Tesi di Laurea - core.ac.uk · TTV è un virus a singolo filamento di DNA circolare a polarità...
95
1 Facoltà di Scienze Matematiche, Fisiche e Naturali Laurea Magistrale in Biotecnologie Molecolari e Industriali Tesi di Laurea Viremia di TTV come marker surrogato dello stato immunologico di pazienti trapiantati di organo solido. Candidata: Relatori: Santi Melissa Prof. Mauro Pistello Dott. Fabrizio Maggi ANNO ACCADEMICO 2011/2012
Transcript of Tesi di Laurea - core.ac.uk · TTV è un virus a singolo filamento di DNA circolare a polarità...

1
Facoltà di Scienze Matematiche, Fisiche e Naturali
Laurea Magistrale in Biotecnologie Molecolari e
Industriali
Tesi di Laurea
Viremia di TTV come marker surrogato dello
stato immunologico di pazienti trapiantati di
organo solido.
Candidata: Relatori: Santi Melissa Prof. Mauro Pistello Dott. Fabrizio Maggi
ANNO ACCADEMICO 2011/2012

2
INDICE
RIASSUNTO pag. 4
ABSTRACT pag. 6
CAPITOLO 1 – INTRODUZIONE
1.1 Torqueteno virus: la scoperta. pag. 8
1.2 Organizzazione genomica di TTV. pag. 9
1.3 Eterogeneità genetica. pag. 14
1.4 Replicazione. pag. 16
1.5 Classificazione. pag. 18
1.6 Patogenesi. pag. 24
1.7 Epidemiologia. pag. 27
1.8 Diagnosi. pag. 30
1.9 Associazione con la patologia. pag. 32
1.10 TTV associato a patologie renali e/o pancreatiche. pag. 35
1.11 Trapianto di organo. pag. 37
CAPITOLO 2 – MATERIALI E METODI
2.1 Pazienti e campioni. pag. 41
2.2 Estrazione del DNA virale. pag. 42
2.3 Real-time PCR. pag. 44
2.4 PCR genogruppo-specifiche. pag. 49
2.5 Corsa elettroforetica su gel di agarosio ed estrazione della banda. pag. 50
2.6 Sequenziamento. pag. 52
2.7 Analisi statistica dei dati. pag. 55
CAPITOLO 3 – RISULTATI
3.1 Presenza e titolo di TTV nel plasma pre-trapianto. pag. 57
3.2 Cinetica dei titoli di TTV nel primo periodo post-trapianto. pag. 58
3.3 Cinetica dei titoli di TTV nel lungo periodo post-trapianto. pag. 60

3
3.4 Titolo di TTV e numero di organi trapiantati. pag. 62
3.5 Titolo di TTV e caratteristiche del donatore. pag. 63
3.6 Cinetica dei titoli di TTV in relazione al tipo di trattamento immunosoppressivo.
pag. 64
3.7 Confronto tra la cinetica dei titoli di TTV e il numero di linfociti totali.
pag. 66
3.8 Andamento dei titoli di TTV in pazienti con o senza riattivazione di CMV.
pag. 68
3.9 Studio dei genogruppi di TTV. pag. 70
CAPITOLO 4 – DISCUSSIONE E CONCLUSIONI pag. 73
CAPITOLO 5 – BIBLIOGRAFIA pag. 78

4
RIASSUNTO
Torque Teno Virus (TTV) è stato identificato nel 1997 da un gruppo di ricercatori
giapponesi che indagavano sulle possibili cause di epatiti post-trasfusionali a eziologia
sconosciuta. Il clone N22 isolato dai loro studi, si rivelò un nuovo virus inizialmente
associato a patologie epatiche. TTV è un virus a singolo filamento di DNA circolare
a polarità negativa, privo di envelope e con genoma di circa 3,8 Kb. È in grado di
indurre viremia cronica nell’80% della popolazione mondiale e, una volta contratta,
l’infezione può persistere nell’organismo anche per molti anni. Attualmente la sua
elevata distribuzione e l’elevato tropismo nell’ospite, lo rendono un virus non
associato ad alcuna una patologia specifica dell’uomo, e per questo definito “orfano”.
Tuttavia studi recenti hanno mostrato un’associazione tra presenza di TTV e alcune
patologie a carico del sistema respiratorio, come lo sviluppo dell’asma. La
replicazione del virus avviene principalmente nelle cellule del sistema immunitario:
intermedi replicativi sono stati identificati in linfociti T e B, natural killer e altri
polimorfonucleati. Alcuni studi hanno ipotizzato una possibile correlazione tra titoli
di TTV e stato del sistema immunitario, come è stato confermato in pazienti
sottoposti a trapianto di organo solido. Questo lavoro di tesi si è concentrato quindi
sullo studio della viremia indotta da TTV in pazienti trapiantati di rene e/o pancreas
che avevano ricevuto diverse terapie immunosoppressive. Lo scopo è stato quello di
validare la viremia di TTV come marcatore surrogato della funzionalità immune in
soggetti sottoposti a diversi regimi di trattamenti immunosoppressivi. I pazienti
arruolati sono stati sottoposti a prelievi longitudinali di sangue e il titolo virale è stato
valutato a livello plasmatico tramite la metodica della real-time PCR. Per un pool
ristretto d’individui, che sono risultati altamente infetti, è stata fatta anche un’analisi

5
dei genogruppi tramite nested-PCR specifiche. I risultati ottenuti mostrano che
effettivamente i titoli di TTV aumentavano nel plasma di soggetti con sistema
immunitario compromesso in seguito alla terapia farmacologica somministrata.
L’analisi dei genogruppi invece, ha confermato l’elevata diversità genetica esistente
nel genere Anellovirus, ma non ha mostrato alcuna correlazione con lo “status” del
sistema immunitario. In conclusione, i dati ottenuti dall’analisi dell’andamento dei
titoli di TTV consentono di affermare che la sua viremia può essere considerata un
valido marker surrogato per lo studio delle condizioni del sistema immunitario e per
la diagnosi delle sue disfunzioni.

6
ABSTRACT
TTV is a single stranded circular DNA virus of about 3.8 kb that was first found in a
Japanese patient with non-A-E hepatitis. TTV is ubiquitous worldwide, it infects
about 80% of adults regardless race, gender, and geographic region. The virus may
persist for years in the infected host, with plasma levels ranging from 102 to 108
copies of viral DNA\ml. At the moment, no human disease has been clearly
associated with this virus; many virologists considered TTV as an orphan virus,
commonly present in humans where it represents a large part of virome. Recent
evidences have demonstrated that TTV viremia is present at higher levels in patients
with compromised immune system than in ones with normal immunity. Starting
from these observations, the aim of the present study was to evaluate the relationship
between TTV loads and status of immune system in patients with solid organ
transplantation over time. TTV viremia was quantified by using an universal real-time
PCR designed on a region of the viral genome highly conserved among all TTVs
known, plasma samples obtained before and at selected time points post-transplant
were tested, and the levels of TTV were correlated with several parameters measured
in the course of clinical follow-up (i.e. type of drug therapy administered, donor’s
characteristics, number of transplant, CMV reactivation, etc…). The results obtained
demonstrate that an impaired immune system promotes TTV replication and
consequently induces increased levels of viremia in transplant patients In conclusion,
the study demonstrates that measuring TTV plasma levels over time might be useful
to define the functionality of immune system in the infected host and help in
determining when the patient’s immune system has recovered a good level of activity.

7
CAPITOLO 1
INTRODUZIONE

8
1.1 Torque Teno virus: la scoperta
Torque Teno Virus è un virus a DNA circolare a singolo filamento privo di
envelope, isolato nel 1997 da un gruppo di ricercatori giapponesi che stavano
effettuando studi su pazienti affetti da epatite post-trasfusionale a eziologia
sconosciuta, per cercare di identificare i possibili agenti che causano questo tipo di
patologia. Tramite un’analisi di rappresentazione differenziale fu isolato un clone di
500 bp, denominato N22, che inizialmente fu ritenuto il responsabile della patologia
(Nishizawa et al., 1997). La tecnica utilizzata è stata sviluppata nel 1993 da Lisitsyn e
consente di mettere a confronto due sequenze di DNA presenti all’interno del
medesimo soggetto: il DNA derivato dai tessuti patologici e quello che proviene dai
tessuti sani (Lisitsyn et Wigler, 1993). Nel caso del clone N22 il materiale genetico è
stato prelevato ed estratto dal siero nel momento in cui il paziente presentava un
picco nei livelli di transaminasi, ed è stato confrontato con il materiale ottenuto
quando le transaminasi mostravano valori nella norma. Il clone fu denominato in
seguito TT virus dalle iniziali del paziente da cui è stato possibile isolarlo e, tramite
un confronto in molteplici banche dati (DDBJ/GenBank/EMBL DNA databases), è
stato visto che non mostrava alcuna omologia con nessun’altra sequenza nota. Studi
successivi hanno permesso il clonaggio di un genoma virale di 3739 nucleotidi (nt)
denominato TA278, che è stato ipotizzato essere a singolo filamento di DNA poiché
sensibile alla DNasi I, alla Mung Bean nucleasi e alla Rnasi A (Okamoto et al., 1998).
Gli studi effettuati da Miyata e collaboratori nel 1999, hanno dimostrato che
nell’isolato TA278 è presente una sequenza di circa 100 nt ricca in GC nella regione
non codificante (Miyata et al, 1999). Tale regione si è rilevata particolarmente
importante poiché forma una struttura secondaria necessaria per la replicazione

9
virale. Tramite la tecnica della PCR inversa, è stato possibile dimostrare la natura
circolare del genoma, perché è stato osservato che le estremità 3’ e 5’ erano legate
l’una con l’altra (Okamoto et al., 1999). Dopo la sua scoperta sono stati intrapresi
numerosi studi su TTV, che però non hanno chiarito totalmente le sue potenzialità
patogenetiche. Molti dubbi devono ancora essere risolti, soprattutto in seguito alla
dimostrata prevalenza del virus nella popolazione mondiale (Charlton et al., 1998;
Simmonds et al., 1998; Naoumov et al., 1998; Prescott et al., 1998; Tanaka et al.,
1998, Woodfield et al., 1998). Questo ha portato alla formulazione di varie ipotesi:
TTV potrebbe rappresentare un agente commensale apatogeno dell’uomo, in quanto
riscontrato indistintamente sia in soggetti malati che nei sani (Griffiths, 1999;
Simmonds, 2002), oppure un virus ancora “orfano” non ancora associato ad alcuna
patologia. Quest’ultima affermazione trova spiegazione anche nel fatto che ad oggi
esistono numerose specie di TTV con alcune di esse assai più rare e meno studiate di
altre. Con il tempo, dunque, potrebbero essere scoperte nuove correlazioni tra
infezione di TTV e specifici stati patologici (Bendinelli et al., 2001).
1.2 Organizzazione genomica di TTV
Data la mancanza di un sistema di coltura cellulare in vitro capace di far replicare
TTV, le informazioni circa le caratteristiche strutturali del virus derivano
principalmente da studi condotti su particelle virali purificate da plasma o da feci. Gli
studi portati avanti da Itoh e collaboratori (2000) hanno permesso di visualizzare la
particella virale al microscopio elettronico: da campioni di siero è stata isolata la
particella virale complessata con le IgG, mentre da campioni fecali il virus è stato

10
isolato dal sovranatante e si ritrova sottoforma di particella libera. TTV è di forma
sferica con un capside a simmetria icosaedrica ed è privo di envelope esterno. Il
diametro del virus è risultato essere compreso tra 30 e 32 nm, come già ipotizzato in
esperimenti che prevedevano l’utilizzo di filtri di policarbonato a porosità
decrescente (Mushahwar et al., 1999). La resistenza ai detergenti (Okamoto et al.,
1999), ai solventi e al calore secco hanno confermato l’assenza di un involucro
lipidico esterno. Tale assenza è stata ulteriormente confermata dalla scoperta di TTV
nella bile di soggetti infetti (Ukita et al., 2000), dove la presenza costante degli acidi
biliari consente la sopravvivenza dei virus che non possiedono una porzione lipidica
esterna. Al contrario del materiale fecale dove le particelle risultano libere, alcuni
lavori riportano l’esistenza nel sangue di particelle virali formanti complessi con
immunoglobuline (Ukita et al., 2000).
Il genoma di TTV è un singolo filamento di DNA circolare approssimativamente di
3.9 kb, ma tale lunghezza può variare leggermente da un genotipo all’altro. A tal
proposito gli studi di Okamoto e collaboratori sono stati fondamentali. Difatti, a
seguito dell’azione della DNasi I e della Rnasi I cui è stato sottoposto l’acido nucleico
estratto dal plasma, la sua sensibilità e resistenza ai due enzimi, rispettivamente,
hanno fatto ipotizzare la presenza di un virus a DNA. Inoltre, testando l’azione della
nucleasi Mung Bean e dell’endonucleasi di restrizione Nde I, è stata confermata la
natura a singolo filamento di DNA (Okamoto et al., 1998; Mushahwar et al., 1999).
Mediante ultracentrifugazione in gradiente di saccarosio è stato possibile costatare
che il virus era privo di envelope poiché la sua densità in tale gradiente si manteneva
costante anche dopo trattamento con tween-80 (Okamoto et al. 1998b).
Successivamente, tramite saggi di ibridazione con molecole di RNA a polarità
positiva e successiva digestione, si è potuto dimostrare che il DNA di TTV aveva una

11
polarità negativa (Mushahwar et al., 1999). Infine, tramite la scoperta della regione di
113 nt ricca in GC e della PCR inversa è stato possibile definire la sua natura
circolare (Okamoto et al., 1999).
Dal punto di vista strutturale, il genoma di TTV può essere suddiviso in una regione
non codificante (UTR) di circa 1.2 kb dove sono concentrati gli elementi regolatori e
una regione codificante (ORFs) approssimativamente di 2.6 kb (Kamada et al., 2004).
Nella regione codificante sono presenti due geni principali, ORF1 e ORF2, localizzati
a livello dell’antigenoma (filamento complementare positivo nel genoma) e sono
parzialmente sovrapposti. ORF1 è il gene più lungo e comprende circa i 2/3
dell’intera lunghezza del genoma. Prendendo come riferimento il ceppo TA278, il
gene ORF1 è trascritto a partire dal nucleotide 589 al 2898 con una capacità
codificante di 770 amminoacidi (aa). La proteina prodotta si suppone abbia
principalmente un ruolo strutturale, a formare cioè il capside, ma probabilmente
assume anche un ruolo nella replicazione del virus in quanto possiede attività
elicasica. Questa ipotesi è stata avvalorata dalla presenza di un dominio idrofilico N-
terminale ricco in arginina, lungo all’incirca 100 aminoacidi, che potrebbe interagire
con il DNA genomico per mediarne il trasporto nel nucleo della cellula ospite (Shoya
et al. 1998; Vihinene-Ranta et al. 1997). All’estremità C-terminale sono presenti due
motivi amminoacidici delle replicasi Rep (molto conservati tra i membri del genere
Circovirus) che vengono sfruttati generalmente dai virus a DNA nella replicazione a
cerchio rotante. Inoltre sono stati evidenziati, nella regione centrale, dei potenziali siti
di glicosilazione e tre regioni ipervariabili (HVR 1, 2 e 3), che risultano diverse a
seconda dell’isolato e conferiscono alla proteina ORF1 proprietà biologiche e
antigeniche variabili (Hijikata et al. 1999).

12
Il gene ORF2 è localizzato tra i nucleotidi 353 e 718 dell’isolato TA278, e codifica
per una proteina non strutturale di circa 150 aa importante per la replicazione virale.
Se la trascrizione inizia con il primo codone ATG, e non con il secondo come
descritto in precedenza, è possibile avere una proteina più lunga, di 202 amminoacidi,
anch’essa con un ruolo replicativo, che sembra essere coinvolta nei meccanismi di
trasmissione dei segnali. Tale proteina possiede una doppia funzione di fosfatasi:
tirosina- e serina/treonina-fosfatasi (Peters et al., 2002). Dall’analisi delle sequenze di
molti isolati, TTV risulta spesso contenere altre ORFs minori, denominate ORF4 e 5,
di cui la prima è altamente conservata mentre la seconda è piuttosto variabile (Tanaka
et al. 2000b; Kakkola et al. 2002). Tali ORFs sono localizzate in prossimità della
porzione C-terminale dell’ORF1 e sono a essa sovrapposte. Sono stati identificati tre
diversi mRNA prodotti da TTV di 3.0, 1.2 e 1.0 kb, aventi polarità negativa e siti
terminali al 3’ e al 5’ in comune (Kamahora et al., 2000). Recentemente è stato
scoperto che il rapporto tra i tre tipi di mRNA è di 60:5:35 e tutti i possibili trascritti
sono stati ritrovati nelle cellule (Qiu et al., 2005). E’ stato ipotizzato che il
meccanismo di splicing per produrre gli mRNA mediante l’utilizzo di diversi schemi
di lettura, agisca sulle regioni ORF conosciute ma anche su ORFs minori. I tre
mRNA si formano grazie ai processi di ricombinazione omologa che avvengono tra
le ORF 1, 2, 4 e 5. In particolare, il messaggero più lungo conterrebbe per intero il
gene ORF1, mentre i due mRNA più corti risulterebbero dalla ricombinazione
dell’ORF2 con l’ORF4 o l’ORF5. I prodotti proteici derivanti dall’ORF3 (o ORF2-4)
e dall’ORF5 potrebbero essere coinvolti nella replicazione del virus o nei meccanismi
di controllo e regolazione della trascrizione virale. In particolare l’ORF3 sembra
codificare per una proteina di 105 aa che può indurre apoptosi in linee cellulari
derivate da carcinoma epatocellulare umano (Kooistra et al. 2004) in modo analogo

13
all’apoptina del virus dell’anemia del pollo (CAV). Dalla sequenza nucleotidica di
alcuni isolati di TTV possono essere dedotte altre ORFs, ma il loro significato
funzionale resta al momento sconosciuto (Yokoyama et al. 2002).
Nonostante l’elevata divergenza nucleotidica che si riscontra nelle regioni codificanti,
le regioni altamente conservate si ritrovano a livello della UTR che contiene i
promotori e i siti di splicing. La regione non codificante è lunga 1,2 kb ed è
caratterizzata dalla presenza di una sequenza costituita per il 90% da guanina e
citosina (Miyata et al., 1999), e forma una struttura secondaria caratterizzata da motivi
ad ansa associati a funzioni regolatorie. All’interno di questa sequenza è presente
inoltre un tratto di 36 nucleotidi che mostra un alto grado di omologia con la stessa
regione in CAV (Miyata et al., 1999). Ciò la rende un dominio di particolare
importanza per la sopravvivenza del virus, poiché è conservato in tutti i ceppi di
TTV. Per questo motivo tali regioni sono state scelte come bersaglio di
amplificazione per la diagnosi molecolare. Nell’UTR, inoltre, sono presenti siti di
legame per fattori di trascrizione quali NF-Kb, SP-1, ATF\CREB e AP-2 (Miyata et
al., 1999). Esperimenti di mutagenesi hanno reso evidente la presenza di alcuni
elementi regolatori, sia negativi sia positivi, con probabile attività cellulo-specifica
(Suzuki et al., 2004). Studi recenti hanno inoltre identificato il sito d’inizio della
trascrizione dei tre mRNA osservati, che sembra essere localizzata 113 nt a valle
rispetto al promotore basale costituito dal dominio TATA box (Kamada et al., 2004).

