Recettori - Pagina di ingresso · può legare anche dei ‘modulatori’ della trasmissione ......
64
Recettori
Transcript of Recettori - Pagina di ingresso · può legare anche dei ‘modulatori’ della trasmissione ......

Recettori

La comunicazione intercellulare: trasmettitori e recettori
La comunicazione tra cellule permette agli organismi di adattarsi in modo coordinato e continuo alle variazioni ‘ambientali’.
Le cellule comunicano mediante mediatori chimici di natura molto varia
Il mediatore viene prodotto e secreto da un tipo cellulare
La cellula cui è destinata la comunicazione, cellula ‘bersaglio’, ha recettori in grado di riconoscere il mediatore

RECETTORI
E’ definita recettore qualsiasi macromolecola funzionale dell’organismo che lega un farmaco
Sono recettori per i farmaci:
* recettori per i neurotrasmettitori e gli ormoni * enzimi * canali ionici * acidi nucleici
I recettori determinano le relazioni quantitative tra dose ed effetti farmacologici
I recettori sono responsabili della selettività dell’azione farmacologica

I recettori sono sempre proteine che legano i trasmettitori (ligandi) ad un sito specifico (sito di legame).
In conseguenza all’interazione recettore-ligando il recettore modifica la sua conformazione; questa modifica di conformazione da’ origine ad una risposta da parte della cellula bersaglio (trasduzione).
Vi sono numerosi meccanismi che provvedono allo ‘spegnimento’ del segnale (interruzione della trasmissione)

Caratteristiche della trasmissione
Ogni recettore riconosce un solo trasmettitore ma può legare anche dei ‘modulatori’ della trasmissione
La correttezza (specificità) del flusso di comunicazioni è assicurata principalmente da tre fattori:
Presenza dei recettori solo sulle cellule bersaglioNel caso di trasmettitori con recettori multipli, la
specificità è assicurata dalla vicinanza della cellula bersaglio al sito di rilascio del trasmettitore
Meccanismi di rimozione del trasmettitore
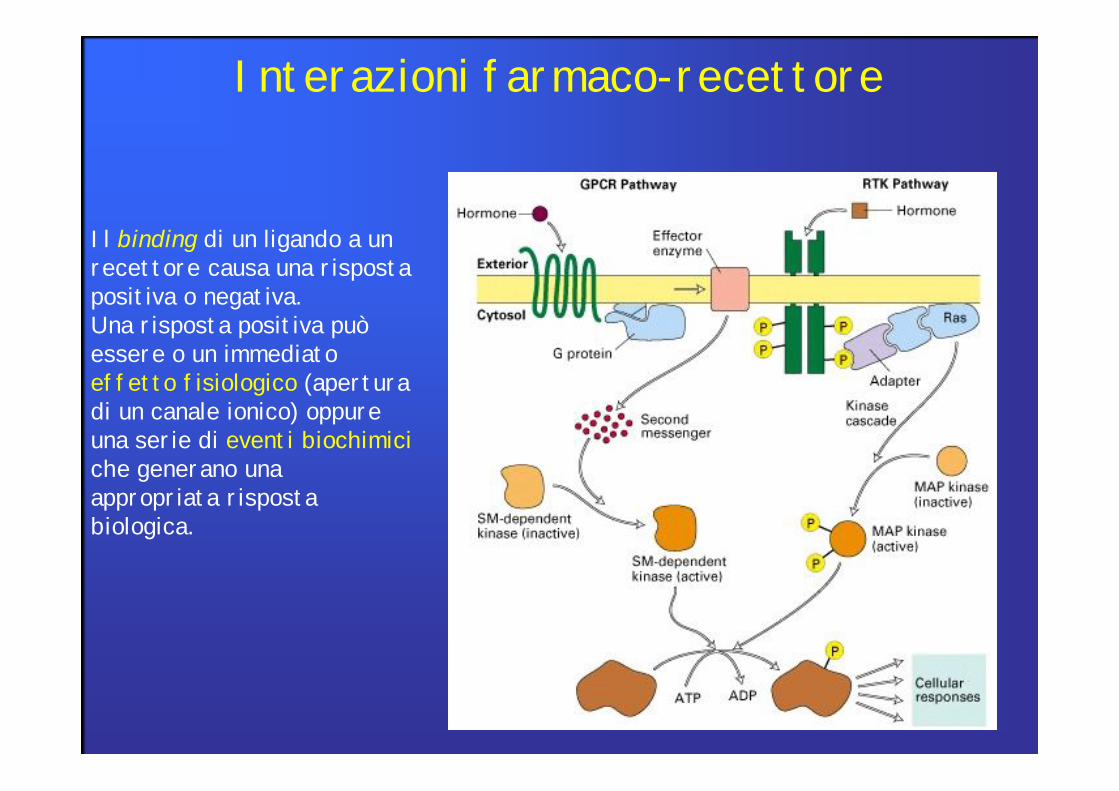
Interazioni farmaco-recettore
Il binding di un ligando a un recettore causa una risposta positiva o negativa.Una risposta positiva può essere o un immediato effetto fisiologico (apertura di un canale ionico) oppure una serie di eventi biochimiciche generano una appropriata risposta biologica.

Ligando e recettore
Farmaco Farmaco-Recettore
Il legame è dipendente dalla concentrazione [C] del farmaco:man mano che [C] aumenta nei fluidi extracellulari, l’equilibrio si sposta a destra e il farmaco si lega al recettore.Quando [C] diminuisce, a causa del metabolismo e dell’escrezione, l’equilibrio si sposta verso sinistra e il complesso si dissocia.
A basse concentrazioni un basso numero di recettori è impegnato dal ligando.
I farmaci con lunga durata d’azione sono quelli che formano legami stabili con il recettore.
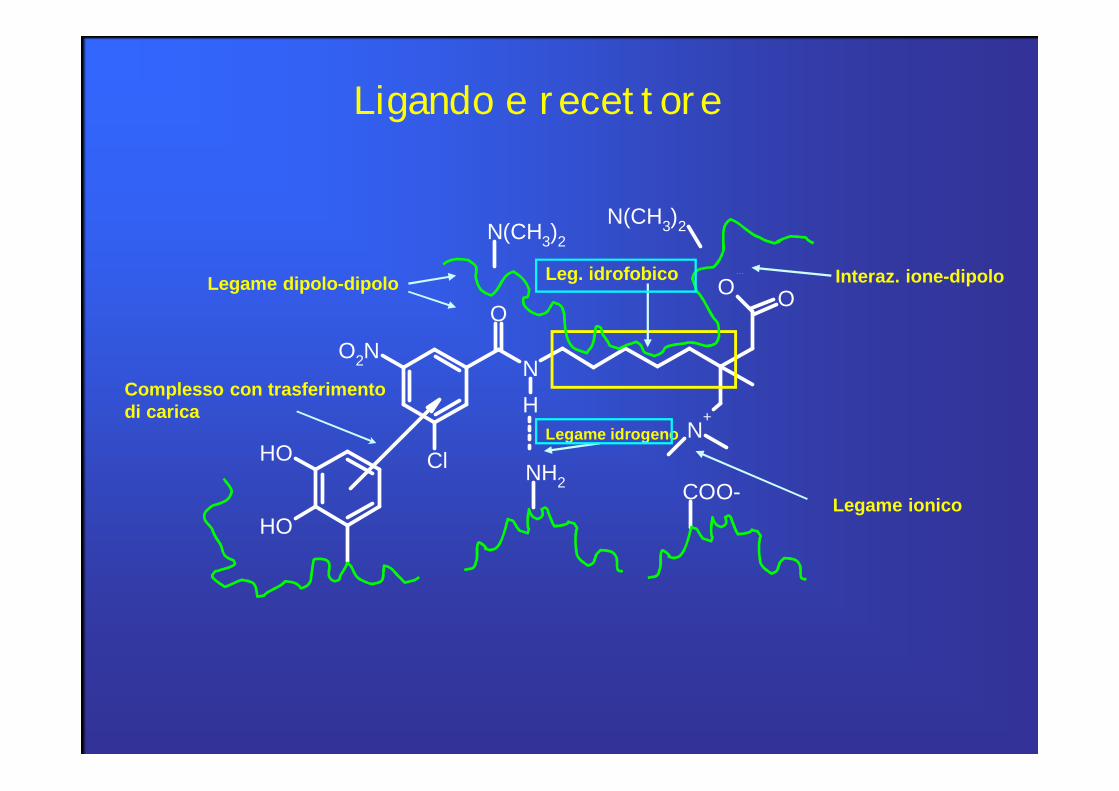
Ligando e recettore
Cl
N
O
O2N
O O
N+H
OH
OHCOO-
N(CH3)2N(CH3)2
NH2
Leg. idrofobico
Legame idrogeno
Legame ionico
Interaz. ione-dipolo
Complesso con trasferimento di carica
Legame dipolo-dipolo

Ligando e recettore
Legame covalente:è di gran lunga il più forte dei legami ligando-recettore. Forma un legame irreversibile. Raramente viene cercato nell’azione di un farmaco (eccezione: chemioterapici nel trattamento del cancro)
Legame ionico o elettrostatico:è molto importante nelle interazioni ligando-recettore in quanto molti gruppi funzionali dei recettori sono ionizzati a pH fisiologico. Si formano interazioni reversibili.
Interazioni elettrostatiche:in forma di forze attrattive ione-dipolo, interazioni dipolo-dipolo e legame H.Formano legami più deboli del legame ionico.Sono le interazioni ligando-recettore più diffuse.
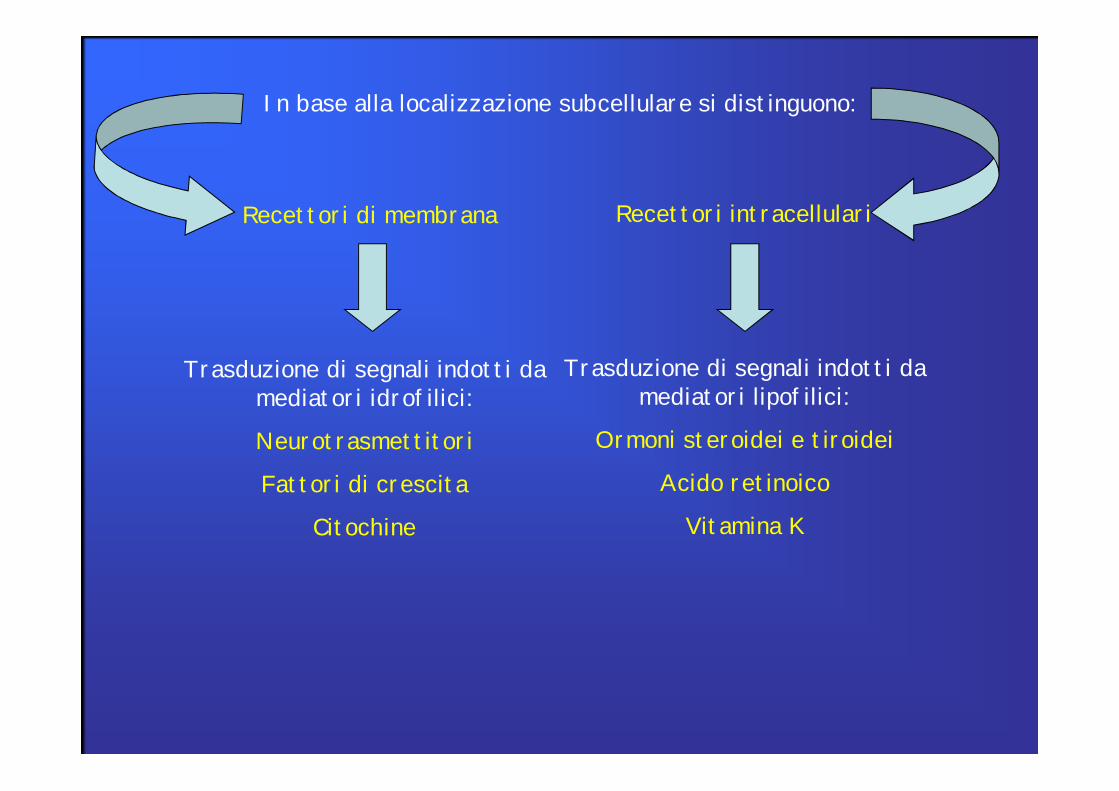
In base alla localizzazione subcellulare si distinguono:
Recettori di membrana Recettori intracellulari
Trasduzione di segnali indotti da mediatori idrofilici:
Neurotrasmettitori
Fattori di crescita
Citochine
Trasduzione di segnali indotti da mediatori lipofilici:
Ormoni steroidei e tiroidei
Acido retinoico
Vitamina K

Recettori di membrana
6 Superfamiglie:
Recettori Canale
Recettori accoppiati a Proteine G
Recettori con attività Guanilato ciclasica
Recettori con attività Tirosin-kinasica
Recettori per le citochine
Recettori per l’adesione cellulare

Canali ionici
Canali operati da ligandiRecettori canali: Canali operati da secondi messaggeri, proteine G, prodotti metabolici
Canali ionici operati dal voltaggio
Canali operati da stimoli sensorialiCanali meccanosensibiliCanali attivati da fotoni o agenti chimici odoranti e gustativi (spesso mediante 2ndi messaggeri)
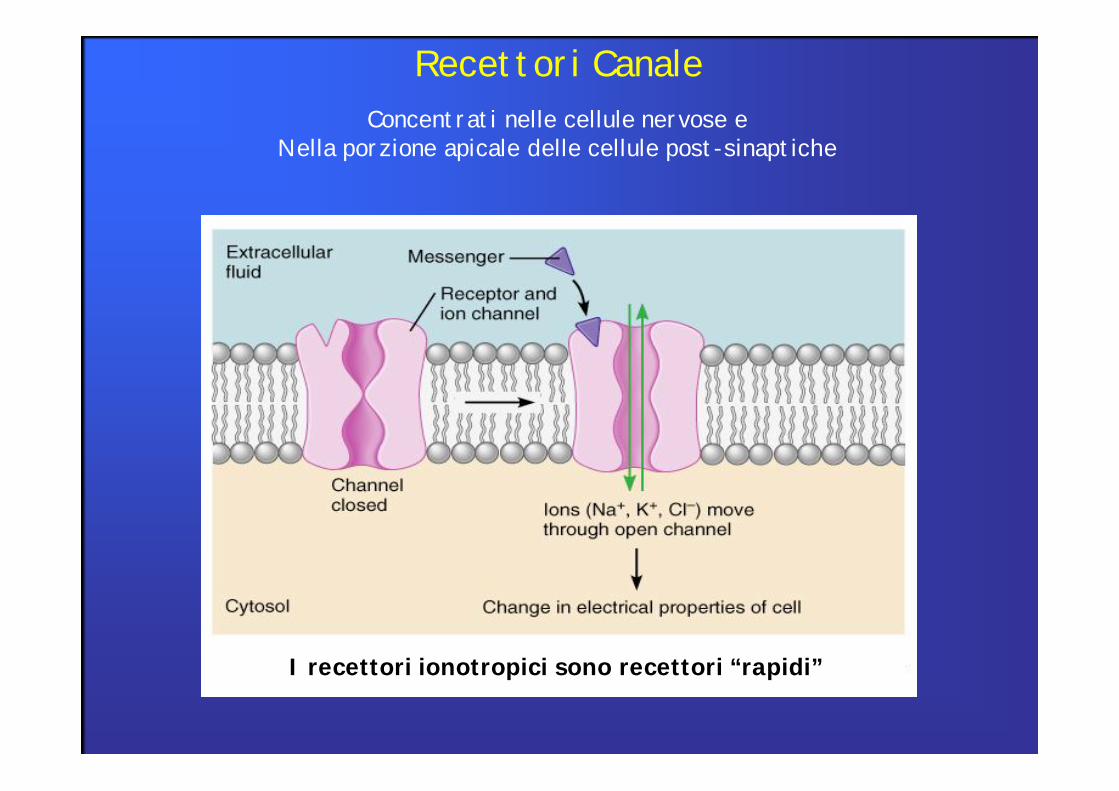
Recettori CanaleConcentrati nelle cellule nervose e
Nella porzione apicale delle cellule post-sinaptiche
I recettori ionotropici sono recettori “rapidi”
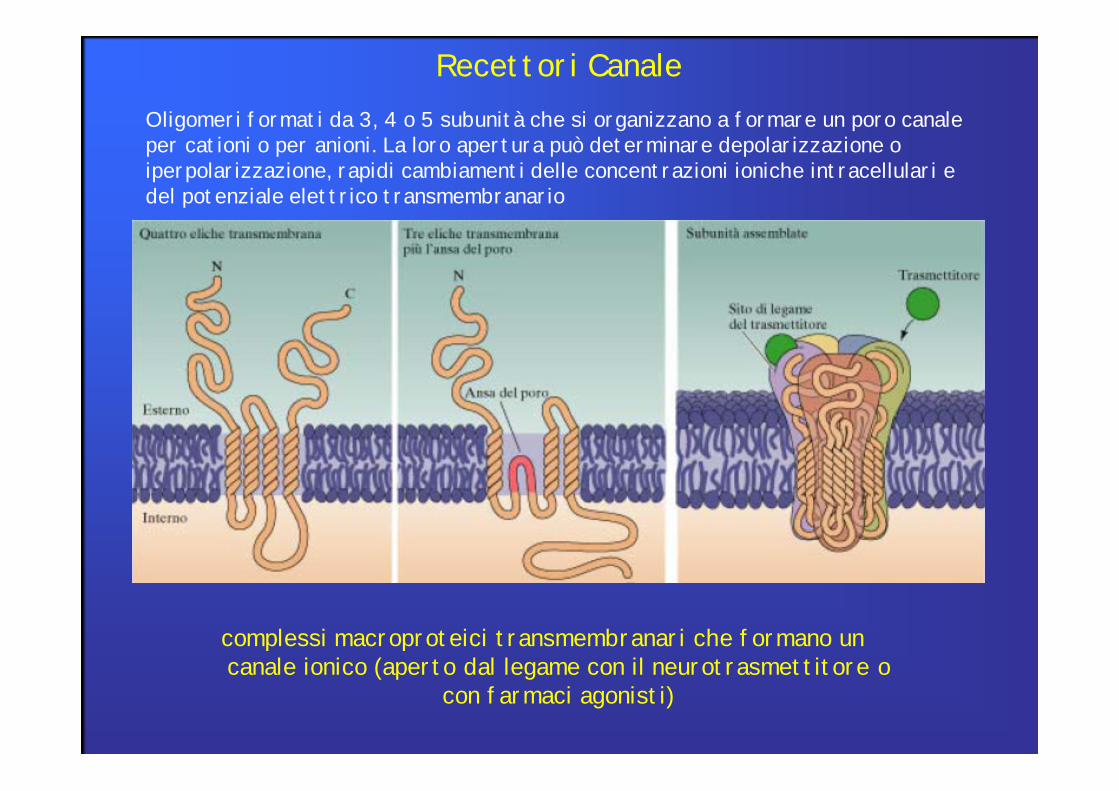
Oligomeri formati da 3, 4 o 5 subunità che si organizzano a formare un poro canale per cationi o per anioni. La loro apertura può determinare depolarizzazione o iperpolarizzazione, rapidi cambiamenti delle concentrazioni ioniche intracellulari e del potenziale elettrico transmembranario
Recettori Canale
complessi macroproteici transmembranari che formano uncanale ionico (aperto dal legame con il neurotrasmettitore o
con farmaci agonisti)

