







Le lingue
Pagine
Legale


Progetto di Ricerca Giacomo Oliveira
1
Messa a punto di un innovativo modello murino umanizzato per lo studio di fattori prognostici molecolari e di meccanismi di immunoevasione nella
leucemia mieloide acuta: relazione finale
Background e scopo del progetto di ricerca
Sebbene gli studi attuali abbiano permesso di descrivere le mutazioni più frequenti nelle
leucemie mieloidi acute (acute myeloid leukemia, AML), la conoscenza dei meccanismi biologici
che permettono alle cellule leucemiche di crescere eludendo il controllo del sistema immunitario
e di resistere ai diversi approcci terapeutici rimane tuttavia parziale. Ciò è dovuto alla difficoltà di
eseguire studi che permettano di definire l’impatto delle lesione genetiche sui meccanismi
biologici delle cellule leucemiche e di correlare i processi deregolati con la prognosi della
patologia. In aggiunta, la grande variabilità inter-paziente spesso limita l’elucidazione di
meccanismi condivisi da molteplici patologie leucemiche. Gli studi funzionali sono spesso
limitati dall’incapacità di mantenere o crescere in vitro cellule leucemiche primarie, nonché
dall’impossibilità di ricostituire artificialmente il microambiente tumorale. D’altro canto, la
trasferibilità nella pratica clinica delle informazioni ottenute da modelli di leucemia murina è
fortemente limitata dalle evidenti barriere di specie.
Con lo scopo di superare queste limitazioni, questo progetto di ricerca si è rivolto allo
sviluppo di un innovativo modello murino umanizzato per (i) studiare fattori prognostici
molecolari e (ii) caratterizzare i meccanismi di immunoevasione nella leucemia mieloide acuta.
Tale modello si propone di coniugare la riproducibilità e il potere analitico dei modelli animali
con l’alta informatività derivante dall’analisi delle leucemie primarie umane.
Risultati
Identificazione di fattori prognostici della leucemia acuta
Cinquantuno leucemie primarie umane
(Tabella 1) sono state infuse in topi
immunodeficienti NSG (NOD SCID gamma
# AML testate 51
Cellule infuse (x106) 9.5
Deplezione linfociti T 86.3% (44)
origine Midollo: 62.7% (32) Sangue periferico 37.3%(19)
Campione fresco 13.7% (7)
Stato della malattia Diagnosi: 72.5% (38)
Post-chemioterapia: 25.5% (12) Altri: 2% (1)
Tabella 1. Caratteristiche delle leucemie testate.

Progetto di Ricerca Giacomo Oliveira
2
chainnull) di 4-8 settimane non irradiati. I linfociti T contenuti nei campioni primari sono stati
depletati con biglie magnetiche per evitare il loro attecchimento e la conseguente reattività
contro i tessuti murini. Dopo l’infusione, la comparsa di cellule leucemiche sul sangue periferico
è stata quantificata settimanalmente in modo tale da monitorare l’attecchimento. Venticinque
delle 51 AML (49%) hanno generato degli xenotrapianti e sono cresciute nei topi (Fig. 1A): ciò
ha permesso di espandere di circa 5 volte le cellule leucemiche primarie (Fig. 1B), fattore
necessario per la loro caratterizzazione e per il loro impiego nei successivi esperimenti. In
particolare la cinetica di attecchimento si è rivelata riproducibile tra i replicati biologici costituiti
0
20
40
60
80
100
leuk
emic
cel
ls (x
106 )
/ mou
se12.4
63.2
25
1433.46
n = Median Fold Increase =
Median Days =
Infused (iv)Harvested form BMHarvested form Spleen
0 25 50 75 100 125 150 200 250 3000
250
500
750
1000200040006000
Days after infusion
leuk
emic
cells
/ µ
L (m
edia
n)
AML#51AML#50AML#49
AML#47AML#44AML#43
AML#33AML#31AML#29AML#27AML#25AML#23AML#21AML#20AML#17AML#16AML#14AML#12AML#8AML#7
AML#1
AML#40
AML#4
AML#39
AML#48
A B
Primary AML
-3 -2 -1 0 1 2-2
-1
0
1
2
Dimension 1
Dim
ensio
n 2
UPN#1
UPN#8
UPN#16
UPN#12
C
NSG xenografts
0 25 50 75 100 1250
250
500
750
1000200040006000
Days after infusion
leuk
emic
cells
/ µ
L
AML#25
Relapse post alloHSCTDiagnosis
D
Figura 1. Caratterizzazione dell’attecchimento delle leucemie nei topi NSG. (A) Cinetica di crescita delle cellule leucemiche misurata su sangue periferico di topi NSG. Ogni colore indica una leucemia primaria differente testata in diversi topi (n da 1 a 15). Ogni leucemia mostra un profilo di crescita paziente specifico. (B) Quantità di cellule leucemiche purificate infuse per ciascun topo (grigio chiaro) e ricavate dalla milza e dal midollo osseo dei topi al sacrificio (grigio scuro). Le barre mostrano i risultati per le 25 leucemie che hanno attecchito nei topi. L’attecchimento nei topi permette l’espansione delle cellule leucemiche. (C) Cinetica di crescita su sangue periferico delle cellule leucemiche di uno stesso paziente prelevate in due timepoint differenti del suo decorso clinico (giallo=diagnosi, blu=recidiva post trapianto allogenico). La capacità di attecchire in topi viene mantenuta in maniera indipendente dallo stadio della malattia. (D) Analisi del profilo di espressione genica di 4 leucemia alla diagnosi (cerchi vuoti) e dopo attecchimento in topi NSG (cerchi pieni). Mentre leucemie appartenenti a pazienti diversi sono molto eterogene, i campioni derivati dai topi si dispongono vicini alle rispettive leucemie di origine, dimostrando come il loro trascrittoma sia simile alle leucemie primarie parentali.

Progetto di Ricerca Giacomo Oliveira
3
da diversi topi NSG; inoltre la capacità di generare xenotrapianti è risultata caratteristica di ogni
AML e paziente-specifica, dal momento che tale proprietà si è mantenuta costante quanto sono
stati testati differenti campioni provenienti da stadi di malattia diversi dello stesso paziente (Fig.