14
Figura 1. Organizzazione genomica di un isolato rappresentativo di TTV. È indicato
l’isolato di riferimento (TA278), la lunghezza in nucleotidi (3853), la porzione
codificante con i quattro geni (ORFs)e la porzione non tradotta (UTR) con la breve
regione ricca in guanina e citosina (GC). (Okamoto, 2009)
1.3 Eterogeneità genetica
Sebbene TTV sia un virus a DNA, presenta una notevole divergenza genetica,
paragonabile ai virus a RNA. La regione più conservata del genoma è quella non
tradotta (UTR) con un’identità che può variare dal 73% tra gli isolati TA278 e
SANBAN, che sono i più lontani da punto di vista filogenetico, fino al 90%. Su
questa regione sono stati disegnati primers in grado di amplificare tutti gli isolati di
TTV, che allo stesso tempo hanno mostrato un’elevata variabilità genetica a livello

15
della regione codificante (Hallett et al. 2000; Hijikata et al. 1999b; Khudyakov et al.
2000; Muljono et al. 2001; Okamoto et al. 1999a, c, d, 2000d, 2001; Peng et al. 2002;
Takahashi et al. 2000a; Ukita et al. 2000). Tale divergenza è particolarmente marcata a
livello dell’ORF1, dove sono presenti tre regioni ipervariabili rispettivamente di 22,
47 e 31 aa, caratterizzate da inserzioni o delezioni multiple di codoni e una
divergenza della sequenza amminoacidica che può raggiungere il 70%. La porzione
ammino-terminale ricca in arginina è molto più conservata, così come altri brevi
frammenti che potrebbero assumere un ruolo fondamentale nella formazione e nella
funzionalità di tale proteina. (Erker et al. 1999; Hijikata et al. 1999; Takahashi et al.
1998b). Nishizawa e collaboratori hanno dimostrato che le regioni HVR presentano
un aumento della variabilità durante la fase acuta dell’infezione, mentre rimangono
più conservate nel momento in cui l’infezione termina, suggerendo dunque che tali
regioni siano necessarie per permettere a TTV di sfuggire alla risposta immunitaria
dell’ospite (Nishizawa et al., 1999). Tuttavia, non è stata riscontrata per tutti gli isolati
di TTV la medesima variabilità all’interno delle HVRs (Ukita et al., 2000). Altre
ipotesi sono state proposte per spiegare l’elevato grado di variabilità tra gli isolati
virali di TTV, tra cui l’uso di polimerasi cellulari con scarsa attività proof-reading
durante il processo replicativo e fenomeni di ricombinazione, che sembra avvengano
più frequentemente a livello dell’UTR. La recente identificazione di virus infettanti
primati escluso l’uomo, indistinguibili da quelli umani, ha fatto supporre che la
necessità di adattarsi ad un nuovo ospite possa rappresentare un importante fattore
che aumenta l’eterogeneità a livello genomico (Romeo et al., 2000; Takayama et al.,
1999; Tanaka et al., 1998). La frequenza con cui avvengono le sostituzioni
nucleotidiche in alcune regioni del genoma (7,3 x 104 per sito per anno nelle HVRs di
SENV) è simile ai virus ad RNA, ovvero circa 10 volte maggiore di quelle di altri

16
virus a DNA, quali il virus dell’epatite B (Umemura et al., 2001). Per avvalorare
l’ipotesi della ricombinazione omologa, in isolati dello stesso genotipo e in isolati di
genotipo diverso, sono stati identificati 19 siti di ricombinazione omologa di cui 13
localizzati all’interno della UTR, che potrebbe quindi rivestire una certa importanza
nell’ampliare la diversità genetica (Worobey, 2000; Biagini et al. 2001b; Manni et al.
2002; Erker et al. 1999; Hijikata et al. 1999), soprattutto se si considera che la
coinfezione con diversi isolati di TTV risulta un evento estremamente comune
(Worobey, 2000; Niel et al. 2000; White et al. 2000). Infine la capacità di TTV di
produrre infezioni croniche caratterizzate da una viremia nel plasma che può durare
anche molti anni, può indurre il sistema immunitario dell’ospite a esercitare una
continua e forte pressione evolutiva (Prescott et al., 1999).
1.4 Replicazione
La maggior parte dei virus a DNA per replicarsi dipende sostanzialmente dal
macchinario replicativo della cellula ospite. I virus infettano le cellule in attiva
divisione o inducono la cellula ospite ad entrare in fase S, prevenendo il processo
apoptotico. Il processo tramite il quale TTV riesce a replicarsi non è stato ancora
compreso appieno. In base alle analogie che TTV condivide con alcuni virus che
infettano gli animali, quali CAV e il Circovirus porcino di tipo 2 (PCV2), si ipotizza
che la replicazione sia strettamente dipendente da enzimi di origine cellulare, in
quanto non esistono geni in grado di codificare per DNA polimerasi o per
retrotrascrittasi. Inoltre, la mancanza di sequenze in grado di alterare il ciclo cellulare
della cellula ospite (Todd et al., 2001) e la capacità dei circovirus di propagarsi nelle

17
cellule linfoidi attivate (Nielsen et al., 2003), spingono a supporre che il loro
meccanismo replicativo dipenda dallo stato di attivazione della cellula, e in particolare
da proteine prodotte durante la fase S del ciclo cellulare (Fields et al., 1996). A
dimostrazione di quanto riportato sopra, TTV è stato ritrovato in cellule in attiva
divisione quali epatociti, cellule del midollo osseo (Okamoto et al., 2000; Okamoto e
Mayumi, 2001) e cellule linfoidi stimolate (Maggi et al., 2001; Mariscal, 2002). Il
processo replicativo in sé avviene probabilmente all’interno del nucleo, mentre le
modalità di assemblaggio del virione non sono ancora note. Come detto in
precedenza, a causa dell’elevata eterogeneità che caratterizza TTV (Umemura et al.,
2002), è probabile che nel processo replicativo sia coinvolta una polimerasi cellulare
con scarsa attività di “correzione delle bozze”. Il rilascio della particella virale matura,
essendo TTV un virus privo di envelope ed involucro lipidico, avviene
probabilmente attraverso la lisi della cellula ospite che quindi va incontro a morte. È
stato ipotizzato che TTV utilizzi, analogamente ai Circovirus, il meccanismo a
cerchio rotante per la replicazione del genoma, che porterebbe alla formazione di un
intermedio replicativo a doppio filamento di DNA. Tale ipotesi è stata avvalorata
dall’esistenza di alcuni motivi amminoacidici, tipici delle proteine Rep, individuati
nella regione ORF1 dei virus caratteristici dell’altra famiglia virale (Erker et al., 1999;
Mushahwar et al., 1999; Tanaka et al., 2000). Anche se nelle cellule del fegato e del
midollo osseo sono stati osservati intermedi replicativi di TTV costituiti da un
doppio filamento di DNA (Okamoto et al., 2000d), nel genoma di TTV manca la
tipica sequenza indispensabile per l’innesco della replicazione secondo il modello
rolling-circle (Erker et al. 1999). Per questo motivo sono state valutate altre ipotesi
come la presenza di un intermedio a RNA ma, non essendo presente nel genoma
alcuna sequenza che codifichi per una retrotrascrittasi virale, tale possibilità risulta

18
poco probabile (Nishizawa et al., 1999). Il genoma di TTV è stato rilevato in molti
tipi di campioni biologici, indicando come esso possa infettare un ampio spettro di
tipi cellulari. Ciò ha fatto pensare che i recettori utilizzati da TTV durante la fase di
riconoscimento cellulare siano probabilmente molecole localizzate a livello di diversi
tessuti, ma non è da escludere l’utilizzo di co-recettori da parte dei differenti
genogruppi e/o genotipi virali.
1.5 Classificazione
Subito dopo la sua scoperta TTV fu classificato all’interno della famiglia Parvoviridae,
in quanto privo di envelope lipidico e con il genoma lineare a singolo filamento di
dimensioni ridotte. Con la scoperta, tramite gli studi di Miyata e collaboratori, della
natura circolare del genoma, è stato necessario rivedere la precedente classificazione.
Esistono altre famiglie virali in cui sono presenti virus a singolo filamento: i virus
batterici appartenenti alla famiglia Microviridae, i virus vegetali della famiglia
Geminiviridae e quelli appartenenti alla famiglia Circoviridae. I Geminivirus e i
Microvirus hanno un genoma con più di una copia di DNA circolare, perciò TTV fu
inserito nella famiglia Circoviridae in quanto aveva mostrato anche una parziale
omologia con CAV (il virus dell’anemia del pollo ora classificato nel genere
Gyrovirus), ipotizzando che fosse il primo rappresentante umano di questa famiglia
(Miyata et al., 1999). Il genoma di CAV è difatti paragonabile a quello di TTV sia per
dimensioni (3200 basi di CAV contro le 3800 di TTV) sia per quanto riguarda la
suddivisione del genoma: entrambi hanno la regione codificante suddivisa in 3
principali ORFs parzialmente sovrapposte, e una regione UTR ricca in GC. In

19
seguito però è stato visto che tra i membri della famiglia Circoviridae e TTV c’erano
notevoli differenze, soprattutto per quanto concerne la sequenza nucleotidica. Negli
anni successivi sono state isolate e caratterizzate numerose sequenze virali associabili
a TTV, che mettono in luce la straordinaria variabilità genetica del virus: tra questi
ricordiamo gli isolati PMV (Hallet et al., 2000), SANBAN (Hijikata et al., 1999) e
YONBAN (Takahashi et al., 2000), con meno del 50% di omologia con il clone di
riferimento TA278. Questa caratteristica, come anche altre, sono uniche tanto da far
considerare TTV come il solo rappresentante di una nuova famiglia, inizialmente
definita Circinoviridae (Mashahwar et al., 1999). Dopo la scoperta di TT-like mini virus
(TTMV), aventi caratteristiche intermedie tra TTV e CAV (il genoma era più piccolo,
di 2800 basi, ma con la stessa organizzazione di TTV), è stato proposto di classificare
i tre virus all’interno di una nuova famiglia definita Paracircoviridae (Takahashi et al.,
2000; Biagini et al. 2001b). Nel corso degli anni sono stati isolati molti altri virus
simili a TTV. Nel 2005 gli studi di Jones e collaboratori portarono alla scoperta di
due nuovi agenti virali aventi un genoma di 2249 nt e 2635 nt, che furono nominati
rispettivamente Small Anellovirus 1 (SAV 1) e Small Anellovirus 2 (SAV 2),
collocabili per le dimensioni del genoma tra TTV e TTMV. I due SAV presentano tra
di loro un’omologia di sequenza pari al 54% e con TTV e TTMV un’omologia
rispettivamente del 32% e del 35%. Questi virus furono classificati nel genere
Anellovirus, poiché possiedono un DNA circolare a singolo filamento con polarità
negativa, una regione codificante suddivisa in più ORFs parzialmente sovrapposte ed
una regione non codificante UTR. Sequenze correlate a SAV sono state poi
identificate nel 2007. Tali sequenze, di circa 3200 nt (cioè di dimensioni intermedie
tra TTV e TTMV) (Ninomiya et al. 2007a; 2007b) appartengono ad un nuovo
anellovirus, definito TTMDV (Torque Teno Midi virus), caratterizzato da un’elevata

20
omologia con SAV, ma da esso differente per la più ampia estensione della regione
non codificante. Ciò ha suggerito che SAV possa essere una forma mutante di
TTMDV dove è avvenuta una delezione.
La classificazione dei membri appartenenti al genere Anellovirus è tuttora piuttosto
complessa. I primi lavori di analisi filogenetiche si basavano sull’amplificazione e
sull’allineamento delle regioni N22 dell’ORF1, e ciò ha permesso in un primo
momento di classificare gli isolati fino a quel momento conosciuti in 23 genotipi
compresi in quattro raggruppamenti filogenetici principali (Okamoto et al., 1999;
Biagini et al., 1999; Ukita et al., 2000). Tuttavia, la breve regione N22 si è rivelata
poco adatta per studi filogenetici e per identificare tutte le specie di TTV nella loro
completezza ed eterogeneità, in quanto specifica per il clone TA278. La successiva
analisi delle sequenze dell’intero genoma e della regione non tradotta UTR, ha
permesso di identificare oltre 40 genotipi e 70 sottotipi. Nel 2002, Peng e
collaboratori suggerirono una nuova classificazione degli isolati di TTV fino ad allora
conosciuti, che prevedeva 5 principali genogruppi, identificati numericamente da 1 a
5, sulla base di divergenze nucleotidiche di almeno il 50% (Peng et al., 2002). Inoltre
era prevista un’ulteriore distinzione in genotipi, identificati con numeri arabi e con
una divergenza nucleotidica del 30%, e in sottotipi, identificati da lettere minuscole e
con una divergenza compresa tra il 15 e il 29%. Seguendo questa nuova
classificazione, l’isolato TA278 (prototipo di TTV) era compreso nel genotipo 1 del
genogruppo 1, dove ritroviamo anche i genotipi dal 2 al 6 (Okamoto et al., 1999). Nel
genogruppo 2, rappresentato dall’isolato PMV, erano inclusi i genotipi 7, 8, 22, 23 e
17 (Hallet et al., 2000). I genotipi dal 9 al 16 (SANBAN), il 18, 19 (SENV-F) e 20
(SENV-G) e altri sette genotipi di SENV (da A ad H) erano inseriti nel genogruppo
3. Infine, nel genogruppo 4 (prototipo YONBAN) si distinguevano nove genotipi,

21
mentre nel genogruppo 5 erano classificati quattro isolati ed il prototipo scelto era
JT33. Studi recenti condotti da Jelcic e collaboratori (2004) hanno permesso di
inserire i ceppi di TTV in un’unica famiglia, definita Anelloviridae. Secondo questa
nuova classificazione, il genogruppo 1 sarebbe inserito nel genere α, i genogruppi 2 e
5 nel genere β, mentre i genogruppi 3 e 4 nei generi γ e δ rispettivamente. Sulla base
di questo nuovo criterio di classificazione, gruppi di TTV con identità di sequenza
inferiore al 50% rappresentano generi separati, all’interno dei quali gli isolati hanno
tra loro un’omologia di sequenza compresa tra il 50% e il 60%. All’interno di ogni
genere, è prevista un’ulteriore classificazione in specie quando un gruppo di isolati di
TTV presentano tra di loro un’identità di sequenza superiore al 60%. Si parla invece
di tipi e sottotipi virali quando l’identità di sequenza arriva rispettivamente al 90% e
al 96% (Jelcic et al., 2004). Nel 2001, l’International Committee on Taxonomy of
Viruses (ICTV) ha assegnato a TTV il nome di Torque Teno virus ed è stato inserito
in una nuova famiglia definita Anelloviridae dove i termini “torques” collana e “tenuis”
sottile, ricordano le caratteristiche del suo genoma. L’ultima classificazione, nonché la
più recente, prevede che all’interno della famiglia Anelloviridae siano compresi undici
generi, ciascuno comprendente un diverso numero di specie.

22
Tabella 1. Classificazione attuale della famiglia Anelloviridae approvata dall’ICTV.
L’ICTV sta attualmente studiando la possibilità di inserire all’interno della famiglia
Anelloviridae altri tre generi: Mutorquevirus, Nutorquevirus e Xitorquevirus, al cui
interno sono presenti isolati scoperti molto recentemente. Contemporaneamente
sono in corso di svolgimento degli studi filogenetici basati sulle sequenze
nucleotidiche che cercano di inserire anche due nuove sottofamiglie: Gyrovirinae e
Tornovirinae. Il genere Gyrovirus è attualmente inserito all’interno della famiglia delle
Circoviridae ed ha come prototipo CAV ma, diversamente da altri membri della stessa
famiglia, contiene elementi virali che mostrano caratteristiche comuni agli anellovirus:
Possiedono un DNA circolare a singolo filamento a polarità negativa molto
simile.

23
La regione non codificante di CAV, come quella di molti altri anellovirus, è ricca
in GC.
Le proteine codificate dall’ORF2 di CAV e di molti anellovirus, contengono una
sequenza che è caratteristica di una proteina tirosin-fosfatasi (PTPasi) e inoltre
condividono un motivo comune: WX7HX3CXCX5H.
Le proteine del capside sia di CAV che degli anellovirus, possiedono una
sequenza amminoacidica che ha un motivo caratteristico delle proteine Rep
(deputate alla replicazione del genoma virale).
Alcuni trascritti derivanti dallo splicing rilevati in CAV e in molti Anellovirus,
sono in grado di indurre apoptosi selettivamente in linee cellulari provenienti da
carcinoma umano.
Proprio sulla base delle sovra descritte considerazioni, il genere Gyrovirus è stato
assegnato alla famiglia Anelloviridae, nella sottofamiglia delle Gyrovirinae.
Considerazioni analoghe possono essere fatte anche per la classificazione della
sottofamiglia Tornovirinae che è composta da virus recentemente identificati nelle
tartarughe marine. Nonostante le caratteristiche comuni delle tre sottofamiglie
(Anello-Gyro-Tornovirinae) vi è comunque un’alta divergenza di sequenza che non
permette ad oggi di avere un albero filogenetico globale che rappresenti l’insieme
della famiglia Anelloviridae.

24
Figura 2. Nuova proposta di classificazione per la famiglia Anelloviridae.
1.6 Patogenesi
Il virus penetra nell’ospite attraverso molteplici porte d’ingresso (il tratto respiratorio,
il sistema gastro-intestinale, l’apparato genitale e la placenta) e dopo poche settimane
è rilevabile nel sangue. Questo è stato dimostrato in diversi lavori in cui è stata
evidenziata la sua presenza in un’ampia varietà di campioni biologici: siero, feci (Itoh
et al., 2000), latte materno, saliva, bile (Hayakawa et al., 2001), secrezioni vaginali,
liquido spermatico, fluido duodenale, liquido sinoviale e fluido cerebrospinale (Deng
et al., 2000; Biagini et al., 2001a; Taijri et al., 2001; Maggi et al., 2001a; Fornai et al.,
2001; Nakagawa et al., 2000; Inami et al., 2000). La viremia è evidente dopo alcune
settimane dall’infezione e spesso rimane evidenziabile anche per lunghi periodi di

25
osservazione (Mushahwar et al., 1999; Tawara et al., 2000). TTV nella specie umana
può comportare una viremia cronica che persiste nell’80% degli individui infettati
(Takayama et al., 1999b; Ali et al., 2002; Lefrère et al., 1999; Matsumoto et al., 1999)
con titoli virali medi di 104 copie (da 102 a 109) di TTV per ml di plasma (Hu et al.,
2005; Pistello et al., 2001) che possono mostrare ampie fluttuazioni o mantenersi
relativamente stabili nel tempo. Uno studio sulla cinetica della clearence di TTV
suggerisce che sia necessaria una produzione maggiore di 1010 virioni al giorno per
mantenere questi livelli di viremia (Maggi et al., 2001b), indicando così che il virus
non rimane latente nella cellula ospite in forma episomiale o integrata come
osservato per altri virus a DNA (Tanaka et al., 2000a; Yu et al., 2002). Grazie
all’ibridazione in situ e alla PCR quantitativa è stato dimostrato che TTV replica nel
fegato (Rodriguez-Iñigo et al. 2000; Ohbayashi et al. 2001; Luo et al. 2000; Nakagawa
et al. 2000; Ukita et al. 1999, Okamoto et al. 2000e). L’escrezione nella bile può essere
la principale fonte di diffusione di TTV tramite il tratto gastrointestinale. Tuttavia
sembra che la replicazione non sia limitata a tale organo, poiché forme replicative di
DNA a doppio filamento e mRNA sono stati ritrovati anche in polmone e pancreas
(Okamoto et al. 2001), midollo osseo (Fanci et al. 2004; Kikuchi et al. 2000;
Okamoto et al. 2000b), milza (Okamoto et al. 2001; Jelcic et al. 2004) e in altri tessuti
linfoidi (Kakkola et al. 2004). Il DNA di TTV è stato rilevato nei PBMC (Barril et al.
2000; Lopez-Alcorocho et al. 2000; Okamoto et al. 1999a, 2000c; Okamura et al.
1999) dove mostra un ampio tropismo essendo presente non solo nei linfociti T e B,
nei monociti e nelle NK (Maggi et al. 2001a; Takahashi et al. 2002; Zhong et al.
2002), ma anche nei granulociti e negli altri polimorfonucleati (Maggi et al. 2001a;
Takahashi et al. 2002). La replicazione avviene con cinetiche paragonabili a quelle di
altri virus persistenti, quali il virus dell’epatite B (HBV), il virus dell’epatite C (HCV) e

26
il virus dell’immunodeficienza umana (HIV). L’emivita di TTV sembra essere di circa
4 ore (Maggi et al., 2001b). Molti studi hanno dimostrato che l’infezione viene
frequentemente contratta durante la prima infanzia. La presenza di co-infezioni è
frequente, con una percentuale del 65% rilevata in bambini di età inferiore ai sette
anni, che può aumentare fino al 90% in soggetti adulti sani (Ukita et al., 2000; Saback
et al., 2002; Maggi et al., 2005). Ciò suggerisce l’esistenza di meccanismi di evasione
immune evoluti da TTV per stabilire infezioni persistenti in individui
immunocompetenti. La persistenza di TTV nell’organismo ospite potrebbe essere
causata dall’incapacità del sistema immunitario di controllare l’infezione, poiché gli
anticorpi anti-TTV prodotti non sembrano in grado, almeno nella maggior parte dei
casi, di eliminare totalmente il virus o di evitare le infezioni sostenute da più
genogruppi (Ukita et al., 2000; Saback et al., 2002; Maggi et al., 2005). Le IgM
compaiono dopo 10-12 settimane, rimanendo diagnosticabili nel sangue per qualche
tempo e poi scompaiono. Le IgG invece sono prodotte dopo 5-11 settimane e
tendono a persistere per anni (Tsuda et al. 2001) o indefinitamente, tanto che
anticorpi verso il virione (Tsuda et al. 1999) o verso la proteina ricombinante ORF1
(Handa et al. 2000; Ott et al. 2000) si ritrovano in soggetti viremici e non. Studi sui
complessi di TTV con le IgG in circolo, hanno mostrato che non esiste alcuna
associazione con patologie da deposito di immunocomplessi quali la glomerulonefrite
e, in alcuni casi, è stata riportata la riduzione del titolo virale in seguito alla
formazione di tali complessi (Nishizawa et al. 1999; Itoh et al. 2000; Tsuda et al.
1999). In generale la produzione di anticorpi non è sufficiente a contrastare
l’infezione: la viremia, infatti, persiste nonostante la presenza di alti titoli anticorpali
e, come si è detto, sono frequenti le superinfezioni con ceppi virali diversi.

