
SPERIMENTAILBIOLAB! - cusmibio.unimi.it BIOCHIMICA.pdf · James B. Sumner che isolò e purificò...
31
1 SPERIMENTA IL BIOLAB IL LATO OSCURO DEGLI ALIMENTI GLI ENZIMI E I LORO INIBITORI NATURALI UNIVERSITA’ DEGLI STUDI DI MILANO
-
Upload
hoangtuong -
Category
Documents
-
view
215 -
download
0
Transcript of SPERIMENTAILBIOLAB! - cusmibio.unimi.it BIOCHIMICA.pdf · James B. Sumner che isolò e purificò...

1
SPERIMENTA IL BIOLAB
IL LATO OSCURO DEGLI ALIMENTI
GLI ENZIMI E I LORO INIBITORI NATURALI
UNIVERSITA’ DEGLI STUDI DI MILANO

2
INDICE
1. CONOSCENZE PROPEDEUTICHE Amminoacidi- proteine- struttura delle proteine-digestione proteica
2. INTRODUZIONE AGLI ENZIMI
- Caratteristiche degli enzimi - Classificazione degli enzimi - Enzimi monomerici e oligomerici - Isoenzimi
3. COME LAVORANO GLI ENZIMI
- Cambiamenti energetici - Catalisi dal punto di vista chimico - Cinetica enzimatica - Fattori che influenzano la velocità di una reazione enzimatica
4. REGOLAZIONE DELL’ATTIVITÀ ENZIMATICA
- Modificazioni della quantità di enzima - Modificazione covalente - Enzimi allosterici
5. INIBIZIONE ENZIMATICA
- Inibitori irreversibili - Inibitori reversibili: competitivi e non competitivi
6. ANTINUTRIENTI CONTENUTI NEGLI ALIMENTI E INIBITORI DELLE PROTEASI
- Classificazione dei fattori antinutrizionali - Metodi per ridurre gli antinutrienti nei vegetali - Inibitori delle proteasi - Inibitori delle proteasi e farmaci
7. PROTOCOLLO SPERIMENTALE
- Storia - Protocollo - Cenni di spettrofotometria - Link utili
8. ATTIVITA’ DI LABORATORIO CHE SI POSSONO SVOLGERE A SCUOLA
- Laboratorio 1: La bromelina - Laboratorio 2: La pepsina - Laboratorio 3: Il vulcano

3
1) CONOSCENZE PROPEDEUTICHE
Amminoacidi- proteine- struttura delle proteine-digestione proteica
2) INTRODUZIONE AGLI ENZIMI
Si comincia a parlare di enzimi nel 1850 con Luois Pasteur, il quale osservò che la fermentazione dello zucchero in alcol è catalizzata da “fermenti” li chiamò enzimi e postulò che fossero inseparabili dalla struttura della cellula vivente. Successivamente: Nel 1897 Eduard Buckner estrasse enzimi attivi da una cellula. Nel 1903 Victor Henri propose per la prima volta la formazione di un complesso Enzima-Substrato come tappa necessaria per iniziare la catalisi. Nel 1913 l’idea di Henri diventò una teoria generale sugli enzimi ad opera di Leonor Michaelis e Maud Menten. Nel 1926 Summer estrasse, isolò e purificò il primo enzima (ureasi da semi di soia) dimostrando la sua natura proteica e concluse che gli enzimi sono proteine ma la sua ipotesi non venne accettata; fu solo tra il 1930 e il 1936, quando Northrop cristallizzò pepsina, tripsina e chimotripsina, che venne stabilita la natura proteica degli enzimi (1946 Nobel Chimica a Sumner, Northrop e Stanley).
Fig. 1. Louis Pasteur (a sinistra) introdusse il termine enzimi nel 1850; Leonor Michaelis e Maud Menten nel 1913 sviluppano la teoria generale sugli enzimi, le tre ultime foto a destra mostrano i vincitori del Nobel per la Chimica nel 1946, rispettivamente James B. Sumner che isolò e purificò l’ureasi e John H. Northrop e Wendell Stanley che per primi cristallizzarono alcuni enzimi.
- Caratteristiche degli enzimi
Gli enzimi sono dei catalizzatori biologici; un catalizzatore è una sostanza che senza subire variazioni produce un incremento della velocità di una reazione chimica. La maggior parte dei catalizzatori biologici sono proteine, esistono anche molecole di RNA coinvolte nelle catalisi, esempio il ribosoma, in questo caso si chiamano ribozimi. Molte reazioni chimiche richiedono - alte temperature ed elevate pressioni, valori estremi di pH e solventi non acquosi - condizioni NON compatibili con quelle fisiologiche. Infatti nelle condizioni fisiologiche, senza l’intervento degli enzimi, i tempi di reazione sarebbero lentissimi e incompatibili con la vita. La catalisi enzimatica è quindi un processo essenziale per gli organismi viventi. L’azione catalizzatrice degli enzimi si differenzia da quella dei catalizzatori non enzimatici per tre caratteristiche: - alta efficienza gli enzimi hanno spesso un’efficienza catalitica prossima alla perfezione: ogni interazione una reazione; producono prodotti con altissime rese– spesso più alte di 95%, e la velocità di reazione catalizzata da un enzima può aumentare di un fattore fino a 1015 rispetto alla reazione non catalizzata - è regolabile
possibilità di variazione da uno stato di bassa o nulla attività dell’enzima ad uno di massima attività mediante l’interazione con altre molecole, attivatori o inibitori
- alta specificicità ogni enzima catalizza, generalmente, una determinata reazione a carico di un substrato specifico. Gli enzimi riconoscono selettivamente i giusti substrati rispetto ad altre molecole. Molti enzimi sono stereospecifici, essendo

4
attivi solo su uno dei due enantiomeri (coppia di entità molecolari che sono immagini speculari ciascuna dell'altra). Altri hanno specificità meno stretta, agiscono su più substrati ma lo fanno con affinità e velocità diverse.
Fig. 2. La molecola del glucosio è chirale; esistono quindi due enantiomeri, l'uno speculare all'altro, il D-glucosio e l'L-glucosio. Dei due, solo il primo (D) è quello utilizzato e prodotto dagli organismi viventi.
Alcuni enzimi richiedono cofattori per la propria attività: - Cofattori inorganici che sono ioni metallici; - Cofattori organici, detti coenzimi. I coenzimi sono vitamine (soprattutto quelle idrosolubili del gruppo B) che vengono trasformate chimicamente in coenzimi ed intervengono nelle reazioni di catalisi insieme ad altri enzimi.
1) Cofattori Inorganici (ioni metallici) 2) Cofattori organici (coenzimi)
Tabella 1. Principali Coenzimi e Cofattori
- Classificazione degli enzimi A causa del crescente numero di enzimi identificati nel 1961 la IUB (International Union of Biochemestry) propose un sistema di classificazione basato sul tipo di reazione catalizzata e sul nome del substrato di ciascun enzima. Gli enzimi sono classificati in 6 classi:

5
Tabella 2. Classificazione internazionale degli enzimi ( nomi delle classi, numeri di codice e tipo di reazione catalizzata).
- Enzimi monomerici e oligomerici ENZIMI MONOMERICI: sono costituiti da una sola catena peptidica (subunità). Ne sono noti pochi. Es.: pepsina, tripsina, elastasi, carbossipeptidasi, trombina, ribonucleasi, lisozima. ENZIMI OLIGOMERICI: sono costituiti da 2 o più subunità, che possono essere uguali o diverse.
tripsina (un'unica subunità) alcool deidrogenasi (2 subunità)
Fig. 3. A sinistra la struttura tridimensionale di un enzima monomerico, la tripsina, e a destra di un enzima oligomerico, l’alcool deidrogenasi.
- Isoenzimi
Sono enzimi esistenti in forme molecolari diverse che catalizzano la stessa reazione sullo stesso substrato, anche se spesso con modalità, affinità e velocità diverse; in diversi tipi cellulari o diversi comparti cellulari.
Per es., la LATTATO DEIDROGENASI (LDH), enzima che catalizza la conversione del lattato in acido piruvico e viceversa, è costituita da 4 subunità, che possono essere di 2 tipi, H (heart) e M (muscle). Ne derivano 5 isoenzimi. La CREATINA CHINASI (CK) catalizza la trasformazione della creatina a fosfocreatina, è costituita da due subunità che possono essere di 2 tipi: B (brain) e M (muscle). Presenta quindi 3 isoenzimi:CK1 (BB) - CK2 MB (cuore) - CK3 MM (cuore, muscolo).
Tabella 3. I cinque isoenzimi della Latico Deidrogenasi ed i tre isoenzimi della Creatina Chinasi e la loro distribuzione nei diversi tessuti.

