Meccanismi psicobiologici di impulsività e propensione all ... · Tendenza ad agire senza...
99
Meccanismi psicobiologici di impulsività e propensione all‟azzardo: Modelli animali e differenze di età Dr. Walter Adriani Sezione di Neuroscienze Comportamentali Dipartimento di Biologia Cellulare e Neuroscienze Istituto Superiore di Sanità, Roma
Transcript of Meccanismi psicobiologici di impulsività e propensione all ... · Tendenza ad agire senza...

Meccanismi psicobiologici di impulsività e propensione
all‟azzardo: Modelli animali e differenze di età
Dr. Walter Adriani
Sezione di Neuroscienze Comportamentali
Dipartimento di Biologia Cellulare e Neuroscienze
Istituto Superiore di Sanità, Roma

E’ un concetto che può essere definito in vari modi:
Incapacità di saper aspettare o di inibire comportamenti inappropriati
Tendenza ad agire senza prevedere le possibili conseguenze
Insensibilità alle conseguenze avverse generate dal proprio agire
Le due principali categorie d’impulsività sono:
Impulsività motoria: incapacità di resistere a un forte impulso, a una pulsione o a una tentazione
Impulsività cognitiva: scelta, fra opzioni, presa senza considerare alternative e/o conseguenze
Il comportamento esageratamente impulsivo è un sintomo clinico significativo in una serie di
patologie psichiatriche, caratterizzate da un controllo inibitorio inappropriato :
Disturbo da deficit di attenzione / iperattività (ADHD)
Disturbi ossessivo-compulsivi
Abuso di sostanze (alcool, fumo, nuove droghe “smart”)
Gioco d‟azzardo patologico
Impulsività

Bambini ADHD
Deficit dell‟attenzione
Impulsività
Eccessiva attività motoria
Rendimento scolastico basso
Difficoltà di relazionarsi con i coetanei
Bassa autostima dovuta a fallimenti ripetuti
Disturbo da deficit dell’attenzione / iperattività (ADHD)
E‟ il più comune tra i disturbi neuro-comportamentali in bambini e adolescenti
La prevalenza stimata è del 4-7% negli USA e dell‟ 1-3% in Europa
É circa 4 volte più frequente nei maschi che nelle femmine
C‟è una elevata co-morbidità con altri disturbi neuropsichiatrici
Nell‟80% dei bambini con ADHD, i sintomi permangono nell‟adolescenza; nel 30-50% di questi casi,
essi persistono anche in età adulta.
Adulti ed adolescenti ADHD
Deficit dell‟attenzione
Impulsività
Propensione al rischio
Gioco d‟azzardo patologico
Abuso di alcool e droghe
Personalità antisociale
Breve panoramica sulla patologia

Gioco d’azzardo patologico (GAP)
Colpisce lo 0.2-5.3% degli adulti nella popolazione mondiale
Inizia sempre più ad essere un problema anche tra gli adolescenti
3.5-8.0% degli adolescenti rientra nei criteri diagnostici del GAP
C‟è co-morbidità con una serie di altri disturbi e con l‟abuso di sostanze
Nel DSM-V: da “Disturbo del controllo degli impulsi, non classificato
altrove” a “Disturbo correlato a sostanza”
Il GAP come una “dipendenza senza sostanza”: tolleranza, crisi di
astinenza, perdita di controllo sul comportamento
Breve panoramica sulla patologia
Sintomi psicologici
• Alterazione dell‟umore
• Alterata autostima
• Aumento della impulsività
• Comportamenti superstiziosi
Sintomi fisiologici
• Ansia correlata a sintomi fisici
• Insonnia
• Mal di testa
• Infarti (dovuti a stress e a sovra-
eccitazione da gioco d‟azzardo)
Impatto sulla società
• Problemi finanziari,
sociali, e legali (inclusi:
fallimento, divorzio, perdita
del lavoro, galera)
• Tentativi di suicidio
• Abuso di sostanze

Propensione all‟azzardo
I vari giochi combinano questi elementi in modi
differenti:
Poker, Black Jack = alea (fortuna) + agon (competizione)
Russian roulette = alea (fortuna) + ilynx (vertigine)
Competizione
P.Es. scacchi
Probabilità, caso
P.Es. slot machine
Abilità
P.Es. imitazione o
pantomima
Vertigine
P.Es. montagne
russe, paracadute
Una definizione di gioco d’azzardo
Gioco che consiste nello scommettere denaro, o altri beni equivalenti, sul futuro esito di un
evento che presenti un margine di incertezza, in vista della vincita di un premio; la vincita è
strettamente legata al caso e non all‟abilità individuale.

Test e scale di personalità:
Barratt Impulsiveness Scale (BIS)
Eysenck Impulsiveness Scale (EIS)
UPPS Impulsive Behaviour Scale
Paradigmi comportamentali (in ambito clinico e sperimentale, inclusi modelli animali):
Marshmallow Test (solo negli umani)
Delay Discounting / Delay Intolerance
Go/No-Go Task
Stop-Signal Reaction Time (SSRT) Task
Differential Reinforcement of Low Response Rate Task (DRL)
Per misurare l‟impulsività
Ritardo nella consegna della ricompensa
Inibizione della risposta

Test e scale di personalità :
South Oaks Gambling Screen (SOGS)
South Oaks Gambling Screen Revised for Adolescents (SOGS-RA)
Gambling Attitudes and Beliefs Survey (GABS)
Canadian Problem Gambling Index (CPGI)
Paradigmi comportamentali (in ambito clinico e sperimentale, inclusi modelli animali):
Iowa Gambling Task (IGT)
Balloon Analogue Risk Task (BART) (solo negli umani)
Probability Discounting Task (PDT)
Per misurare la propensione all‟azzardo
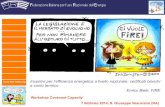
Aree cerebrali coinvolte nella impulsività e propensione all‟azzardo
Corteccia pre-frontale
Elaborazione e attuazione delle risposte
comportamentali più evolute
Inibizione attiva delle risposte sottocorticali
Striato dorsale
Apprendimento di nuove procedure e di
strategie abitudinarie
Formarsi e stabilirsi di “riti” e abitudini
Comportamenti semi-automatici innati e/o
acquisiti
Striato ventrale (nucleus accumbens)
Gratificazione correlata alla ricompensa
Valutazione del valore della ricompensa
Sostiene e motiva le pulsioni che portano al
raggiungimento degli obiettivi

Corteccia pre-frontale
Coinvolta nella risoluzione di scelte binarie o multiple, attraverso la pianificazione, regolazione sulla
base del risultato (feedback) e inibizione dei comportamenti concorrenti (Dalley et al. 2004)
Promuove l‟attenzione cognitiva sia verso le caratteristiche dello stimolo, sia verso le caratteristiche
del compito, supportando i cambiamenti attentivi e la flessibilità cognitiva / comportamentale
Striato dorsale
Classicamente considerato come promotore del comportamento abitudinario, innato o acquisito
Lo striato dorso-laterale è coinvolto nell‟espressione dei “riti” e delle abitudini; lo striato dorso-mediale
svolge un ruolo cruciale nel supportare la flessibilità comportamentale - permettendoci di comportarci
diversamente da quanto suggerito in prima istanza dal nostro istinto (Ragozzino 2003; Yin et al. 2004)
Striato ventrale
Consiste nel nucleo accumbens (NAcc) e nel tubercolo olfattivo
Coinvolto nei processi di rinforzo: svolge la valutazione “affettiva” delle caratteristiche dell‟esito di una
scelta, e la modulazione (feedback) delle scelte successive (Cardinal et al. 2004; Christakou et al. 2004)
Il NAcc svolge un ruolo cruciale nella motivazione a compiere sforzi volti a raggiungere gli obiettivi,
determinando il massimo sforzo sostenibile (Salamone et al. 2005; Salamone et al. 2007)
Aree cerebrali coinvolte nella impulsività e propensione all‟azzardo
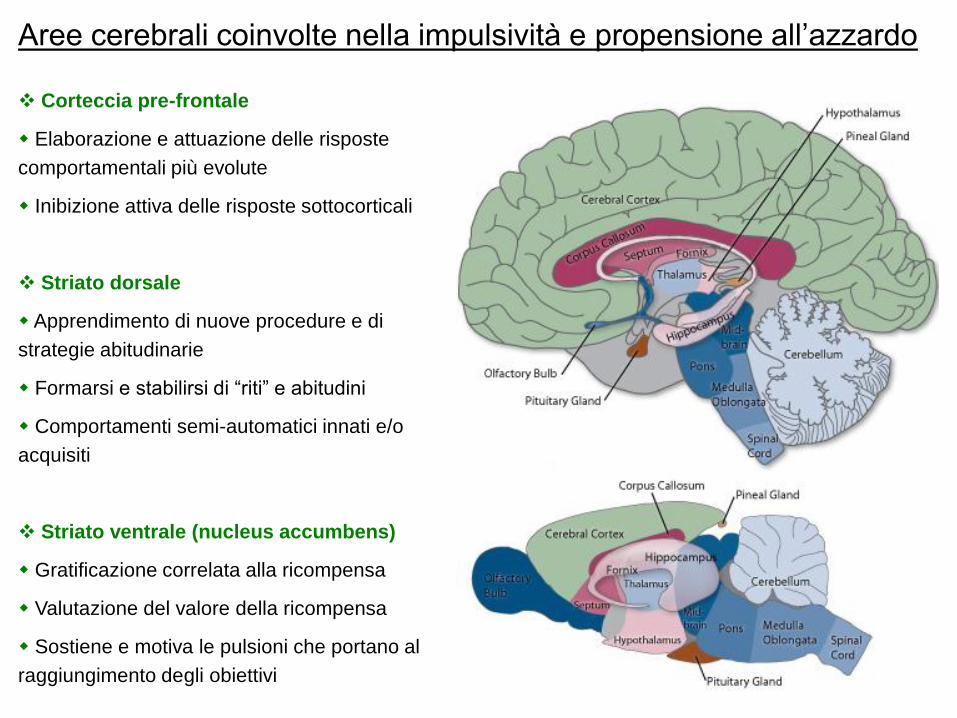
Sistemi neurochimici di trasmissione
Sistema Dopaminergico Sistema Serotoninergico
Neurotrasmettitore: Dopamina, DA
Origine: Ventral tegmentum; Substantia nigra
Target: Nucleus accumbens; Striato dorsale
Neurotrasmettitore: Serotonina, 5-HT
Origine: Raphe nuclei (mesencefalo)
Target: Corteccia pre-frontale

