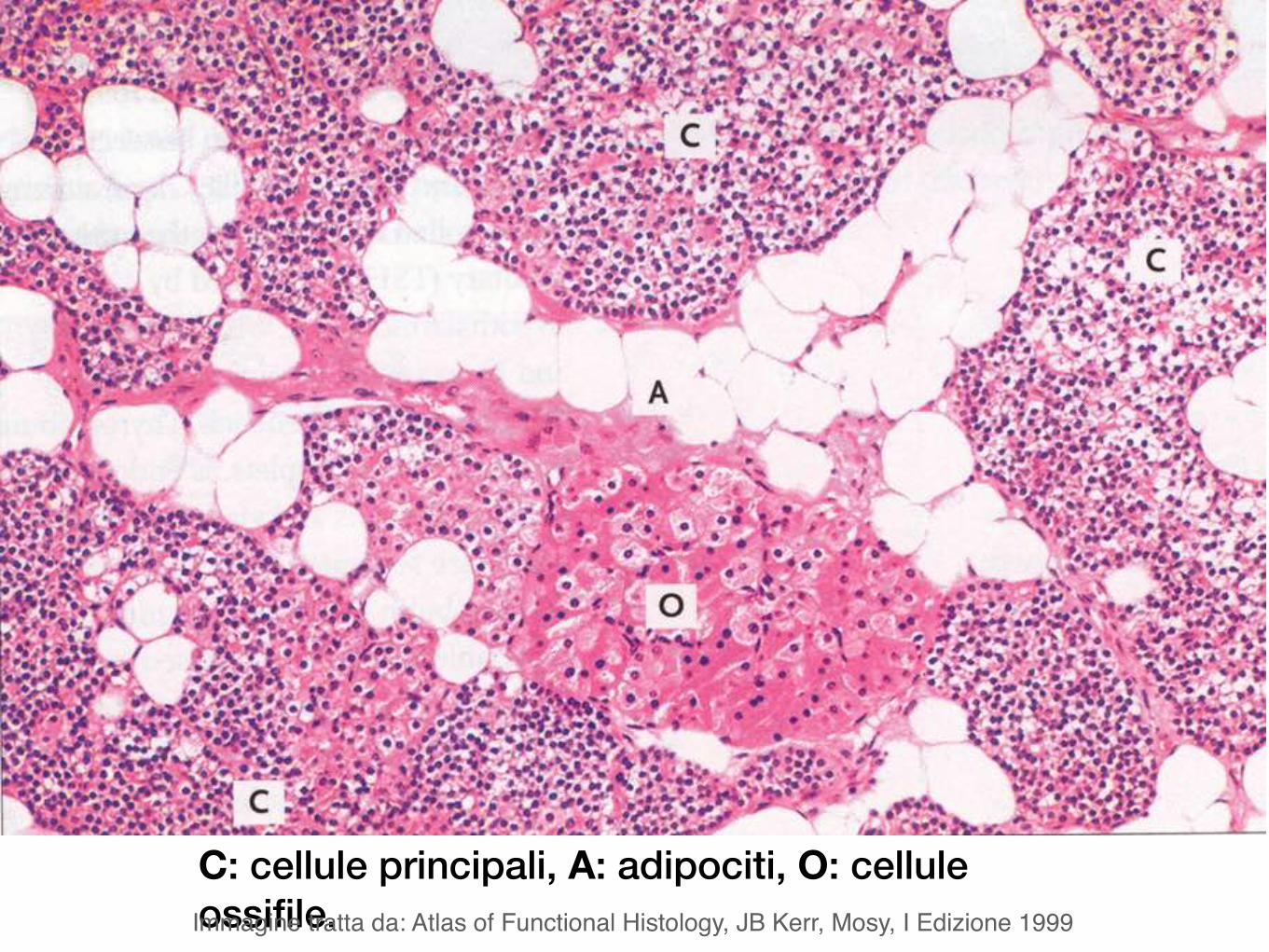
C: cellule principali, A: adipociti, O: cellule ossifile. Immagine tratta da: Atlas of Functional Histology, JB Kerr, Mosy, I Edizione 1999

Le cellule principali della paratiroide producono paratormone, un peptide che controlla le concentrazioni di calcio e fosfato nel sangue.
Il paratormone aumenta il riassorbimento di tessuto osseo incrementando così la concentrazione di calcio e fosfato nel sangue.
