






Le lingue
Pagine
Legale


;
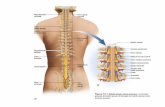
1 MIDOLLO SPINALE Il midollo spinale innerva il tronco e gli arti attraverso i nervi spinali e le loro ramificazioni periferiche, riceve le fibre afferenti primarie dai recettori periferici estesamente localizzati nelle strutture somatiche, viscerali e invia fibre motorie ai muscoli scheletrici. Contiene inoltre i corpi cellulari di tutti i neuroni pregangliari responsabili dell’innervazione del muscolo cardiaco, della muscolatura liscia e delle ghiandole nonché i neuroni pregangliari per l’innervazione parasimpatica della muscolatura liscia dei visceri. Il midollo spinale occupa i due terzi superiori del canale vertebrale; si continua cranialmente con il bulbo e si restringe caudalmente nel cono midollare, dal cui apice discende un filamento di tessuto connettivo, il filo terminale, che raggiunge la por zione dorsale del primo segmento vertebrale coccigeo. Il midollo si estende dal margine superiore dell’atlante alla giunzione tra la 1
a e la 2
a vertebra lombare.
Durante lo sviluppo, la colonna vertebrale si allunga più rapidamente del midollo spinale, per tale motivo esiste una discrepanza crescente tra il livello anatomico dei segmenti midollari e le loro vertebre corrispondenti.
Al 23° giorno, la colonna vertebrale e il midollo spinale si trovano allo stesso livello e il midollo termina a livello dell’ultima vertebra coccigea; questa disposizione permane fino al 3° mese di vita fetale. Alla nascita, il midollo termina a livello del margine inferiore della 2
a vertebra lombare, talvolta può raggiungere la 3
a vertebra lombare.
Nell’adulto, si ritiene che il midollo spinale termini a livello del disco intervertebrale tra il corpo della 1a e della 2
a vertebra lombare, che
si trova un poco sopra al livello della piega del gomito in adduzione, cioè a lato del corpo. Tuttavia, esiste una grande variabilità per quanto riguarda il livello dove termina il midollo spinale (1
a ‐ 2
a vertebra lombare; 12
a vertebra toracica; 2
a ‐3
a vertebra lombare.
Il midollo spinale varia anche in riferimento al suo diametro trasversale affusolandosi craniocaudalmente e presentando dei rigonfiamenti.
Il rigonfiamento cervicale è l’origine dei nervi spinali che innervano gli arti superiori; esso si estende da C3 a T2. Il rigonfiamento lombare è l’origine dei nervi spinali che innervano gli arti inferiori; esso si estende da L1 a S3. Le fessure e i solchi si estendono lungo la maggior parte della superficie esterna. Una fessura mediana anteriore, un solco mediano posteriore e un setto dividono quasi nettamente il midollo in una metà destra e sinistra. La fessura mediana anteriore si estende lungo l’intera superficie ventrale; il solco mediano posteriore si trova dorsalmente; lateralmente esistono due solchi posterolaterali come anteriormente esistono due solchi anterolaterali.
RADICI DORSALI E VENTRALI Le radici dorsali e ventrali dei nervi spinali sono in continuità con il midollo spinale. Attraversano lo spazio subaracnoideo e la dura madre separatamente, unendosi nei pressi o all’interno dei fori intervertebrali per formare i nervi spinali, che sono tutti misti (contengono la componente motrice somatica, sensitiva somatica e viscerale, sia sensitiva che motoria insieme alla componente autonoma). Poiché il midollo spinale è più breve della colonna vertebrale, le radici spinali più caudali discendono a tratti di varia lunghezza attorno ed oltre il midollo per raggiungere i fori corrispondenti. Per questo formano un fascio divergente di radici dei nervi spinali, la cauda equina, che si raccoglie attorno al filo terminale. Le radici ventrali dei nervi spinali sono motorie, mentre quelle dorsali sono sensitive.