Neurotrasmettitore: Dopaminae Neurotrasmettitore: Serotonina
Sistemi neurochimici di trasmissione

Sistema di rinforzo e ricompensa
Circuito costituito da neuroni dopaminergici
che, dall‟ Area Tegmentale Ventrale (VTA),
vanno ad innervare il Nucleus Accumbens e
parte della Corteccia pre-frontale
Questo importante sistema neurochimico è
chiamato sistema meso-limbico e agisce in
parallelo con quello meso-corticale
Altro sistema, implicato nell‟apprendimento
procedurale, è quello nigro-striatale
Il sistema della gratificazione nei disturbi di controllo degli impulsi
Alterata percezione della ricompensa vs. alterata sensibilità al rinforzo
Corteccia pre-frontale (mediale e orbitale) e area ventro-striatale (Nucleus Accumbens): regolano i
processi motivazionali, di gratificazione e di “aspettativa” (valore attribuito a rinforzi futuri)
I circuiti cortico-striatali dorso-mediale e dorso-laterale regolano le azioni semi-automatiche
Entrambi i sistemi, dipendenti dall‟attività della dopamina, sono attivati dagli psicostimolanti
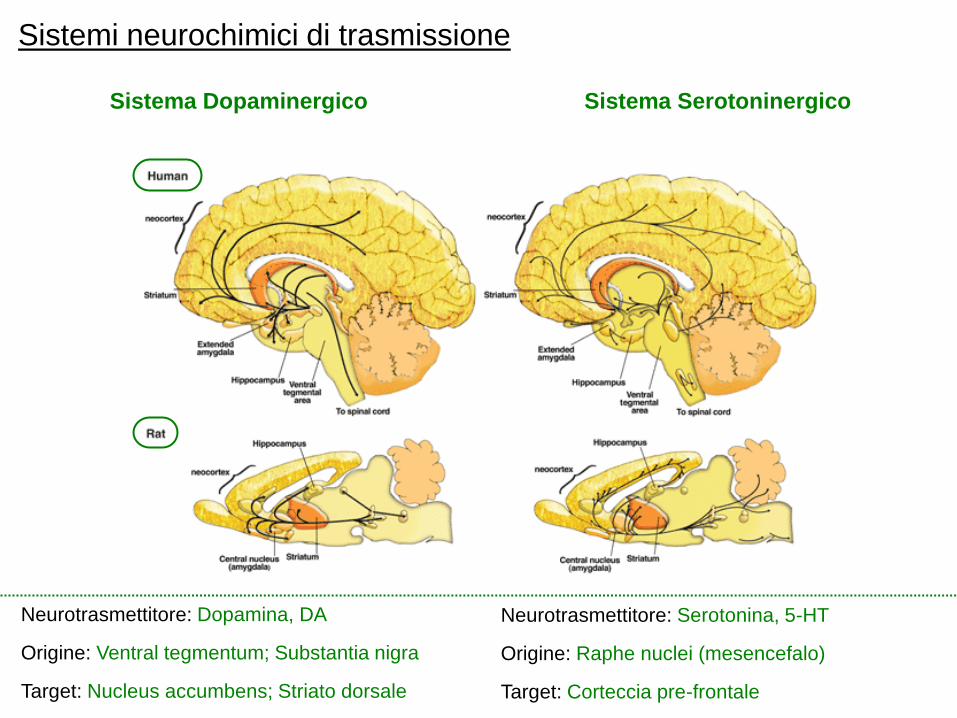
Le proiezioni meso-limbiche e meso-corticali, che originano dalla VTA
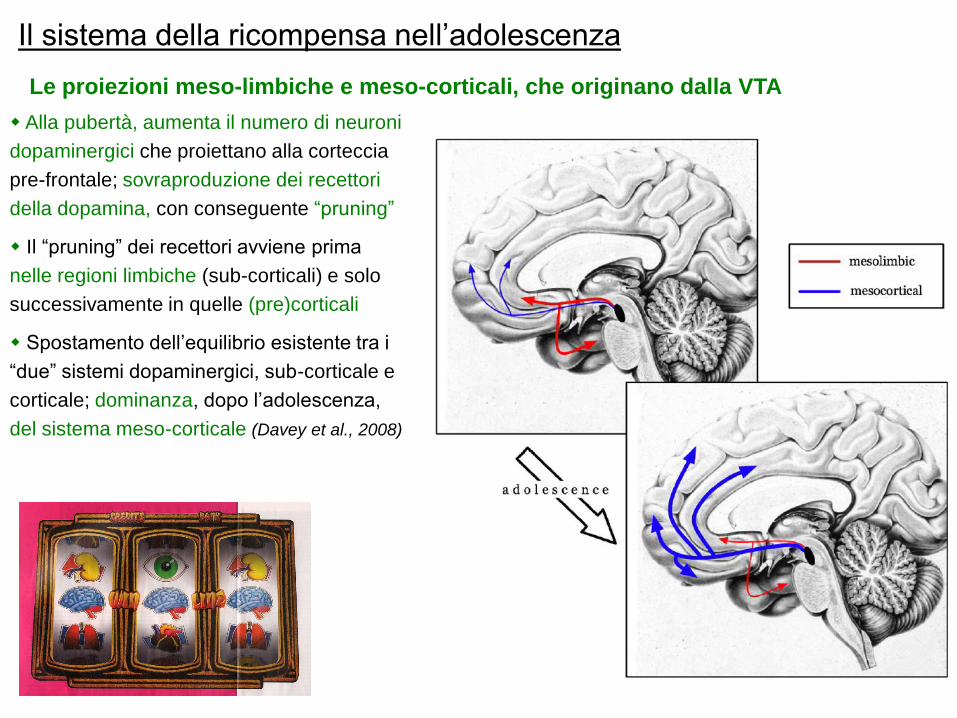
Il sistema della ricompensa nell‟adolescenza
Alla pubertà, aumenta il numero di neuroni
dopaminergici che proiettano alla corteccia
pre-frontale; sovraproduzione dei recettori
della dopamina, con conseguente “pruning”
Il “pruning” dei recettori avviene prima
nelle regioni limbiche (sub-corticali) e solo
successivamente in quelle (pre)corticali
Spostamento dell‟equilibrio esistente tra i
“due” sistemi dopaminergici, sub-corticale e
corticale; dominanza, dopo l‟adolescenza,
del sistema meso-corticale (Davey et al., 2008)
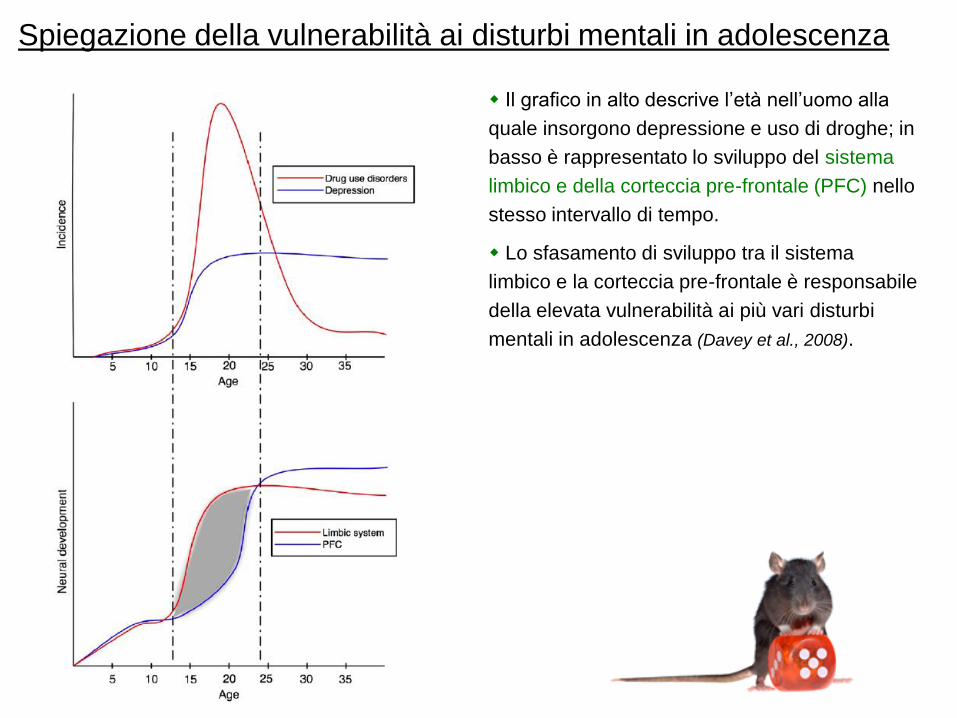
Spiegazione della vulnerabilità ai disturbi mentali in adolescenza
Il grafico in alto descrive l‟età nell‟uomo alla
quale insorgono depressione e uso di droghe; in
basso è rappresentato lo sviluppo del sistema
limbico e della corteccia pre-frontale (PFC) nello
stesso intervallo di tempo.
Lo sfasamento di sviluppo tra il sistema
limbico e la corteccia pre-frontale è responsabile
della elevata vulnerabilità ai più vari disturbi
mentali in adolescenza (Davey et al., 2008).