27
Sebbene sia un evento abbastanza raro, in letteratura sono stati descritti anche eventi
di infezione che si sono risolti anche dopo mesi o anni dal contagio (Lefrere et al.,
1999; Matsumoto et al., 1999; Yuki et al., 1999), e una rapida eliminazione del virus è
stata osservata in scimpanzé sperimentalmente infettati (Mushahwar et al., 1999).
Tale fenomeno non è chiaro e potrebbe essere spiegato da: una ridotta immissione in
circolo del virus, un aumento della sua eliminazione, una modificazione delle
sequenze virali che non permettono più la rilevazione del DNA o dalla possibilità che
il virus stabilisca uno stato di latenza. A tal proposito è stato osservato che
l’apparente scomparsa del virus dal plasma non è necessariamente associata alla
completa eradicazione dell’infezione (Maggi et al., 2001c; Pollicino et al., 2002).
1.7 Epidemiologia
TTV è globalmente diffuso nel mondo e può essere rilevato, sotto forma di infezione
cronica, nel sangue di circa 2/3 della popolazione generale indipendentemente da
origine etnica, età, condizioni socio-economiche o altre variabili (Biagini et al., 2000;
Biagini et al., 2001a; Matsubara et al., 2001; Niel e Lampe, 2001). I primi studi di
prevalenza si basavano sulla regione ORF1 utilizzata come bersaglio di
amplificazione nelle reazioni di PCR. I risultati ottenuti confermavano l’elevata
prevalenza di TTV ma con percentuali diverse in base alla zona considerata (Niel et
al., 1999): molto bassa in Stati Uniti e Nord Europa e molto elevata in Africa e Sud
America (Prescott e Simmonds, 1998). La successiva introduzione della PCR
universale, basata sull’amplificazione della regione non tradotta (UTR), ha mostrato
però una distribuzione mondiale senza differenze significative tra le varie aree

28
geografiche (Biagini et al., 2000; Biagini et al., 2001a; Matsubara et al., 2001; Niel e
Lampe, 2001). In particolare è stato rilevato che TTV è molto diffuso in Asia, con
una prevalenza del 90%, mentre è risultato meno diffuso in Nord America e in
Oceania (40%). È opportuno precisare che l’uso della PCR permette di rilevare solo
le forme attive dell’infezione, sia acute sia croniche, ma non è in grado di evidenziare
i casi di infezione pregressa. Quindi, si può ipotizzare che le percentuali di prevalenza
siano sottostimate, in quanto la presenza di TTV è stata confermata in diversi tessuti
anche in assenza di viremia (Maggi et al., 2001c; Pollicino et al., 2002). I dati
disponibili per TTMV mostrano similarità con quelli ottenuti per TTV (Biagini et al.,
2000; 2001). In tutti gli studi sopracitati non è stata rilevata alcuna associazione tra la
prevalenza di TTV e il quadro clinico dei soggetti, il sesso e/o altre caratteristiche
della popolazione analizzata. Tuttavia, alcuni lavori hanno evidenziato che i livelli di
prevalenza del virus tendono ad aumentare con il passare degli anni, raggiungendo
l’apice in età adulta (Vasconcelos et al., 2002; Maggi et al., 2003a; Kazi et al., 2000;
Ninomiya et al., 2008; Chen et al., 1999; Maggi et al., 1999; Oguchi et al., 1999).
Comunque, è opportuno sottolineare che nella prima infanzia il virus è già
ampiamente diffuso (Davidson et al., 1999; Hsieh et al., 1999; Vasconcelos et al.,
2002; Maggi et al., 2003). Una così elevata prevalenza dell’infezione presuppone che
TTV sia altamente contagioso e che la sua diffusione avvenga attraverso molteplici
vie di trasmissione. Tra queste, quelle sicuramente dimostrate includono la via
parenterale (per l’elevata percentuale di positività nei soggetti esposti al sangue o agli
emoderivati come gli emofilici, emodializzati, politrasfusi, talassemici, e
tossicodipendenti); la via oro-fecale (per la presenza di TTV nelle feci di soggetti
viremici); e la via materno-fetale (Morrica et al., 2000; Okamura et al. 1999; Hsieh et
al., 1999; Yokozaki et al., 1999a). Il genoma di TTV è stato ritrovato anche in

29
campioni di latte materno (Davidson et al., 1999). È stata ipotizzata, ma non ancora
confermata, la trasmissione di TTV per via sessuale, a causa del ritrovamento del
virus in liquido vaginale, liquido spermatico e saliva (Gerner et al., 2000; Liu et al.,
2000; Matsubara et al., 2000; Matsubara et al., 2001; Biagini et al., 2001a; Chan et al.,
2001; Fornai et al., 2001; Davidson et al., 1999). Lavori recenti hanno confermato la
capacità di TTV di replicarsi nell’epitelio respiratorio, nei polmoni, nelle tonsille e
nelle adenoidi (Pollicino et al., 2002), e ciò suggerisce anche un coinvolgimento della
via aerea per la trasmissione virale. Particolare rilievo assume il fatto che molti
animali da allevamento (bovini, suini, ovini e pollame) e animali domestici (cani e
gatti) sono con frequenza portatori di virus identici o molto simili ai TTV nell’uomo
(Leary et al., 1999; Okamoto et al., 2002), e potrebbero favorire una trasmissione
interspecifica. Non è ancora chiaro se i diversi genotipi/genogruppi di TTV siano
trasmessi con una diversa efficienza ma appare evidente che alcuni sono più diffusi di
altri, e si potrebbe pensare che la diversa distribuzione dipenda dall’area geografica
e/o dal tipo di popolazione esaminata: ad esempio, il genotipo 1 appare più diffuso
in Asia anziché nei Paesi occidentali (Shibata et al., 2001; Umemura et al., 2001a), e i
genotipi 22 e 23 sono prevalenti in Indonesia, ma praticamente assenti in Giappone
(Muljono et al., 2001). Tali scoperte tendono a suggerire una trasmissione
differenziale dei diversi genotipi/genogruppi. Studi di prevalenza dei diversi genotipi
di TTV e TTMV hanno evidenziato una più ampia diffusione di alcuni genogruppi
rispetto ad altri, come emerge in un lavoro di recente pubblicazione in cui i
genogruppi 1 e 3 sono stati ritrovati con maggiore frequenza nella popolazione
generale rispetto al 4 e 5, mentre il genogruppo 2 è quello più raro (Maggi et al,
2006).

30
1.8 Diagnosi
La diagnosi di infezione da TTV risulta ancora abbastanza primitiva e basata soltanto
su metodi di analisi molecolare quali l’amplificazione degli acidi nucleici (one-step
PCR, Nested-PCR e Real-time PCR), poiché non esistono, ad oggi, né un sistema di
coltura cellulare in grado di supportare la replicazione virale (Kakkola et al, 2007), né
test sierologici per la determinazione degli anticorpi e degli antigeni virali.
Inizialmente, quando non era ancora nota l’enorme eterogeneità del genoma virale
dei vari isolati di TTV, fu scelta la porzione N22 a livello della ORF1 come bersaglio
per l’amplificazione tramite nested o semi-nested PCR (PCR N22). Questa regione si
è rilevata non idonea in quanto riusciva ad amplificare solo i genotipi dall’1 al 6 (Itoh
et al., 1999). Tale limitazione è stata in parte superata utilizzando un set multiplo di
primer (Davison et al., 1999; Desai et al., 1999). In seguito sono stati disegnati altri
primers sulla regione UTR più conservata (Okamoto e Mayumi, 2001), tuttora
utilizzati (PCR UTR o universale). Utilizzando la Real-time PCR universale è
possibile identificare la quasi totalità degli isolati di TTV: successive reazioni di PCR
consentono di identificare genogruppi specifici, tramite amplificazione di regioni
diverse del genoma (Maggi et al., 2001b) e, in questo caso, il sequenziamento è
necessario per confermare i risultati ottenuti. L’introduzione della PCR universale ha
aumentato notevolmente la percentuale di campioni analizzati che sono risultati
positivi grazie ad un’aumentata sensibilità: studi di Takahashi hanno dimostrato un
miglioramento dal 23 al 92%, mentre nelle prove effettuate da Itoh e collaboratori è
stato evidenziato un aumento delle reazioni positive dal 20 al 95% (Takahashi et al.,
1998; Itoh et al., 1999). Tutte le metodiche sopracitate consentono l’individuazione
della sola forma virale in attiva replicazione all’interno dell’ospite: come già

31
accennato, questo comporta una sottostima dell’effettiva prevalenza di TTV nella
popolazione mondiale ritenendo dunque necessario sviluppare nuovi test sierologici
in grado di identificare forme pregresse d’infezione. Tuttavia sono stati eseguiti alcuni
saggi di immunoblotting ed immunoprecipitazione combinata con la PCR. Per
adesso, l’unico sistema che permetta di ottenere degli antigeni, sembra consistere
nell’utilizzo di particelle virali intere o proteine derivanti dalle ORF1 e ORF2 inserite
all’interno di vettori e fatte esprimere nelle cellule batteriche. E’ in questo modo che
in siero umano sono state identificate IgG rivolte contro il virus, permettendo così di
valutare la percentuale di TTV immunocomplessato. Inoltre, utilizzando come
antigene il virione intero estratto da campioni fecali, sono state rilevate anche IgM
specifiche (Nishizawa et al., 1999; Tsuda et al., 1999; Handa et al., 2000; Ott et al.,
2000). Tuttavia, tali metodi non hanno fornito risultati riproducibili considerando che
nei diversi studi i soggetti sani risultati positivi per anticorpi anti-TTV di tipo G
variavano dal 10 al 98% (Handa et al., 2000; Ott et al., 2000; Tsuda et al., 2001;
Kakkola et al., 2002). Ciò suggerisce che il tipo di antigene utilizzato è in grado di
influenzare notevolmente la prestazione del saggio sierologico. La messa a punto di
metodi sierologici sensibili e specifici consentirebbe di individuare tutti i soggetti che
durante la loro vita sono già venuti a contatto con il virus e che al momento
dell’indagine sono risultati negativi per la presenza di TTV.

32
1.9 Associazione con la patologia
In generale, le interazioni che gli anellovirus sono in grado di stabilire con l’ospite
sono ancora poco conosciute. Quando TTV fu scoperto, grazie al lavoro intrapreso
da Nishizawa e collaboratori, fu considerato come la possibile causa di epatiti non A-
G fulminanti, che spesso colpivano pazienti che avevano subito trasfusioni di sangue
a seguito di un intervento chirurgico (Nishizawa et al., 1997). Ben presto però è stato
scoperto che la prevalenza di TTV era estremamente alta nella popolazione
mondiale, senza alcuna discriminazione di sesso, età, etnia, condizioni economiche
e/o altro. Questi dati hanno messo in dubbio l’effettivo potenziale patogenetico di
TTV, che attualmente alcuni ricercatori hanno definito apatogeno, mentre altri
addirittura lo hanno inquadrato come un semplice commensale della flora microbica
umana (Griffiths, 1999; Mushahwar, 2000). Ulteriori studi sono quindi necessari per
chiarire a fondo tale condizione.
Ad oggi, gli sforzi effettuati per trovare una correlazione con una patologia specifica
sono tra i più svariati. Alcuni lavori hanno proposto delle correlazioni deboli tra TTV
e patologie quali: il diabete, la fibrosi polmonare idiopatica, l’artrite reumatoide ed
emodializzati con bassi livelli di emoglobina (Hirata et al., 1998; Bando et al., 2001;
Boysen et al., 2003). Altri studi, condotti su anemia aplastica, crioglobulinemia, lupus
eritematoso e psoriasi, non hanno ancora dimostrato alcuna correlazione significativa
(Maggi et al., 1999; Kikuchi et al., 2000; Cacoub et al., 2003).
Per quanto riguarda l’associazione di TTV con la patologia epatica, è presente un
numero maggiore di studi. In letteratura sono riportati lavori volti a sostenere
l’associazione tra lo sviluppo di epatocarcinomi e alti titoli di TTV, che avevano

33
valore prognostico nei pazienti con epatite cronica da HCV (Tokita et al., 2002). Tale
condizione può essere spiegata assumendo che TTV possa svolgere un effetto
negativo sulla progressione della malattia cronica, oppure che un sistema immunitario
compromesso da HCV determini l’aumento del titolo di TTV, come avviene nel caso
di HIV, in cui a bassi valori dei CD4+ corrispondono alti titoli di TTV (Christensen
et al., 2000). Il lavoro di Zein e collaboratori (1999) ha dimostrato che TTV infetta in
misura maggiore pazienti con cirrosi scompensata piuttosto che quelli affetti da
epatite cronica e cirrosi compensata. Esistono dati certi sulla sua replicazione nel
fegato ma, al momento, non può essere considerato un virus responsabile di forme di
epatite (Matsumoto et al., 1999; Kao et al., 2000; Shang et al., 2000; Ali et al., 2002).
D’altro canto sono stati pubblicati lavori che fanno emergere dei dubbi, in quanto
animali quali scimpanzé, naturalmente o sperimentalmente infettati con TTV, non
mostrano sempre un danno a livello epatico (Mushahwar et al., 1999; Verschoor et
al., 1999).
Relativamente alle patologie respiratorie sono stati approfonditi molti aspetti
sull’insorgenza dell’asma in bambini affetti da bronchiettasie o malattie respiratorie
acute, dove sono stati rilevati titoli virali molto più elevati rispetto a soggetti sani o
con malattie meno gravi (Maggi et al., 2003a, 2003b; Pifferi et al., 2006). La presenza
di elevati titoli di TTV nell’epitelio nasale e livelli alterati delle spirometrie di soggetti
asmatici (Pifferi et al., 2005), ha fatto supporre un collegamento con il virus, che
potrebbe stimolare la risposta immunitaria da parte dei linfociti T helper-2: com’è
noto, questi ultimi rappresentano uno dei meccanismi di sviluppo dell’asma (Pifferi et
al., 2005).

34
Alcuni studi hanno mostrato la replicazione di TTV a livello di midollo osseo, fatto
che è stato correlato con lo sviluppo di anemia aplastica associata a epatite ad
eziologia sconosciuta (Kikuchi et al., 2000; Miyamoto et al., 2000). I dati ottenuti
però sono piuttosto contrastanti e non dimostrano un’effettiva significatività. Il DNA
di TTV si può ritrovare in linfociti circolanti e nei linfonodi di pazienti con linfomi a
cellule B e linfomi di Hodgkin (Garbuglia et al., 2003). Oltre al tumore epatocellulare,
polmonare e ai linfomi, TTV è stato ritrovato in altri tessuti neoplastici, ma non
esiste ancora un’associazione plausibile tra infezione da TTV e trasformazione
cellulare o tumorigenesi.
Molto interessanti si sono rivelati gli studi volti a dimostrare una correlazione tra
presenza di TTV e condizione del sistema immunitario: è stato ipotizzato che un
sistema immunitario compromesso favorisca un aumento della replicazione virale.
Diversi lavori hanno evidenziato livelli elevati di TTV in pazienti
immunocompromessi quali: pazienti HIV+, pazienti con patologie tumorali o
soggetti con patologie immunomediate come l’artrite reumatoide (Christensen et al.,
2000; Shibayama et al., 2002; Zhong et al., 2001). L’incremento della viremia di TTV
è stato rilevato anche in individui a cui erano stati somministrati farmaci
immunosoppressivi prima di essere sottoposti a trapianto d’organo (Shang et al.,
2000). Tutto ciò suggerisce un possibile ruolo del sistema immunitario nel modulare
la replicazione del virus: grazie agli studi descritti, è stato possibile dimostrare che nel
corso del progressivo indebolimento della risposta immunitaria dell’ospite, si
dovrebbe assistere ad un aumento della carica virale (Simmonds, 2002).
Le similarità con i circovirus animali hanno inoltre suggerito l’eventualità che alcune
delle complicanze ematologiche rilevate in pazienti immunocompromessi come la

35
neutropenia, la trombocitopenia e l’anemia, possano in qualche misura dipendere
dall’incremento dei livelli di TTV nel midollo osseo (Kikuchi et al., 2000; Tokita et
al., 2001c; Simmonds, 2002). L’elevata eterogeneità che caratterizza l’intera famiglia
delle Anelloviridae (TTV, TTMV e TTMDV) potrebbe comportare la presenza di
genotipi che assumono un ruolo differente in relazione al particolare tipo di
patologia.
1.10 TTV associato a patologie renali e/o pancreatiche
Per quanto riguarda le patologie renali e/o pancreatiche, esistono pochi lavori che
hanno cercato di evidenziare una qualche correlazione tra la presenza di infezione da
TTV e patologie relative a tali organi. Relativamente alle malattie pancreatiche,
Tomasiewicz e collaboratori hanno osservato in due pazienti TTV-positivi, ricoverati
per un’epatite ad eziologia sconosciuta, lo sviluppo di un cancro al pancreas
(Tomasiewicz et al., 2005). Tale relazione è difficile da confermare ma può essere
utilizzata come punto di partenza per studi volti a comprendere meglio il ruolo di
TTV nella formazione della neoplasia. Più numerose sono le informazioni che
provengono da studi effettuati su pazienti con patologie renali. A tal proposito, è
stata dimostrata una prevalenza di TTV molto più elevata in soggetti sottoposti ad
emodialisi rispetto ai controlli sani, e spesso si ritrovano delle co-infezioni con HCV
e HBV (Gallian et al., 1999; Rivanera et al., 2008; Chattopadhyay et al., 2005).
Solitamente il virus non mostra un effetto clinico importante nei pazienti, ma rimane
la possibilità che possa aggravare la malattia epatica causata da HCV quando ci si
trova dinanzi a un’infezione multipla (Rivanera et al., 2008). È stato inoltre

36
evidenziato, in un esperimento condotto in vitro, un possibile ruolo di TTV nel
generare alterazioni in cellule renali, simili a quelle osservate nella sindrome nefrosica
(Yokoyama et al., 2002). A tale scopo sono stati creati topi transgenici per le varie
ORF di TTV e, tra gli undici topi studiati, due mostravano alterazioni a livello renale:
entrambi portavano porzioni diverse dell’ORF1. L’effetto più o meno rilevante nel
livello di malformazione delle cellule renali era dovuto non tanto alla presenza del
transgene, ma piuttosto alla quantità di proteina prodotta. Altri studi sono stati
condotti su soggetti sottoposti a trapianto renale: la percentuale di positivi può
variare dal 33% nelle popolazioni indiane e brasiliane, al 66% come osservato in un
gruppo di pazienti giapponesi (Michitaka et al., 2000; Yokosuka et al., 2000; Abraham
et al., 2003). I risultati ottenuti mostrano che alcuni individui acquisiscono
un’infezione da TTV solo dopo il trapianto e spesso non esiste una correlazione con
fattori di rischio quali età, livello di ALT, numero di trasfusioni oppure i mesi
trascorsi dopo il trapianto. Anche in questo caso il virus si trova in un contesto di co-
infezione insieme con altri virus epatici (Abraham et al., 2003). Un successivo studio,
condotto su un gruppo di pazienti sottoposti a trapianto renale, ha sottolineato la
persistenza di isolati del genogruppo 1 in questo organo (Szladek et al., 2003).
Tuttavia, i risultati descritti sopra non ci permettono di affermare con certezza
l’effettivo coinvolgimento di TTV nel causare tali patologie, e sono necessari ulteriori
approfondimenti.

37
1.11 Trapianto di organo
Con il termine trapianto è indicata una procedura chirurgica di sostituzione di cellule,
tessuti o organi non funzionanti in un paziente ricevente, e la loro sostituzione con la
componente omologa proveniente da un soggetto donatore. Nella pratica clinica il
trapianto è utilizzato come unica possibilità di cura per molte patologie degenerative,
non aggredibili con ulteriori terapie. Esistono più tipologie di trapianto, distinguibili
in base alle caratteristiche del donatore o al tipo di trapianto stesso. Si parla di
trapianto ortotopico quando l'organo originario degenerato viene rimosso e quello del
donatore viene piazzato nella stessa posizione anatomica, mentre viene definito
eterotopico quando il nuovo organo è affiancato a quello vecchio non più funzionante,
che però rimane localizzato nella medesima posizione. Inoltre, è possibile fare
un’ulteriore distinzione in:
Autotrapianto, quando il ricevente e il donatore sono la stessa persona e il rischio
di rigetto è assente. È un’opzione che si presenta utile nel caso di interventi
programmati ed è spesso associata all’autotrasfusione.
Allotrapianto, quando il ricevente e il donatore appartengono alla stessa specie, ma
il rischio di rigetto aumenta fortemente.
Isotrapianto, quando ricevente e donatore sono due individui geneticamente
identici come nel caso dei gemelli omozigoti, dove il rischio di rigetto è eliminato.
Xenotrapianto, quando il ricevente e il donatore appartengono a specie diverse e
quindi l’organo o tessuto trapiantato deve essere opportunamente trattato in
modo da eliminare, almeno in parte, la risposta immunologica.
L’animale più studiato e utilizzato per questo tipo di trapianti è il maiale, poiché
presenta caratteristiche anatomiche molto simili all’uomo. Il problema principale a

38
cui va incontro un soggetto ricevente è quello del rigetto, condizione patologica in
cui il sistema immunitario riconosce l’organo trapiantato come non-self alla stregua di
virus e batteri. Il rigetto può essere più o meno lieve e può compromettere o meno la
funzionalità del nuovo organo e la vita stessa dell’individuo. Per eliminare o
comunque diminuire la risposta del sistema immunitario esistono diverse strategie:
tutte mirano alla riduzione della potenza della reazione immunitaria, scatenata dal
riconoscimento da parte dei linfociti T di molecole definite alloantigeni presenti
sull’organo trapiantato. Solitamente sono utilizzate combinazioni di farmaci definiti
appunto anti-rigetto che possono essere suddivisi nelle seguenti classi:
Inibitori della calcineurina: Ciclosporine e Tacrolimus. Questi farmaci
impediscono l’attivazione da parte della calcineurina del fattore di trascrizione
NFAT (Nuclear Factor of Activated T cells), fondamentale per attivare una
cascata di segnali che porta alla trascrizione di alcuni geni in grado di produrre
proteine attivatrici dei linfociti T quali IL-2, IL-4, TNF-α.
Inibitori di mTOR: Sirolimus e Everolimus. Impediscono l’attività di mTOR
(mammalian target of rapamycin), una proteina chinasi coinvolta nella
regolazione di crescita cellulare, proliferazione, motilità, sopravvivenza delle
cellule, sintesi proteica e trascrizione.
Antiproliferativi: Azatioprina e Micofenolato mofetil. Sono tossine metaboliche
che uccidono preferibilmente i linfociti proliferanti e che quindi sono venuti a
contatto con gli antigeni.
Corticosteroidi: Prednisolone e Idrocortisone. Sono ottimi antinfiammatori e
sono spesso usati in combinazione con altri farmaci per la terapia anti-rigetto.
Anticorpi: possono essere monoclonali, come il Basiliximab, o policlonali, come
la globulina antitimocitaria (ATG). Il Basiliximab è un anticorpo chimerico (topo-

39
uomo) che si lega alla catena α (CD25) dell’IL-2 inibendo la divisione dei linfociti
T; l’ATG invece è ottenuta dall’immunizzazione di conigli o cavalli e mira a
distruggere i linfociti T. Spesso vengono somministrati in associazione con i
glucocorticoidi.

