12.metabolismo lipidi [modalit compatibilit ] · punto di regolazione metabolica. Regolazione...
30
Metabolismo Lipidi
Transcript of 12.metabolismo lipidi [modalit compatibilit ] · punto di regolazione metabolica. Regolazione...
![Page 1: 12.metabolismo lipidi [modalit compatibilit ] · punto di regolazione metabolica. Regolazione covalente mediante fosforilazione ... della biosintesi degli acidi grassi e del colesterolo,](https://reader031.fdocumenti.com/reader031/viewer/2022022709/5c65ab8009d3f2a86e8cf785/html5/thumbnails/1.jpg)
Metabolismo Lipidi
![Page 2: 12.metabolismo lipidi [modalit compatibilit ] · punto di regolazione metabolica. Regolazione covalente mediante fosforilazione ... della biosintesi degli acidi grassi e del colesterolo,](https://reader031.fdocumenti.com/reader031/viewer/2022022709/5c65ab8009d3f2a86e8cf785/html5/thumbnails/2.jpg)
Digestione ed assorbimento dei lipidi della dieta
![Page 3: 12.metabolismo lipidi [modalit compatibilit ] · punto di regolazione metabolica. Regolazione covalente mediante fosforilazione ... della biosintesi degli acidi grassi e del colesterolo,](https://reader031.fdocumenti.com/reader031/viewer/2022022709/5c65ab8009d3f2a86e8cf785/html5/thumbnails/3.jpg)
Il principale pigmento biliare è la BILIRUBINA, derivata dalla biliverdina,
prodotta dalla degradazione dell’EME
nell’intestino è convertita in stercobilinogeno e poi stercobilina (marrone)
nel fegato la bilirubina è trasformata in urobilinogeno e urobilina che passa
nelle urine (gialla)
Se vi sono troppi pigmenti biliari in circolo � ITTERO
![Page 4: 12.metabolismo lipidi [modalit compatibilit ] · punto di regolazione metabolica. Regolazione covalente mediante fosforilazione ... della biosintesi degli acidi grassi e del colesterolo,](https://reader031.fdocumenti.com/reader031/viewer/2022022709/5c65ab8009d3f2a86e8cf785/html5/thumbnails/4.jpg)
![Page 5: 12.metabolismo lipidi [modalit compatibilit ] · punto di regolazione metabolica. Regolazione covalente mediante fosforilazione ... della biosintesi degli acidi grassi e del colesterolo,](https://reader031.fdocumenti.com/reader031/viewer/2022022709/5c65ab8009d3f2a86e8cf785/html5/thumbnails/5.jpg)
![Page 6: 12.metabolismo lipidi [modalit compatibilit ] · punto di regolazione metabolica. Regolazione covalente mediante fosforilazione ... della biosintesi degli acidi grassi e del colesterolo,](https://reader031.fdocumenti.com/reader031/viewer/2022022709/5c65ab8009d3f2a86e8cf785/html5/thumbnails/6.jpg)
![Page 7: 12.metabolismo lipidi [modalit compatibilit ] · punto di regolazione metabolica. Regolazione covalente mediante fosforilazione ... della biosintesi degli acidi grassi e del colesterolo,](https://reader031.fdocumenti.com/reader031/viewer/2022022709/5c65ab8009d3f2a86e8cf785/html5/thumbnails/7.jpg)
![Page 8: 12.metabolismo lipidi [modalit compatibilit ] · punto di regolazione metabolica. Regolazione covalente mediante fosforilazione ... della biosintesi degli acidi grassi e del colesterolo,](https://reader031.fdocumenti.com/reader031/viewer/2022022709/5c65ab8009d3f2a86e8cf785/html5/thumbnails/8.jpg)
Acidi grassi
Componenti deilipidi di riserva (triacilgliceroli)lipidi di membrana
Molecole combustibili infegatomuscolo scheletricocuore
I LIPIDI ALIMENTARI FORNISCONO ALL’ORGANISMOca IL 25% DELLE CALORIE NECESSARIE GIORNALMENTEinfatti l’ossidazione degli acidi grassi si verifica in quasi tutti i tessuti (a parte cervello, tessuto nervoso e globuli rossi) e soprattutto nel cuore.
