
Ricerche microscopiche e submicroscopiche sull'area ... · mediante un meccanismo che risulta perb...
23
Zeitschrift f/Jr Anutomie und Entwicklungsgeschichte 123, 397~419 (1962) Istituto di Anatomiu Umana NormMe dell~ Universits di Puvi~ (Direttore: Prof. G. PALU~I) Ricerche microscopiche e submicroscopiche sull'area ematogena del blastoderma di pollo Di LuI(~I CATTANEO Con 16 Figure nel testo (Pervenuto alla redazione il 28 maggio 1962) Scopo del lavoro e riassunto bibliografico Esistono tuttora in merito all'area ematogena numerosi problemi non ancora chiariti o differentemente interpretati che riguardano non solo dettagli marginMi del complesso argomento, ma anche questioni di importanza fondamentale. Nel presente lavoro sull'area ematogena de1 blastoderma di pollo ho limitato lo studio ad alcuni di tall problemi ed ho particolarmente preso in esame: la derivazione e la modalit~ di formazione delle isole del sangue, la costituzione attorno a queste delle primitive pareti vasali, infine la struttura submicroscopica delle cellule ehe compongono le isole medesime. Espongo in breve iorma riassuntiva i dati bibliografici inerenti agli argomenti trattati nel corso di questo lavoro. Per quanto riguarda il primo punto, e ciob la derivazione delle cellule che nell'ambito dell'area ematogena danno luogo alle isole dcl sangue, il K6LLIKE~ (1882), il Rt~CK~RT (1888--1889) ed il vA~r DE~ ST~IC~T (1892) sostengono che esse prendono origine direttamente dM mesoderma. AnMoga veduta esprimono l'I-Iis (1900) ed il RUFF%~I (1913 e 1925), quantunque essi affermino trattarsi di cellule mesodermiche particolari gis a quel fine predeterminate; anzi, I'HIS designa il complesso di tMi cellule col home di angioblasto e ritiene che questo si addentri anche nell'embrione e lo compenetri interamente per formare in esso il sangue ed i vasi. La natura mesodermica delle isole del sangue ~ stata in linea di massima accettata dalla maggior parte degli Autori successivi (HILTO~r 1913; MILLER e McW]-IOl~TEI~ 1914; C~IA~UGT 1932; G~ODZI~S~:I 1933; STO~TI 1935; B~AUS ENGELHAI~DT 1940; t)~SA 1944; ecc.), ma mentre alcuni (WETZEL 1929; G~OD- ZI~CSKI 1934) ritengono che dette cellule derivino pifl propriamente dM materiale della linea primitiva, numerosi altri (D~CTSC~AKOFF 1907, 1908 e 1915--1916; STOCKAde) 1915; I~EA~r 1917; MAXI~OW 1927; ecc.) sostengono invece che esse traggono origine dal mesenchima. Altri l%icercatori infine, come il G~X~E~ (1907) che studi6 l'area em~togena degli uccelli e lo STOH~ jr. (1931) che esaminb quella degli anfibi anuri, riferiseono che le cellule delle isole del sangue sono di derivazione entodermica. Quest'ultima evenienza b ritenuta possibile, sia pure in misura alquanto limitata, anche da alcuni di quegli Autori (RffCKE~T e 1V[oLLIEI~1906; Pv,~SA 1944) che locahzzano la normale sede di origine del sangue nel mesoderma ; il P~s~ attribuisce clb alia indeterminatezza dei confini che inizialmente pub esistere tra l'entoderma ed il mesoderma.
Transcript of Ricerche microscopiche e submicroscopiche sull'area ... · mediante un meccanismo che risulta perb...

Zeitschrift f/Jr Anutomie und Entwicklungsgeschichte 123, 397~419 (1962)
Istituto di Anatomiu Umana NormMe dell~ Universits di Puvi~ (Direttore: Prof. G. PALU~I)
Ricerche microscopiche e submicroscopiche sull'area ematogena del blastoderma di pollo
Di LuI(~I CATTANEO
Con 16 Figure nel testo
(Pervenuto alla redazione il 28 maggio 1962)
Scopo del lavoro e riassunto bibliografico Esistono tu t tora in merito all 'area ematogena numerosi problemi non ancora
chiariti o differentemente interpretati che riguardano non solo dettagli marginMi del complesso argomento, ma anche questioni di importanza fondamentale. Nel presente lavoro sull 'area ematogena de1 blastoderma di pollo ho limitato lo studio ad alcuni di tall problemi ed ho particolarmente preso in esame: la derivazione e la modalit~ di formazione delle isole del sangue, la costituzione attorno a queste delle primitive pareti vasali, infine la s trut tura submicroscopica delle cellule ehe compongono le isole medesime.
Espongo in breve iorma riassuntiva i dati bibliografici inerenti agli argomenti t r a t t a t i nel corso di questo lavoro.
Per quanto riguarda il primo punto, e ciob la derivazione delle cellule che nell 'ambito dell 'area ematogena danno luogo alle isole dcl sangue, il K6LLIKE~ (1882), il Rt~CK~RT (1888--1889) ed il vA~r DE~ ST~IC~T (1892) sostengono che esse prendono origine direttamente dM mesoderma. AnMoga veduta esprimono l'I-Iis (1900) ed il RUFF%~I (1913 e 1925), quantunque essi affermino t ra t tars i di cellule mesodermiche particolari gis a quel fine predeterminate; anzi, I 'HIS designa il complesso di tMi cellule col home di angioblasto e ritiene che questo si addentri anche nell 'embrione e lo compenetri interamente per formare in esso il sangue ed i vasi. La natura mesodermica delle isole del sangue ~ stata in linea di massima accet tata dalla maggior parte degli Autori successivi (HILTO~r 1913; MILLER e McW]-IOl~TEI~ 1914; C~IA~UGT 1932; G~ODZI~S~:I 1933; STO~TI 1935; B~AUS ENGELHAI~DT 1940; t )~SA 1944; ecc.), ma mentre alcuni (WETZEL 1929; G~OD- ZI~CSKI 1934) ritengono che dette cellule derivino pifl propriamente dM materiale della linea primitiva, numerosi altri (D~CTSC~AKOFF 1907, 1908 e 1915--1916; STOCKAde) 1915; I~EA~r 1917; MAXI~OW 1927; ecc.) sostengono invece che esse traggono origine dal mesenchima. Altri l%icercatori infine, come il G~X~E~ (1907) che studi6 l 'area em~togena degli uccelli e lo STOH~ jr. (1931) che esaminb quella degli anfibi anuri, riferiseono che le cellule delle isole del sangue sono di derivazione entodermica. Quest 'ult ima evenienza b ri tenuta possibile, sia pure in misura alquanto limitata, anche da alcuni di quegli Autori (RffCKE~T e 1V[oLLIEI~ 1906; Pv,~SA 1944) che locahzzano la normale sede di origine del sangue nel mesoderma ; il P ~ s ~ attribuisce clb alia indeterminatezza dei confini che inizialmente pub esistere t ra l 'entoderma ed il mesoderma.

398 Lurer C2~TTANEO :
La modalitg di formazione delle isole del sangue 6 stata illustrata partieolar- mente dal R v s s I ~ I (1913 e 1925) e dal MAxI~OW (1927), ma in modo net tamente differente. I1 RVSFI~r afferma ehe il mesoderma dell'area ematogena h inizial- mente aderente all 'ectoderma, ma che poi si sposta sull 'entoderma dove le sue eellule si stratifieano in pih file ed inglobano sferule di vitello ; eompaiono allora in esso numerose cavits seriate che dilaminano la sua parte superficiale nella somato- pleura e nelia splancnopleura e causano contemporaneamente la disgregazione della restante sua parte helle isole del sangue, le quali si t rovano eosl comprese fra ]a splanenopleura e l 'entoderma. I1 MAXIMOW sostiene invece che cellule mesen- ehimah migrano dal eontorno periferieo del mesoderma embrionale nell 'area emato- gena dove si organizzano nelle isole del sangue ; in un seeondo tempo, al di sopra di queste si forma ]a splanenopleura ehe diventa per esse sorgente di nuove cellule mescnehimali. Secondo il MAxI~ow le isole neoformate sarebbero collegate da ponti di eellule fusate, secondo il Bt~EMER (1914) inveee sarebbero direttamente congiunte fra loro per le estremit~.
Anehe le opinioni sulla formazione delle primitive p~reti vasali attorno alle isole del sangue sono discordanti. I1 RUFFI~I (1913 e 1925) infatti afferma ehe nel blastoderma di pol]o dette pareti sono inizia]mente costituite dalla splancnopleura, mediante un meccanismo che risulta perb oscuro; il MAxI~ow (1927) sostiene invece, sempre a proposito del blastoderma di pollo, ehe esse sono formate dalle cellule periferiehe delle isole del sangue; lo STOCKAt~D (1915) infine relativamente all 'area ematogen~ dei teleostei, il FS.D~RICI (1926) ed il Goss (1928--1929) per quella degli anfibi ed il GRODZINSKI (1934) per quella degli ueeelli riferiscono the le pareti vasali originano da una trasformazione endoteliale delle cellule mesen- ehimali poste in prossimit~ delle isole del sangue. Le isole del sangue incluse nei vasi neoformati si disgregano, seeondo molti Autori (MAxIMow 1927; CI~IA~UG~ 1932; STO~TI 1935; ecc.), per il formarsi in esse di vacuoli eontenenti plasma sanguigno elaborato datle loro stesse eellule, seeondo il I~UrFI~I (1925) inveee per il progressivo instaurarsi della funzionalith del euore che determinerebbe una corrente di liquido sempre pifi forte entro i vasi; (a questo proposito 6 da ricordare che ]e rieerche dell 'OHvo [1925] sull'embrione di pollo dimostrarono che ,,la fun- zione eardiaea si istituisce gradualmente ed 6 certo che nelle prime ore l 'effetto meccanieo 6 assai scarso").
Per quanto infine si riferisce a]l 'ultrastruttura delle eellule delle isole del saugue mancano attualmente, a quanto mi consta, ricerche in proposito.
Materiale e tecniche
I1 presente lavoro ~ stato compiuto complessivamente su quarantotto blasto- dermi di po]lo (Gallus bankiva L., domestico), prelevati d a uova incubate a 380--390 C.
Per le riccrche di microscopia luce, i blastodermi sono stati raceolti tsa la 14 a e la 44 ~ ora di incubazione, di ora in ora per gli stadi pifi precoci, ciog fino alla 24 a ora, e di due ore in due ore per gli stadi pifi avanzati. I1 prelicvo dei blastodermi, perchg questi rimanessero perfet tamente distesi, 6 stato ia t to secondo 1~ metodica consigliata dal RvSFINI (1927) che eonsiste, come noto, nell 'appoggiare sul tuorlo dell'uovo, attorno al blastoderma, un anellino di carta paraffinata entro cui viene messo il liquido fissatore che conferisce cosi una certa consistenza al blastoderma mcdesimo prima ehe questo venga rimosso mediante l'uso di una spatola. I blasto-