40
CAPITOLO 2
MATERIALI E METODI

41
2.1 Pazienti e campioni
Per questo lavoro di tesi sono stati selezionati 114 soggetti adulti, suddivisi
principalmente in base al tipo di organo trapiantato: 51 di essi avevano subito
trapianto di organo solido (rene e/o pancreas) da donatore vivente, 63 erano soggetti
trapiantati con organo ottenuto da cadavere. In base alla tipologia di trapianto i
pazienti sono stati suddivisi in cinque gruppi principali:
1. N° 37: trapianto di rene (KTA);
2. N° 19: trapianto simultaneo di rene e pancreas (SPK);
3. N° 2: trapianto di pancreas (PTA);
4. N° 5: trapianto di rene e successivamente di pancreas (PAK);
5. N°51: trapianto di rene da donatore vivente (LDKT).
La popolazione in esame può essere suddivisa anche secondo il tipo di trattamento
farmaceutico ricevuto; in base alla terapia infatti, i pazienti sono stati trattati con due
diversi farmaci immunosoppressivi fondamentali per la terapia anti-rigetto:
l’anticorpo monoclonale Basiliximab (N°83) o la globulina anti-timocitaria ATG
(N°31), un anticorpo policlonale.
Tutti i pazienti, dopo aver firmato il consenso per essere inseriti nello studio, sono
stati sottoposti a prelievi longitudinali di sangue a partire dal momento antecedente al
trapianto, definito “tempo zero”. Gli altri prelievi sono stati eseguiti durante il
follow-up. I campioni sono stati suddivisi in due gruppi di studio: il primo, composto
dai prelievi di ciascun paziente effettuati in un breve periodo di tempo dopo il
trapianto che va da 7 a 14 giorni (“short term”), mentre il secondo, composto dai

42
prelievi effettuati a partire da 1 mese dal trapianto fino a raggiungere i due anni dallo
stesso (“long term”).
I campioni di sangue sono stati centrifugati a 1800 x g per 10 minuti. In questo modo
è stato possibile separare la parte corpuscolata, che rimane nella porzione inferiore
della provetta, dal plasma, localizzato più in superficie. Il plasma è stato quindi
prelevato e aliquotato all’interno di eppendorf da 500 µl, che sono state subito
trasportate presso il nostro laboratorio e conservate a -80°C fino al momento del
loro utilizzo.
2.2 Estrazione del DNA virale
Per effettuare l’estrazione del DNA virale a partire dal campione di plasma è stato
utilizzato il kit commerciale QIAamp DNA mini kit (QIAgen, Chatsworth, CA), che
consente la purificazione di tutto il DNA totale (genomico, mitocondriale e virale).
L’estrazione può essere eseguita a partire da diverse tipologie di campione tra cui
sangue intero, plasma, siero, linfociti, fluidi corporei, colture cellulari, tessuti e
campioni forensi. Seguendo il protocollo indicato, abbiamo utilizzato 200 µl di
campione per ogni estrazione effettuata: se il volume a nostra disposizione era
inferiore, è stata aggiunta soluzione fisiologica fino al raggiungimento della quantità
prevista dal kit. Il campione è stato dunque posto all’interno di eppendorf da 1.5 ml e
sono stati addizionati 20 µl di proteinasi K (20 mg/ml) e 200 μl di buffer AL per
effettuare la lisi cellulare. Al fine di ottenere una buona omogeneizzazione e
agevolare la lisi del campione, è stato necessario vortexare per alcuni secondi la
miscela, con conseguente incubazione a 56°C per 10 minuti. Trascorso questo tempo

43
è stata effettuata una rapida centrifugata per eliminare eventuali gocce evaporate fino
al tappo della eppendorf e successivamente sono stati aggiunti 200 µl di etanolo
assoluto (96-100%) per permettere la precipitazione degli acidi nucleici. Nuovamente
la miscela è stata vortexata e centrifugata, e poi rapidamente trasferita all’interno delle
colonne da 2 ml fornite dal kit (QIAamp spin columns). Queste particolari colonne
contengono all’interno una membrana in gel di silice che possiede un’elevata affinità
per gli acidi nucleici: il DNA difatti è stato in precedenza trattato in modo tale da
essere in grado di legarsi a tali membrane, grazie alle giuste condizioni di pH e salinità
del lisato. La colonnina è posta sopra un apposito tubo di scarico, dove possono
fluire le proteine e tutti gli altri possibili agenti contaminanti che potrebbero
interferire con le successive reazioni di amplificazione. E’ stata dunque eseguita una
prima centrifugata di 1 minuto a 8000 rpm (6000 x g) e il filtrato così ottenuto è stato
eliminato. Successivamente sono stati eseguiti due lavaggi della membrana con due
diversi tamponi in modo da aumentare la purezza del DNA ad esso legato ed
eliminare i contaminanti rimasti. Sono stati aggiunti 500 µl del primo tampone, AW1
(20 mM di NaCl, 2 mM di Tris-HCl, pH 7.5 e etanolo al 57%), e a seguito di una
centrifuga di 1 minuto a 8000 rpm (6000 x g) il filtrato è stato di nuovo scartato. Si
sono aggiunti 500 µl del secondo tampone, AW2, che differisce dal precedente solo
nel contenuto di alcool (etanolo 70%). A seguito di un'ulteriore centrifugata per 3
minuti alla massima velocità (14.000 rpm, 20.000 x g) il filtrato è stato nuovamente
scartato. Per eliminare in modo definitivo qualsiasi residuo di tampone
eventualmente rimasto nel filtro, abbiamo eseguito una nuova centrifugata a vuoto
alla massima velocità per un minuto. A questo punto si procede con l’eluizione del
DNA in una nuova eppendorf da 1,5 ml utilizzando 50 µl di buffer AE (10 mM Tris-
HCl, 0.5 mM EDTA, pH 9) il quale, mediante incubazione a temperatura ambiente

44
per almeno un minuto, seguita da una centrifugata a 8000 rpm (6000 x g) per un
minuto, permette al DNA di essere rilasciato dalla membrana di gel di silice. L’eluato
così ottenuto è stato conservato a -20°C fino al momento del suo utilizzo.
2.3 Real-time PCR
Il DNA estratto dai campioni longitudinali di plasma ottenuti prima e dopo il
trapianto, è stato testato per verificare la presenza del DNA virale. A tale scopo è
stata utilizzata la tecnica della Real-Time PCR che, oltre a verificare la presenza o
meno del virus nel campione in esame, è in grado di quantificare in modo molto
accurato le copie di genoma di TTV. La Real-Time PCR è una metodica che
permette di monitorare l’andamento della reazione mentre è ancora in svolgimento, e
i dati che si ottengono sono utilizzati per effettuare una quantificazione del
frammento amplificato. Il principio su cui si fonda la tecnica è uguale a quello su cui
si basano le altre tipologie di PCR: il DNA è amplificato da reazioni a catena della
DNA-polimerasi e, nel caso della PCR quantitativa, dopo ogni ciclo di amplificazione
il DNA viene quantificato. In questo contesto è stata utilizzata la tecnica Taqman,
basata sull’utilizzo di una sonda complementare alle sequenze target posta tra i siti di
legame dei primers senso e antisenso. Questa sonda è marcata su entrambe le
estremità: al 5’ è presente il fluorocromo R “reporter” (6-carbossi-fluorescina, FAM)
mentre al 3’ c’è una molecola Q “quencer” (6-carbossi-tetrametil-rodamina, TAMRA).
Solitamente il reporter è un fluoroforo ad alta energia che emette fluorescenza, mentre
il quencer è un fluoroforo a bassa energia che spegne la fluorescenza emessa dal
reporter. Quando la sonda è libera in soluzione, forma una struttura secondaria a

45
forcina e il quencer, trovandosi molto vicino al reporter, ne blocca la fluorescenza
assorbendo l’energia emessa. Quando la sonda si appaia con il DNA (ss)
complementare, l’attività esonucleasica 5’-3’ della Taq polimerasi la degrada,
allontanando così il quencer dal reporter che emette fluorescenza. Tale segnale è captato
da un apposito strumento che fornisce un’indicazione, in tempo reale,
dell’amplificazione del tratto di DNA ricercato. Sulla base del meccanismo sovra
descritto, la fluorescenza emessa è dunque direttamente proporzionale alla quantità
del DNA target presente nel campione esaminato, e dipende dai cicli ripetuti di PCR
eseguiti. L’andamento della reazione può essere monitorato tramite diagrammi di
fluorescenza che si basano sul numero di cicli svolti. Normalmente assume un
andamento di tipo sigmoide, con raggiungimento di un plateau finale tipico delle
reazioni di PCR, provocato dal progressivo esaurimento dei substrati (Figura 2.2).
L’incremento della fluorescenza è stato misurato ed interpretato dallo strumento
ICycler iQ5 (Bio-Rad Laboratories) costituito da: termociclizzatore, lampada laser per
indurre la fluorescenza, e software specifici per rilevare l’emissione di fluorescenza. Il
software calcola anche il ciclo soglia Ct, ovvero il ciclo a cui si osserva un aumento
significativo della fluorescenza, momento d’inizio della fase esponenziale dei prodotti
di PCR. Il sistema elabora una curva di taratura sulla base di alcuni standards, per i
quali è noto il numero di copie della sequenza di DNA target. Sull’asse delle ascisse è
riportata la concentrazione nota degli standards e sull’asse delle ordinate il
corrispondente ciclo soglia (Figura 2.3). Ciascun campione inserito è quantificato
utilizzando tale retta come riferimento, risalendo dal ciclo soglia alla quantità di DNA
presente.

46
Figura 2.2: Curve di amplificazione.
Figura 2.3: Retta di taratura costruita sulla base degli standards.

47
La miscela di reazione ha un volume finale di 25 µl ed è così composta:
Universal Master Mix, concentrazione finale 1X, costituita da polimerasi
AmpliTaq Gold™, enzima AmpErase uracil-N-glycosilase (UNG) e nucleotidi,
in cui dTTP è sostituito da dUTP, incorporato nella sintesi TaqMan al posto di
dTTP.
Primer senso AMT_S, concentrazione finale 0,9 µM.
(5’-GTGCCGIAGGTGAGTTTA-3’)
Primer antisenso AMT_AS, concentrazione finale 0,9 µM.
(5’-AGCCCGGCCAGTCC-3’)
Sonda AMT_P, concentrazione finale 0,1 µM.
(5’-FAM- TCAAGGGGCAATTCGGGCT -TAMRA-3’)
5 µl del DNA estratto.
X µl di acqua sterile, per raggiungere il volume finale di 25 µl.
L’enzima AmpErase (UNG), contenuto nella Universal Master Mix, agisce sul DNA
a doppio filamento andando a tagliare l’uracile, impedendo di conseguenza alla
polimerasi di continuare la sintesi di DNA e rendendo sensibile l’acido nucleico
all’idrolisi acido-basica. Per rendere possibile l’impiego dell’enzima UNG, la reazione
Taqman sostituisce dUTP al dTTP, cosicché eventuali contaminanti che si trovano
nel tubo di reazione vengono riconosciuti ed eliminati prima che la seduta di PCR
abbia inizio. L’enzima UNG è attivo grazie all’utilizzo di una temperatura di 50°C alla
miscela di reazione e viene in seguito inibito dall’elevata temperatura di
denaturazione prima che si svolga la reazione di PCR. Inoltre, per verificare che la
miscela preparata non abbia subito contaminazioni, si inseriscono nella reazione dei

48
controlli negativi che contengono, oltre alla mix, 5 µl di acqua sterile al posto del
DNA . Il protocollo di amplificazione prevede:
1 ciclo a 50°C per 2 minuti, per permettere all’enzima UNG di attivarsi e che sarà
poi inattivato dalle alte temperature nella fase successiva;
1 ciclo a 95°C per 10 minuti, dove viene attivata l’AmpliTaq Gold™ polimerasi;
45 cicli a 95°C per 15 secondi e a 50°C per 30 secondi, grazie ai quali si
favoriscono rispettivamente denaturazione e fase di ibridazione dei primers, e
successiva estensione.
Il limite di sensibilità di questa metodica è pari a 1,0 x 102 copie di DNA per ml di
plasma o per μg di DNA estratto.
In questo lavoro di tesi, al fine di valutare la presenza di TTV sono stati utilizzati dei
primers costruiti all’interno della regione UTR poiché risulta la più conservata tra
tutte le numerosissime specie di TTV esistenti (Figura 2.5). Ogni campione è stato
testato in doppio e i risultati dubbi sono stati più volte ripetuti.
Figura 2.4: Schema della regione UTR utilizzata come bersaglio per l’amplificazione.

49
2.4 PCR genogruppo-specifiche
Tra i pazienti risultati positivi a TTV in Real-Time PCR, 10 sono stati esaminati per
la determinazione dei genogruppi del virus implicati nell’infezione. A tale scopo sono
state effettuate 5 reazioni di PCR differenti, basate sull’amplificazione di 5 diverse
regioni genomiche in grado di discriminare tutti i genogruppi di TTV. Per aumentare
la specificità delle reazioni di amplificazione, sono state eseguite delle nested-PCR
condotte in due step successivi: i prodotti della prima reazione sono stati utilizzati
come DNA bersaglio in una seconda, basata sull’utilizzo di primers più interni
rispetto a quelli usati nel primo passaggio. Per tutte e 5 le reazioni di PCR
genogruppo-specifiche il protocollo utilizzato è lo stesso, sebbene differisca per due
aspetti: le coppie di primers e le rispettive Temperature di annealing sono specifiche
per ciascun genogruppo. Per il primo step la miscela, con volume finale di 50 µl per
tubo, è così composta:
Buffer di reazione (10 nM Tris-HCl pH 8,2, 50 mM KCl, 1,5 mM MgCl2);
Primer senso esterno con concentrazione finale di 0.6 µM;
Primer antisenso esterno con concentrazione finale di 0.6 µM;
Nucleotidi con concentrazione finale di 0.2 mM;
Taq polimerasi 0.025 U/µl;
5 µl di DNA estratto;
x µl di acqua sterile deionizzata a volume.
Preparata la miscela, sono stati effettuati 35 cicli con il seguente profilo termico:
Fase di denaturazione: 94 °C per 45 secondi;

50
Fase di annealing dei primer: temperatura di annealing di ciascuna coppia di
primer per 30 secondi;
Fase di estensione: 72°C per 1 minuto.
Al termine dei 35 cicli la temperatura è stata mantenuta a 72°C per 15 minuti, in
modo da consentire alla polimerasi di completare eventuali frammenti incompleti.
Dunque è stata preparata la miscela per il secondo step di PCR uguale alla precedente
tranne che per i primers utilizzati, che si appaiavano ad una regione più interna
rispetto ai primi. Alla miscela sono stati aggiunti 5 µl di prodotto derivato dal primo
step, ed è stata effettuata una nuova amplificazione per 25 cicli e con una diversa
temperatura di annealing specifica per i nuovi primers. I saggi sono stati eseguiti
mediante l’uso di termociclizzatori automatici Biorad T100TM Thermo Cycler.
2.5 Corsa elettroforetica su gel di agarosio ed estrazione
della banda
Gli amplificati ottenuti dalle PCR genogruppo-specifiche sono stati analizzati
mediante corsa elettroforetica su gel di agarosio che viene preparato ad una
concentrazione dell’1%. Il prodotto di PCR è miscelato con il “loading buffer”
(0,25% blu di bromofenolo; 0,25% xilene cianolo; 30% glicerolo), colorante con la
duplice funzione di facilitare la visualizzazione della corsa elettroforetica e di
“appesantire” i campioni, agevolandone la deposizione sul fondo del pozzetto del gel
grazie alla presenza del glicerolo. Come tampone di corsa viene usato il TAE 1X e i
campioni sono caricati nei singoli pozzetti. Si applica un voltaggio compreso tra 80 e
95 mV per circa 30 minuti. Il gel viene quindi osservato tramite un transilluminatore

51
grazie alla presenza del bromuro d’etidio, un intercalante del DNA che posto sotto
una luce ultravioletta, con lunghezza d’onda compresa tra 254 e 306 nm, emette una
tipica luce rosa-arancione. Le bande di DNA potranno essere confrontate con un
marker di peso molecolare corso contemporaneamente al campione in modo da
determinarne la lunghezza. Il frammento di nostro interesse può essere isolato da gel
utilizzando un bisturi sterile e, dopo aver determinato il suo peso con una bilancia di
precisione, si procede con l’estrazione del DNA utilizzando il kit commerciale
GenElute Extraction Kit (SIGMA), che permette di estrarre con facilità frammenti di
DNA lineari di dimensioni variabili (da 50 pb a 10 kb). Si aggiungono 3 volumi di
buffer Gel Solubilization Solution per ogni volume di gel, per ottenere la completa
solubilizzazione dell’agarosio e il successivo legame del DNA alla membrana nella
colonna. Il buffer contiene un indicatore di pH che semplifica la visualizzazione del
pH ottimale per il legame al DNA. Si procede quindi con un’incubazione della
soluzione ad una temperatura di 56 °C per 10 minuti. Si aggiunge un volume di
isopropanolo per aumentare la resa dell’estrazione e l’intera miscela è trasferita
all’interno delle colonnine di estrazione fornite dal kit, precedentemente preparate
tramite aggiunta di 500 μl di Column Preparation Solution per favorire il legame del
DNA alla membrana. Poi si esegue un lavaggio della membrana mediante aggiunta
di 700 µl di Wash Solution e si centrifuga a 13000 rpm per un minuto. La soluzione
di lavaggio contiene etanolo, il quale elimina residui di sali ed altre impurità
eventualmente presenti. Rimosso l’eluato, si effettua una centrifugata a 13000 rpm
per 1 minuto per eliminare qualsiasi traccia di etanolo rimasta, che potrebbe
compromettere la successiva eluizione del DNA. A questo punto le colonnine sono
trasferite in un nuovo tubo di raccolta e la membrana è reidratata con 50 µl di
Elution Solution (10 mM TRIS/HCl; pH 9). Si lascia ad incubare per 1 minuto e poi

52
si centrifuga a 13000 rpm per un altro minuto. L’eluato così ottenuto contiene il
frammento di DNA purificato.
2.7 Sequenziamento
I prodotti di PCR, una volta estratti dal gel, sono stati sequenziati per confermare il
genogruppo di TTV presente e identificare il tipo di isolato virale. Le sequenze sono
state analizzate mediante strumento ABI PRISM 373 DNA Sequencer (PE,
Biosystem) che si basa sul metodo enzimatico di Sanger. Quest’ultimo utilizza
nucleotidi modificati e marcati (dideossitrifosfato, ddNTPs) che causano la
terminazione della reazione di sintesi in posizioni specifiche. La reazione è allestita
utilizzando un filamento di DNA denaturato che viene suddiviso in quattro tubi di
reazione diversi, e a ciascuno vengono aggiunti: un primer per iniziare la reazione, la
DNA polimerasi, i 4 nucleotidi (dNTPs), e uno dei quattro dideossinucleotidi in
quantità stechiometricamente inferiore per permettere un’elongazione sufficiente del
filamento e poi terminare la reazione di polimerizzazione. Le quattro reazioni
vengono fatte correre su gel di poliacrilammide, che consente la risoluzione di
frammenti che differiscono anche per una sola base. Le bande sono visualizzate
tramite lastra autoradiografica o tramite luce UV a seconda del tipo di marcatura dei
ddNTPs, e la sequenza è letta direttamente dalla lastra o dal gel. Il nostro strumento è
automatizzato e vengono utilizzati dei ddNTPs marcati con fluorofori diversi che
emettono differente fluorescenza. Il campione unico di DNA viene corso in un
singolo pozzetto (o capillare), e un laser colpisce il campione in modo tale che ogni
fluoroforo emetta la propria fluorescenza che sarà captata da un rilevatore. Un

53
apposito software elaborerà i dati ottenuti, dando come risultato la sequenza intera in
forma grafica a quattro colori. La reazione di sequenza prevede una fase di
amplificazione seguita da un processo di elettroforesi. La reazione di amplificazione
utilizza una miscela già pronta denominata Mix Terminator Ready Reaction,
contenente tutti gli elementi necessari alla reazione a cui vengono aggiunti il primer e
il campione da sequenziare. Tale miscela contiene la AmpliTaq DNA polimerasi FS,
un tampone Tris-HCl pH 9, MgCl2, deossinucleosidi trifosfato (dATPs, dCTPs,
dUTPs, dGTPs) e i quattro dideossinucleotidi marcati. L’enzima è associato ad una
pirofosfatasi stabile al calore che determina la scissione del pirofosfato inorganico
generato nella formazione del legame fosfodiestere durante l’estensione del filamento
di DNA, impedendone così l’accumulo. Un’alta concentrazione di questa molecola
favorirebbe la reazione inversa alla polimerizzazione: i nucleosidi marcati potrebbero
essere staccati dal filamento in crescita e sostituiti da quelli non marcati. La miscela di
reazione del volume totale di 20 μl è così composta:
4 μl di Terminator Ready Mix;
3,2 μl di primer diluito 1:20;
X μl di campione da valutare in seguito alla concentrazione di DNA presente;
Y μl di acqua sterile deionizzata per raggiungere il volume totale.
La miscela viene in seguito introdotta in un termociclizzatore e sottoposta a un
programma di 25 cicli secondo il seguente schema:
Fase di denaturazione: 96 °C per 10 secondi;
Fase di annealing del primer: 50 °C per 5 secondi;
Fase di estensione: 60 °C per 4 minuti.