1C). L’analisi del trascrittoma tramite microarray ha permesso di evidenziare come il profilo di
espressione sia caratteristico di ciascun paziente, ma anche come tale profilo sia riproducibile
negli animali (Fig. 1D). In casi selezionati (n=6), le cellule leucemiche attecchite sono state
reinfuse in altri topi riceventi, in modo tale da realizzare trapianti seriali, e il loro profilo di
espressione genica è stato analizzato tramite microarray. In particolare abbiamo osservato che
leucemie fortemente adattate all’ambiente murino mostrano una cinetica di crescita più
aggressiva (Fig. 2A). Tale comportamento correla con una perturbazione dell’espressione
genica e con l’acquisizione di un profilo specifico caratterizzato dalla prevalente deregolazione
leuk
emic
cel
ls /u
L
0 25 50 75 100 125 1500
250
500
750
1000
2000
4000
6000 AML#1
0 25 50 75 100 125 1500
250
500
750
1000
2000
4000
6000 AML#8
0 25 50 75 100 125 1500
250
500
750
1000
2000
4000
6000 AML#12
0 25 50 75 100 125 1500
250
500
750
1000
2000
4000
6000 AML#16
Days after infusion
A
cell cycleRNA or DNA metabolismCell metabolismResponse to stress Immune processesCellular organizationSignaling
B
C Upregulated genes DescriptionARHGAP5 Rho GTPase Activating Protein 5AURKA Aurora kinase AAZU1 Azurocidin1C2orf44 chr2 open reading frame 44CCDC34 Coiled-Coil Domain Containing 34CDC20 Cell Division Cycle 20CENPF Centromere Protein FCTSG Cathepsin GDLGAP5 Discs Large (Drosophila) homolog-associated protein 5MPO MyeloperoxidaseNCOA5 Nuclear receptor coactivator 5PHF23 PHD Finger Protein 23PPP1R10 Protein Phosphatase 1, Regulatory Subunit 10PRTN3 Proteinase 3 +serpine 1 serpine 2PSRC1 Proline/Serine-Rich Coiled-Coil 1RNASE3 Ribonuclease, RNAse A family, 3RNASEH2C Ribonuclease H2 subunit CTMEM177 Transmembrane Protein 177TP53RK TP53 Regulating Kinase1 +p53TRIM13 Tripartite Motif Containing 13WDR4 WD repeat domainZBTB3 Zinc Finger And BTB Domain Containing 3ZNF627 Zinc Finger Protein 627ZNF788 Zinc Finger Protein 788
Downregulated genes DescriptionATF3 Activating Transcription Factor 3 CD44 CD44 moleculeCEP170 Centrosomal Protein 170kDACHMP1B Charged Multivesicular Body Protein 1BFAM65B Family With Sequence Similarity 65, Member BIDS Iduronate 2-SulfatasKLF6 Kruppel-Like Factor 6 +TP53KRT8P9 Keratin 8 Pseudogene 9LOC649431LOC653171LOC728098MYADM Myeloid-Associated Differentiation MarkerPPTC7 PTC7 protein posphatase homologSTK17B Serine/Threonine Kinase 17bTIPARP TCDD-Inducible Poly(ADP-Ribose) PolymeraseUBE2A Ubiquitin-Conjugating Enzyme E2AVIM VimentinYPEL5 Yippe-like5
Figura 2. Trapianti seriali delle leucemie attecchite nei topi NSG e identificazione dei geni associati ad una maggiore aggressività in vivo . (A) Cinetica di crescita delle cellule leucemiche misurata su sangue periferico di topi NSG in seguito a trapianti seriali. Con l’aumentare dei trapianti seriali, la stessa leucemia mostra una cinetica di crescita più aggressiva. (B) Classificazione dei processi deregolati da ciascuna leucemia in seguito all’attecchimento nel topo. La maggiore parte dei geni deregolati appartiene alla regolazione del ciclo cellulare e al metabolismo degli acidi nucleici ad esso collegato. (C) Geni deregolati stabilmente da tutte le leucemie in seguito a trapianto seriale in topi NSG. Sono elencati 24 geni upregolati (rosso) e 18 geni downregolati (blu).
1st recipients 2nd recipients 3rd recipients 4th recipients
0 25 50 75 100 125 1500
250
500
750
1000
2000
4000
6000 AML#25
0 25 50 75 100 125 1500
250
500
750
1000
2000
4000
6000 AML#20
Progetto di Ricerca Giacomo Oliveira
4
di processi coinvolti nella proliferazione cellulare (Fig. 2B). Sono stati identificati 42 geni
deregolati da tutte le leucemie in seguito ad attecchimento nel topo e correlati con un aumento
dell’aggressività delle AML nel nostro modello (Fig. 2C). Tra questi spiccano geni legati alla
mitosi cellulare (AURKA, CDC20), alla riorganizzazione della cromatina (CENPF, CEP170) e al
differenziamento mieloide (come AZU1, MPO, MYADM, CTSG), la cui deregolazione è stata
spesso associata ad una maggiore aggressività delle cellule tumorali (Zhang et al, Clin Cancer
Res 2013; Jin, Oncogene 2013 et al.; Katayama et al., Cancer Cell 2012). Il nostro modello
fornisce quindi un contesto idoneo per lo studio di tali geni nell’ambito della leucemia mieloide
acuta.
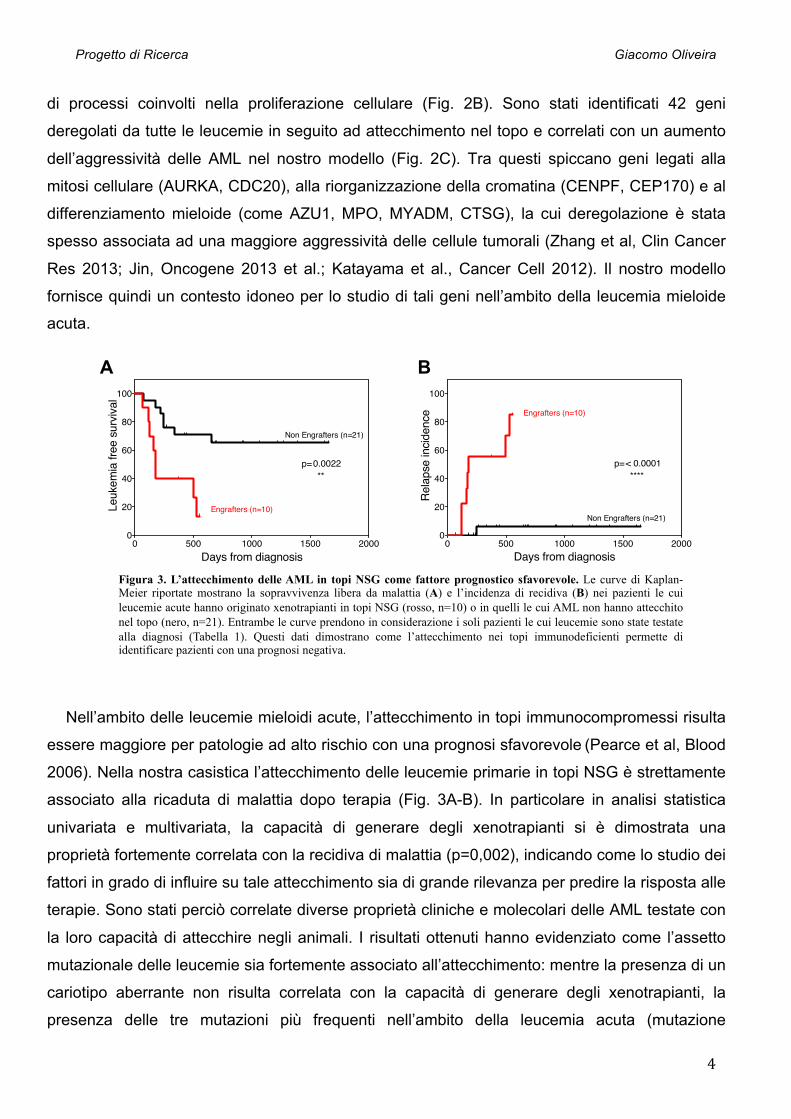
Nell’ambito delle leucemie mieloidi acute, l’attecchimento in topi immunocompromessi risulta
essere maggiore per patologie ad alto rischio con una prognosi sfavorevole (Pearce et al, Blood
2006). Nella nostra casistica l’attecchimento delle leucemie primarie in topi NSG è strettamente
associato alla ricaduta di malattia dopo terapia (Fig. 3A-B). In particolare in analisi statistica
univariata e multivariata, la capacità di generare degli xenotrapianti si è dimostrata una
proprietà fortemente correlata con la recidiva di malattia (p=0,002), indicando come lo studio dei
fattori in grado di influire su tale attecchimento sia di grande rilevanza per predire la risposta alle
terapie. Sono stati perciò correlate diverse proprietà cliniche e molecolari delle AML testate con
la loro capacità di attecchire negli animali. I risultati ottenuti hanno evidenziato come l’assetto
mutazionale delle leucemie sia fortemente associato all’attecchimento: mentre la presenza di un
cariotipo aberrante non risulta correlata con la capacità di generare degli xenotrapianti, la
presenza delle tre mutazioni più frequenti nell’ambito della leucemia acuta (mutazione
0 500 1000 1500 20000
20
40
60
80
100
Days from diagnosis
Leuk
emia
free
sur
vival
0.0022p=**
Non Engrafters (n=21)
Engrafters (n=10)
0 500 1000 1500 20000
20
40
60
80
100
Days from diagnosis
Rela
pse
incid
ence
Non Engrafters (n=21)
Engrafters (n=10)
< 0.0001p=****
A B
Figura 3. L’attecchimento delle AML in topi NSG come fattore prognostico sfavorevole. Le curve di Kaplan-Meier riportate mostrano la sopravvivenza libera da malattia (A) e l’incidenza di recidiva (B) nei pazienti le cui leucemie acute hanno originato xenotrapianti in topi NSG (rosso, n=10) o in quelli le cui AML non hanno attecchito nel topo (nero, n=21). Entrambe le curve prendono in considerazione i soli pazienti le cui leucemie sono state testate alla diagnosi (Tabella 1). Questi dati dimostrano come l’attecchimento nei topi immunodeficienti permette di identificare pazienti con una prognosi negativa.

