Neher%e%Sakmann:%patch&clamp& Come si misura la … · 2018-11-09 · Proprietà funzionali della...
16
Neher e Sakmann: patch clamp Come si misura la corrente da singolo canale?
Transcript of Neher%e%Sakmann:%patch&clamp& Come si misura la … · 2018-11-09 · Proprietà funzionali della...
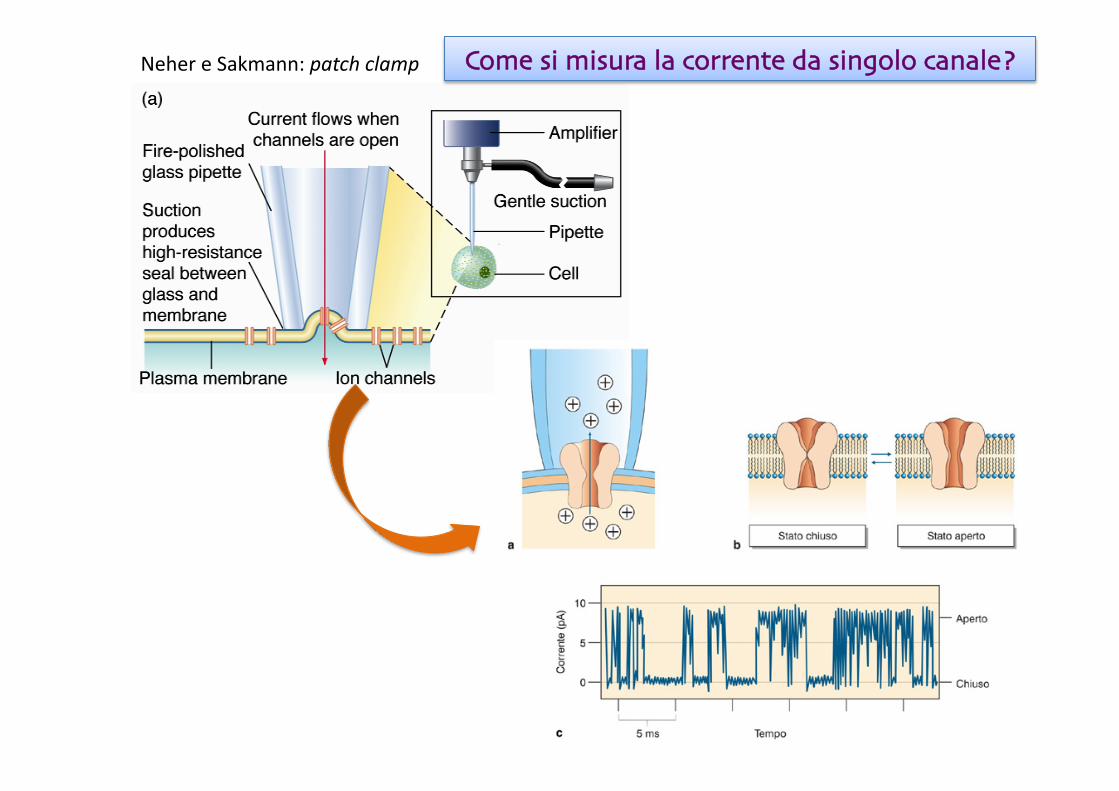
Neher e Sakmann: patch clamp Come si misura la corrente da singolo canale?