1° classe RECETTORI DELLA SUPERFAMIGLIA DEIRECETTORI NICOTINICI (recettore nicotinico muscolare e
neuronale, recettore GABAa, recettore della glicina e recettore 5HT3 serotonina)
2° classe RECETTORI IONOTROPI PER IL GLUTAMMATO(recettori NMDA, AMPA e Kainato)
3° classe RECETTORI APERTI DA NUCLEOTIDI CICLICI(recettori cAMP e cGMP)
4° classe RECETTORI IONOTROPI DELL’ATP (P2X)
4 classi di recettori canale
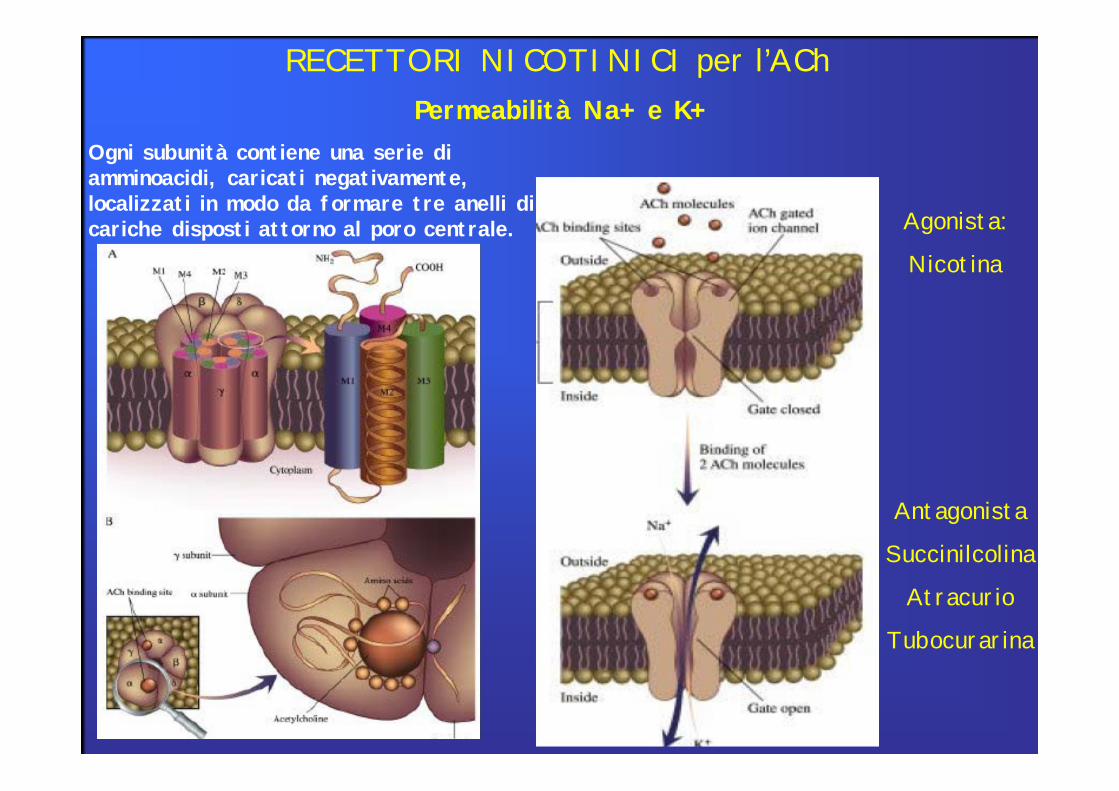
Ogni subunità contiene una serie di amminoacidi, caricati negativamente, localizzati in modo da formare tre anelli di cariche disposti attorno al poro centrale.
Permeabilità Na+ e K+
RECETTORI NICOTINICI per l’ACh
Agonista:
Nicotina
Antagonista
Succinilcolina
Atracurio
Tubocurarina
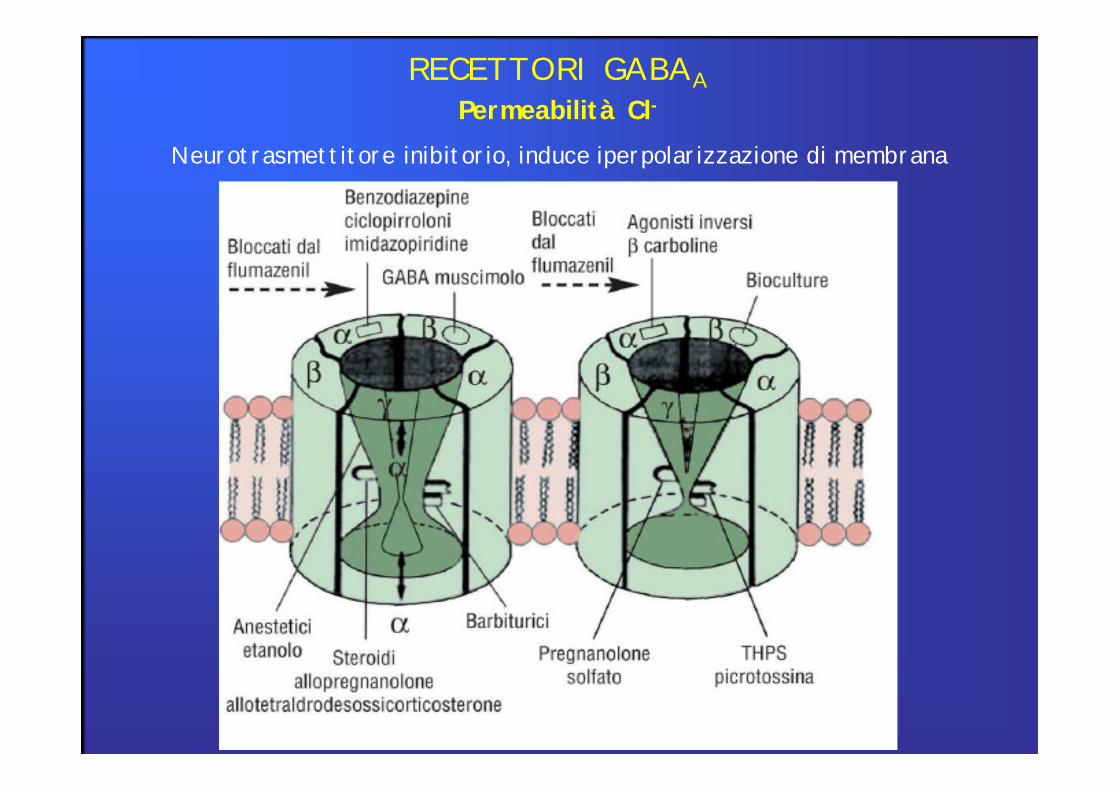
Permeabilità Cl-RECETTORI GABAA
Neurotrasmettitore inibitorio, induce iperpolarizzazione di membrana
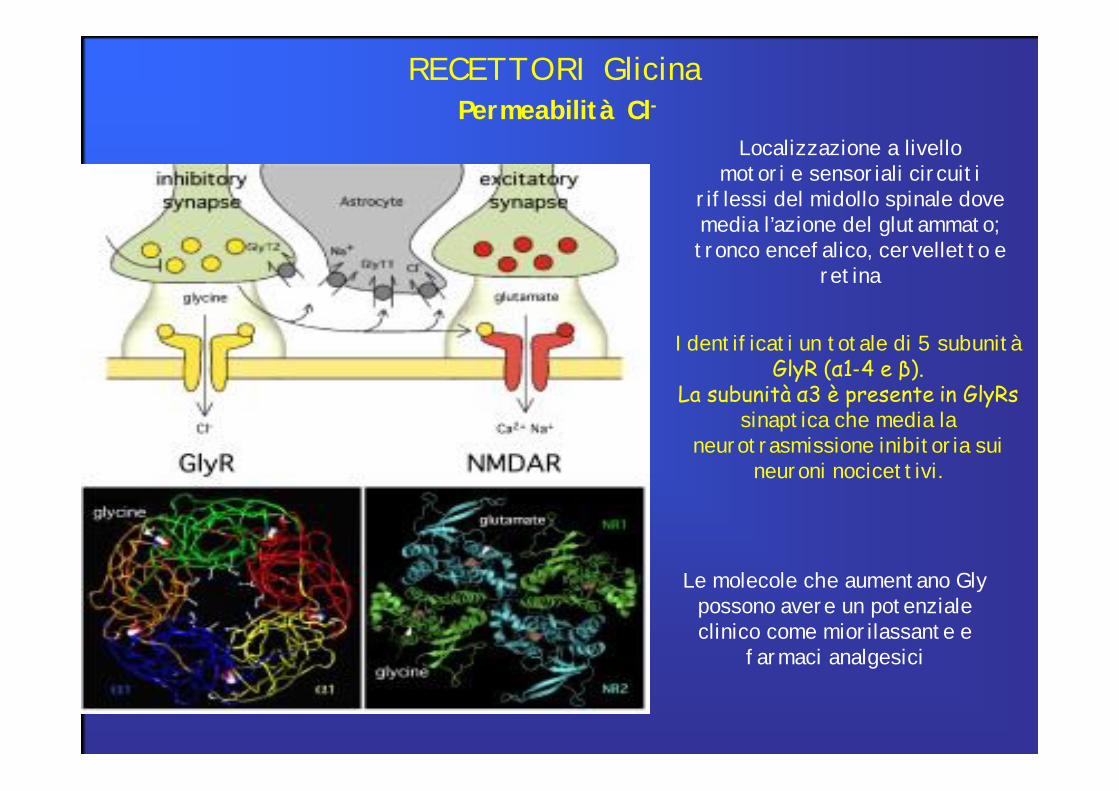
Permeabilità Cl-RECETTORI Glicina
Localizzazione a livello motori e sensoriali circuiti
riflessi del midollo spinale dove media l’azione del glutammato;tronco encefalico, cervelletto e
retina
Identificati un totale di 5 subunità GlyR (α1-4 e β).
La subunità α3 è presente in GlyRs sinaptica che media la
neurotrasmissione inibitoria sui neuroni nocicettivi.
Le molecole che aumentano Gly possono avere un potenziale clinico come miorilassante e
farmaci analgesici
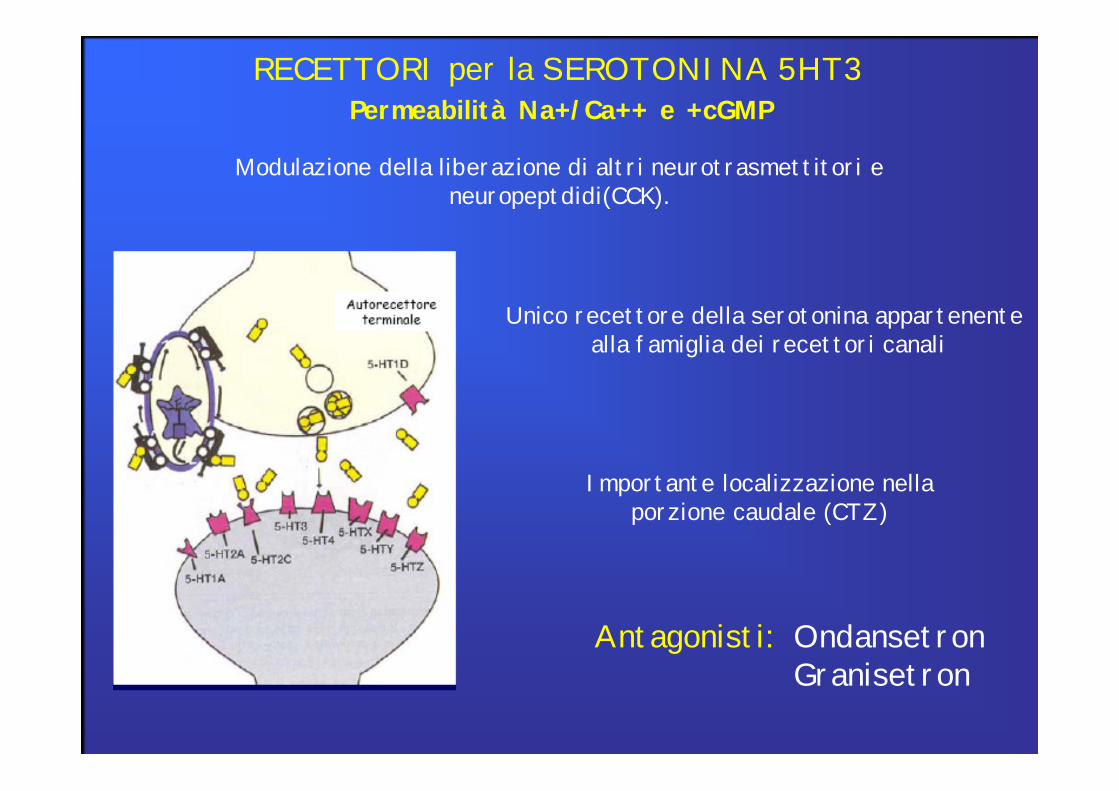
Permeabilità Na+/Ca++ e +cGMPRECETTORI per la SEROTONINA 5HT3
Modulazione della liberazione di altri neurotrasmettitori e neuropeptdidi(CCK).
Antagonisti: OndansetronGranisetron
Importante localizzazione nella porzione caudale (CTZ)
Unico recettore della serotonina appartenente alla famiglia dei recettori canali
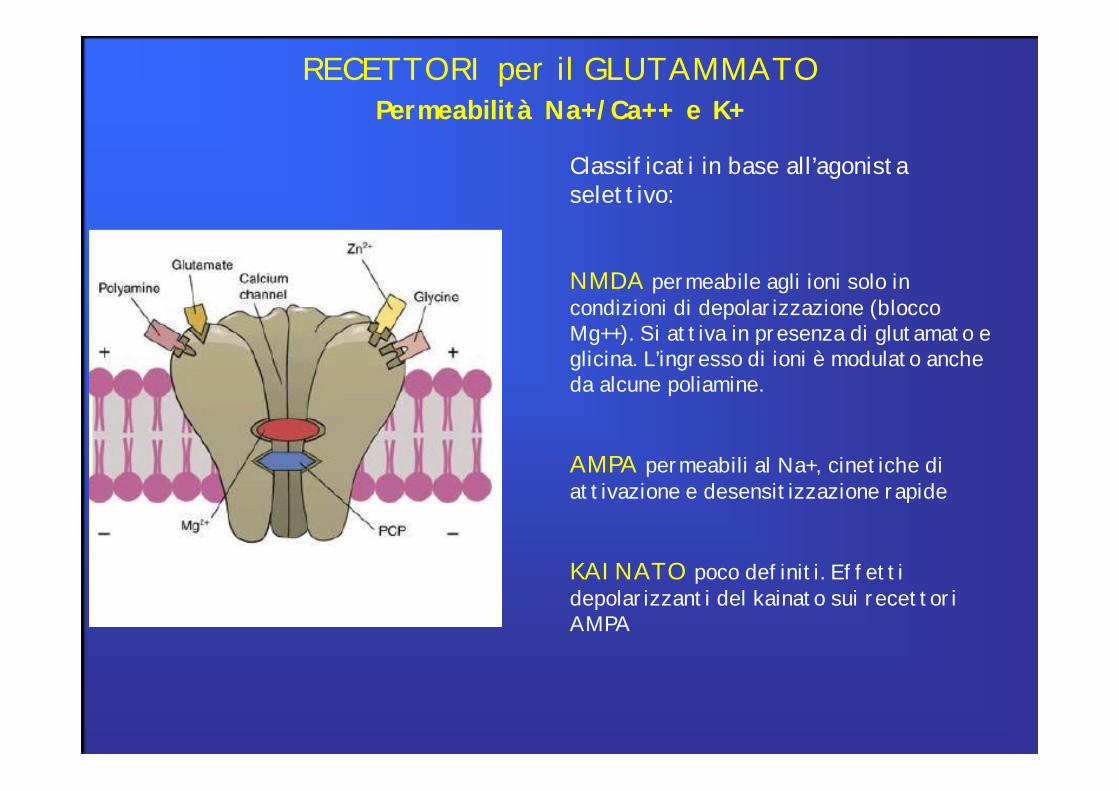
Permeabilità Na+/Ca++ e K+RECETTORI per il GLUTAMMATO
Classificati in base all’agonista selettivo:
NMDA permeabile agli ioni solo in condizioni di depolarizzazione (blocco Mg++). Si attiva in presenza di glutamato e glicina. L’ingresso di ioni è modulato anche da alcune poliamine.
AMPA permeabili al Na+, cinetiche di attivazione e desensitizzazione rapide
KAINATO poco definiti. Effetti depolarizzanti del kainato sui recettori AMPA
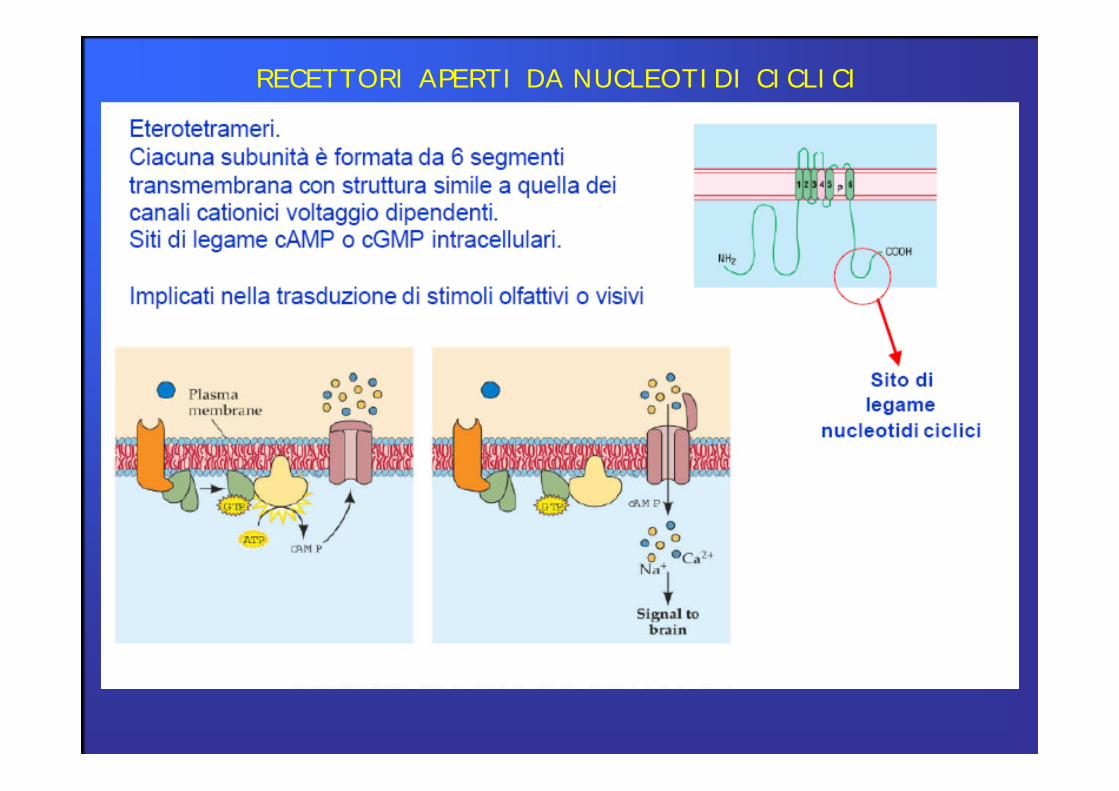
RECETTORI APERTI DA NUCLEOTIDI CICLICI
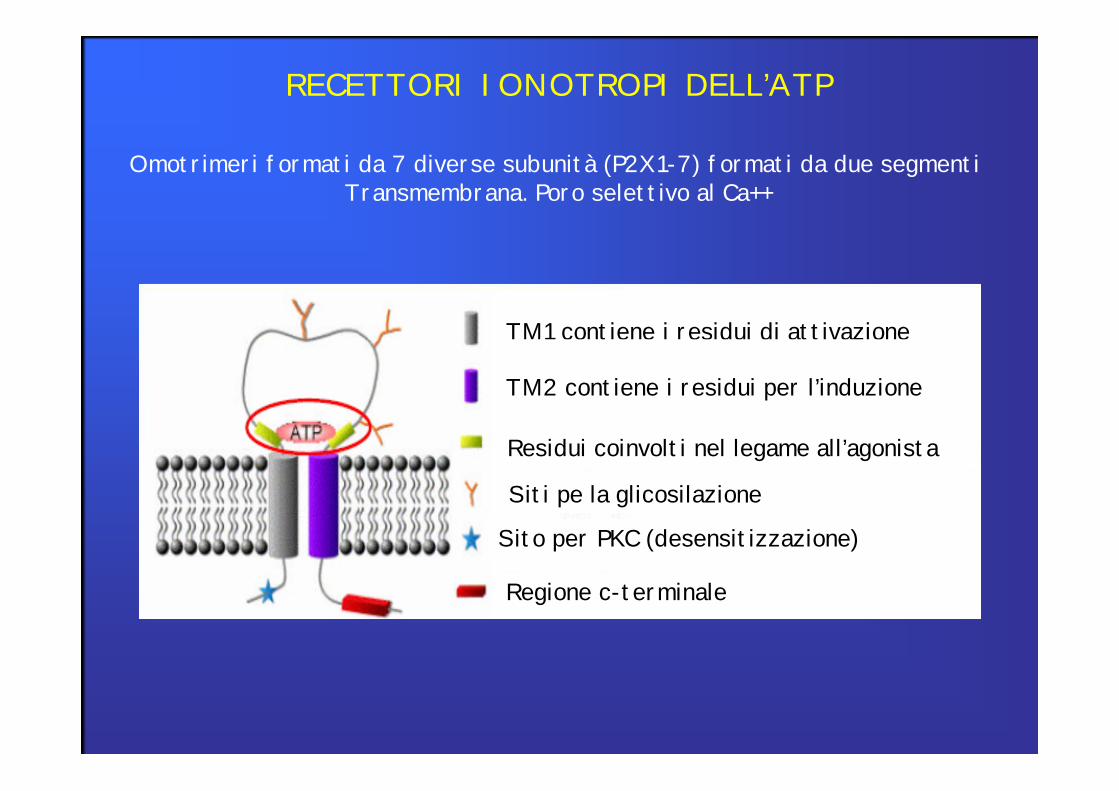
RECETTORI IONOTROPI DELL’ATP
Omotrimeri formati da 7 diverse subunità (P2X1-7) formati da due segmenti Transmembrana. Poro selettivo al Ca++