Nel rene il paratormone aumenta la formazione di vitamina D attiva che è secreta nel sangue e facilita l’assorbimento di calcio a livello intestinale.
Ormoni prodotti dalla paratiroide
Apparato endocrinoGhiandole surrenali
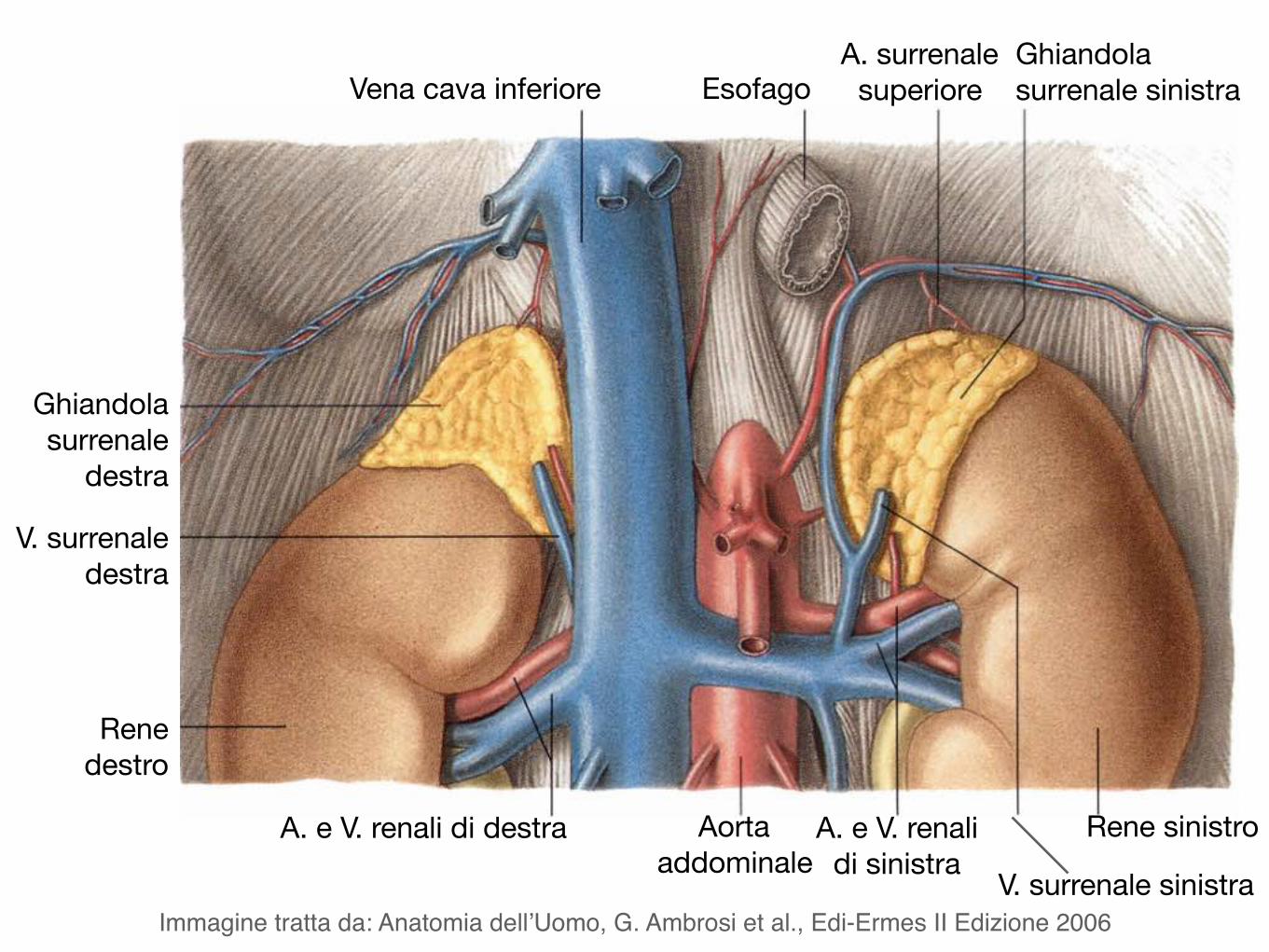
Vena cava inferiore EsofagoA. surrenale
superioreGhiandola surrenale sinistra
Ghiandola surrenale
destra
V. surrenale destra
Rene destro
A. e V. renali di destra Aorta addominale
A. e V. renali di sinistra
Rene sinistro
V. surrenale sinistraImmagine tratta da: Anatomia dell’Uomo, G. Ambrosi et al., Edi-Ermes II Edizione 2006