6
Gli isoenzimi possono essere usati come indicatori diagnostici.
Esempio nell’infarto del miocardio si ha necrosi delle cellule muscolari, i cui enzimi allora si riversano in circolo, in particolare la: CK2 e LDH1
Fig. 4. A sinistra il grafico dell’andamento dell’attività della CK2 e LDH nelle prima 24 ore dopo l’infarto. Nella figura di destra un rappresentazione delle variazioni quantitative, nel siero, dei varie forme isoenzimatiche della LDH in caso di infarto del miocardio e di epatite acuta.
La determinazione dell’attività catalitica di numerosi enzimi (dosaggi enzimatici) è di uso comune nella diagnostica medica. Esempi di enzimi che hanno un rilevante valore diagnostico: - glutammico-ossalacetico transaminasi (GOT): aumenta nel siero marcatamente e rapidamente dopo un infarto; - glutammico-piruvico transaminasi (GPT): aumenta nel siero in seguito a lesioni epatiche; - fosfatasi acida: aumenta nel siero in seguito a carcinoma prostatico con metastasi; - fosfatasi alcalina: aumenta nel siero in seguito a rachitismo, iperfunzionamento delle ghiandole paratiroidi ecc. 3) COME LAVORANO GLI ENZIMI
Il meccanismo di azione degli enzimi può essere trattato da tre punti di vista:
- analizzando i cambiamenti energetici che avvengono nel corso della reazione - esaminando il modo in cui il sito attivo favorisce la catalisi dal punto di vista chimico - studiando la cinetica della reazione catalizzata
- Cambiamenti energetici
La velocità di una reazione, misurata come quantità di prodotto ottenuto in un tempo definito, dipende dall’energia di attivazione. L'energia di attivazione è l'energia minima necessaria a un sistema per innescare una reazione chimica, perché questa possa avvenire, è necessaria la collisione di due o più molecole opportunamente orientate e dotate di un minimo livello di energia (l'energia di attivazione, appunto), tale da permettere la collisione malgrado le forze elettriche repulsive generate dalle loro nubi di elettroni esterne. Gli enzimi abbassano l’energia di attivazione, come si osserva in figura 5, aumentando così la velocità delle reazioni chimiche.
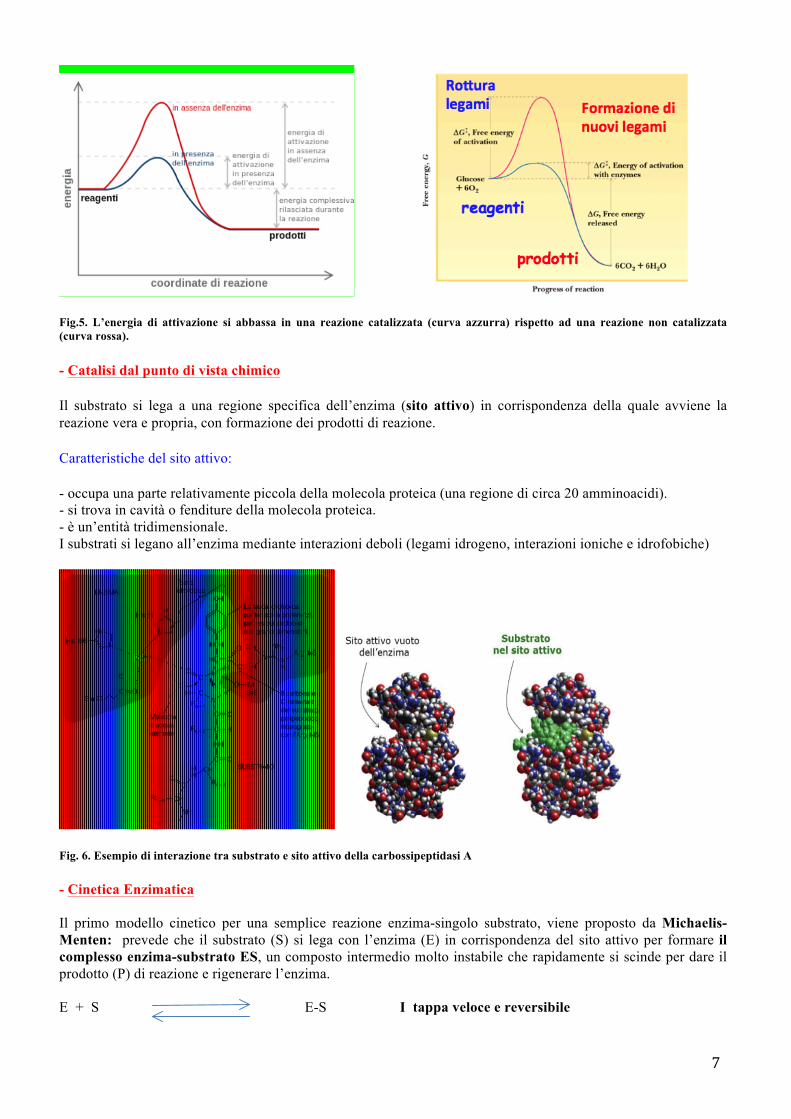
7
Fig.5. L’energia di attivazione si abbassa in una reazione catalizzata (curva azzurra) rispetto ad una reazione non catalizzata (curva rossa).
- Catalisi dal punto di vista chimico
Il substrato si lega a una regione specifica dell’enzima (sito attivo) in corrispondenza della quale avviene la reazione vera e propria, con formazione dei prodotti di reazione.
Caratteristiche del sito attivo:
- occupa una parte relativamente piccola della molecola proteica (una regione di circa 20 amminoacidi). - si trova in cavità o fenditure della molecola proteica. - è un’entità tridimensionale. I substrati si legano all’enzima mediante interazioni deboli (legami idrogeno, interazioni ioniche e idrofobiche)
Fig. 6. Esempio di interazione tra substrato e sito attivo della carbossipeptidasi A
- Cinetica Enzimatica Il primo modello cinetico per una semplice reazione enzima-singolo substrato, viene proposto da Michaelis-Menten: prevede che il substrato (S) si lega con l’enzima (E) in corrispondenza del sito attivo per formare il complesso enzima-substrato ES, un composto intermedio molto instabile che rapidamente si scinde per dare il prodotto (P) di reazione e rigenerare l’enzima. E + S E-S I tappa veloce e reversibile
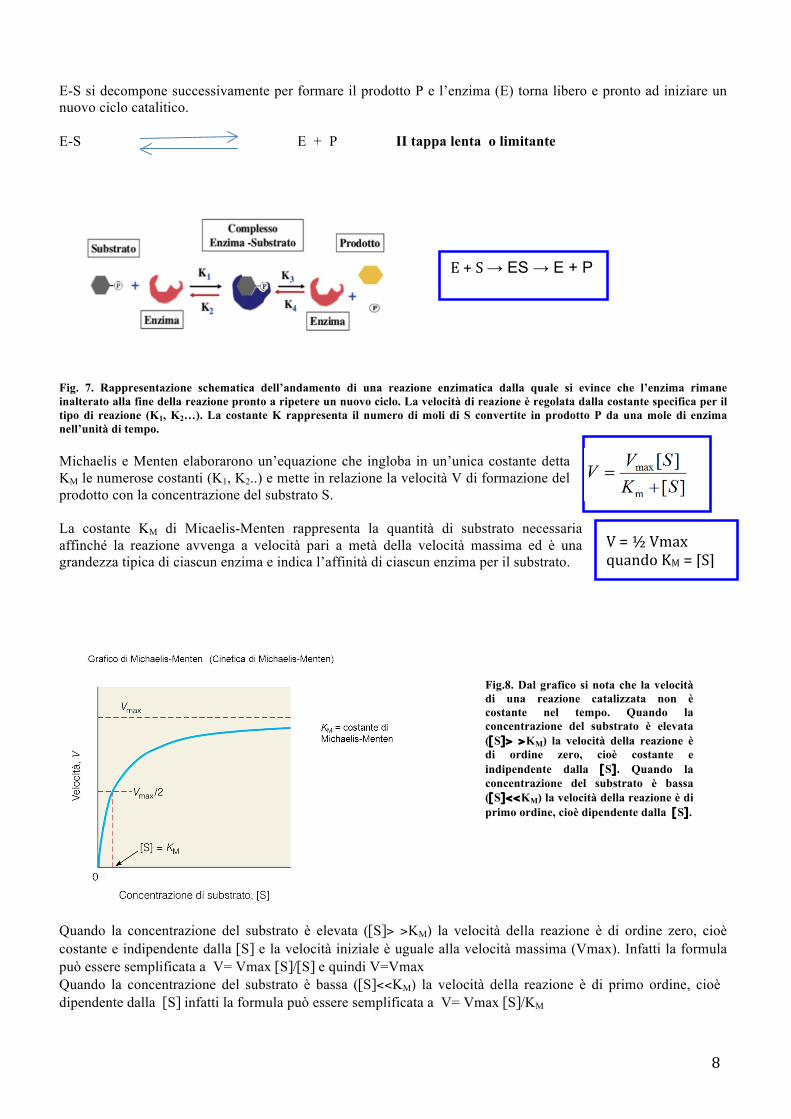
8
E-S si decompone successivamente per formare il prodotto P e l’enzima (E) torna libero e pronto ad iniziare un nuovo ciclo catalitico. E-S E + P II tappa lenta o limitante
Fig. 7. Rappresentazione schematica dell’andamento di una reazione enzimatica dalla quale si evince che l’enzima rimane inalterato alla fine della reazione pronto a ripetere un nuovo ciclo. La velocità di reazione è regolata dalla costante specifica per il tipo di reazione (K1, K2…). La costante K rappresenta il numero di moli di S convertite in prodotto P da una mole di enzima nell’unità di tempo. Michaelis e Menten elaborarono un’equazione che ingloba in un’unica costante detta KM le numerose costanti (K1, K2..) e mette in relazione la velocità V di formazione del prodotto con la concentrazione del substrato S. La costante KM di Micaelis-Menten rappresenta la quantità di substrato necessaria affinché la reazione avvenga a velocità pari a metà della velocità massima ed è una grandezza tipica di ciascun enzima e indica l’affinità di ciascun enzima per il substrato.
Quando la concentrazione del substrato è elevata ([S]> >KM) la velocità della reazione è di ordine zero, cioè costante e indipendente dalla [S] e la velocità iniziale è uguale alla velocità massima (Vmax). Infatti la formula può essere semplificata a V= Vmax [S]/[S] e quindi V=Vmax Quando la concentrazione del substrato è bassa ([S]<<KM) la velocità della reazione è di primo ordine, cioè dipendente dalla [S] infatti la formula può essere semplificata a V= Vmax [S]/KM
E + S → ES → E + P
Fig.8. Dal grafico si nota che la velocità di una reazione catalizzata non è costante nel tempo. Quando la concentrazione del substrato è elevata ([S]> >KM) la velocità della reazione è di ordine zero, cioè costante e indipendente dalla [S] . Quando la concentrazione del substrato è bassa ([S]<<KM) la velocità della reazione è di primo ordine, cioè dipendente dalla [S] .
V = ½ Vmax quando KM = [S]