54
Il prodotto di PCR così ottenuto deve essere purificato prima di procedere con
l’elettroforesi utilizzando colonnine Centri-Sep (Princeton Separation), costituite da
un gel liofilizzato che deve essere idratato con 800 μl di acqua sterile deionizzata.
Dopo aver lasciato le colonnine ad idratare per circa 30 minuti, si elimina l’acqua in
eccesso per sgocciolamento e, successivamente, eseguendo una centrifugata a 650 x g
per 2 minuti. A questo punto si trasferisce la miscela proveniente dalla reazione di
amplificazione al centro delle colonnine contenenti il gel idratato e si centrifuga
nuovamente a 650 x g per 2 minuti, ottenendo così il campione purificato dalle
impurità. Cinque μl del campione sono diluiti con 15 μl di acqua sterile deionizzata e
il DNA viene sottoposto a denaturazione alla temperatura di 94°C per circa 2 minuti;
subito dopo i tubini vengono posti in ghiaccio per impedire ai singoli filamenti di
DNA di appaiarsi di nuovo. A questo punto i campioni sono pronti per il processo di
elettroforesi. I frammenti di DNA denaturati sono sottoposti all’analisi eseguita dal
sequenziatore automatico. Inizialmente, sia il capillare che l’elettrodo sono immersi nel
tampone di corsa (Genetic Analyzer Buffer) in modo da generare la differenza di potenziale,
poi vengono a contatto con il primo campione. L’iniezione di quest’ultimo all’interno del
capillare avviene in seguito al flusso di corrente che si genera dal catodo verso l’anodo.
Dopo, il catodo e il capillare sono nuovamente immersi nel tampone, così da generare una
nuova differenza di potenziale che consente la migrazione dei frammenti di DNA attraverso
il polimero contenuto nel capillare (Performancer Optimizer Polymer 6%, POP-6). I
frammenti di DNA separati giungono quindi, in tempi diversi, nella “zona finestra” del
capillare, dove vengono a contatto con il laser con il conseguente eccitamento del
fluorocromo inserito che emette una tipica fluorescenza. Lo spettro di emissione di ciascun
fluorocromo è analizzato e convertito da segnale luminoso a segnale digitale dal programma
ABI PRISM DNA Sequencing Analysis Software. In questo modo il nucleotide finale di
ciascun frammento è identificato dal fluorocromo all’estremità 3’, e i frammenti di lunghezza

55
diversa permettono di discriminare gradatamente tutti i nucleotidi. Alla fine della corsa, la
sequenza di bande di DNA marcato viene visualizzata in un unico grafico detto
elettroferogramma, caratterizzato da una successione di picchi di 4 colori diversi,
corrispondenti alle emissioni fluorescenti dei diversi fluorocromi.
2.8 Analisi statistica dei dati
I titoli di TTV ottenuti mediante Real-Time PCR sono stati convertiti in scala
logaritmica in base 10 per approssimare i valori ad una distribuzione normale. Le
differenze tra le medie e le distribuzioni sono state valutate mediante l’uso del test t
di Student. Il valore di p è stato ritenuto significativo se inferiore a 0,05.

56
CAPITOLO 3
RISULTATI

57
3.1 Presenza e titolo di TTV nel plasma pre-trapianto
Come introdotto in Materiali e Metodi, la popolazione dello studio era costituita da
114 soggetti per ciascuno dei quali è stato possibile analizzare un campione di plasma
immediatamente prima del trapianto. Di questi soggetti, 51 erano soggetti trapiantati
di rene e/o pancreas che avevano ricevuto l’organo da donatore vivente, mentre 63
avevano ricevuto l’organo da cadavere. Tutti risultavano immunocompromessi a
causa della terapia di condizionamento effettuata e 32 erano al loro secondo
trapianto. Presenza e titolo di TTV sono stati valutati tramite una real-time PCR, con
primers universali disegnati su un frammento della UTR del genoma virale altamente
conservata tra tutti gli anellovirus, e dunque in grado di rilevare tutti gli isolati di TTV
ad oggi conosciuti. I risultati ottenuti nel plasma pre-trapianto dei diversi soggetti
esaminati sono riportati in Tabella 3.1. Come mostrato, TTV era altamente presente
in tutti i soggetti analizzati con una prevalenza media totale del 94% e piccole
oscillazioni, non abbastanza significative, che andavano dall’84 al 100% tra i vari tipi
di trapianto. L’analisi delle cariche virali dimostrava che i titoli di TTV si
concentravano intorno ad un valore medio di 4.0 log copie per ml di plasma, con
variazioni inter-gruppo che fluttuavano da 3.7 a 4.2 log. Anche in questo caso i titoli
del virus risultavano molto simili e non si evidenziavano differenze statisticamente
significative.

58
Tabella 3.1: Presenza e titolo di TTV nel plasma pre-trapianto.
*Log copie per ml di plasma ± deviazione standard
3.2 Cinetica dei titoli di TTV nel breve periodo post-
trapianto
La prima analisi effettuata è stata quella di valutare l’andamento della carica virale di
TTV subito dopo il trapianto, in particolare nei primi 14 giorni. Come mostrato in
Figura 3.1 il titolo medio di TTV, dall’iniziale valore di circa 3.8 log copie/ml, subiva
un rapido e significativo calo nei primi sette giorni post-trapianto. Ad una settimana
dall’operazione infatti, il titolo virale era di 2.7 log con una diminuzione di circa 1,5
logaritmi rispetto al valore basale. Poi, la carica di TTV mostrava un modesto

59
incremento, fino a raggiungere al quattordicesimo giorno un valore pari a 3.0 log
copie per ml.
Figura 3.1: Andamento medio della carica di TTV nei primi 14 giorni post-trapianto.
Un andamento simile era osservabile anche esaminando le singole tipologie di
trapianto. Come mostrato nel grafico in Figura 3.2, per ogni gruppo considerato
l’andamento era riconducibile a quello medio totale mostrato nella figura precedente.
Tuttavia, alcune piccole differenze risultavano evidenti: nel caso del trapianto
simultaneo di rene e pancreas o nel gruppo di individui che hanno ricevuto l’organo
da persona vivente, il calo del titolo di TTV era più ampio. Per altre tipologie, tale
diminuzione era più contenuta come nel caso dei trapiantati di rene, dove al
quattordicesimo giorno era presente un calo di soli 0.3 log. Tali differenze comunque
non si rivelavano statisticamente significative.
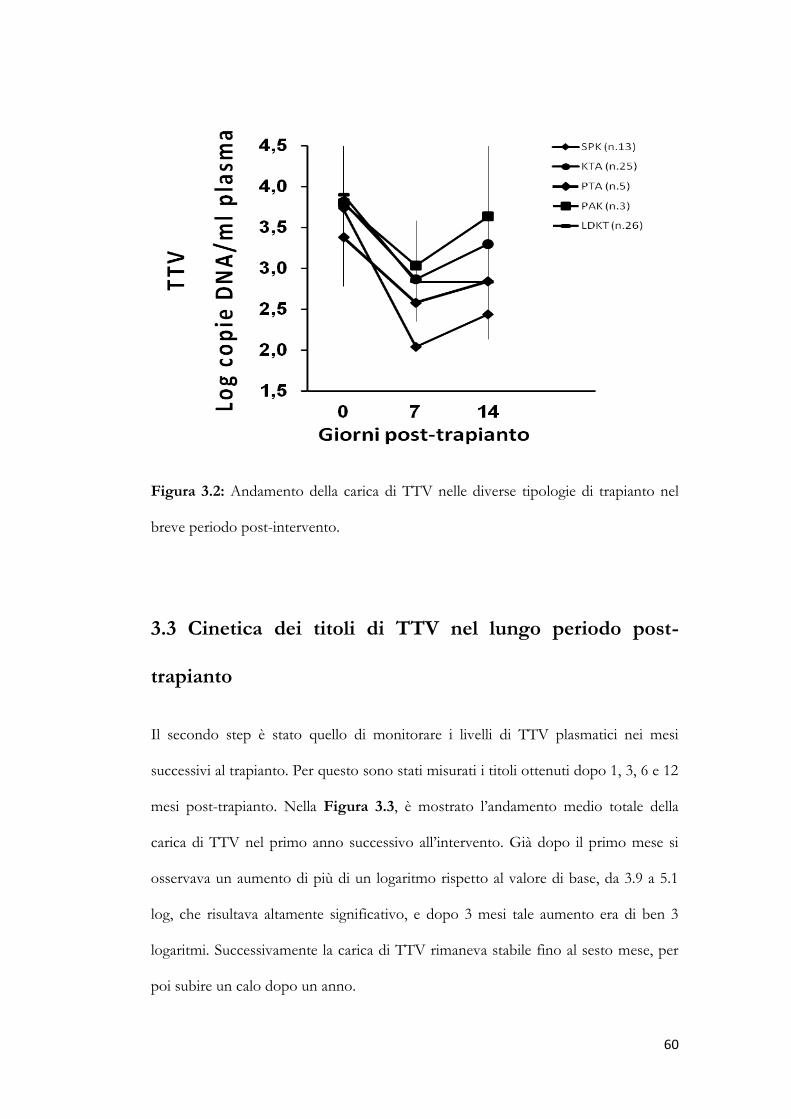
60
Figura 3.2: Andamento della carica di TTV nelle diverse tipologie di trapianto nel
breve periodo post-intervento.
3.3 Cinetica dei titoli di TTV nel lungo periodo post-
trapianto
Il secondo step è stato quello di monitorare i livelli di TTV plasmatici nei mesi
successivi al trapianto. Per questo sono stati misurati i titoli ottenuti dopo 1, 3, 6 e 12
mesi post-trapianto. Nella Figura 3.3, è mostrato l’andamento medio totale della
carica di TTV nel primo anno successivo all’intervento. Già dopo il primo mese si
osservava un aumento di più di un logaritmo rispetto al valore di base, da 3.9 a 5.1
log, che risultava altamente significativo, e dopo 3 mesi tale aumento era di ben 3
logaritmi. Successivamente la carica di TTV rimaneva stabile fino al sesto mese, per
poi subire un calo dopo un anno.
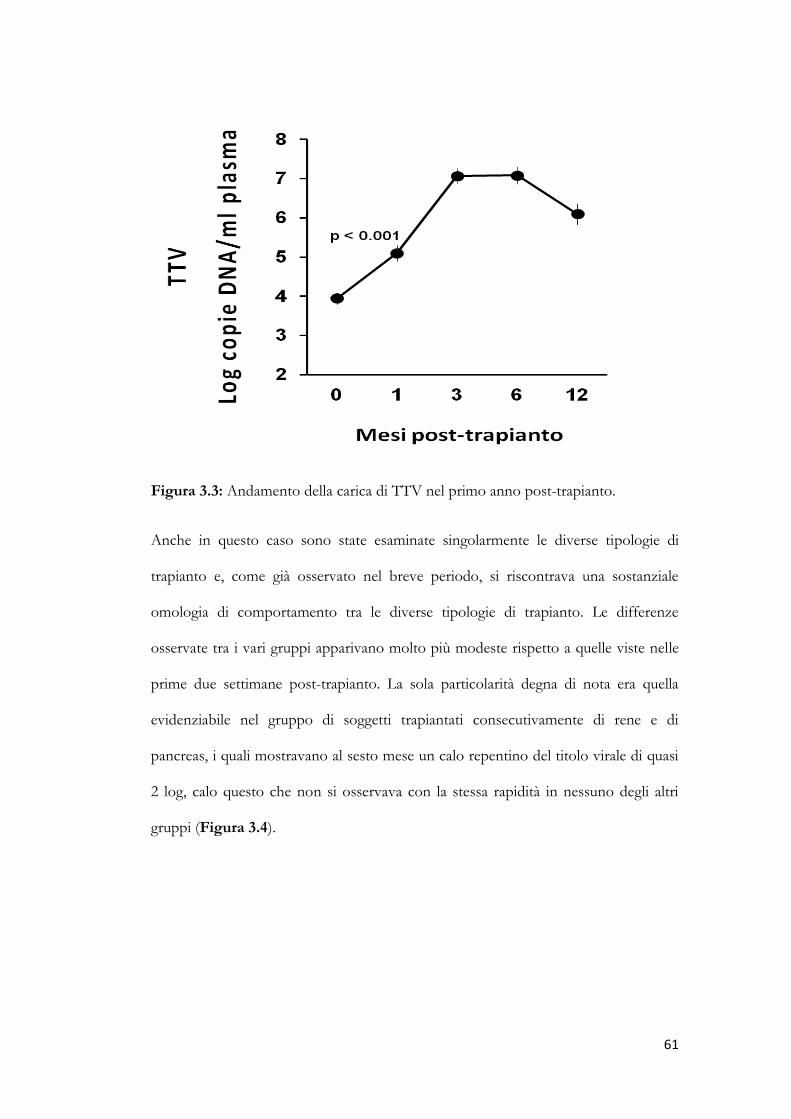
61
Figura 3.3: Andamento della carica di TTV nel primo anno post-trapianto.
Anche in questo caso sono state esaminate singolarmente le diverse tipologie di
trapianto e, come già osservato nel breve periodo, si riscontrava una sostanziale
omologia di comportamento tra le diverse tipologie di trapianto. Le differenze
osservate tra i vari gruppi apparivano molto più modeste rispetto a quelle viste nelle
prime due settimane post-trapianto. La sola particolarità degna di nota era quella
evidenziabile nel gruppo di soggetti trapiantati consecutivamente di rene e di
pancreas, i quali mostravano al sesto mese un calo repentino del titolo virale di quasi
2 log, calo questo che non si osservava con la stessa rapidità in nessuno degli altri
gruppi (Figura 3.4).

62
Figura 3.4: Andamento della carica di TTV nelle diverse tipologie di trapianto nel
primo anno post-intervento.
3.4 Titolo di TTV e numero di organi trapiantati
I pazienti sono stati suddivisi a seconda del numero di organi trapiantati per poter
valutare se vi fosse una correlazione tra titolo di TTV e stato del sistema
immunitario. Per questo è stato eseguito un confronto tra i soggetti sottoposti a due
interventi, e quindi con un sistema immunitario sottoposto maggiormente a stress,
con quelli che ne hanno subito solo uno. Come osservato in Figura 3.5, i soggetti
che avevano subito sia il trapianto di rene che di pancreas mostravano titoli di TTV
più elevati rispetto a quelli che avevano subito la sostituzione di un solo organo.
Sebbene l’andamento generale della carica di TTV appariva assai simile, al terzo mese
post-trapianto si osservava una differenza statisticamente significativa.
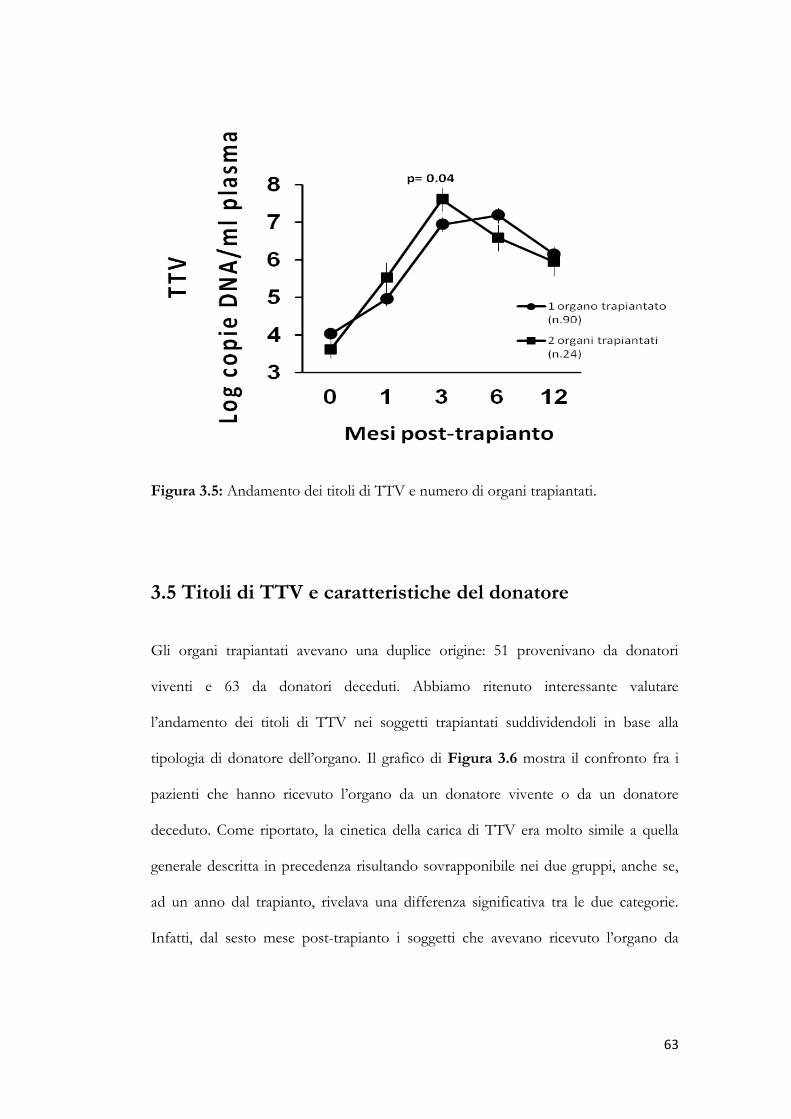
63
Figura 3.5: Andamento dei titoli di TTV e numero di organi trapiantati.
3.5 Titoli di TTV e caratteristiche del donatore
Gli organi trapiantati avevano una duplice origine: 51 provenivano da donatori
viventi e 63 da donatori deceduti. Abbiamo ritenuto interessante valutare
l’andamento dei titoli di TTV nei soggetti trapiantati suddividendoli in base alla
tipologia di donatore dell’organo. Il grafico di Figura 3.6 mostra il confronto fra i
pazienti che hanno ricevuto l’organo da un donatore vivente o da un donatore
deceduto. Come riportato, la cinetica della carica di TTV era molto simile a quella
generale descritta in precedenza risultando sovrapponibile nei due gruppi, anche se,
ad un anno dal trapianto, rivelava una differenza significativa tra le due categorie.
Infatti, dal sesto mese post-trapianto i soggetti che avevano ricevuto l’organo da