MENINGI Le meningi sono tre e a partire dall’esterno verso l’interno del cranio e della colonna vertebrale sono distinte in: dura madre, aracnoide e pia madre. DURA MADRE In alcune aree all’interno del cranio la dura madre può essere distinta dall’endosti, ma a livello della base cranica i due strati sono fusi e aderenti all’osso. All’interno della colonna vertebrale, la dura madre è separata dai tessuti che rivestono il canale vertebrale ed è separata da essi da uno spazio detto spazio epidurale. L’accesso a questo spazio, di solito nella regione lombare, è indispensabile per la somministrazione di anestetici e analgesici e per l’endoscopia. Lo spazio caudale è usato maggiormente per le iniezioni di analgesici. Lo spazio subdurale è uno spazio che si può creare nel midollo normale poiché l’aracnoide e la dura sono strettamente connesse. ARACNOIDE L’aracnoide spinale è in continuità con quella cranica; è separata dalla terza meninge da uno spazio detto spazio subaracnoideo dove passa il LCR. PIA MADRE La pia madre è l’ultima meningi a contatto con il midollo spinale e si approfonda nelle fessure presenti a livello midollare.

;
2 ORGANIZZAZIONE INTERNA In sezione trasversale. Il midollo spinale è diviso in maniera incompleta in due metà simmetriche da un solco mediano posteriore e da una fessura mediana anteriore. Il midollo spinale è costituito da una porzione esterna di sostanza bianca e da una parte centrale di sostanza grigia. La quantità di sostanza grigia riflette il numero di neuroni presenti, essendo proporzionalmente maggiore a livello dei rigonfiamenti cervicale e lombare contenente i neuroni che innervano gli arti. La quantità di sostanza bianca è maggiore a livello cervicale e diminuisce progressivamente ai livelli inferiori poiché i fasci discendenti perdono fibre in senso caudale mentre quelli ascendenti ne accumulano in senso craniale. Al centro della sostanza grigia del midollo spinale e per tutta la lunghezza del midollo spinale, si estende un minuscolo canale centrale rivestito da un epitelio cilindrico ciliato (ependima) e contenente il LCR.
SOSTANZA GRIGIA DEL MIDOLLO SPINALE La sostanza grigia presenta una forma a farfalla o a “H”. E’ costituita dai corpi cellulari e dai dendriti dei neuroni multipolari; i neuroni possono essere intersegmentali, cioè contenuti all’interno di un singolo neuromero, o intersegmentali se le loro ramificazioni si prolungano lungo più neuromeri. Essa consta di quattro masse cellulari collegate far loro, i due corni anteriori e i due corni posteriori, che si proiettano verso la parte ventrale e dorsale del midollo spinale. La sostanza grigia che circonda il canale centrale e unisce le due metà del midollo costituisce le commissure grigie anteriore e posteriore. Il corno posteriore è il punto i n cui terminano le fibre afferenti , entrate nel midollo spinale tramite le radici posteriori dei nervi spinali. Il corno anteriore contiene neuroni efferenti i cui assoni si portano nelle radici anteriori dei nervi spinali , mentre a livello toracico e lombare superiore è presente n piccolo corno laterale, che contiene i corpi cellulari dei neuroni pregangliari.
GRUPPI CELLULARI NEURONALI DEL MIDOLLO SPINALE In sezione trasversale, la sostanza grigia è organizzata in dieci strati cellulari, noti come lamine di Rexed. Tali lamine , definite sulla base della morfologia, della grandezza, delle caratteristiche citologiche e della densità dei neuroni, sono numerate secondo un ordine postero‐anteriore. Le lamine I‐IV si trovano nella parte dorsale del corno posteriore e rappresentano il principale punto di terminazione delle fibre afferenti cutanee e dei loro collaterali.
CORNO POSTERIORE E’ un’area di terminazione delle fibre afferenti che entrano nel midollo attraverso le radici posteriori dei nervi spinali. Ad esempio la colonna di Clarke si trova alla base del corno posteriore. Lungo la maggior parte del midollo spinale, la colonna di Clarke, procede vicina al cordone posteriore, a cui può inviare alcune proiezioni. Inizia dall’ottavo neuromero cervicale e il terzo o il quarto neuromero lombare.