t~icerche microscopiche e submicroscopiche sull'area ematogena del blastoderma 399
dermi sono stati fissati con uno dei seguenti liquidi: 1% 3 seguito da trattamento con KMserling II , Susa, Carnoy; sono stati inclusi in paraffina previo passaggio in metilbenzoato-eelloidina e poi sezionati in serie, quelli degli stadi pifi preeoci totalmente, quelli pih avanzati invece limitatamente all'urea ematogena. Le sezioni seriate sono state colorate in parte con emMlume-eosiua ed in parte con ematossilina ferrica. Sono anche stati allestiti preparati Iissati e colorati con acido osmico per studiare la distribuzione delle sferule di vitello; nouchg preparati fissati in formMina neutra e colorati con la reazione al bleu di Prussia di PE~LS per la dimostrazione in queste ultime del ferro trivMente.
Le osservazioni di microscopia elettronica sono state effettuate su blastodermi della 24 a, 26 a', 32 a e 40 ~ ora di incubazione, fissati per un'ora circa con soluzione di acido osmico MI'1% tamponata a pH 7,5--7,8. Anche in questo caso i blasto- dermi sono stati fissati sul tuorlo dell'uovo prima del loro prelievo, seeondo la gi~ ricordata metodica del RvFFI~I; durante la fissazione le uova sono state mantenute in frigorifero alla temperatura di 40 C. I pezzi sono stati inclusi in metacrilato e sezionati con l 'ultramicrotomo LKB rood. ,,Ultrotome". Le sezioni, aleune delle quMi colorate con soluzione satura di aeetato di uranile, sono state esaminate e fotografate con microscopio Zeiss EM 8 e con microscopio Siemens Elmiskop I.
0sservazioni personali
1. Processi evolutivi dell'area ematogena
Premetto che i fatti da me rilevati sulla formazione delle isole del sangue e dei primitivi vasi si discostano da quanto ~ riferito dalla maggior parte dei gicerca- tori, tra i quMi il MAxI~ow; essi sono invece fondamentMmente in accordo con le osservazioni del I~UFFI~I, osservazioni che purtroppo sono rimaste pressochb ignorate dagli Autori che si occuparono dell'argomento. Pertanto, anche se alcuni dei fatti non saranno del tutto nuovi, ne dar5 pur tuttavia una descrizione abba- stanza dettagliata pereh~ cib servirs oltre che a dimostrare l'esattezza dei dati del gUFrI~I, ad ampliare questi e a d integrarli in molti punti, nonchg a rendere note le diverse eta del blastoderma in cui i vari fenomeni si verfficano, dato che il t~UFFI~ ds di essi una descrizione globMe senza riferire specifieatamente in quMi stadi essi si instaurano.
Nella descrizione che segue mi rfferirb sopratutto a quanto si osserva nelle sezioni seriate trasversMi; inoltre, poichb specie nei primi stadi i processi evolutivi dell'area ematogena si instaurano dapprima nella zona posta subito dietro la linea primitiva e pih tardivamente in quella perfferica, ove necessario dirb separata- mente di queste due zone.
Nei blastodermi della 14 a e 15 a ora di incubazione (nei quMi la linea primitiva si estende circa per la met~ posteriore dell'area chiara) la parte di area opaca retrostante la linea primitiva appare costituita (fig. 1): in superficie, dMl'ecto- derma formato da uu'unica fila di cellule; subito sotto, dal mesoderma che ancora molto arretrato nel suo sviluppo ed ~ semplicemente rappresentato da cellule rotondeggianti immediatamente aderenti alia superiicie inferiore dell'ecto- derma; ancora pi~ profondamente e separato dallo strato precedente da uno spazio relativamente ampio, dall'entoderma che ~ costituito da una fila di cellule adagiate sul vitello. I tre foglietti aderiscono fra loro in corrispondenza dell'orlo germinativo
400 LUIGI CATTANEO :
.~ ............... .............. ,-o.,-~~'!e~Q:-@;;~;~~i'~;; l':...:i~ ;~~;~";;;~
~. e e . �9 � 9 4 , ", .e. ,~.,,, . ..... [ . . . . . . �9
e/7 ................................ ;....... ,:.....,,~ ..... ,...,. ? w
3
~ I .~; ~-'.~ .~. ~.;~:~;~.~. ....
" -i]. �9 ...................... ~::~. ~ ;..~,,;;~:7 ~; .....
C,S
~i ."~ ~.....,~'-..~. [~.., ..... 'i .... ~::"~,~ r.., %~ ............ ,.-~ ~.~;.' e :6~-: e �9 ~,
[~.;~i~" ~ ~' '~ ~ .",d~ '"'=~,~-,~;: :~':,'.~; ..........
r..":" :; e*. 0 ~ Wo ~ .;~-~! ..... '~I.=~; ................ .................. ~ ................... ;;' . .o~..~ ..........
I"~ ................ ; ...... ~ o" ~;
.." ~ ' " : : ~ ! : 72~ -~ ' ..... / ' . .
~.I" . - . ' . ; " ; . ~ .... ~, �9 I...e ~ .e "...e "?---.,-.-.-, ............... :: �9 .. ~ /
7 .......... "t ............. !;....e.*~ ........... '
7.. '~~ ~~.~ . , ~
~ ,~2
~ o �9 �9 o ~ e ~ �9 �9 �9
*.e.~:ie;~:~; O i ~ , -..,~ - . . ~ d,.~ ........ '. .......
6
~ . a ~ ~ ~.~......, . = , . , ; ~ . :~ . .~ . . ~...~..,~ . . . . .
......... . . . . . . . . . ~ ~ � 9 . ' " ~ Q . . . ~ � 9 ~ ........ ., . ~ . ~ ~Q., .;:~.~,O" - : , m \ . O ! ~ : ~ . ~
~ - : . �9 �9 - ~ v . - 0 ~ " ~ - . ~ . ,
,. ....... �9 ~. . ;~ . . .~ . , )~ ,~ i~ - ...... ..~:;:~:.i @
........ ;"".... ........... . @ i.;~*~0 + O i ",. ~ ' . ....
6
eC
en
eC
C5'
el/
~C SO
C sp
i
e/7
ec 5o
c
sp
C/7
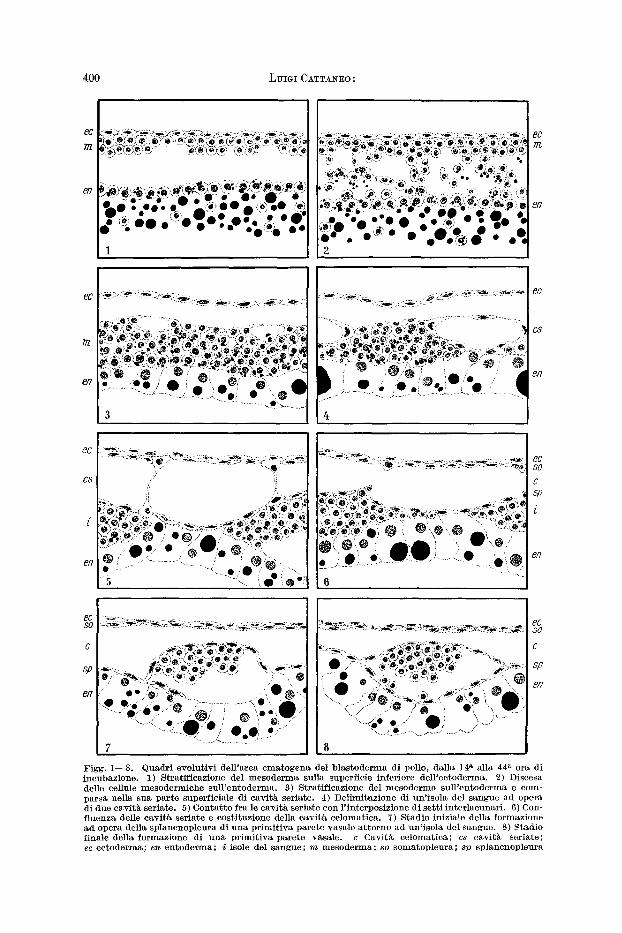
Figg . 1 - -8 . Quadr i evo lu t lv i del l 'a rea e m a t o g e n a del b l a s t o d e r m a di polio, dalla 14 �9 al ia 44 a e ra di incubazione. 1) St ra t i f icaz ione del m e s o d e r m a sulla superf icie infer iore de l l ' e c tode l~a . 2) Discesa delle cellule mesode rmiehe S~lll'entodel~na. 3) S t ra t i f icaz ione del m e s o d e r m a su l l ' en tode rma e com- pa r sa nella sua pa r t e superf ie ia le di cav i t~ ser iate . 4) Del imi taz ione di un ' i so la del s angue a4 opera di due cav i t~ seriate. 5) Conta t to f ra le cavit& ser ia te con l ' in terposiz ione di se t t i in ter lacunar i . 6) Con- f luenza delle cav i t~ ser ia te e eost i tuzione delia cav i t~ celomat iea . 7) S tadio iniziale della formazione ad opera della splancnopleura di u n a p r i m i t i v a pa re te vasa le a t to rno ad ~n ' i so la de1 sangue. 8) Stadio f inale della form~zione di n n a p r i m i t i v a pa re te vasale , c Cavi t~ ce lomat ica ; c s cav i t~ ser ia te ; e c ee t o d e r ma ; e n e n t o d e r m a ; i isole de1 sangue ; m m e s o d e m a ; s o somatop leu ra ; s p sp lancnopleura