Lo sviluppo della sostanza grigia corticale segue un modello regionale specifico. La sostanza grigia
diminuisce man mano che il cervello matura. In figura, lo sviluppo del cervello è riassunto in 5 immagini
in cui si nota il passaggio da rosso = meno maturo a viola = più maturo (Lenroot & Giedd, 2006).
Differenze di età negli esseri umani: MRI anatomica

Differenze di sesso ed età nell‟impulsività degli umani
(A) Grafico sulla sensation-seeking ed impulsività
in funzione dell‟età. (B) Grafico che illustra l‟attività
in due regioni del cervello, implicate nel feedback
(la valutazione dei risultati), durante un compito di
controllo cognitivo; si noti l‟andamento durante lo
sviluppo adolescenziale (Casey & Jones, 2010).
Gli effetti della deplezione acuta di triptofano
(ATD) sul tipo di risposta in uomini e donne.
Uno stile di risposta < 1 indica una risposta
impulsiva, mentre > 1 indica una risposta
equilibrata (Walderhaug et al., 2007).
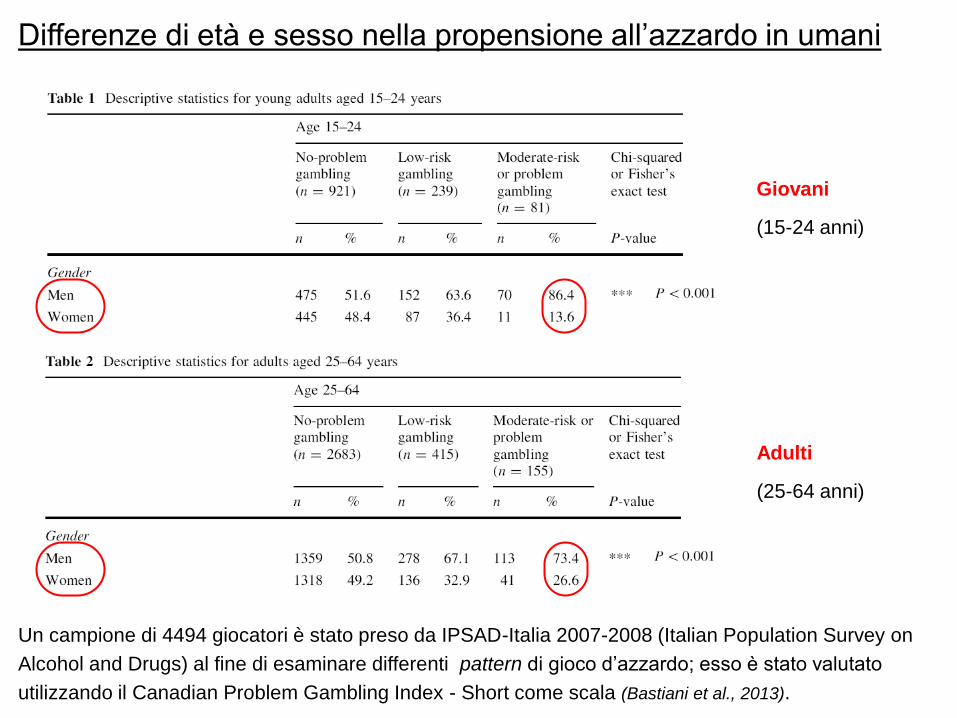
Differenze di età e sesso nella propensione all‟azzardo in umani
Un campione di 4494 giocatori è stato preso da IPSAD-Italia 2007-2008 (Italian Population Survey on
Alcohol and Drugs) al fine di esaminare differenti pattern di gioco d‟azzardo; esso è stato valutato
utilizzando il Canadian Problem Gambling Index - Short come scala (Bastiani et al., 2013).
Giovani
(15-24 anni)
Adulti
(25-64 anni)

Differenze di età e sesso in propensione all‟azzardo in umani
Distribuzione dei soggetti italiani, registrati per la terapia contro il gioco d‟azzardo patologico nel 2011,
suddivisi per sesso ed età (totale di 4544 soggetti, di cui l‟ 82% sono uomini ed il 18% donne).

Gli adolescenti umani possono presentare dei comportamenti devianti e/o a rischio, tra
cui la ricerca e l‟utilizzo di sostanze psicoattive (e altre fonti di “sensazioni”).
Il ritardo nella maturazione tra la corteccia pre-frontale e le aree sub-corticali (limbiche) ha
un ruolo importante e può spiegare la manifestazione di comportamenti “sregolati”.
Negli ultimi vent‟anni, il contributo della pubertà, con tutti i suoi rimodellamenti nei
processi neurobiologici e neuroendocrinologici importanti per lo sviluppo, ha ricevuto
un‟attenzione crescente nella investigazione sperimentale.
E‟ quindi importante caratterizzare il comportamento di ricerca della novità e del rischio
nei roditori di laboratorio, durante l‟adolescenza e/o utilizzando modelli specifici.
Riassumendo

Modelli animali in neuroscienze comportamentaliModelli animali
Permettono di indagare le relazioni tra il cervello e il
comportamento, con l‟obiettivo di approfondire la nostra
conoscenza sui comportamenti normali e anormali, e sui
sottostanti processi neuro-cerebrali e neuro-endocrinologici
(van der Staay, 2006)
Permettono di analizzare alcune di queste relazioni in
condizioni controllate (per es., condizioni di stabulazione
standardizzate)
Perché i modelli animali sono necessari?
Per ottenere quelle informazioni che non possono essere
acquisite in altri modi (per es., manipolazioni genetiche ed
ambientali, trattamenti farmacologici, sostanze psicoattive,
ecc.)
Approcci in vitro non possono riprodurre le interazioni in
sistemi viventi complessi, come le relazioni tra il cervello e il
comportamento

Validità dei modelli animali
La distinzione tra differenti forme di validità è utile al fine di identificare debolezze e limitazioni di un
modello: una scarsa validità determina un aumento del numero di falsi positivi e falsi negativi nella
ricerca di base, limitando in tal modo la possibilità di sviluppare nuove strategie terapeutiche (Willner
1984, 1995, 2002; van der Staay 2006, 2009).
Categorie di validità
Validità interna (affidabilità e replicabilità): si riferisce alla qualità intrinseca della valutazione
sperimentale condotta nel modello animale.
Validità di “facciata” (validità apparente): indica la similarità fenomenologica tra il modello e la
condizione alla quale il modello si riferisce.
Un animale modello con elevata validità predittiva permette l‟estrapolazione dell‟effetto, esercitato
da una particolare manipolazione sperimentale (o farmaco), dal modello alla specie umana.
Validità di costrutto: fa riferimento al grado di similarità biologica, tra i meccanismi alla base del
comportamento nel modello animale e quelli alla base del comportamento nella condizione che il
modello si propone di riprodurre.
Validità esterna (generalizzabilità): fa riferimento alla misura in cui i risultati ottenuti usando un
particolare modello animale possono essere generalizzati ad altre specie più simili all‟uomo.