![Page 9: 12.metabolismo lipidi [modalit compatibilit ] · punto di regolazione metabolica. Regolazione covalente mediante fosforilazione ... della biosintesi degli acidi grassi e del colesterolo,](https://reader031.fdocumenti.com/reader031/viewer/2022022709/5c65ab8009d3f2a86e8cf785/html5/thumbnails/9.jpg)
![Page 10: 12.metabolismo lipidi [modalit compatibilit ] · punto di regolazione metabolica. Regolazione covalente mediante fosforilazione ... della biosintesi degli acidi grassi e del colesterolo,](https://reader031.fdocumenti.com/reader031/viewer/2022022709/5c65ab8009d3f2a86e8cf785/html5/thumbnails/10.jpg)
Fonti di acidi grassi
•Lipidi di deposito (trigliceridi, tessuto adiposo)
•Lipidi sintetizzati dal fegato ed esportati
•Lipidi introdotti con la dieta
•Eccesso di proteine e carboidrati (dieta)
![Page 11: 12.metabolismo lipidi [modalit compatibilit ] · punto di regolazione metabolica. Regolazione covalente mediante fosforilazione ... della biosintesi degli acidi grassi e del colesterolo,](https://reader031.fdocumenti.com/reader031/viewer/2022022709/5c65ab8009d3f2a86e8cf785/html5/thumbnails/11.jpg)
Il livello degli acidi grassi liberi nel plasma (FFA o NEFA) è sottoposto a controllo ormonale mediante regolazione della lipolisi nel tessuto adiposo
Il grasso viscerale, compreso quello che si può depositare nel fegato, cuore e muscoli è un tessuto adiposo “disfunzionale”, iperlipolitico e resistente all’effetto anti-lipolitico dell’insulina.
![Page 12: 12.metabolismo lipidi [modalit compatibilit ] · punto di regolazione metabolica. Regolazione covalente mediante fosforilazione ... della biosintesi degli acidi grassi e del colesterolo,](https://reader031.fdocumenti.com/reader031/viewer/2022022709/5c65ab8009d3f2a86e8cf785/html5/thumbnails/12.jpg)
Gli ormoni che principalmente regolano il metabolismo energetico sono:
i polipeptidi insulina (segnale dell’avvenuta assunzione di cibo e quindisegnale di alto livello di glucoso nel sangue) e glucagone (segnale di basso livello di glucoso nel sangue, quindi nel digiuno ; il suo livello può essere alto anche nel diabete inizialmente poiché l’insulina normalmente inibisce la sua liberazione) il glucorticoide cortisolo (aumenta la produzione di glucoso via gluconeogenesi)la catecolamina adrenalina (segnale di eccitazione e quindi segnale che energia, e quindi anche glucoso, è immediatamente necessaria)
Ma anche altri ormoni influenzano i livelli di glucosio ematico
+++ secreti dall’ipofisi anteriore: ACTH e GH per es. La somministrazione prolungata di GH porta al diabete in quanto, determinando iperglicemia, stimola la secrezione diinsulina causando a lungo andare l’esaurimento delle cell. Beta del pancreas endocrino (secernenti insulina)
+++ ormone tiroideo
L’ipoglicemia prolungata deve essere evitata perché potenzialmente letale per il cervello
![Page 13: 12.metabolismo lipidi [modalit compatibilit ] · punto di regolazione metabolica. Regolazione covalente mediante fosforilazione ... della biosintesi degli acidi grassi e del colesterolo,](https://reader031.fdocumenti.com/reader031/viewer/2022022709/5c65ab8009d3f2a86e8cf785/html5/thumbnails/13.jpg)
EFFETTO DELL’INSULINA SULL’UTILIZZAZIONE DEL GLUCOSIO
energia (glicolisi)50%