64
vivente mostravano un calo più rapido del titolo di TTV rispetto a quelli che avevano
ricevuto l’organo da cadavere.
Figura 3.6: Andamento della carica di TTV a seconda delle caratteristiche dei
donatori.
3.6 Cinetica dei titoli di TTV in relazione al tipo di
trattamento immunosoppressivo
Come già anticipato in Materiali e Metodi, la popolazione dello studio aveva subito
un trattamento immunosoppressivo con due diversi farmaci: 31 pazienti erano stati
trattati con ATG mentre agli altri 83 era stato somministrato Basiliximab. I due
farmaci possiedono una diversa potenza immunosoppressiva: infatti l’ATG ha una
maggiore capacità di agire sul sistema immunitario rispetto al Basiliximab. Per
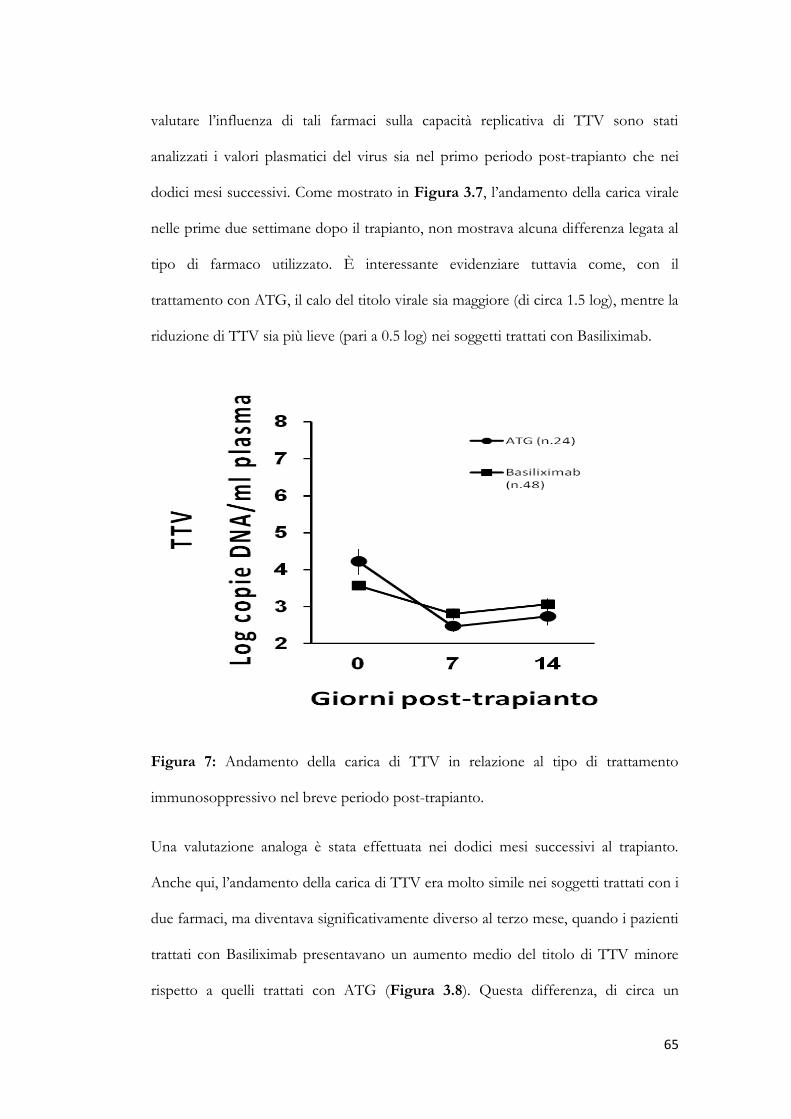
65
valutare l’influenza di tali farmaci sulla capacità replicativa di TTV sono stati
analizzati i valori plasmatici del virus sia nel primo periodo post-trapianto che nei
dodici mesi successivi. Come mostrato in Figura 3.7, l’andamento della carica virale
nelle prime due settimane dopo il trapianto, non mostrava alcuna differenza legata al
tipo di farmaco utilizzato. È interessante evidenziare tuttavia come, con il
trattamento con ATG, il calo del titolo virale sia maggiore (di circa 1.5 log), mentre la
riduzione di TTV sia più lieve (pari a 0.5 log) nei soggetti trattati con Basiliximab.
Figura 7: Andamento della carica di TTV in relazione al tipo di trattamento
immunosoppressivo nel breve periodo post-trapianto.
Una valutazione analoga è stata effettuata nei dodici mesi successivi al trapianto.
Anche qui, l’andamento della carica di TTV era molto simile nei soggetti trattati con i
due farmaci, ma diventava significativamente diverso al terzo mese, quando i pazienti
trattati con Basiliximab presentavano un aumento medio del titolo di TTV minore
rispetto a quelli trattati con ATG (Figura 3.8). Questa differenza, di circa un
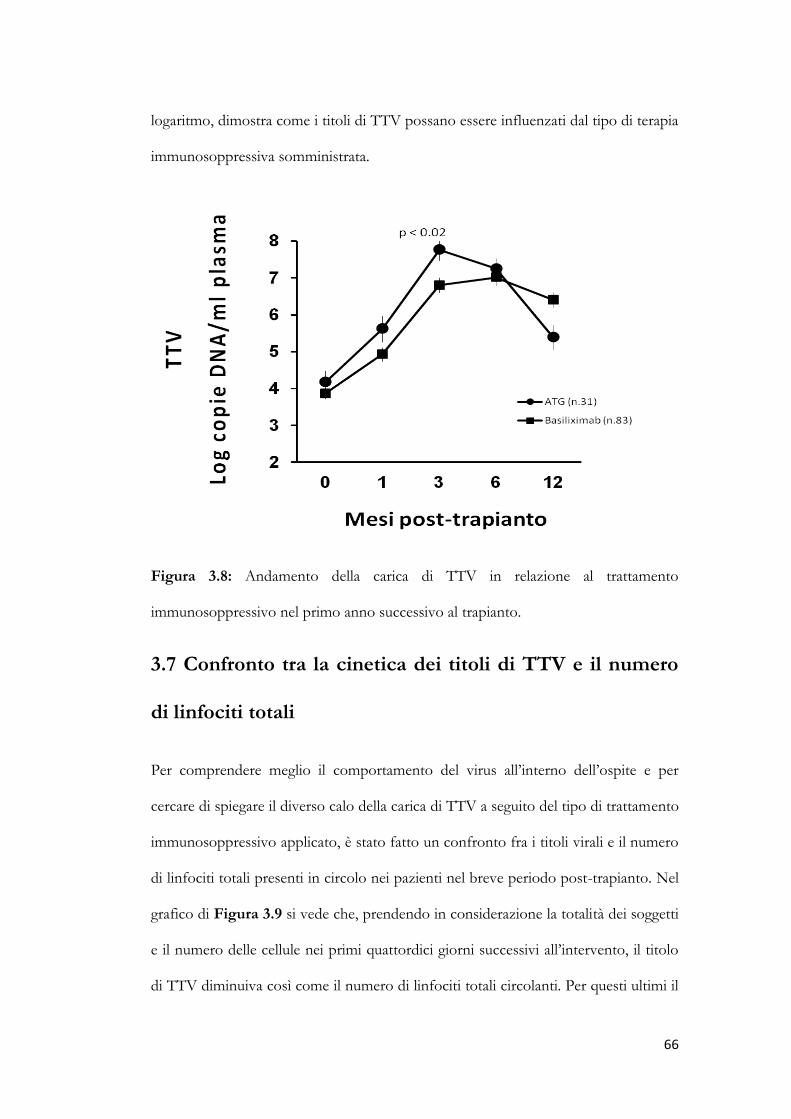
66
logaritmo, dimostra come i titoli di TTV possano essere influenzati dal tipo di terapia
immunosoppressiva somministrata.
Figura 3.8: Andamento della carica di TTV in relazione al trattamento
immunosoppressivo nel primo anno successivo al trapianto.
3.7 Confronto tra la cinetica dei titoli di TTV e il numero
di linfociti totali
Per comprendere meglio il comportamento del virus all’interno dell’ospite e per
cercare di spiegare il diverso calo della carica di TTV a seguito del tipo di trattamento
immunosoppressivo applicato, è stato fatto un confronto fra i titoli virali e il numero
di linfociti totali presenti in circolo nei pazienti nel breve periodo post-trapianto. Nel
grafico di Figura 3.9 si vede che, prendendo in considerazione la totalità dei soggetti
e il numero delle cellule nei primi quattordici giorni successivi all’intervento, il titolo
di TTV diminuiva così come il numero di linfociti totali circolanti. Per questi ultimi il
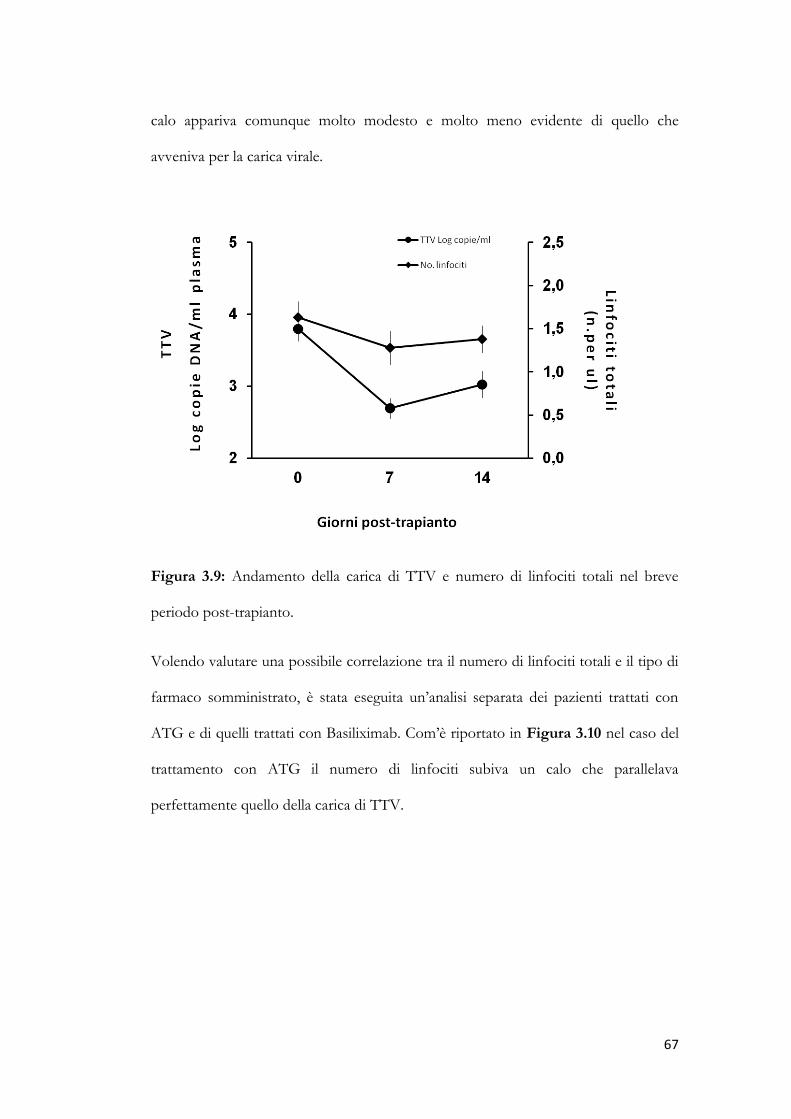
67
calo appariva comunque molto modesto e molto meno evidente di quello che
avveniva per la carica virale.
Figura 3.9: Andamento della carica di TTV e numero di linfociti totali nel breve
periodo post-trapianto.
Volendo valutare una possibile correlazione tra il numero di linfociti totali e il tipo di
farmaco somministrato, è stata eseguita un’analisi separata dei pazienti trattati con
ATG e di quelli trattati con Basiliximab. Com’è riportato in Figura 3.10 nel caso del
trattamento con ATG il numero di linfociti subiva un calo che parallelava
perfettamente quello della carica di TTV.
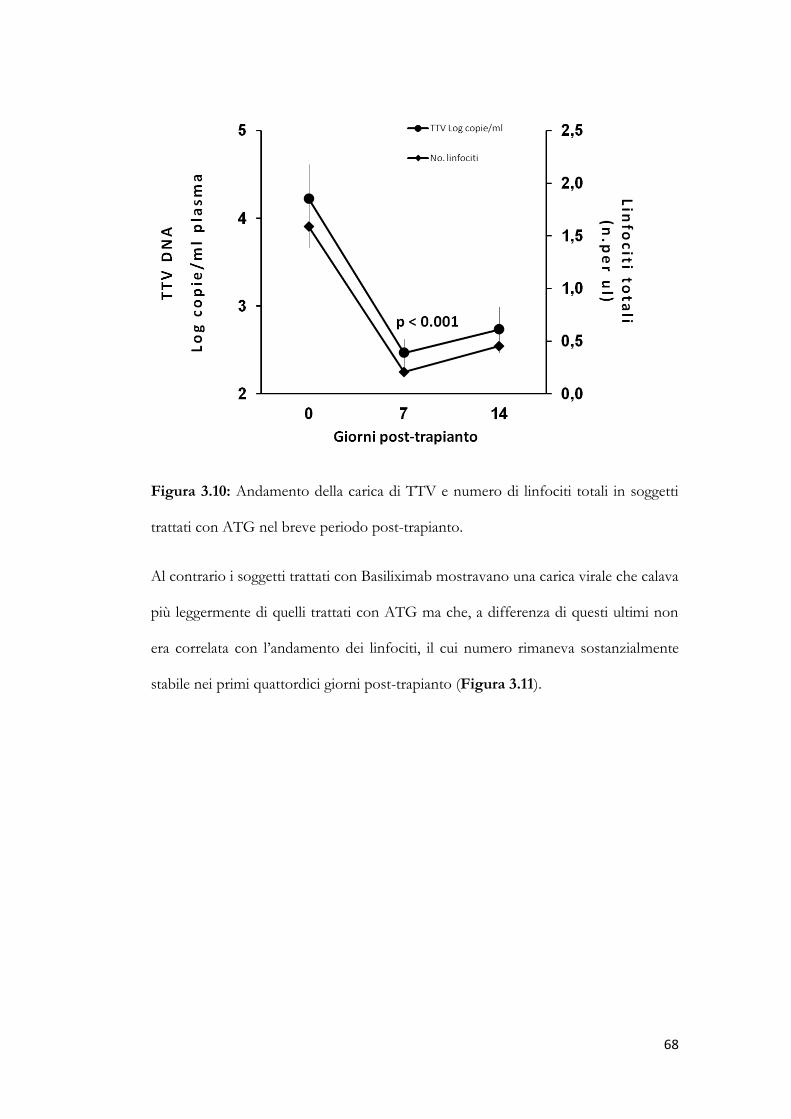
68
Figura 3.10: Andamento della carica di TTV e numero di linfociti totali in soggetti
trattati con ATG nel breve periodo post-trapianto.
Al contrario i soggetti trattati con Basiliximab mostravano una carica virale che calava
più leggermente di quelli trattati con ATG ma che, a differenza di questi ultimi non
era correlata con l’andamento dei linfociti, il cui numero rimaneva sostanzialmente
stabile nei primi quattordici giorni post-trapianto (Figura 3.11).

69
Figura 3.11: Andamento della carica di TTV e numero di linfociti in soggetti trattati
con Basiliximab nel breve periodo post-trapianto.
3.8 Andamento dei titoli di TTV in pazienti con o senza
riattivazione di CMV
Il citomegalovirus umano (CMV) è un virus appartenente alla famiglia Herpesviridae
che produce un’infezione comunemente asintomatica e ampiamente diffusa fra la
popolazione. Una volta contratto il citomegalovirus persiste nell’individuo in uno
stato di latenza la cui sede principale è costituita dalle cellule ematiche mononucleate.
In alcune situazioni si può osservare il fenomeno della riattivazione dell’infezione
latente la quale può verificarsi a tutte le età. In particolar modo può avvenire
riattivazione in seguito a malattie debilitanti, malattie con stati di immunodepressione
o in seguito a trattamenti farmacologici immunosoppressivi conseguenti a un

70
trapianto d’organo. In questi ultimi soggetti in particolare, la riattivazione di
un’infezione citomegalica latente o la trasmissione dell’infezione con l’organo del
donatore, possono essere legate a fenomeni di rigetto. In quest’analisi è stata valutata
la cinetica della carica di TTV a seguito di riattivazione o meno dell’infezione da
CMV. Come si vede in Figura 3.12, i pazienti che non avevano subito riattivazione
di CMV nei primi tre mesi post-trapianto mostravano un andamento della carica di
TTV che subiva un calo più marcato a partire dal terzo mese. I pazienti che invece
sperimentavano la riattivazione avevano una cinetica della carica di TTV
significativamente diversa e più in linea con quella mostrata in Figura 3.3.
Figura 3.12: Andamento della carica di TTV in seguito o meno a riattivazione di
CMV.

71
3.9 Studio dei genogruppi di TTV
Nel 2002 Peng e collaboratori proposero la classificazione di tutti gli isolati di TTV in
5 genogruppi principali, identificati con i numeri da 1 a 5, basandosi su una
divergenza nucleotidica minima del 50%. Prendendo in considerazione questa
classificazione, sono stati selezionati 10 pazienti che presentavano dopo il trapianto
un titolo virale molto elevato e sui campioni di questi soggetti sono state eseguite
PCR genogruppo-specifiche per tutti e cinque i genogruppi noti. La Tabella 3.2
mostra per ogni paziente il titolo e i genogruppi di TTV rilevati prima e dopo il
trapianto. Dal punto di vista esclusivamente numerico si osservava che 3 soggetti
(CM, LM e PF) presentavano dopo il trapianto un numero più elevato di genogruppi
di TTV rispetto al pre-trapianto; altri 3 soggetti (CF, MG e PC) mostravano una
riduzione della loro complessità genetica, mentre tutti gli altri rimanevano pressoché
stabili geneticamente. La tipologia dei genogruppi era variabile: a parità di numero,
alcuni soggetti perdevano alcuni genogruppi e ne acquisivano altri dopo l’intervento
(AD, CAM, PA). I genogruppi 1, 3 e 4 avevano una prevalenza maggiore rispetto agli
altri. Essi comparivano nella quasi totalità dei soggetti studiati e spesso erano
mantenuti anche dopo il trapianto. Al contrario, il genogruppo 2 non è stato
identificato in nessun campione analizzato. Questi risultati sono in linea con i dati
presenti in letteratura.
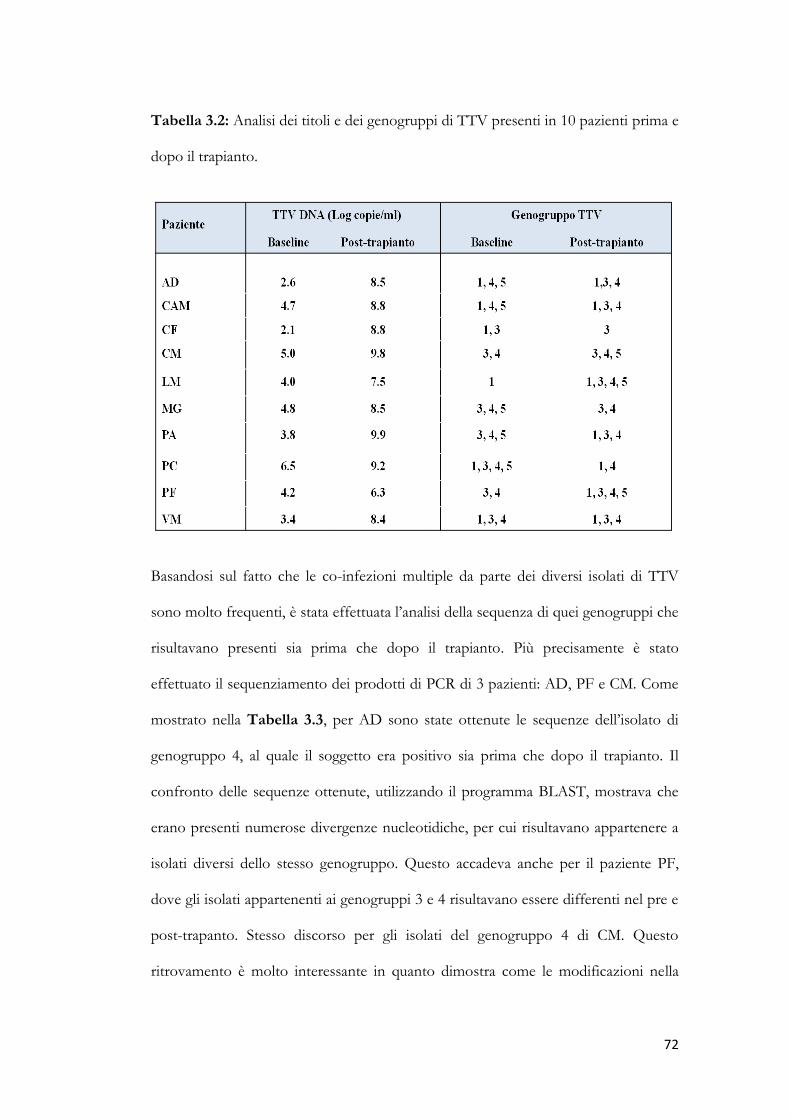
72
Tabella 3.2: Analisi dei titoli e dei genogruppi di TTV presenti in 10 pazienti prima e
dopo il trapianto.
Basandosi sul fatto che le co-infezioni multiple da parte dei diversi isolati di TTV
sono molto frequenti, è stata effettuata l’analisi della sequenza di quei genogruppi che
risultavano presenti sia prima che dopo il trapianto. Più precisamente è stato
effettuato il sequenziamento dei prodotti di PCR di 3 pazienti: AD, PF e CM. Come
mostrato nella Tabella 3.3, per AD sono state ottenute le sequenze dell’isolato di
genogruppo 4, al quale il soggetto era positivo sia prima che dopo il trapianto. Il
confronto delle sequenze ottenute, utilizzando il programma BLAST, mostrava che
erano presenti numerose divergenze nucleotidiche, per cui risultavano appartenere a
isolati diversi dello stesso genogruppo. Questo accadeva anche per il paziente PF,
dove gli isolati appartenenti ai genogruppi 3 e 4 risultavano essere differenti nel pre e
post-trapanto. Stesso discorso per gli isolati del genogruppo 4 di CM. Questo
ritrovamento è molto interessante in quanto dimostra come le modificazioni nella
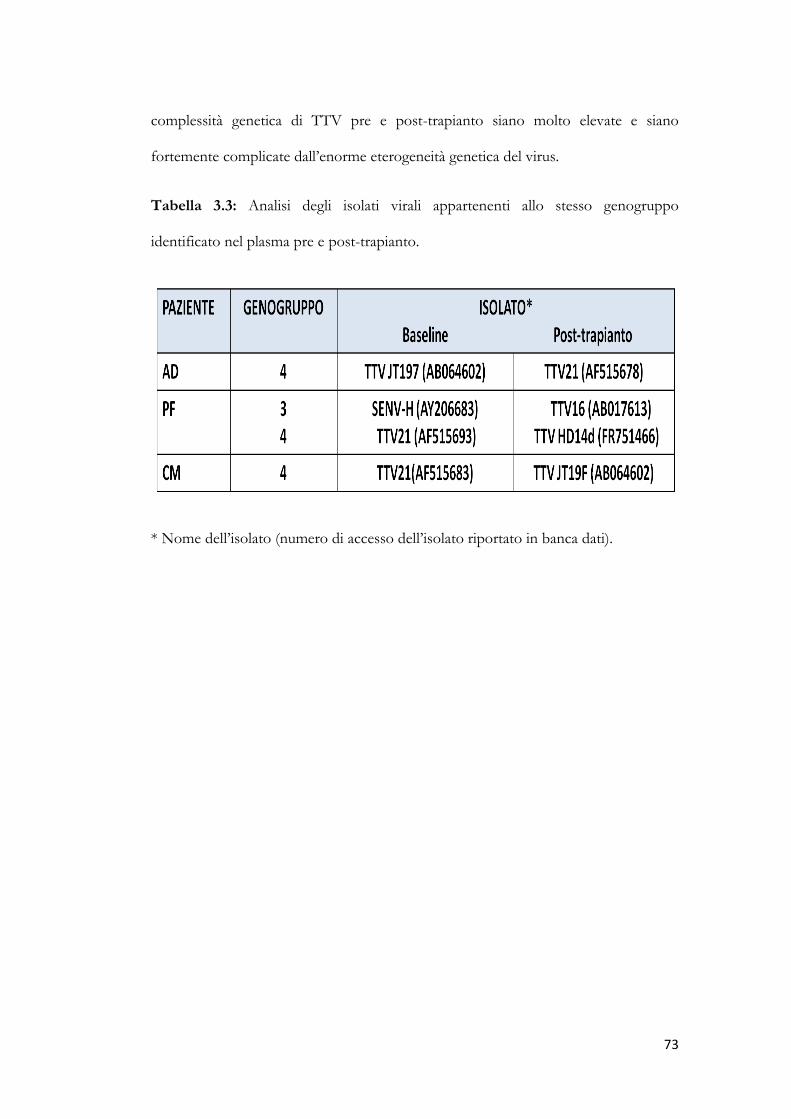
73
complessità genetica di TTV pre e post-trapianto siano molto elevate e siano
fortemente complicate dall’enorme eterogeneità genetica del virus.
Tabella 3.3: Analisi degli isolati virali appartenenti allo stesso genogruppo
identificato nel plasma pre e post-trapianto.
* Nome dell’isolato (numero di accesso dell’isolato riportato in banca dati).