Ricerche microscopiche e submicroscopiche sull'area ematogena del blastoderma 401
e determinano eosi la ehiusura, sui lati, dello spazio interposto f ra i l mesoderma e l 'entodenna.
Nei blastodermi prelevati alla 16 a ora di incubazione lo strato mesodermieo ispessito a eausa della spiceata attivit~ mitotiea delle sue eellule. Inoltre, lo spazio interposto Ira il mesoderma e l 'entoderma contiene sferule di vitello ehe sono sopratut to abbondanti in vieinanza dell'orlo germinativo.
Nei blastodermi della 17 a ora di ineubazione (nei quali il nodo di HENSEN g in via di formazione all 'estremit~ eefaliea della linea primitiva) lo spazio interposto f r a i l mesoderma e l 'entoderma g oeeupato, oltre ehe da pieeole sferule di vitello, da numerosissime eellule singole o riunite in gruppetti, in eontinuazione da un lato con quelle dello strato mesodermieo applieato all 'eetoderma e dall 'altra con cellule di analogo aspetto che sono stratifieate sul foglietto entodermieo (fig. 2); tali eellule presenti nello spazio in parte hanno forma rotondeggiante, in parte inveee sono fornite di aleuni prolungamenti ehe, almeno apparentemente, le fanno assomigliare agli elementi mesenehimali; molte di esse inoltre eontengono nel eitoplasma sferule di vitello. Tut to eib b evidente presso l 'estremit~ eaudMe della linea primitiva; nella parte periferiea dell 'area opaea inveee, lo spazio sovraento- dermico b libero da eellule ed il mesoderma permane sotteso all 'eetoderma. Alla 18 a e 19 ~ ora di incubazione, le immagini ora deseritte si ripetono anehe alla peri- Ieria dell 'area opaca. I1 fat to dunque sostenuto dal RUFFI~I dello spostamento del mesoderma dMl'ectoderma sull 'entoderma corrisponde pienamente a realtY.
Nei blastodermi prelevati alla 20 a ora eli incubazione (nei quali la eorda dorsale b rieonoscibile eranialmente al nodo di H~NSl~N) il foglietto ectodermico ~ rimasto in superficie completamente solo; il mesoderma si b infatti ricostituito al di sopra dell 'entoderma e risulta di pifi file di cellule s tret tamente stipate, le quali contengono inclusion di vitello e sono di forma globosa, tranne nello strato superfieiale dove in genere sono appiatt i te; t ra l 'eetoderma ed il mesoderma b interposto uno spazio privo sia di eellule che di sferule di vitello. L 'entoderma infine b eostituito ora da un'uniea fila di eellule molto voluminose eontenenti grosse sferule vitelline.
Nei blastodermi della 21 a ora di incubazione, in vicinanza dell 'estremit~ eaudale della linea primitiva, alcune delle cellule pifi superficiali del mesoderma, clog prossime allo spazio sottoeetodermico, si differenziano dalle Mtre per avere un eorpo di forma fusata fornito eli prolungamenti e per essere disposte in modo tale da delimitare pieeole fessure irregolari oppure lacune un po' pifi ampie di forma ovalare con asse maggiore parallelo alla superficie del mesoderma stesso (fig. 3); in eorrispondenza degli angoli di queste eavit~ spesso si t rovano cellule che con le loro espansioni parteeipano alla costituzione sia della parete rivolta verso lo spazio sottoectodermico che di quella applieata alla massa mesodermica. Nei blasto- dermi prelevati alla 22 a ed alla 23 a ora di incubazione, le lacune presenti nel meso- derma vieino Mla linea primitiva si presentano pifi voluminose; esse fanno in alto lieve sporgenza entro lo spazio sottoeetodermico ed in basso sono aHondate nell 'am- masso mesodermico ehe pertanto, in loro eorrispondenza, si presenta assottigliato; nuove pieeole laeune sono pure in eorso di differenziazione alla sommit/~ delle zone pifi spesse del mesoderma. Anche alla periferia dell 'area ematogena sono ora scavate, nella parte superfieiale del mesoderma, laeune analoghe alle preeedenti. I reperti rivelano quindi che le lacune in questione hanno un rapido e notevole sviluppo per cui in breve volgere di tempo, aumentando eli numero ed espanden- dosi, invadono tu t to il mesoderma dell 'area ematogena; esse, per tali loro caratte-

402 Lvmi CATTAN~O"
ristiche di aspetto e di comportamento, sono senz'altro da identifiearsi con le eavitg seriate descritte dal RUFFINI.
Nei blastodermi della 24 a era di incubazione (nei quali 6 gig presente il primo pale di somiti) le cavitg seriate ingranditesi hanno raggiunto inferiormente l 'ento- derma (fig. 4); pertanto, il mesoderma si presenta scomposto in numerosi ammassi cellulari intervallati dalle oavitg medesime: sono le primitive isole del sangue o isole di WOLFF. Le pareti delle cavitg seriate, ohe sono formate da alcune cellule appiatt i te ed allungate, aderiseono in basso alle isole del sangue lore contigue e, nell'intervallo Ira queste, all 'entoderma; in alto fanno invece sporgenza nello spazio sottoectodermico. Le cellule delle isole del sangue sono ancora caratterizzate dalla presenza nel lore citoplasma di inclusioni di vitello.
Nei blastodermi della 26 a era di incubazione (nei quali sono presenti 2- -3 paia di somiti) le cavitg seriate, essendo ulteriormente aumentate di grandezza, si pre- sentano a eontatto fra lore entre lo spazio sottoectodermico; quelle poi tra esse che hanno assunto rapporti pih estesi risultano separate solo da sottili setti cellu- lari appiattiti , disposti vertiealmente (fig. 5). Quest 'ultimo quadro 6 particolar- mente evidente nei blastodermi della 28 a era di incubazione (con 6--7 paia di somiti) ; in questi anzi, a livello dell 'area ematogena prossima all 'area embrionale, si riscontra gig una involuzione di tali setti con eonseguente confluenza delle relative cavitg seriate in un'uniea eavitg (fig. 6).
Nei blastodermi raccolti verso la 30 a era di ineubazione (nei quali vi sono in genere 8 paia di somiti) l'involuzione dei setti t ra le cavitg seriate 6 pressoeh6 totale; si riscontra pertanto un'unica grande cavitg circoscritta in alto da una lamina di cellule appiattite, risultante dalla unione delle pareti superiori delle singole eavitg seriate, ed in basso da un 'al t ra lamina pure di cellule appiatt i te ehe, essendo formata dalle pareti inferiori delle cavitg seriate medesime, aderisce alle isole del sangue e, negli intervalli ira queste, all 'entoderma. Come dimostrano sia le sezioni trasversali ehe le longitudinali dei blastodermi in esame, tali due lamine cellulari che circoscrivono la cavitg neoformata sono in diretta oontinuitg rispetti- vamente la superiore con la somatopleura e l'inferiore con ]a splanenopleura dell 'area embrionale ; esse sono quindi la somatopleura e la splancnopleura dell'area ematogena e pertanto l ' interposta cavitg rappresenta la cavitg celomatica dell 'area ematogena medesima.
Dalla successione dei fatt i finora esposti risu]ta dunque the il mesoderma del- l 'area ematogena dg origine fondamentalmente in superficie alla somatopleura ed alla splancnopleura, mentre con tu t t a la restante parte forma le isole del sangue.
I~ei blastodermi della 32 a e della 34 ~ era di ineubazione (aventi 9--10 paia di somiti) la somatopleura ~ applicata alla superfieie inferiore dell 'ectoderma, mentre la splanenopleura permane aderente alle isole del sangue e, ira queste, all'ento- derma. Le isole del sangue, le cui eellule si presentano era prive di inelusioni vitelline, appaiono fra lore distanziate maggiormente che negli stadi preeedenti; inoltre, molte di esse non sono pih stret tamente applicate all 'entoderma, m a n e sono separate per l'interposizione di una cavitg, forse causata da un proeesso di secrezione. (Per alcune isole questo fatto era gig rintraeciabile, almeno parzial- mente, in precedenti stadi.) Lg dove le isole sono net tamente staccate, la splaneno- pleura prolilera nei punti in eui si solleva per rivestirle e dg eosi origine a eellule

Ricerche microscopiche e submicroscopiche sull'area ematogena del blastoderm~ 403
lamellari ehe si portano al di sotto delle isole medesime, aderendo all'entoderma (fig. 7); dove poi tali eellule neoformate gig eostituiseono uno strato eontinuo sotto le isole, queste ultime risultano raeehiuse entro eavitg le eui pareti sono formate interamente dalle eellule appiattite della splanenopleura (fig. 8). Queste cavitg neoformate sono i primitivi vasi sanguigni dell'area ematogena; le isole del sangue ehe vi sono contenute sono ancora eompatte e stanno strettamente applieate alla
parete superiore, ]aseiando libera la parte inferiore del lume.
Nei blastodermi raceolti tra la 36 ~ e la 44 a ora di ineubazione (ton 10--14 paia di somiti) si vede ehe tutte le isole del sangue restano via via incluse entro nuovi vasi ehe si formano seeondo le modalitg ora deseritte. Lg dove il processo di Iormazione delle primitive pareti vasali si ~ gig espletato, la splanenoplenra appare come una lamina di eellule appiattite, sdoppiata ad intervalli per avvolgere le isole del sangue. Delle isole intluse nei vasi neoformati, aleune sono ancora eom- patte; altre permangono tali solo nella loro parte alta che aderisce al tontorno superiore della parete vasale, mentre nella parte inferiore, sporgente nel lume, presentano piceole fessure tra le tellule che allontanano queste Ira loro; si ha quindi l'impressione ehe il distaeco e la migrazione delle eellule ematiehe avvenga prima dal lato inferiore dell'isola, ore forse si raccoglie entro il vaso neoformato il plasma setreto in loeo o ehe vi giunge da altra parte. Altre isole infine sono pressoeh& eompletamente sgretolate e le loro eellule sono sparse nel lume vasale; tale infatti ~ il quadro ehe si osserva specie negli stadi pitt avanzati.
Da quanto g stato detto, risulta quindi the i primi vasi dell'area ematogena sono formati dalla splancnopleura e ehe essi inizialmente sono delimitati da un unieo strato di eellule appiattite; risulta inoltre ehe le isole del sangue ineluse nei vasi neoformati dapprima sono antora eompatte e stanno aderenti al eontorno superiore delle pareti vasali e ehe in seguito, per l'instaurarsi di pieeoli spazi nella loro eompagine, a tomineiare dalla parte inferiore sporgente nel lume si seompon- gono in eellule singole ehela eorrente plasmatica trasporterg nel eireolo.
2. Ricerca istochimica del /erro helle s/erule di vitello
t to gig fatto notare che le eellule mesodermiehe dell'area ematogena, per un eerto periodo della loro evoluzione, presentano nel eitoplasma numerose inclusioni di vitello.
Questo fatto, gig osservato dal GR/ieE~ (1907) e dal I~UFFINI (1913 e 1925), fu da quest'ultimo Autore attribuito ad una eessione di vitello da parte delle eellule entodermiehe a quelle mesodermiche allorquando queste ultime si stratifitano sulle prime; come si g visto per6, le mie osservazioni dimostrano in proposito ehe le eellule del mesoderma gig prima comineiano ad assumere vitello, quando nel trasmigrare dall'ectoderma sull'entoderma attraversano l'interposto spazio essendo in questo eontenute numerose sfere viteliine ectopiehe the vi sono entrate dalla zona dell'orlo germinativo. I1 mesoderma dell'area ematogena dunque ha tra le sue primarie attivitg quella di inglobare il vitello in notevole misura.
Sembrandomi ehe una eosl abbondante assunzione di vitello ad opera di eellule destinate nella maggior parte a diventare elementi ematici non potesse essere tausata dal solo bisogno di nutrimento come fu affermato dal lgVFFI~I, e eon-
Z. Anat. Entwickl. Gesch., ]3d. 123 28