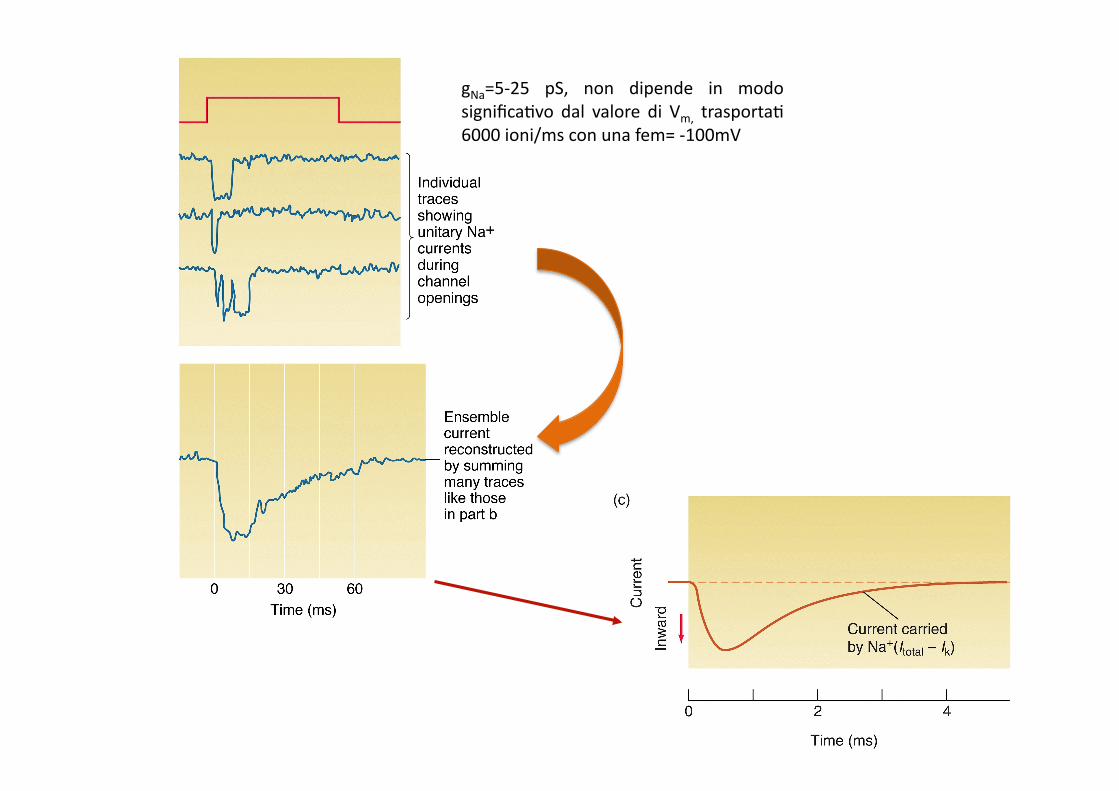
gNa=5-‐25 pS, non dipende in modo significa9vo dal valore di Vm, trasporta9 6000 ioni/ms con una fem= -‐100mV

Aperture stocastiche del canale del Na+ voltaggio-dipendente: gli eventi di apertura sono più frequenti all’inizio dello stimolo depolarizzante
c a
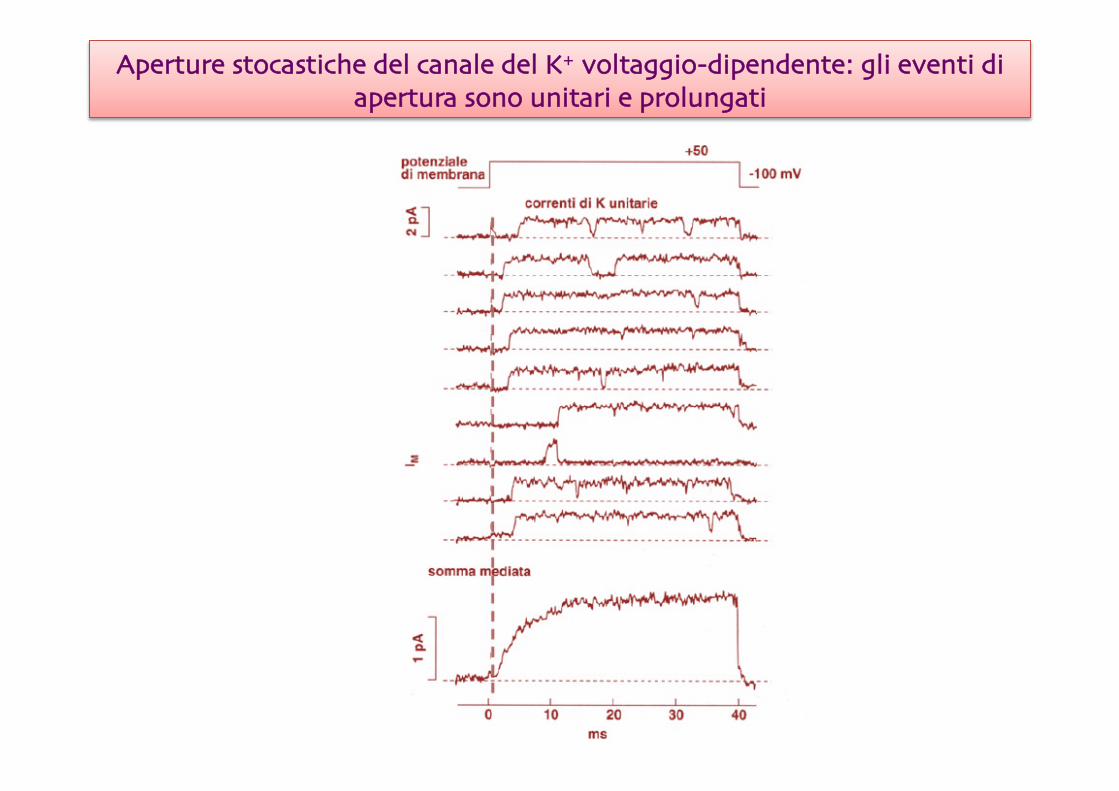
Aperture stocastiche del canale del K+ voltaggio-dipendente: gli eventi di apertura sono unitari e prolungati

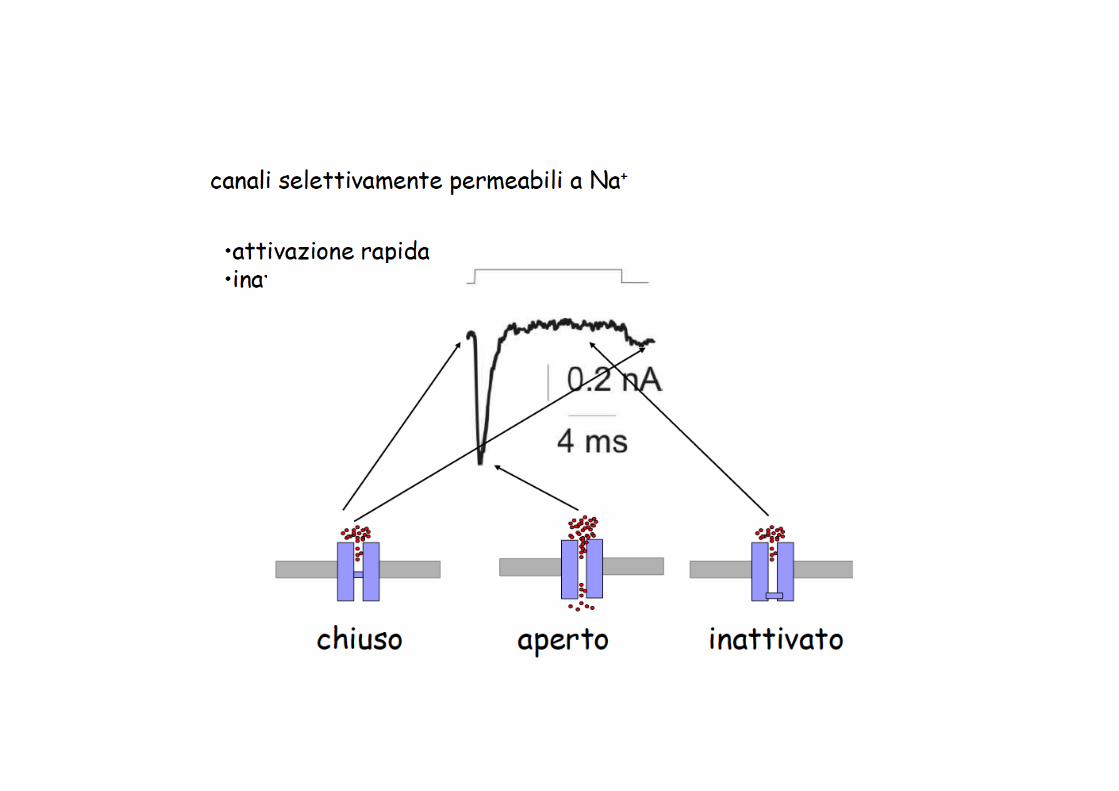

Proprietà funzionali della membrana dipendenti dai canali del Na+ voltaggio-dipendenti
Valore soglia Uno stimolo soglia deve fornire approssimativamente una quantità minima di carica (q), che può essere ottenuta sia aumentando l’ampiezza della corrente stimolante (I), sia la durata del tempo (t), essendo la carica il prodotto di questi due fattori
I dt q =
Ma il livello soglia del potenziale non è una costante assoluta e può cambiare con
le condizioni della stimolazione!
Questo rivela un’importante modalità di modulazione dell’attività nervosa q/C E =
Si può ritenere che questa carica si depositi sulle piastre del condensatore di membrana, alterandone la differenza di potenziale E, in prima approssimazione secondo la relazione
Quindi si parlerà di una quantità minima di carica che deve entrare nella cellula per raggiungere un livello minimo di depolarizzazione per innescare un potenziale d’azione (livello soglia).