CORNO LATERALE Il corno laterale è un piccolo processo laterale della sostanza grigia, localizzato fra il corno posteriore e quello anteriore. Inizia dall’ottavo neuromero cervicale o dal primo toracico e arriva al secondo o terzo neuromero lombare. Al suo interno sono contenuti i corpi cellulari dei neuroni pregangliari, i quali si sviluppano nel midollo spinale embrionale dorsolateralmente al canale centrale per poi spostarsi lateralmente. Gli assoni dei neuroni del corno laterale passano nelle radici spinali anteriori e nei rami comunicanti bianchi per raggiungere il tronco del simpatico. Un sistema simile può essere individuato a livello sacrale, ma, a differenza della colonna toracolombare, non forma un processo laterale ben evidente e dà origine alle fibre nervose pregangliari del parasimpatico.

;
3 CORNO ANTERIORE Il corno anteriore presenta neuroni di dimensione variabile; i corpi cellulari più grandi sono quelli dei motoneuroni α, i cui assoni escono dalle radici anteriori per innervare le fibre extrafusali dei muscoli scheletrici; vi sono anche corpi cellulari più piccoli tra cui quelli dei motoneuroni γ che innervano le fibre intrafusali dei fusi neuromuscolari. I motoneuroni del corno anteriore seguono un’ organizzazione somatotopica. La disposizione di base prevede che il gruppo cellulare mediale innervi la muscolatura assiale, mentre quello laterale la muscolatura appendicolare.
La colonna motrice mediale si estende lungo tutto il midollo spinale e i suoi neuroni innervano i gruppi muscolari epiassiali e ipoassiali. In particolare, i muscoli epiassiali includono il muscolo erettore della colonna (che estendono testa e colonna vertebrale), mentre i muscoli ipoassiali i muscoli prevertebrali, del collo, i muscoli intercostali e i muscoli della parete anteriore dell’addome (che flettono il collo e il tronco). I muscoli epiassiali sono innervati da rami delle radici dorsali dei nervi spinali, mentre gli ipoassiali da rami delle radici ventrali dei nervi spinali. I motoneuroni che innervano i muscoli epiassiali sono situati in posizione ventrale, mentre quelli che innrvano la muscolatura ipoassiale si trovano in posizione dorsale. La colonna motrice laterale si trova solo a livello dei rigonfiamenti cervicale e lombare del midollo spinale, dove è indispensabile per l’innervazione degli arti. ORGANIZZAZIONE SOMATOTOPICA DEI MOTONEURONI:
Arto superiore: a)I motoneuroni che innervano i muscoli intrinseci dell’arto superiore si trovano in posizione dorsale rispetto alla colonna motrice laterale e quelli che innervano i muscoli della mano si trovano ancora più dorsalmente;
I motoneuroni che innervano i muscoli estrinseci dell’arto superiore; quelli del cingolo scapolare sono invece nella parte ventrale.
b) i muscoli prossimali degli arti sono innervati dai motoneuroni più rostrali del rigonfiamento corrispondente rispetto a quelli che innervano i muscoli distali. Ad esempio i motoneuroni che innervano i muscoli della mano si trovano nei neuromeri C8‐T1, mentre quelli che innervano i muscoli della spalla sono localizzati ai livelli C5‐C6.