Un confronto tra sviluppo negli umani e nei ratti (Andersen,
2003).
Sviluppo umani-ratti a confronto

“Impulsività motoria” (azione impulsiva) può essere valutata con paradigmi che non sono basati sulla
scelta, e che ricordano il classico“Go/No-Go Task” utilizzato negli umani.
La valutazione della “impulsività cognitiva” (scelta impulsiva) richiede paradigmi che vertono su scelte
binarie o multiple; uno dei più utilizzati è il “delay-discounting”.
Soggetti impulsivi sono molto intolleranti a situazioni in cui la ricompensa è ritardata: premi più
piccoli ma che arrivano immediatamente sono preferiti rispetto a quelli più grandi, ma che arrivano
solo dopo un ritardo (Thiebot et al., 1985; Logue, 1988; Evenden & Ryan, 1996; 1999; Bizot et al., 1999)
Il “five-choice serial reaction time task (5CSRTT)”: inizialmente sviluppato per studiare l‟attenzione,
implica aspetti di inibizione comportamentale, tra cui: risposte premature, perseverative (Carli et al., 1983).
Il “unpredictable operant conditioning schedule (variable interval-15, VI-15)” : ci consente di
misurare le risposte “liberamente” emesse durante un periodo refrattario variabile (Coppens et al., 2012).
Il “conditioned locomotor activity to food” : un paradigma non operante che consente di misurare
la disinibizione comportamentale (Matthews et al., 1996; Winstanley et al., 2004).
Lo studio dell‟impulsività in modelli animali

Studio della propensione all‟azzardo in modelli animali
Molti paradigmi operanti sono stati sviluppati per studiare l‟(in)tolleranza verso l‟incertezza e/o la
propensione all‟azzardo (Mobini et al., 2000; Cardinal & Howes, 2005; Adriani et al., 2006; Wilhelm & Mitchell, 2008).
“Probabilistic Delivery Task (PDT)”: scelta tra una ricompensa di cibo piccola e sicura, oppure una
ricompensa grande che verrà consegnata o meno sulla base del caso - quindi, potrà arrivare oppure
no in maniera random (Adriani & Laviola, 2006; Adriani et al., 2006).
“Iowa Gambling Task (IGT)”: scelta tra bassa probabilità di ottenere un premio grande o elevata
probabilità di ottenere un premio piccolo (van den Bos et al., 2006).
“Risky Decision-Making Task (RDT)”: scelta tra una ricompensa di cibo piccola ma sicura, oppure
una più grande ma associata al rischio di ottenere in maniera random anche una punizione (per es.,
una lieve scossa elettrica) (Simon et al., 2009).

“Incertezza”, “Rischio”, “Perdita”
“Incertezza” e “Rischio” non sono sinonimi
Il “rischio di perdere” è diverso dal “non riuscire a vincere”
“Rischio”: implica una conseguenza apertamente negativa (per es., lieve scossa elettrica alle zampe)
“Rischio di perdere”: quando una scommessa non va a buon fine, si hanno meno risorse rispetto alla
situazione di partenza
“Non riuscire a vincere”: assenza di qualsiasi ulteriore guadagno, situazione che causa “frustrazione”
Molti paradigmi di scelta operante (Adriani & Laviola, Cardinal and Howes, 2005; Mobini et al., 2000 van den
Bos et al., 2006) hanno a che fare esclusivamente con il “non riuscire a vincere” (per es., omissione della
ricompensa, oppure: consegna di un premio sgradevole): non v‟è nessun rischio di finire la sessione in
condizione di svantaggio rispetto alla situazione di partenza
Ogni caso di insuccesso è un evento “sfortunato” - che causa una “frustrazione” - ma non si tratta
necessariamente di un “rischio”
L‟attrazione verso una ricompensa incerta può essere correlata alla “propensione all‟azzardo”, ma
non riflette necessariamente il concetto di “propensione al rischio”

paradigmi operanti per lo studio di
Impulsività: Intolerance to Delay (ID) task
Propensione all‟azzardo: Probabilistic Delivery (PD) task
Lo studio della impulsività e della propensione all‟azzardo nel nostro
laboratorio
Classiche gabbie operanti o pannelli operanti all‟interno delle gabbie di stabulazione

Classiche gabbie operanti
(Skinner-box)
pannelli operanti
all’interno delle gabbie di
stabulazione

A
A
B B
C C
D D
A: Fori per il nose-poking
B: Luce, indica:
Durata del ritardo (ID task)
Flash (4 s) tra il nose-poke e la
consegna del rinforzo (PD task)
C: scomparto, dove vengono
rilasciati i pellet di cibo
D: luce - illumina lo scomparto
dei pellet, e segnala la durata
del timeout (TO)
Rappresentazione schematica di un apparato
Timeout (TO): un breve periodo
(di circa 30 s) successivo alla
consegna del cibo, in cui ogni
ulteriore nose-poking viene
registrato, ma non ha alcuna
conseguenza (detti, pertanto,
nose-pokes inadeguati)
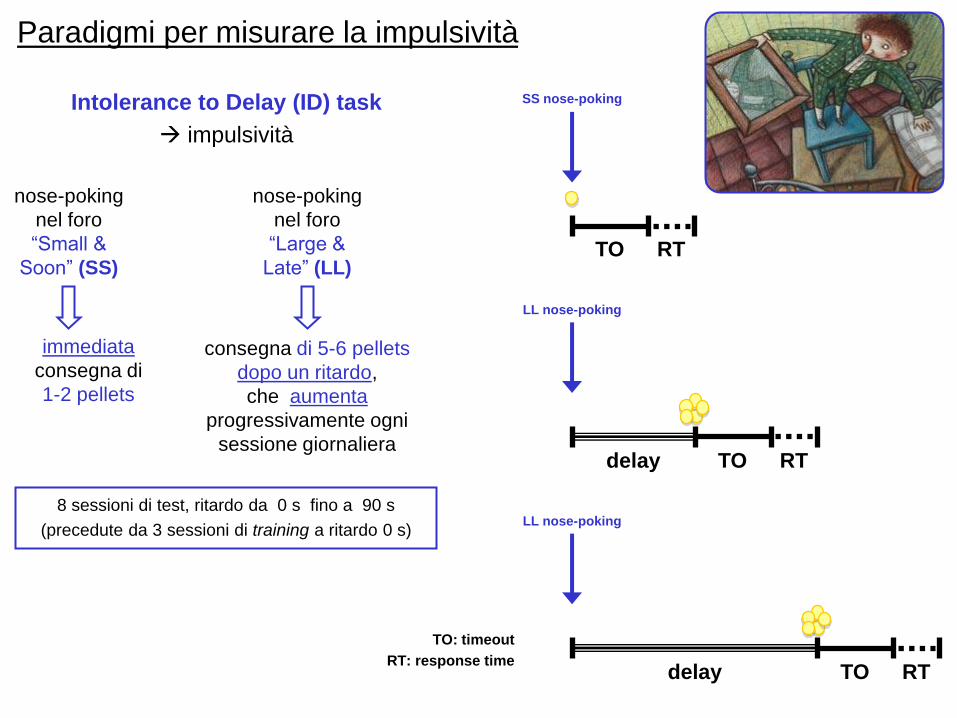
Paradigmi per misurare la impulsività
nose-poking
nel foro
“Small &
Soon” (SS)
immediata
consegna di
1-2 pellets
nose-poking
nel foro
“Large &
Late” (LL)
consegna di 5-6 pellets
dopo un ritardo,
che aumenta
progressivamente ogni
sessione giornaliera
TO RT
SS nose-poking
delay TO RT
LL nose-poking
delay TO RT
LL nose-poking8 sessioni di test, ritardo da 0 s fino a 90 s
(precedute da 3 sessioni di training a ritardo 0 s)
Intolerance to Delay (ID) task
impulsività
TO: timeout
RT: response time