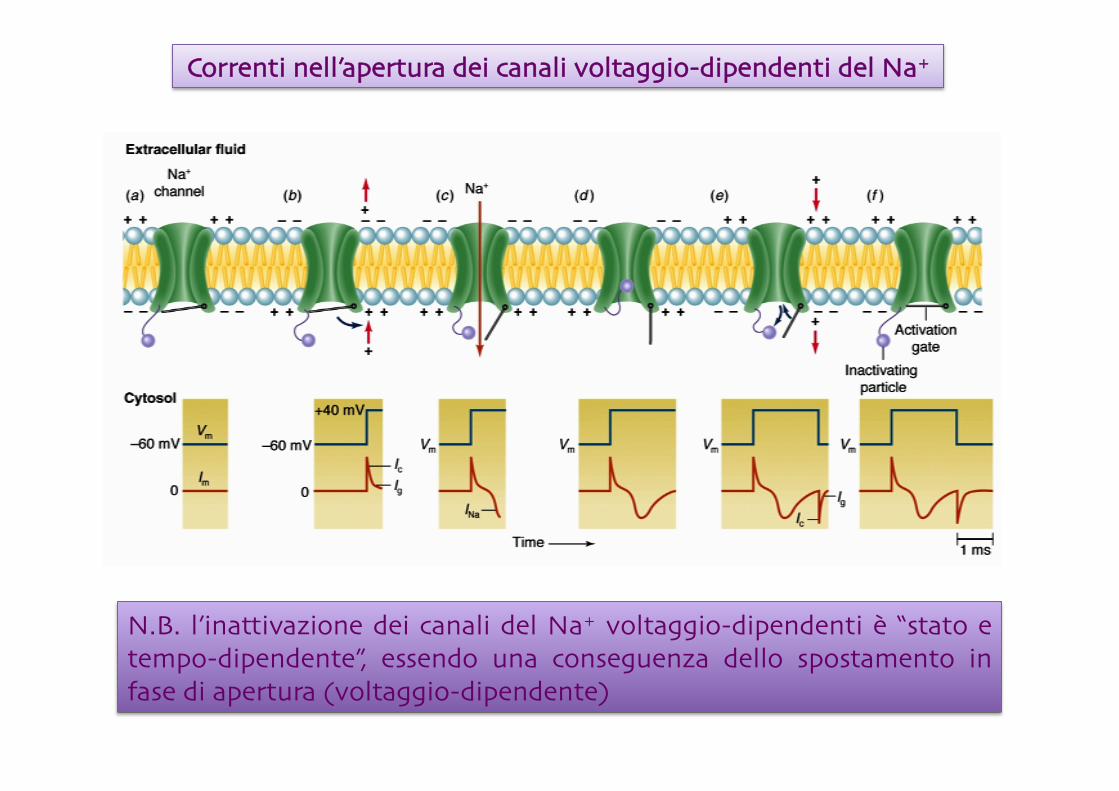
Correnti nell’apertura dei canali voltaggio-dipendenti del Na+
N.B. l’inattivazione dei canali del Na+ voltaggio-dipendenti è “stato e tempo-dipendente”, essendo una conseguenza dello spostamento in fase di apertura (voltaggio-dipendente)
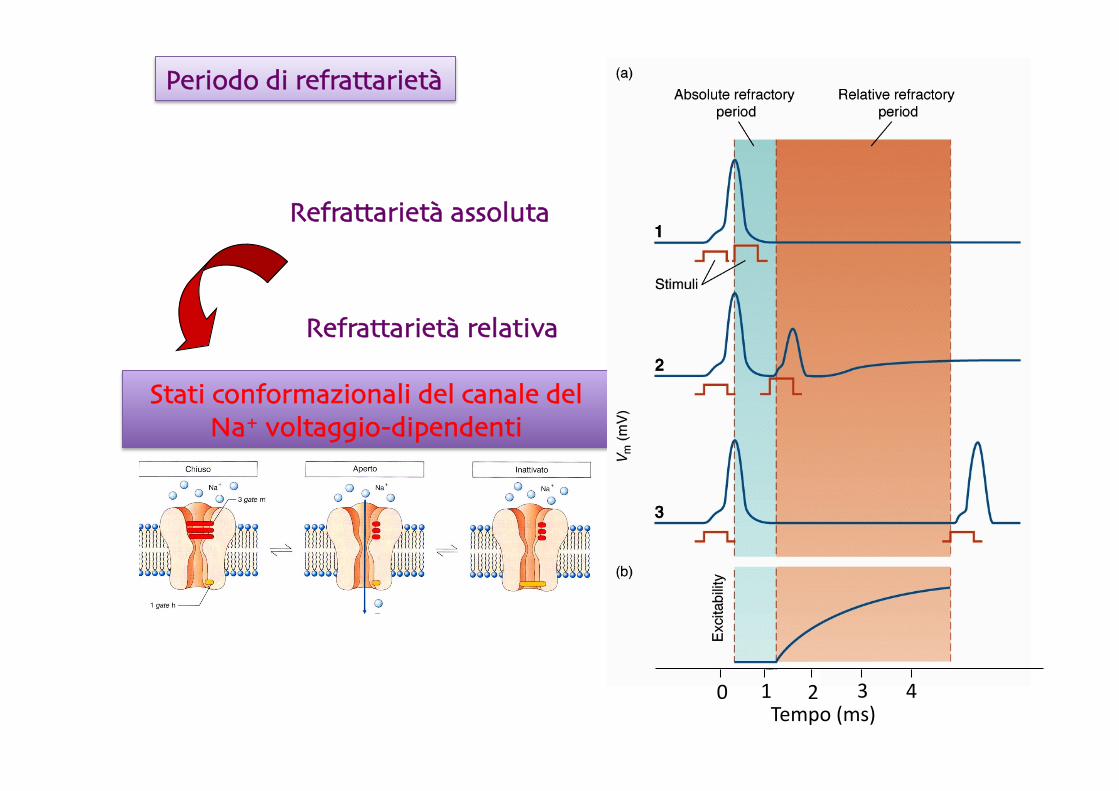
Refrattarietà assoluta
Refrattarietà relativa
Periodo di refrattarietà
Stati conformazionali del canale del Na+ voltaggio-dipendenti
0 1 2 3 4 Tempo (ms)
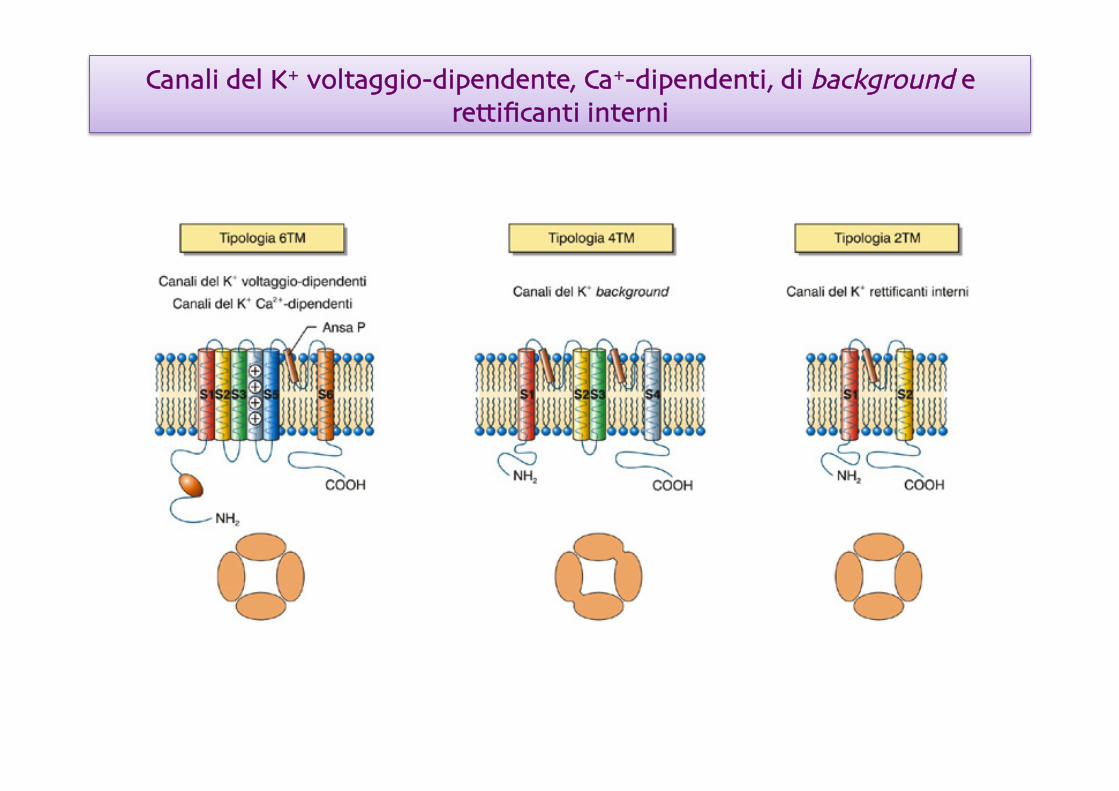
Canali del K+ voltaggio-dipendente, Ca+-dipendenti, di background e rettificanti interni
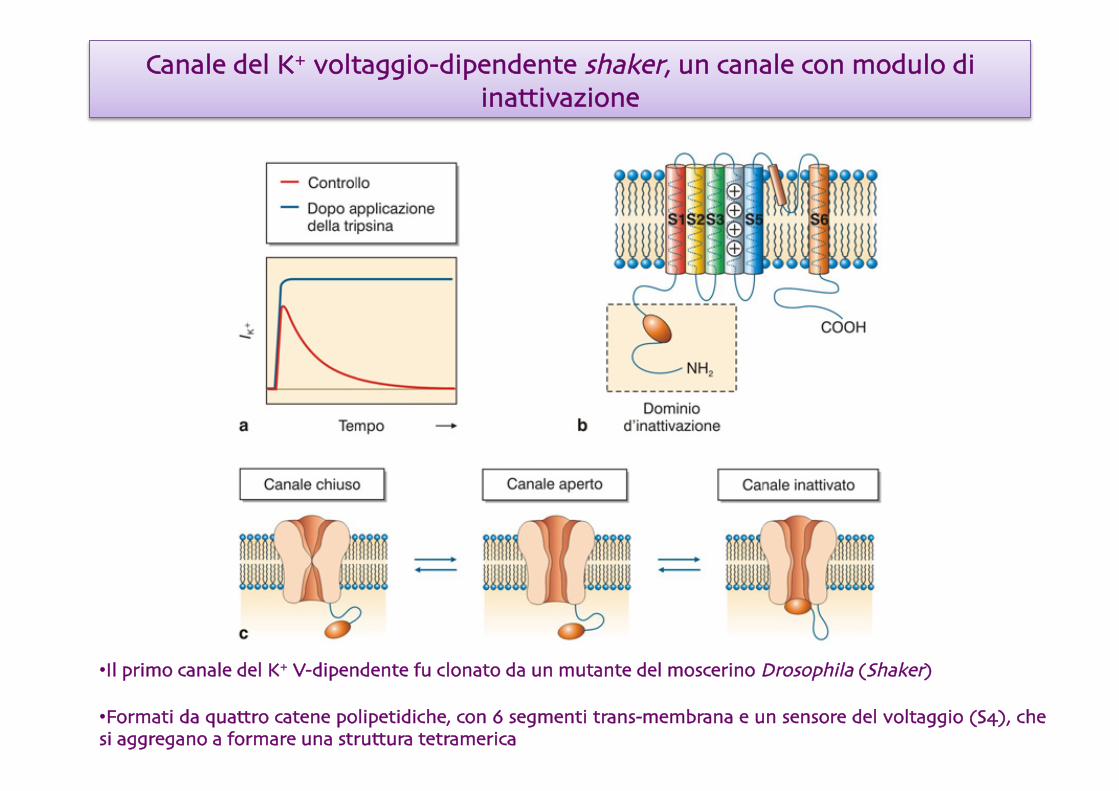
Canale del K+ voltaggio-dipendente shaker, un canale con modulo di inattivazione