Progetto di Ricerca Giacomo Oliveira
5
puntiforme del gene DNMT3A, mutazione puntiforme del gene NPM1 e duplicazione tandem
interna del gene FLT3) è significativamente associata all’attecchimento in topi NSG (Fig. 4A).
Inoltre la co-occorren za di queste tre mutazioni ha un notevole impatto sul tasso di
attecchimento (Fig. 4B). Questi dati dimostrano come il carico mutazionale delle leucemie (ed in
particolare le mutazioni dei geni DMNT3A, NPM1 e FLT3) influenzi la capacità delle cellule
leucemiche di generare degli xenotrapianti in topi immunodeficienti e permetta di identificare
AML ad alto rischio prognostico.
0 25 50 75 100 125 1500
25
50
75
100
Time of engraftment (days)
% o
f eng
rafte
d AM
L
p = 0.0883
p = 0.0134
p = 0.0026
3
2
1
0
# of most common mutations(FLT3ITD, DNMT3A mut, NPM1mut)
# of most common mutation
(FLT3 ITD, NPM1 mut, DNMT3A mut)
Most commonly mutated genes in AML
FLT3-ITDNPM1 mutDNMT3A mut
FLT3 wtNPM1 wtDNMT3A wt
0 25 50 75 100 125 1500
25
50
75
100
Time of engraftment (days)
% o
f eng
rafte
d AM
L
p = 0.0295
p = 0.0244
p = 0.0113
A
B
Figura 4. Associazione tra il profilo mutazionale e trascrizionale delle leucemia e l’attecchimento in topi immunodeficienti. (A-B) Curve Kaplan-Meier relative al tasso di attecchimento delle leucemie testate alla diagnosi in relazione al tipo (A) o al numero (B) delle mutazioni più ricorrenti nelle AML. I risultati dimostrano che la copresenza delle tre mutazioni si associa a un maggior tasso di attecchimento. (C) Analisi della varianza dell’espressione genica delle leucemie primarie capaci di generare xenotrapianti (arancione) o leucemie che non attecchiscono nei topi (blu). I simboli uguali identificano dei replicati tecnici di uno stesso campione, mentre simboli diversi rappresentano campioni appartenenti a pazienti diversi. Le leucemie in grado di attecchire nel topo mostrano un profilo di espressione genica più omogeneo. (D) Heatmap rappresentante il set di 136 geni differenzialmente espressi tra leucemie in grado o meno di generare xenotrapianti.
-100 -50 0 50 100-100
-50
0
50
100
PC1: 59% expl. var.
PC2:
41%
exp
l. va
r.
Engrafters Non EngraftersC
FOCA_Diagn Engrafted_2
FOCA_Diagn Engrafted_2
MABI_postCT Engraft_3
MABI_postCT Engraft_3
PLGA_Diagn Engrafted_3
PLGA_Diagn Engrafted_3
JARA_postCT Engraft_2JARA_postCT Engraft_2M
OG
E_postCT Engraft_3M
OG
E_postCT Engraft_3PIAG
_Diagn Engrafted_3PIAG
_Diagn Engrafted_3M
AGI_postCT Engraft_1
MAG
I_postCT Engraft_1FRLA_Diagn Engrafted_1FRLA_Diagn Engrafted_1M
ACE_Diagn Engrafted_1M
ACE_Diagn Engrafted_1ZIDO
_Diagn Engrafted_2ZIDO
_Diagn Engrafted_2BADI_Diagn Non Engraft_2BADI_Diagn Non Engraft_2BAPI_Diagn Non Engraft_3BAPI_Diagn Non Engraft_3CAEL_Diagn Non Engraft_1CAEL_Diagn Non Engraft_1M
AEL_Diagn Non Engraft_2M
AEL_Diagn Non Engraft_2ANAN_Diagn Non Engraft_1ANAN_Diagn Non Engraft_1CALU_postCT Engraft_2CALU_postCT Engraft_2O
ZAR_postCT Engraft_1O
ZAR_postCT Engraft_1TATI_Diagn Non Engraft_3TATI_Diagn Non Engraft_3
SERPINB8AK3L1HS.10862CHD7HOMER3ST3GAL6ITSN1ACSL1SAP30COL5A1SOX18LOC283340RNF144BVNN1CLIP4IGF2BP3VWA5ALDLRAD2BCAS4SULT1A1LOC728047SLC29A2AFAP1L2CYP2C8GPR27EFHC2ITGA2BZNF154ACY3INPP4BGCH1FRMD4BTMEM144DDEF2KLHL3PNMA3LSP1C4ORF32LOC641825ACP6PPM1HLOC391475CARD11ST6GALNAC4IGJPRDM16GLI2LOC647336DOCK1C9ORF58LOC730051JAM3LCTANGPT2PLS1GPR12SLC24A3RPS6KA2TDRD9TRAT1HALMFAP4CYP2E1LOC643911FLJ22536MC1REPHB4LOC401237ALDH1A1LRRN2GPRC5CMPZL2C6ORF192ASAP2GSTO2GATA1ZNF135C20ORF54ZNF418TNFRSF25ABCG1RGS12HOPXSMARCA1CMTM2TSPAN13ARHGEF5LOR2A9PSLC22A5SLC24A4ATP8A1KDELC1MOSC2ACPL2PGAP1TGFAVASNACVR1BCDKN2AABLIM1MMRN1C1ORF150HS.25318H2AFY2AMICA1SORT1NYNRINCD7KLRG1ALDOCLOC286367SPINK2GPR114C11ORF21SPNS3GPR56FAM30AKIAA0125AIF1LSPINT2PADI4CPNE3ZNF185FCER1ASEPT5SH3PXD2AHCP5LTBERMP1RTN4GUCY1A3S100ZS100A10PLIN2FLNBSFRS2B
6
8
10
12
14
FOCA_Diagn Engrafted_2
FOCA_Diagn Engrafted_2
MABI_postCT Engraft_3
MABI_postCT Engraft_3
PLGA_Diagn Engrafted_3
PLGA_Diagn Engrafted_3
JARA_postCT Engraft_2JARA_postCT Engraft_2M
OG
E_postCT Engraft_3M
OG
E_postCT Engraft_3PIAG
_Diagn Engrafted_3PIAG
_Diagn Engrafted_3M
AGI_postCT Engraft_1
MAG
I_postCT Engraft_1FRLA_Diagn Engrafted_1FRLA_Diagn Engrafted_1M
ACE_Diagn Engrafted_1M
ACE_Diagn Engrafted_1ZIDO
_Diagn Engrafted_2ZIDO
_Diagn Engrafted_2BADI_Diagn Non Engraft_2BADI_Diagn Non Engraft_2BAPI_Diagn Non Engraft_3BAPI_Diagn Non Engraft_3CAEL_Diagn Non Engraft_1CAEL_Diagn Non Engraft_1M
AEL_Diagn Non Engraft_2M
AEL_Diagn Non Engraft_2ANAN_Diagn Non Engraft_1ANAN_Diagn Non Engraft_1CALU_postCT Engraft_2CALU_postCT Engraft_2O
ZAR_postCT Engraft_1O
ZAR_postCT Engraft_1TATI_Diagn Non Engraft_3TATI_Diagn Non Engraft_3