74
CAPITOLO 4
DISCUSSIONE E CONCLUSIONI

75
Dal momento della sua scoperta avvenuta nel 1997 in un paziente giapponese affetto
da epatite criptogenica (Nishizawa et al., 1997), Torque Teno Virus (TTV) è stato
oggetto di numerosissimi studi per comprenderne i meccanismi di replicazione e il
ruolo patogenetico. E’ stato così dimostrato che il virus è estremamente diffuso
(Bendinelli et al., 2001) e in grado di indurre una viremia cronica nell’80% della
popolazione generale (Takayama et al., 1999b; Matsumoto et al., 1999). La sua reale
diffusione potrebbe comunque essere sottostimata, poiché le attuali metodiche usate
per determinarne l’infezione, sono in grado di evidenziare solo quelle in atto ma non
le infezioni pregresse che sono state risolte (Biagini et al., 2001; Matsubara et al.,
2001). Studi recenti hanno dimostrato un ruolo di TTV nella patogenesi di alcune
importanti malattie, quali quelle acute e/o croniche a carico del sistema respiratorio
(Maggi et al., 2003b) fra cui l’asma (Pifferi et al., 2005). Le cellule in cui il virus
sembra principalmente replicare sono quelle del sistema immunitario: intermedi
replicativi sono stati rilevati in linfociti T e B (Yu et al., 2002), nelle cellule natural
killer e nei polimorfonucleati (Maggi et al., 2001a). A conferma di ciò, l’evidenza che,
nei pazienti sottoposti a trapianto, si ha un calo significativo della carica virale dovuta
alla drastica riduzione delle cellule immuni a seguito della terapia somministrata
(Kanda et al., 1999).
Scopo di questo lavoro di tesi è stato quello di valutare l’andamento delle cariche di
TTV in pazienti sottoposti a trapianto di organo solido (rene e/o pancreas) e di
verificare se tale andamento fosse in qualche modo correlato con alcune
caratteristiche dell’organo trapiantato e con il tipo di terapia immunosoppressiva
somministrata. Nello studio la percentuale di pazienti con infezione da TTV prima
del trapianto è risultata oltre il 90%, una prevalenza estremamente elevata ma

76
perfettamente in linea con i dati riportati in letteratura. Di tutti i pazienti arruolati,
avevamo a disposizione campioni di sangue multipli ottenuti longitudinalmente nel
periodo post-trapianto: in particolare, campioni prelevati entro i primi 14 giorni dal
trapianto e campioni ottenuti nei mesi successivi fino ad un anno dall’intervento.
Come dimostrato dai dati, il titolo di TTV subiva un calo significativo nella prima
settimana post- trapianto, cominciava a risalire lentamente già nella seconda
settimana e subiva un notevole e significativo incremento a partire dal primo mese
post-trapianto. Raggiunto il picco al sesto mese, il livello ematico di TTV tendeva
successivamente a stabilizzarsi. Per trovare una spiegazione al repentino calo iniziale
della viremia di TTV, è stata effettuata una misurazione della quantità di linfociti
totali circolanti nei primi 14 giorni post-trapianto. I due schemi di trattamento, ATG
versus basiliximab, a cui erano stati sottoposti i pazienti arruolati nello studio
mostravano un diverso effetto sulle cellule periferiche circolanti. L’ATG, più potente,
determinava un calo repentino di tutti i linfociti, calo che invece non si osservava nei
pazienti a cui era stato somministrato basiliximab, che come è noto agisce inattivando
e non eliminando i linfociti T, principali responsabili delle reazioni anti-rigetto. In
accordo con quanto detto sopra, una diminuzione molto più significativa della
viremia di TTV si osservava nei pazienti trattati con ATG, mentre il blocco della
funzione dei linfociti T indotta dal basiliximab comportava una riduzione dei titoli di
TTV molto più blanda. Ciò sembra confermare l’ipotesi che TTV possa replicare in
cellule del sangue periferico, preferenzialmente in linfociti T, ma che soltanto
l’abbattimento del numero e non il semplice blocco della loro funzionalità, possa
determinare il drop della carica del virus circolante. Inoltre, il ripristino del numero
delle cellule periferiche osservato dalla seconda settimana in poi, spiega il perché del
cellulare trova il terreno più ideale per la sua replicazione.

77
L’analisi della relazione fra viremia di TTV e status di immunosoppressione del
paziente trapiantato è stata condotta analizzando i dati in vari modi. Dapprima, è
stata fatta una valutazione distinguendo i pazienti in base al numero di organi
trapiantati: la carica di TTV è risultata più elevata nel gruppo di pazienti che avevano
subito almeno due trapianti d’organo, con una differenza che rispetto alla carica
virale osservata nei pazienti mono-trapiantati risultava statisticamente significativa al
sesto mese post-intervento. Il doppio trapianto è certamente più debilitante e più
immunosoppressivo del singolo trapianto e quindi probabilmente più adatto a creare
quel particolare status di ipofunzionalità immune che potrebbe favorire una più
intensa replicazione di TTV. Ciò è in linea con quanto osservato in molti lavori circa
un aumento della carica di TTV in soggetti con sistema immunitario particolarmente
compromesso (Simmonds et al., 2002; Shang et al., 2000; Burra et al., 2008). In
seguito i pazienti trapiantati sono stati suddivisi sulla base del fatto che avessero
ricevuto l’organo da donatore vivente o da cadavere. E’ noto che il sistema
immunitario di pazienti che hanno ricevuto un organo da vivente recupera più
rapidamente la propria funzionalità rispetto a quanto avviene dopo un trapianto
d’organo da cadavere. Questo perché i tempi di attesa per il trapianto sono molto più
ristretti e ciò comporta un minor periodo di dialisi che, a lungo andare, può causare
episodi di uremia e una successiva maggiore immunosoppressione. Anche in questo
caso, l’andamento della viremia di TTV risultava diverso: più precisamente, i soggetti
con trapianto d’organo da vivente mostravano, a partire dal sesto mese dal trapianto,
un più netto calo della viremia di TTV, calo che, ad un anno, risultava statisticamente
significativo rispetto a quello osservato in chi aveva ricevuto l’organo da cadavere.
Altro aspetto preso in considerazione è stato quello di valutare fra i soggetti
trapiantati chi avesse subito o meno una riattivazione da CMV entro 3 mesi dal

78
trapianto. Come è noto, la riattivazione da CMV avviene a seguito di particolari
condizioni legate essenzialmente a disfunzionalità del sistema immunitario e quindi
distinguere chi ha riattivazione da chi non l’ha, equivale probabilmente a discriminare
pazienti con sistema immune più o meno alterato. I risultati mostravano un
andamento della carica di TTV sovrapponibile nei due gruppi di pazienti fino al terzo
mese. Successivamente i soggetti che non subivano riattivazione da CMV,
evidenziavano un aumento meno significativo di TTV nei mesi successivi rispetto a
chi invece aveva positività per CMV. Anche questo dato si sposa bene con l’idea
generale che un sistema fortemente immunocompromesso facilita l’aumento della
viremia di TTV.
Infine è stata fatta anche una valutazione della variabilità genetica di TTV in un
gruppo di campioni ottenuti da pazienti selezionati, allo scopo di valutare se
l’aumento della carica virale in seguito al trapianto potesse dipendere dall’acquisizione
di qualche particolare genogruppo del virus. I risultati ottenuti hanno dimostrato
l’estrema variabilità genetica del virus anche in questi soggetti, variabilità che subisce
fluttuazioni nel tempo sia in termini di complessità che di diversità. Tuttavia non è
stato rilevato nessuno specifico trend (acquisizione e o eliminazione di uno specifico
genogruppo, aumento o riduzione della variabilità genetica, ecc…) che potesse essere
responsabile delle fluttuazioni della viremia.
I risultati ottenuti ci consentono di affermare che la misura della viremia di TTV può
effettivamente essere utile come marcatore surrogato dello status del sistema
immunitario in pazienti sottoposti a trapianto di organo solido. Tale misura potrebbe
essere importante per definire al meglio le dosi e i tempi della terapia farmacologica
da somministrare ad ogni singolo paziente e per certificare in modo oggettivo il
ripristino di una normale funzionalità del sistema immunitario del paziente.

79
CAPITOLO 5
BIBLIOGRAFIA

80
Abraham P, John GT, Raghuraman S, Radhakrishnan S, Thomas PP, Jacob CK, Sridharan G, GB virus C/hepatitis G virus and TT virus infections among high risk renal transplant recipients in India, GB virus C/hepatitis G virus and TT virus infections among high risk renal transplant recipients in India. J Clin Virol. 2003 Sep;28(1):59-69.
Ali S, Fevery J, Peerlinck K, Verslype C, Schelstraete R, Gyselinck F, Emonds MP, Vermylen J, Yap SH, (2002), TTV infection and its relation to serum transaminases in apparently healthy blood donors and in patients with clotting disorder who have been investigated previously for hepatitis C virus and GBV-C/HGV infection in Belgium. J Med Virol 66:561-566.
Bando M, Ohno S, Oshikawa K, Takahashi M, Okamoto H and Sugiyama Y (2001). Infection of TT virus in patients with idiopathic pulmonary fibrosis. Respir Med 95: 935-942.
Barril G, Lopez-Alcorocho JM, Bajo A, et al (2000) Prevalence of TT virus in serum and peripheral mononuclear cells from a CAPD population. Perit Deal Int 20:65-68.
Bendinelli M, Pistello M, Maggi F, Fornai C, Freer G, Vatteroni M, 2001. Molecular properties, biology and clinical implications of TT virus, a recently identified widespread infectious agent of man. Clin. Microbiol. Rev, 14:98-113.
Biagini P, Gallian P, Attoui H, Cantaloube JF, de Micco P and de Lamballerie X. (1999). Determination and phylogenetic analysis of partial sequences from TT virus isolates. J Gen Virol 80:419-424.
Biagini P, Gallian P, Touinssi M, Cantaloube JF, Zapitelli JP, de Lamballerie X and de Micco P (2000). High prevalence of TT virus infection in French blood donors revealed by the use of three PCR systems. Transfusion 40:590-595.
Biagini P, Gallian P, Attoui H, Cantaloube JF, Touinssi M, de Micco P and de Lamballerie X (2001a), Comparison of systems performance for TT virus detection using PCR primer sets located in non-coding and coding regions of the viral genome. J Clin Virol 22:91-99.
Biagini P, Gallian P, Attoui H, Touinssi M, Cantaloube J, de Micco P and de Lamballerie X (2001b). Genetic analysis of full-

81
length genomes and subgenomic sequences of TT virus-like mini virus human isolates. J Gen Virol 82:379-383.
Boysen T, Christensen JK, Madsen CD, Eugen-Olsen J, Christensen LS, Moller-Sorensen H, Weber A, Ladefoged SD and Krogsgaard K (2003) Presence and significance of TT virus in Danish patients on maintenance hemodialysis. Scand J Urol Nephrol 37:259-264.
Cacoub P, Rosenthal E, Gerolami V, Hausfater P, Ghillani P, Sterkers Y, Thibault V, Khiri H, Piette JC and Halfon P (2003), Transfusion associated TT virus coinfection in patients with hepatitis C virus is associated with type II mixed cryoglobulinemia but not with B-cell non-Hodgkin lymphoma. Clin Microbiol Infect 9: 39-44.
Chan PK, Tam WH, Yeo W, Cheung JL, Zhong S, Cheng AF (2001) High carriage rate of TT virus in the cervices of pregnant women. Clin Infect Dis. 1; 32(9):1376-1377.
Charlton M, Adjei P, Poterucha J, Zein N, Moore B, Therneau T,
Krom R, Wiesner R, (1998), TT-virus infection in North American blood donors, patients with fulminant hepatic failure, and
cryptogenic cirrhosis. Hepatology 28: 839–842.
Chattopadhyay S, Rao S, Das BC, Singh NP, Kar P (2005), Prevalence of transfusion-transmitted virus infection in patients on maintenance hemodialysis from New Delhi, India, Hemodial Int. 2005 Oct;9(4):362-6.
Chen C, Wallis R: Two mechanisms for mannose-binding protein modulation of the activity of its associated serine proteases. J Biol Chem 2004, 279:26058-26065.
Christensen J, Eugen-Olsen J Sorensen M, Ullum H, Gjedde SB, Pedersen, BK, Nielsen JO and Krogsgaard K (2000) Prevalence and prognostic significance of infection with TT virus in patients infected with human immunodeficiency virus. J Infect Dis 181:1796-1799.
Davidson F, MacDonald D, Mokili JL, Prescott LE, Graham S and Simmonds P (1999), Early acquisition of TT virus (TTV) in an area endemic for TTV infection. J Infect Dis 179: 1070-1076.
Deng X, Terunuma H, Handema R, Sakamoto M, Kitamura T, Ito M, Akahane Y, (2000), Higher prevalence and viral load of TT virus in

82
saliva than in the corresponding serum: another possible transmission route and replication site of TT virus. J Med Virol. 2000 Dec; 62(4):531-7.
Desai SM, Muerhoff AS, Leary TP, Erker JC, Simons JN, Chalmers ML, Birkenmeyer LG, Pilot-Matias TJ, Mushahwar IK, Prevalence of TT virus infection in US blood donors and populations at risk for acquiring parenterally transmitted viruses, J Infect Dis. 1999 May;179(5):1242-4.
Erker JC, Leary TP, Desai SM, Chalmers ML and Mushahwar IK (1999) Analyses of TT virus full-length genomic sequences. J Gen Virol 80:1743-1750.
Fanci R, De Santis R, Zakrzewska K, et al. (2004), Presence of TT virus DNA in bone marrow cells from hematologic patients. New Microbiol 27:113-117.
Fields N, In Fields N, Kinpe DM, Howley PM, Chanok MR, Melnick JL, Monath TP, Roizman B, Straus SE, Fields Virology 3rd ed.-1996. Lippincot–Raven, Philadelphia.
Fornai C, Maggi F, Vatteroni ML, Pistello M and Bendinelli M (2001), High prevalence of TT virus (TTV) and TTV-like minivirus in cervical swabs. J Clin Microbiol 39:2022-2024.
Galibert F, Mandart E, Fitoussi F, Tiollais P, Charnay P: Nucleotide sequence of the hepatitis B virus genome (subtype ayw) cloned in E.coli. Nature 1979; 281:646–650.
Gillian P, Borland Y, Elmer M, Racal D, de Micro P, Biogenic P, Simon S, Bouchouareb D, Mourey C, Roubicek C, Touinssi M, Cantaloube JF, Dussol B, de Lamballerie X (1999), TT virus infection in French hemodialysis patients: study of prevalence and risk factors, J Clin Microbiol. 1999 Aug; 37(8):2538-42.
Garbuglia A, Iezzi T, Capobianchi MR, et al (2003) Detection of TT virus in lymph node biopsies of B-cell lymphoma and Hodgkin’s disease, and its association with EBV infection. Int J Immunopathol Pharmacol 16:109-118.

83
Gerner P, Oettinger R, Gerner W, Falbrede J and Wirth S (2000) Mother-to-infant transmission of TT virus: prevalence, extent and mechanism of vertical transmission. Pediatr Infect Dis J 19:1074-1077.
Griffiths P (1999), Time to consider the concept of a commensal virus? J Gen Virol 78: 1795-1799.
Hallett R, Clewley JP, Bobet F, McKiernan PJ and Teo CG (2000), Characterization of a highly divergent TT virus genome. J Gen Virol 81:2273-2279.
Handa A, Dickstein B, Young NS and Brown KE (2000), Prevalence of the newly described human circovirus, TTV, in United States blood donors. Transfusion 40:245-251.
Hayakawa, E., Y. Edamoto, D. Xin, H. T. Tran, Y. Iwaki, Y. Sato, T. Sata, and K. Abe. 2001. Detection of TT virus in human bile juice. Jpn. J. Infect. Dis. 54:127-128.
Hijikata M, Takahashi K and Mishiro S (1999b) Complete circular DNA genome of a TT virus variant (isolate name SANBAN) and 44 B 101 partial ORF2 sequences implicating a great degree of diversity beyond genotypes. Virology 260:17-22.
Hirata D, Kaneko N, Iwamoto M, Yoshio T, Okazaki H, Mimori A, Masuyama J and Minota S (1998) Infection with an unenveloped DNA virus (TTV) associated with non-A to G hepatitis in patients with rheumatoid arthritis. Br J Rheumatol 37:1361-1362.
Hsieh S, Wu YH, Ho YP, Tsao KC, Yeh CT and Liaw YF (1999), High prevalence of TT virus infection in healthy children and adults and in patients with liver disease in Taiwan. J Clin Microbiol 37:1829-1831.
Hu Y, Al-Moslih MI, Al Ali MT, et al (2005), Molecular detection method for all known genotypes of TT virus (TTV) and TTV-like viruses in thalassemia patients and healthy individuals. J Clin Microbiol 43:3747-3754.
Itoh K., Masaharu Takahashi, Masato Ukita, Tsutomu Nishizawa and Hiroaki Okamoto, Influence of Primers on the Detection of TT Virus DNA by Polymerase Chain Reaction, J Infect Dis. (1999) 180.

84
Itoh Y, Takahashi M, Fukuda M, Shibayama T, Ishikawa T, Tsuda F, Tanaka T, Nishizawa T, Okamoto H (2000), Visualization of TT virus particles recovered from the sera and feces of infected humans. Biochem and Biophys Res Commun 279: 718-724.
Inami T, Konomi N, Arakawa Y, Abe K. High prevalence of TT virus DNA in human saliva and semen. J Clin Microbiol. 2000 Jun; 38(6):2407-8.
Jelcic I, Hotz-Wagenblatt A, Hunziker A, Zur Hausen H and de Villiers EM (2004) Isolation of multiple TT virus genotypes from spleen biopsy tissue from a Hodgkin's disease patient: genome reorganization and diversity in the hypervariable region. J Virol 78:7498-7507.
Jones M, Kapoor A, Lukashov VV, Simmonds P, Hecht F, Delwart E (2005), New DNA viruses identified in patients with acute viral infection syndrome. J Virol 79:8230-8236.
Kakkola L, Hedman K, Vanrobaeys H, Hedman L and SoderlundVenermo M (2002) Cloning and sequencing of TT virus genotype 6 and expression of antigenic open reading frame 2 proteins. J Gen Virol 83:979-990.
Kakkola L, Kaipio N, Hokynar K, et al (2004) Genoprevalence in human tissues of TT-virus genotype 6. Arch Virol 149:1095-1106.
Kakkola L, Tommiska J, Boele LC, Miettinen S, Blom T, Kekarainen T, Qiu J, Pintel D, Hoeben RC, Hedman K, Söderlund-Venermo M (2007) Construction and biological activity of a full-length molecular clone of human Torque teno virus (TTV) genotype 6, FEBS J. 2007 Sep;274(18):4719-30. Epub 2007 Aug 22.
Kamada K, Kamahora T, Kabat P and Hino S (2004) Transcriptional regulation of TT virus: promoter and enhancer regions in the 1.2-kb noncoding region. Virology 0:341-348.
Kamahora T, Hino S and Miyata H (2000) Three spliced mRNAs of TT virus transcribed from a plasmid containing the entire genome in COS1 cells. J Virol 74:9980-9986.
Kazi A, Miyata H, Kurokawa K, Khan MA, Kamahora T, Katamine S and Hino S (2000) High frequency of postnatal transmission of TT virus in infancy. Arch Virol 145:535-540.

85
Kikuchi K, Miyakawa H, Abe K, Kako M, Katayama K, Fukushi S and Mishiro S (2000), Indirect evidence of TTV replication in bone marrow cells, but not in epatocytes, of a subacute hepatitis/aplastic anemia patient. J Med Virol 61:165-170.
Kooistra K, Zhang YH, Henriquez NV, Weiss B, Mumberg D, and Noteborn MH (2004) TT virus-derived apoptosis-inducing protein induces apoptosis preferentially in hepatocellular carcinoma-derived cell. J Gen Virol 85:1445-1450.
Leary TP, Erker JC, Chalmers ML, Desai SM and Mushahwar IK (1999) Improved detection systems for TT virus reveal high prevalence in humans, non-human primates and farm animals. J Gen Virol 80:2115-2120.
Lefrère J, Roudot-Thoraval F, Lefrere F, Kahfer A, Mariotti M, Lerable J, Thauvin M, Lefevre G, Rouger P and Girot R (1999), Natural history of the TT virus infection through follow-up of TTV DNA-positive multipletransfused patients. Blood 95:347-351.
Lisitsyn N and Wigler M (1993) Cloning the differences between two complex genomes. Science 259:946-951.
Lopez-Alcorocho JM, Mariscal LF, de Lucas S, Rodriguez-Inigo E, Casqueiro M, Castillo I, Bartolome J, Herrero M, Manzano ML, Pardo M and Carreno V (2000) Presence of TTV DNA in serum, liver and peripheral blood mononuclear cells from patients with chronic hepatitis. J Viral Hepat 7:440-447.
Luo K, Liang W, He H, et al (2000) Experimental infection of nonenveloped DNA virus (TTV) in rhesus monkey. J Med Virol 61:159-164.
Maggi F, Fornai C, Morrica A, Casula F, Vatteroni ML, Marchi S, Ciccorossi P, Riente L, Pistello M and Bendinelli M (1999), High prevalence of TT virus viremia in italian patients, regardless of age, clinical diagnosis, and previous interferon treatment. J Infect Dis 180:838-842.
Maggi F, Fornai C, Zaccaro L, Morrica A, Vatteroni ML, Isola P, Marchi S, Ricchiuti A, Pistello M and Bendinelli M (2001a), TT virus (TTV) loads associated with different peripheral blood cell types and evidence for TTV replication in activated mononuclear cells. J Med Virol 64:19019-4.

86
Maggi F, Pistello M, Vatteroni M, Presciuttini S, Marchi S, Isola P, Fornai C, Fagnani S, Andreoli E, Antonelli G and Bendinelli M (2001b) Dynamics of persistent TT virus infection, as determined in patients treated with alpha interferon for concomitant hepatitis C virus infection. J Virol 75:11999-2004.
Maggi F., Pistello M., Vatteroni M. L., Presciuttini S., Marchi S., Isola P., Fornai C., Fagnani S., Andreoli E., Antonelli G., Bendinelli M. 2001c. Dynamics of persistent TT virus infection, as determined in patients treated with alpha interferon for concomitant hepatitis C virus infection. Journal of Virology 75, 11999-12004.
Maggi F, Pifferi M, Fornai C, Andreoli E, Tempestini E, Vatteroni M, Presciuttini S, Marchi S, Pietrobelli A, Boner A, Pistello M and Bendinelli M. (2003a) TT virus in the nasal secretions of children with acute respiratory diseases: relations to viremia and disease severity. J Virol 77:2418-2425.
Maggi F, Pifferi M, Tempestini E, Fornai C, Lanini L, Andreoli E, Vatteroni M, Presciuttini S, Pietrobelli A, Boner A, Pistello M and Bendinelli M (2003b), TT virus loads and lymphocyte subpopulations in children with acute respiratory diseases. J Virol 77:9081-9083.
Maggi F, Tempestini E, Lanini L, Andreoli E, Fornai C, Giannecchini S, Vatteroni M, Pistello M, Marchi S, Ciccorossi P, Specter S and Bendinelli M. (2005) Blood levels of TT virus following immune stimulation with influenza or hepatitis B vaccine. J Med Virol 75:358-365.
Maggi F, Andreoli E, Lanini L, Meschi S, Rocchi J, Fornai C, Vatteroni ML, Pistello M, Bendinelli M (2006), Rapid increase in total Torquetenovirus (TTV) plasma viremia load reveals an apparently transient superinfection by a TTV of a novel group genotype. J Clin Microbiol 44: 2571-2574.
Manni F, Rotola A, Caselli E, Bertorelle G and Di Luca D (2002) Detecting recombination in TT virus: a phylogenetic approach. J Mol Evol 55:563-572.
Mariscal L, Lopez-Alcorocho JM, Rodriguez-Inigo E, Ortiz-Movilla N, de Lucas S, Bartolome J and Carreno V (2002), TT virus replicates in stimulated but not in non stimulated peripheral blood mononuclear cells. Virology 301:121-129.