404 LuIGI CATTANEO ."
siderando ehe nel tuorlo dell 'uovo 6 presente anche il ferro 1, ho supposto ehe il proeesso di assunzione del vitello potesse essere in relazione con la neeessit~ the tall eellule hanno di proeurarsi il ferro per la fu tura sintesi del l 'emoglobina; ho quindi r i tenuto anz i tu t to oppor tuno rieereare is toehimieamente il ferro entro le sferule vitelline ~. Poieh6 6 noto dai lavori dell'HILL (1931) e del TOMPSST~ (1934) ehe il ferro presente nel l 'uovo 6 allo stato tr ivalente, per la sua dimostrazione h o f a t t o uso della reazione di PERLe (con il ferroeianuro di potassio) ehe 6 spe- cifies per il ferro ferrieo, eseguendola su sezioni di blastodermi ehe erano stat i fissati con formalina neu t ra e prelevati eonservando anehe una parte del sotto- s tante vitello. Orbene, i preparat i eosl allestiti d imostrano ehe tu t to il vitello, eompreso quello intraeellulare, assume la t ipiea eolorazione verde-azzurra del bleu di Prussia ehe indiea la presenza in esso del ferro tr ivalente. Tali preparat i farebbero perb supporre ehe il ferro non abbia una distr ibuzione uniforme entro il vitello ; infa t t i le sfere vitelline, anche in una medesima sezione, non sono tu t te reat t ive in uguale misura, essendo aleune pih eolorate ed altre meno, e most rando inoltre nel loro stesso ambi to zone di differente reatt ivi t~.
3. Studio ultrastrutturale delle cellule delle isole del sangue
Come gis premesso, sono state s tudiate al mieroseopio elettronico le isole del sangue di blastodermi prelevati alla 24~,alla 26 a, alla 32 a eal la 40 a ora di ineubazione.
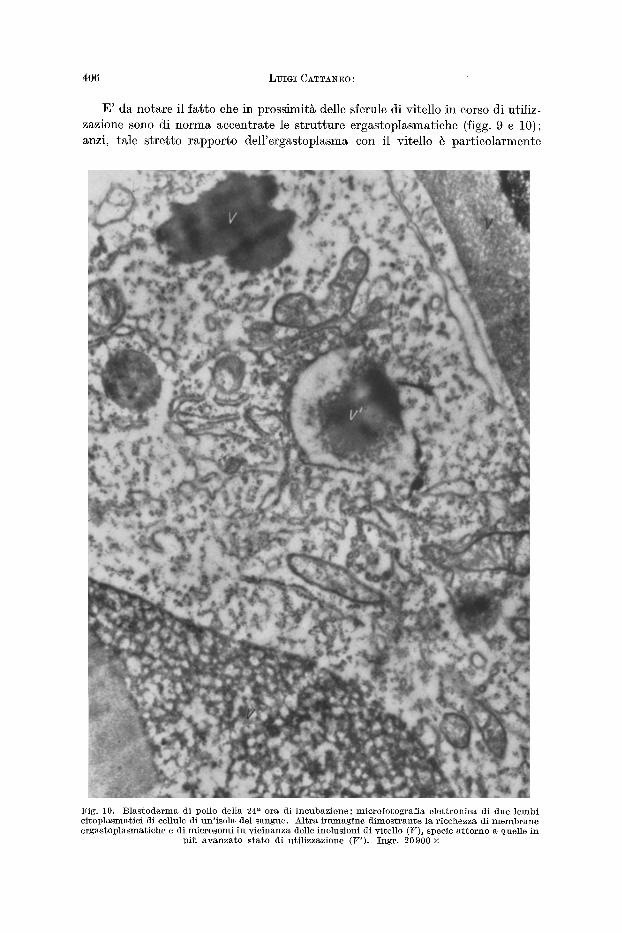
I1 citoplasma delle eellule delle isole del sangue dei blastodermi della 2 4 ~ 2 6 a ora di ineubazione, helle mierofotografie elettroniehe, appare ehiaro, eosparso di mierosomi e presenta molt i mi tocondri e numerose s t ru t tu re ergastoplasmatiche. lVia eib ehe fondamenta lmente caratterizza il eitoplasma di tall eellule g la pre- senza in esso di inelusioni vitelline.
Le inelusioni vitell ine (figg. 9 e 10) si presentano come masse osmiofile non omogenee, ma lormate di mater ia l i di differente densit~ var iamente dis t r ibui t i ; appaiono inoltre w ive di membrane l imi tan t i ehe le eireoserivino dal resto del ei toplasma a. Le inelusioni di vitello sono molto differenti Ira loro sia per dimensioni ehe per aspetto ; infat t i , accanto ad aleune voluminose e eompat te con forma sferica e contorni regolari, se ne t rovano moire altre che hanno eseavazioni marginal i pifl o meno prolonde, oppure ehe presentano nella loro eompagine zone di rarefazione o di vacuolizzazione, ta lune ampie e eireoscritte ad una sola par te del l 'ammasso vitellino, talal t re pieeole e numerosissime ehe danno alle sferule medesime quasi un aspetto spugnoso; si r iscontrano inoltre alcune masserelle di vitello molto piccole con eontorni irregolari e vaeuolizzate. In te rpre to queste immagin i di erosioni marginal i e di vaeuolizzazione in te rna come l 'espressione morfologiea del
1 La quantits di ferro eontenuta in gr. 100 di tuorlo di uovo di gallina sarebbe di rag. 6,13 seeondo i da~i di McCANcE e WIDDOWSON (1948), sarebbe invece di mg. 7,20 in base ai dati pubblieati dall'Isti~uto Nazionale dell'Alimentazione di ROMA (1961). Seeondo I'HILL (1931) ed il TOMt'SETT (1934) il ferro ~ presente nel tuorlo dell'uovo in forma inorganiea, forse come idrato ferrico colloidale; anche per il LENTI (1939) ~ SOttO forma inorganica; seeondo il MeFARLA~E (1932) invece, sarebbe in combinazione organiea.
2 E' nota dalla Letteratura (WAssE~A~ 1910--1911; V. D. MA~ZA, E. 2r e CIIIOSA 1932; BERTtIIER 1948) la possibilits di dimostrare istoehimieamente il ferro entro il vitello.
3 In base a ricerche di microscopia elettronica eondotte su differenti territori dell'area embrionale del blas~oderma di pollo, alcuni Autori (BELLAIRS 1958; BALINSKY e WALTtIER 1961) riferiscono della possibile esistenz~ di una membrana attorno a taluni inclusi vitellini presen~i helle cellule.