9
La KM indica la concentrazione del substrato [S] alla quale la V della reazione è ½ Vmax . Il valore della costante è espresso in Molarità (M). L’equazione di Michaelis – Menten descrive il comportamento cinetico di molti enzimi. Tutti i parametri cinetici presenti nell’equazione di Michaelis-Menten possono essere determinati sperimentalmente con sufficiente precisione eccetto Vmax che rappresenta un valore asintotico. I valori di KM e di Vmax possono variare notevolmente a un enzima all’altro; sono funzioni tipiche di ogni enzima e possono servire per valutare e confrontare l’efficienza catalitica di enzimi diversi. La KM può variare anche per substrati diversi dello stesso enzima. Essa viene in genere usata come indicazione dell’affinità di un enzima per il suo substrato: più basso è il valore di KM , più bassa sarà la concentrazione di substrato che permette di raggiungere un valore di velocità di reazione pari alla metà della velocità massima, il che indica un’alta affinità dell’enzima per il suo substrato. Viceversa un alto valore di KM indica che sarà necessario più substrato per raggiungere una velocità di reazione pari alla metà della velocità massima, il che significa una minore affinità dell’enzima per il substrato. Si definisce “ Numero di turnover” (Kcat ) il numero di molecole di substrato convertite in prodotto per molecola di enzima per unità di tempo quando l’enzima è saturato con il substrato; questo valore rappresenta l’efficienza catalitica. Tabella 4. Valori di Km di alcuni enzimi e Valori di Turnover per alcuni enzimi. All’aumentare del valore di Km diminuisce l’affinità dell’enzima per il substrato. All’aumentare della Kcat aumenta l’efficienza catalitica
Valori di Km per alcuni Enzimi Enzima Substrato KM (mM) Anidrasi carbonica CO2 12 Esochinasi Glucosio, Fruttosio 0,15; 1,5
beta-galattosidasi Lattosio 4 Treonin deaminasi Treonina 5 Piruvato carbossilasi
HCO3-
Piruvato 1.0 0.4
Penicillinasi Benzilpenicillina 0.05 Lisozima Esa-N-acetilglucosammina 0.006
- Fattori che influenzano la velocità di una reazione enzimatica La velocità di una reazione catalizzata è influenzata oltre che dalla concentrazione del substrato (la velocità massima si raggiunge nel momento in cui l’enzima è saturato dal substrato) anche dal pH e dalla temperatura. Ogni enzima ha un pH ottimale e una temperatura ottimale, se diminuisce o aumenta, allora l'attività enzimatica diminuisce. Se ci si discosta ulteriormente dai valori ottimali, avviene la denaturazione: punto di non ritorno, se si denatura difficilmente torna alla forma nativa. Il cambiamento del pH altera il grado di ionizzazione sia del substrato che quella dell’enzima modificando il modo di combinarsi dell’uno con l’altro.
Valori di Kcat ( numero di turnover) per alcuni enzimi Enzima Kcat (sec-1) Catalasi 40.000.000 Anidrasi carbonica 1.00 1.000.000 Acetilcolinesterasi 141 14.000 Penicillinasi 2.000 Lattato deidrogenasi 1 1.000 Chimotripsina 100 100 DNA polimerasi I 15 15 Lisozima 0.5 0,5
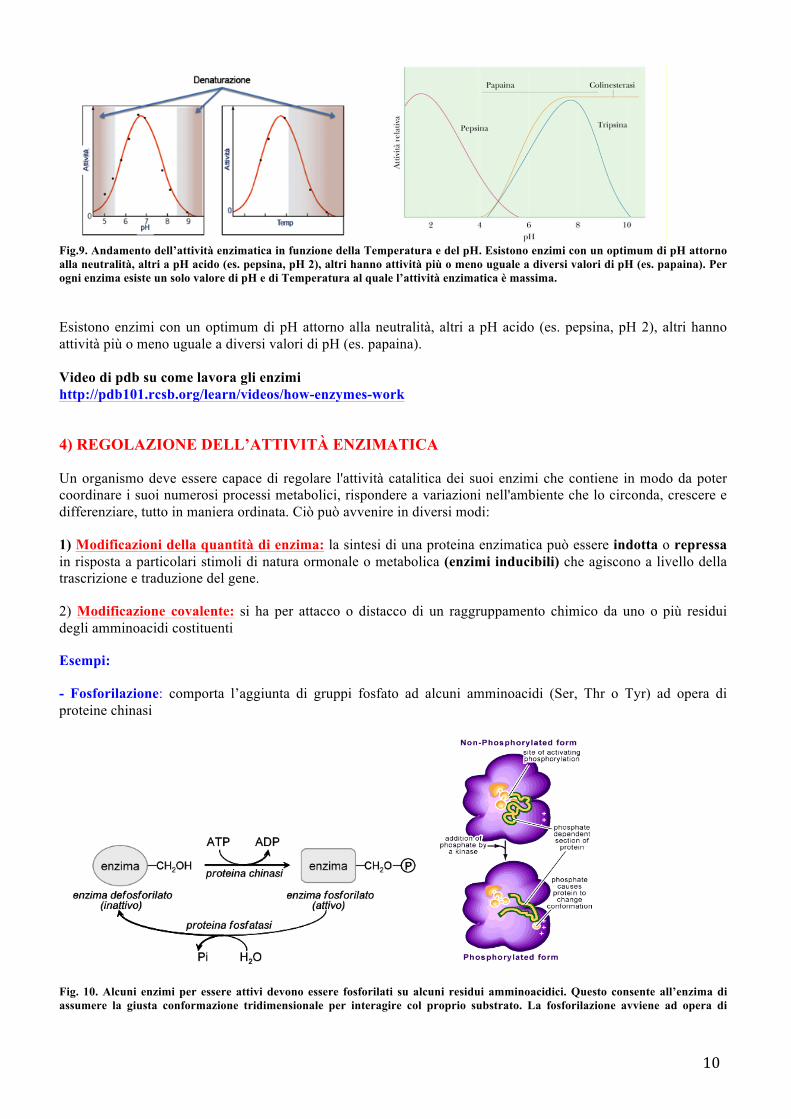
10
Fig.9. Andamento dell’attività enzimatica in funzione della Temperatura e del pH. Esistono enzimi con un optimum di pH attorno alla neutralità, altri a pH acido (es. pepsina, pH 2), altri hanno attività più o meno uguale a diversi valori di pH (es. papaina). Per ogni enzima esiste un solo valore di pH e di Temperatura al quale l’attività enzimatica è massima. Esistono enzimi con un optimum di pH attorno alla neutralità, altri a pH acido (es. pepsina, pH 2), altri hanno attività più o meno uguale a diversi valori di pH (es. papaina).
Video di pdb su come lavora gli enzimi http://pdb101.rcsb.org/learn/videos/how-enzymes-work
4) REGOLAZIONE DELL’ATTIVITÀ ENZIMATICA
Un organismo deve essere capace di regolare l'attività catalitica dei suoi enzimi che contiene in modo da poter coordinare i suoi numerosi processi metabolici, rispondere a variazioni nell'ambiente che lo circonda, crescere e differenziare, tutto in maniera ordinata. Ciò può avvenire in diversi modi:
1) Modificazioni della quantità di enzima: la sintesi di una proteina enzimatica può essere indotta o repressa in risposta a particolari stimoli di natura ormonale o metabolica (enzimi inducibili) che agiscono a livello della trascrizione e traduzione del gene.
2) Modificazione covalente: si ha per attacco o distacco di un raggruppamento chimico da uno o più residui degli amminoacidi costituenti
Esempi:
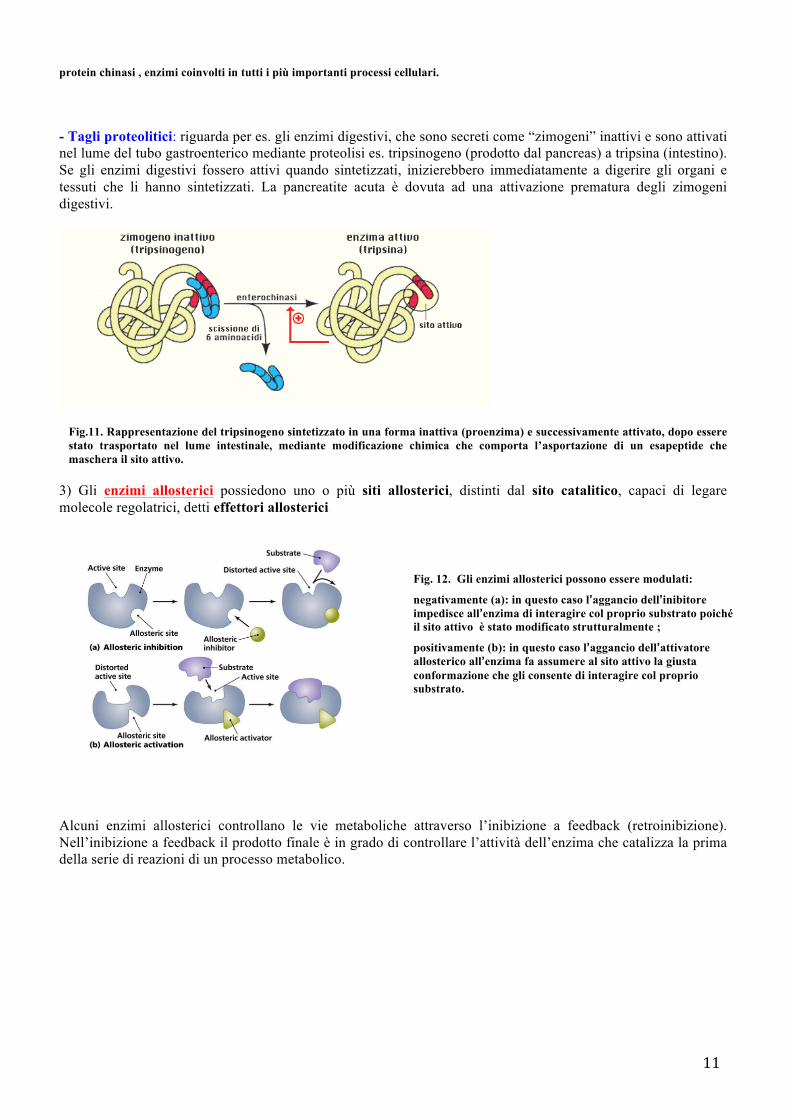
- Fosforilazione: comporta l’aggiunta di gruppi fosfato ad alcuni amminoacidi (Ser, Thr o Tyr) ad opera di proteine chinasi
Fig. 10. Alcuni enzimi per essere attivi devono essere fosforilati su alcuni residui amminoacidici. Questo consente all’enzima di assumere la giusta conformazione tridimensionale per interagire col proprio substrato. La fosforilazione avviene ad opera di
11
protein chinasi , enzimi coinvolti in tutti i più importanti processi cellulari.
- Tagli proteolitici: riguarda per es. gli enzimi digestivi, che sono secreti come “zimogeni” inattivi e sono attivati nel lume del tubo gastroenterico mediante proteolisi es. tripsinogeno (prodotto dal pancreas) a tripsina (intestino). Se gli enzimi digestivi fossero attivi quando sintetizzati, inizierebbero immediatamente a digerire gli organi e tessuti che li hanno sintetizzati. La pancreatite acuta è dovuta ad una attivazione prematura degli zimogeni digestivi.
Fig.11. Rappresentazione del tripsinogeno sintetizzato in una forma inattiva (proenzima) e successivamente attivato, dopo essere stato trasportato nel lume intestinale, mediante modificazione chimica che comporta l’asportazione di un esapeptide che maschera il sito attivo.
3) Gli enzimi allosterici possiedono uno o più siti allosterici, distinti dal sito catalitico, capaci di legare molecole regolatrici, detti effettori allosterici
Alcuni enzimi allosterici controllano le vie metaboliche attraverso l’inibizione a feedback (retroinibizione). Nell’inibizione a feedback il prodotto finale è in grado di controllare l’attività dell’enzima che catalizza la prima della serie di reazioni di un processo metabolico.
Fig. 12. Gli enzimi allosterici possono essere modulati: negativamente (a): in questo caso l’aggancio dell’inibitore impedisce all’enzima di interagire col proprio substrato poiché il sito attivo è stato modificato strutturalmente ; positivamente (b): in questo caso l’aggancio dell’attivatore allosterico all’enzima fa assumere al sito attivo la giusta conformazione che gli consente di interagire col proprio substrato.