Probabilistic Delivery (PD) task
Propensione all‟azzardo
Paradigmi per misurare la propensione all‟azzardo
11 sessioni di test, da una probabilità del 99% fino
ad arrivare al 9% (precedute da 2 sessioni di
training a probabilità pari al 99%)
TO RT
SS nose-poking
TO RT
LLL nose-poking
LLL nose-poking
TO RT
X
nose-poking
nel foro
“Small &
Sure” (SS)
sicura
consegna di
1-2 pellets
nose-poking nel
foro “Large &
Luck-Linked”
(LLL)
consegna (o no) di 5-6 pellets,
a seconda del livello di probabilità
percentuale “p”, che diminuisce
progressivamente ogni giorno
TO: timeout
RT: response time

Intolerance to
Delay (ID) task
Probabilistic
Delivery (PD) task
TO: timeout
RT: response time
Il punto di indifferenza
Entrambe le scelte sono
identiche, da un punto di
vista puramente matematico,
in termini di guadagno totale

Differenze di età nella propensione all‟azzardo

Comportamento di propensione all‟azzardo leggermente aumentato nei ratti adolescenti (pnd 36-49)
rispetto agli adulti (pnd 67-80) .
Livelli di propensione all’azzardo
-60
-50
-40
-30
-20
-10
0
middle adolescents adults
age
slo
pe
of
pre
fere
nc
e-o
dd
s c
urv
e
#P = 0.07
0
25
50
75
100
0 5 10 15 20 25 30
experienced odds
% c
ho
ice o
f th
e larg
e r
ew
ard
GAMBLING PART(n = 12)
(n = 14)
0
2
4
6
8
10
50 33 25 20 17 14 11
level of probability "p" (%)
n in
ad
eq
ua
te n
os
e-p
ok
es
x t
ria
l
middle adolescents
adults

Frequenza negli adulti paragonabile a quanto riscontrato in precedenza in ratti adulti testati
individualmente (Zoratto et al., 2012); frequenza sorprendentemente bassa negli adolescenti.
Frequenza di risposte
inadeguate, come indice di
“frustrazione”
* **** **
** ** ** **** ** **
0
2
4
6
8
10
50 50 33 33 25 25 20 20 17 17 14 14 11 11
level of probability "p" (%)
n in
ad
eq
ua
te n
os
e-p
ok
es
x t
ria
l
GAMBLING PART
(n = 12)
(n = 14)
0
2
4
6
8
10
50 33 25 20 17 14 11
level of probability "p" (%)
n in
ad
eq
ua
te n
os
e-p
ok
es
x t
ria
l
middle adolescents
adults

Considerando i tratti comportamentali tipici dell‟adolescenza, ci aspettavamo che i ratti adolescenti
dovessero essere molto più soggetti all‟ attrazione verso una ricompensa incerta
Nose-poke inadeguati in reazione all‟omissione della ricompensa sono un indice di frustrazione; gli
adolescenti potrebbero dunque essere “insensibili” alla perdita di una ricompensa.
La risposta cerebrale e comportamentale alla punizione è ridotta? improbabile!
Aumento del valore soggettivo del rinforzo secondario? ruolo delle luci …
Fori di nose-poking
Luce, nei
due fori di
nose-
poking
Luce, di
“casa”
Deposito di cibo
Luce, del
deposito
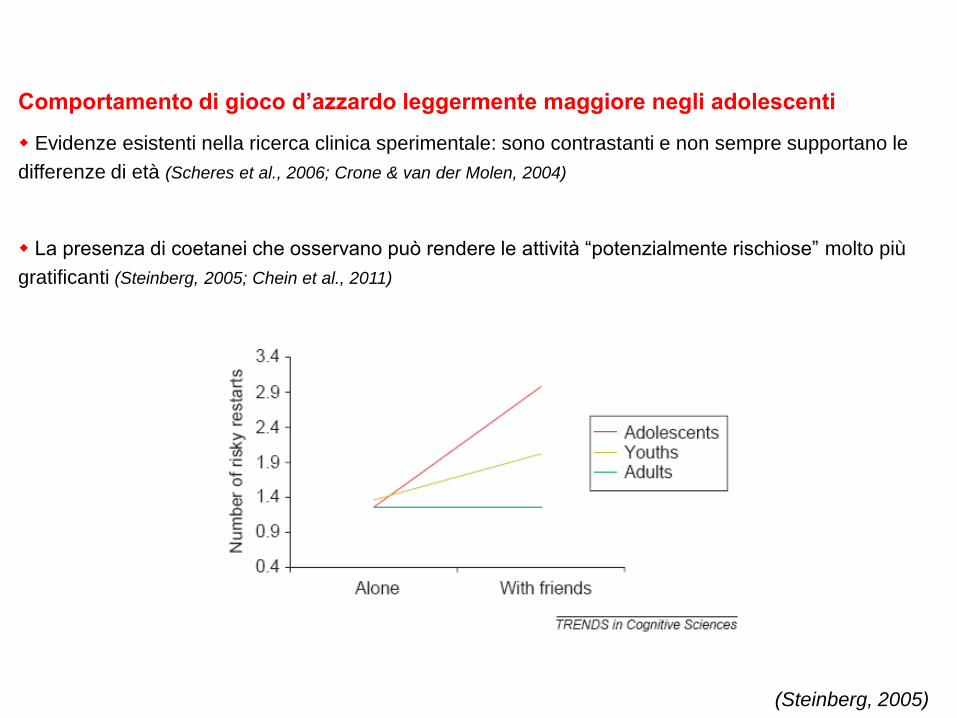
Comportamento di gioco d’azzardo leggermente maggiore negli adolescenti
Evidenze esistenti nella ricerca clinica sperimentale: sono contrastanti e non sempre supportano le
differenze di età (Scheres et al., 2006; Crone & van der Molen, 2004)
La presenza di coetanei che osservano può rendere le attività “potenzialmente rischiose” molto più
gratificanti (Steinberg, 2005; Chein et al., 2011)
(Steinberg, 2005)
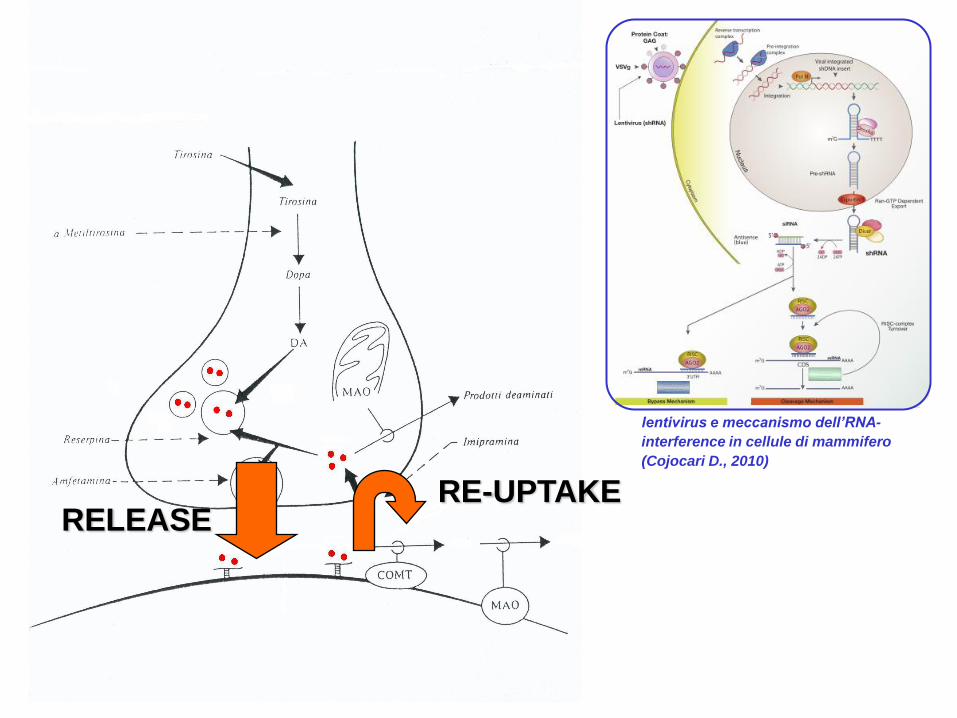
RELEASERE-UPTAKE
lentivirus e meccanismo dell’RNA-
interference in cellule di mammifero
(Cojocari D., 2010)

Farmaci usati per il trattamento dell‟ADHD: metilfenidato (Ritalin)

Manipolazione dell‟espressione del DAT con lentivirus

Delay-intolerance task. I ratti
hanno libera scelta tra:
- un solo pellet di cibo, ma
che viene consegnato
immediatamente, o
- cinque pellets di cibo,
consegnati con un certo
ritardo che aumenta
progressivamente.
DAT+
SIL
GFP
choice to delay
MODELLO DI IMPULSIVITÁ /
PROPENSIONE ALL’AZZARDO
Livelli di impulsività cognitiva
Adriani W, Boyer F, Gioiosa L, Macrì S, Dreyer J-L, Laviola G (2009) Neuroscience, 159: 47-58.