l~icerche microscopiche e submicroscopiehe sull'area ematogena del bl~stoderma 405
processo di riassorbimento del vitello nelle sue varie fasi di attuazione e ritengo pertanto ehe le masserelle pih pieeole, irregolari e vacuo[izzate, rappresentino il residuo di sferule vitelline quasi eompletamente riassorbite.
Fig . 9~ B l a s t o d e r m a di pol io de l ia 24 s~ o ra d i i n c n b a z i o n e ; m i e r o f o t o g r a f i ~ e l e t t r e n i c g di u n a p o r z i o n e d[ c i t o D l a s m a eli u n a ce l lu la d i u n ' i s o l a deI s a n g u e . A d d e n s ~ m e n t o d i e l e m e n t i e r g a s t o I ) l a s r n a t i c i loca liz- z a t i t r a i n c l u s i o n i d i v i t e l lo (V) i n co r so d i u t i l i z z a z i o n e ; ne l l a p a r t e p e r i f e r i c a de[ v i t e l lo sono e e c u m u l a t i
g r a n u l i d i f e r r i t i n a ( f recce) . I n g r . 29 600 •
28*
4 0 6 L u I G I r :
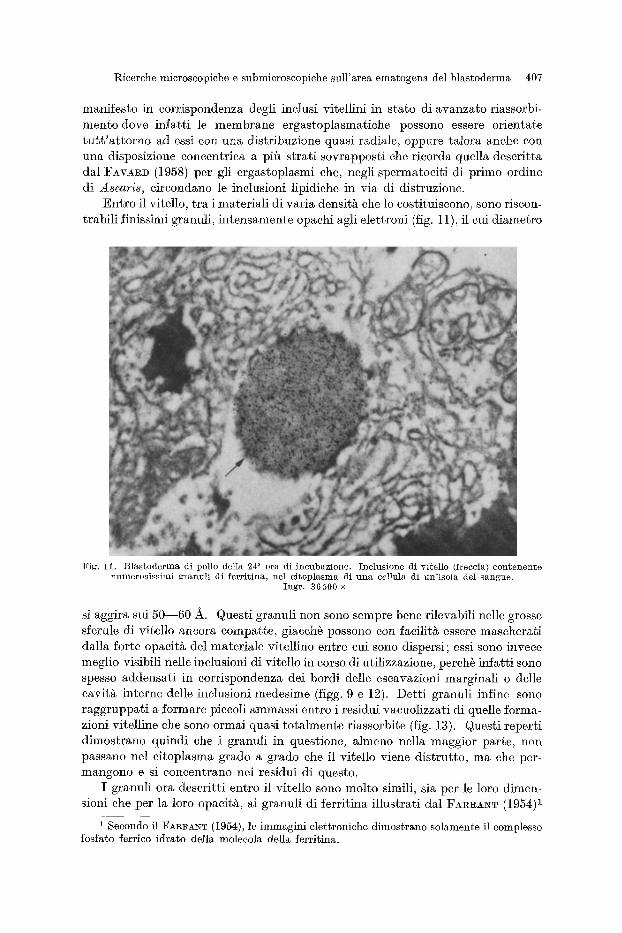
E' da notate il fatto che in prossimit~ delle sferule di vitello in corso di utiliz- zazione sono di norma accentrate le strutture ergastoplasmatiche (figg. 9 e 10); anzi, tale stretto rapporto dell'ergastoplasma con il vitello 5 particolarmente
F i g . 10. B l a s t o d e r m a di pol lo de l ia 24 a o r a di i n c u b a z i o n e ; m i c r o f o t o g r a f i a e l e t t r o n i c ~ d i d u e l e m b i citoplasmatici di eelluIe di un'isola del sangue. Aftra immagine dimostrante la rieehezza di membrane ergastoplasmatiche e di microsorai in ~rieinanza deIIe inclusioni di vitello (V), specie attorno a quelle in
pilk a v a n z a t o s t a t o d i u t i l i z z a z i o n e (V'). I n g r . 20900 •
I~ieerche microscopiehe e submicroscopiche sull'area ematogena del blastoderma 407
manifesto in corrispondenza degli inclusi vitellini in stato di avanzato riassorbi- mento dove infatt i le membrane ergastoplasmatiehe possono essere orientate t u t t ' a t t o rno ad essi con una distribuzione quasi radiale, oppure talora anehe con una disposizione eoneentriea a pifi strati sovrappost i ehe rieorda quella deseritta dal FAVA~D (1958) per gli ergastoplasmi ehe, negli spermatoeit i di primo ordine di Ascaris, eireondano le inelusioni lipidiehe in via di distruzione.
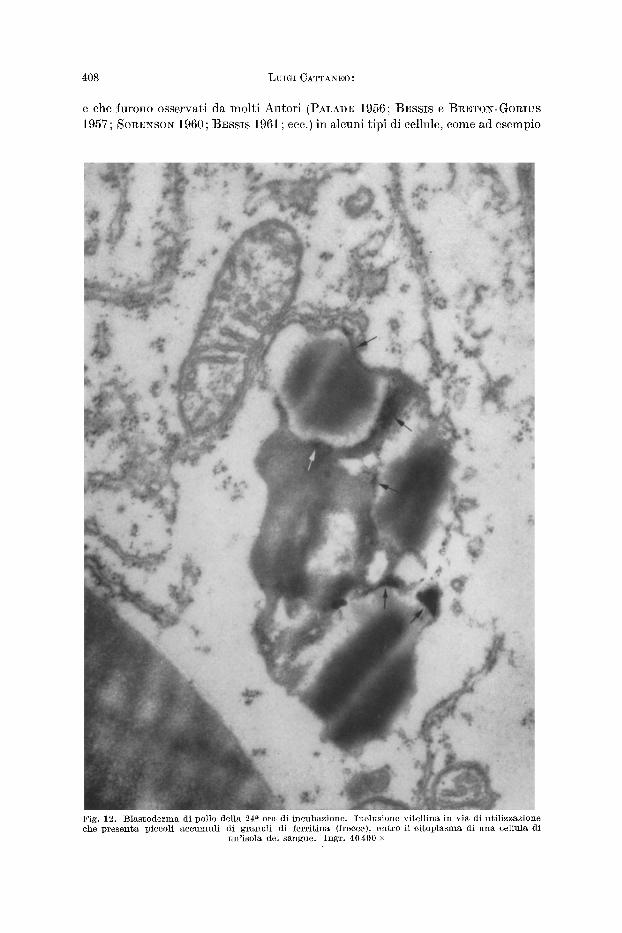
En t ro il vitello, t ra i materiali di varia densitg ehe lo eostituiseono, sono riseon- trabili finissimi granuli, intensamente opaehi agli elettroni (fig. 11), il eui diametro
Fig. II. Blastoderma di pollo della 24 a ora di incubazione. Inclusione di vitello (freccia) contenente numerosissimi granuli di ferritina, nel citoplasma di una cellula di un'isol~ del sangue.
Ingr. 36500 •
si aggira sui 50--60 A. Questi granuli non sono sempre bene rilevabili nelle grosse sferule di vitello ancora compatte, giacch~ possono con faeilit~ essere maseherati dalla forte opacit~ del materiale vitellino entro eui sono dispersi; essi sono invece meglio visibili nelle inelusioni di vitello in eorso di utilizzazione, perch6 infatt i sono spesso addensat i in eorrispondenza dei bordi delle eseavazioni marginali o delle eavit~ interne delle inelusioni medesime (figg. 9 e 12). Det t i granuli infine sono raggruppat i a formare pieeoli ammassi entro i residui vaeuolizzati di quelle forma- zioni vitelline ehe sono ormai quasi to ta lmente riassorbite (fig. 13). Questi reperti dimostrano qnindi the i granuli in questione, almeno nella maggior parte, non passano nel ci toplasma grado a grado che il vitello viene distrutto, ma ehe per- mangono e si concentrano nei residui di questo.
I granuli ora descritti entro il vitello sono molto simili, sia per le loro dimen- sioni che per la loro opacitg, ai granuli di ferritina illustrati dal FAI%RANT (195~) 1
1 Secondo il FAI~RANT (1954), le immagini elettroniche dimostrano solamente il complesso fosfato ferrico idrato della molecola della ferritina.
408 LU~G~ CATTANEO :
e the furono osservati da mol t i Autori (PALAI)E 1956; BESSlS e BI~ETO~-GOl~IUS 1957 ; SORE~SON 1960; BESSIS 1961 ; eec.) in alouni t ipi di cellule, come ad esempio
a Fig . 12. B l a s t o d e r m a di pol io del la 24 ora d i i ncubaz ione . I n c l u s i o n e v i t e l l i n a i n v i a di u t i l i z zaz ione che P r e s e n t s p iecol i a e e u m u l i d i g r a n u l i d i f e r r i t i n a (frecce), e n t r o i l c i t o p l a s m a d i u n a ee l lu la d i
u n ' i s o l a del sangue . I n g r . 40400 •
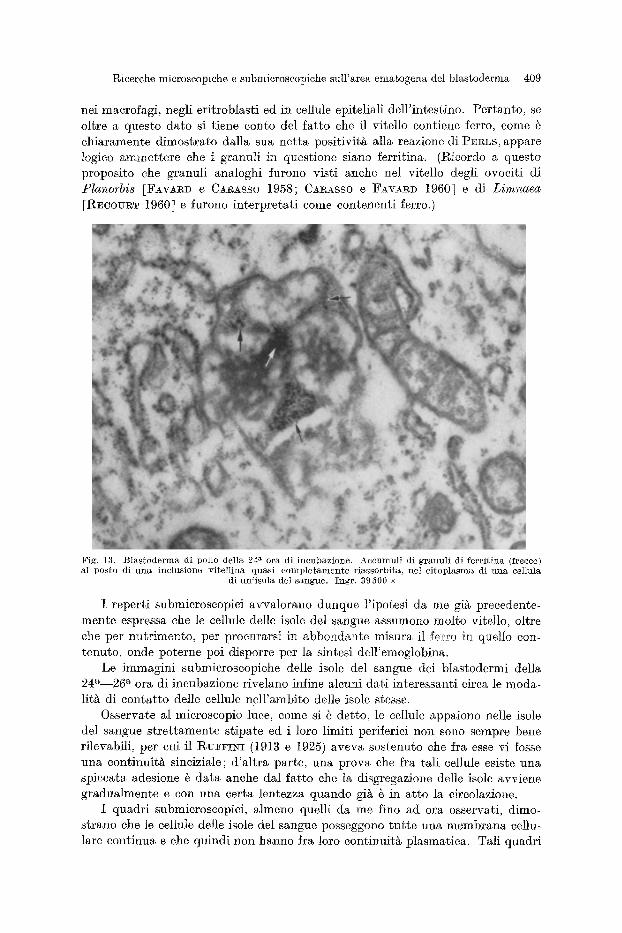
II~icerche microscopiche e submicroscopiche sull'area emagogena del blastoderma 409
nei maerofagi, negli eritroblasti ed in eellule epiteliali dell'intestine. Pertanto, se oltre a questo date si tiene eonto del fat to ehe il vitello eontiene ferro, come @ ehiaramente dimostrato dalla sua netta positivitg alla reazione di PSRLS, appare logieo ammettere ehe i granuli in questione siano ferritina. (Rieordo a questo proposito ehe granuli analoghi furono visti anehe nel vitello degli ovoeiti di P l a n o r b i s [FAvA~D e CA~ASSO 1958; CARASSO e FAVARD 1960] e di Limnaec~
[REcouRT 1960] e furono interpretati come eontenenti ferro.)
Fig . 13. B l a s t o d e r m a di pol lo del la 24 a e r a di i n c u b a z i o n e . Accnnqnl i d i g r a n n l i di fcrr i t ins~ ( f recce) al p o s t o d i u n a i n c l u s i o n c v i t e l l i n a q u a s i e o m p l e t a m e n t e r i a s s o r b i t a , ~e l c i t o p l a s r a a d i u n a cel lula
di u n ' i s o l a de l s a n g a e . I n g r , 39500 •
I reperti submieroseopiei avvalorano dunque l'ipotesi da me gig precedente- mente espressa ehe le celiule delle isole del sangue assumono molto vitello, oltre the per nutrimento, per procur~rsi in abbondante misura iI ferro in quello con- tenure, onde poterne poi disporre per la sintesi dell'emoglobina.
Le immagini submieroseopiehe delle isole del sangue dei blastodermi dells 2 4 ~ 2 6 ~ ors di ineubazione rivelano infine aleuni dati interessanti circa le moda- litg di eontatto delle eellule nell 'ambito delle isole stesse.
Osservate al mieroseopio luee, come si 6 detto, le eellule appaiono helle isole del sangue s tret tamente stipate ed i lore limiti periferiei non sono sempre bene rilevabili, per eui il RvFFI~I (1913 e 1925) aveva sostennto ehe fra esse vi fosse una eontinuitg sineiziale; d 'al t ra parte, una prova ehe Ira tali eellule esiste una spieeata adesione 6 data anehe dal fatto ehe ls disgregazione delle isole avviene gradualmente e con una eerta lentezza quando gig 6 in at to la eireolazione.
I quadri submieroseopiei, Mmeno quelli da me fine ad era osservati, dimo- strano ehe Ie eellule delle isole del sangue posseggono tnt te una membrana eellu- lark continua e ehe quincli non hanno ira lore eontinuitg plasmatiea. Tali quadri