40%Glucosio ingerito trasformato in grassi
10%trasformato in glicogeno
L’insulina:incrementa la glicolisi epatica (stimolando gli enz. chiavecome esochinasi) e quindi l’utilizzo del glucosio (quindiabbassa la glicemia)diminuisce l’attività della glucoso-6-fosfato fosfatasi epaticaquindi il glucosio non può essere liberato nel sangue(sempre abbasssamento della glicemia)
![Page 14: 12.metabolismo lipidi [modalit compatibilit ] · punto di regolazione metabolica. Regolazione covalente mediante fosforilazione ... della biosintesi degli acidi grassi e del colesterolo,](https://reader031.fdocumenti.com/reader031/viewer/2022022709/5c65ab8009d3f2a86e8cf785/html5/thumbnails/14.jpg)
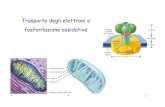
L’enzima acil CoA sintetasi (= acido grasso tiokinasi) sta sulla membrana mitocondriale est.Accoppiata avviene una reazione catalizzata da una pirofosfatasi:PPi 2 Pi (altamente esoergonica) che sposta l’equilibrio della reaz. globale della Acil CoA sintasi verso destra Quindi vengono rotti 2 legami fosfoanidridici ad alta energia(altro motivo ricorrente nelle reazioni biosintetiche)
![Page 15: 12.metabolismo lipidi [modalit compatibilit ] · punto di regolazione metabolica. Regolazione covalente mediante fosforilazione ... della biosintesi degli acidi grassi e del colesterolo,](https://reader031.fdocumenti.com/reader031/viewer/2022022709/5c65ab8009d3f2a86e8cf785/html5/thumbnails/15.jpg)
Gli ac. grassi con - di 12 C possono attraversare la m. mitoc. int. senza l’aiuto della carnitina. Gli altri:
La carnitina è un aminoacido non
proteico sintetizzato nel fegato e nel
rene a partire da lys e met. Il 97%
però di carnitina si trova nei muscoli
scheletrici. Negli alimenti si trova
soprattutto nella carne.
![Page 16: 12.metabolismo lipidi [modalit compatibilit ] · punto di regolazione metabolica. Regolazione covalente mediante fosforilazione ... della biosintesi degli acidi grassi e del colesterolo,](https://reader031.fdocumenti.com/reader031/viewer/2022022709/5c65ab8009d3f2a86e8cf785/html5/thumbnails/16.jpg)
+ 2 e
![Page 17: 12.metabolismo lipidi [modalit compatibilit ] · punto di regolazione metabolica. Regolazione covalente mediante fosforilazione ... della biosintesi degli acidi grassi e del colesterolo,](https://reader031.fdocumenti.com/reader031/viewer/2022022709/5c65ab8009d3f2a86e8cf785/html5/thumbnails/17.jpg)
![Page 18: 12.metabolismo lipidi [modalit compatibilit ] · punto di regolazione metabolica. Regolazione covalente mediante fosforilazione ... della biosintesi degli acidi grassi e del colesterolo,](https://reader031.fdocumenti.com/reader031/viewer/2022022709/5c65ab8009d3f2a86e8cf785/html5/thumbnails/18.jpg)
β ossidazione acidi grassi(n° pari atomi C)
rendimento in ATP
Per ogni ciclo di beta- ossidazione (4 reazioni):1 Acetil CoA1 FADH21 NADH
Per ogni acetile che entra nel ciclo dell’acido cit rico3 NADH x 2,5 ATP = 7,5 ATP1 FADH2 x 1,5 ATP = 1,5 ATP1 GTP (ATP) = 1 ATPTotale = 10 ATP
Esempio: Acido palmitico (saturo, 16 atomi C )7 cicli di β ossidazione -8 acetil CoA (x 10 ATP) → 80 ATP- 7 FADH2 (x 1,5 ATP) → 10,5 ATP- 7 NADH (x 2,5 ATP) → 17,5 ATP
108 ATP-2 ATP (utilizzati per attivare l’ acido grasso)
totale 106 ATP
Contro circa 30 ATP