Scelta (%) della ricompensa grande ma incerta (per es., un pellet di cibo di sicuro vs. cinque pellets, consegnati o
meno, al progressivo diminuire delle probabilità).
Il compito è stato ripetuto 3 volte: ratti esposti alla
doxyciclina (doxy) nella seconda volta, per spegnere qualsiasi over-espressione
esogena del DAT.
A sinistra: media delle 3 replicazioni. A destra: ultimo punto rappresentato (p=6%)
per la prima (no doxy), seconda (sotto doxy) e
terza (no doxy) replicazione.
+DOXY
DAT+
SIL
DAT+SIL
GFP
Replication
choice
Percent (%) of
MODELLO DI IMPULSIVITÁ /
PROPENSIONE ALL’AZZARDO
Livelli di propensione
all’azzardo nel ratto

Conclusioni
In generale, il miglioramento di test quali ID e PD, e lo sviluppo di modelli animali innovativi, sono
essenziali per aumentare le conoscenze circa i meccanismi neurobiologici che caratterizzano disturbi
neuropsichiatrici, e per lo sviluppo di nuovi approcci terapeutici .
Impulsività e propensione all‟azzardo vengono modulati dall‟ età, dal sesso, da fattori genetici
(polimorfismi), da cambiamenti ambientali (stress, alterazione della dieta, trattamento farmacologico),
che agiscono andando a modificare dei marcatori specifici fondamentali del sistema dopaminergico e
serotoninergico.
Obiettivi futuri
Studi preclinici sul trattamento con farmaci attualmente usati per:
Giocatori d‟azzardo: antagonisti degli oppioidi (naltrexone,
nalmefene); inibitori selettivi di ricaptazione della serotonina,
SSRI (paroxetine, fluvoxamine)
Morbo di Parkinson: levodopa; terapie per il ripristino della
dopamina (agonisti di recettori della dopamina)
Lo svolgimento dei protocolli, effettuabili direttamente nelle gabbie
di stabulazione, ci può permettere di migliorare sia il benessere degli
animali sia la qualità dei dati sperimentali.

Hikikomori
Forme estreme di ritiro sociale sono state identificate in Giappone nei primi anni „90
I giovani si isolano nelle loro case, o camerette, per mesi o addirittura per anni
Alcuni credono che questo fenomeno possa diffondersi in tutto il mondo
Mancanza di una solida ricerca clinica
by Maggie Jones, The New York Times (January 15, 2006)
“ … spesso stanno svegli tutta la notte, ad usare
Internet, a giocare con il computer, o a guardare la
TV, e poi dormono tutto il giorno … ”

Grazie per l’attenzione
Si ringrazia, per il prezioso contributo:
Francesca Zoratto
Giovanni Laviola
Funding source:
- Italian Ministry of Health, with “under 40” Young-Investigator Project “ADHD-sythe” and
EU-FP7 “Prio-Med-Child” ERAnet Project “NeuroGenMRI” (both coordinated as PI by WA)
- The Dipartimento Politiche Antidroga, Presidenza del Consiglio dei Ministri (grant to GL)
Immagine (ratto di laboratorio) proveniente dalla
biblioteca “Understanding Animal Research”


SPETTROSCOPIA DI RISONANZA
MAGNETICA
Dati ottenuti da:
ISS (dr.ssa R. Canese)
Dorsal Striatum
Nucleus Accumbens
Prefrontal Cortex

SPETTROSCOPIA DI RISONANZA
MAGNETICA in:
striato, n. accumbens, corteccia prefrontale
I principali metaboliti possono essere così raggruppati:
tCr/PCr= total / phospho- creatine tCho = total choline
NAA = N-acetyl-aspartate Glx = glutamine + glutamate
Tau = taurine Ins = inositols phosphates
NAAtCrtCho
Glx
tCr
GluInsTau

SPETTROSCOPIA RISONANZA MAGNETICA
in: striato, n.accumbens, corteccia pre-frontale
Parametri metabolici in dStr e NAcc di ratti adulti precedentemente
inoculati con vettori lentivirali (DAT+ and/or SIL) a livello di NAcc.
Dorsal Striatum PCr/tot (%) Phospho-creatine Total creatine
GFP control 65.4±3.4 % 6.29±0.54 9.86±0.41
DAT+ 63.0±2.7 % 6.60±0.30 * 10.46±0.10 *
SIL 64.8±2.6 % 6.24±0.52 9.57±0.46
DAT+SIL 65.5±2.1 % 6.97±0.35 * 10.62±0.29 *
N. Accumbens PCr/tot (%) Phospho-creatine Total creatine
GFP control 64.3±1.5 % 6.49±0.17 10.10±0.26
DAT+ 67.5±2.3 % 6.72±0.27 9.96±0.22
SIL 59.7±3.4 % * 5.63±0.44 * 9.56±0.26
DAT+SIL 59.1±3.6 % * 5.84±0.32 * 9.80±0.26
Livelli di metaboliti indicati in unità arbitrarie, riferiti al segnale d‟acqua non soppresso. (*) p
< .05 confrontato con ratti di controllo GFP. Solo ratti DAT+SIL presentano cambiamenti in
entrambe le aree.

Interessante come, creatina e/o fosfo-creatina (metaboliti
bioenergetici) erano “up-regulated” in dStr e contrariamente
“down-regulated” in NAcc di ratti DAT+SIL. La diversa
regolazione di queste due aree non genera un comportamento
impulsivo e/o di propensione al rischio.
Coerentemente con il ruolo funzionale di queste due aree del
prosencefalo, gli animali DAT+SIL possono presentare un
aumento della loro abilità di coping utilizzando nuove strategie
comportamentali (dStr), ed essere meno guidati dall’attrattività
della ricompensa o dalle caratteristiche salienti del test (NAcc).
COMPORTAMENTO OPERANTE E
SPETTROSCOPIA MR
Ruolo di striato dorsale vs n. accumbens nell’autocontrollo

Differenze di sesso nella propensione all‟azzardo
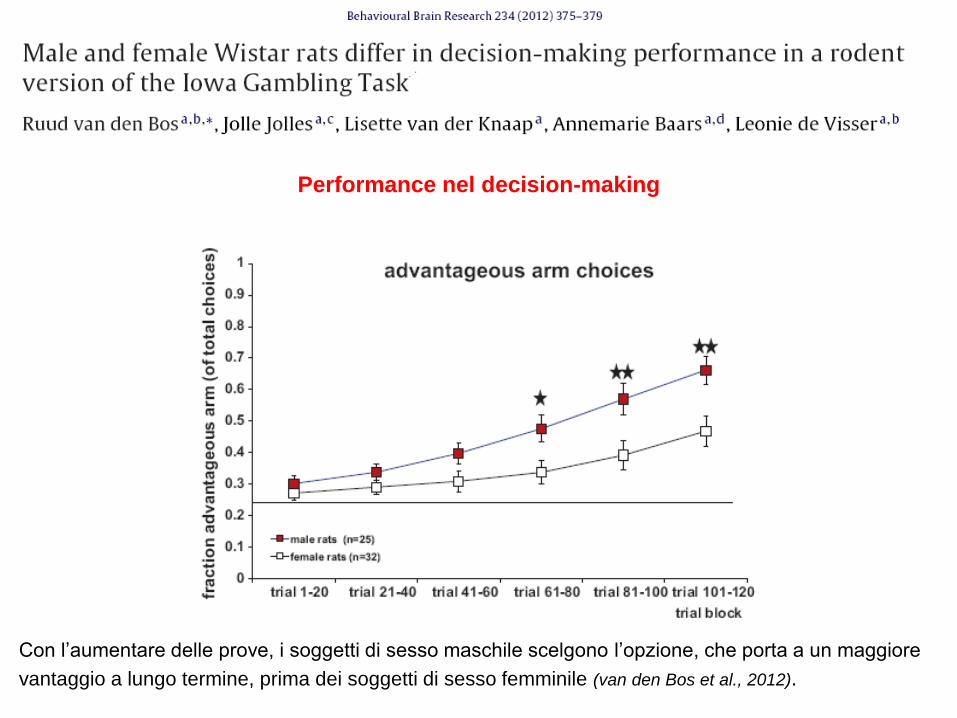
Con l‟aumentare delle prove, i soggetti di sesso maschile scelgono l‟opzione, che porta a un maggiore
vantaggio a lungo termine, prima dei soggetti di sesso femminile (van den Bos et al., 2012).
Performance nel decision-making