4 1 0 L U I G I CATTAN]~O :
elettronici rivelano perb i seguenti due fatti che sono di per s5 sufficienti a spiegare la stretta adesione che esiste Ira ]e cellule: 1) i contorni cellulari non sono retti-
F i g . 14. B l a s t o d e r m a d i pol io del la 24 a o ra d i i n c u b a z i o n e . I c o n f i n i c e n u l a r i d e n e d u e eel lule p a r z i a l - m e n t e e o m p r e s e ne l la m i e r o f o t o g r a f i a elett l~oniea p r e s e n t a n o i n t e r d i g i t a z i o n i ( f recc ia ) i n g r a n a t e f r a loro ; 1o spaz io i n t e r c e l l u l a r e a p p a r e a t r a t t i pi t t a m p i o . I n e n t r a m b i i l e m b i c i t o p l a s m a t i c i s i r i s c o n t r a n o m i t o o o n d r i , e l e r a e n t i e r g a s t o p l a s m a t i e i , mie roso~ni s p a r s i e d i n e l u s i o n i d i v i t e l lo (V) i n eorso di
u t f l i z zaz ione . I n g r . 23800 •
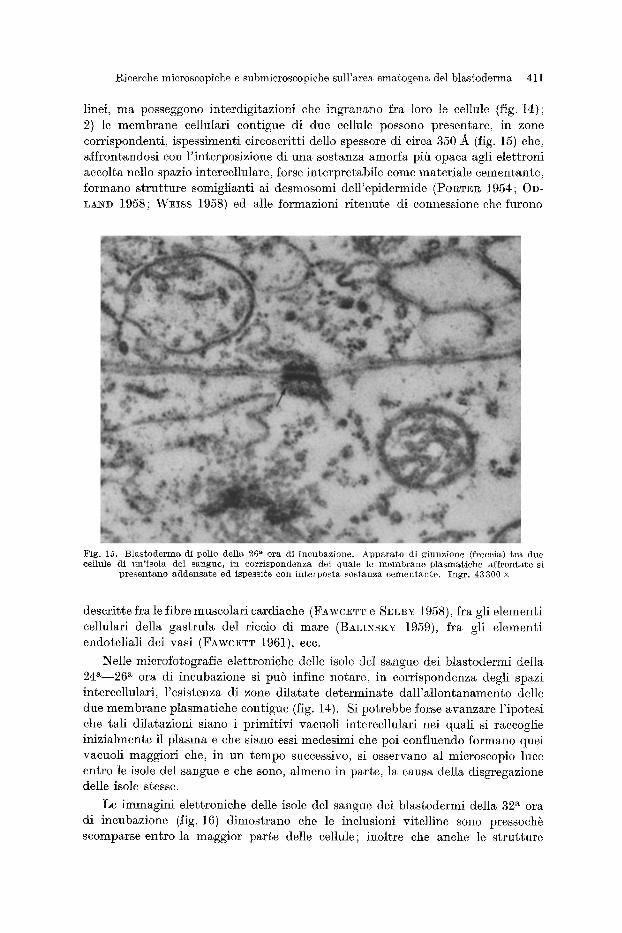
gicerche microscopiche e submicroscopiche sull'a.rea ematogena del blastoderma 411
linei, ma posseggono interdigitazioni ehe ingranano fra loro le cellule (fig. 14); 2) le membrane eellulari eontigue di due eellule possono presentare, in zone eorrispondenti, ispessimenti eireoseritti dello spessore di circa 350 ~ (fig. 15) ehe, aifrontandosi con l'interposizione di una sostanza amorfa pih opaea agli elettroni accolta nello spazio intereellulare, forse interpretabile come materiale eementante, Iormano strutture somiglianti ai desmosomi dell'epidermide (Po~TnR 1954; 0D- LA~D 1958; Wmss 1958) ed alle formazioni ritenute di eonnessione ehe furono
Fig. 15. B la s tode rma di pollo della 26 a ora di incubazione. Appa ra to di g iunzione (freccia) t r a due cellule di un ' i so la del sangue, in corr i spondenza del quale lc m e m b r a n e p lasmat i che aff~:ontate si
p resen tano addensa t e ed ispessi te con in t e rpos t a sos tanza cemen tan te . Ingr . 43300 x
descritte fra le fibre muscolari cardiache ( F A ~ r C E T T e SELBY 1958), fra gli elementi cellulari della gastrula del riccio di mare (BALI~SK~r 1959), Ira gli elementi endotelia]i dei vasi (FAwcETT 1961), ecc.
Nelle microfotogr~fie elettroniche delle isole del sangue dei blastodermi della 24 a 26 a ora di incubazione si pub infine notare, in corrispondenza degli spazi intercellulari, Fesistenza di zone dilatate determinate dalFallontanamento delle due membrane plasmatiche contigue (fig. 14). Si potrebbe forse avanzare l 'ipotesi che tali dilatazioni siano i primitivi vacuoli intercellulari nei quali si raccoglie inizialmente il plasma e che siano essi medesimi che poi confluendo formano quei vacuoli maggiori che, in un tempo successivo, si osservano al microscopio luce entro le isole del sangue e che sono, Mmeno in parte, la causa della disgregazione delle isole stesse.
Le immagini elettroniehe delle isole del sangue dei blastodermi della 32 a ora di incubazione (fig. 16) dimostrano ehe le inclusioni vitelline sono pressoch6 seomparse entro la maggior parte delle cellule; inoltre ehe anehe le s trut ture
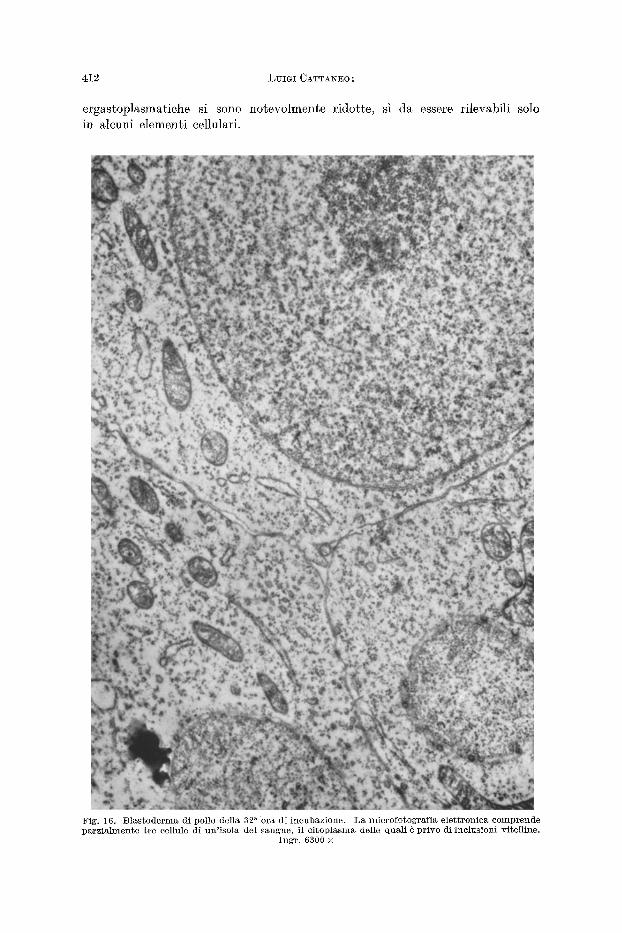
4 1 2 L U I G I CATTAbTEO :
ergastoplasmatiche si sono notevo lmente ridotte, sl da essere rilevabili solo in alcuni e lement i cellulari.
Fig . 16. B l a s t o d e r m a 4 i pollo del la 32 a o r a d i i n c u b a z i o n e . L a microfofGograf ia e l e t t r o n i c ~ c o m p r e n d e p a r z i a h n e n t e ~re cel lnle d i ~ n ' i s o l a de l s a n g n e , i l c i t o p l a s m a delle qua l i ~ p r i v o d i i n c l u s i o n i v i t e l l i ne .
I n g r . 6300 •

Ricerche microscopiche e submicroscopiche sull'area ematogena del blastoderma 413
I reperti submicroscopici ottenuti dallo studio al microscopic elettronieo delle isole del sangue dei blastodermi prelevati alla 40 ~ era di incubazione dimostrano che le eellule si sono iatte rotondeggianti e che molte di esse presentano era nel citoplasma la zona di GOLOL Le cellule inoltre per la maggior parte sono ampia- mente distanziate tra lore e solo alcune conservano ancora un contatto reciproco in tratti limitati del lore contorno.
Considerazioni e conclusioni
Nel corse del presente lavoro sono staff studiati i primi stadi dello sviluppo dell'area ematogena del blastoderma di pollo con particolare considerazione per alcuni dei molteplici e complessi fenomeni che in essa si verifieano.
I processi che si susseguono nell'area ematogena dallo stadio iniziale fine a quello di 14 somiti (44 a era di incubazione), in base alle mie ricerche, possono essere cosi riassunti.
Alla 1~ a era di incubazione, allorquando la linea primitiva si estende solo lunge la metg posteriore dell'area chiara, il mesoderma dell'area opaca 6 rappresen- rate caudalmente alla linea medesima da pochi strati di cellule globose applicati alla superficie profonda dell'ectoderma, separati dall'entoderma mediante l'inter- posizione di uno spazio che si estende fine all'orlo germinativo dove i foglietti embrionali prendono contatto ira lore. Tall cellule mesodermiche, che progressiva- mente aumentano di numero, nel periodo compreso a un incirca tra la comparsa del node di HE~sE~r e la formazione della corda dorsale abbandonano il contatto con l'ectoderma e, attraversando a gruppi o isolatamente l'interposto spazio, discen- done sopra l 'entoderma sul quale ricostituiscono il foglietto mesodermico che, per l 'attiva proliferazione dei suoi elementi, va poi rapidamente espandendosi alla periferia ed aumenta di spessore sl da risultare formate da pih file di cellule stretta- mente avvicinate.
Nel mesoderma dell'area opaea si formano allora, probabilmente per un pro- cesso di secrezione, le eavitg seriate di RuF~I~I. Queste iniziano come piccole lacune intercellulari localizzate nella parte superficiale del mesoderma; tosto per6, ingrandendosi in ogni sense, esse si spingono anehe in profonditg fine a ragginngere l 'entoderma e cosl suddividono il mesoderma stesso in numerosi raggruppamenti eellulari the sono le primitive isole del sangue o isole di WOLFF. II fatto coincide a u n dipresso con la comparsa nell'embrione del primo paio di somiti (24 a era di ineubazione).
Le grosse cavitg seriate, che sono circoseritte da una parete di cellule appiattite, aderiscono inferiormente alle isole del sangue e, negli intervalli fra queste, all'ento- derma; in alto invece, fanno sporgenza entre lo spazio sottoectodermieo dove sono tra lore a eontatto, separate solo da sottili setti di eellule allungate. Successi- vamente, circa allo stadio di 6--8 somiti ( 2 8 ~ 3 0 a era di ineubazione), per la involuzione di tali setti interlacunari le eavitg seriate confluiscono in una eavitg nniea che ~ il celoma dell'area ematogena, il qnale pertanto resta circoseritto dai contorni superiori e da quelli inferiori delle pareti delle singole cavitg seriate ehe, separatisi, sono divenuti rispettivamente la somatopleura e la splancnopleura dell'area medesima. Pertanto, in conseguenza della formazione delle due lamine del mesoderma laterale, le isole del sangue vengono ad essere rivestite superior- mente dalla splanenopleura.

414 LumI CATTANEO"
A cominciare dallo stadio di 8--10 somiti ( 3 0 ~ 3 4 a ora di incubazione), le isole del sangue, che fino a questo momento si erano mantenute strettamente aderenti all'entoderma, vanno allentando i loro rapporti con quest'ultimo foglietto e, per il formarsi al di sotto di esse di cavit~ forse di secrezione, grado a grado se ne staccano. Allora la splancnopleura, prolilerando nei punti dove si solleva per rivestire le isole, produce cellule appiatt~te che si spingono al di sotto delle isole medesime scorrendo sull'entoderma; in tale modo la splancnopleura viene a circondare le isole del sangue anehe inferiormente, racchiudendole entro cavits Sono queste i primitivi vasi sanguigni dell'area ematogena; in essi le isole sono inizialmente compatte ed aderiscono alla parete supcriore, indi vanno ineontro a disgregazione per il formarsi di vacuoli e di spazi Ira le cellule.
Passando ora ad un'analisi critica di alcuni iatti riscontrati nel corso di questo lavoro, vorrei anzitutto sofiermarmi sulla origine delle primitive isole del sangue. I miei dati in proposito, come si ~ visto, non lasciano alcun dubbio sulla deriva- zione diretta delle isole del sangue dal foglietto mesodermico dell'area ematogena; essi dunque, mentre sono in pieno accordo con quanto disse il RUFFI~L nettamente contrastano con l'opinione di quegli Autori che, seguendo le vedute della DA~T- SC~AKOFF e del MAxIMOW, sostengono la formazione delle isole del sangue per opera di cellule del mesenchima migrate dal contorno del mesoderma cmbrionale entro l'area ematogena. Se 6 pur vero infatti ehe nello stadio che va dalla forma- zione del nodo di HE~SE~ alla comparsa della linea primitiva ]e cellule destinate a formare le isole sono sparse Ira l 'ectoderma e l 'entoderma dell'area opaea retro- stante la linea primitiva si da poter sembrare elementi mescnchimali in corso di immigrazione, risulta per6 chiaramente comprovato che dette cellule gi~ in pre- cedenza erano presenti nell'area opaca dove iormavano il foglietto mesodermieo a ridosso dell'ectoderma, e che tale loro atteggiamento migrante 6 l'espressione del fatto che esse stanno abbandonando il rapporto con l'ectoderma per trasferirsi sopra l'entoderma dove poi, prima di dare luogo alle isole del sangue, rieostituiranno nuovamente fl ioglietto mesodermico.
A]tro fatto che intendo qui considerare riguarda l'assunzione del vitello da parte delle cellule destinate ad evolvere in elementi del sangue; questo ienomeno, come si disse, era gis stato osservato nei blastodermi degli uceelli dal G~:~P~, che l 'aveva perb intcrpretato come la prova della derivazione entodermica delle pri- mitive eellule ematiche, e indi dal RU~FI~I che inveee l 'aveva riportato nella sua giusta luce considerandolo una tipica attivit~ de] mesoderma dell'area ematogena. Le mie osservazioni dimostrano che le cellule mesodermiche cominc]ano ad inglobare le sferule vitelline prima ancora di essersi stratificate sull'entoderma, come invece affermato dal RUFFI~I, ma gi~ quando, nel trasmigrare dall'ectoderma sull'entoderma, esse attraversano lo spazio interposto che eontiene numerosissime sfere ectopiche di vitello in eontinuit~ con quelle presenti nella zona dell'orlo germinativo.
Nel corso delle mie ricerche ho soprattutto cercato di stabilire la eausa dell'as- sunz~one di vitello da parte delle future eellule ematiche, sembrandomi c h e l a spiegazione del bisogno di nutrimento data dal t~UFFINI, pur essendo plausibile, non potesse giustificare pienamente fl fatto che il vitello venga inglobato in quantits tale da essere riscontrabile in Iorma cosi manifesta entro il citoplasma, tenuto presente che le altre cellule del blastoderma, se si prescinde da quelle entodermiche