SERPINB8AK3L1HS.10862CHD7HOMER3ST3GAL6ITSN1ACSL1SAP30COL5A1SOX18LOC283340RNF144BVNN1CLIP4IGF2BP3VWA5ALDLRAD2BCAS4SULT1A1LOC728047SLC29A2AFAP1L2CYP2C8GPR27EFHC2ITGA2BZNF154ACY3INPP4BGCH1FRMD4BTMEM144DDEF2KLHL3PNMA3LSP1C4ORF32LOC641825ACP6PPM1HLOC391475CARD11ST6GALNAC4IGJPRDM16GLI2LOC647336DOCK1C9ORF58LOC730051JAM3LCTANGPT2PLS1GPR12SLC24A3RPS6KA2TDRD9TRAT1HALMFAP4CYP2E1LOC643911FLJ22536MC1REPHB4LOC401237ALDH1A1LRRN2GPRC5CMPZL2C6ORF192ASAP2GSTO2GATA1ZNF135C20ORF54ZNF418TNFRSF25ABCG1RGS12HOPXSMARCA1CMTM2TSPAN13ARHGEF5LOR2A9PSLC22A5SLC24A4ATP8A1KDELC1MOSC2ACPL2PGAP1TGFAVASNACVR1BCDKN2AABLIM1MMRN1C1ORF150HS.25318H2AFY2AMICA1SORT1NYNRINCD7KLRG1ALDOCLOC286367SPINK2GPR114C11ORF21SPNS3GPR56FAM30AKIAA0125AIF1LSPINT2PADI4CPNE3ZNF185FCER1ASEPT5SH3PXD2AHCP5LTBERMP1RTN4GUCY1A3S100ZS100A10PLIN2FLNBSFRS2B
6
8
10
12
14
FOCA_Diagn Engrafted_2
FOCA_Diagn Engrafted_2
MABI_postCT Engraft_3
MABI_postCT Engraft_3
PLGA_Diagn Engrafted_3
PLGA_Diagn Engrafted_3
JARA_postCT Engraft_2JARA_postCT Engraft_2M
OG
E_postCT Engraft_3M
OG
E_postCT Engraft_3PIAG
_Diagn Engrafted_3PIAG
_Diagn Engrafted_3M
AGI_postCT Engraft_1
MAG
I_postCT Engraft_1FRLA_Diagn Engrafted_1FRLA_Diagn Engrafted_1M
ACE_Diagn Engrafted_1M
ACE_Diagn Engrafted_1ZIDO
_Diagn Engrafted_2ZIDO
_Diagn Engrafted_2BADI_Diagn Non Engraft_2BADI_Diagn Non Engraft_2BAPI_Diagn Non Engraft_3BAPI_Diagn Non Engraft_3CAEL_Diagn Non Engraft_1CAEL_Diagn Non Engraft_1M
AEL_Diagn Non Engraft_2M
AEL_Diagn Non Engraft_2ANAN_Diagn Non Engraft_1ANAN_Diagn Non Engraft_1CALU_postCT Engraft_2CALU_postCT Engraft_2O
ZAR_postCT Engraft_1O
ZAR_postCT Engraft_1TATI_Diagn Non Engraft_3TATI_Diagn Non Engraft_3
SERPINB8AK3L1HS.10862CHD7HOMER3ST3GAL6ITSN1ACSL1SAP30COL5A1SOX18LOC283340RNF144BVNN1CLIP4IGF2BP3VWA5ALDLRAD2BCAS4SULT1A1LOC728047SLC29A2AFAP1L2CYP2C8GPR27EFHC2ITGA2BZNF154ACY3INPP4BGCH1FRMD4BTMEM144DDEF2KLHL3PNMA3LSP1C4ORF32LOC641825ACP6PPM1HLOC391475CARD11ST6GALNAC4IGJPRDM16GLI2LOC647336DOCK1C9ORF58LOC730051JAM3LCTANGPT2PLS1GPR12SLC24A3RPS6KA2TDRD9TRAT1HALMFAP4CYP2E1LOC643911FLJ22536MC1REPHB4LOC401237ALDH1A1LRRN2GPRC5CMPZL2C6ORF192ASAP2GSTO2GATA1ZNF135C20ORF54ZNF418TNFRSF25ABCG1RGS12HOPXSMARCA1CMTM2TSPAN13ARHGEF5LOR2A9PSLC22A5SLC24A4ATP8A1KDELC1MOSC2ACPL2PGAP1TGFAVASNACVR1BCDKN2AABLIM1MMRN1C1ORF150HS.25318H2AFY2AMICA1SORT1NYNRINCD7KLRG1ALDOCLOC286367SPINK2GPR114C11ORF21SPNS3GPR56FAM30AKIAA0125AIF1LSPINT2PADI4CPNE3ZNF185FCER1ASEPT5SH3PXD2AHCP5LTBERMP1RTN4GUCY1A3S100ZS100A10PLIN2FLNBSFRS2B
6
8
10
12
14
FOCA_Diagn Engrafted_2
FOCA_Diagn Engrafted_2
MABI_postCT Engraft_3
MABI_postCT Engraft_3
PLGA_Diagn Engrafted_3
PLGA_Diagn Engrafted_3
JARA_postCT Engraft_2JARA_postCT Engraft_2M
OG
E_postCT Engraft_3M
OG
E_postCT Engraft_3PIAG
_Diagn Engrafted_3PIAG
_Diagn Engrafted_3M
AGI_postCT Engraft_1
MAG
I_postCT Engraft_1FRLA_Diagn Engrafted_1FRLA_Diagn Engrafted_1M
ACE_Diagn Engrafted_1M
ACE_Diagn Engrafted_1ZIDO
_Diagn Engrafted_2ZIDO
_Diagn Engrafted_2BADI_Diagn Non Engraft_2BADI_Diagn Non Engraft_2BAPI_Diagn Non Engraft_3BAPI_Diagn Non Engraft_3CAEL_Diagn Non Engraft_1CAEL_Diagn Non Engraft_1M
AEL_Diagn Non Engraft_2M
AEL_Diagn Non Engraft_2ANAN_Diagn Non Engraft_1ANAN_Diagn Non Engraft_1CALU_postCT Engraft_2CALU_postCT Engraft_2O
ZAR_postCT Engraft_1O
ZAR_postCT Engraft_1TATI_Diagn Non Engraft_3TATI_Diagn Non Engraft_3
SERPINB8AK3L1HS.10862CHD7HOMER3ST3GAL6ITSN1ACSL1SAP30COL5A1SOX18LOC283340RNF144BVNN1CLIP4IGF2BP3VWA5ALDLRAD2BCAS4SULT1A1LOC728047SLC29A2AFAP1L2CYP2C8GPR27EFHC2ITGA2BZNF154ACY3INPP4BGCH1FRMD4BTMEM144DDEF2KLHL3PNMA3LSP1C4ORF32LOC641825ACP6PPM1HLOC391475CARD11ST6GALNAC4IGJPRDM16GLI2LOC647336DOCK1C9ORF58LOC730051JAM3LCTANGPT2PLS1GPR12SLC24A3RPS6KA2TDRD9TRAT1HALMFAP4CYP2E1LOC643911FLJ22536MC1REPHB4LOC401237ALDH1A1LRRN2GPRC5CMPZL2C6ORF192ASAP2GSTO2GATA1ZNF135C20ORF54ZNF418TNFRSF25ABCG1RGS12HOPXSMARCA1CMTM2TSPAN13ARHGEF5LOR2A9PSLC22A5SLC24A4ATP8A1KDELC1MOSC2ACPL2PGAP1TGFAVASNACVR1BCDKN2AABLIM1MMRN1C1ORF150HS.25318H2AFY2AMICA1SORT1NYNRINCD7KLRG1ALDOCLOC286367SPINK2GPR114C11ORF21SPNS3GPR56FAM30AKIAA0125AIF1LSPINT2PADI4CPNE3ZNF185FCER1ASEPT5SH3PXD2AHCP5LTBERMP1RTN4GUCY1A3S100ZS100A10PLIN2FLNBSFRS2B
6
8
10
12
14
Non Engrafters
Engrafters
FOCA_Diagn Engrafted_2
FOCA_Diagn Engrafted_2
MABI_postCT Engraft_3
MABI_postCT Engraft_3
PLGA_Diagn Engrafted_3
PLGA_Diagn Engrafted_3
JARA_postCT Engraft_2JARA_postCT Engraft_2M
OG
E_postCT Engraft_3M
OG
E_postCT Engraft_3PIAG
_Diagn Engrafted_3PIAG
_Diagn Engrafted_3M
AGI_postCT Engraft_1
MAG
I_postCT Engraft_1FRLA_Diagn Engrafted_1FRLA_Diagn Engrafted_1M
ACE_Diagn Engrafted_1M
ACE_Diagn Engrafted_1ZIDO
_Diagn Engrafted_2ZIDO