Impulsivity and home-cage activity are decreased by
lentivirus-mediated silencing of serotonin transporter
in the rat hippocampus
Zoratto F., Laviola G., Adriani W.
Neuroscience Letters (submitted)
Modificazione dell‟espressione del SERT mediante lentivirus

Soggetti sperimentali
attività locomotoria spontanea:
Ratti maschi adulti
All‟interno delle gabbie di stabulazione, 50
giorni dopo l‟inoculo
Registrazione continua automatica (per 15
giorni) usando sensori ad infrarossi (20 Hz)
Giornata media calcolata su intervalli di 5
giorni consecutivi
Test di Intolleranza al ritardo (ID):
Analizzati in classiche gabbie operanti,
durante la fase di buio del ciclo
Lieve restrizione alimentare
test: 8 giorni, 1 sessione al giorno (40 min),
timeout 30 s
Gruppi sperimentali
Lenti-SERT (n=11): inoculo di un lentivirus attivo
controlli (n=6): inoculo di un lentivirus inattivato con
alte temperature
Inoculo bilaterale (volume di 1 µl) a livello
dell‟ippocampo; AP −3.3, ML ±2.0, DV −4.0
0
20
40
60
80
100
LV-siSERT1 LV-siSERT2 LV-siSERT3 mix
avera
ged
SE
RT
exp
ressio
n l
evel
(%)
In-vitro quantification of SERT silencing
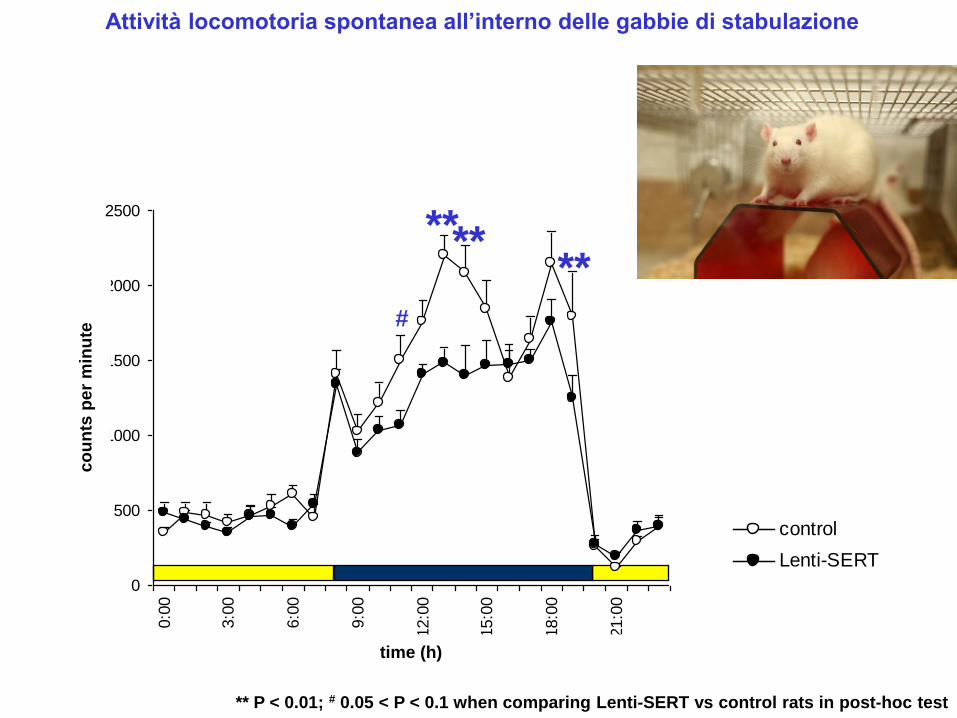
Attività locomotoria spontanea all’interno delle gabbie di stabulazione
** P < 0.01; # 0.05 < P < 0.1 when comparing Lenti-SERT vs control rats in post-hoc test
0
500
1000
1500
2000
2500
0:0
0
3:0
0
6:0
0
9:0
0
12:0
0
15:0
0
18:0
0
21:0
0
time (h)
co
un
ts p
er
min
ute
control
Lenti-SERT ****
#
**
co
un
ts p
er
min
ute
time (h)
0,0
0,5
1,0
1,5
2,0
2,5
0 7 15 30 45 60 75 90
delay (s)
n i
nad
eq
uate
no
se-p
okes x
tri
al
control
Lenti-SERT

Livelli di impulsività cognitiva
* P < 0.05 when comparing Lenti-SERT vs control rats in post-hoc test
0
20
40
60
80
100
0 7 15 30 45 60 75 90
delay (s)
% c
ho
ice o
f th
e l
arg
e r
ew
ard
(L
L)
control
Lenti-SERT
*
chance level
% c
ho
ice o
f th
e l
arg
e r
ew
ard
delay (s)
0,0
0,5
1,0
1,5
2,0
2,5
0 7 15 30 45 60 75 90
delay (s)
n i
nad
eq
uate
no
se-p
okes x
tri
al
control
Lenti-SERT

Livelli di impulsività motoria
* P < 0.05 quando confrontiamo ratti Lenti-SERT vs ratti di controllo
0,0
0,5
1,0
1,5
2,0
2,5
0 7 15 30 45 60 75 90
delay (s)
n i
nad
eq
uate
no
se-p
okes x
tri
al
control
Lenti-SERT *
n i
na
de
qu
ate
no
se
-po
kes x
tri
al
delay (s)
0,0
0,5
1,0
1,5
2,0
2,5
0 7 15 30 45 60 75 90
delay (s)
n i
nad
eq
uate
no
se-p
okes x
tri
al
control
Lenti-SERT

La funzionalità ridotta del SERT ha un impatto significativo sulla attività e sui profili
di impulsività
Questi profili confermano la relazione inversa esistente tra 5-HT e impulsività, come riportata da studi
effettuati con varie manipolazioni a livello del sistema serotoninergico.
Considerando la complessità del sistema serotoninergico, potrebbe non essere sorprendente la
correlazione tra la variante allelica “s” e l‟incremento della impulsività, riportata nella ricerca clinica.
L‟associazione tra il genotipo s/s e la diminuzione dell‟espressione di SERT, nel sistema nervoso
centrale umano, non è stata ancora dimostrata del tutto.

Differenze di sesso nella impulsività

I maschi di controllo sono meno in grado, rispetto alle femmine, di inibire il comportamento di nose
poking durante il ritardo. E‟ interessante notare che il profilo mostrato dai maschi trattati con BPA è
paragonabile a quello espresso dalle femmine (Adriani et al., 2003).
Livelli di impulsività motoria

All‟aumentare dei ritardi, topi di sesso femminile scelgono l‟opzione economicamente più vantaggiosa
prima dei topi maschi (Koot et al., 2009).
Livelli di impulsività cognitiva
indifferent
point

Differenze di età nella impulsività
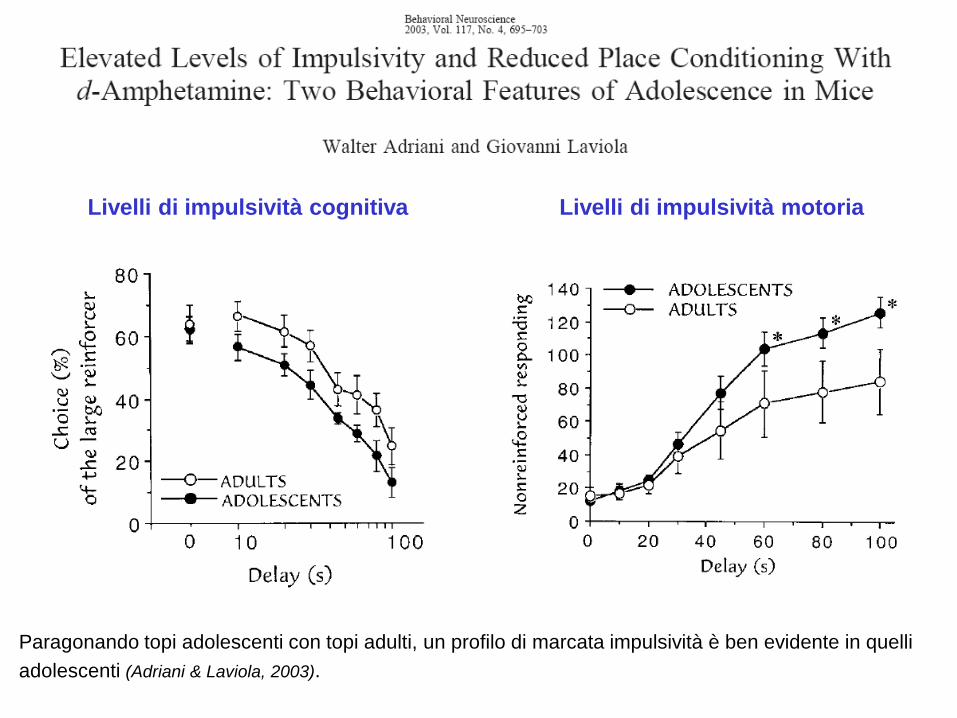
Paragonando topi adolescenti con topi adulti, un profilo di marcata impulsività è ben evidente in quelli
adolescenti (Adriani & Laviola, 2003).
Livelli di impulsività cognitiva Livelli di impulsività motoria