Arto inferiore:
I motoneuroni che innervano l’arto inferiore mostrano una disposizione analoga a livello del midollo lombosacrale
RIFLESSI SPINALI Le connessioni del midollo spinale e del tronco encefalico entrano in gioco in numerosi riflessi attraverso i quali le funzioni periferiche sono modulate e regolate in maniera automatica o autonoma in risposta agli stimoli in arrivo. Si parla di archi riflessi diretti quando non sono presenti interneuroni fra il neurone afferente ed efferente; in caso contrario gli archi riflessi sono definiti come indiretti. Questi archi sono controllati da vie discendenti che provengono sia dalla corteccia cerebrale, sia dal tronco encefalico. Alterazioni patologiche dei fasci discendenti causano alterazioni di questi riflessi, che sono, routinariamente, testati all’esame neurologico. Durante lo sviluppoi meccanismi di controllo discendenti sopprimono quelli denominati riflessi spinali primitivi , come il riflesso estensorio plantare e quello di prensione. Quando i meccanismi di controllo sono danneggiati, questi riflessi si riattivano e ricompaiono come segni di patologia del SNC (ad esempio il riflesso di Babinski). RIFLESSO DA STIRAMENTO E’ il riflesso più semplice essendo mediato solamente da un neurone afferente e da un neurone efferente. La componente afferente origina da recettori di stiramento associati alle fibre muscolari intrafusali localizzate all’interno dei fusi neuromuscolari. Le terminazioni sensitive di queste cellule recettoriali danno origine a fibre afferenti che entrano nel midollo spinale, ove prendono contatto diretto con i motoneuroni α, innervanti il medesimo muscolo. I motoneuroni α dei muscoli antagonisti sono simultaneamente inibiti da connessioni collaterali inibitorie da parte di interneuroni. Riflesso gamma Così come i motoneuroni α innervano le fibre extrafusali, le fibre intrafusali sono innervate da motoneuroni γ. La loro attivazione aumenta la sensibilità allo stiramento e sul tono muscolare. Come i motoneuroni α, i motoneuroni γ, sono influenzati da fasci discendenti che originano nel tronco encefalico e nella corteccia cerebrale. Nelle patologie che coinvolgono il SNC e il SNP determinano cambiamenti nei riflessi e nel tono muscolare. Riflesso da flessione Stimoli dolorosi agli arti determinano un riflesso di allontanamento flessorio mediato da un riflesso polisinaptico, nel quale i neuroni afferenti ed efferenti sono mediati da interneuroni. L’attivazione dei nocicettori va quindi ad attivare indirettamente l’attivazione dei motoneuroni per i muscoli flessori dell’arto. L’attivazione della flessione può avvenire a diversi livelli sulla base dell’intensità dello stimolo. Le connessioni che decussano nel lato opposto del midollo spinale attivano i motoneuroni estensori e producono il cosiddetto riflesso estensorio crociato. Normalmente tutti gli stimoli cutanei provocano un riflesso flessorio, ma tranne che per gli stimoli dolorifici, questi riflessi vengono inibiti da fasci discendenti. In seguito alla perdita di tali connessioni inibitorie, anche uno stimolo innocuo può causare un riflesso flessorio. Ricordare che il riflesso di Babinski è un riflesso estensorio plantare, che fa seguito alla risposta flessoria di allontanamento dell’arto inferiore in stimolazione alla pianta del piede.

;
4 SOSTANZA BIANCA DEL MIDOLLO SPINALE La sostanza bianca del midollo spinale circonda la parte centrale di sostanza grigia
VIE ASCENDENTI Colonna dorsale Il cordone posteriore è costituito da due grossi fasci ascendenti, il fascicolo gracile e cuneato, denominati anche colonne dorsali.
Il fascicolo gracile origina a livello dell’estremità caudale del midollo spinale e continua fino a livello cervicale dove contiene una più alta percentuale di fibre afferenti di origine cutanea più che di quelli propriocettive in quanto la maggior parte di queste abbandonano il fascicolo nei metameri inferiori per contrarre sinapsi con i neuroni della colonna di Clarke. La sensibilità propriocettiva dell’arto inferiore infatti raggiunge il talamo passando per la colonna di Clarke.
Gli assoni del fascicolo gracile terminano nel nucleo gracile nella parte posteriore mediale del bulbo. Il fascicolo cuneato origina a livello medio toracico ed è localizzato lateralmente al fascicolo gracile. E’ composto per la maggior parte da
fibre afferenti primarie delle radici posteriori toraciche e cervicale ed a livello cervicale contiene afferenze che provengono principalmente dall’arto superiore e dai recettori cutanei e profondi.