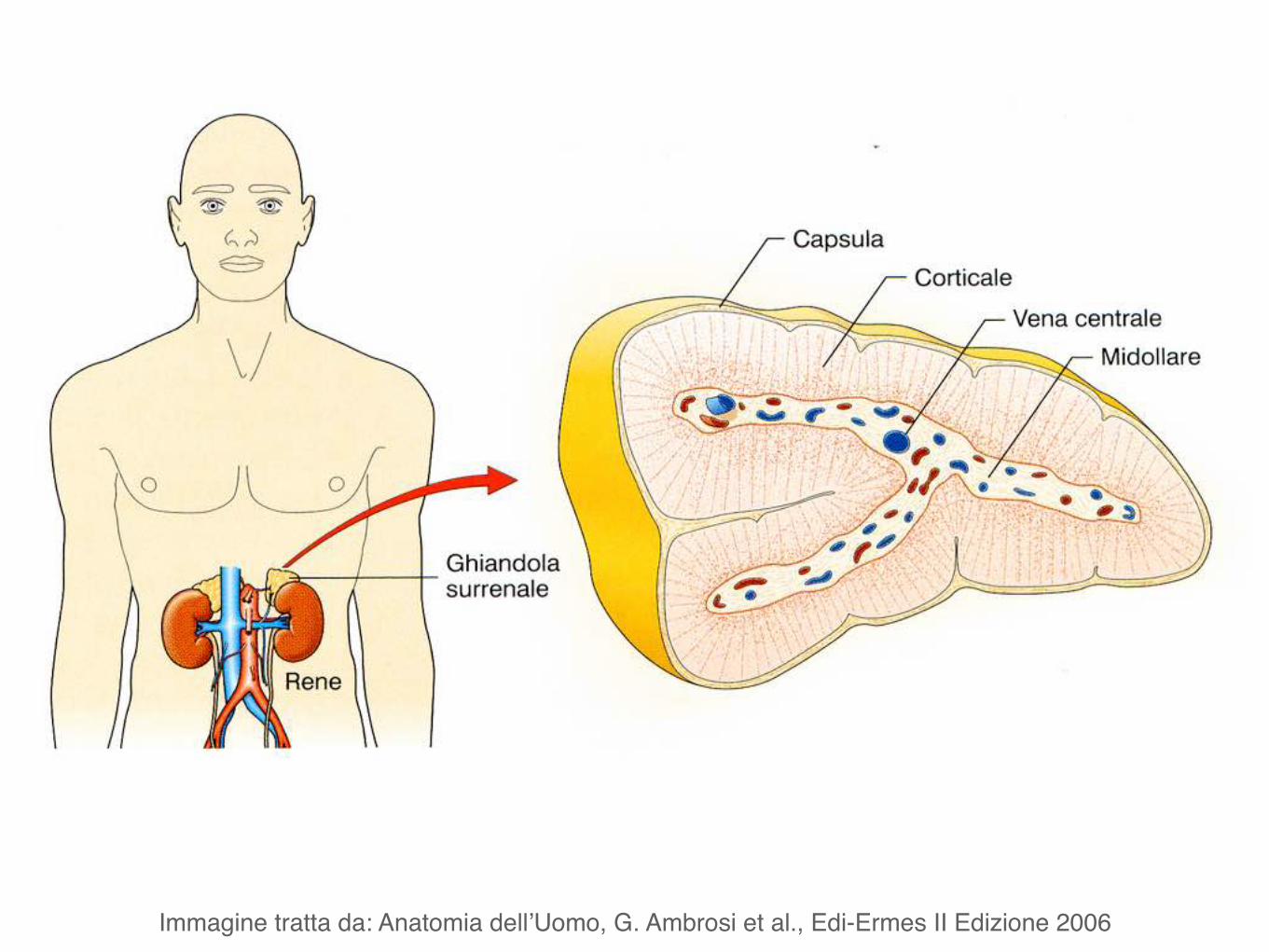
Immagine tratta da: Anatomia dell’Uomo, G. Ambrosi et al., Edi-Ermes II Edizione 2006
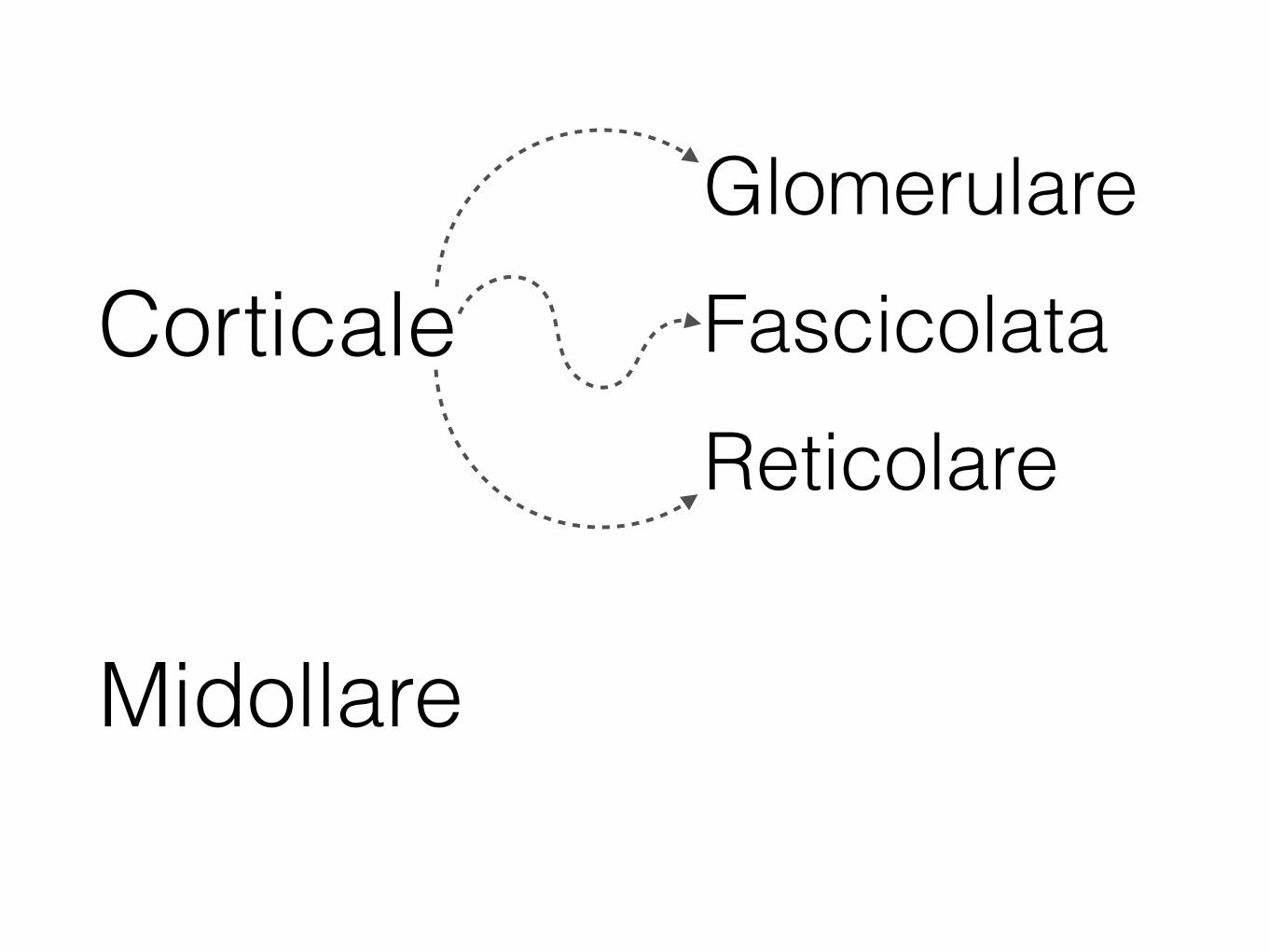
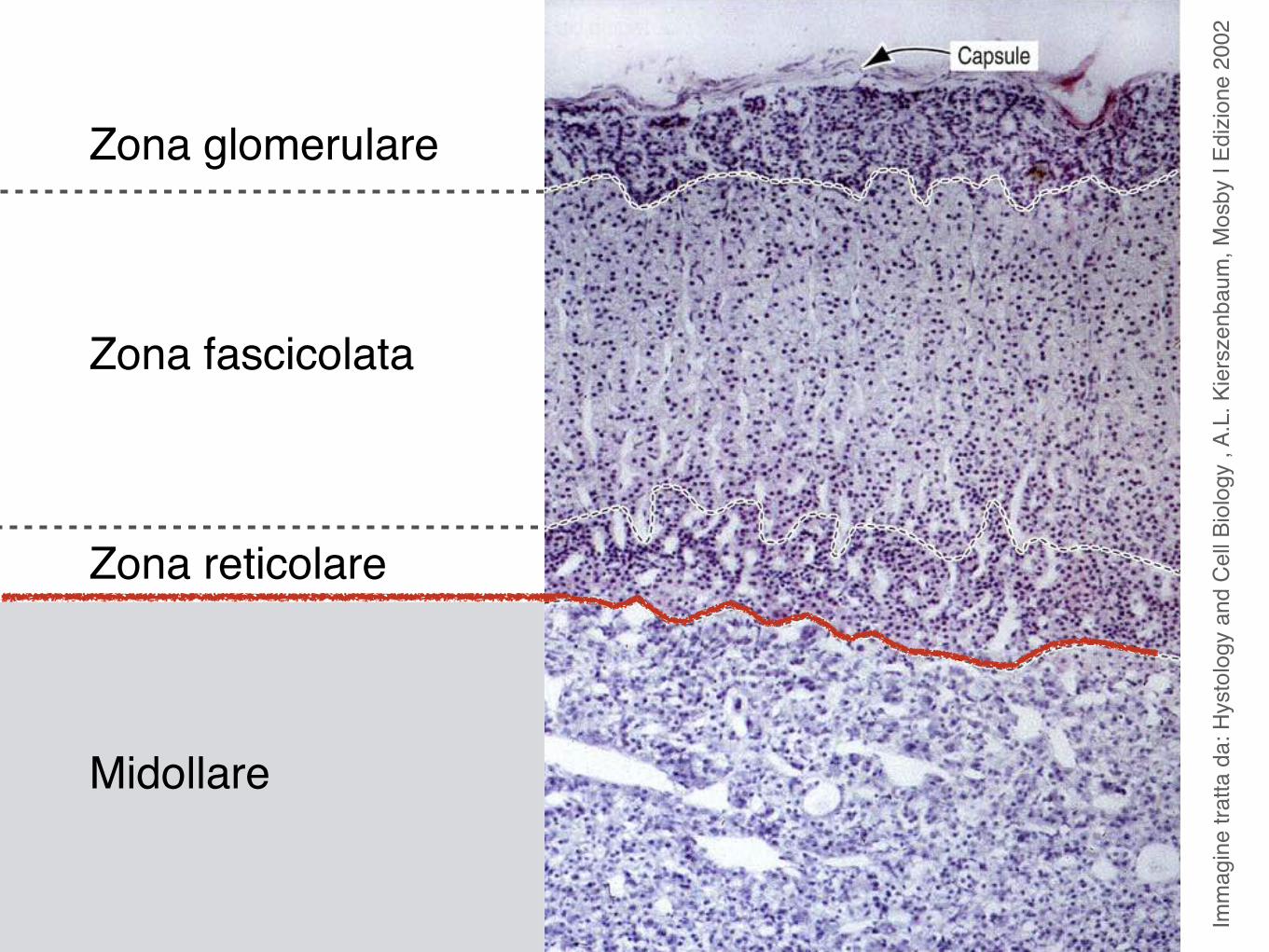
Corticale
Midollare
Glomerulare
ReticolareFascicolata
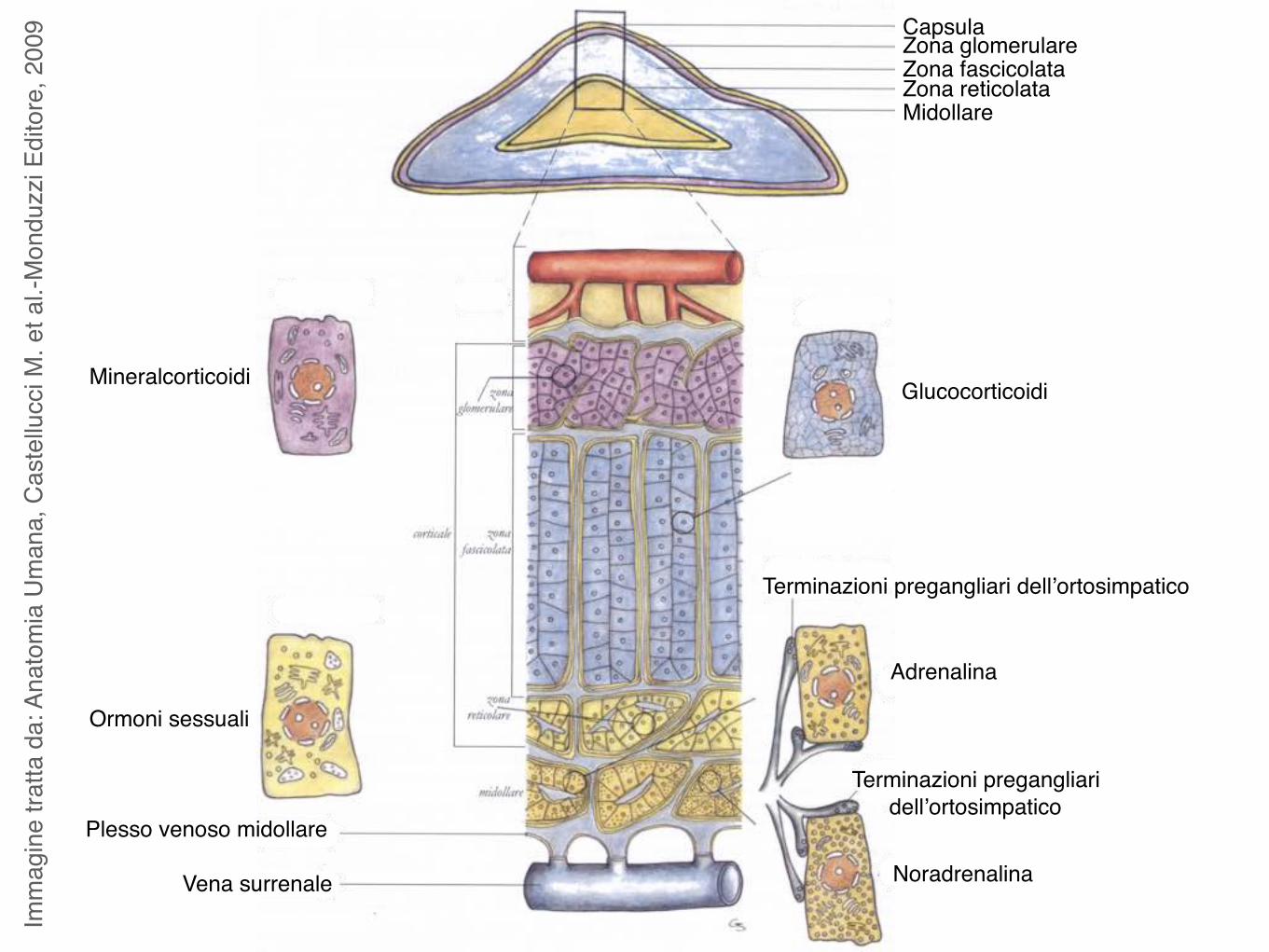
Imm
agin
e tra
tta d
a: A
nato
mia
Um
ana,
Cas
tellu
cci M
. et a
l.-M
ondu
zzi E
dito
re, 2
009 Capsula
Zona glomerulareZona fascicolataZona reticolataMidollare