TM1 contiene i residui di attivazione
TM2 contiene i residui per l’induzione
Residui coinvolti nel legame all’agonista
Siti pe la glicosilazione
Sito per PKC (desensitizzazione)
Regione c-terminale
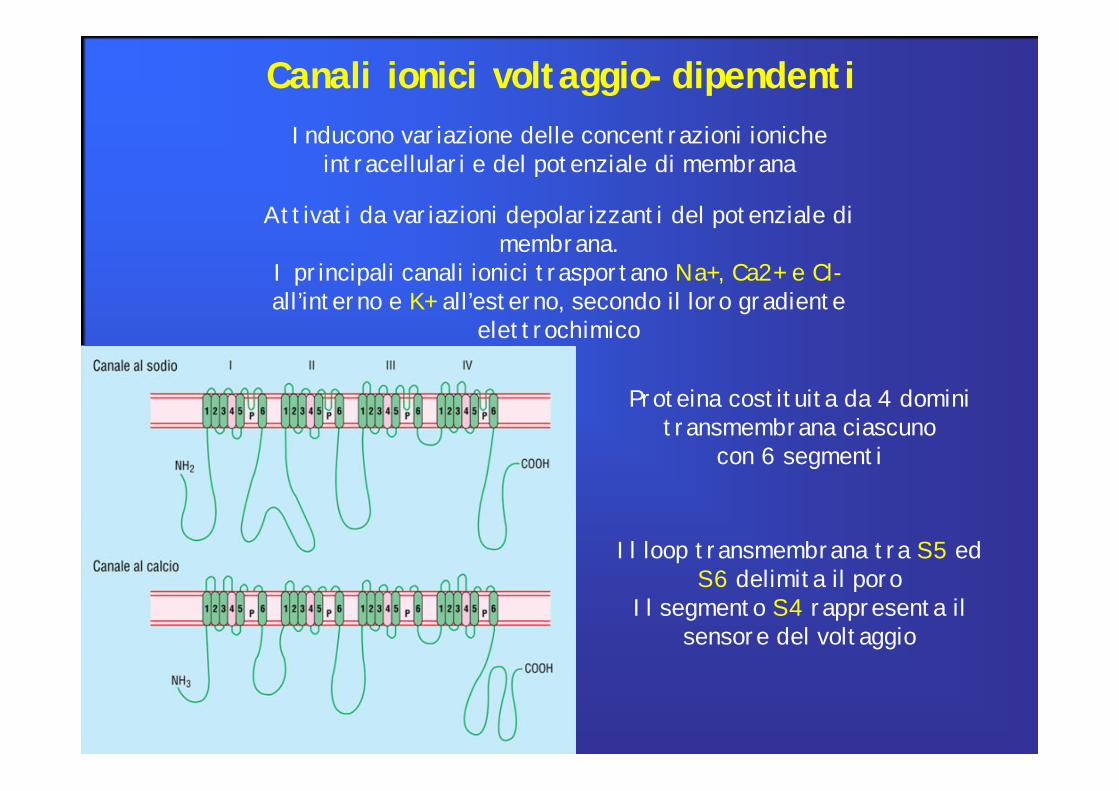
Canali ionici voltaggio-dipendentiInducono variazione delle concentrazioni ioniche
intracellulari e del potenziale di membrana
Attivati da variazioni depolarizzanti del potenziale di membrana.
I principali canali ionici trasportano Na+, Ca2+ e Cl-all’interno e K+ all’esterno, secondo il loro gradiente
elettrochimico
Proteina costituita da 4 domini transmembrana ciascuno
con 6 segmenti
Il loop transmembrana tra S5 edS6 delimita il poro
Il segmento S4 rappresenta ilsensore del voltaggio
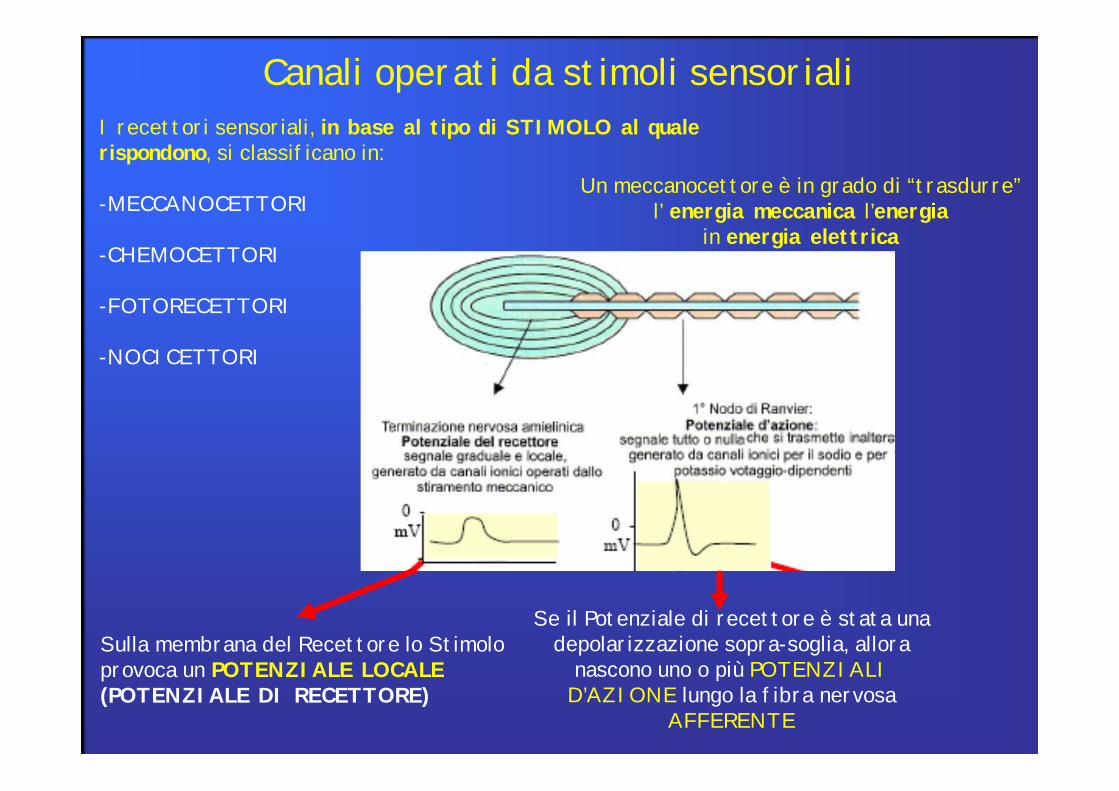
I recettori sensoriali, in base al tipo di STIMOLO al quale rispondono, si classificano in:
-MECCANOCETTORI
-CHEMOCETTORI
-FOTORECETTORI
-NOCICETTORI
Canali operati da stimoli sensoriali
Sulla membrana del Recettore lo Stimolo provoca un POTENZIALE LOCALE(POTENZIALE DI RECETTORE)
Se il Potenziale di recettore è stata una depolarizzazione sopra-soglia, allora
nascono uno o più POTENZIALI D’AZIONE lungo la fibra nervosa
AFFERENTE
Un meccanocettore è in grado di “trasdurre” l’ energia meccanica l’energia
in energia elettrica
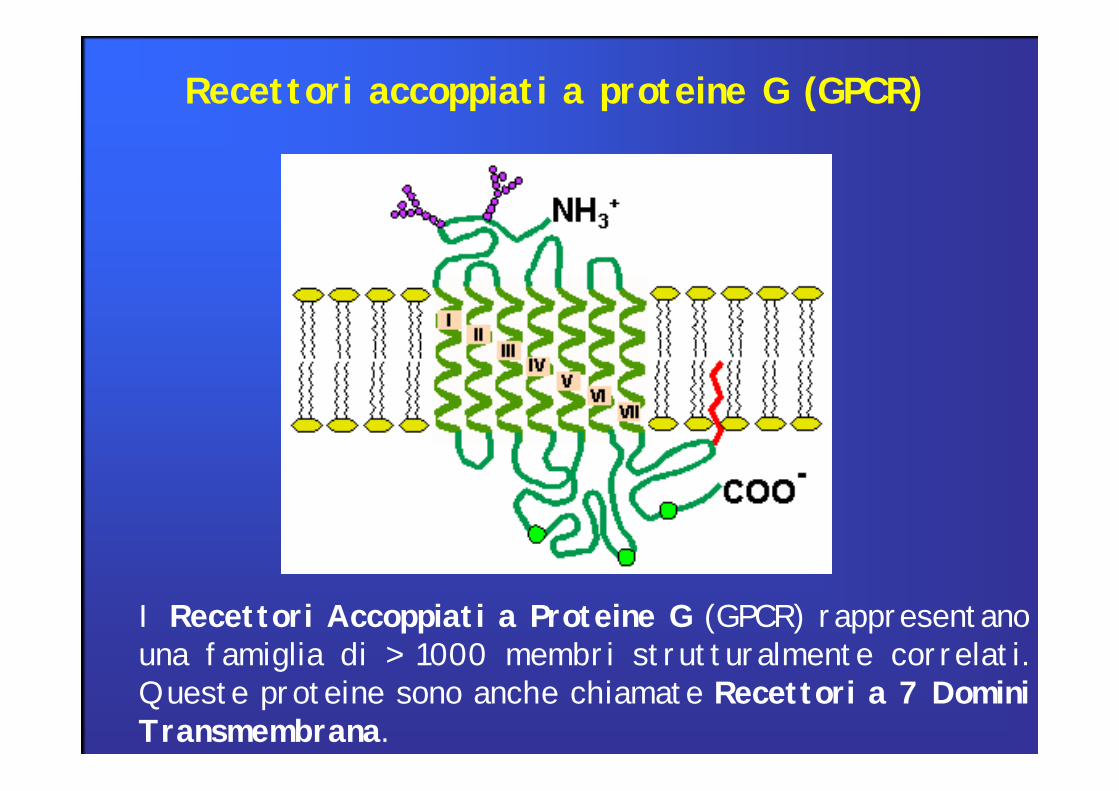
I Recettori Accoppiati a Proteine G (GPCR) rappresentanouna famiglia di > 1000 membri strutturalmente correlati.Queste proteine sono anche chiamate Recettori a 7 DominiTransmembrana.
Recettori accoppiati a proteine G (GPCR)
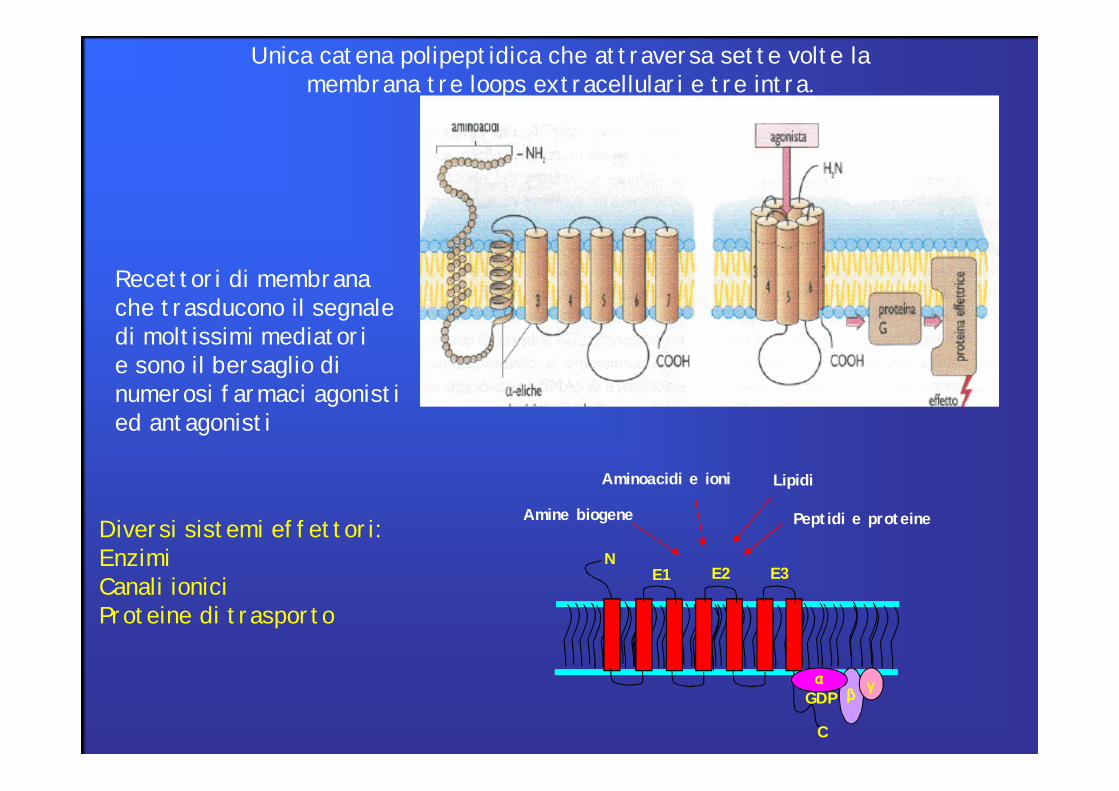
Recettori di membranache trasducono il segnaledi moltissimi mediatorie sono il bersaglio dinumerosi farmaci agonistied antagonisti
Unica catena polipeptidica che attraversa sette volte la membrana tre loops extracellulari e tre intra.
Diversi sistemi effettori:EnzimiCanali ioniciProteine di trasporto
N
C
E1 E2 E3
αβ γ
GDP
Amine biogene
Aminoacidi e ioni Lipidi
Peptidi e proteine
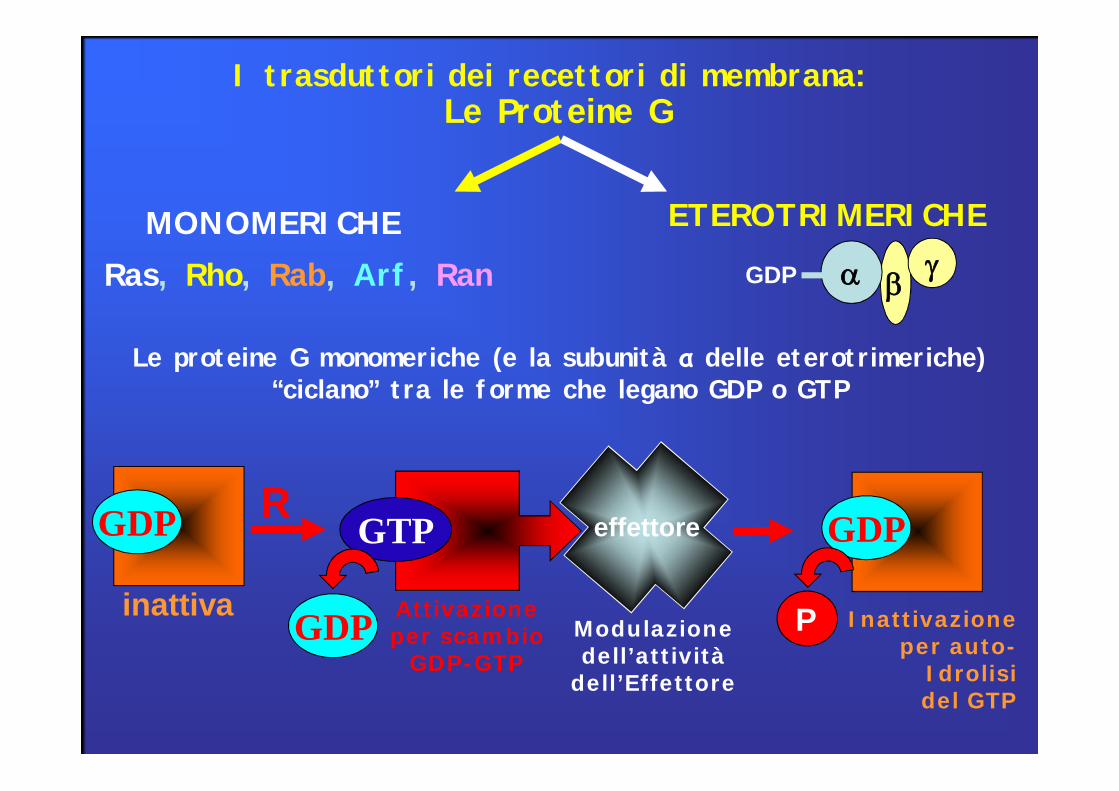
Le proteine G monomeriche (e la subunità α delle eterotrimeriche) “ciclano” tra le forme che legano GDP o GTP
GDP
inattiva
I trasduttori dei recettori di membrana: Le Proteine G
Modulazione dell’attività
dell’Effettore
effettoreR
GTP
Attivazioneper scambio
GDP-GTPGDP Inattivazione
per auto-Idrolisidel GTP
GDP
P
MONOMERICHERas, Rho, Rab, Arf, Ran
ETEROTRIMERICHE
bgaGDP