Molti assoni che salgono nel fascicolo cuneato terminano nel nucleo cuneato nella parte posteriore laterale del bulbo rispetto al nucleo gracile. Alcuni terminano nel nucleo cuneato laterale (accessorio o esterno); i neuroni che originano da questo nucleo si proiettano al cervelletto attraverso il fascio cuneocerebellare.
Gli assono che originano dai nuclei gracile e cuneato della colonna dorsale si incrociano nella decussazione sensitiva (decussazione del lemnisco mediale) andando a formare il lemnisco mediale. Quindi si portano al nucleo ventrale posterolaterale del talamo e da qui altri neuroni si proiettano alla corteccia somatosensitiva del lobo parietale. Alcuni neuroni della colonna dorsale formano le fibre arcuate esterne che si portano al cervelletto. L’alto grado di organizzazione somatotopica presente nella colonna dorsale è mantenuta anche quando i fasci raggiungono la corteccia somatosensitiva. I fasci che originano dal nucleo gracile trasportano la sensibilità dell' arto inferiore, quelli del nucleo cuneato l’arto superiore e la sensibilità del tronco è in una posizione intermedia tra i due. Per quanto riguarda la sensibilità cutanea e profonda, i fasci che trasportano la prima sono superficiali rispetto a quelli che trasportano la sensibilità profonda (recettori tattili vibrazioni).
Fasci spinocerebellari Vi sono due fasci spinocerebellari: dorsale e ventrale. Essi trasportano una sensibilità cutanea (esterocettiva) e propriocettiva al cervelletto per la coordinazione del movimento. Le fibre mieliniche di grosso calibro sono associate al fascio spinocerebellare dorsale, quelle di piccolo calibro al fascio spinocerebellare ventrale. I due fasci trasportano informazioni che provengono dal tronco e dagli arti inferiori; l’equivalente del fascio spinocerebellare dorsale per l’arto superiore è il fascio cuneocerebellare: questo perché la Colonna di Clarke in cui i due fasci spinocerebellari fanno sinapsi non è più presente a livello cervicale. Il fascio spinocerebellare dorsale origina ai neuroni della colonna di Clarke, alla base del corno posteriore ei neuromeri toracici e raggiunge i nuclei cerebellari per lo più omolaterali trasportando, quindi, informazioni omolaterali; il fascio spinocerebellare ventrale si sposta controlateralmente nel midollo spinale, ma gran parte delle fibre si incrocia nuovamente nella sostanza bianca del cervelletto in modo che la distribuzione delle fibre ai nuclei cerebellari e alla corteccia cerebellare ricalchi quelle delle fibre del fascio dorsale (informazioni omolaterali). Anche il fascio cuneocerebellare è principalmente non crociato.
Fasci spinotalamici I fasci spinotalamici sono costituiti da neuroni che trasmettono la sensibilità termica, dolorifica e tattile. Le cellule di origine si trovano in varie lamine di tutti i neuromeri del midollo spinale e decussano nella commissura ventrale della sostanza bianca per proseguire come fasci spinotalamici laterali. Mentre i fasci che trasportano una sensibilità termodolorifica decussano subito, quelli che trasportano altre sensibilità possono salire per alcuni neuromeri prima di decussare. SI possono (per alcuni studiosi) distinguere due fasci: fascio spinotalamico anteriore e fascio spinotalamico laterale.