12
5) INIBIZIONE ENZIMATICA Molti enzimi, oltre che con i propri substrati, cofattori e prodotti, possono combinarsi anche con altre molecole formando complessi che perdono in parte o totalmente la capacità di legare il substrato e/o di catalizzare la reazione. Queste molecole sono indicate come inibitori ed inibizione enzimatica il fenomeno di inattivazione dell’enzima. Oltre ad avere una grande importanza farmacologica (molti farmaci sono inibitori enzimatici: statine, AZT, aspirina, penicilline, ACE inibitori ecc.) lo studio degli inibitori ha fornito importanti informazioni sul meccanismo di azione degli enzimi. Gli inibitori sono divisi in due classi: - inibitori irreversibili che inibiscono l’enzima legandosi a questo in modo stabile, spesso covalente, e quindi irreversibilmente - inibitori reversibili che si combinano con l’enzima per mezzo di legami deboli e possono quindi essere allontanati lasciando l’enzima nuovamente attivo. Gli inibitori irreversibili si combinano o distruggono un gruppo funzionale dell’enzima, essenziale per la sua attività catalitica. Poiché gli stessi gruppi funzionali sono presenti nel sito attivo di numerosi enzimi, uno stesso inibitore irreversibile può agire inattivando gruppi anche molto ampi di enzimi Molte sostanze altamente tossiche sono in realtà inibitori irreversibili di enzimi; per esempio composti organofosforici quali il diisopropilfluorofosfato (DIPF), anche noti come gas nervini, devono la loro estrema tossicità alla capacità di combinarsi irreversibilmente nel sito attivo dell’enzima acetilcolinesterasi (enzima che idrolizza il neurotrasmettitore acetilcolina in acetato e colina), che viene inattivato, con grave compromissione della funzionalità del sistema nervoso. Allo stesso modo, questi composti organofosforici inattivano numerosi altri enzimi come la tripsina, la chimotripsina, la fosfoglucomutasi e molte esterasi.
Fig. 13. Esempio di inibizione a feedback secondo cui il prodotto di una via metabolica inibisce l’attività del primo enzima della via metabolica stessa. Il primo enzima della via metabolica è un enzima allosterico, esso possiede un sito anche per il prodotto finale oltre che per il suo specifico substrato. Quando il prodotto finale della via metabolica si lega all'enzima, questo non potrà più legarsi al suo substrato e tutta la via sarà interrotta. Quando il prodotto finale sarà tutto esaurito, l’enzima allosterico potrà riprendere la sua funzione. E’ un meccanismo che permette di regolare l’attività degli enzimi di una via metabolica in base alla disponibilità del prodotto.

13
Fig. 14. Composti orgonofosforici: Sarin è una neurotossina, nota per essere stata rilasciata da terroristi nel 1955 nella metro di Tokio; Parathion e Malathion sono due insetticidi efficaci perché molto tossici per gli insetti
Alcuni sali di Arsenico, Piombo, Mercurio e altri metalli pesanti devono la loro tossicità alla capacità di legarsi covalentemente con gruppi –SH dei residui di cisteina; conseguentemente tali residui non possono più formare i legami disolfuro indispensabili per stabilizzare la struttura tridimensionale di molti enzimi e nel caso siano presenti nel sito attivo, partecipare al processo catalitico. Una classe particolare d’inibitori irreversibili è costituito dagli inibitori suicidi ossia composti la cui reattività aumenta quando si trovano nel sito attivo di un particolare enzima. L’inattivatore suicida viene convertito dall’enzima in un composto molto reattivo che si combina in modo irreversibile con l’enzima stesso inattivandolo completamente. La produzione di questo tipo d’inibitori è uno dei campi che la moderna ricerca farmacologica sta esplorando per ottenere nuovi agenti terapeutici efficaci e privi di effetti collaterali. Esempio la Penicillina inattiva irreversibilmente la glicopeptide transpeptidasi, un enzima chiave della sintesi del peptidoglicano della parete batterica. La conformazione della penicillina può entrare facilmente nel sito attivo dell’enzima dove si lega molto stabilmente, appena l’enzima tenta di rompere il legame peptidico contenuto nella molecola di penicillina, questa si lega covalentemente e quindi irreversibilmente al sito attivo dell’enzima stesso inibendo la sintesi della parete batterica.
Fig. 15. A sinistra la molecola di penicillina con l’anello b-lattamico instabile; a destra l’enzima transpeptidasi batterica capace di rompere il legame consentendo alla penicillina di legarsi irreversibilmente all’enzima. La maggior parte dei batteri resistenti alla penicillina esprimono la penicillasi una beta-lattamasi che inattiva la penicillina rompendo l’anello beta-lattamico e impedendo alla penicillina di legarsi alla transpeptidasi.
Gli inibitori reversibili possono ridurre l’attività degli enzimi in due modi:
- legandosi nel sito attivo al posto del substrato (inibitori competitivi) - legandosi solo al complesso ES e quindi in un sito diverso dal sito attivo (inibitori non competitivi)
Gli inibitori competitivi sono molecole con struttura simile a quella della molecola di substrato. L’inibitore compete con il substrato per il legame al sito attivo dell’enzima. L’inibitore unito all’enzima non subisce la catalisi e può essere rimosso aggiungendo altro substrato. Un inibitore competitivo modifica la KM e lascia inalterata la Vmax. In realtà, l’affinità dell’enzima per il substrato, alla presenza dell’inibitore, cambia solo apparentemente: infatti, data la competizione per il sito attivo tra substrato ed inibitore, in presenza dell’inibitore è necessaria una concentrazione di substrato maggiore perché l’enzima catalizzi la reazione con una velocità semimassimale; in altre parole è come se l’affinità dell’enzima per il substrato venisse a diminuire.

14
Farmaci come inibitori competitivi: statine, sulfamidici, metotrexate, AZT (Azidotimidina, detta anche Zidovutina).
L’etanolo viene utilizzato come inibitore competitivo nella terapia dell’avvelenamento da metanolo o da glicoletilenico (anticongelante dei radiatori).
Fig.16. Inibizione competitiva. Un inibitore competitivo si lega al sito attivo dell’enzima in competizione con le molecole di substrato. Ciò provoca un aumento del valore della Km mentre il valore della Velocità massima resta invariato.
Fig. 17. Il metotrexate è un analogo del diidrofolato, coenzima della diidrofolato reduttasi (enzima coinvolto nella biosintesi dei nucleotidi). Si lega all’enzima 1000 volte più saldamente del suo substrato naturale. Viene usato come antitumorale.
15
Fig. 18. Il glicol etilenico e il metanolo vengono ossidati dall’alcool deidrogenasi (ADH) in composti tossici per l’organismo (acido ossalico e formaldeide rispettivamente). L’etanolo, normalmente ossidato ad acetaldeide dall’ADH, è un inibitore competitivo di ADH. La terapia consiste in una infusione intravenosa di etanolo che rallenta la formazione del metabolita tossico in modo da permettere un’escrezione lenta in concentrazioni sufficientemente basse da non provocare danni cellulari.
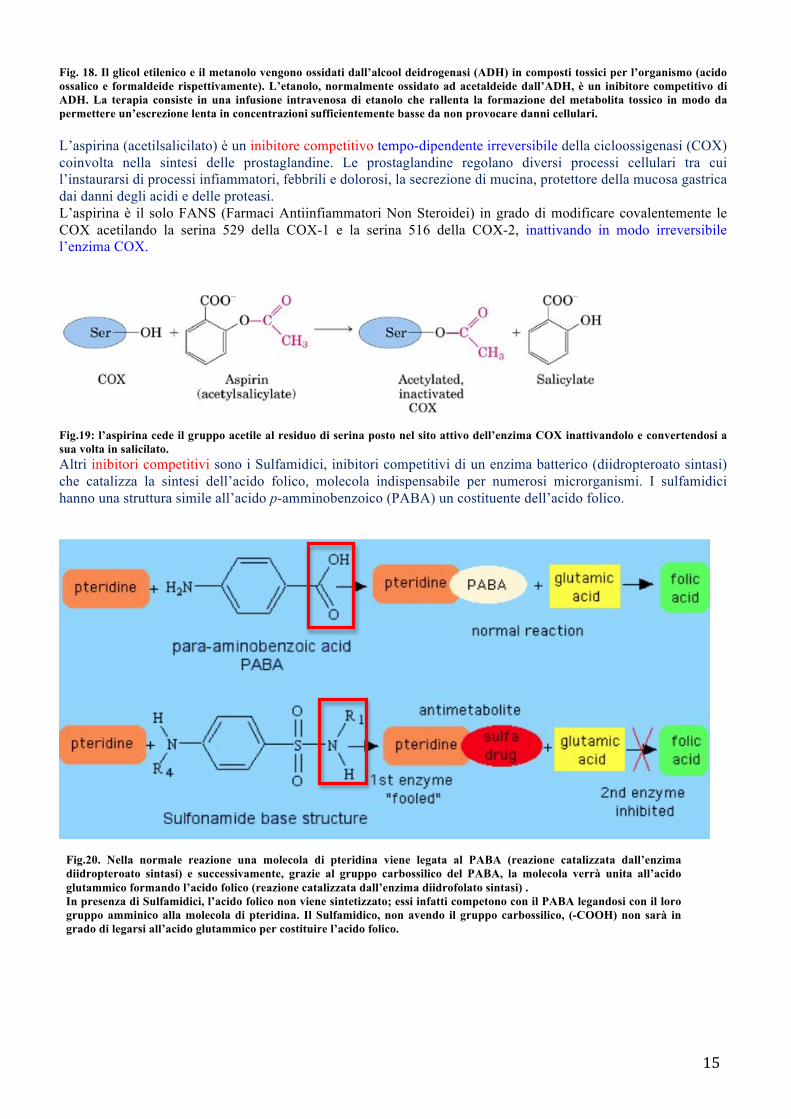
L’aspirina (acetilsalicilato) è un inibitore competitivo tempo-dipendente irreversibile della cicloossigenasi (COX) coinvolta nella sintesi delle prostaglandine. Le prostaglandine regolano diversi processi cellulari tra cui l’instaurarsi di processi infiammatori, febbrili e dolorosi, la secrezione di mucina, protettore della mucosa gastrica dai danni degli acidi e delle proteasi. L’aspirina è il solo FANS (Farmaci Antiinfiammatori Non Steroidei) in grado di modificare covalentemente le COX acetilando la serina 529 della COX-1 e la serina 516 della COX-2, inattivando in modo irreversibile l’enzima COX.
Fig.19: l’aspirina cede il gruppo acetile al residuo di serina posto nel sito attivo dell’enzima COX inattivandolo e convertendosi a sua volta in salicilato. Altri inibitori competitivi sono i Sulfamidici, inibitori competitivi di un enzima batterico (diidropteroato sintasi) che catalizza la sintesi dell’acido folico, molecola indispensabile per numerosi microrganismi. I sulfamidici hanno una struttura simile all’acido p-amminobenzoico (PABA) un costituente dell’acido folico.
Fig.20. Nella normale reazione una molecola di pteridina viene legata al PABA (reazione catalizzata dall’enzima diidropteroato sintasi) e successivamente, grazie al gruppo carbossilico del PABA, la molecola verrà unita all’acido glutammico formando l’acido folico (reazione catalizzata dall’enzima diidrofolato sintasi) . In presenza di Sulfamidici, l’acido folico non viene sintetizzato; essi infatti competono con il PABA legandosi con il loro gruppo amminico alla molecola di pteridina. Il Sulfamidico, non avendo il gruppo carbossilico, (-COOH) non sarà in grado di legarsi all’acido glutammico per costituire l’acido folico.