• Il primo canale del K+ V-dipendente fu clonato da un mutante del moscerino Drosophila (Shaker)
• Formati da quattro catene polipetidiche, con 6 segmenti trans-membrana e un sensore del voltaggio (S4), che si aggregano a formare una struttura tetramerica

Canali del potassio voltaggio-attivati
Tre sottofamiglie (8 membri)
Conducono :
• corrente macroscopica rettificante ritardata (DRK), no intattivante, sebbene dopo diverse centinaia di ms di stimolazioni vada incontro ad una lentissima inattivazione
• corrente macroscopica transiente (corrente di tipo A), si instaura rapidamente e si inattiva;
• corrente di tipo M di più lenta attivazione e non inattivante (scoperta nei neuroni parasimpatici)
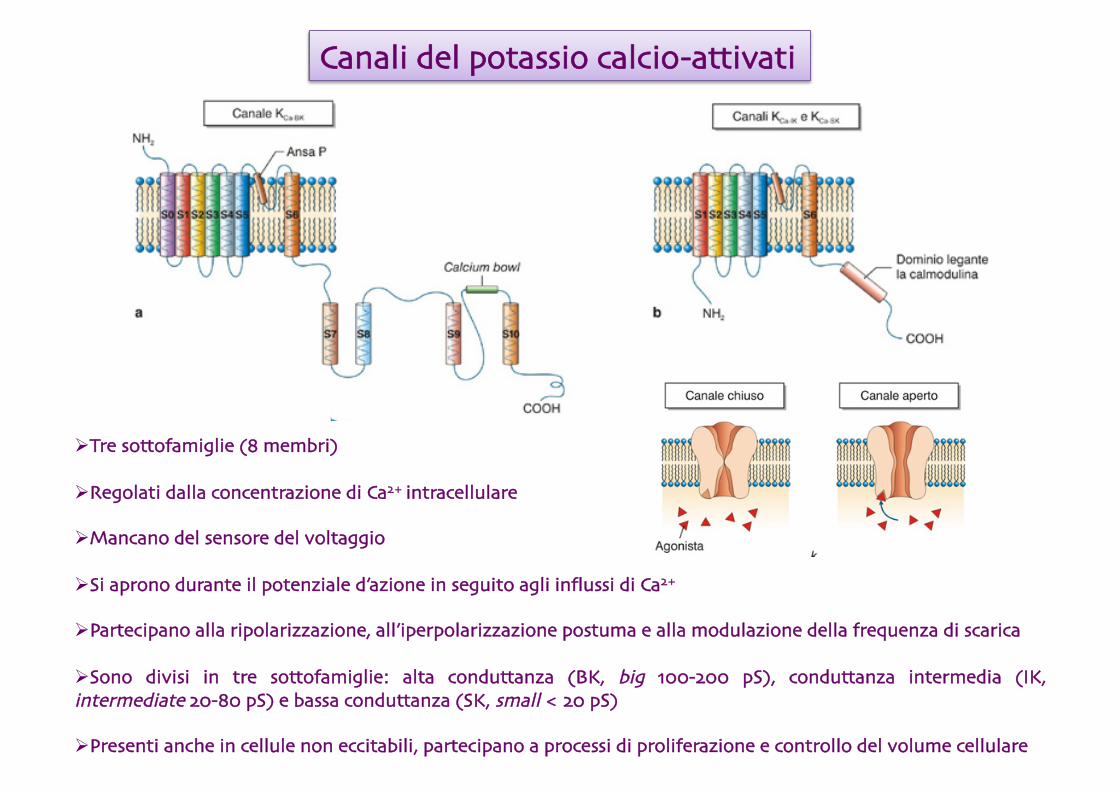
Canali del potassio calcio-attivati