87
Matsubara H, Michitaka K, Horiike N, Yano M, Akbar SM, Torisu M and Onji M (2000) Existence of TT virus DNA in extracellular body fluids from normal healthy Japanese subjects. Intervirology 43:16-19.
Matsubara H, Michitaka K, Horiike N, Kihana T, Yano M, Mori T and Onji M (2001) Existence of TT virus DNA and TTV-like mini virus DNA in infant cord blood: mother-to-neonatal transmission. Hepatol Res 21:280-287.
Matsumoto A, Yeo AE, Shih JW, Tanaka E, Kiyosawa K and Alter HJ (1999), Transfusion-associated TT virus infection and its relationship to liver disease. Hepatology 30:283-288.
Michitaka K, Horiike N, Matsubara H, Masumoto T, Toshino A, Oka A, Ohoka H, Yokoyama M, Onji M, TT virus infection among renal transplant recipients, Hepatol Res. 2000 Aug;18(2):122-131.
Miyamoto M, Takahashi H, Sakata I, et al (2000) Hepatitisassociated aplastic anemia and transfusion-trasmitted virus infection. Intern Med 39:1068-1070.
Miyata H, Tsunoda H, Kazi A, Yamada A, Khan MA, Murakami J, Kamahora T, Shiraki K and Hino S (1999), Identification of a novel GC-rich 113- nucleotide region to complete the circular, singlestranded DNA genome of TT virus, the first human circovirus. J Virol 73:3582-3586.
Morrica A, Maggi F, Vatteroni ML, Fornai C, Pistello M, Ciccorossi P, Grassi E, Gennazzani A and Bendinelli M (2000) TT virus: evidence for transplacental transmission. J Infect Dis 181:803-804.
Muljono DH, Nishizawa T, Tsuda F, Takahashi M and Okamoto H (2001) Molecular epidemiology of TT virus (TTV) and characterization of two novel TTV genotypes in Indonesia. Arch Virol 146:1249-1266.
Mushahwar IK, Erker JC, Muerhoff AS, Leary TP, Simons JN, Birkenmeyer LG, Chalmers, ML, Pilot-Matias TJ, Desai SM (1999), Molecular and biophysical characterization of TTV virus: evidence for a new virus family infecting humans. Proc Natl Acad Sci 96: 3177–3182.
Mushahwar IK (2000) Recently discovered blood-borne viruses: are they hepatitis viruses or merely endosymbionts? J Med Virol 62:399-404.

88
Nakagawa N, Ikoma J, Ishihara T, Yasui-Kawamura N, Fujita N, Iwasa M, Kaito M, Watanabe S, Adachi Y. Biliary excretion of TT virus (TTV). J Med Virol. 2000 Aug; 61(4):462-7.
Niel C, de Oliveira JM, Ross RS, Gomes S A, Roggendorf M and Viazov S (1999) High prevalence of TT virus infection in Brazilian blood donors. J Med Virol 57:259-263.
Niel C, Saback FL and Lampe E (2000) Coinfection with multiple TT virus strains belonging to different genotypes is a common event in healthy Brazilian adults. J Clin Microbiol 38:1926-1930.
Niel C and Lampe E (2001) High detection rates of TTV-like mini virus sequences in sera from Brazilian blood donors. J Med Virol 65:199-205.
Nielsen J, Vincent IE, Bøtner A, Ladekaer-Mikkelsen AS, Allan G, Summerfield A, McCullough KC (2003) Association of lymphopenia with porcine circovirus type 2 induced postweaning multisystemic wasting syndrome (PMWS), Vet Immunol Immunopathol. 2003 May 12;92 (3-4):97-111.
Ninomiya M, Nishizawa T, Tkahashi M, et al. (2007a) Identification and genomic characterization of a novel human torque teno virus of 3.2 kb. JGen Virol 57:259-263.
Ninomiya M, Takahashi M, Shimosegawa T, Okamoto H (2007b) Analysis of the entire genomes of fifteen torque teno midi virus variants classifiable into a third group of genus Anellovirus. Arch Virol 152(11):1961-1975.
Ninomiya M, Takahashi M, Nishizawa T, et al (2008) Development of PCR assays with nested primers specific for differential detection of three human anelloviruses and early acquisition of dual or triple infection during infancy. J Clin Microbiol 46:507-514.
Nishizawa T, Okamoto H, Konishi K, Yoshizawa H, Miyakawa Y, Mayumi M (1997), A novel DNA virus (TTV) associated with elevated transaminase levels in posttrasfusion hepatitis of unknow etiology. Biochem Biophis Res Commun 241: 92-97.
Nishizawa T, Okamoto H, Tsuda F, Aikawa T, Sugai Y, Konishi K, Akahane Y, Ukita M, Tanaka T, Miyakawa Y and Mayumi M (1999), Quasispecies of TT virus (TTV) with sequence divergence in

89
hypervariable regions of the capsid protein in chronic TTV infection. J Virol 73:9604-9608.
Oguchi T, Tanaka E, Orii K, Kobayashi M, Hora K and Kiyosawa K (1999) Transmission of and liver injury by TT virus in patients on maintenance hemodialysis. J Gastroenterol 34:234-240.
Ohbayashi H, Tanaka Y, Ohoka S. et al (2001) TT virus is shown in the liver by in situ hybridization with a PCR-generated probe from the serum TTV-DNA. J Gastroenterol Hepatol 16:424-428.
Okamoto H, Okada S, Sugiyama Y, Yotsumoto S, Tanaka T, Yoshizawa H, Tsuda F, Miyakawa Y, Mayumi M: The 5’-terminal sequence of the hepatitis C virus genome. Jpn J Exp Med 1990; 60:167–177.
Okamoto H, Nishizawa T, Kato N, Ukita M, Ikeda K, Iizuka H, Miyakawa Y, Mayumi M (1998), Molecular cloning and characterization of a novel DNA virus (TTV) associated with post-transfusion hepatitis of unknow etiology. Hepatol Res 10: 1-16.
Okamoto H, Kato N, Iizuka H, Tsuda F, Miyakawa Y, Mayumi M: Distinct genotypes of a nonenveloped DNA virus associated with post-transfusion non-A to G hepatitis (TT virus) in plasma and peripheral blood mononuclear cells. J Med Virol 1999; 57:252–258.
Okamoto H, Nishizawa T, Ukita M (1999b) A novel unenveloped DNA virus (TT virus) associated with acute and chronic non-A to G hepatitis. Intervirology 42:196-204.
Okamoto H, Takahashi M, Nishizawa T, et al. (2000), Replicative forms of TT virus DNA in bone marrow cells. Biochem Biophys Res Commun 270: 657–662.
Okamoto H, Nishizawa T, Tawara A, Takahashi M, Kishimoto J, Sai T and Sugai Y (2000b) TT virus mRNAs detected in the bone marrow cells from an infected individual. Biochem Biophys Res Commun 279:700-707.
Okamoto H, Takahashi M, Kato N, Fukuda M, Tawara A, Fukuda S, Tanaka T, Miyakawa Y and Mayumi M (2000c) Sequestration of TT virus of restricted genotypes in peripheral blood mononuclear cells. J Virol 74:10236-10239.

90
Okamoto H, Takahashi M, Nishizawa T, Tawara A, Sugai Y, Sai T, Tanaka T and Tsuda F (2000d) Replicative forms of TT virus DNA in bone marrow cells. Biochem Biophys Res Commun 279:700-707.
Okamoto H, Ukita M, Nishizawa T, et al (2000e) Circular doublestranded forms of TT virus DNA in the liver. J Virol 74:5161-5167.
Okamoto H, Nishizawa T, Takahashi M, Asabe S, Tsuda F and Yoshikawa A (2001) Heterogeneous distribution of TT virus of distinct genotypes in multiple tissues from infected humans. Virology 88:358-368.
Okamoto H. (2009) History of discoveries and pathogenicity of TT viruses. Curr Top Microbiol Immunol 331:1-20. Review.
Okamoto H and Mayumi M (2001) TT virus: virological and genomic characteristics and disease associations. J Gastroenterol 36:519-529.
Okamura A, Yoshioka M, Kubota M, Kikuta H, Ishiko H and Kobayashi K (1999) Detection of a novel DNA virus (TTV) sequence in peripheral blood mononuclear cells. J Med Virol 58:174-177.
Ott C, Duret L, Chemin I, Trepo C, Mandrand B and Komurian Pradel F (2000), Use of TT virus ORF-1 recombinant protein to detect anti-TT virus antibodies in human sera. J Gen Virol. 81:29492958.
Peng Y, Nishizawa T, Takahashi M, Ishikawa T, Yoshikawa A and Okamoto H (2002) Analysis of the entire genomes of thirteen TT virus variants classifiable into the fourth and fifth genetic groups, isolated from viremic infants. Arch Virol 147:21-41.
Peters M, Jackson DC, Crabb BS and Browning GF (2002). Chicken anemia virus VP2 is a novel dual specificity protein phosphatase. J Biol Chem 277:39566-39573.
Pifferi M, Maggi F, Caramella D, et al (2006), High torquetenovirus loads are correlated with bronchiectasis and peripheral airflow limitation in children. Pediatr Infect Dis J 25:804-808.
Pifferi M, Maggi F, Andreoli E, et al (2005) Associations between nasal torquetenovirus load and spirometric indices in children with asthma. J Infect Dis 192:1141-1148.

91
Pistello M, Morrica A, Maggi F, Vatteroni ML, Freer G, Fornai C, Casula F, Marchi S, Ciccorossi P, Rovero P and Bendinelli M (2001), TT virus levels in the plasma of infected individuals with different hepatic and extrahepatic pathology. J Med Virol 63:189-195.
Pollicino T, Raffa G, Squadrito G, Costantino L, Cacciola I, Brancatelli S, Alafaci C, Florio MG and Raimondo G (2002) TT virus has ubiquitous diffusion in human body tissues: analyses of paired serum and tissue samples. J Viral Hephat 10:95-102.
Prescott LE and Simmonds P (1998), Global distribution of transfusion-transmitted virus. N Engl J Med 339:776-777.
Prescott LE, MacDonald DM, Davidson F, Mokili J, Pritchard DI, Arnot DE, Riley EM, Greenwood BM, Hamid S, Saeed AA, McClure MO, Smith DB, Simmonds P, Sequence diversity of TT virus in geographically dispersed human populations, J Gen Virol. 1999 Jul; 80 (Pt 7):1751-8.
Qiu J, Kakkola L, Cheng F, et al. Human circovirus TT virus genotype 6 expresses six proteins following transfection of a full-length clone. J Virol 2005; 79:6505–6510.
Rivanera D, Lozzi MA, Idili C, Lilli D, Prevalence of TT virus infection in Italian dialysis patients, Pathol Biol (Paris). 2009 Feb; 57(1):97-100. doi: 10.1016/j.patbio.2008.07.014. Epub 2008 Oct 7.
Rodriguez-Inigo E, Casqueiro M, Bartolome J, Ortiz-Movilla N, Lopez-Alcorocho JM, Herrero M, Manzarbeitia F, Oliva H and Carreno V (2000) Detection of TT virus DNA in liver biopsies by in situ hybridization. Am J Pathol 156:1227-1234.
Romeo R, Hegerich P, Emerson SU, Colombo M, Purcell RH, Bukh J (2000) High prevalence of TT virus (TTV) in naive chimpanzees and in hepatitis C virus-infected humans: frequent mixed infections and identification of new TTV genotypes in chimpanzees J Gen Virol. 2000 Apr; 81(Pt 4):1001-7.
Saback F, Gomes SA and Niel C (2002) High frequency of mixed TT virus infections in healthy adults and children detected by a simplified heteroduplex mobility assay. J Virol Methods 101:117-125.
Shibata M, Wang RY, Yoshiba M, Shih JW, Alter HJ and Mitamura K (2001) The presence of a newly identified infectious agent (SEN virus)

92
in patients with liver diseases and in blood donors in Japan. J Infect Dis 184:400-404.
Shibayama T, Masuda G, Ajisawa A, Takahashi M, Nishizawa T, Tsuda F and Okamoto H (2001) Inverse relationship between the titre of TT virus DNA and the CD4 cell count in patients infected with HIV. Aids 15:563-570.
Shoya Y, Kobayashi T, Koda T, Ikuta K, Kakinuma M and Kishi M (1998) Two proline-rich nuclear localization signals in the aminoand carboxyl-terminal regions of the Borna disease virus phosphoprotein. J Virol 72:9755-9762.
Simmond P (2002), TT virus infection: a novel virus-host relationship. J Med Microbiol 51: 455-458.
Suzuki T, Suzuki R, Li J, Hijkata M, Matsuda M, Li TC, Matsuura Y, Mishiro S and Miyamura T (2004) Identification of Basal Promoter and Enhancer elements in a untraslated region of the TT virus genome. J Virol 78:10820-10824.
Szládek G, Juhász A, Asztalos L, Szöke K, Murvai M, Szarka K, Veress G, Gergely L, Kónya J, Persisting TT virus (TTV) genogroup 1 variants in renal transplant recipients, Arch Virol. 2003 May; 148(5):841-51.
Tajiri H, Tanaka T, Sawada A, Etani Y, Kozaiwa K, Mushiake S, Mishiro S. Three cases with TT virus infection and idiopathic neonatal hepatitis. Intervirology. 2001; 44(6):364-9.
Takahashi K, Ohta Y and Mishiro S (1998) Partial -2, 4 kb sequences of TT virus (TTV) genome from eight Japanese isolates: diagnostic and phylogenetic implication. Hepatol Res 12:111-120.
Takahashi M, Asabe S, Gotanda Y, Kishimoto J, Tsuda F, Okamoto H. TT virus is distributed in various leukocyte subpopulations at distinct levels, with the highest viral load in granulocytes. Biochem Biophys Res Commun 2002; 290: 242–248.
Takayama S, Yamazaki S, Matsuo S and Sugii S (1999), Multiple infection of TT virus (TTV) with different genotypes in Japanese hemophiliacs. Biochem Biophys Res Commun 256:208-211.
Tanaka Y, Mizokami M, Orito E, Ohno T, Nakano T, Kato T, Kato H, Mukaide M, Park YM, Kim BS, Ueda R New genotypes of

93
TT virus (TTV) and a genotyping assay based on restriction fragment length polymorphism, FEBS Lett. 1998 Oct 23; 437(3):201-6.
Tanaka Y, Mizokami M, Orito E, Nakano T, Kato T, Ding X, Ohno T, Ueda R, Sonoda S,Tajima K, Miura T, Hayami M (1999) A new genotype of TT virus (TTV) infection among Colombian native Indians. J Med Virol 57:264–268.
Tanaka Y, Mizokami M, Orito E, Ohno T, Nakano T, Kato T, Iida S and Ueda R (2000a) Lack of integrated TT virus (TTV) genomes in cellular DNA in infected human hematopoietic cells. Leuk Lymphoma 38:411-417.
Tanaka Y, Orito E, Ohno T, Nakano T, Hayashi K, Kato T, Mukaide M, Iida S and Mizokami M (2000b) Identification of a novel 23kDa protein encoded by putative open reading frame 2 of TT virus (TTV) genotype 1 different from the other genotypes. Arch Virol 145:1385-1398.
Tawara A, Akahane Y, Takahashi M, Nishizawa T, Ishikawa T, Okamoto H (2000), Transmission of human TT virus of genotype 1a to chimpanzees with fecal supernatant or serum from patients with acute TTV infection. Biochem Biophys Res Commun 278:470-476.
Todd D, McNulty MS, Adair BM and AllanGM (2001) Animal circoviruses. Adv Virus Res 57:1-70.
Tokita H, Murai S, Kamitsukasa H, Yagura M, Harada H, Hebisawa A, Takahashi M, Okamoto H (2001a), Influence of TT virus on the histopathological features of nonalcoholic fatty liver disease. Hepatol Res 19:197-211.
Tomasiewicz K, Modrzewska R, Lyczak A, Krawczuk G (2005), TT virus infection and pancreatic cancer: relationship or accidental coexistence, World J Gastroenterol. 2005 May 14; 11(18):2847-9.
Tsuda F, Okamoto H, Ukita M, Tanaka T, Akahane Y, Konishi K, Yoshizawa H, Miyakawa Y, Mayumi M (1999), Determination of antibodies to TT virus (TTV) and application to blood donors and patients with post-transfusion non-A to G hepatitis in Japan. J Virol Methods 77:199-206.

94
Tsuda F, Takahashi M, Nishizawa T, Akahane Y, Konishi K, Yoshizawa H and Okamoto H (2001) IgM class antibodies to TT virus (TTV) in patients with acute TTV infection. Hepatol Res 19:1-11.
Ukita M, Okamoto H, Nishizawa T, Tawara A, Takahashi M, Iizuka H, Miyakawa Y and Mayumi M (2000) The entire nucleotide sequences of two distinct TT virus (TTV) isolates (TJN01 and TJN02) remotely related to the original TTV isolates. Arch Virol 145:1543-1559.
Ukita M, Okamoto H, Kato N, et al (1999), Excretion into bile of a novel unenveloped DNA virus (TT virus) associated with acute and chronic non-A-G hepatitis. J Infect Dis 179:1245-1248.
Ukita M, Okamoto H, Nishizawa T, Tawara A, Takahashi M, Iizuka H, Miyakawa Y and Mayumi M (2000) The entire nucleotide sequences of two distinct TT virus (TTV) isolates (TJN01 and TJN02) remotely related to the original TTV isolates. Arch Virol 145:1543-1559.
Umemura T, Alter HJ, Tanaka E, Yeo AE, Shih JW, Orii K, Matsumoto A, Yoshizawa K and Kiyosawa K (2001a) Association between SEN virus infection and hepatitis C in Japan. J Infect Dis 184:1246-51.
Umemura K, Saka M, Nakagawa T, Kawasaki N, Thiel S, Jensenius JC, Kawasaki T, L-MBP is expressed in epithelial cells of mouse small intestine. J Immunol 2002, 169:6945-6950.
Vasconcelos H, Cataldo M and Niel C (2002) Mixed infections of adults and children with multiple TTV-like mini virus isolates. J Med Virol 68:291-298.
Verschoor EJ, Langenhuijzen S and Heeney JL (1999) TT viruses (TTV) of non-human primates and their relationship to the human TTV genotypes. J Gen Virol 80:2491-2499.
Vihinene-Ranta M, Kakkola L, Kalela A, Vilja P and Vuento M (1997) Characterization of a nuclear localization signal of canine parvovirus capsid proteins. Eur J Biochem 250:389-394.
White PA, Li Z, Zhai X, Marinos G and Rawlinson WD (2000) Mixed viral infection identified using heteroduplex mobility analysis (HMA). Virology 271:382-389.

95
Worobey M (2000) Extensive homologous recombination among widely divergent TT viruses. J Virol 74:7666-7670.
Yokoyama H, Yasuda J, Okamoto H and Iwakura Y (2002), Pathological changes of renal epithelial cells in mice transgenic for the TT virus ORF1 gene. J Gen Virol 83:141-150.
Yokosuka O, Ikeuchi T, Kanda T, Kawai S, Imazeki F, Saisho H, Mazzalli M, Filho GA, Nishimura NF, Soares EC, The prevalence of TT virus infection in renal transplant recipients in Brazil, Transplantation. 2000 Oct 27; 70(8):1194-7.
Yokozaki S, Fukuda Y, Nakano I and Katano Y (1999), TT virus: a mother-to-child transmitted rather than bloodborne virus. Blood 93:3569-3570.
Yu Q, Shiramizu B, Nerurkar VR, Hu N, Shikuma CM, Melish ME, Cascio K, Imrie A, Lu Y and Yanagihara R (2002), TT virus: preferential distribution in CD19(+) peripheral blood mononuclear cells and lack of viral integration. J Med Virol 66:276-284.
Yuki N., Kato M., Masuzawa M., Ishida H., Inoue T., Tabata T., Matsushita Y., Kishimoto H., Sasaki Y., Hayashi N., Hori M (1999), Clinical implications of coinfection with a novel DNA virus (TTV) in hepatitis C virus carriers on maintenance hemodialysis. Journal of Medical Virology 59, 431-436.
Zein NN, Arslan M, Li H, et al (1999), Clinical significance of TT virus infection in patients with chronic hepatitis C. Am J Gastroenterol 94:3020-3027.
Zhong S, Yeo W, Tang MW, Lin XR, Mo F, Ho WM, Hui P, Johnson P J (2001), Gross elevation of TT virus genome load in the peripheral blood mononuclear cells of cancer patients. Ann N Y Acad Sci 945:84-92.
Zhong S, Yeo W, Tang M, Liu C, Lin XR, Ho WM, Hui P and Johnson PJ (2002), Frequent detection of the replicative form of TT virus DNA in peripheral blood mononuclear cells and bone marrow cells in cancer patients. J Med Virol 66:428-434.