Plesso venoso midollare
Vena surrenale
Terminazioni pregangliari dell’ortosimpatico
Terminazioni pregangliari dell’ortosimpatico
Adrenalina
Noradrenalina
GlucocorticoidiMineralcorticoidi
Ormoni sessuali
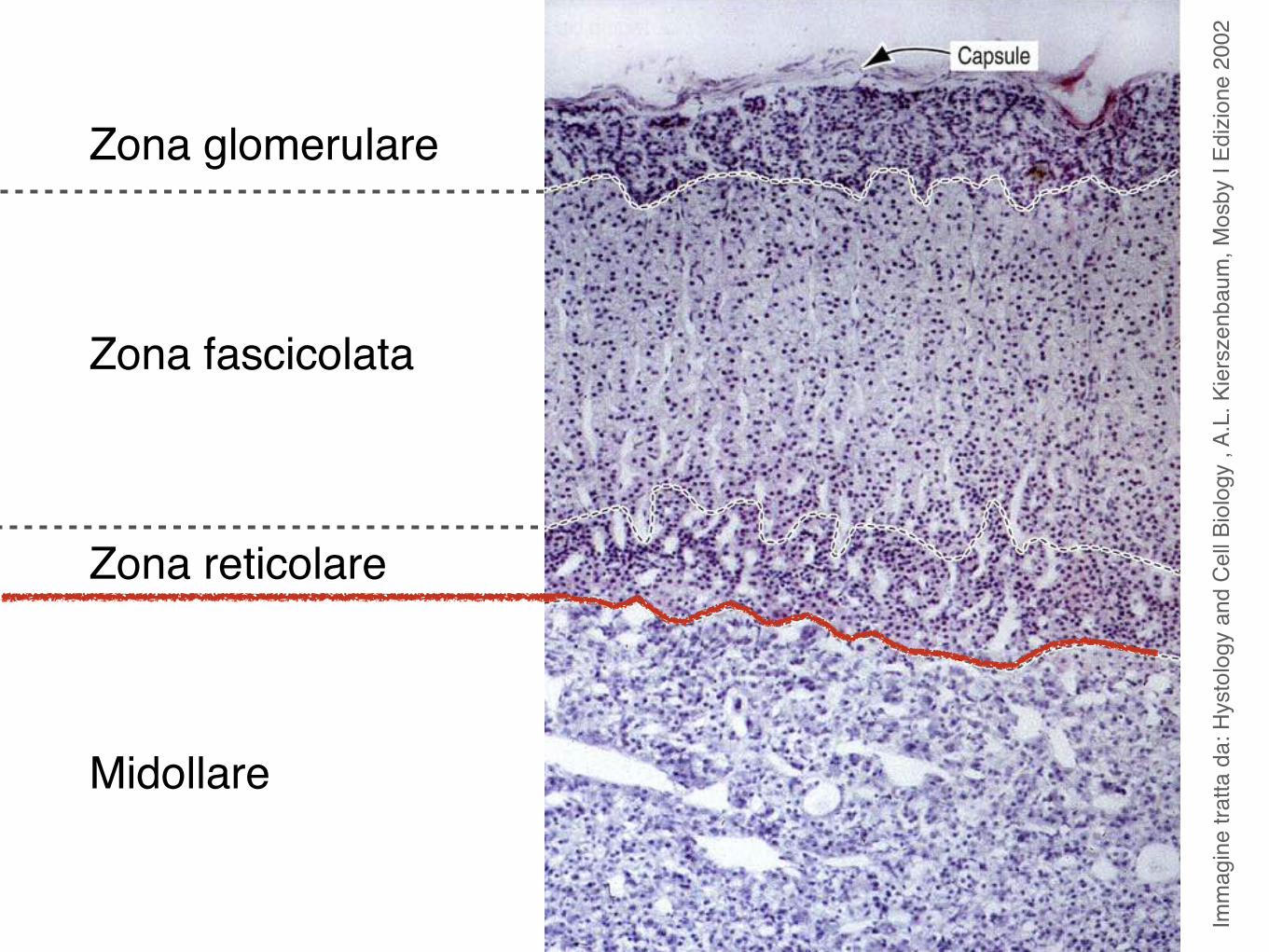
Imm
agin
e tra
tta d
a: H
ysto
logy
and
Cel
l Bio
logy
, A.
L. K
iers
zenb
aum
, Mos
by I
Edizi
one
2002
Zona glomerulare
Zona fascicolata
Zona reticolare
Midollare
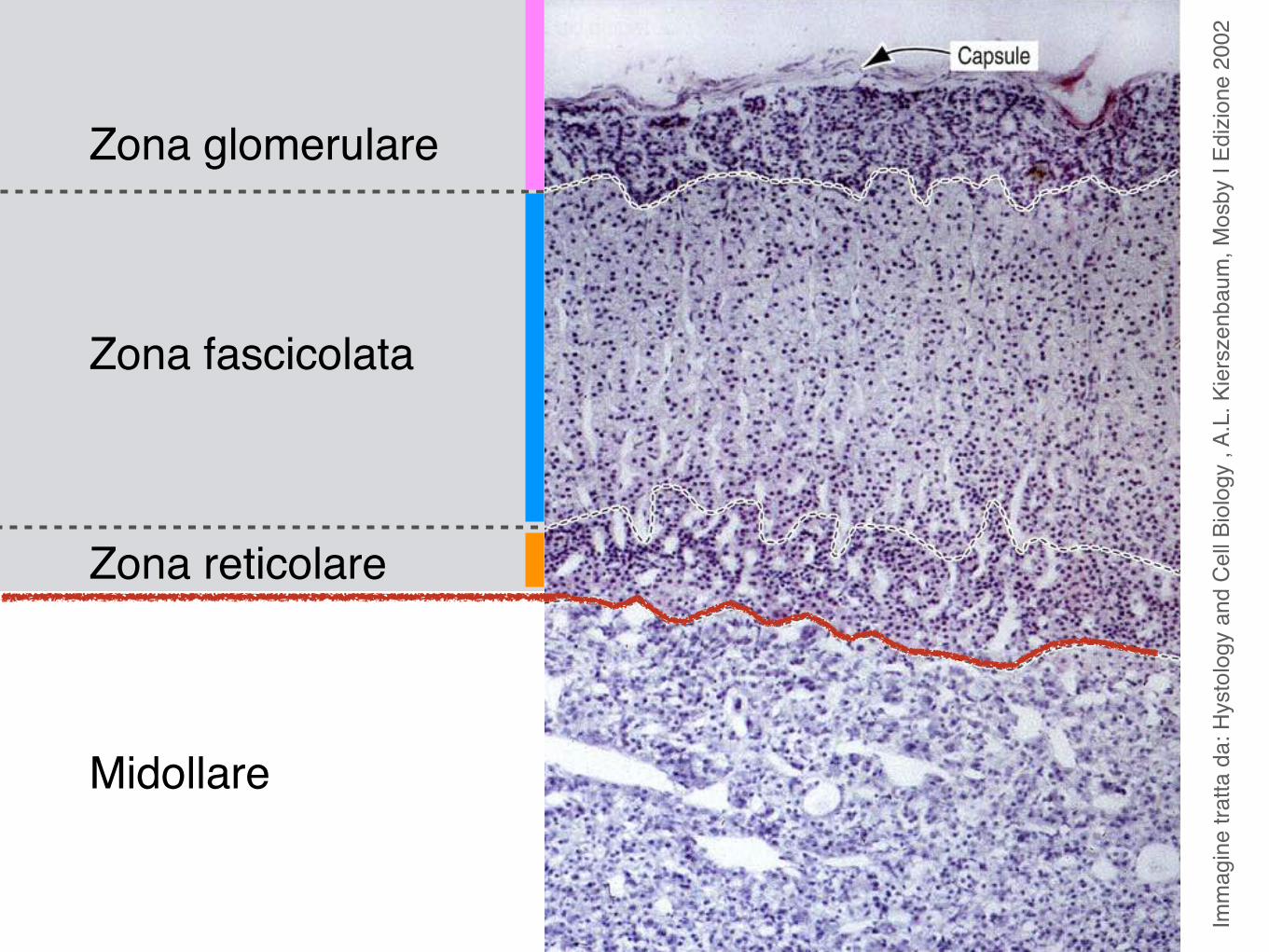
Imm
agin
e tra
tta d
a: H
ysto
logy
and
Cel
l Bio
logy
, A.