16
I sulfamidici non sono nocivi per l’uomo, che non possiede l’enzima che catalizza la sintesi dell’acido folico e per ottenere questa sostanza dipende dall’apporto alimentare o dalla sintesi da parte della flora batterica intestinale.
Il farmaco antigottoso Allopurinolo è un inibitore competitivo della xantina ossidasi, enzima coinvolto nel catabolismo delle basi puriniche. La gotta è causata da alti livelli di acido urico nel sangue e nei tessuti, prodotto da un carico troppo alto di purine che si deposita nelle giunture delle articolazioni e nei tubuli renali sotto forma di cristalli di urato di sodio.
ACE inibitori sono inibitori competitivi usati come farmaci nella terapia dell'ipertensione arteriosa, del post-infarto del miocardio e dell'insufficienza cardiaca cronica. L'enzima convertitore dell'angiotensina (ACE: Angiotensin converting enzyme) fa parte di una cascata regolatrice della pressione arteriosa (fig.22 ). Appartiene alla famiglia delle esopeptidasi e in particolare delle carbossipeptidasi, ha il compito di tagliare gli ultimi due amminoacidi dell’angiotensina II convertendola nella proteina attiva angiotensina I. L’enzima ACE è coinvolto nel sistema di controllo del volume dei liquidi extracellulari e della vascolarizzazione delle arterie. La mancata regolazione dell'enzima porta a un alto tenore di vasocostrizione responsabile dell'ipertensione. Gli ACE inibitori sono farmaci inibitori dell'enzima di conversione dell'angiotensina (ACE)
Fig.22. Nella figura è rappresentato una cascata di segnali che portano alla regolazione della pressione arteriosa. In rosso sono indicati le molecole che vengono utilizzate contro l’ipertensione, sono una serie di farmaci che portano al blocco della vasocostrizione; tra questi ci sono gli ACE-inibitori che bloccano l’azione dell’enzima Angiotensin Converting Enzyme.
Fig. 21. Azione del farmaco Allopurinolo inibitore competitivo dell’enzima xantina ossidasi. Esso impedisce la formazione dell’acido urico prevenendo la gotta.
17
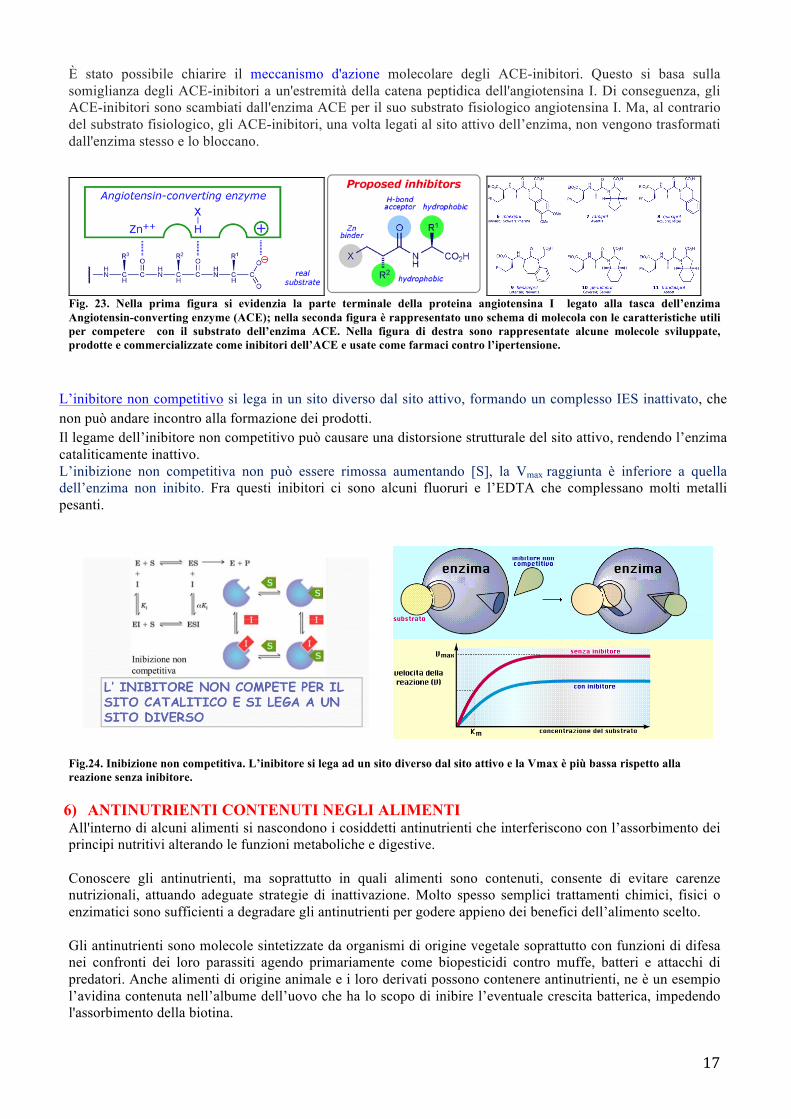
È stato possibile chiarire il meccanismo d'azione molecolare degli ACE-inibitori. Questo si basa sulla somiglianza degli ACE-inibitori a un'estremità della catena peptidica dell'angiotensina I. Di conseguenza, gli ACE-inibitori sono scambiati dall'enzima ACE per il suo substrato fisiologico angiotensina I. Ma, al contrario del substrato fisiologico, gli ACE-inibitori, una volta legati al sito attivo dell’enzima, non vengono trasformati dall'enzima stesso e lo bloccano.
Fig. 23. Nella prima figura si evidenzia la parte terminale della proteina angiotensina I legato alla tasca dell’enzima Angiotensin-converting enzyme (ACE); nella seconda figura è rappresentato uno schema di molecola con le caratteristiche utili per competere con il substrato dell’enzima ACE. Nella figura di destra sono rappresentate alcune molecole sviluppate, prodotte e commercializzate come inibitori dell’ACE e usate come farmaci contro l’ipertensione.
L’inibitore non competitivo si lega in un sito diverso dal sito attivo, formando un complesso IES inattivato, che non può andare incontro alla formazione dei prodotti. Il legame dell’inibitore non competitivo può causare una distorsione strutturale del sito attivo, rendendo l’enzima cataliticamente inattivo. L’inibizione non competitiva non può essere rimossa aumentando [S], la Vmax raggiunta è inferiore a quella dell’enzima non inibito. Fra questi inibitori ci sono alcuni fluoruri e l’EDTA che complessano molti metalli pesanti.
Fig.24. Inibizione non competitiva. L’inibitore si lega ad un sito diverso dal sito attivo e la Vmax è più bassa rispetto alla reazione senza inibitore.
6) ANTINUTRIENTI CONTENUTI NEGLI ALIMENTI All'interno di alcuni alimenti si nascondono i cosiddetti antinutrienti che interferiscono con l’assorbimento dei principi nutritivi alterando le funzioni metaboliche e digestive. Conoscere gli antinutrienti, ma soprattutto in quali alimenti sono contenuti, consente di evitare carenze nutrizionali, attuando adeguate strategie di inattivazione. Molto spesso semplici trattamenti chimici, fisici o enzimatici sono sufficienti a degradare gli antinutrienti per godere appieno dei benefici dell’alimento scelto. Gli antinutrienti sono molecole sintetizzate da organismi di origine vegetale soprattutto con funzioni di difesa nei confronti dei loro parassiti agendo primariamente come biopesticidi contro muffe, batteri e attacchi di predatori. Anche alimenti di origine animale e i loro derivati possono contenere antinutrienti, ne è un esempio l’avidina contenuta nell’albume dell’uovo che ha lo scopo di inibire l’eventuale crescita batterica, impedendo l'assorbimento della biotina.
18
- Classificazione dei fattori antinutrizionali - Fattori che hanno un effetto depressivo sulla digestione proteica e sulla utilizzazione delle proteine (inibitori della tripsina e della chimotripsina, lectine o emoagglutinine, composti fenolici, saponine). Gli inibitori della tripsina (un enzima pancreatico che permette di digerire le proteine) sono contenuti in legumi, patate e albumi delle uova quando questi alimenti sono consumati crudi: il calore, e dunque la cottura, risolve il problema. - Fattori che hanno un effetto negativo sulla digestione dei carboidrati (inibitori dell’amilasi, composti polifenolici, fattori di flautolenza); - Fattori che hanno un effetto negativo sulla digestione e sulla utilizzazione dei minerali (glicosinolati, acido ossalico, acido fitico, tannini): i fitati ad esempio inibiscono l'assorbimento di calcio, magnesio, zinco, rame e ferro e si trovano soprattutto in semi, frutta secca, cereali e legumi. Gli ossalati riducono invece l'assorbimento del calcio e si trovano in verdure come spinaci, peperoni verdi, barbabietole, rabarbaro, melanzane e anche nel cacao. I tannini sono una classe di polifenoli che, nonostante apportino vari benefici per via delle loro proprietà antiossidanti, se assunti in grandi quantità possono interferire con la corretta digestione e assimilazione di vari nutrienti tra cui vitamine e minerali (calcio e ferro), e inibiscono l'azione di alcuni enzimi digestivi. Si trovano nel caffè, nel tè, nel vino, nei cachi e nell'uva. - Fattori che inattivano le vitamine o causa un incremento nel fabbisogno di vitamine (es. l’avidina, il cui nome è dovuto all’avidità con cui sequestra la biotina (vitamina B8 o vitamina H), rendendola indisponibile per l’organismo umano; presente nell’albume crudo); - Fattori che stimolano il sistema immunitario (antigeni).
Tabella 6. Principali fattori antinutrizionali presenti in alcuni vegetali utilizzati come alimenti