Ricerche microscopiche e submicroscopiche sull'area ematogena del blastoderma 415
dell'area opaca che anehe in stadi avanzati presentano grosse sfere vitelline, one sono wive oppure ne contengono ma in misura molto pifi scarsa, pure nutrendosi anch'esse ovviamente a spese di sostanze provenienti dal vitello. La spiegazione del fenomeno 6 da ricerearsi, a mio avviso, nei seguenti due rilievi emersi nel corso delle mie rieerche : 1) il trattamento delle sezioni dei blastodermi con la reazione al bleu di Prussia di PEALS eolora le sferule di vitello sia estra- che intraeellulari, la quale cosa dimostra in esse l'esistenza di ierro trivalente; 2) lo studio submicroscopico delle cellule delle isole del sangue dei blastodermi della 2 4 ~ 2 6 a ora di incubazione rivela entro le inelusioni vitelline la presenza di granuli di ferritina i quali poi, a seguito del riassorbimento delle inelusioni medesime, restano riuniti in piecoli aeeumuli entro il citoplasma. Lo studio al mieroseopio elettronico del vitello ineluso helle cellule delle isole del sangue dei blastodermi di tale epoea ha appunto dimostrato come i granuli di ferritina non si disperdano nel eitoplasma via via che il vitello viene riassorbito, ma come essi vadano invece addensandosi nella parte periferica dei residui vitellini fino a quando, espletata anehe l'utilizzazione di questi ultimi, rimangono entro le cellule, riuniti in piecoli aecumuli.
In base a tall constatazioni ritengo di potere ammettere ehe le cellule del mesoderma dell'area opaca destinate ad evolvere in elementi del sangue inglobino le sferule vitelline, oltre ehe per assumere nutrimento, per proeurarsi il ferro ehe nel vitello g eontcnuto e che dovr~ a loro servire per la sintesi dell'emoglobina. Considerata dunque sotto questo aspetto, l'assunzione del vitello rappresenterebbe per eosl dire la prima manifestazione morfologica della differenziazione in senso ematopoietico del mesoderma dell'area ematogena. (Non posso eseludere the anche algre cellule congenenti vitello, come quelle dell'entoderma dell'area opaca, siano in grado di accumulate granuli di ferritina, non possedendo infatti per ora elementi sufficienti a riguardo.)
I reperti ultrastrutturali hanno anche dimostrato l'utilizzazione del vitello da parte delte cellule delle isole del sangue, ponendo sotto nuova luce la parteeipa- zione dell'ergastoplasma a questo proeesso. Le sferule vitelline contenute nelle cellule delle isole dei blastodermi della 24a--26 a ora di ineubazione vengono infatti risolte dal mieroscopio elcttronieo in zone di varia forma e grandezza e soprattutto di diversa densitY, ed 6 apparso chiaro che tale disomogeneit~ g da attribuire a stadi diversi della loro utilizzazione; orbene, le strutture ergastoplasmatiche sono apparse tanto pifi sviluppate quanto pifi prossime alle inelusioni vitelline in eorso di distruzione, la quale eosa farebbe appunto supporre the esse siano in relazione col processo di utilizzazione del vitello stesso. Una eonferma di el5 sarebbe data anehe dal fa%o ehe le strutture ergastoplasmatiehe sono risultate essere molto ridotte oppure assenti in quelle eellule in cui il vitello intraeitoplasmatico era stato eompletamente riassorbito, come ad esempio in cellule delle isole del sangue dei blastodermi della 32 a ora di ineubazione.
Le ricerche submicroscopiche hanno infine offerto interessanti rilievi sulle connessioni delle eellule entro le isole del sangue: nei blastodermi della 24~--26 a ora di incubazione, dette cellule non hanno fra loro eontinuit~ plasmatiea, come qualehe Autore (RUFFINI) aveva sostenuto in base ad osservazioni di microseopia luce, esse tuttavia sono strettamente eonnesse sia da interdigitazioni ehe s'ingra- nano vieendevolmente, sia da formazioni assomiglianti ai desmosomi che sono eostituite da ispessimenti eireoseritti di due membrane plasmatiehe eontigue,

416 LI;IGI C.~TTANEO :
affrontati Ira loro e eementati da una sostanza amorfa posta nello spazio inter- cellulare.
L'ult imo punto sul quale vorrei riehiamare l 'attenzione ~ quello relativo alla genesi dei primitivi vasi sanguigni dell 'area ematogena; g infatti noto, come ho gi~ riferito, che sull 'argomento esistono pareri alquanto diseordi, Mcuni Autori ritenendo detti vasi formati dalla splanenopleura, altri dalle eellule del mesen- ehima, altri infine da una trasformazione endoteliale delle eellule pih superfieiali delle isole del sangue. I risultati da me ottenuti dimostrano ehe i primitivi vasi estraembrionali sono Iormati dalla splancnopleura; come si ~ visto infatti, allor- quando le isole del sangue si staeeano dall 'entoderma, la splancnopleura ehe gi~ le rivestiva superiormente prolifera in loro eorrispondenza e le avvolge eosl anehe al di sotto, delimitando attorno a eiaseuna di esse un lume vasale. Le mie osserva- zioni eoneordano dunque con quanto afiermb il RuFrI~I, ehe g appunto l 'Autore ehe sostenne la derivazione dei vasi estraembrionali dalla splanenopleura; esse perb dimostrano anche la modalits mediante cui il foglietto viseerale del meso- derma laterMe forma le pareti vasali, modalits the il I~UFFIm non fu in grado di osservare perch,, come Egli medesimo riferisee, non gli Iu possibile studiare blasto- dermi in stadi di sviluppo sufiieientemente vieini. Dai miei reperti risulta infine chela parete dei primitivi vasi ~ formata da uno strato unieo di eellule appiattite, e non da due strati come sostenuto dal t~UFFI~I; g probabile che il dato di questo Autore si riierisea a blastodermi in stadi di sviluppo pifi progrediti di quelli da me studiati.
Zusammen~assung Die Untersuchungen fiber das extraembryonale h/imatogene Gebiet des Blasto-
derms beim ttfihnchen wurden unternommen, um folgende strittige Probleme zu 15sen: I-Ierkunft und Bildungsweise der Blutinseln und Beschaffenheit der primi- riven GefgBwi~nde, sowie die submikroskopische Zellstruktur der Blutinseln.
Ieh habe das Blastoderm vom t Iuhn yore Stadium des Primitivstreifens ab his zum Stadium yon 14 Somiten untersucht (yon der 14. bis zur 44. Inkubations- stunde) und habe folgende Resultate erhMten.
Aus der Betrachtung der mit H/imalaun-Eosin und mit Eisenh/imatoxylin gel/~rbten Serienschnitte geht hervor, 1. dab die Blutinseln direkt vom Mesoderm herstammen, wie I~VFFI~I feststellt, und nicht vom Mesenchym, wie MAxIMow und viele andere behaupten, und erst recht nicht yore Entoderm, wie G~31P~ meint; 2. dag die Vorg/~nge, die zur Bildung der besagten Inseln ffihren, in Iol- gender Reihenfolge stattiinden: anfangs (Stadium der 14.--16. Std) hat das Niesoderm Kontak t zur Unterfl~che des Ektoderms; bMd (17.--19. Std) legt es sich dem Entoderm an; dann (20.--24. Std) erscheinen im Mesoderm serienweise angeordnete }Iohlr/iume, die es in Blutinseln zerlegen; sp/~ter (yon der 28. Std an) bilden die W/inde dieser Itohlr/iume die Somatopleura und die Splanehnopleura u n d e s ist letztere, die endiich die Bildung der primitiven Gefs um die Blutinseln herum entstehen ]~gt. Auch diese Resultate beweisen und erweitern die Angaben yon I~VFFIm und stehen haupts~chlich im Gegensatz zu den Be- hauptungen all der anderen Autoren, die der Existenz der serienweise angeord- neten I tohlrgume nicht Rechnung tragen und die auBerdem die primitiven Gef/il~- w/inde yon den peripherischsten Zellen der Inseln oder yon Mesenchymzellen ableiten.