L. K
iers
zenb
aum
, Mos
by I
Edizi
one
2002
Zona glomerulare
Zona fascicolata
Zona reticolare
Midollare
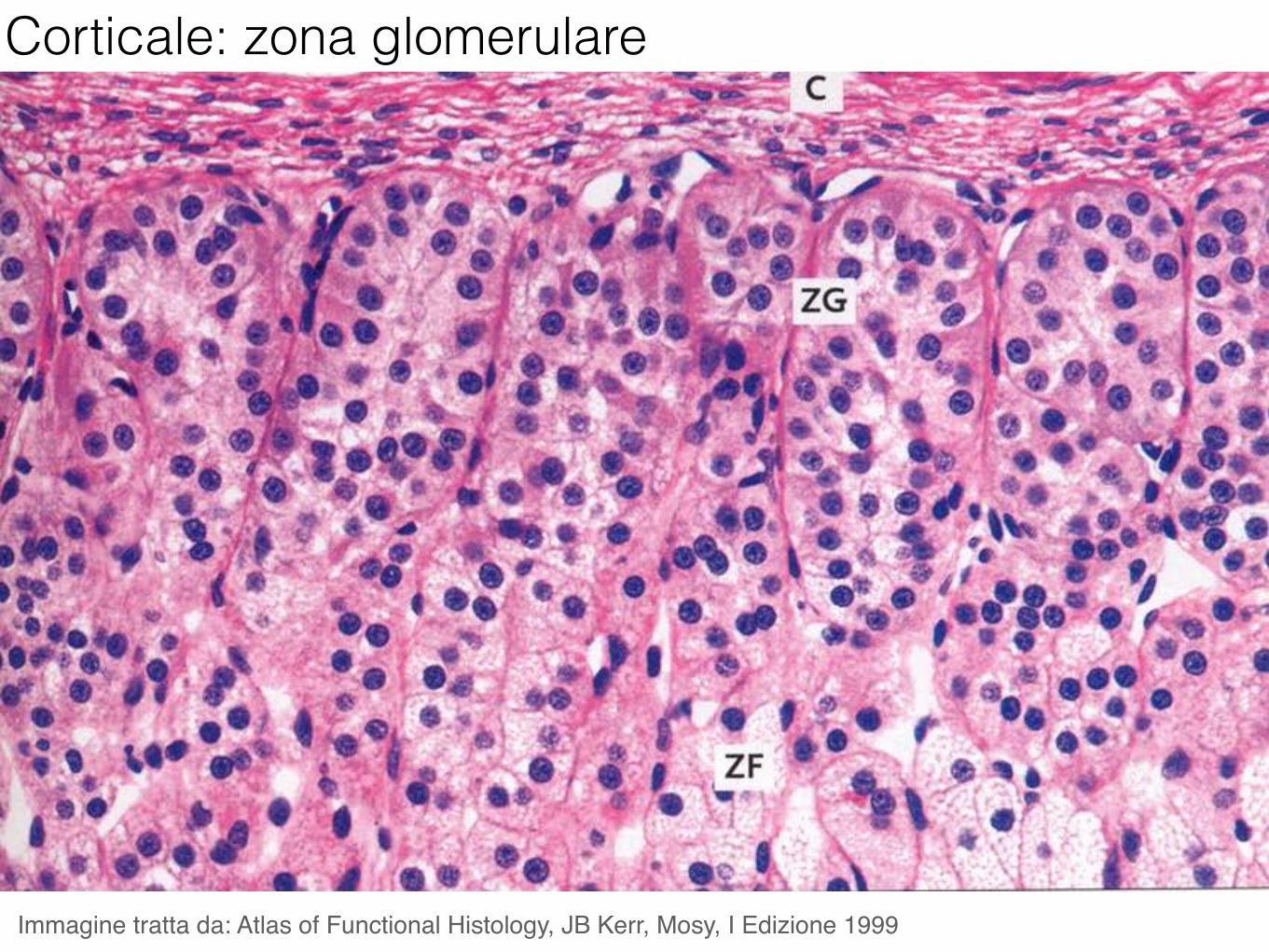
Corticale: zona glomerulare
Immagine tratta da: Atlas of Functional Histology, JB Kerr, Mosy, I Edizione 1999
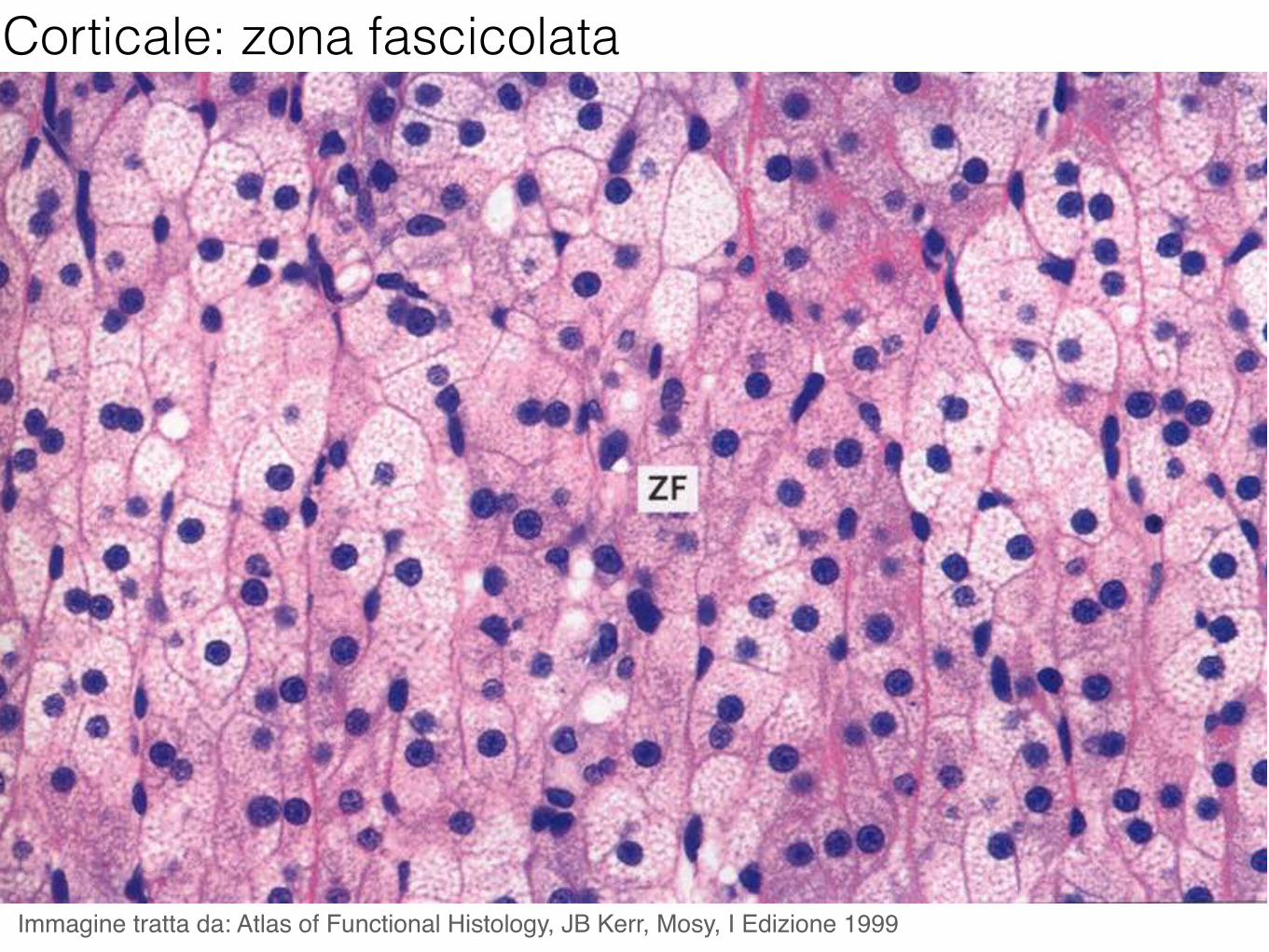
Corticale: zona fascicolata
Immagine tratta da: Atlas of Functional Histology, JB Kerr, Mosy, I Edizione 1999
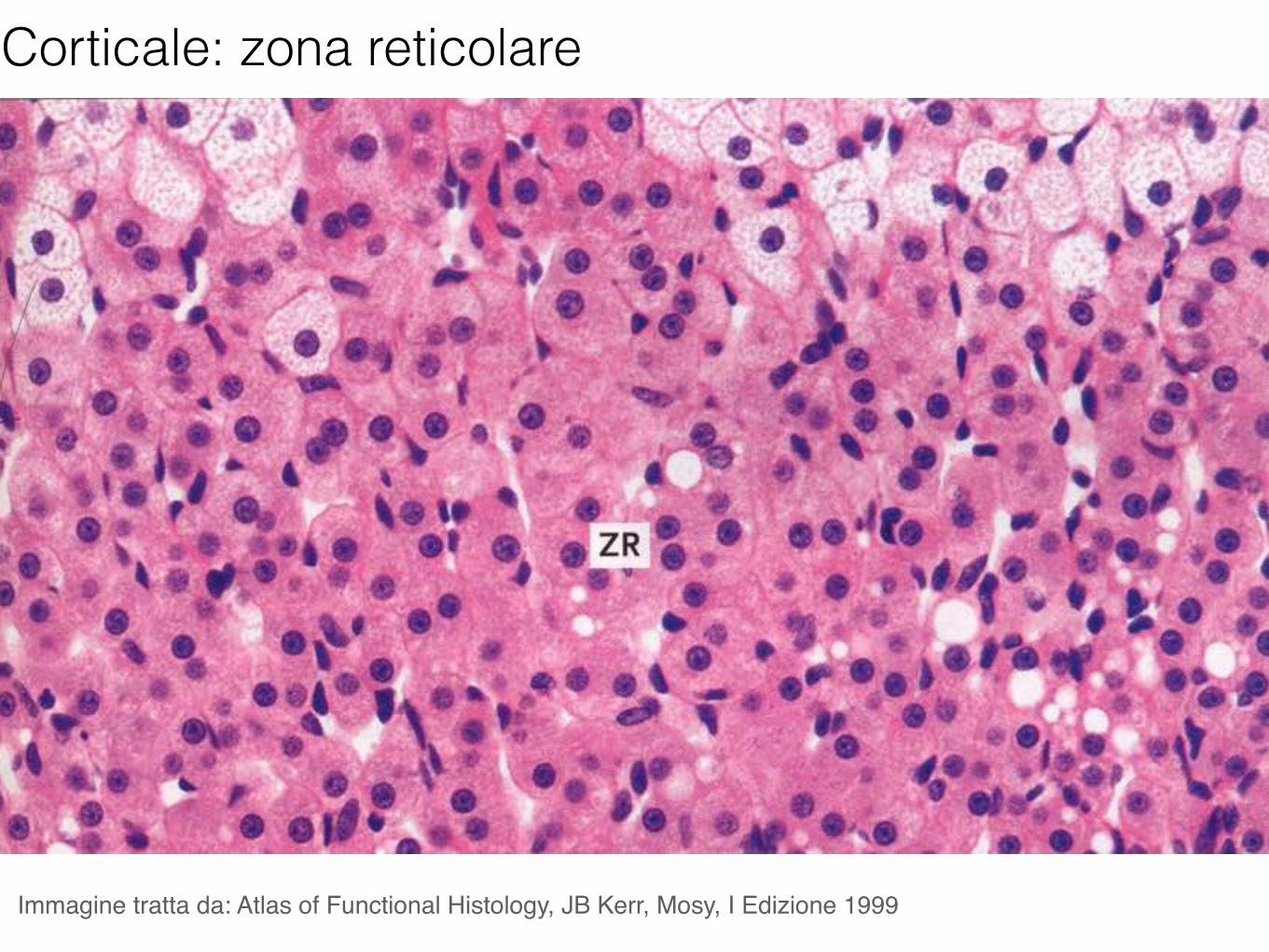
Immagine tratta da: Atlas of Functional Histology, JB Kerr, Mosy, I Edizione 1999
Corticale: zona reticolare

La corticale del surrene produce 3 classi di ormoni steroidei: mineralcorticoidi, glucocorticoidi e androgeni.