Tre sottofamiglie (8 membri)
Regolati dalla concentrazione di Ca2+ intracellulare
Mancano del sensore del voltaggio
Si aprono durante il potenziale d’azione in seguito agli influssi di Ca2+
Partecipano alla ripolarizzazione, all’iperpolarizzazione postuma e alla modulazione della frequenza di scarica
Sono divisi in tre sottofamiglie: alta conduttanza (BK, big 100-200 pS), conduttanza intermedia (IK, intermediate 20-80 pS) e bassa conduttanza (SK, small < 20 pS)
Presenti anche in cellule non eccitabili, partecipano a processi di proliferazione e controllo del volume cellulare
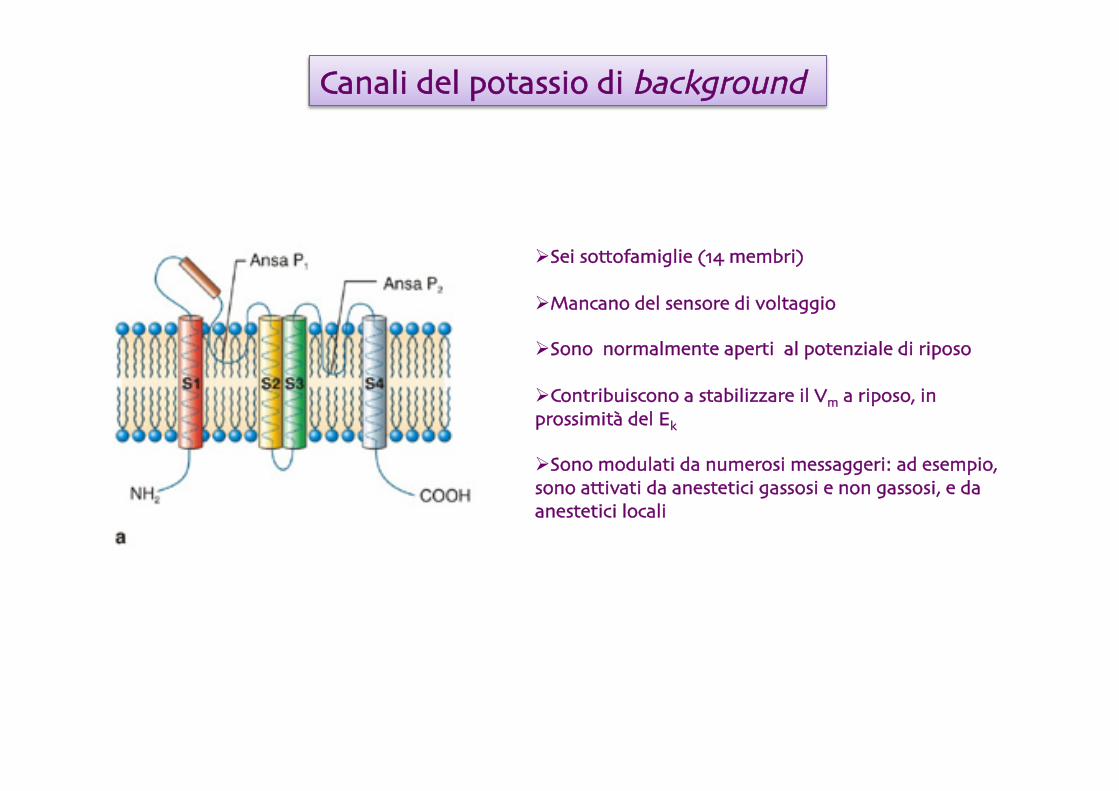
Sei sottofamiglie (14 membri)
Mancano del sensore di voltaggio
Sono normalmente aperti al potenziale di riposo
Contribuiscono a stabilizzare il Vm a riposo, in prossimità del Ek