_Diagn Engrafted_2BADI_Diagn Non Engraft_2BADI_Diagn Non Engraft_2BAPI_Diagn Non Engraft_3BAPI_Diagn Non Engraft_3CAEL_Diagn Non Engraft_1CAEL_Diagn Non Engraft_1M
AEL_Diagn Non Engraft_2M
AEL_Diagn Non Engraft_2ANAN_Diagn Non Engraft_1ANAN_Diagn Non Engraft_1CALU_postCT Engraft_2CALU_postCT Engraft_2O
ZAR_postCT Engraft_1O
ZAR_postCT Engraft_1TATI_Diagn Non Engraft_3TATI_Diagn Non Engraft_3
SERPINB8AK3L1HS.10862CHD7HOMER3ST3GAL6ITSN1ACSL1SAP30COL5A1SOX18LOC283340RNF144BVNN1CLIP4IGF2BP3VWA5ALDLRAD2BCAS4SULT1A1LOC728047SLC29A2AFAP1L2CYP2C8GPR27EFHC2ITGA2BZNF154ACY3INPP4BGCH1FRMD4BTMEM144DDEF2KLHL3PNMA3LSP1C4ORF32LOC641825ACP6PPM1HLOC391475CARD11ST6GALNAC4IGJPRDM16GLI2LOC647336DOCK1C9ORF58LOC730051JAM3LCTANGPT2PLS1GPR12SLC24A3RPS6KA2TDRD9TRAT1HALMFAP4CYP2E1LOC643911FLJ22536MC1REPHB4LOC401237ALDH1A1LRRN2GPRC5CMPZL2C6ORF192ASAP2GSTO2GATA1ZNF135C20ORF54ZNF418TNFRSF25ABCG1RGS12HOPXSMARCA1CMTM2TSPAN13ARHGEF5LOR2A9PSLC22A5SLC24A4ATP8A1KDELC1MOSC2ACPL2PGAP1TGFAVASNACVR1BCDKN2AABLIM1MMRN1C1ORF150HS.25318H2AFY2AMICA1SORT1NYNRINCD7KLRG1ALDOCLOC286367SPINK2GPR114C11ORF21SPNS3GPR56FAM30AKIAA0125AIF1LSPINT2PADI4CPNE3ZNF185FCER1ASEPT5SH3PXD2AHCP5LTBERMP1RTN4GUCY1A3S100ZS100A10PLIN2FLNBSFRS2B
6
8
10
12
14
D

Progetto di Ricerca Giacomo Oliveira
6
Infine, con lo scopo di identificare le peculiarità del trascrittoma delle leucemie più aggressive
ed in grado di attecchire nei topi, il profilo di espressione genica delle AML primarie in grado di
generare degli xenotrapianti è stato confrontato con quello di leucemie che sono prive di tale
proprietà. L’analisi della varianza dell’espressione genica ha permesso di visualizzare che,
sebbene le leucemie provenienti da pazienti diversi siano molto eterogenee, le AML in grado di
attecchire nei topi risultano più simili tra loro di quanto siano le altre leucemie (Fig. 4C). Dal
confronto dei due gruppi sperimentali è stato possibile identificare un set di 136 geni espressi in
maniera differenziale dalle leucemie in grado di generare xenotrapianti e che permetta quindi di
identificare le leucemie con una prognosi sfavorevole (Fig. 4D).
Messa a punto di un modello di pressione immunologica in vivo
La disponibilità di leucemie umane in grado di attecchire in topo immunodeficientici ha
permesso di studiare direttamente in vivo l’efficacia della pressione immunologica nel controllo
delle cellule tumorali e i cambiamenti messi in atto dalle leucemie mieloidi acute nell’interazione
col sistema immunitario umano. A tal scopo abbiamo messo a punto un modello che ricapitoli
una pressione immune nei confronti delle cellule leucemiche: una volta attecchite in topi NSG,
le cellule leucemiche sono state sottoposte in vivo ad una pressione immunologica costituita da
infusioni seriali di linfociti T umani. In particolare, per mimare una diversa intensità della
pressione immune, sono stati utilizzati linfociti T provenienti da diversi donatori e dotati di un
differente grado di compatibilità nei confronti delle leucemie (linfociti T autologhi, HLA-identici,
HLA compatibili da donatore da registro, aploidentici o completamente scorrelati), in modo tale
da ottenere un diverso grado di allo-reattività. Per ottenere numeri sufficienti per il trattamento di
diversi topi, le cellule T dei differenti donatori sono state attivate con biglie antiCD3/CD28 ed
espanse in presenza di IL-7 e IL-15, in modo tale da preservarne la policlonalità e le potenzialità
effettrici (Kaneko, Blood 2009). Un gruppo di controllo ha ricevuto la sola leucemia, in assenza
di trattamento linfocitario. I risultati ottenuti su 4 leucemie differenti mostrano che in nessuno dei
casi analizzati le cellule autologhe sono state in grado di controllare la crescita delle cellule
leucemiche (Fig. 5, linee verdi). A seconda della leucemia testata, l’utilizzo di linfociti T HLA-
identiche ha consentito di ottenere un controllo della malattia parziale o duraturo (Fig. 5, linee
blu). Al contrario un completo arresto della crescita delle cellule leucemiche è stato raggiunto
con l’infusione di linfociti T con una singola incompatibilità dell’HLA di classe seconda (Fig. 5,
line viola). Infine l’utilizzo di linfociti T completamente o parzialmente scorrelati (Fig. 5, linee
rosa e rosse) ha comportato la completa scomparsa delle cellule leucemiche circolanti nel

Progetto di Ricerca Giacomo Oliveira
7
sangue periferico degli animali trattati. Tale modello dimostra come il riconoscimento alloreattivo
mediato dai linfociti T rappresenta una delle maggiori forze che determinano un effetto
antileucemico delle terapie cellulari che utilizzano linfociti da donatore.