prodotti dalla respirazione
partendo da 1 glucosio
![Page 19: 12.metabolismo lipidi [modalit compatibilit ] · punto di regolazione metabolica. Regolazione covalente mediante fosforilazione ... della biosintesi degli acidi grassi e del colesterolo,](https://reader031.fdocumenti.com/reader031/viewer/2022022709/5c65ab8009d3f2a86e8cf785/html5/thumbnails/19.jpg)
Degradazione acidi grassi(n°dispari atomi di carbonio)
intermedio ciclo di Krebs
Beta ossidazione
Propionil CoA Acetil CoA
![Page 20: 12.metabolismo lipidi [modalit compatibilit ] · punto di regolazione metabolica. Regolazione covalente mediante fosforilazione ... della biosintesi degli acidi grassi e del colesterolo,](https://reader031.fdocumenti.com/reader031/viewer/2022022709/5c65ab8009d3f2a86e8cf785/html5/thumbnails/20.jpg)
Corpi chetonici
• I corpi chetonici vengono sintetizzati nei mitocondri del fegato a partire da acetil-CoA prodotto dalla β-ossidazione degli acidi grassi.
![Page 21: 12.metabolismo lipidi [modalit compatibilit ] · punto di regolazione metabolica. Regolazione covalente mediante fosforilazione ... della biosintesi degli acidi grassi e del colesterolo,](https://reader031.fdocumenti.com/reader031/viewer/2022022709/5c65ab8009d3f2a86e8cf785/html5/thumbnails/21.jpg)
![Page 22: 12.metabolismo lipidi [modalit compatibilit ] · punto di regolazione metabolica. Regolazione covalente mediante fosforilazione ... della biosintesi degli acidi grassi e del colesterolo,](https://reader031.fdocumenti.com/reader031/viewer/2022022709/5c65ab8009d3f2a86e8cf785/html5/thumbnails/22.jpg)
![Page 23: 12.metabolismo lipidi [modalit compatibilit ] · punto di regolazione metabolica. Regolazione covalente mediante fosforilazione ... della biosintesi degli acidi grassi e del colesterolo,](https://reader031.fdocumenti.com/reader031/viewer/2022022709/5c65ab8009d3f2a86e8cf785/html5/thumbnails/23.jpg)
![Page 24: 12.metabolismo lipidi [modalit compatibilit ] · punto di regolazione metabolica. Regolazione covalente mediante fosforilazione ... della biosintesi degli acidi grassi e del colesterolo,](https://reader031.fdocumenti.com/reader031/viewer/2022022709/5c65ab8009d3f2a86e8cf785/html5/thumbnails/24.jpg)
Biosintesi degli acidi grassi
1. La sintesi ha luogo nel citoplasma(la degradazione nella matrice mitocondriale).
2. Utilizza acetil CoA come precursore.3. Il riducente nella sintesi è il NADPH (gli
ossidanti nella degradazione sono il NAD+ e il FAD).
5. Enzima: acido grasso sintetasi.6. La sintesi è particolarmente attiva nel:-fegato-tessuto adiposo-ghiandola mammaria
![Page 25: 12.metabolismo lipidi [modalit compatibilit ] · punto di regolazione metabolica. Regolazione covalente mediante fosforilazione ... della biosintesi degli acidi grassi e del colesterolo,](https://reader031.fdocumenti.com/reader031/viewer/2022022709/5c65ab8009d3f2a86e8cf785/html5/thumbnails/25.jpg)
La biosintesi degli acidi grassi saturi fino a 16 C negli animali e nel
lievito avviene nel citosol, è una biosintesi riduttiva che utilizza il
NADPH come cofattore riducente, che nel citosol si trova in un rapporto
di circa 75 sul NADP, quindi alto rispetto al NADH il cui rapporto sul
NAD è invece 8 x 10-4.
Il NADPH necessario proviene dalla via del pentoso fosfato.