Sono modulati da numerosi messaggeri: ad esempio, sono attivati da anestetici gassosi e non gassosi, e da anestetici locali
Canali del potassio di background
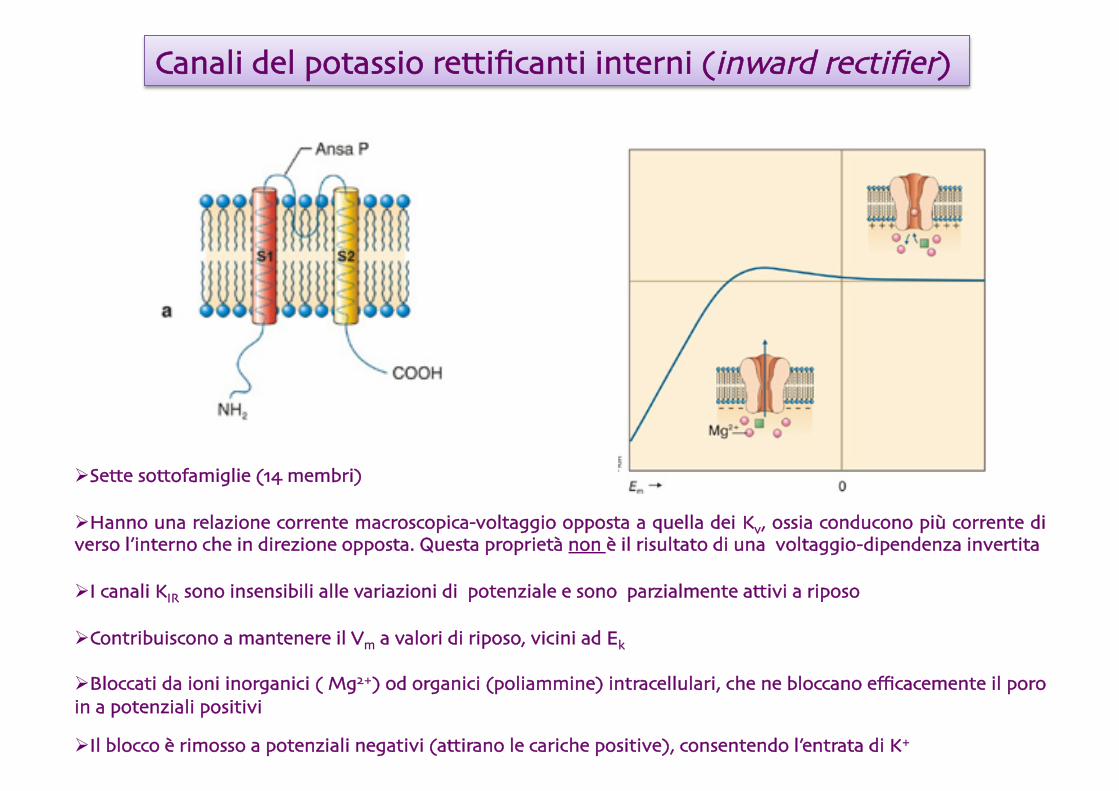
Canali del potassio rettificanti interni (inward rectifier)
Sette sottofamiglie (14 membri)
Hanno una relazione corrente macroscopica-voltaggio opposta a quella dei Kv, ossia conducono più corrente di verso l’interno che in direzione opposta. Questa proprietà non è il risultato di una voltaggio-dipendenza invertita
I canali KIR sono insensibili alle variazioni di potenziale e sono parzialmente attivi a riposo
Contribuiscono a mantenere il Vm a valori di riposo, vicini ad Ek
Bloccati da ioni inorganici ( Mg2+) od organici (poliammine) intracellulari, che ne bloccano efficacemente il poro in a potenziali positivi
Il blocco è rimosso a potenziali negativi (attirano le cariche positive), consentendo l’entrata di K+

Riassumendo: Caratteristiche uniche del potenziale d’azione
Il potenziale di membrana si inverte di segno e l’eccedenza si avvicina al potenziale di equilibrio del Na+ (+55 mV - +60 mV)
Durante un potenziale d’azione la conduttanza della membrana cambia, mentre la sua capacità rimane costante
Il potenziale d’azione è un evento “tutto o nulla”
3
4
5
3 4 5
Il potenziale d’azione è un evento rigenerativo (non decade con la distanza)
A differenza dei potenziali elettrotonici, i potenziali d’azione non si sommano tra loro (conseguenza del periodo di refrattarietà)