Ricerche microscopiche e submicroscopiche sull'area ematogena del blastoderma 417
,Nachdem ich beobachte t hat ,e , dab die Zellen des Mesoderms bereits zur
17. Std zahlreiche Dotterkfigelchen in ihrem Cytoplasma en tha l ten u n d nachdem
ich histochemisch ( P ~ L s Reakt ion) in diesen Kfigelchen t r ivalentes Eisen nach- gewiesen hatte , sollte an Ul t ra ie inschn i t t en festgestellt werden, aui welehe Weise das Dot te rmater ia l verwertet wird und was mi t dem in ihm en tha l t enen Eisen gesehieht.
Die elektronenmikroskopischen Un~ersuehungen zeigen die wichtige Rolle des Ergastoplasmas bei der Verwendung der in den Blutinselzellen en tha l t enen Dotter- kfigelehen ; sic beweisen auBerdem, dab in denDotterkf igelehen zahlreiche Ferr i t in- kSrnchen vorhanden sind, die zu kleinen Anhgufungen vereint im Cytoplasma b]eiben, naehdem das Dot te rmater ia l resorbiert worden ist. Diese Tatsache lgBt vermuten , dab die Dot te raufnahme seitens der Blutinselzellen haupt.sgchlioh der Besehaffung yon Eisen ffir die kfinftige Hgmoglobinsynthese dient. Ferner zeigen die elektronenmikroskopisehen Untersuehungen , dab der anfangs bestehende enge Zusammenhang der Blutinselzellen un te re inander auf fingerf6rmig ine inander verfloehtene Ausbuch tungen (Interdlgi ta t ionen) der Zellgrenzen und auf Ver- d iekungen der durch Ki t t subs t anz ve rbundenen l~lasmamembranen zurfiekzu- ffihren ist.
B i b l i o g r a f i a
BALzxs~:Y, B. I. : An electron microscopic investigation of the mechanisms of adhesion of the cells in a sea urchin blastul~ and gastrula. Exp. Cell Res. 16, 429--433 (1959).
- - , and H. WALT~E~: The immigration of presumptive mesoblast from the primitive streak in the chick as studied with the electron microscope. Aeta Embryol. Morph. exp. 4, 261--283 (1961).
BELLAIgS, R. : The conversion of yolk into cytoplasm in the chick blastoderm as shown by electron microscopy. J. Embryol. exp. Morph. 6, 149--161 (1958).
BERTI=IIER, J. : Le fer darts l'ovog6n~se chez Pl~norbis corneus L. Bull. biol. Fran. Belg. 8~, 61--78 (1948).
BEssls, M. : The blood cells and their formation. In a. BRACKET and A. E. MIRSKY, The cell. Biochemistry, Physiology, Morphology, Voh V: Specialized cells, Part II, p. 163~217. New York and London: Academic Press 1961.
-- , et J. BRETOI"T-GoRIUS : Trois aspects du ferdans des coupes d'organes examinees au micro- scope 6lectronique (ferritine et d~riv6, dans les cellules intestinales, les ~rythroblastes et les eellules r6ticulaires). C. R. Acid. Sci. (Paris) 24a, 1271--1272 (1957).
- - - Iron particles in normal erythroblasts and normal and pathological erythroeytes. J. biophys, biochem. Cytol. 3, 503--504 (1957).
BRAus-EIqGELI=IARDT, A. : Untersuehungen zur Ermittlung der Entstehung der roten Blutzellen in der Embryonalentwioklung der Urodelen. Wilhelm Roux' Arch. Entwickl.-Meeh. Org. 140, 741--789 (1940).
BREME1% J. L.: The easiest blood-vessels in man. Amer. J. Anat. 16, 447--~75 (1914). CARASSO, N., et P. FAVARD : Vitellogen~se de la planorbe. Ultrastructure des plaquettes vitel-
lines. In: Vierter Internat. Kongr. ffir Elektronenmikroskopie. Berlin 10.--17. Sept. 1958. Verhandlungen, Bd. II: Bio]ogiseh-Medizinischer Tell, S. 431~435. Berlin-GSt- tingen-Heidelberg: Springer 1960.
C~IAeCoI, G. : Trattato di embriologi~ con particol~re riguardo alla storia dello sviluppo dei mammiferi e dell'uomo, Parte II: La formazione dell'embrione. Milano : See. ed. libr. 1932.
DANTSOttAKOFF, W. : D~ber das erste Auftreten der Blutelemente im Hfihnerembryo. Fol. haemat. (Leipzig) 4, Suppl., 159--166 (1907).
- - Untersuchungen fiber die Entwlcklung des Blutes und Bindegewebes bei den VSgeln. I. Die erste Entstehung der Blutzellen beim giihnerembryo und der Dottersack als blut- bildendes Organ. Anat. H. 37, 471--589 (1908).
- - Origin of the blood cells. Development of the haematopoetic organs and regeneration of the blood cells from the standpoint of the monophyletic school. Anat. 1%ec. 1O, 397~414 (1915--1916).

418 LwG~ CATTAI~EO **
FARRAZ~T, J. L. : An electron microscopic study of ferritin. Biochim. biophys. Acta 13, 569--576 (1954).
FAVARD, P.: L'origine ergastoplasmique des granules prot~iques dans les spermatocytes d'Ascaris. C. 1%. Acad. Sci. (Paris) 247, 531--533 (1958).
- - , et N. CARASSO: Origine et ultrastrueture des plaquettes vitellines de la planorbe. Arch. Anat. micr. Morph. exp. 47, 211--234 (1958).
FAWCETT, D. W.: Intercellular bridges. Exp. Cell l%es., Suppl. 8, 174--187 (1961). - - , and C. C. SELBY: Observations on the fine structure of the turtle atrium. J. biophys.
biochem. Cytol. 4, 63--72 (1958). FEDERICI, E . : Recherches ~xperimentales sur le potentialit~s de l 'ilot sanguin chez l 'embryon
de Rana ]usca. Arch. Biol. (Liege) 36, 465--487 (1926). Goss, CH. M. : Experimental removal of the blood island of Amblystoma punctatum embryos.
J . exp. Zool. 52, 45--63 (1928--1929). GRIPER, L. : Untersuchungen fiber die Herzbildung der V6gel. Wilhelm Roux' Arch. Ent-
wickl.-Mech. Org. 24, 375--410 (1907). GRODZI~SKI, Z. : ~ber die Entwicklung yon unterkfihlten Itfihnereiern. Wilhelm l%oux' Arch.
Entwickl.-Mech. Org. 129, 502--521 (1933). - - Weitere Untersuchungen fiber den EinfluB der Unterkfihlung auf die Entwicklung der
Hfihnereier. Wilhelm Roux' Arch. Entwickl.-Meeh. Org. 131, 653--671 (1934). HILL, R.: A method for the estimation of iron in biological material. Proc. roy. Soc. B 197,
205--214 (1931). HILTON, W. A. : The development of the blood on the transformation of some of the early
vitelline vessels in amphibia. J. Morph. 24, 339--382 (1913). His, W. : Lecithoblast und Angioblast der Wirbelthiere. Histogenetische Studien. Abh. kgl.
s~chs. Ges. Wiss. (Leipzig), math.-phys. K1. 26, 171--328 (1900). KhLLIKER, A. : Embryologie ou Trait6 complet du dSveloppement de l 'homme et des animaux
sup~rieurs. (Trad. par A. SChnEIDER.) Paris: Reinwald 1882. LENTI, C. : Stato chimico del ferro nell 'uovo di pollo fecondato e neU'embrione durante lo
sviluppo. Arch. Sci. biol. 25, 1--6 (1939). MARZA, V.D. , E. MARZA et L. CHIOSA: Etude histochimique du fer dans l 'ovaire de poule.
Bull. Hist. appl. 9, 213--225 (1932). MAXIMOW, A. : Bindegewebe und blutbildende Gewebe. In v. MhLLE~TDORFS ttandbuch der
mikroskopischen Anatomie des ~enschen, Bd. II , Teil I : Epithel- und Drfisengewebe. Bindegewebe und blutbildende Gewebe. Blur, S. 232--583. Berlin: Springer 1927.
McCA~cE, R.A. , and E. M. WIl)nowsozq: Cir. in HuTcHIsoz~'s Food and the principles of dietetics. Revised by V. H. MOTTR~ and G. GRAHA~I. Ed. X. London: Arnold & Co. 1948.
MGFARLANE, W. D. : The alkaline hydrolysis of lecitho-vitellin with particular reference to the separation of an organic fraction containing iron and copper. Biochem. J. 26, 1061--1071 (1932).
MILLER, A. M., and J. E. McWttoRTEt~: Experiments on the development of blood vessels in the area pellucida and embryonic body of the chick. Anat. Rec. 8, 203--227 (1914).
ODLA~TD, G. F. : The fine structure of the interrelationship of cells in the human epidermis. J. biophys, biochem. Cytol. 4, 529--538 (1958).
OLIVO, O. : Sulrinizio della funzione eontrattfle del euore e dei miotomi dell'embrione di pollo in rapporto alla loro differenziazione morfologica e strutturale. Arch. exp. Zellforsch. 1, 427--500 (1925).
PALAnE, G. E. : The endoplasmic reticulum. J. biophys, biochem. Cytol. 2, No 4,Part 2 Suppl., 85--98 (1956).
P~SA, A. : Trattato di embriologia generale. Mflano: Garzanti 1944. PORTE~, R. : Observations on the submicroscopic structure of animal epidermis. Anat. Rec.
118, Abst., 433 (1954). REAGAn, F. P. : Experimental studies on the origin of vascular endothelium and of erythro-
cytes. Amer. J. Anat. 21, 39--175 (1917). RECocR% A. : Some aspects of the oogenesis of the pondsnail Limnaea stagnalis L. In:
Vierter internat. Kongr. ffir Elektronenmikroskopie. Berlin 10.--17. Sept. 1958. Ver- handlungen, Bd. I I : Biologiseh-Mediziniseher Tefl, S. 427--431. Berlin-Ghttingen-tteidel- berg: Springer 1960.

lZicerche microscopiche e sublnieroscopiche sull 'area ematogena del blastoderma 419
I:~0CKERT, J. : ~Tber die Ents tehung der endothelialen Anlagen des Herzens und der ersten Gef~l~stgmme bei Selachier-Embryonen. Biol. Zbl. 8, 385--399, 417--430 (1888--1889).
- - , u. S. ]V[OLLIER: Die erste En ts tehung der Gefg6e und des Blutes bei Wirbeltieren. In O. HEETWIG, Handbuch der vergleichenden und experimentellen Entwickelungslehre der Wirbeltiere, Bd. I, Tell I, S. 1019--1278. Jena : Gustav Fischer 1906.
RUFFI•I, A. : L'origine, la sede e le dffferenziazioni dell 'abbozzo del sangue e dei vasi sanguigni nel blastoderma di pollo. Bios 1, 5- -19 (1913). Fisiogenia. La biodinamica dello sviluppo ed i fondamental i problemi morfologici dell'em- briologia generale. Milano: Vallardi ]925.
- - Processi di teenica embriologica ed istologica. Bologna: Cappelli 1927. Sol~s~sos, G. D. : An electron microscopic s tudy of hematopoiesis in the liver of the fetal
rabbit . Amer. J. Anat. 106, 2 7 ~ 0 (1960). STOCKARD, C. I~. : The origin of blood and vascular endothelium in embryos wi thout a circula-
t ion of the blood and in the normal embryo. Amer. J. Anat. 18, 227--327 (1915). ST/3I~I~ jr., Pg. : Beobachtungen zur Organentwicklung bei erythrozytenfreien Amphibien-
larven. Wilhelm l~oux' Arch. Entwicld.-Mech. Org. 124, 707--746 (1931). STOI~TI, E. : Studio sull 'ematopoiesi nella v i ta embrionale. Ricerche morfologiche e speri-
mental i su: Mammiferi, Uccelli ed Anfibi. Par te I. I1 periodo preepatico. Arch. zooh ital. 21, 2 4 1 ~ 2 6 (1935).
STI~ICtlT, 0. VAN DER: Nouvelles recherches sur la gengse des globules rouges et de globules blancs du sang. Arch. Biol. (Ligge) 12, 199--344 (1892).
TOMPSETT, S. L. : Thiolaeetie acid as a reagent for the determinat ion of the inorganic iron content of certain biological materials. Biochem. J. ~o8, 1536--1543 (1934).
WASSERMAnn, F. : Uber den makro- und mikrochemischen Eisennachweis im Dot te r des Hiihnereies. Anat. H. 42, 281--310 (1910--1911).
WEISS, P.: Cell contact. Int . Rev. Cytol. 7, 3 9 1 ~ 2 3 (1958). WETZEL, I~. : Untersuchungen am IIi ihnchen. Die Entwieklung des Keims wghrend der ersten
beiden Bruttage. Wilhelm l%oux' Arch. Entwickl.-Mech. 0rg. 119, 188--321 (1929).
Dott. LuIGI CATTANEO, Istituto di Anatomia Umana Normale della Universit&, Pavia, via Forlanini 8 (Italia)
Z. Anat. Entwickh Gesch., Bd. 123 29


