Il fascio spinotalamico laterale è situato nel cordone laterale, medialmente rispetto al fascio spinocerebellare ventrale. L’evidenza clinica indica che esso trasporta una sensibilità termodolorifica. Essi raggiungono il nucleo ventrale posterolaterale del talamo e continuano per la corteccia somatosensitiva nel lobo parietale del telencefalo. Il fascio spinotalamico anteriore è localizzato nel cordone anteriore e sulla base dell’evidenza clinica provvede alla sensibilità tattile protopatica e alla barestesia. . Essi raggiungono il nucleo ventrale posterolaterale del talamo dopo aver decorso parallelamente al lemnisco mediale e continuano per la corteccia somatosensitiva nel lobo parietale del telencefalo. Meccanismi del dolore Le connessioni ascendenti sono soggette a controllo e modulazione ad opera di connessioni infraspinali e di vie discendenti dal tronco encefalico e dalla corteccia cerebrale. Questo è importaste ad esempio per la percezione del dolore. La teoria del controllo del cancello propone un controllo delle vie in entrata nocicettive e di altro tipo. Considerando questa teoria le fibre afferenti di grosso calibro provenienti dai recettori tattili e dai recettori annessi ai peli eccitano si a i grossi neuroni della lamina IV, da cui originano le fibre spinotalamiche sia gli interneuroni della sostanza gelatinosa di Rolando, mentre le fibre afferenti amieliniche di piccolo calibro eccitano la lamina IV ma inibiscono gli interneuroni. Si presume che gli interneuroni inibiscano tutte le fibre afferenti del fascio In un sistema del genere della bassa attività delle fibre di piccolo calibro inibiscono gli interneuroni e prevengono l’inibizione delle cellule del fascio da parte di questi ultimi. Il “cancello” per la lamina IV è aperto alla trasmissione degli impulsi provenienti dalle fibre di grosso calibro. La trasmissione massiva di impulsi da parte delle fibre di grosso calibro determinerebbe prima una eccitazione delle cellule della lamina IV ma successivamente questa attività cesserebbe quando gli interneuroni chiudono il cancello. Il dolore quindi è dovuto ad uno sbilanciamento tra i vari impulsi afferenti, in presenza di un traffico di impulsi sproporzionato lungo le fibre afferenti di piccolo calibro.

;
5 Via spinoreticolare Queste fibre sono intercalate a quelle del fascio spinotalamico e risalgono ventrolateralmente nel midollo spinale. E’ presente in tutti i neuromeri del midollo spinale con prevalenza da parte dei neuromeri cervicali; la via si proietta al nucleo reticolare omo e controlaterale del talamo e sembra avere un ruolo importante nella percezione del dolore.
Via spinomesencefalica La via spinomesencefalica è costituita da fasci ascendenti che vanno dal midollo spinale a varie regioni del mesencefalo. Tra questi fasci i più importanti sono il fascio spinotettale che decorre insieme al fascio spinotalamico laterale, le fibre che terminano nella sostanza grigia periacqueduttale del mesencefalo, fibre che si portano al nucleo parabrachiale, ai nuclei pretettali. Le cellule di origine sono localizzate in tutto il midollo spinale con una prevalenza a livello cervicale e lombosacrale(in particolare lamina V) e sono per la maggior parte controlaterali anche se esiste un contingente ipsilaterale. La maggior parte dei neuroni trasportano informazioni dolorifiche.
Fascio spinoolivare Il fascio origina dalle lamine più profonde della sostanza grigia, risalgono tra il cordone anteriore e il cordone laterale e terminano nei nuclei olivari accessori mediale e dorsale. Trasporta informazioni muscolari propriocettive e di recettori cutanei
VIE DISCENDENTI Le vie discendenti del midollo spinale originano da neuroni della corteccia cerebrale o del tronco encefalico. Sono associate al controllo del tono muscolare, del movimento e della postura.
Fasci corticospinali e corticobulbare Il fascio corticospinale (piramidale) o e corticobulbare (fascio genicolato) originano dai neuroni della corteccia cerebrale e proiettano in massima parte controlateralmente ai neuroni del midollo spinale e del tronco encefalico.
La maggior parte delle fibre corticospinali e corticobulbari originano dal cellule situate nella corteccia dell’area motoria primaria (area 4) e nell’are premotoria (area 6). Le fibre corticospinali e corticobulbari attraversano il braccio posteriore della capsula interna e il suo ginocchio rispettivamente.