I substrati iniziali della biosintesi sono acetilCoA e malonilCoA
All’acido grasso in allungamento vengono poi aggiunte, una alla volta,altre
molecole di malonilCoA, l’allungamento però è di 2 C alla volta x cui a ogni
ciclo viene liberata CO2
O
S
O
O-
CoA
![Page 26: 12.metabolismo lipidi [modalit compatibilit ] · punto di regolazione metabolica. Regolazione covalente mediante fosforilazione ... della biosintesi degli acidi grassi e del colesterolo,](https://reader031.fdocumenti.com/reader031/viewer/2022022709/5c65ab8009d3f2a86e8cf785/html5/thumbnails/26.jpg)
Il C metilenico del malonile, trovandosi tra un
carbonile e un carbossile, può essere un buon
nucleofilo che attacca il C=O dell’acetile nella fase
di condensazione.
La decarbossilazione del malonile facilita la
condensazione.
Per la sintesi di un palmitato occorrono quindi
1 Acetil CoA, 7 malonil CoA, 14 NADPH e oltre al
palmitato si producono 7 CO2, 8 CoA e 14 NADP,
non tenendo conto della stechiometria degli H+ e
delle molecole d’acqua che si producono.
![Page 27: 12.metabolismo lipidi [modalit compatibilit ] · punto di regolazione metabolica. Regolazione covalente mediante fosforilazione ... della biosintesi degli acidi grassi e del colesterolo,](https://reader031.fdocumenti.com/reader031/viewer/2022022709/5c65ab8009d3f2a86e8cf785/html5/thumbnails/27.jpg)
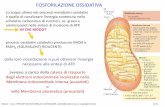
Il malonilCoA è prodotto a partire da acetilCoA e bicarbonato nella
reazione catalizzata dall’Acetil-CoA carbossilasi, reazione in cui è
necessario ATP x l’attivazione del bicarbonato e reazione che limita la
velocità della sintesi degli acidi grassi. L’enzima è infatti un importante
punto di regolazione metabolica.
Regolazione covalente mediante fosforilazione
inibisce l’entrata degli acili dentro il mitocondrio x la ββββ ossidazione
![Page 28: 12.metabolismo lipidi [modalit compatibilit ] · punto di regolazione metabolica. Regolazione covalente mediante fosforilazione ... della biosintesi degli acidi grassi e del colesterolo,](https://reader031.fdocumenti.com/reader031/viewer/2022022709/5c65ab8009d3f2a86e8cf785/html5/thumbnails/28.jpg)
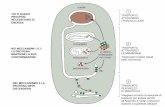
La membr. mitocondriale int. è impermeabile all’AcetilCoA, sicchè
l’Acetil CoA, per uscire dal mitocondrio e poter essere substrato
della biosintesi degli acidi grassi e del colesterolo, prima condensa
con l’ossalacetato a formare il citrato nella 1° reaz. del ciclo di
Krebs, poi attraverso un suo trasportatore viene portato nel
citosol, dove la citrato liasi lo scinde consumando ATP e CoA a dare
nuovamente Acetil CoA e ossalacetato.
![Page 29: 12.metabolismo lipidi [modalit compatibilit ] · punto di regolazione metabolica. Regolazione covalente mediante fosforilazione ... della biosintesi degli acidi grassi e del colesterolo,](https://reader031.fdocumenti.com/reader031/viewer/2022022709/5c65ab8009d3f2a86e8cf785/html5/thumbnails/29.jpg)
Quando c’è molto glucosio avviene
•sia la sintesi degli acidi grassi
•che la sintesi del colesterolo, sempre a
partire da AcetilCoA
![Page 30: 12.metabolismo lipidi [modalit compatibilit ] · punto di regolazione metabolica. Regolazione covalente mediante fosforilazione ... della biosintesi degli acidi grassi e del colesterolo,](https://reader031.fdocumenti.com/reader031/viewer/2022022709/5c65ab8009d3f2a86e8cf785/html5/thumbnails/30.jpg)