Nonostante l’efficacia dei linfociti T allogenici nel controllare la crescita della malattia, le
cellule leucemiche sono in grado di eludere dal controllo del sistema immunitario, come
dimostrato clinicamente dalla recidiva di malattia in seguito a trapianto di cellule staminali da
No T cells
Autologous T cells
Class II mismatched T cells
Haploidentical T cells
Fully mismatched T cells
HLA-Identical T cells
Treated with:
20 40 60 800
50
100
1000
2000
3000AML#26
20 40 60 800
50
100
500100015002000
AML#16
20 40 60 800
50
100
500
1000AML#8
20 40 60 80 1000
200
400
500
750
1000AML#1
Days after infusion of leukemic cells
leuk
emic
cel
ls / μL
No T cells
Autologous T cells
Class II mismatched T cells
Haploidentical T cells
Fully mismatched T cells
HLA-Identical T cells
Treated with:
No T cells
Autologous T cells
Class II mismatched T cells
Haploidentical T cells
Fully mismatched T cells
HLA-Identical T cells
Treated with:
Figura 5. Pressione immunologica mediata in vivo dai linfociti T sulle cellule leucemiche attecchite nei topi NSG. I grafici mostrano le conte delle cellule leucemiche misurate sul sangue periferico di topi immunodeficienti in cui è stata fatta attecchire una specifica AML. Una volta stabilita la presenza della leucemia in vivo, i topi hanno ricevuto una o più infusione di 5x106 linfociti T (freccia nera) provenienti da differenti donatori con un diverso grado di disparità genetica nei confronti della malattia. In particolare i differenti gruppi sperimentali (n>4) hanno ricevuto una pressione immunologica costituita da: linfociti T autologhi (verde), linfociti T HLA-identici (blu), linfociti T da donatore da registro con una disparità in uno degli HLA di classe II (viola), linfociti aploidentici (rosa) e linfociti completamente scorrelati (rosso). Per ciascuna delle 4 leucemie testate, un gruppo ha ricevuto la sola leucemia, senza trattamento linfocitario (giallo). I dati mostrano l’attività antileucemica della pressione immune somministrata nei confronti della leucemia: mentre i linfociti T autologhi risultano inefficaci e i linfociti T HLA-identici lo sono solo parzialmente, una singola disparità dei geni HLA è sufficiente a determinare l’eradicazione delle cellule leucemiche dal sangue dei topi, dimostrando come in tale modello l’alloreattività sia la sorgente principale dell’azione antileucemica.
Progetto di Ricerca Giacomo Oliveira
8
donatore allogenico. Nel nostro modello abbiamo rilevato due casi sperimentali che indicano
come le cellule leucemiche siano in grado di persistere anche in presenza della pressione
immune mediata dai linfociti T. In un primo caso, il trattamento con linfociti T HLA-identici si è
0 50 100 150 200 250 300 350 400 450 5000
100
200
300
400
10000200003000040000
Treated withHaploidentical T cells
1st recipient NSG 2nd recipient NSG 3rd recipient NSG 4th recipient NSGDays
cells
/ µL
0 50 100 150 200 250 300 350 400 450 5000
100
200
300
400
10000200003000040000
Treated withHLA-mismatched T cells
1st recipient NSG 2nd recipient NSG 3rd recipient NSG 4th recipient NSGDays
cells
/ µL
T cells Leukemic cells DLILegend: CD3 depletion Transfer of leukemic cells in recipient NSG mice
A
C
B
T cells Leukemic cells DLILegend:
0 50 100 1500
100
200
300
400
10000200003000040000
Days ce
lls / µL
0 50 100 1500
100
200
300
400
10000200003000040000
Days
cells
/ µL
UntreatedTreated with
HLA-Identical T cells
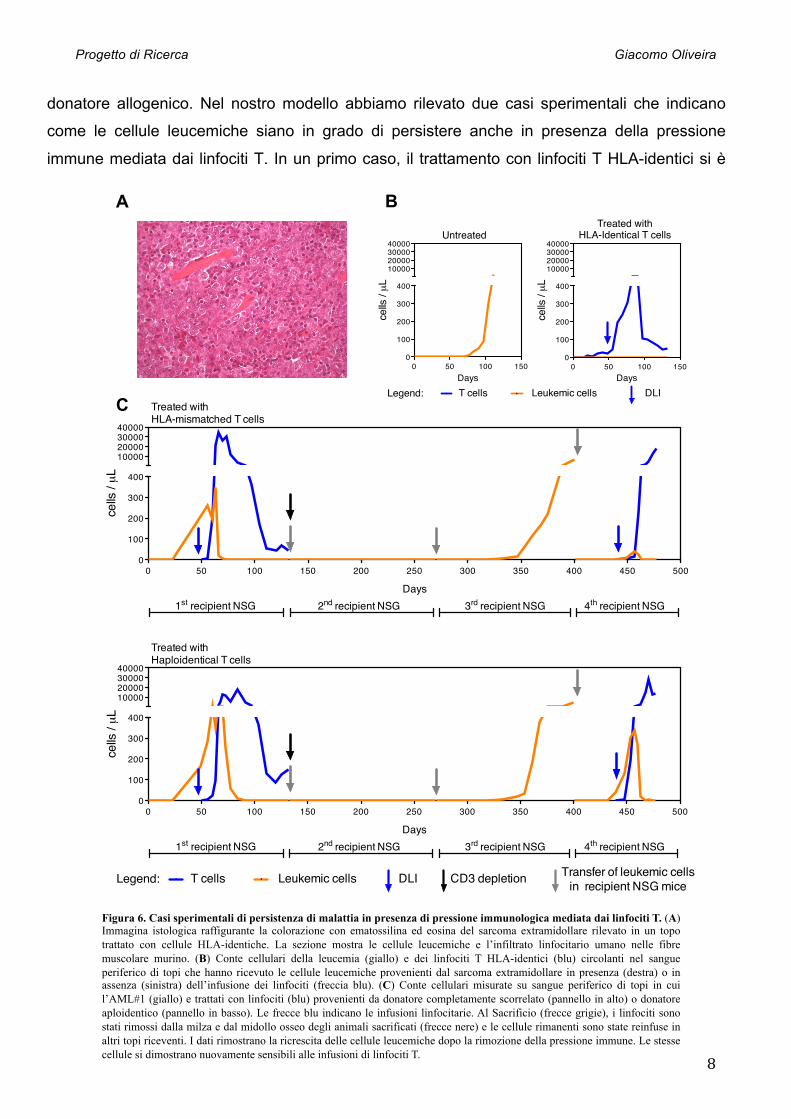
Figura 6. Casi sperimentali di persistenza di malattia in presenza di pressione immunologica mediata dai linfociti T. (A) Immagina istologica raffigurante la colorazione con ematossilina ed eosina del sarcoma extramidollare rilevato in un topo trattato con cellule HLA-identiche. La sezione mostra le cellule leucemiche e l’infiltrato linfocitario umano nelle fibre muscolare murino. (B) Conte cellulari della leucemia (giallo) e dei linfociti T HLA-identici (blu) circolanti nel sangue periferico di topi che hanno ricevuto le cellule leucemiche provenienti dal sarcoma extramidollare in presenza (destra) o in assenza (sinistra) dell’infusione dei linfociti (freccia blu). (C) Conte cellulari misurate su sangue periferico di topi in cui l’AML#1 (giallo) e trattati con linfociti (blu) provenienti da donatore completamente scorrelato (pannello in alto) o donatore aploidentico (pannello in basso). Le frecce blu indicano le infusioni linfocitarie. Al Sacrificio (frecce grigie), i linfociti sono stati rimossi dalla milza e dal midollo osseo degli animali sacrificati (frecce nere) e le cellule rimanenti sono state reinfuse in altri topi riceventi. I dati rimostrano la ricrescita delle cellule leucemiche dopo la rimozione della pressione immune. Le stesse cellule si dimostrano nuovamente sensibili alle infusioni di linfociti T.

Progetto di Ricerca Giacomo Oliveira
9
dimostrato efficace nel controllare la crescita delle cellule leucemiche; in uno degli animali
trattati, tale efficacia è stata però accompagnata dallo sviluppo di un sarcoma extramidollare
costituito da cellule leucemiche umane e da un infiltrato linfocitario umano all’intermo nel
tessuto muscolare murino (Fig. 6A). Per verificare se ciò fosse il risultato di un evento di
immunoevasione della AML#16 nei confronti della pressione immune, le cellule leucemiche del
sarcoma sono state purificate e infuse in topi riceventi secondari (Fig. 6B). Tali cellule sono
state in grado di ricrescere in assenza di trattamento, ma si sono dimostrate nuovamente
sensibili all’infusione degli stessi linfociti T HLA-identici. In un secondo caso abbiamo dimostrato
la persistenza di malattia anche in presenza di una pressione immunologica altamente allo-
reattiva: in topi con AML#1 e trattati con linfociti T parzialmente o completamente scorrelati, la
presenza di cellule leucemiche residue è stata vagliata rimuovendo i linfociti T dal midollo e
dalla milza degli animali sacrificati e trapiantando in modo seriale le cellule rimanenti. In modo
del tutto inaspettato, abbiamo osservato che alcune rare cellule leucemiche sono sopravvissute
al trattamento, sono state trasferite nei topi secondari e sono ricresciute in seguito a trapianti
seriali (Fig. 6C). Una volta riapplicata la pressione immune, tali cellule si sono nuovamente
dimostrate sensibili al trattamento linfocitario e sono state eradicate dal sangue periferico. Tali
osservazioni suggeriscono che alcune cellule leucemiche possono persistere nel nostro modello
sotto il controllo dei linfociti T, e sono in grado di riemergere in seguito alla rimozione della
pressione immune.