19
Tabella 7. Effetti in vivo dei vari fattori antinutrizionali (FAN). Concludendo si può affermare che la presenza di antinutrienti negli alimenti non deve intimorire, bensì deve sicuramente essere un campanello di allarme per tutti coloro che scegliendo regimi alimentari monotoni e poco diversificati, devono aver cura di applicare piccoli trucchi che permettano di assimilare i macro e i micronutrienti introdotti quotidianamente con la dieta, onde evitare di andare incontro a possibili rischiose carenze. - Metodi per ridurre gli antinutrienti nei vegetali Per ridurre il livello di composti antinutrizionali all'interno degli alimenti vegetali si possono utilizzare diverse tecniche: ammollo, germinazione, fermentazione, cottura a vapore, bollitura o altri metodi di cottura, e/o una combinazione di alcuni di questi metodi. Lista degli antinutrienti e dei metodi di eliminazione ● Fitati (acido fitico): ammollo, germinazione, fermentazione; ● Lectine: ammollo, cottura a vapore, ebollizione o altro metodo di cottura, fermentazione; ● Tannini: ammollo, ebollizione; ● Inibitori della proteasi: ammollo, germinazione, bollitura. ● Ossalato di calcio: ammollo, ebollizione. https://www.piuvivi.com/alimentazione/antinutrienti-cosa-sono-come-inibire-ridurre-nei-cibi.html
- INIBITORI DELLE PROTEASI Le proteasi sono enzimi che tagliano le proteine in punti specfici: es. tripsina taglia tra K e R (lisina e arginina) mentre la chimotripsina taglia a livello di amminoacidi aromatici F,Y,W (fenilalanina, tirosina e triptofano). Dopo un pranzo, il tripsinogeno (precursore inattivo dell’enzima) prodotto dal pancreas e rilasciato nel lume dell’intestino, subisce una proteolisi e diventa tripsina (enzima attivo). La tripsina è una serina proteasi (il nome deriva dal fatto che hanno sempre un residuo di serina nel sito attivo) capace anche di attivare altre serina proteasi come la chimotripsina, l’elastasi e altra tripsina (tramite autocatalisi).

20
Gli inibitori naturali delle proteasi possono essere suddivisi in base alle classi di proteasi che sono in grado di inibire. Tra i più studiati ci sono gli inbibitori delle serina proteasi: - Inibitori di Kunitz - Inibitori di Bowman-Birk Gli inibitori di Kunitz, hanno peso molecolare di circa 20.000 Dalton, contengono da 170 a 200 amminoacidi e due ponti zolfo. Nelle piante hanno un ruolo importante nella difesa delle piante contro i patogeni e come regolatori di proteinasi endogene. Si trovano nei legumi, principalemente nella soia. Sono dei forti inibitori della tripsina e quindi impediscono la corretta digestione delle proteine. Vengono inattivati dai succhi gastrici e dalla cottura (i due ponti di solfuro sono termolabili). Inibitori di Bowman-Birk sono più piccoli, peso 8000 Dalton, hanno molte cisteine capaci di formare legami covalenti, hanno struttura più stabile e sono quindi più termostabile e resistono alla cottura. Sono inibitori competitivi, con una grande affinità per il substrato, e sono potenti inibitori della tripsina e della chimotripsina sono degli antinutrienti.
Fig.26. A sinistra sequenza amminoacidica di un Inibitore di Kunitz (A) e struttura tridimensionale (B) e a destra Inibitore di Bowman-Birk con struttura tridimensionale (A) e sequenza amminoacidica (B) in cui sono indicati i legami covalenti tra gli amminoacidi Cisteina (C). Se nel cibo sono presenti inibitori della tripsina, essi si legano fortemente al sito attivo dell’enzima bloccandolo, alterando così la corretta digestione di certe proteine. In modelli murini, alimentati con una dieta ricca di soia, gli effetti negative degli inibitori delle proteasi includono: riduzione della digeribilità proteica, iperplasia e ipertrofia pancreatica.
- INIBITORI DELLE PROTEASI E FARMACI
Fig.25. Rappresentazione della proteolisi. Le proteasi sono enzimi che idrolizzano il legame peptidico che si viene a formare tra gli amminoacidi di una proteina. (a) Struttura di un peptide. (b) Le endopeptidasi sono delle proteasi che idrolizzano i legami peptidici che si trovano all’interno della proteina. (c) Le esopeptidasi idrolizzano i legami peptidici che si trovano alle due estremità della catena peptidica, liberando singoli amminoacidi. Si distinguono in Carbossipeptidasi, se idrlizzano i legami peptidici a partire dall’estremità –C terminale, e Aminopeptidasi se idrolizzano i legami peptidici a partire dall’estremità –N terminale.

21
La Terapia antivirale ad alta efficacia oggi maggiormente utilizzata nella terapia dell’AIDS si chiama HAART (Highly active antiretroviral therapy), il protocollo è basato sulla combinazione di farmaci antivirali con diverso meccanismo di azione e bersaglio:
• Inibitori della trascrittasi inversa (nucleosidici, nucleotidici e non nucleosidici), • Inibitori della proteasi virale, • Inibitori dell’integrasi, • Inibitori della fusione e dell’ingresso virale.
La combinazione di tre di questi farmaci consente di bloccare il virus a diverse fasi del ciclo replicativo e di neutralizzare gli effetti di singole mutazioni che inducono resistenza a uno dei farmaci, che possono insorgere durante la terapia. La terapia diventerebbe inefficace solo nel caso in cui, in uno stesso virione, si accumulassero 3 mutazioni distinte, ognuna delle quali induce resistenza a uno dei 3 farmaci, evento la cui frequenza è statisticamente inferiore al numero di virioni circolanti presenti in un soggetto AIDS )
prevenire i meccanismi di resistenza che possono insorgere per la mutazione del virus nel corso del tempo.
Gli inibitori della proteasi rappresentano la classe più potente tra i farmaci antivirali; essi hanno come obiettivo l’enzima virale proteinasi. L’inibizione della proteasi virale previene il taglio delle poliproteine gag-pol, da cui derivano le proteine strutturali necessarie per l’assemblamento dei virioni maturi. Le particelle virali che vengono prodotte in presenza del farmaco inibitore della proteasi virale, non sono infettive. Tra questi farmaci annoveriamo il saquinavir, il ritonavir, l’indinavir e il nelfinavir (sono analoghi del substrato quindi inibitori competitivi)
È noto da tempo che le terapie a base di inibitori della proteasi riducono l'incidenza del sarcoma di Kaposi, un tumore maligno dei vasi sanguigni che, prima dell’introduzione di questa classe di farmaci, colpiva di frequente le persone malate di AIDS, e rallentano la sua progressione, nel caso in cui il tumore si è manifestato. Si stanno sperimentando i farmaci anti-HIV per la cura di forme tumorali.
Inibitori proteasi e cancro Le metastasi sono cellule maligne che si staccano dal tumore originario e si diffondono in altri organi dove possono riprodursi e generare nuove localizzazioni tumorali. Nelle generazione delle metastasi è determinante l’ azione di diverse proteasi extracellulari. L’azione di questi enzimi proteolitici causa la degradazione della matrice extracellulare facendo aumentare la capacità di migrazione delle cellule tumorali e quindi la diffusione del tumore. Le varie proteasi hanno ruoli distinti nella “cascata metastatica” che comprende: distacco, invasione dei vasi, migrazione, adesività, angiogenesi tumorale e anche un ruolo nella capacità del sistema immunitario di rilevare il cancro. Gli inibitori delle proteasi possono quindi diventare dei farmaci efficaci contro i tumori.
Fig.27. Nel caso dell’HIV, l’inibizione della proteasi impedisce il rilascio di singole proteine dalle gag e la successiva maturazione di particelle virali infettive.

22
Collettivamente, i geni delle proteasi e i loro inibitori naturali sono conosciuti come "degradome". Il progetto Cancerdegradome, finanziato dall'UE, sta studiando il "degradome" per definire nuovi bersagli molecolari per la progettazione di farmaci contro il cancro. (http://cordis.europa.eu/result/rcn/89066_it.html)
Fig.28. Anticorpi nel tentativo di intercettare cellule metastatiche in circolo.