Le proteine G sono una famiglia di molecole proteiche, chiamate così per la loro interazione con i nucleotidi guaninici GTP e GDP
Ogni proteina G è formata da 3 subunità (α, β, γ)20 isoforme di subunità α5 isoforme di subunità β10 isoforme di subunità γ
N.B. la diversa composizione regola la selettività di accoppiamento tra proteine G e recettoriGTP e GDP si legano alla subunità α, che ha attività GTPasica intrinseca (GTP -> GDP)
β, γ sono-associate a formare il complesso βγ-idrofobiche-ancorate alla superficie citoplasmatica della membrana
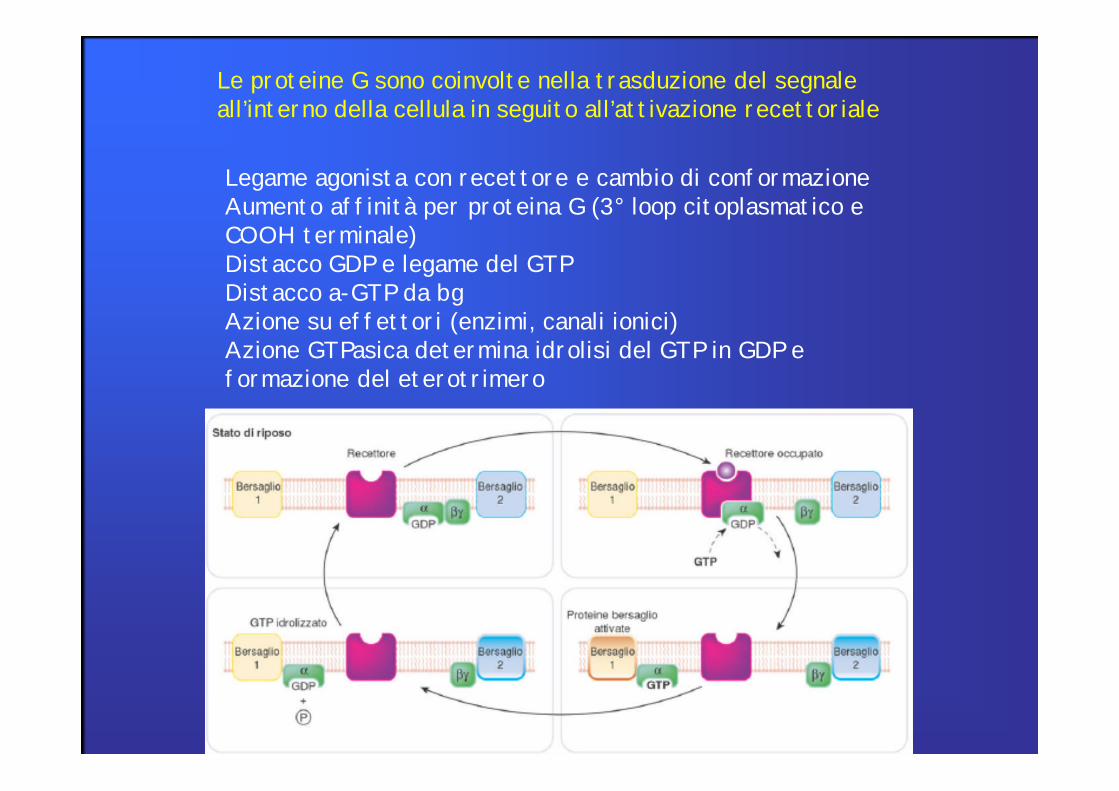
Le proteine G sono coinvolte nella trasduzione del segnale all’interno della cellula in seguito all’attivazione recettoriale
Legame agonista con recettore e cambio di conformazioneAumento affinità per proteina G (3° loop citoplasmatico e COOH terminale)Distacco GDP e legame del GTPDistacco a-GTP da bgAzione su effettori (enzimi, canali ionici)Azione GTPasica determina idrolisi del GTP in GDP e formazione del eterotrimero
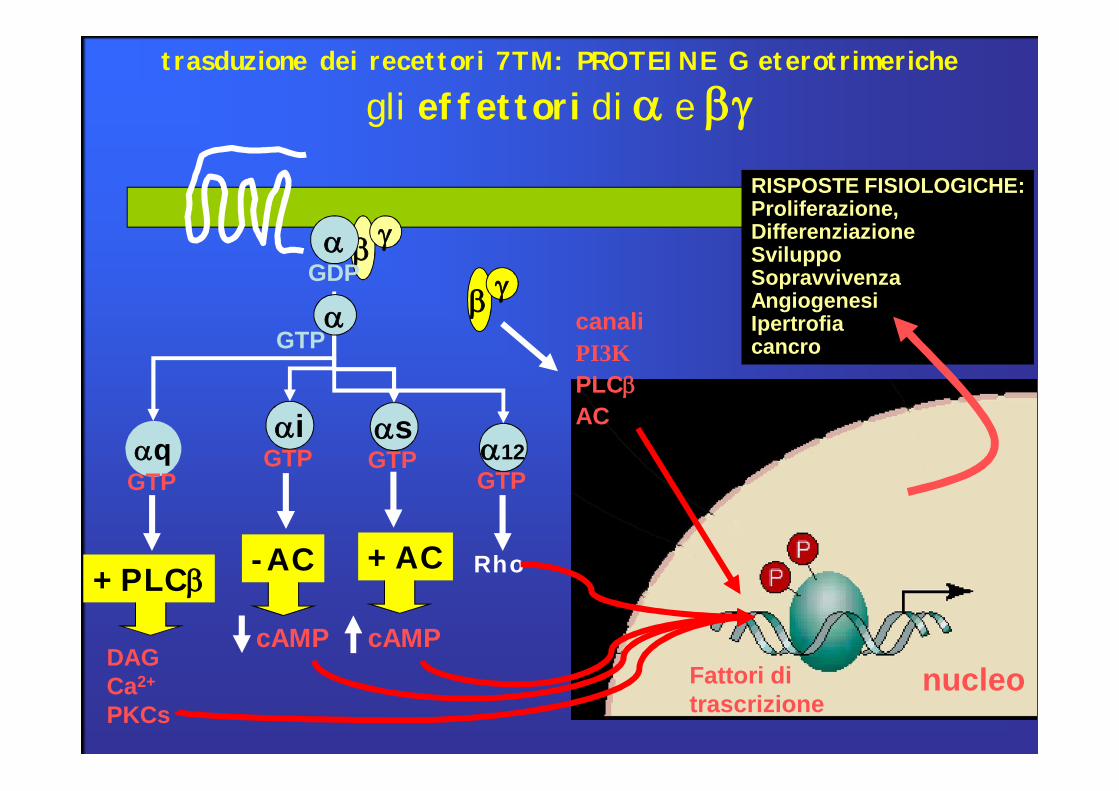
gli effettori di a e bgtrasduzione dei recettori 7TM: PROTEINE G eterotrimeriche
b gaGDP
a12GTP
Rho
regolazione dellaespressione genica
aiGTP
cAMP
-AC
asGTP
+AC
cAMP
aqGTP
DAGCa2+
PKCs
+PLCb
nucleoFattori ditrascrizione
b gcanaliPI3KPLCbAC
RISPOSTE FISIOLOGICHE:Proliferazione, DifferenziazioneSviluppoSopravvivenzaAngiogenesiIpertrofiacancro
aGTP
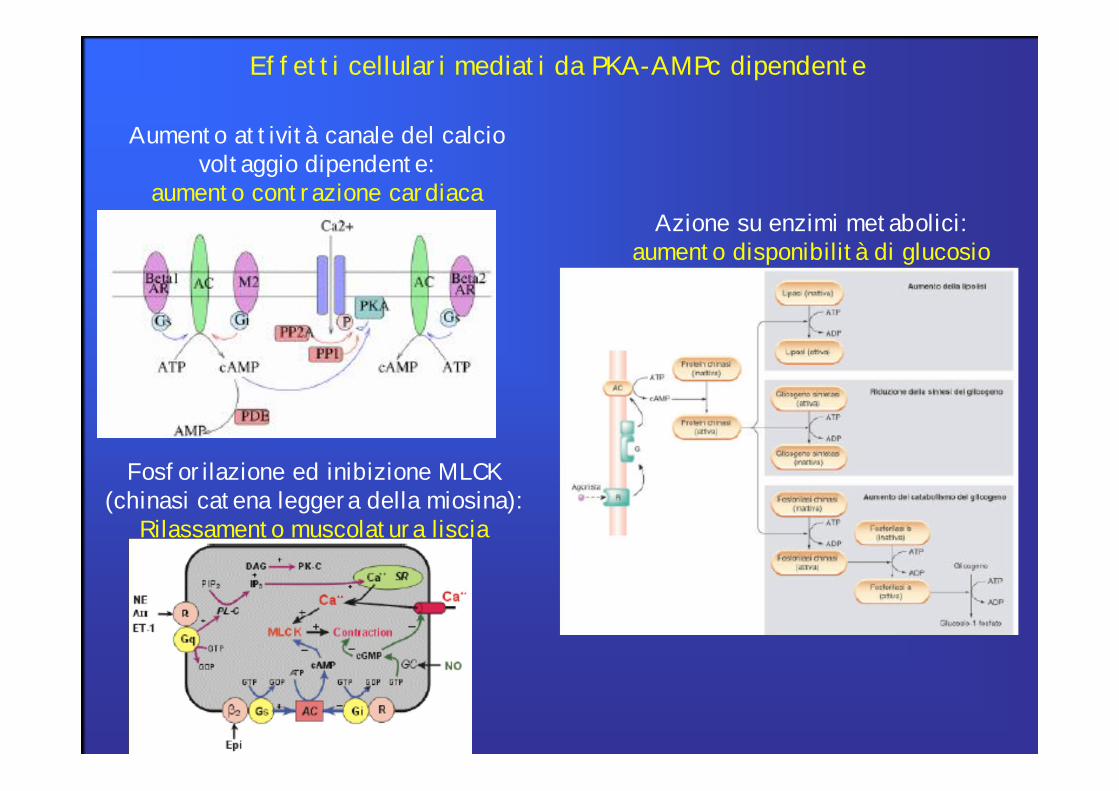
Aumento attività canale del calcio voltaggio dipendente:
aumento contrazione cardiacaAzione su enzimi metabolici:
aumento disponibilità di glucosio
Fosforilazione ed inibizione MLCK(chinasi catena leggera della miosina):
Rilassamento muscolatura liscia
Effetti cellulari mediati da PKA-AMPc dipendente
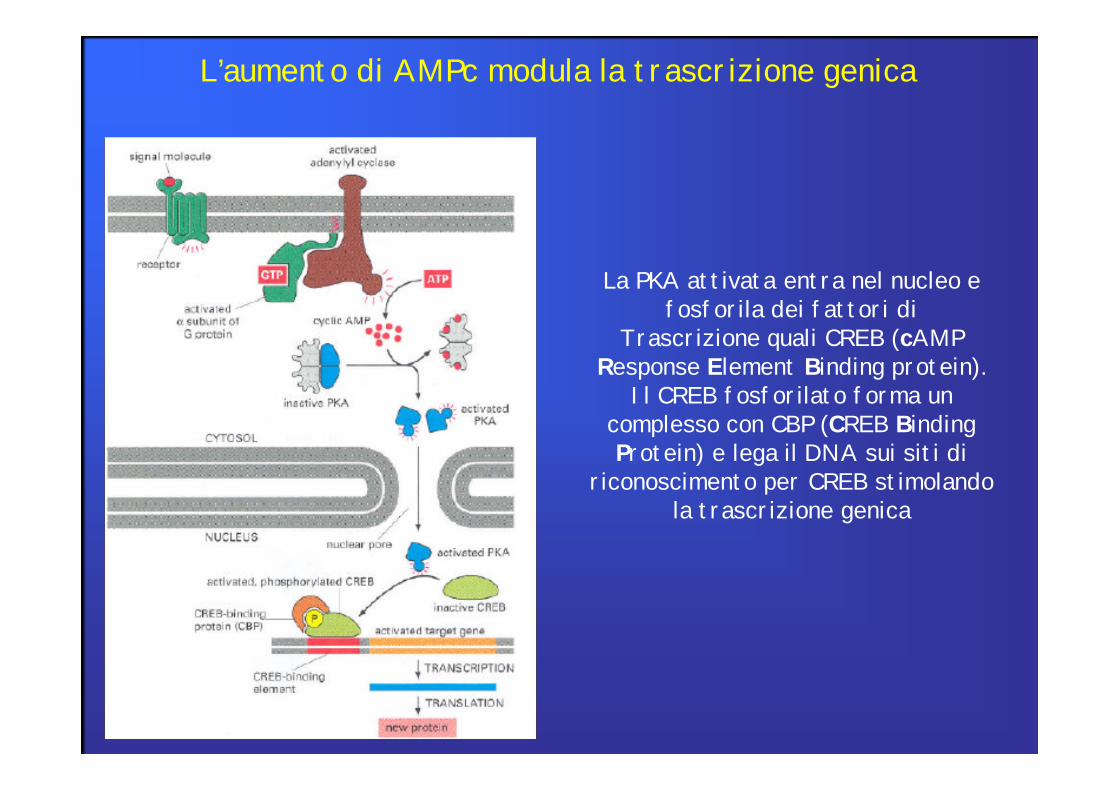
La PKA attivata entra nel nucleo efosforila dei fattori di
Trascrizione quali CREB (cAMPResponse Element Binding protein).
Il CREB fosforilato forma uncomplesso con CBP (CREB BindingProtein) e lega il DNA sui siti di
riconoscimento per CREB stimolandola trascrizione genica
L’aumento di AMPc modula la trascrizione genica
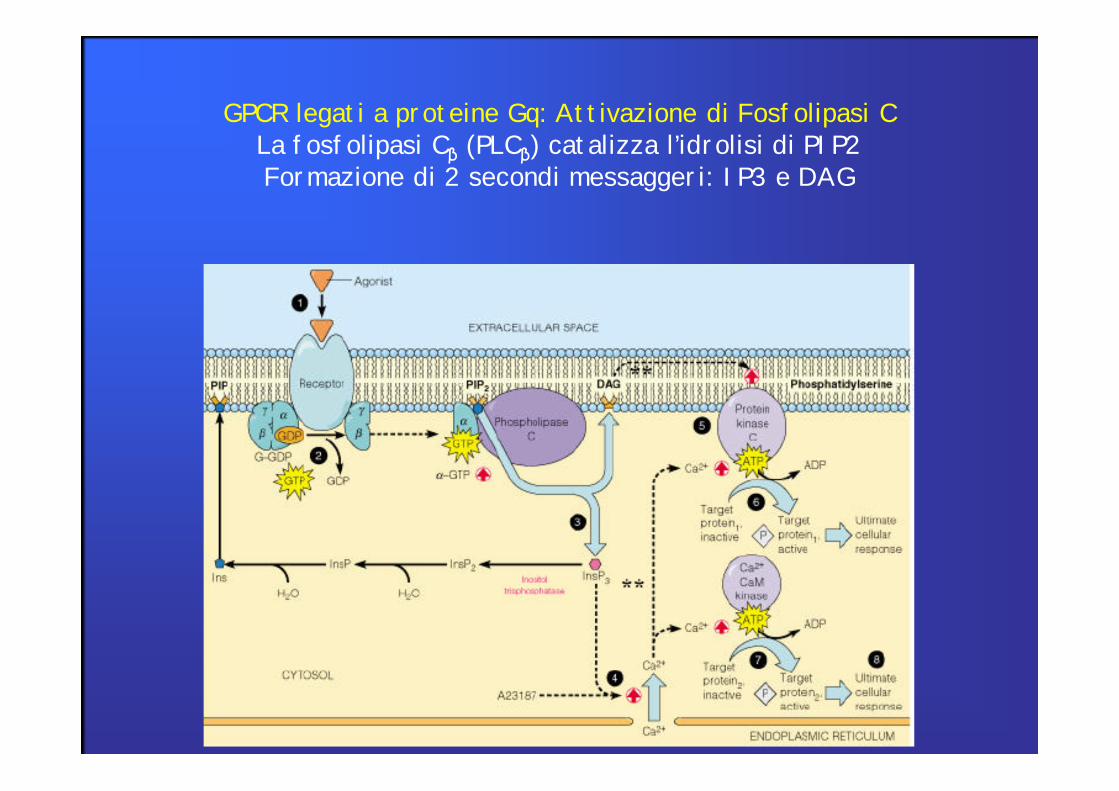
GPCR legati a proteine Gq: Attivazione di Fosfolipasi CLa fosfolipasi Cβ (PLCβ) catalizza l’idrolisi di PIP2 Formazione di 2 secondi messaggeri: IP3 e DAG
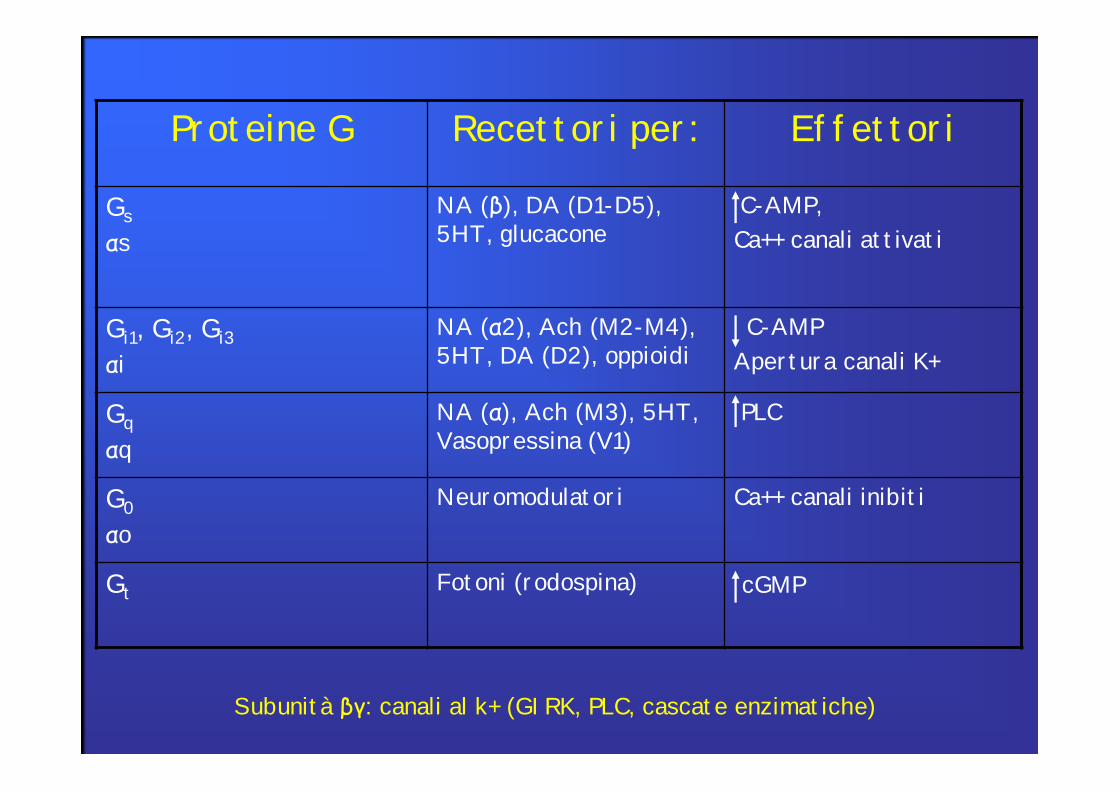
Proteine G Recettori per: Effettori
Gs αs
NA (β), DA (D1-D5), 5HT, glucacone
C-AMP, Ca++ canali attivati
Gi1, Gi2, Gi3αi
NA (α2), Ach (M2-M4), 5HT, DA (D2), oppioidi
C-AMPApertura canali K+
Gq αq
NA (α), Ach (M3), 5HT, Vasopressina (V1)
PLC
G0 αo
Neuromodulatori Ca++ canali inibiti
Gt Fotoni (rodospina) cGMP
Subunità βγ: canali al k+ (GIRK, PLC, cascate enzimatiche)