Infine, per determinare come la pressione immunologica mediata dai linfociti T influenzi il
profilo di espressione delle cellule leucemiche, abbiamo effettuato un’analisi del trascrittoma
delle cellule leucemiche persistenti dopo trattamento. Per questo motivo, le cellule dell’AML#1
sono state purificate dalla milza e dal midollo di topi trattati con linfociti T autologhi o HLA-
identici e sono state confrontate tramite microarray con cellule leucemiche isolate da topi non
trattati. Data la completa eradicazione, tale studio non è stata effettuata per i topi trattati con
linfociti T scorrelati. L’analisi della varianza dell’espressione genica ha dimostrato che il
trattamento con linfociti T determina una perturbazione del profilo trascrizionale della AML presa
in considerazione (Fig. 7A). Sono stati identificati 2383 geni comunemente deregolati in seguito
a trattamento con linfociti T autologhi e HLA-identici. Interrogando le liste annotate di Gene
Ontology (GO) e Gene Set Enrichment Analysis (GSEA) sono stati identificati i processi
maggiormente influenzati dalla pressione immune. La maggior parte di questi è costituita da
processi immuni (Fig. 7B): i geni maggiormente deregolati risultano essere correlati alla risposta
agli interferoni, come molte subunità del proteasoma e dell’immunoproteasoma (PSMD1,
PSMB8, PSME2, IP11s regulator, 26S proteasome, 20S core, HSP90), o annoverano molecole
Progetto di Ricerca Giacomo Oliveira
10
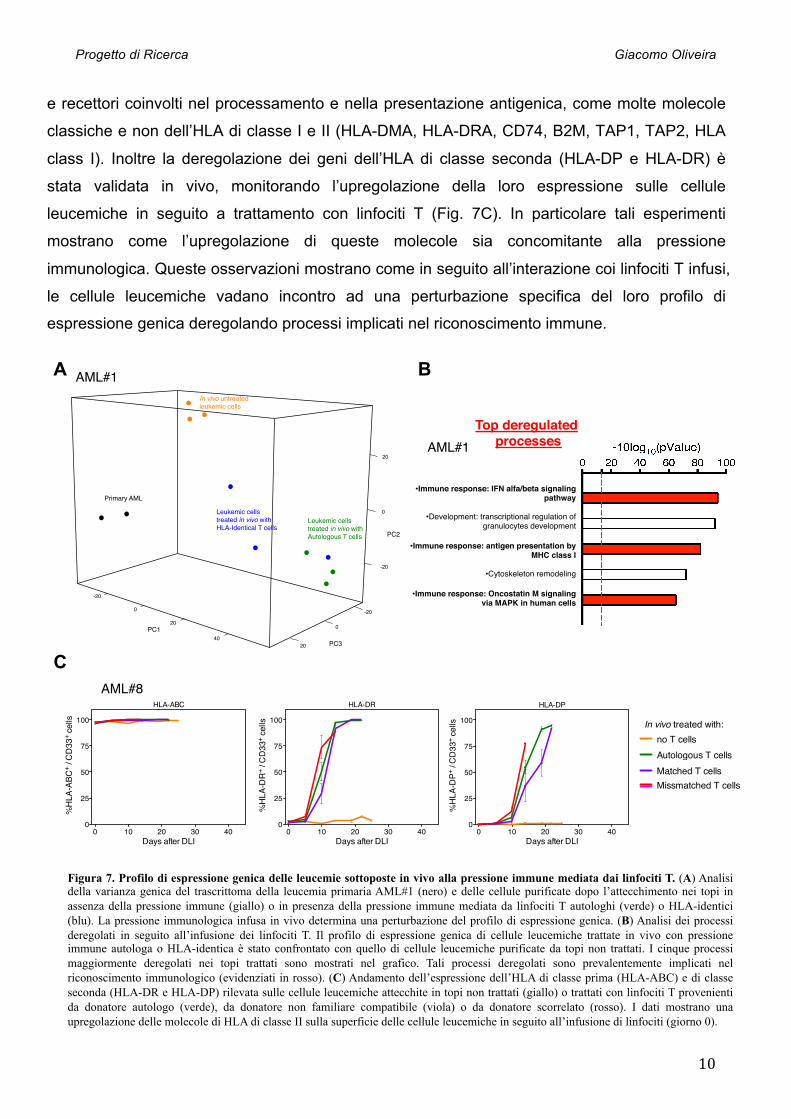
e recettori coinvolti nel processamento e nella presentazione antigenica, come molte molecole
classiche e non dell’HLA di classe I e II (HLA-DMA, HLA-DRA, CD74, B2M, TAP1, TAP2, HLA
class I). Inoltre la deregolazione dei geni dell’HLA di classe seconda (HLA-DP e HLA-DR) è
stata validata in vivo, monitorando l’upregolazione della loro espressione sulle cellule
leucemiche in seguito a trattamento con linfociti T (Fig. 7C). In particolare tali esperimenti
mostrano come l’upregolazione di queste molecole sia concomitante alla pressione
immunologica. Queste osservazioni mostrano come in seguito all’interazione coi linfociti T infusi,
le cellule leucemiche vadano incontro ad una perturbazione specifica del loro profilo di
espressione genica deregolando processi implicati nel riconoscimento immune.
20
0
-20
-20
0
20
-20
0
20
40PC1
PC3
PC2
Leukemic cells treated in vivo with Autologous T cells
Leukemic cells treated in vivo with HLA-Identical T cells
In vivo untreated leukemic cells
Primary AML• Immune response: IFN alfa/beta signaling
pathway
• Development: transcriptional regulation of granulocytes development
• Immune response: antigen presentation by MHC class I
• Cytoskeleton remodeling
• Immune response: Oncostatin M signaling via MAPK in human cells
Top deregulated processes
A B
C
no T cellsAutologous T cellsMatched T cellsHaploidentical T cellsMissmatched T cells
In vivo treated with:
AML#8
0 10 20 30 400
25
50
75
100
HLA-ABC
Days after DLI
%H
LA-A
BC+ /
CD
33+ c
ells
0 10 20 30 400
25
50
75
100
HLA-DR
Days after DLI
%H
LA-D
R+ /
CD
33+ c
ells
AML#1
AML#1
0 10 20 30 400
25
50
75
100
HLA-DP
Days after DLI
%H
LA-D
P+ / C
D33
+ cel
ls no T cellsAutologous T cellsMatched T cellsHaploidentical T cellsMissmatched T cells
In vivo treated with:
Figura 7. Profilo di espressione genica delle leucemie sottoposte in vivo alla pressione immune mediata dai linfociti T. (A) Analisi della varianza genica del trascrittoma della leucemia primaria AML#1 (nero) e delle cellule purificate dopo l’attecchimento nei topi in assenza della pressione immune (giallo) o in presenza della pressione immune mediata da linfociti T autologhi (verde) o HLA-identici (blu). La pressione immunologica infusa in vivo determina una perturbazione del profilo di espressione genica. (B) Analisi dei processi deregolati in seguito all’infusione dei linfociti T. Il profilo di espressione genica di cellule leucemiche trattate in vivo con pressione immune autologa o HLA-identica è stato confrontato con quello di cellule leucemiche purificate da topi non trattati. I cinque processi maggiormente deregolati nei topi trattati sono mostrati nel grafico. Tali processi deregolati sono prevalentemente implicati nel riconoscimento immunologico (evidenziati in rosso). (C) Andamento dell’espressione dell’HLA di classe prima (HLA-ABC) e di classe seconda (HLA-DR e HLA-DP) rilevata sulle cellule leucemiche attecchite in topi non trattati (giallo) o trattati con linfociti T provenienti da donatore autologo (verde), da donatore non familiare compatibile (viola) o da donatore scorrelato (rosso). I dati mostrano una upregolazione delle molecole di HLA di classe II sulla superficie delle cellule leucemiche in seguito all’infusione di linfociti (giorno 0).