Le fibre corticospinali raggiungono il midollo spinale, mentre quelle corticobulbari i nuclei somatomotori del tronco encefalico. Nel bulbo in parte le fibre corticospinali (75‐90% delle fibre) decussano a livello delle piramidi (le piramidi sono prominenze visibili sulla superficie ventrale del bulbo) e vanno a costituire il fascio piramidale crociato che raggiunge il cordone laterale del midollo; la quota di fibre rimanenti non decussa e raggiunge il midollo spinale nel suo cordone anteriore (fascio piramidale diretto).
Il fascio piramidale crociato contiene anche fibre che si spostano controlateralmente nei neuromeri spinali e che raggiungono di conseguenza l’area omolaterale di innervazione.
Il fascio piramidale diretto contiene una quota di fibre che a livello midollare si sposta controlateralmente raggiungendo di conseguenza l’area controlaterale di innervazione.
I due fasci sono associati al movimento preciso degli arti.
Fascio rubrospinale Il fascio rubrospinale origina da neuroni collocati nel nucleo rosso, una porzione ovoidale del tegmento mesencefalico, decussano a livello del tegmento e scendono nel cordone laterale del midollo spinale, anteriormente alle fibre del fascio corticospinale laterale
La sua funzione sembrerebbe simile a quella assunta dalle fibre del fascio corticospinale laterale.
Fasci vestibolospinali I fasci vestibolospinali sono due, il fascio vestibolospinale mediale e il laterale, entrambi originano a livello del solco bulbopontino, nelle pareti laterali e nel pavimento del quarto ventricolo, dove si trovano i nuclei vestibolari.
Il fascio vestibolospinale mediale origina dal nucleo vestibolare mediale, tuttavia alcuni neuroni sono collocati anche nel nucleo vestibolare
inferiore o nel laterale. Il fascio, che per la maggior parte decussa ma presenta anche fibre che restano omolaterali, attraversa il fascicolo longitudinale mediale nel cordone anteriore del midollo spinale e termina in esso sino ai neuromeri mediotoracici,
La funzione del fascio è quella di inibire i motoneuroni che innervano i muscoli assiali del collo e della parte superiore del dorso. Il fascio vestibolospinale laterale origina da neuroni che compongono il nucleo vestibolare laterale o nucleo di Deiters, presso il solco
bulbopontino; discende omolateralmente nella sostanza banca del midollo spinale, poi si sposta nel cordone anteriore del midollo nei neuromeri inferiori.
La funzione del fascio è quella di eccitare i motoneuroni dei muscoli estensori del collo, del dorso e degli arti e, attraverso interneuroni inibitori, di inibire i motoneuroni dei muscoli flessori degli arti corrispondenti.
Fasci reticolospinali Anche i fasci reticolo spinale sono due, il fascio reticolospinale mediale e il laterale, la loro caratteristica comune è la loro origine nella formazione reticolare del tronco. Le loro funzioni sono piuttosto diversificate, aiutano a realizzare i movimenti stereotipati, mantengono il tono muscolare, sono coinvolti nella regolazione della postura e nel controllo della sensibilità dolorifica (in particolare associato al fascio laterale).

;
6 Il fascio reticolospinale mediale origina dalla formazione reticolare mediale pontina e bulbare. Le fibre originatesi dalla formazione
reticolare pontina discendono omolateralmente nel cordone anteriore del midollo spinale, mentre le fibre della formazione reticolare bulbare discendono sia omolateralmente che controlateralmente nel cordone anteriore e in quello laterale. Percorrono tutto il midollo spinale e terminano nei neuromeri lombosacrali, in particolare nelle lamine VII e VIII per le fibre che originano dal ponte e VI e VII per quelle dal bulbo. Sono fibre eccitatorie per i muscoli del tronco e degli arti.
Il fascio reticolo spinale laterale origina dalla formazione reticolare pontina, nel bulbo le sue fibre decussano, portandosi nel cordone laterale controlaterale, poi scendono per l'intera lunghezza del midollo spinale. Terminano nelle lamine I, V e VI della sostanza grigia. Sono fibre eccitatorie per i muscoli del tronco, inibitorie per quelli del collo, sembrano inoltre essere coinvolte
Top Related