trasduzione dei recettori 7TM: PROTEINE G monomeriche
Piccole proteine intracellulari (20-40 kD) in grado di legare ed idrolizzare il GTP
•Attività indipendente dall’attivazione dei classici GPCR ma spesso dovuta all’attivazione di altri tipi recettoriali (ad es. Recettori-kinasi)
•Lo scambio GDP-GTP (attivazione) è regolato da interazione con specifiche proteine, quali ad esempio SOS
•Le proteine G monomeriche attivate interagiscono con sistemi effettori intracellulari

Ras: Attivazione della cascata MAP chinasi ERK-1 e 2Rap-1: Inibitore di Ras e attivazione MAP chinasi ERK-
1 e 2 Ras indipendenteRac: Controllo risposta a stress cellulare (apoptosi da
MAP kinasi JNK)Rab: Trasporto e esocitosi vescicoleRho: Assemblaggio del citoscheletroARF: Assemblaggio apparato Golgi
EF-2: Regolazione sintesi proteica nei ribosomiRan: Scambio di RNA e proteine tra nucleo e
ciotoplasma
trasduzione dei recettori 7TM: PROTEINE G monomeriche
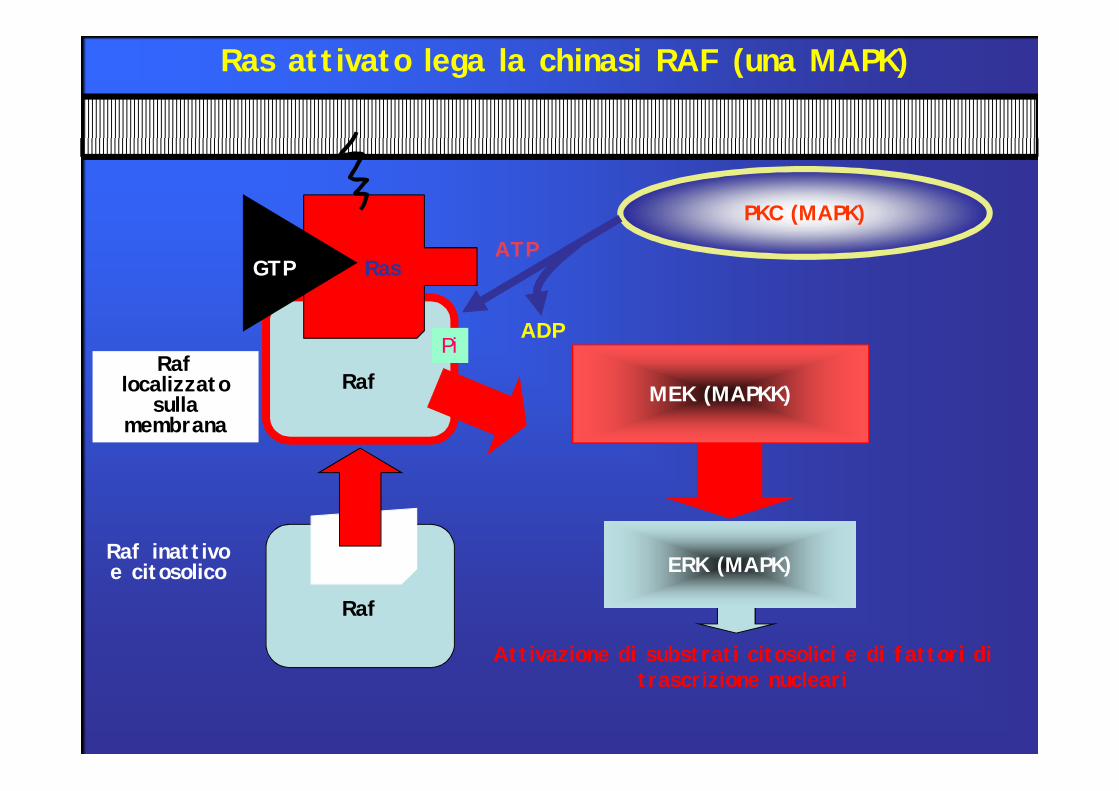
Attivazione di substrati citosolici e di fattori di trascrizione nucleari
RafRaf
localizzato sulla
membrana
ERK (MAPK)
MEK (MAPKK)
PKC (MAPK)
Pi
ATP
ADP
Ras attivato lega la chinasi RAF (una MAPK)
GTP Ras
Raf inattivo e citosolico
Raf

Spegnimento della subunità a della proteina G
La subunita α è dotata di una attività GTPasica intrinseca: idrolizza GTP a GDP e Pi e si riassocia alle subunità βγ per riformare la proteina G inattiva

Le proteine RGS (regolatrici della segnalazione della
proteina G)
• Sono proteine che attivano la GTPasi • Ruolo cruciale nello spegnimento di risposte
mediate da proteine G• Circa 25 proteine codificate nel genoma
umano: ognuna interagisce con una serie di proteine G
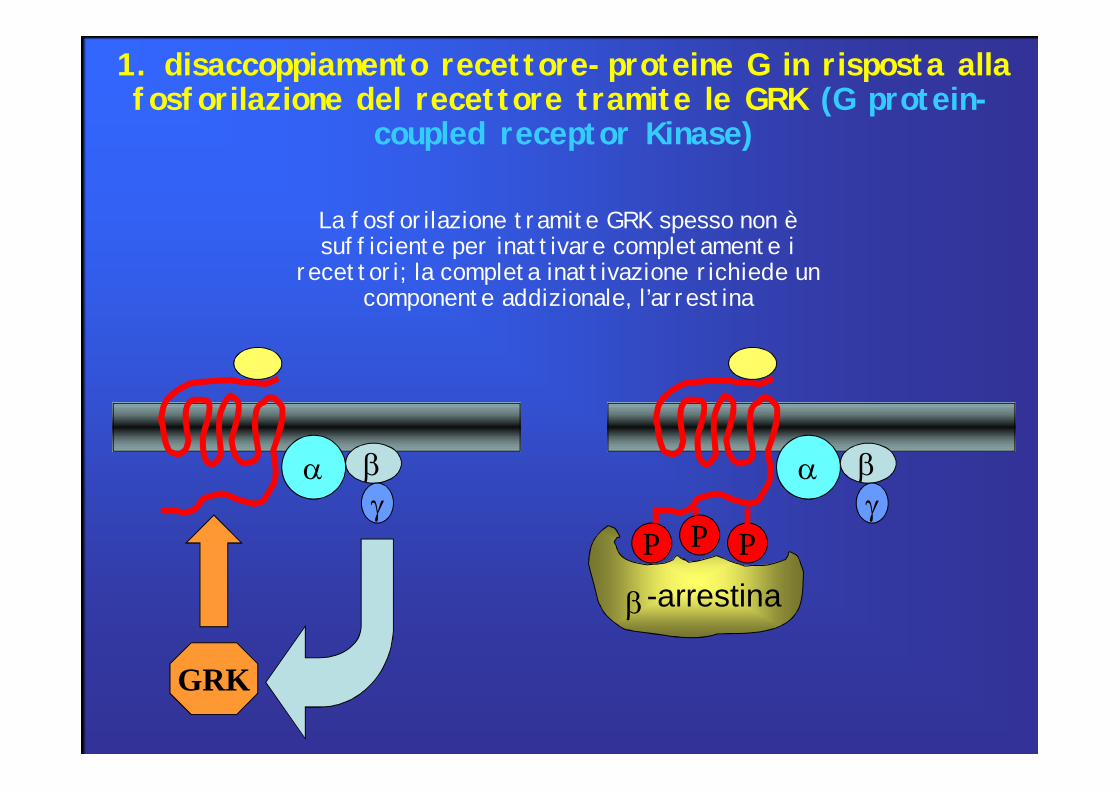
GRK
P PP
-arrestina
La fosforilazione tramite GRK spesso non è sufficiente per inattivare completamente i
recettori; la completa inattivazione richiede un componente addizionale, l’arrestina
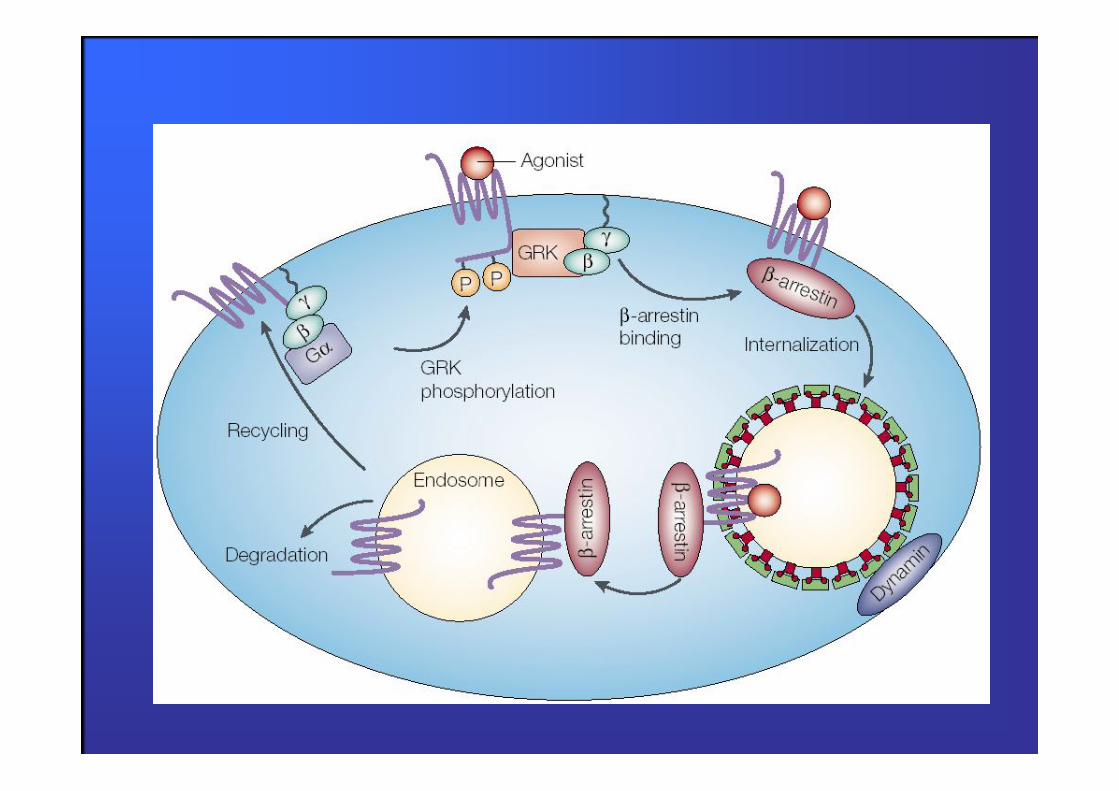
1. disaccoppiamento recettore-proteine G in risposta alla fosforilazione del recettore tramite le GRK (G protein-
coupled receptor Kinase)