Progetto di Ricerca Giacomo Oliveira
11
Conclusioni e prospettive future
Nel corso del progetto abbiamo sviluppato e messo a punto un modello murino umanizzato
con lo scopo di caratterizzare i fattori prognostici molecolari nell’ambito della leucemia mieloide
acuta. I risultati ottenuti dimostrano che l’attecchimento delle AML in topi immunodeficienti
costituisce una proprietà che correla con una prognosi infausta: lo studio delle variabili che
influenzano tale proprietà ha così permesso di definire come la co-presenza di mutazioni nei
geni più frequentemente colpiti nell’ambito delle leucemie mieloidi acute (DNMT3A, NMP1 e
FLT3) si associ ad un elevata capacità di attecchire negli animali testati. In linea con queste
osservazioni, altri lavori hanno riportato un effetto sfavorevole della co-presenza di tali
mutazioni sulla prognosi dei pazienti (Dohner et al, ASH 2015). Confrontando il profilo
trascrizionale di AML in grado o meno di generare xenotrapianti abbiamo individuare un set di
geni deregolato nelle cellule leucemiche in grado di attecchire nei topi. Tale gruppo di geni
potrebbe permettere di stratificare i pazienti in gruppi con differente prognosi: questa ipotesi
verrà valutata successivamente prendendo in considerazione database pubblici e analizzando
come la deregolazioni dei geni da noi identificati sia un grado di stratificare l’outcome dei
pazienti. Inoltre verranno validati e studiati in dettaglio i geni che maggiormente definiscono
questa “signature” specifica: tale studio potrebbe costituire la base per lo sviluppo di nuove
strategie terapeutiche volte a colpire i geni deregolati nelle leucemie associati ad un fenotipo più
aggressivo.
La studio delle leucemie attecchite nei topi ha permesso di dimostrare come le caratteristiche
principali delle cellule leucemiche umane rimangano inalterate nei topi immunodeficienti e come
tutti i parametri osservati (crescita delle cellule e profilo trascrizionale) siano riproducibili: ciò ci
ha permesso di sfruttare questo modello per determinare il potenziale antileucemico dei linfociti
T e per studiare come la pressione immunologica da essi esercitata vada a scolpire il profilo
delle cellule leucemiche. Il modello umanizzato messo a punto in un contesto privo di fattori
confondenti (quali l’immunosoppressione e le reazioni avverse di GvHD), ha permesso di
evidenziare come il potenziale leucemico dei linfociti T sia proporzionale alla disparità HLA tra
donatore e leucemia. La pressione immune, inoltre, determina una perturbazione specifica del
profilo trascrizionale delle cellule leucemiche che è basato sulla deregolazione di processi
immuni fondamentali quali il processamento e la presentazione antigenica. Inoltre in diversi casi
sperimentali abbiamo documentato la capacità di alcune rare cellule leucemiche di persistere in
presenza della pressione immune. Il nostro modello permette di studiare a fondo come la
pressione immunologica allogenica sia in grado di eliminare la malattia e come alcune cellule

Progetto di Ricerca Giacomo Oliveira
12
leucemiche siano in grado di persistere in sua presenza. Nel corso degli esperimenti non
abbiamo però identificato delle varianti leucemiche in grado di acquisire una resistenza
permanente e capaci di evadere il controllo del sistema immune. Ciò potrebbe essere dato dalla
scarsa eterogeneità clonale delle leucemie attecchite nei topi e da una pressione immunologica
troppo forte mediata dai linfociti T infusi. A tal proposito sono già state identificate delle modalità
sia per indurre instabilità genetica nelle cellule leucemiche, favorendo così l’insorgere di cloni
immunoresistenti, sia per controllare la reattività della pressione immune somministrata. In
particolare le cellule leucemiche possono essere modificate con vettori lentivirali genotossici in
grado di indurre instabilità genetica ed espressione aberrante dei geni colpiti (Cesana et al, Mol
Ther 2014). Abbiamo verificato che vettori lentivirali contenenti un gene reporter (GFP) siano in
grado si essere trasdurre efficientemente le leucemie testate e che tale processo non influenzi
la capacità di attecchimento e di crescita delle cellule in vivo (Fig. 8). Applicando una pressione
immunologica su tali cellule leucemiche si potrebbero identificare delle varianti leucemiche
capaci di eludere la risposta immune; inoltre, analizzando dove il vettore virale genotossico si è
inserito nel DNA delle cellule leucemiche, è possibile identificare i geni deregolati, e quindi
coinvolti nell’evasione dalla pressione mediata dai linfociti T. Al tempo stesso verranno valutate
delle modalità con cui controllare la forza della reazione linfocitaria: in tal caso la
somministrazione di agenti immunosoppressivi selettivi per i linfociti T potrebbe costituire una
valida opzione per attenuare la crescita delle cellule antileucemiche. Inoltre i linfociti T possono
essere ingegnerizzati con geni suicidi, la cui attivazione permette di eliminare in maniera
selettiva le cellule, controllando così la loro reattività. Questi approcci verranno valutati e messi
a punti nel corso del progetto, con l’obiettivo di identificare nel nostro modello umanizzato eventi
di immunoevasione. La caratterizzazione molecolare e biologica di tali fenomeni potrà
consentire lo studio, in un sistema controllato, dei processi indispensabili per acquisire la
capacità di eludere il controllo del sistema immunitario.

Progetto di Ricerca Giacomo Oliveira
13
0
20
40
60
80
100
% o
f GFP
+ ce
lls / l
euke
mic
cel
ls
Blood Spleen BM0
20
40
60
80
100
% o
f GFP
+ ce
lls / l
euke
mic
cel
ls
Blood Spleen BM
Spleen BM0
1
2
3
4
5
6
VCN
Spleen BM0
1
2
3
4
5
6
VCN
AML#1 AML#16
0 25 50 75 1000
200
400
600
800
1000
2000
4000
6000
8000
Days after infusion of leukemic cells
leuk
emic
cel
ls / μ
L
0 25 50 75 1000
200
400
600
800
1000
2000
4000
6000
8000
Days after infusion of leukemic cells
leuk
emic
cel
ls / μ
L
AML#1 AML#16A
B
CFigura 8. Trasduzione e attecchimento delle cellule leucemiche in topi NSG. (A) Crescita delle cellule leucemiche trasdotte con un vettore lentivirale (verde) o non trasdotte (blu) in seguito alla loro infusione in topi NSG. I dati mostrano che il processo di trasduzione non va ad inficiare la crescita delle due AML testate. (B) Percentuale di cellule trasdotte (GFP positive) rilevata su sangue periferico, nella milza e nel midollo osseo degli animali che hanno ricevuto cellule leucemiche trasdotte (verde) o non trasdotte (blu). (C) Quantità di vettore integrato nelle cellule trasdotte ed isolate dalla milza o dal midollo osseo degli animali. I dati mostrano come le cellule leucemiche isolate dai topi possano essere trasdotte con una buona efficienza con vettori lentivirali per poi essere infuse ed espanse in topi immunodeficenti.
AML#1 AML#16
Top Related