23
7) PROTOCOLLO SPERIMENTALE STORIA
La soia (Glicine max) è un legume originario dell’Asia, i suoi semi sono ampiamente utilizzati in alimentazione animale e umana perché contengono elevate quantità di olio (20%) e proteina (40%), sono infatti i semi con più elevato contenuto proteico. Il suo impiego diretto è limitato dalla presenza di alcuni fattori antinutrizionali, che inibiscono la completa utilizzazione delle proteine per gli animali e l’uomo. Tali fattori, presenti nella soia cruda, solitamente vengono, in parte, eliminati dopo l’estrazione dell’olio con un trattamento termico delle farine ad alte temperature (100 -‐105 ° C). I principali tipi d’inibitori presenti nella soia cruda sono il Kunitz trypsin inibitor e il Bowman-‐Birk i quali, limitando l’azione della tripsina e della chimotripsina, causano un minor sviluppo corporeo rilevabile in particolare nei monogastrici alimentati con soia cruda. Le varietà commerciali di soia e i suoi derivati hanno un’attività antitripsinica variabile che dipende anche dalla modalità di trattamente dei semi e dei prodotti a base di soia. Si hanno a disposizione vari tipi di latte tra cui il “latte di soia” e si vuole determinare presenza e la quantità di inibitori della tripsina.
L’attività della tripsina attiva è misurata utilizzando come substrato L-BAPA (benzoil-L-arginina-para-nitroanilide). La tripsina idrolizzando il legame peptidico (indicato dalla freccia rossa) presente nella molecola BAPA, forma un prodotto di colore giallo (para-nitro-anilina) la cui quantità può essere misurata spettrometricamente a 410n. L’attività degli inibitori della tripsina è valutata aggiungendo alla reazione l’inibitore e valutando la riduzione di p-nitro-aniline prodotto.
inibitori legati alla tripsina (le catene rosa)
24
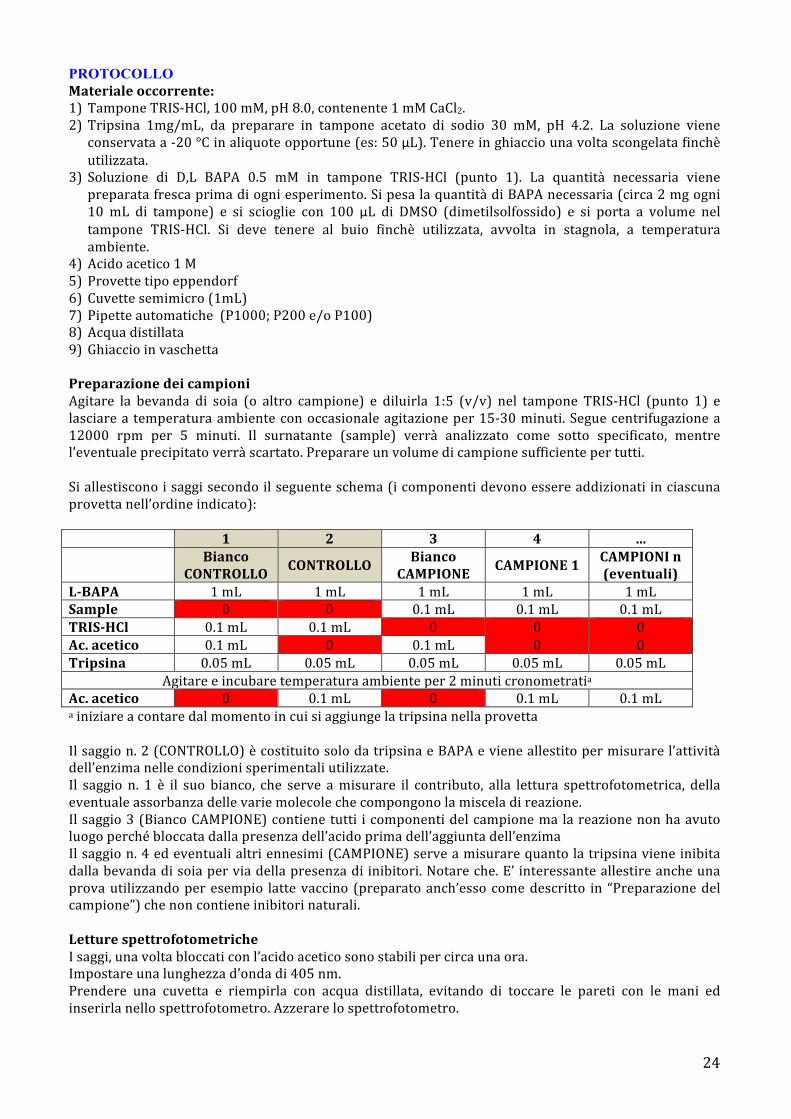
PROTOCOLLO Materiale occorrente: 1) Tampone TRIS-‐HCl, 100 mM, pH 8.0, contenente 1 mM CaCl2. 2) Tripsina 1mg/mL, da preparare in tampone acetato di sodio 30 mM, pH 4.2. La soluzione viene conservata a -‐20 °C in aliquote opportune (es: 50 µL). Tenere in ghiaccio una volta scongelata finchè utilizzata.
3) Soluzione di D,L BAPA 0.5 mM in tampone TRIS-‐HCl (punto 1). La quantità necessaria viene preparata fresca prima di ogni esperimento. Si pesa la quantità di BAPA necessaria (circa 2 mg ogni 10 mL di tampone) e si scioglie con 100 µL di DMSO (dimetilsolfossido) e si porta a volume nel tampone TRIS-‐HCl. Si deve tenere al buio finchè utilizzata, avvolta in stagnola, a temperatura ambiente.
4) Acido acetico 1 M 5) Provette tipo eppendorf 6) Cuvette semimicro (1mL) 7) Pipette automatiche (P1000; P200 e/o P100) 8) Acqua distillata 9) Ghiaccio in vaschetta Preparazione dei campioni Agitare la bevanda di soia (o altro campione) e diluirla 1:5 (v/v) nel tampone TRIS-‐HCl (punto 1) e lasciare a temperatura ambiente con occasionale agitazione per 15-‐30 minuti. Segue centrifugazione a 12000 rpm per 5 minuti. Il surnatante (sample) verrà analizzato come sotto specificato, mentre l’eventuale precipitato verrà scartato. Preparare un volume di campione sufficiente per tutti. Si allestiscono i saggi secondo il seguente schema (i componenti devono essere addizionati in ciascuna provetta nell’ordine indicato):
1 2 3 4 …
Bianco CONTROLLO CONTROLLO Bianco
CAMPIONE CAMPIONE 1 CAMPIONI n (eventuali)
L-‐BAPA 1 mL 1 mL 1 mL 1 mL 1 mL Sample 0 0 0.1 mL 0.1 mL 0.1 mL TRIS-‐HCl 0.1 mL 0.1 mL 0 0 0 Ac. acetico 0.1 mL 0 0.1 mL 0 0 Tripsina 0.05 mL 0.05 mL 0.05 mL 0.05 mL 0.05 mL
Agitare e incubare temperatura ambiente per 2 minuti cronometratia Ac. acetico 0 0.1 mL 0 0.1 mL 0.1 mL a iniziare a contare dal momento in cui si aggiunge la tripsina nella provetta Il saggio n. 2 (CONTROLLO) è costituito solo da tripsina e BAPA e viene allestito per misurare l’attività dell’enzima nelle condizioni sperimentali utilizzate. Il saggio n. 1 è il suo bianco, che serve a misurare il contributo, alla lettura spettrofotometrica, della eventuale assorbanza delle varie molecole che compongono la miscela di reazione. Il saggio 3 (Bianco CAMPIONE) contiene tutti i componenti del campione ma la reazione non ha avuto luogo perché bloccata dalla presenza dell’acido prima dell’aggiunta dell’enzima Il saggio n. 4 ed eventuali altri ennesimi (CAMPIONE) serve a misurare quanto la tripsina viene inibita dalla bevanda di soia per via della presenza di inibitori. Notare che. E’ interessante allestire anche una prova utilizzando per esempio latte vaccino (preparato anch’esso come descritto in “Preparazione del campione”) che non contiene inibitori naturali. Letture spettrofotometriche I saggi, una volta bloccati con l’acido acetico sono stabili per circa una ora. Impostare una lunghezza d’onda di 405 nm. Prendere una cuvetta e riempirla con acqua distillata, evitando di toccare le pareti con le mani ed inserirla nello spettrofotometro. Azzerare lo spettrofotometro.

25
Effettuare in successione la lettura di tutti i saggi allestiti e riportare i valori di assorbanza nella seguente tabella.
1 (BT) 2 (T) 3 (B1) 4 (C1) 5 (Bn) 6 (Cn)
Bianco CONTROLLO CONTROLLO Bianco
CAMPIONE 1 CAMPIONE 1 Bianco CAMPIONE n
CAMPIONI n (eventuale)
Abs N.B. Di norma le assorbanze (A) non devono superare il valore di 1. Nel caso fossero maggiori di tale valore ridurre proporzionalmente il tempo di incubazione o il volume di tripsina. Al contrario, se il valore della lettura del saggio 2 risultasse inferiore a 0.2 aumentare il tempo di incubazione. ATTENZIONE: se si riducono o si aumentano i tempi di incubazione o le quantità di enzima tale la variazione deve essere effettuata per tutti i saggi allestiti (cioè non solo per quei saggi che risultassero al primo tentativo fuori scala). Calcoli Eseguire i calcoli della percentuale di inibizione utilizzando la formula:
Dove: i = percentuale di inibizione T = Assorbanza tripsina controllo BT= Assorbanza Bianco della tripsina controllo C1 = Assorbanza campione 1 B1 = Assorbanza Bianco del campione 1
(T-‐BT) -‐ (C1-‐B1) i = x 100
(T-‐BT)

26
CENNI DI SPETTROFOTOMETRIA La spettroscopia (o spettrofotometria) è una tecnica di analisi che permette di ricavare informazioni sulle proprietà strutturali dei corpi, studiando l’interazione della materia con l’energia elettromagnetica.
Quando una radiazione elettromagnetica interagisce con la materia si possono verificate tre fenomeni: riflessione, trasmissione e assorbimento, che si possono verificare tutti contemporaneamente. I metodi di analisi si basano sul fatto che la lunghezza d'onda delle radiazioni emesse o assorbite sono caratteristiche delle varie sostanze e ciò consente di effettuare analisi qualitative, inoltre l'intensità delle radiazioni emesse o assorbite dipendono dalla quantità di sostanza e quindi consente di effettuare analisi quantitative. La legge di Lambert-Beer si mette in relazione l’assorbanza con la concentrazione della soluzione; dalla formula si evince che l’assorbanza (A) è direttamente proporzionale alla concentrazione (C) della soluzione contenuta nella cuvetta. Quindi costruendo una retta di taratura, misurando l’assorbanza di quantità note di una sostanza, si può dedurre la concentrazione della stessa sostanza in un campione , misurandone l’assorbanza tramite uno spettrofotometro.
La legge di lambert-Beer è valida per soluzioni diluite la cui molarità M è inferiore a 0,01 mol/L. In questo modo è possibile utilizzare la spettrofotometria come tecnica di analisi qualitativa e quantitativa.
LINK UTILI
http://pdb101.rcsb.org/learn/videos/how-enzymes-work https://www.youtube.com/watch?v=ok9esggzN18 http://pdb101.rcsb.org/learn/videos/how-enzymes-work
ε è il coefficiente di assorbimento (estinzione) molare, è una grandezza che dipende dal tipo di solvente, dalla lunghezza d'onda utilizzata e dalla specie chimica che dà l'assorbimento;
-‐ l è il cammino ottico ovvero lo spessore della soluzione contenuta nella cuvetta Viene misurato in cm.
C è la concentrazione della soluzione contenuta nella cuvetta. La sua unità di misura è espressa in termini di molarità M (M = mol/L)
L’insieme di tutte le radiazioni elettromagnetiche costituiscono lo spettro elettromagnetico che copre tutto l’intervallo compreso fra le onde radio (λ ˃1 m) e i raggi cosmici (λ ˃10-13 m)