RECETTORI AD ATTIVITA’ ENZIMATICA
Recettori con attività tirosin-chinasica intrinseca
Attivazione recettoriale determina l’attivazione dell’attività enzimatica ed il trasferimento di gruppi fosfato su residuidi tirosina:
Enzimi regolati da ligandi endogeni
Recettori per ormoni peptidici (insulina) e per fattori di crescita (IGF; VEGF;
PDGF).
Regolano attività metaboliche, crescita, differenziazione e sviluppo

1. Recettori chinasici che agiscono fosforilando i residui di serina e treonina:Recettore per TGF-β
2. Recettori con altre attività enzimaticheDominio intracellulare è una guanilciclasi che sintetizza GMP ciclico che agisce come 2° messaggero (simile all’ cAMP, attiva una PKG).Recettore per il peptide natriuretico atriale
3. Recettori chinasici senza dominio catalitico intracellulare: legano o attivano chinasi di membrana o citoplasmatiche.Recettori per peptidi neurotroficiRecettori per antigeni a subunità multipla sui linfociti T e B
RECETTORI AD ATTIVITA’ ENZIMATICA
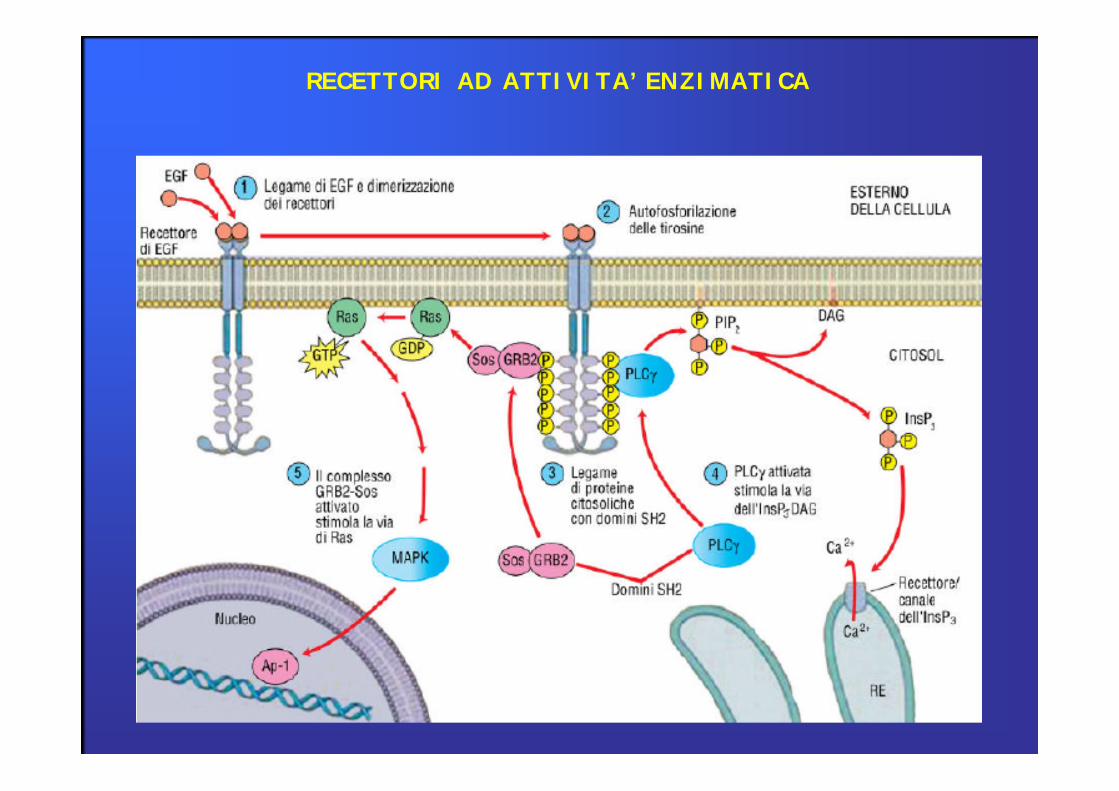
RECETTORI AD ATTIVITA’ ENZIMATICA
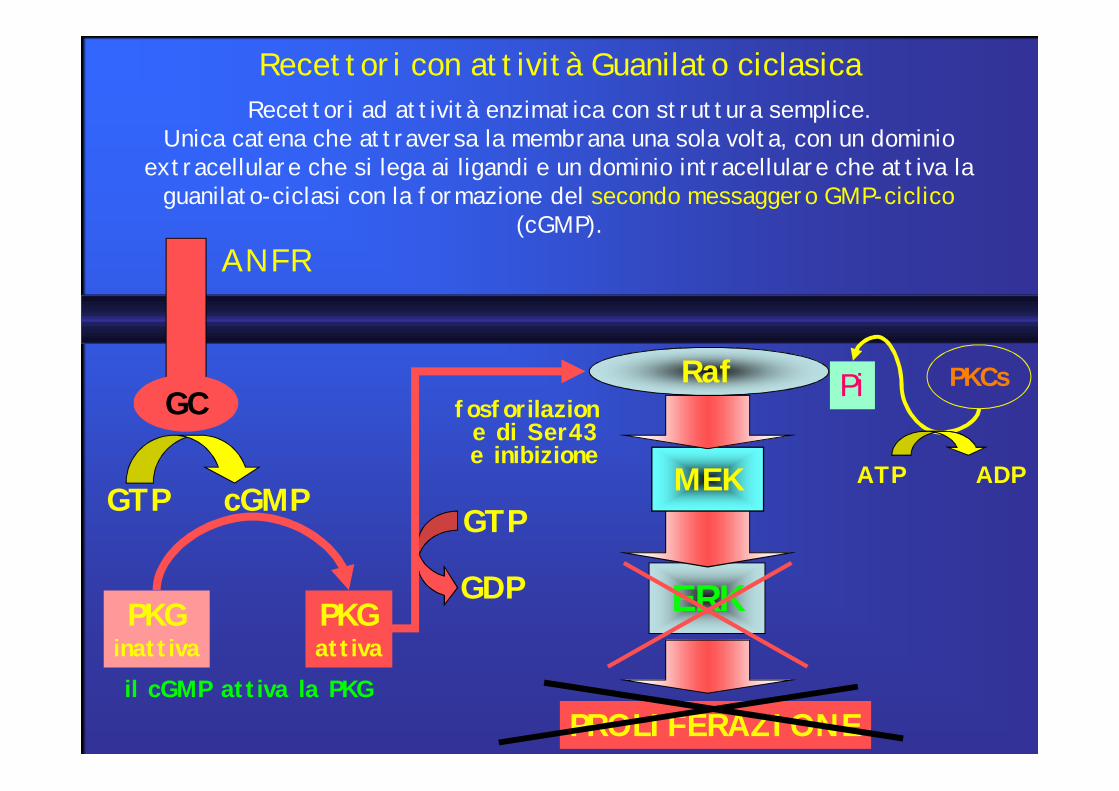
GC
ANFR
GTP cGMP
il cGMP attiva la PKG
PKGinattiva
PKGattiva
ERK
MEK
PKCsPiRaf
ATP ADP
PROLIFERAZIONE
GTP
GDP
fosforilazione di Ser43e inibizione
Recettori con attività Guanilato ciclasicaRecettori ad attività enzimatica con struttura semplice.
Unica catena che attraversa la membrana una sola volta, con un dominio extracellulare che si lega ai ligandi e un dominio intracellulare che attiva la
guanilato-ciclasi con la formazione del secondo messaggero GMP-ciclico(cGMP).

Recettori con attività Guanilato ciclasica
• Nel rene controllo del trasporto ionico e la ritenzione d’acqua, mediante il fattore natriuretico atriale (AFN). L’AFN giunge al rene dove attiva la guanilil-ciclasi nelle cellule dei dotti collettori; l’aumento di cGMP incrementa la velocità di escrezione di Na+ e di conseguenza di acqua.
•Nell’intestino, attivazione da parte peptide intestinale, la guanilina, che regola la secrezione di Cl- nell’intestino. Anche bersaglio di un’endotossina prodotta da E. coli e da altri batteri Gram-negativi. L’icremento della concentrazione di cGMP prodotto dall’endotossina aumenta la secrezione di Cl- e di conseguenza diminuisce il riassorbimento di acqua, causando diarrea.
•Nel muscolo liscio il cGMP determina rilassamento. La guanilil-ciclasi citosolica con un gruppo eme saldamente legato è attivata dall’ossido di azoto (NO). L’aumento di cGMP determina vasodilatazione. Nel cuore, inoltre, il cGMP riduce la forza contrattile stimolando una pompa ionica che espelle dal citosol ioni calcio.
•Nel cervello il cGMP è coinvolto nei processi di sviluppo e funzionamento.
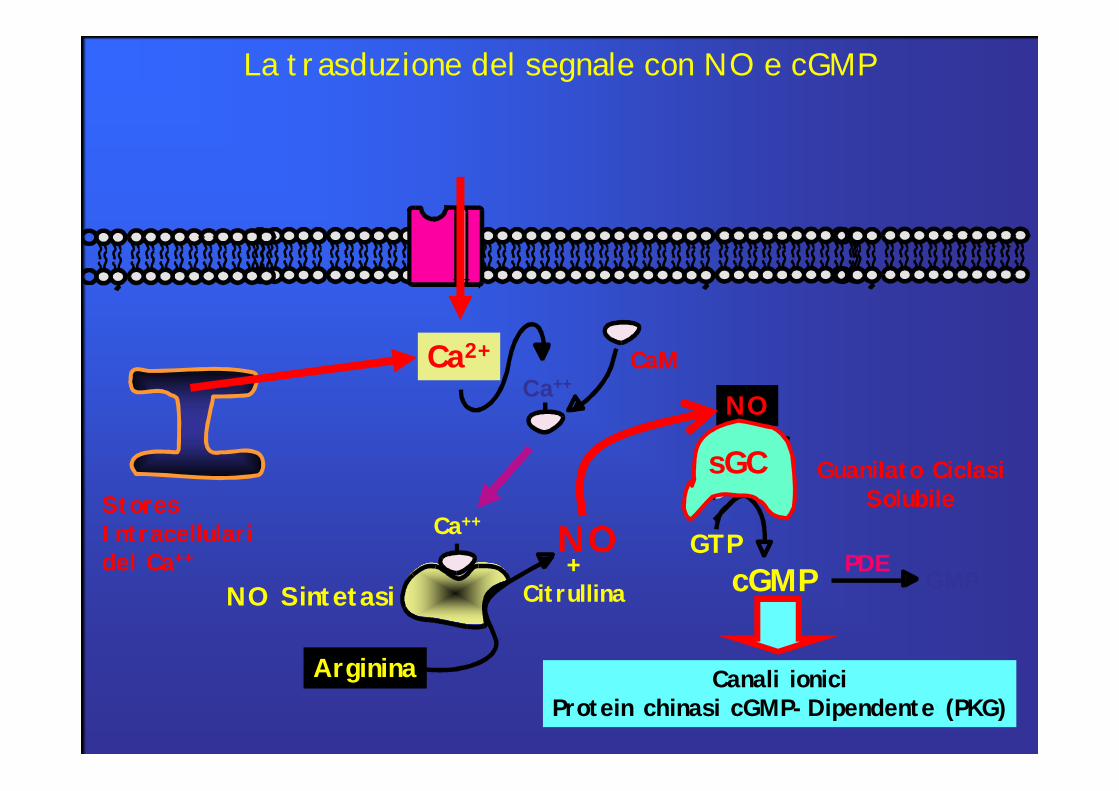
StoresIntracellulari del Ca++
La trasduzione del segnale con NO e cGMP
Ca2+
NO
Arginina
+CitrullinaNO Sintetasi
Ca++
Ca++CaM
cGMPGTP
NO
Guanilato Ciclasi Solubile
sGC
Canali ioniciProtein chinasi cGMP-Dipendente (PKG)
GMPPDE
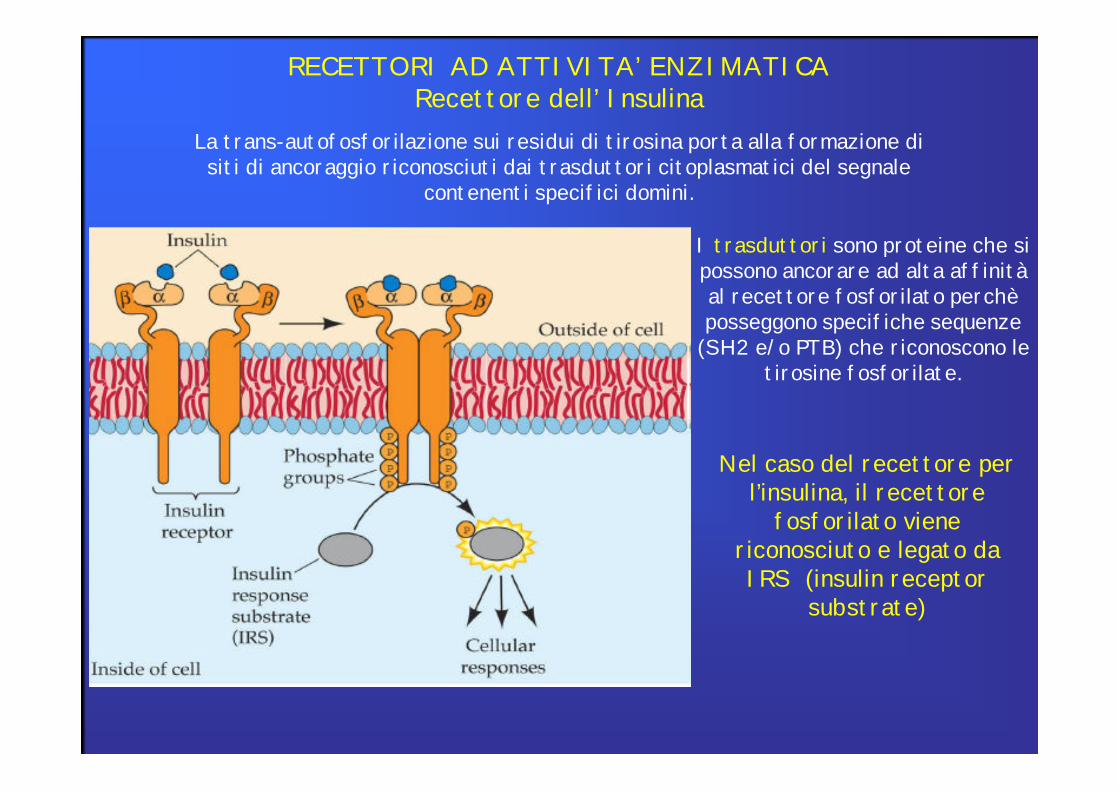
RECETTORI AD ATTIVITA’ ENZIMATICARecettore dell’ Insulina
La trans-autofosforilazione sui residui di tirosina porta alla formazione di siti di ancoraggio riconosciuti dai trasduttori citoplasmatici del segnale
contenenti specifici domini.
I trasduttori sono proteine che sipossono ancorare ad alta affinitàal recettore fosforilato perchèposseggono specifiche sequenze
(SH2 e/o PTB) che riconoscono le tirosine fosforilate.
Nel caso del recettore per l’insulina, il recettore
fosforilato viene riconosciuto e legato da IRS (insulin receptor
substrate)
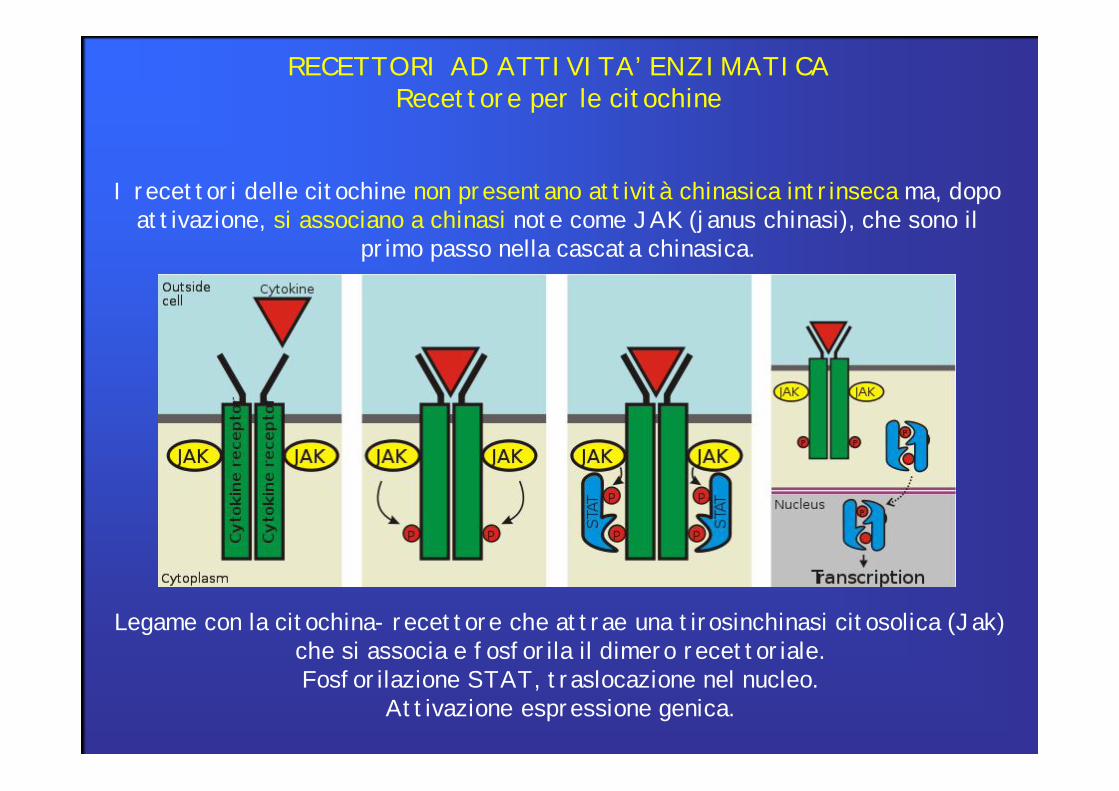
RECETTORI AD ATTIVITA’ ENZIMATICARecettore per le citochine
I recettori delle citochine non presentano attività chinasica intrinseca ma, dopo attivazione, si associano a chinasi note come JAK (janus chinasi), che sono il
primo passo nella cascata chinasica.
Legame con la citochina- recettore che attrae una tirosinchinasi citosolica (Jak) che si associa e fosforila il dimero recettoriale.Fosforilazione STAT, traslocazione nel nucleo.
Attivazione espressione genica.