Il principale mineralcorticoide è l’aldosterone, prodotto nella zona glomerulare. La sua funzione è controllare il volume dei liquidi dell’organismo incrementando l’assorbimento di sodio nei reni agendo sulle cellule del tubulo contorto distale. Viene prodotto in seguito a stimolazione mediante angiotensina.
Il pricipale glucocorticoide è il cortisolo, prodotto nella zona fascicolata. Il cortisolo influisce sulla sintesi di carboidrati, proteine e grassi:
diminuisce la sintesi proteica aumenta la sintesi di glicogeno mobilizza gli acidi grassi e il glicerolo del tessuto adiposo
Gli androgeni secreti nella zona reticolare sono il deidroepiandrosterone (DHA) ed il suo solfato.
Ormoni prodotti dalla corticale
Imm
agin
e tra
tta d
a: H
ysto
logy
and
Cel
l Bio
logy
, A.
L. K
iers
zenb
aum
, Mos
by I
Edizi
one
2002
Zona glomerulare
Zona fascicolata
Zona reticolare
Midollare
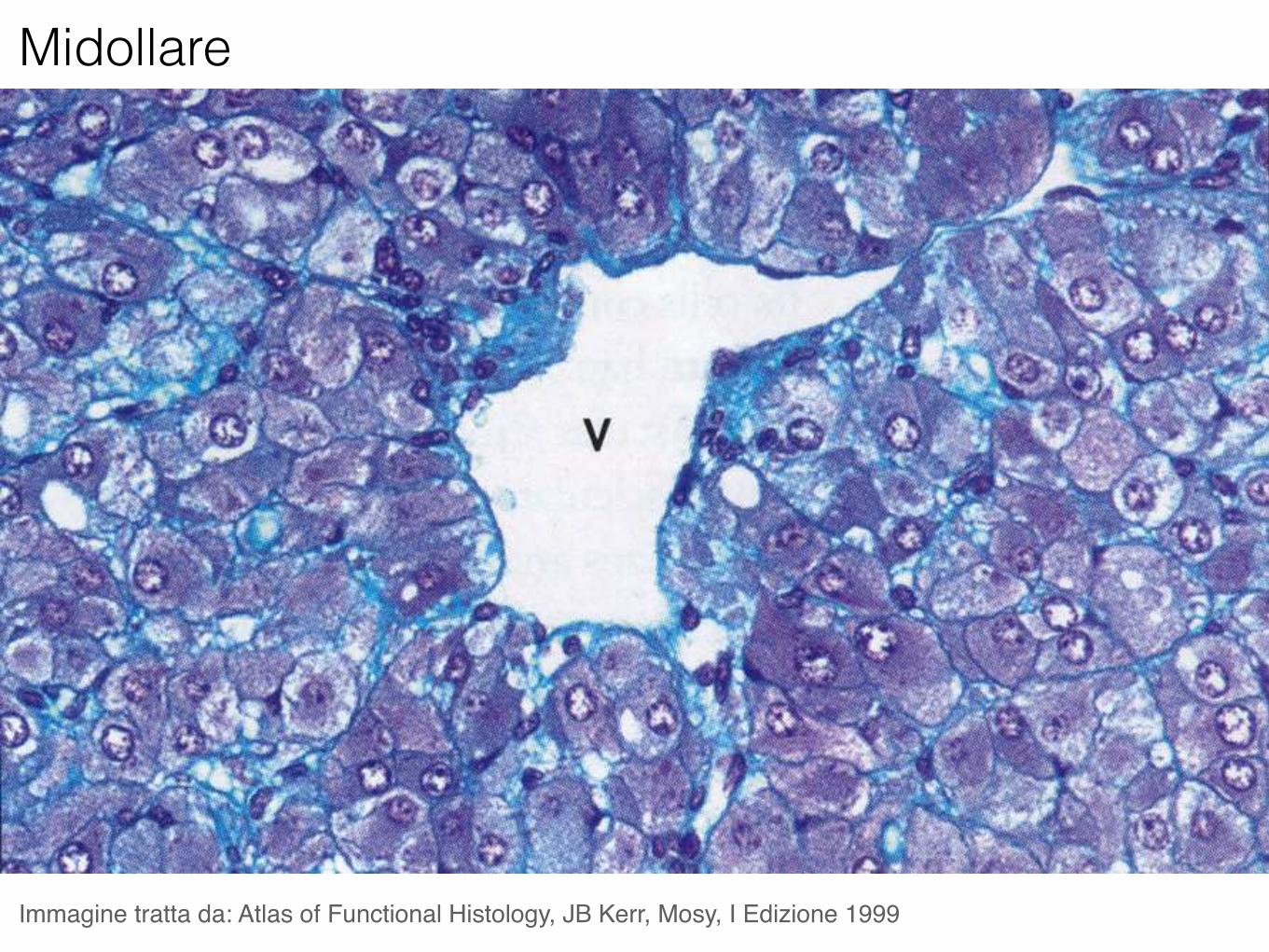
Midollare
Immagine tratta da: Atlas of Functional Histology, JB Kerr, Mosy, I Edizione 1999

Le cellule della midollare del surrene sono neuroni postgangliari modificati e la loro attività secernente ricade sotto il controllo nervoso, attraverso i nervi splancnici.
Le cellule della midollare secernono adrenalina (75%) e noradrenalina (25%).
Ormoni prodotti dalla midollare

Benchè siano molto simili adrenalina e noradrenalina hanno effetti molto diversi:
l’adrenalina aumenta la frequenza e la gittata cardiaca ed il metabolismo;
la noradrenalina non influenza frequenza e gittata cardiaca e metabolismo ma causa un intenso rialzo della pressione arteriosa dovuto a costrizione dei vasi.