27
Attività di laboratorio che si possono svolgere a scuola Laboratorio 1 La Bromelina L’ananas è un frutto dalle particolari proprietà poichè contiene un enzima, la bromelina, dall'azione proteolitica che impedisce, ad esempio, la solidificazione delle gelatine di origine animale. Tale azione è paragonabile a quanto avviene nello stomaco ad opera della pepsina, enzima responsabile della digestione delle proteine. Materiali: 1 arancia 1 fetta d’ananas naturale 1 fetta d’ananas in scatola 1 busta di fogli di colla di pesce 4 capsule Petri 1 becker da 1L 1 becker da 50 mL 1 bacchetta di vetro 1 mestolo 3 coltelli 1 pennarello Piastra riscaldante Procedimento: 1. prendere la gelatina ed immergerla per 10 minuti in un becker da 50 mL con acqua fredda 2. tagliare l’arancia, l’ananas crudo e l’ananas in scatola in cubetti e metterli in quattro capsule Petri diverse,
utilizzando per il taglio coltelli differenti 3. scrivere su ogni capsula Petri il contenuto 4. trascorsi i 10 minuti, strizzare i fogli di gelatina e trasferirli in un becker contenente 200 mL di acqua
bollente 5.mescolare fino a completa solubilizzazione della gelatina, versarla nelle 4 Petri fino a riempirle 6.appoggiare le quattro capsule sul bancone ed attendere 5 minuti in frigorifero per accelerare il
raffreddamento 7.lasciar riposare per 40 minuti ed osservare il risultato. Osservare le capsule Petri e rispondere alle seguenti domande : a) In quali capsule Petri la gelatina è riuscita a solidificare? Perchè? b) In quali non ha solidificato? Perché?
GELATINA Arancia Ananas al naturale Ananas in scatola Controllo
SI
NO

28
Laboratorio 2 La pepsina
La pepsina è un enzima contenuto nel succo gastrico sotto forma di zimogeno, forma inattiva dello stesso enzima, che viene attivato in ambiente acido. Ha la proprietà di scindere le proteine in grossi frammenti peptidici (peptoni); la sua attività si svolge a pH variabile tra 1,6 e 4,4, secondo l'origine del l’enzima e la natura del substrato. La pepsina digerisce la maggior parte delle proteine, fatta ad eccezione della mucina, alcune varietà di protamine e la cheratina; inoltre le proteine denaturate vengono scisse più facilmente e rapidamente di quelle integre. La pepsina viene prodotta dalle cellule principali dello stomaco sotto forma di pepsinogeno, pro-enzima che si trasforma in pepsina in seguito a un processo provocato dall'acido cloridrico del succo gastrico o dalla pepsina stessa. La pepsina svolge un ruolo importante ma non indispensabile nella digestione delle proteine alimentari, perché anche i succhi intestinali sono in grado di scindere le proteine in peptoni.
Materiali:
Uovo, pepsina, acido cloridrico (HCl), termometro, becher, carta da filtro, acqua, fornellino, 4 provette, pentolino.
Procedimento:
Versare l’albume dell’uovo nel becher con acqua, scaldare la miscela, mescolando, fino ad ottenere un liquido lattiginoso. Filtrare liquido ottenuto e versarne un po’ in ognuna delle 4 provette.
Nella prima provetta aggiungere mezzo cucchiaino di pepsina e 5 gocce di HCl; nella 2° solo HCl; nella 3° solo pepsina e nella 4 non aggiungere nulla (controllo).
Scaldare le provette a bagnomaria nel pentolino fino a 37°C circa. Aspettare circa un’ora.
Cosa osservi? Come ti spieghi i risultati ottenuti?
Colore del liquido Pepsina+ 5 gocce di HCl HCl Pepsina Controllo
lattiginoso
trasparente

29
Laboratorio 3
La Catalisi L'ossigeno, elemento molto importante per la vita degli esseri viventi, può essere trasformato, durante i processi metabolici delle cellule, in composti molto reattivi e dannosi per le cellule. Una sostanza prodotta in tracce durante questi processi è il perossido di idrogeno ( H2O2), acqua ossigenata, che viene, però, prontamente trasformata e neutralizzata da reazioni chimiche che avvengono nelle cellule stesse. L’enzima catalasi o perossidasi accelera la reazione di decomposizione dell’acqua ossigenata in acqua (H2O) e ossigeno (O2) che si libera sottoforma di bolle. Quando versiamo alcune gocce di acqua ossigenata su una ferita,le cellule lesionate liberano molecole di catalasi che intervengono in massa a scomporre l’H2O2 : in questo modo si libera molto ossigeno allo stato nascente, attivissimo nell’ossidare le sostanze organiche con cui entra in contatto. Nel caso di una ferita l’ossigeno che si libera ossida, uccidendoli, i batteri introdotti insieme allo sporco: da qui proviene la sua azione antisettica (disinfettante). Materiali: Fegato crudo e cotto Mela Acqua ossigenata Idrossido di sodio(NaOH) Acido cloridrico (HCl) 5 provette Pipette pasteur Coltello spatola Procedimento: 1. Introdurre in tre provette circa 3 cm di fegato crudo, fegato cotto e mela. 2. In un'altra provetta inserire fegato crudo + NaOH e nell’altra fegato crudo + HCl 3. Aggiungere a tutte le provette 2-3 mL di acqua ossigenata 4. Osservare le reazioni. Domande: In quali alimenti è contenuto l’enzima catalasi? Per quale motivo lo trovi in queste cellule? Come hai fatto a capirlo? L’ambiente basico e quello acido favoriscono o inibiscono l’eventuale azione enzimatica?
Catalasi Fegato crudo Fegato cotto Mela Fegato crudo + HCl
Fegato crudo + NaOH
si
si

30
Laboratorio 4 IL VULCANO: PRESENZA DELLE PROTEINE NEGLI ALIMENTI
MATERIALE
Bicarbonato di sodio Aceto bianco 3 uova crude 1 bottiglia di plastica da mezzo litro a collo stretto 1 piatto fondo di plastica 3 bicchieri da tavola di plastica (180 ml) 3 bicchieri da tavola trasparenti di plastica (180ml) 3 bicchieri piccoli tipo per caffè di plastica (80 ml) Coloranti alimentari o colori ad acquarello (facoltativi) 1 cucchiaio da tavola 1 cucchiaino Alcool incolore per liquori Garza sterile o non a forma quadrata Acqua demineralizzata per stireria
Procedimento:
1. posizionate una bottiglia di plastica da mezzo litro vuota senza tappo a collo stretto, pulita e senza etichetta, al centro di un piatto fondo;
2. rompete due uova crude, separate gli albumi dell'uovo dai tuorli e versateli separatamente in due bicchieri da tavola di plastica;
3. versate l'albume nella bottiglia, aggiungete un cucchiaio da tavola colmo di bicarbonato di sodio (eventualmente aiutati con un imbuto);
4. agitate il contenuto della bottiglia per mescolare il bicarbonato di sodio e l’albume; 5. aggiungete (facoltativo) alcune gocce o più di colorante nel miscuglio bicarbonato-albume; 6. riempite 1 bicchiere piccolo di plastica quasi pieno di aceto bianco; 7. versate velocemente l'aceto nella bottiglia; si formerà il vulcano in eruzione!
Si formerà un’ intensa schiuma bianca oppure colorata (se avete aggiunto coloranti alimentari) che fuoriesce dalla bottiglia. Quando il vulcano si addormenta (fine dell'eruzione) provate a risvegliarlo (inizio dell'eruzione): versate velocemente nella stessa bottiglia un altro bicchiere piccolo quasi pieno di aceto bianco e agitate il contenuto per alcuni secondi con movimenti rotatori. Spiegazione L'aceto provoca con il bicarbonato di sodio una reazione chimica: la formazione dell'anidride carbonica e, quasi subito, anche la coagulazione dell'albumina, la proteina del bianco d'uovo, sotto forma di schiuma (coagulazione simile a quella che si ottiene quando il bianco d'uovo viene ripetutamente sbattuto meccanicamente). Il gas per effetto della pressione esercitata spinge fuori la schiuma.
Approfondimenti
La denaturazione delle proteine (coagulazione) comporta la distruzione di tutte le funzioni vitali delle cellule
sia animali sia vegetali; con il termine coagulazione si indica un liquido che sta per rapprendersi, o cagliarsi
(per es. la caseina, proteina del latte, quando coagula si separa sotto forma di grumi bianchi solidi).

31
Le proteine vengono denaturate (coagulate) in vari modi:
• con il calore: per es. l'uovo sodo o la crosta del pane non sono altro che proteine coagulate;
• meccanicamente: sbattendo l'albume si forma una soffice schiuma nevosa;
• chimicamente: nel nostro caso con l'acido acetico dell'aceto ma anche con altri composti chimici come
l'alcool.
L 'assunzione di alcool ha un effetto distruttivo sulle proteine del nostro organismo?
Sì, perché provoca la coagulazione delle proteine e con esse anche la distruzione delle nostre cellule.
Provate a fare questo semplice esperimento:
1. rompete un uovo crudo e separate l’albume dal tuorlo;
2. versate la metà dell’albume in un bicchiere da tavola di plastica trasparente;
3. aggiungete una quantità di 10 volte superiore di acqua demineralizzata, agitate energicamente (15-20
secondi) con un cucchiaino: si forma una sospensione albuminosa;
4. posizionate per esteso sul bordo di un bicchiere piccolo un doppio strato di garza;
5. debordate gli angoli della garza lungo il bicchiere e teneteli racchiusi con una mano per impedire alla
garza di cadere nel bicchiere;
6. filtrate la sospensione albuminosa attraverso la garza fino a riempire metà bicchiere; il liquido filtrato è
la nostra soluzione albuminosa;
7. mescolate in un altro bicchiere da tavola trasparente tutto il liquido filtrato con la stessa quantità di
alcool chiaro per liquori.
8. osservate contro luce la coagulazione (formazione di fiocchi bianchi di albume coagulato) dell'albume a
opera dell'alcool. Hai così verificato il motivo della pericolosità dell'alcool !
Hanno partecipato alla stesura della dispensa, sotto la supervisione di Giovanna Viale, Cinzia Grazioli e Livia Pirovano e Alesio Scarafoni: Prof.ssa Maria Luisa Erba Prof. Romualdo Esposito Prof.ssa Maria Teresa Oliveira Prof.ssa Ilenia Santi

















![00 Indice [V-X]...mour, Mark Knopfler, Robert Smith, Michael Stipe, Mike Mills, Paul Hewson, Dave Evans, Thom Yorke, Gordon Sumner, Steven Morrissey e Johnny Marr per essermi stati](https://static.fdocumenti.com/doc/165x107/5ea3ebd50cda7b51f95eb4b0/00-indice-v-x-mour-mark-knopfler-robert-smith-michael-stipe-mike-mills.jpg)