Approcci farmacologici:Coinvolgimento di alcuni di questi recettori in disturbi metabolici e in processi tumorali.
Inibitori interazione ligando-recettoreAnticorpi monoclonali contro epitopi extracellulari deirecettori per fattori di crescitaHerceptin : anticorpo recettore EGFAvastin : anticorpo recettore VEGF (inibitore di angiogenesi)
Inibitori attività catalitica tirosin-chinasicaComposti naturali (quercetina, genisteina) mimano il compostointermedio che si forma tra ATP ed il residuo di tirosina durante lareazione chinasica (molto tossici)Composti di sintesi (gefitinib per EGRF; PTK787 per VEGFR; etc)
Inibitori della farnesil-transferasi:Enzima che catalizza la farnesilazione di Ras
RECETTORI AD ATTIVITA’ ENZIMATICA

Recettori per l’adesione cellulareTrasmissione del segnale all’interno della cellula per la regolazione di funzioni quali
Movimento, proliferazione, sopravvivenza cellulare e trascrizione genica
Immunoglobuline (ICAM-1, ICAM-2, VCAM, CD31)Mediano le interazione tra linfociti e cellule endoteliali
Caderine partecipano alla formazione delle“giunzioni aderenti”
Selectine espresse sui leucociti e cellule endoteliali.Migrano dal sangue all’interstizio sottoendoteliale in presenza di infezione o danno tissutale.
Integrine mediano l’adesione delle cellule alleproteine della matrice extracellulare
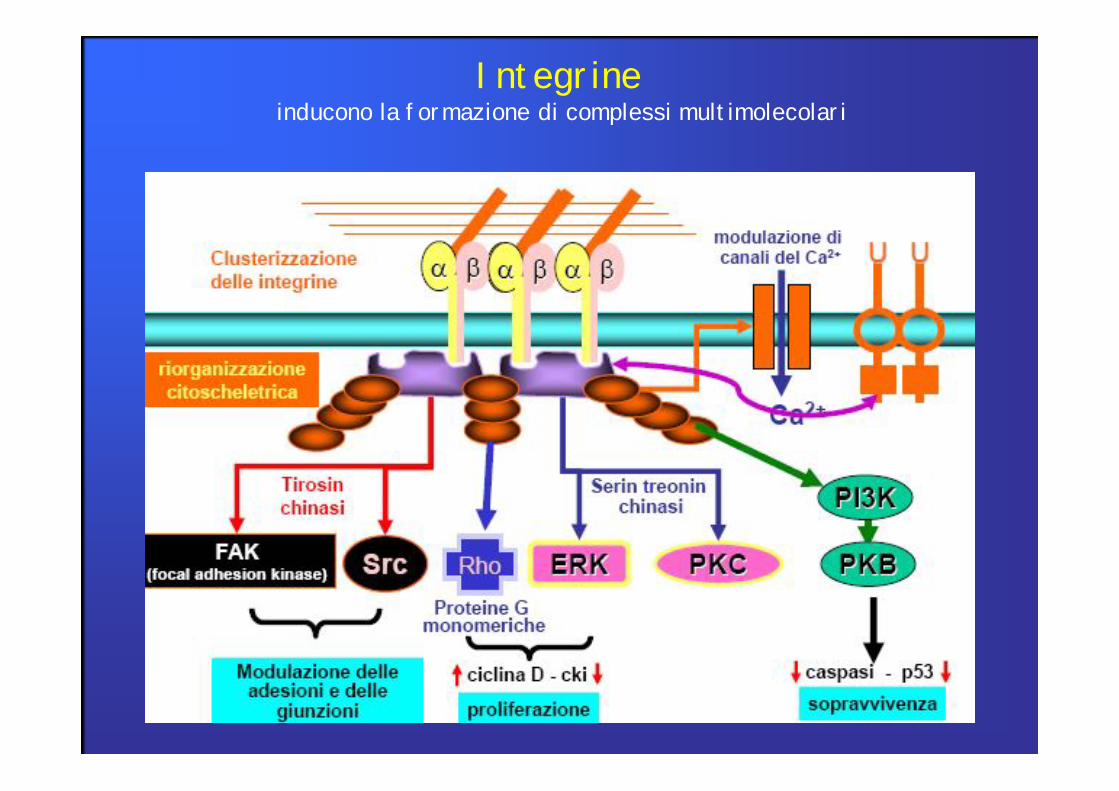
Integrineinducono la formazione di complessi multimolecolari

N
Recettorenucleare
1. Recettori per gli ormoni STEROIDEIaldosterone, cortisolo, estradiolo, progesterone, testosterone
2. Recettore per la vitamina D3 attivata:VDR (1,25-diOH-colecalciferolo)
3. J2R (15-deoxyD12,14-prostaglandina J2)
4. Recettori per i retinoidiRXR (9-cis-acido retinoico)RAR (all-trans-acido retinoico)
5. Recettori epaticiLXR ((7a-) (20-) (22-) (24-) (27-)-OH-colesterolo
6. Recettori per gli ormoni tiroideiTR (T3, T4)
Recettori Nucleari
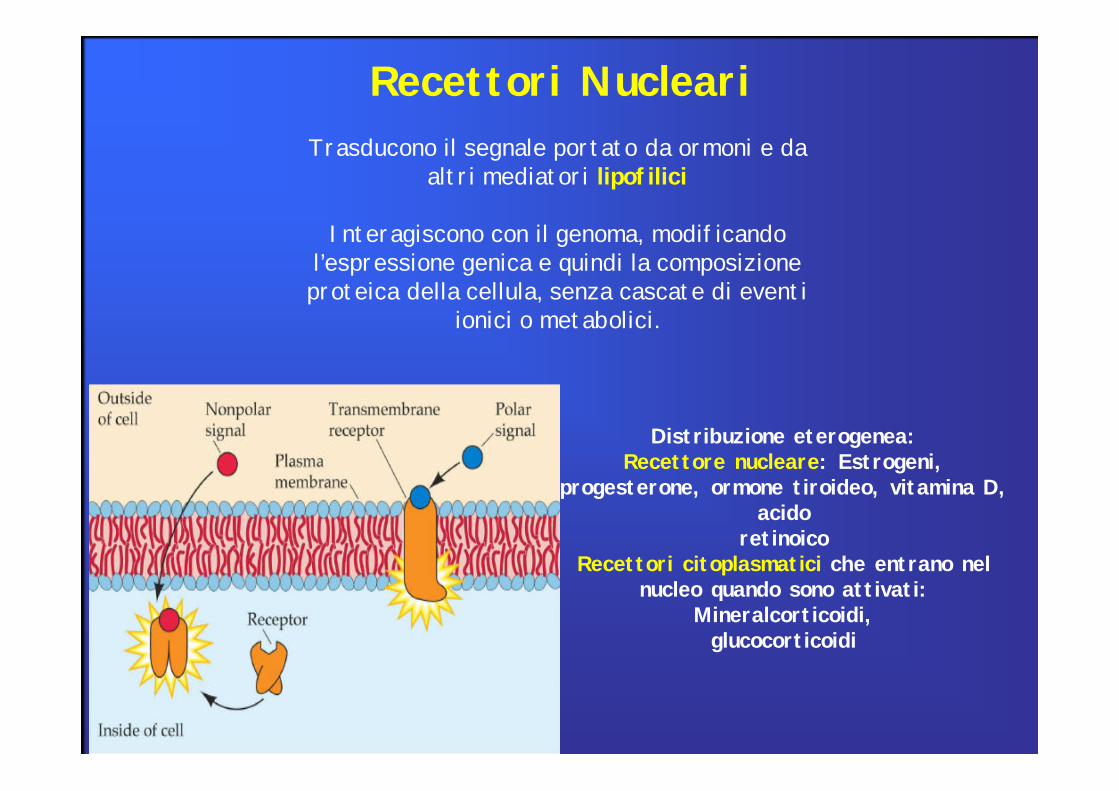
Trasducono il segnale portato da ormoni e da altri mediatori lipofilici
Interagiscono con il genoma, modificando l’espressione genica e quindi la composizione proteica della cellula, senza cascate di eventi
ionici o metabolici.
Distribuzione eterogenea:Recettore nucleare: Estrogeni,
progesterone, ormone tiroideo, vitamina D, acido
retinoicoRecettori citoplasmatici che entrano nel
nucleo quando sono attivati: Mineralcorticoidi,glucocorticoidi
Recettori Nucleari
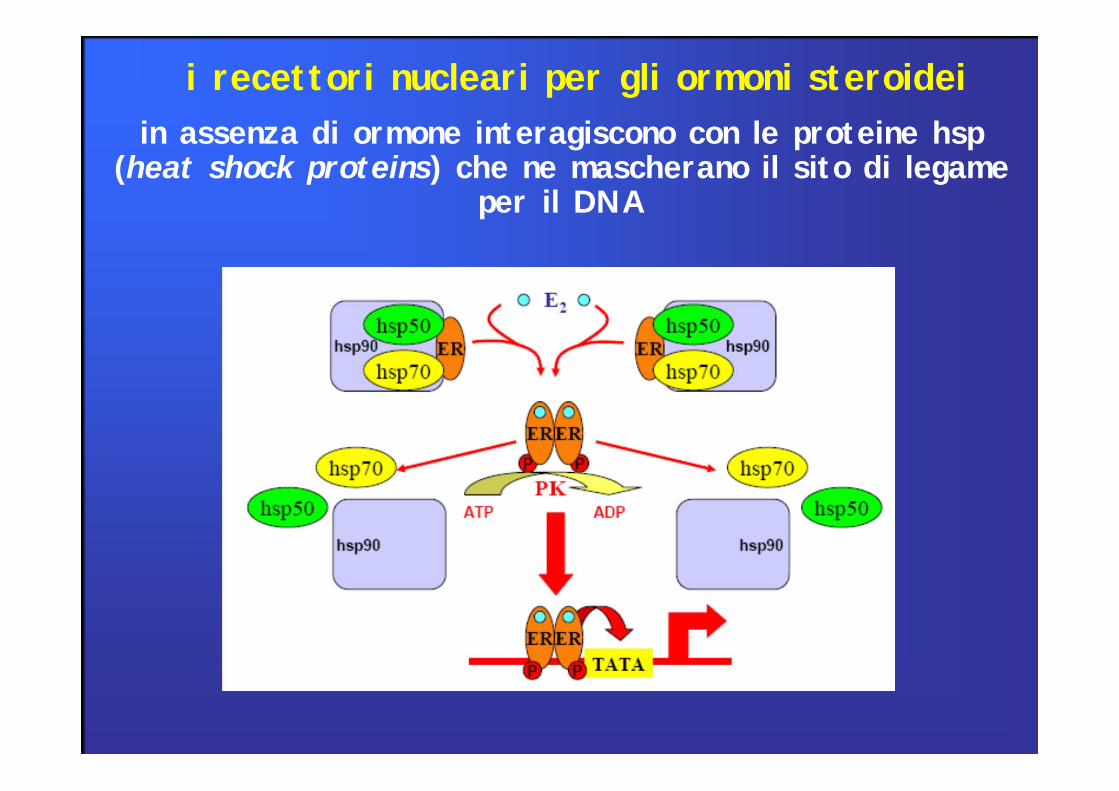
in assenza di ormone interagiscono con le proteine hsp (heat shock proteins) che ne mascherano il sito di legame
per il DNA
i recettori nucleari per gli ormoni steroidei
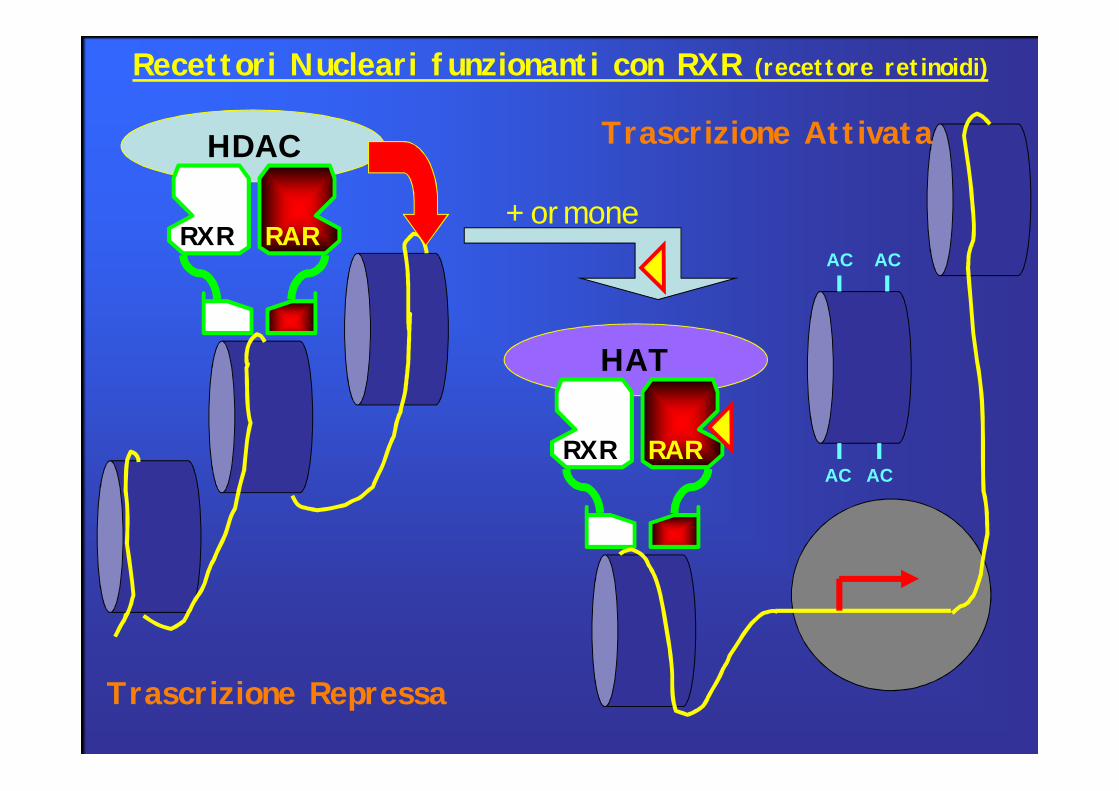
HDAC
RXR RAR
Trascrizione Repressa
HAT
RXR RAR
+ ormoneAC AC
AC AC
Trascrizione Attivata
Recettori Nucleari funzionanti con RXR (recettore retinoidi)
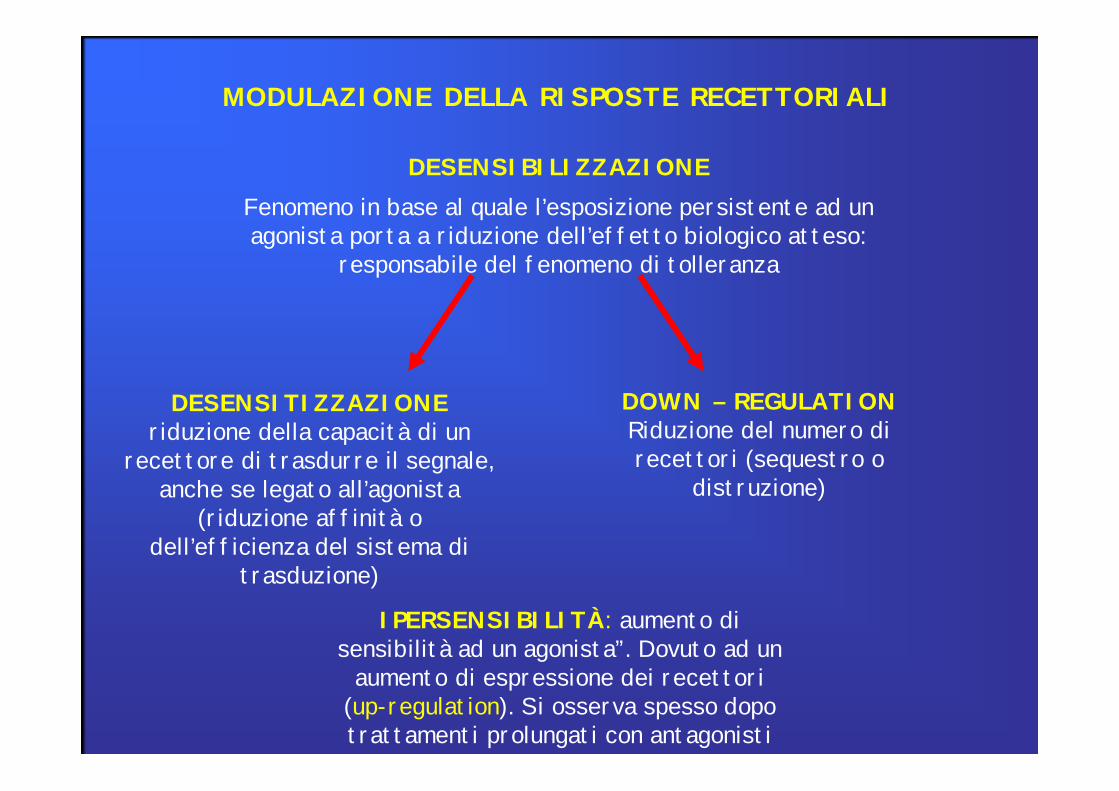
MODULAZIONE DELLA RISPOSTE RECETTORIALI
DESENSIBILIZZAZIONEFenomeno in base al quale l’esposizione persistente ad un agonista porta a riduzione dell’effetto biologico atteso:
responsabile del fenomeno di tolleranza
DESENSITIZZAZIONEriduzione della capacità di un
recettore di trasdurre il segnale,anche se legato all’agonista
(riduzione affinità odell’efficienza del sistema di
trasduzione)
DOWN – REGULATIONRiduzione del numero direcettori (sequestro o
distruzione)
IPERSENSIBILITÀ: aumento di sensibilità ad un agonista”. Dovuto ad un
aumento di espressione dei recettori(up-regulation). Si osserva spesso dopo trattamenti prolungati con antagonisti
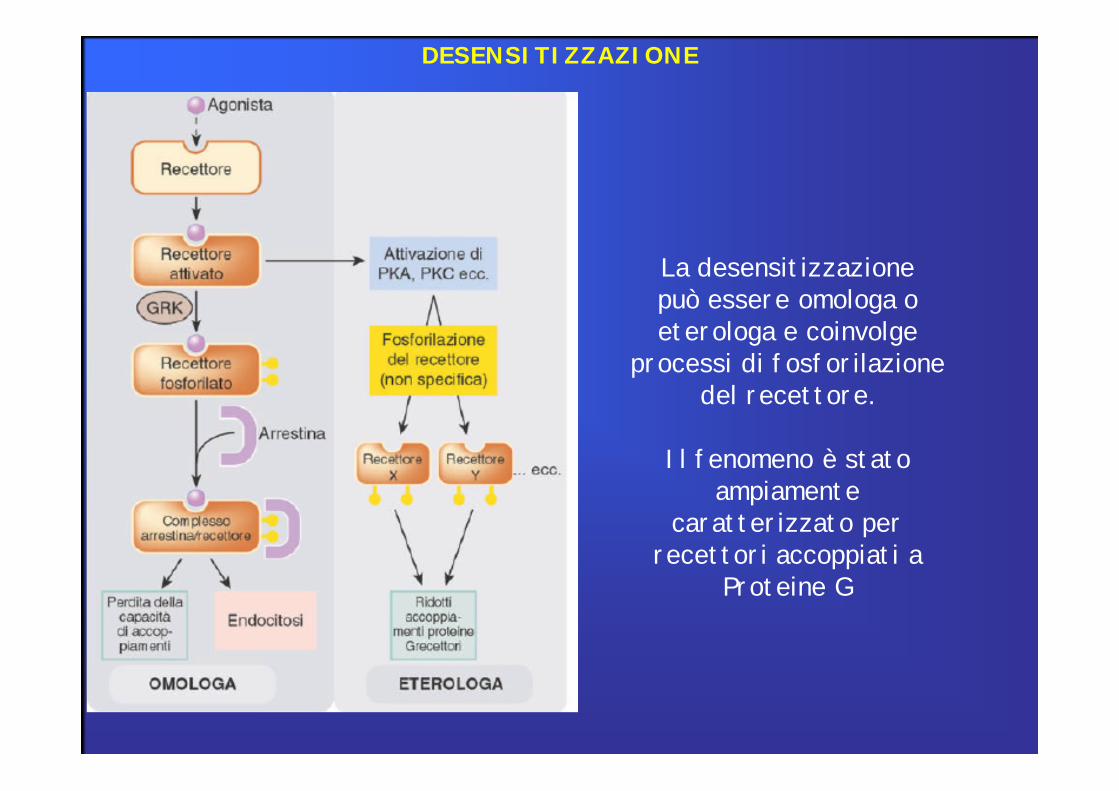
La desensitizzazionepuò essere omologa oeterologa e coinvolge
processi di fosforilazionedel recettore.
Il fenomeno è statoampiamente
caratterizzato per recettori accoppiati a
Proteine G
DESENSITIZZAZIONE
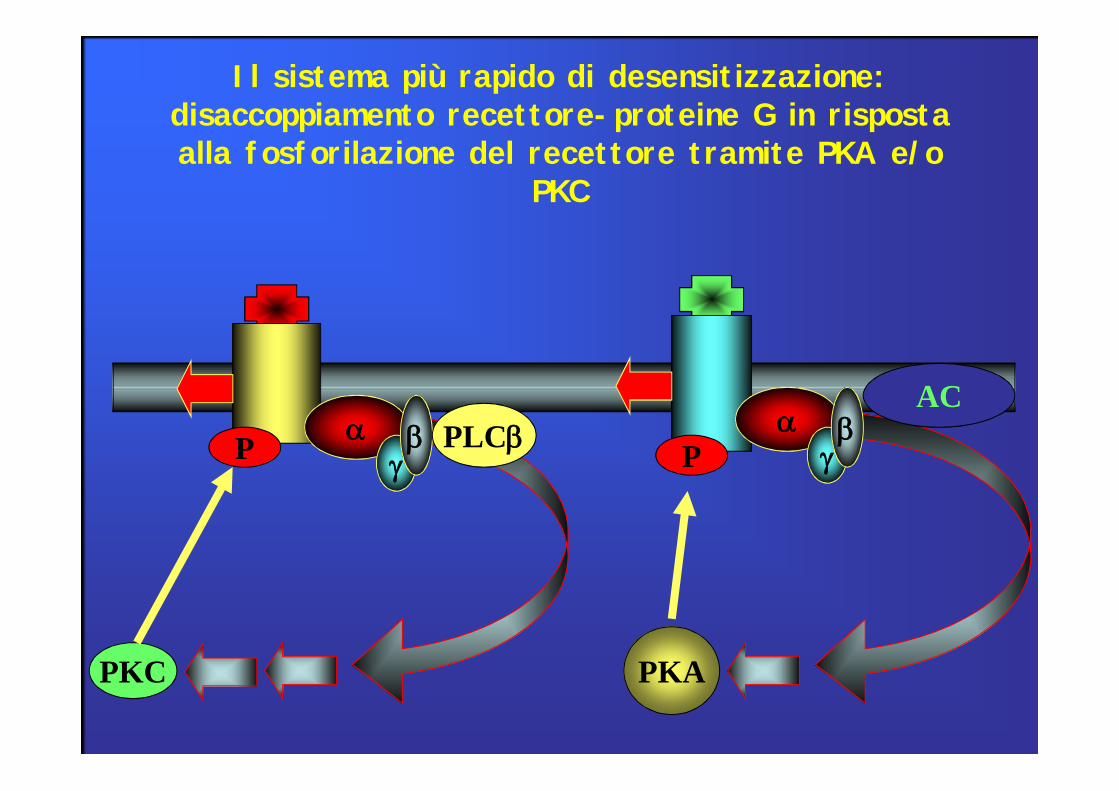
PKC PKA
agb a
gb
ACPLCbP P
Il sistema più rapido di desensitizzazione: disaccoppiamento recettore-proteine G in risposta alla fosforilazione del recettore tramite PKA e/o
PKC