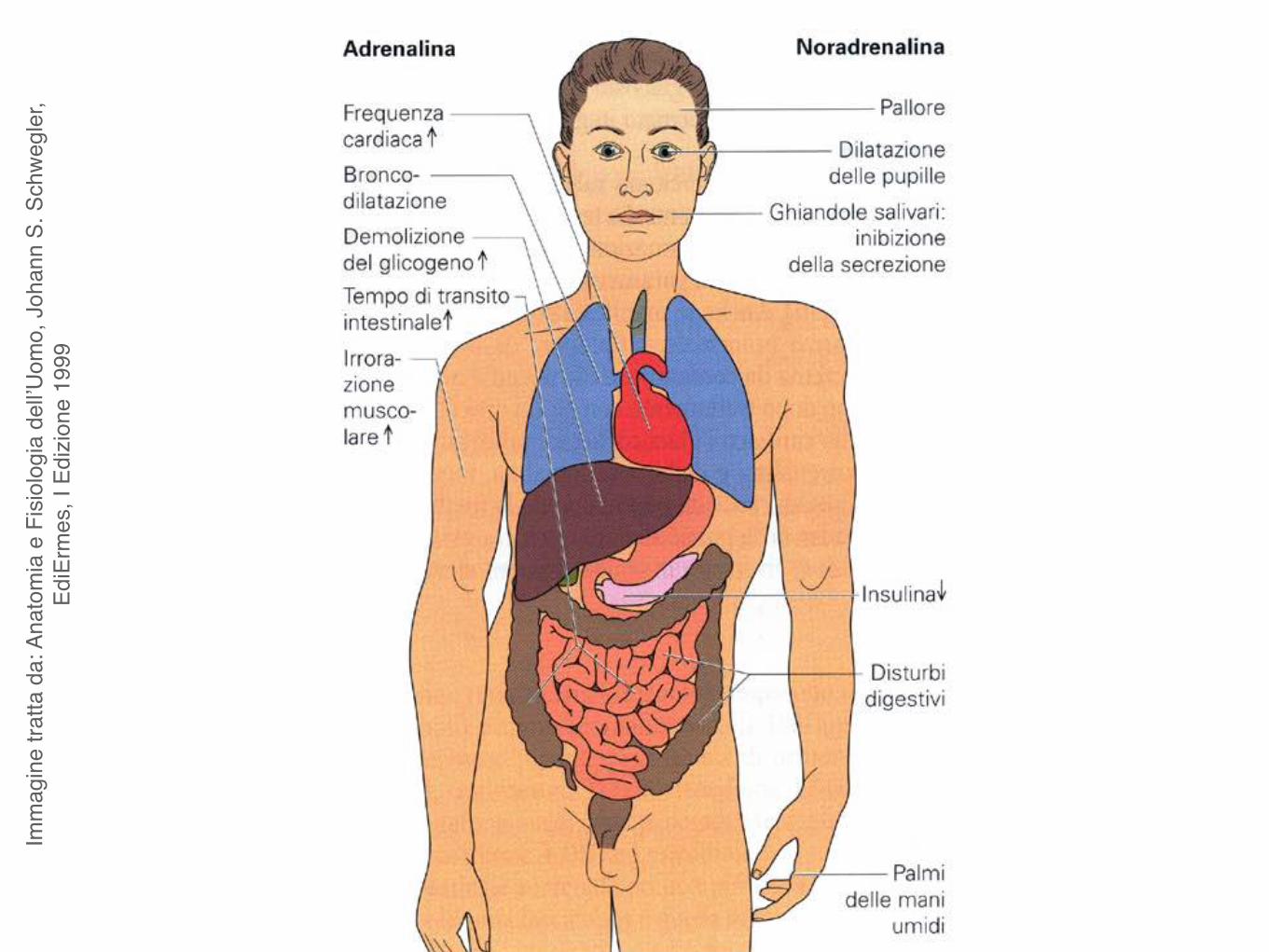
Imm
agin
e tra
tta d
a: A
nato
mia
e F
isiol
ogia
del
l’Uom
o, J
ohan
n S.
Sch
wegl
er,
EdiE
rmes
, I E
dizio
ne 1
999
Apparato endocrinoPancreas endocrino
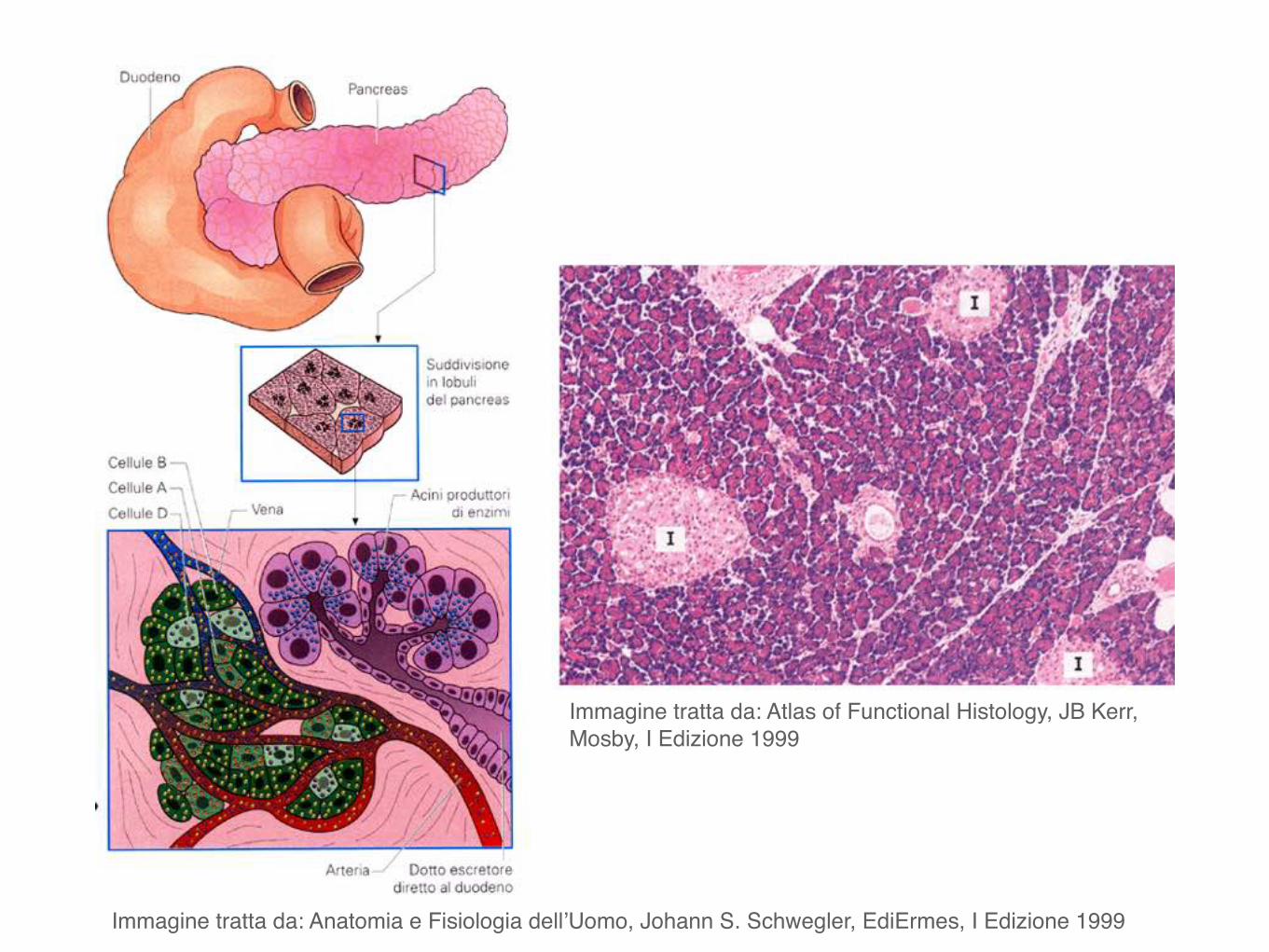
Immagine tratta da: Anatomia e Fisiologia dell’Uomo, Johann S. Schwegler, EdiErmes, I Edizione 1999
Immagine tratta da: Atlas of Functional Histology, JB Kerr, Mosby, I Edizione 1999
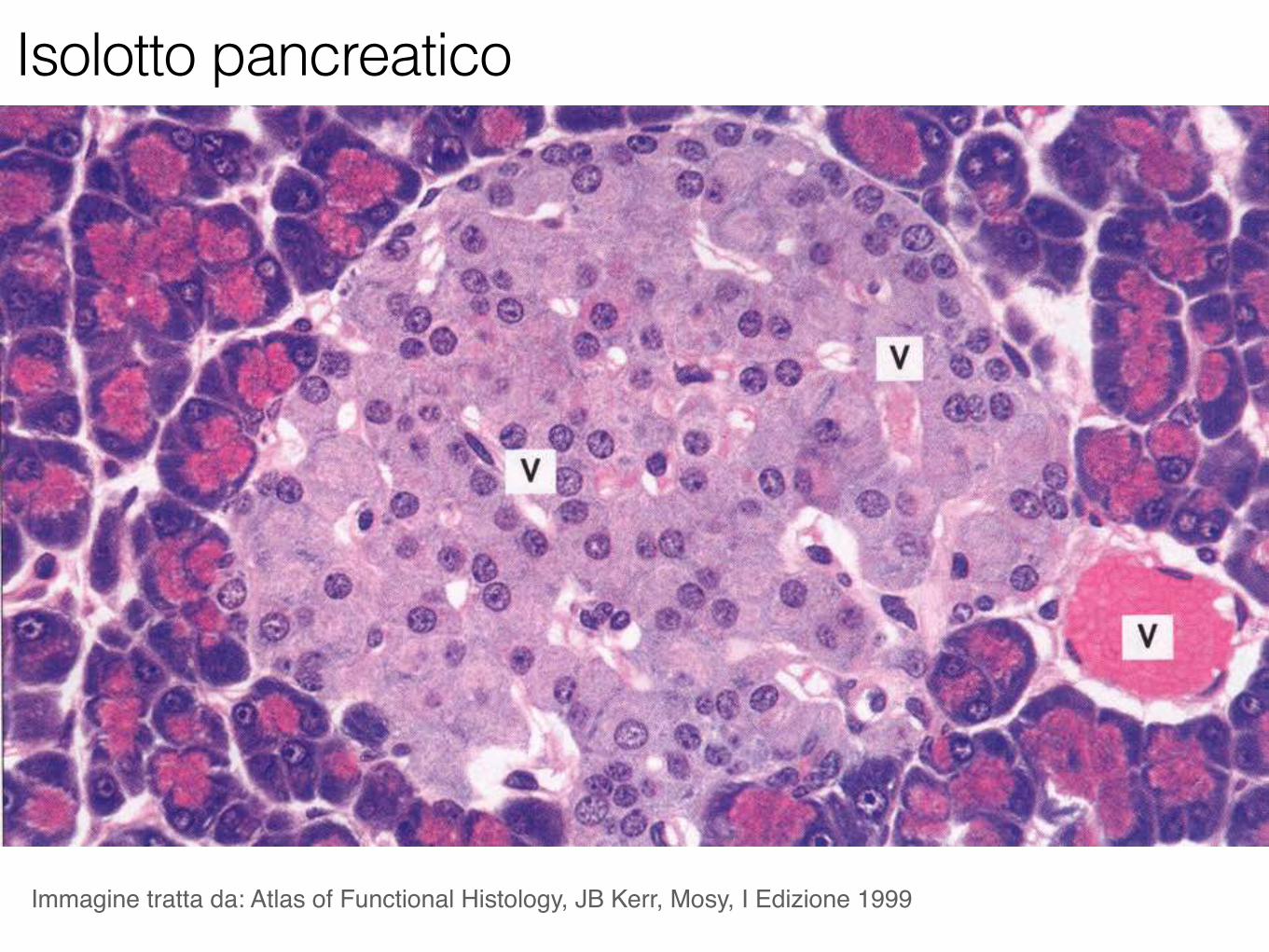
Immagine tratta da: Atlas of Functional Histology, JB Kerr, Mosy, I Edizione 1999
Isolotto pancreatico

Cellule α: rappresentano il 20% delle cellule delle isole pancreatiche. Producono l’ormone glucagone.
Cellule β: rappresentano il 70% delle cellule delle isole pancreatiche. Producono l’ormone insulina.
Cellule δ: sono più rare, <10% del totale delle cellule delle isole pancreatiche. Producono l’ormone somatostatina che, per via paracrina, inibisce la sintesi di insulina e glucagone.
Tipi cellulari costituenti le isole di Langerhans
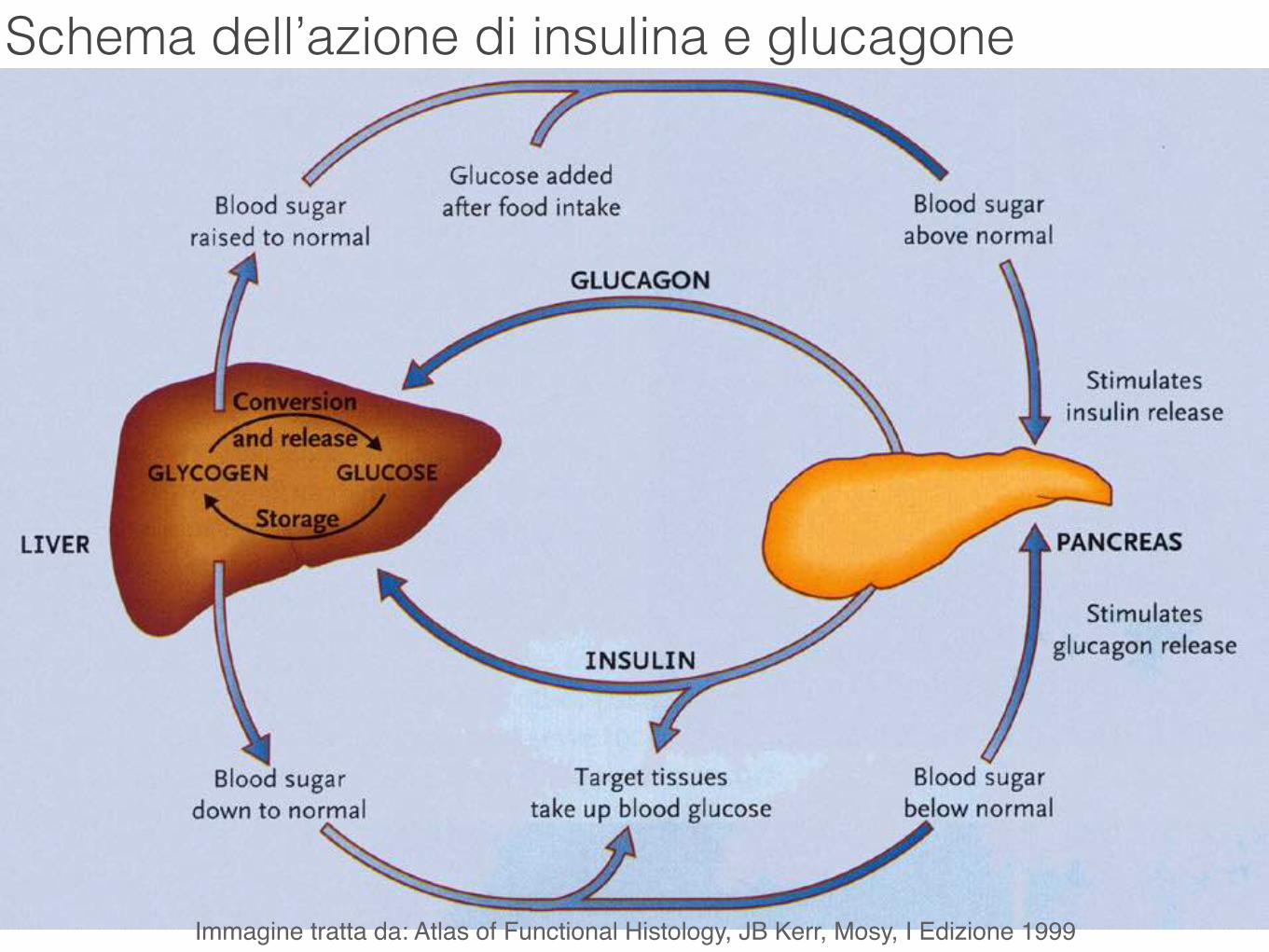
Schema dell’azione di insulina e glucagone
Immagine tratta da: Atlas of Functional Histology, JB Kerr, Mosy, I Edizione 1999

Sistema gastroentero- pancreatico (GEP)

Il tratto gastrointestinale contiene 16 o più tipi di cellule neuroendocrine che producono più di 30 ormoni diversi: lo stomaco contiene:
- le cellule G che secernono gastrina - le cellule ECL (tipo-enterocromaffini) che secernono istamina - le cellule D che secernono somatostatina;
l’intestino tenue contiene:
- le cellule S che producono secretina che stimola la produzione di fluido ricco di HCO3- nel pancreas,
- le cellule I che producono colecistochinasi (CCK) che stimola la produzione di enzimi pancreatici,
- le cellule K che secernono GIP un peptide che stimola il rilascio dell’insulina,
- le cellule M che secernono motilina che stimola la contrazione della muscolatura liscia.
Sistema gastroenteropancreatico (GEP)

Altri organi ad attività endocrina

Rene secerne renina che ha un’azione ormone-simile nel sistema renina-angiotensina, secerne eritropoietina che stimola la produzione di eritrociti nel midollo osseo nel rene si ha la conversione della vitamina D nella sua forma attiva.
Placenta produce gonadotropina corionica che mantiene le funzioni del corpo luteo all’inizio della gravidanza secerne lattogeno placentale che stimola lo sviluppo del seno.

Cuore Le cellule mioendocrine del cuore,site principalmente nelle auricole atriali e nel setto interventricolare sintetizzano un pro-ormone che dà origine a ad una serie di ormoni peptidici: cardionatrina, cardiodilatina, atriopeptina e polipeptide natriuretico atriale (ANP).
Nell’insieme essi hanno una potente azione diuretica e aumentano l’escrezione di sodio, inibiscono la secrezione di aldosterone da parte del surrene e inducono rilassamento della parete dei vasi.