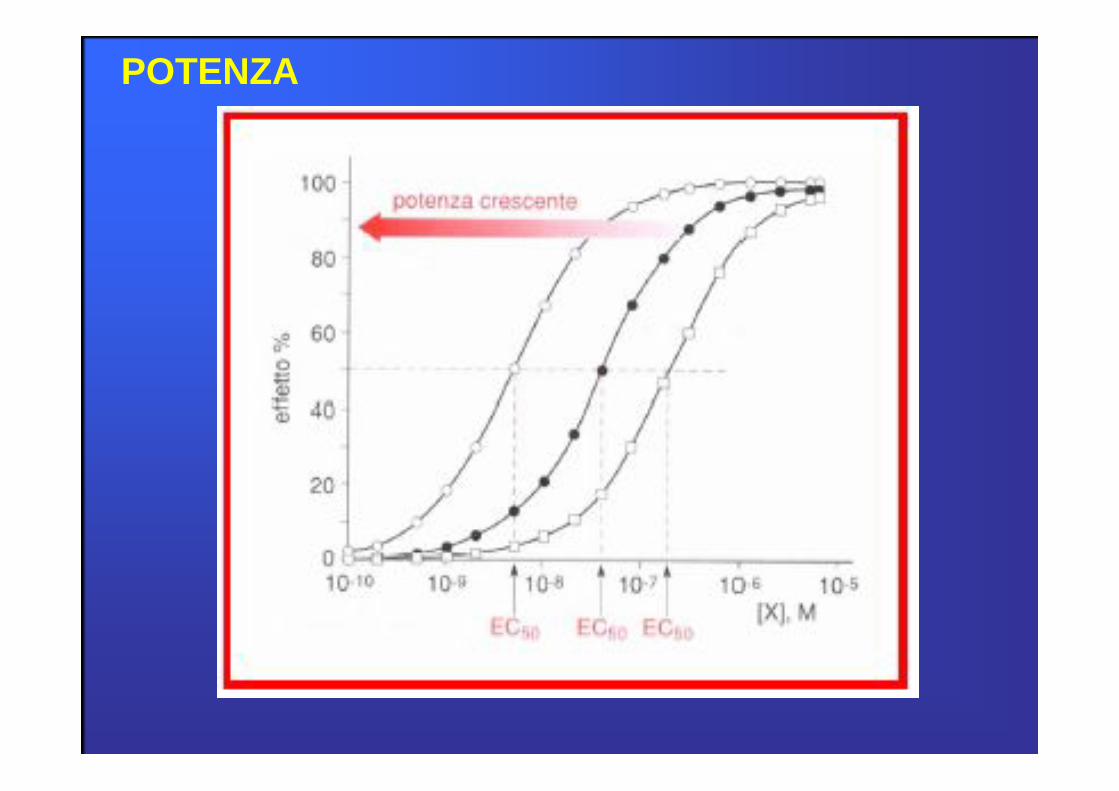
POTENZA
La potenza è indice della dipendenza dell’effetto dalla dose del farmaco.
E’ uguale alla affinità del farmaco per il recettore.
La posizione della curva dose-risposta sull’asse delle ascisse rispecchia la potenza del farmaco.
La potenza influenza la dose necessaria per ottenere un determinato effetto.
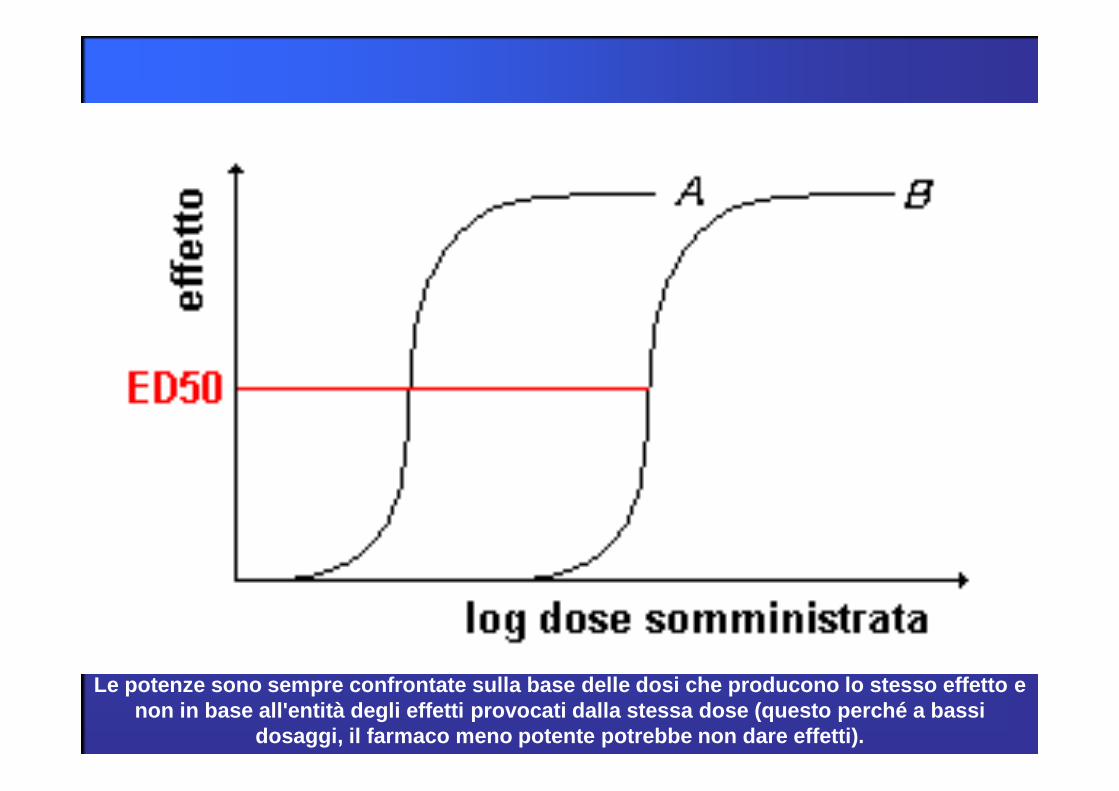
Le potenze sono sempre confrontate sulla base delle dosi che producono lo stesso effetto e non in base all'entità degli effetti provocati dalla stessa dose (questo perché a bassi
dosaggi, il farmaco meno potente potrebbe non dare effetti).
POTENZA

EFFICACIA
Si definisce EFFICACIA di un farmaco l’entità massima dell’effetto che esso può
indurre.
L’altezza della curva dose-risposta rispecchia l’efficacia.
L’efficacia non dipende dalla dose, ma è indice dell’attività intrinseca del farmaco
nell’indurre un determinato effetto.
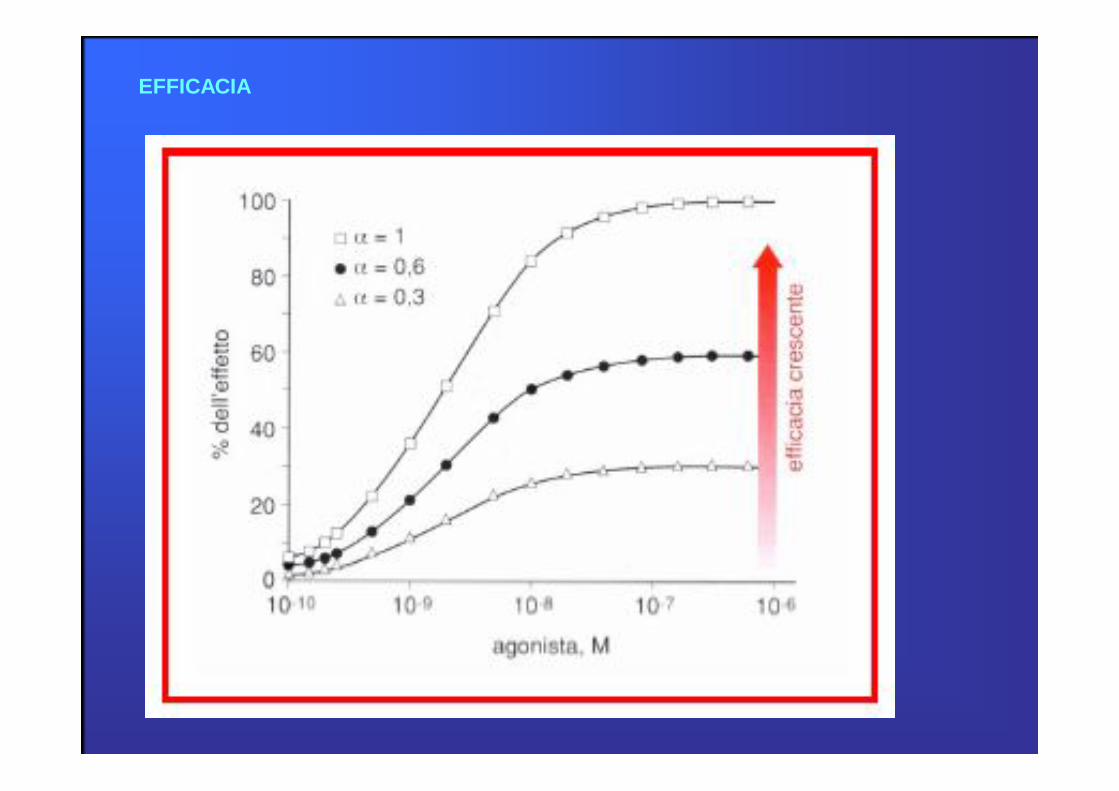
EFFICACIA