EFFETTI PERMANENTI DOVUTI A
SOMMINISTRAZIONE DI MPH IN ADOLESCENTI
Somministrazione di metilfenidato (MPH, Ritalin) può
determinare la modulazione del comportamento impulsivo
attraverso:
- un incremento permanente della neurotrasmissione di 5-
HT(7).
- cambi funzionali permanenti all’interno di circuiti fronto-
striatali.
DISEGNO SPERIMENTALEIn ratti adolescenti (dai 30 ai 44 giorni di vita) viene somministrato MPH (2
mg/kg i.p.) o una soluzione salina per 14 giorni, e poi, diventati adulti,
vengono testati, per :
1) cambiamenti a livello di impulsività (intolleranza al ritardo)
2) blocco degli effetti del MPH usando un antagonista di 5-HT(7)
3) ottenimento degli effetti “MPH-like” usando un agonista di 5-HT(7)
4) risonanza magnetica: spettrometria, DTI, fMRI, connettività

Tolleranza al
ritardo = minore
impulsività
INTOLERANCE TO REWARD DELAY
Delay (sec)
0 7.5 15 30 45 60 75
% choic
e of the
large re
inforcer
40
45
50
55
60
65
70
75
80
85
90
95
100
saline
MPH
**
*
EFFETTO A
LUNGO
TERMINE DEL
MPH
RATTI ANALIZZATI
DURANTE IL PERIODO
DELL’ADOLESCENZA , UN
MESE DOPO IL
PRETRATTAMENTO CON
MPH
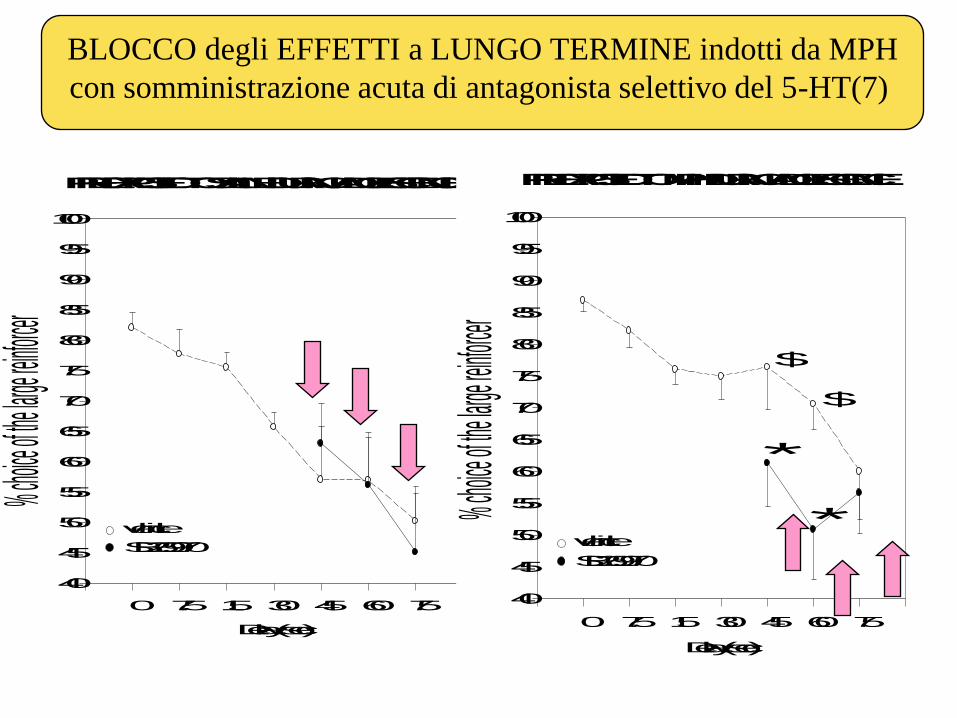
PRE-EXPOSED TO SALINE DURING ADOLESCENCE
Delay (sec)
0 7.5 15 30 45 60 75
% choic
e of the
large re
inforcer
40
45
50
55
60
65
70
75
80
85
90
95
100
vehicle
SB 269970
PRE-EXPOSED TO MPH DURING ADOLESCENCE
Delay (sec)
0 7.5 15 30 45 60 75
% choic
e of the
large re
inforcer
40
45
50
55
60
65
70
75
80
85
90
95
100
vehicle
SB 269970
*
*
$
$
BLOCCO degli EFFETTI a LUNGO TERMINE indotti da MPH
con somministrazione acuta di antagonista selettivo del 5-HT(7)
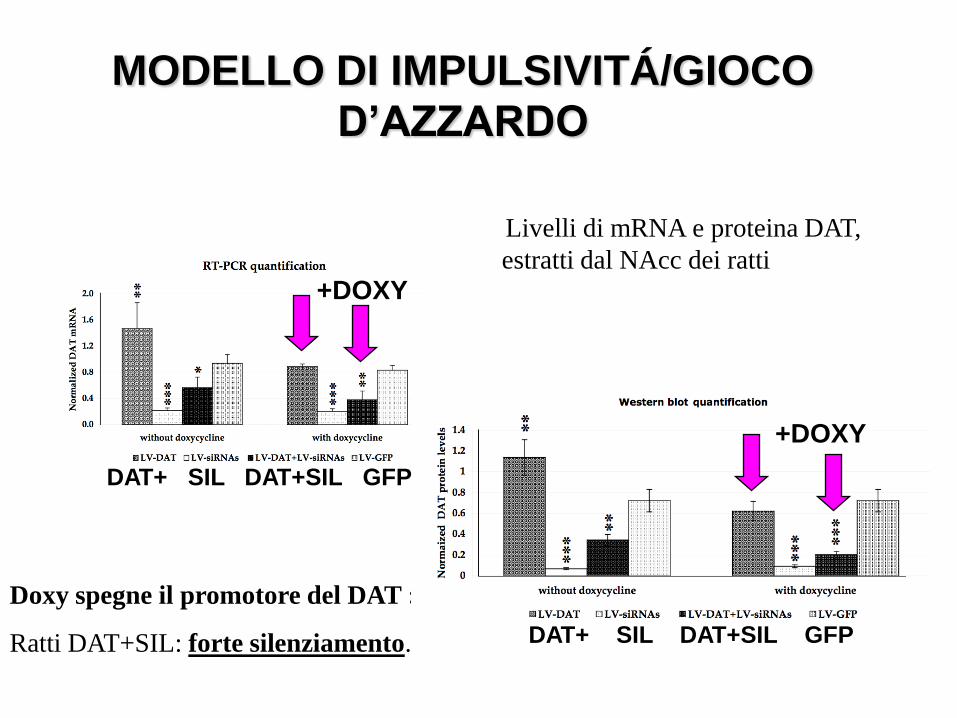
Livelli di mRNA e proteina DAT,
estratti dal NAcc dei ratti
Doxy spegne il promotore del DAT :
Ratti DAT+SIL: forte silenziamento.
DAT+ SIL DAT+SIL GFP
DAT+ SIL DAT+SIL GFP
+DOXY
+DOXY
MODELLO DI IMPULSIVITÁ/GIOCO
D’AZZARDO

MODELLO DI IMPULSIVITÁ /
PROPENSIONE ALL’AZZARDO
DAT+ DAT+SIL
+DOXY
Livelli di
controllo
(colorazione
DAT)
Adriani W, Boyer F, Gioiosa L, Macrì S, Dreyer J-L, Laviola G (2009) Neuroscience, 159: 47-58.
Efficacia di lentivirus inoculati nel Nacc di ratti: Lenti-DAT con o senza Lenti-DAT- siRNAs
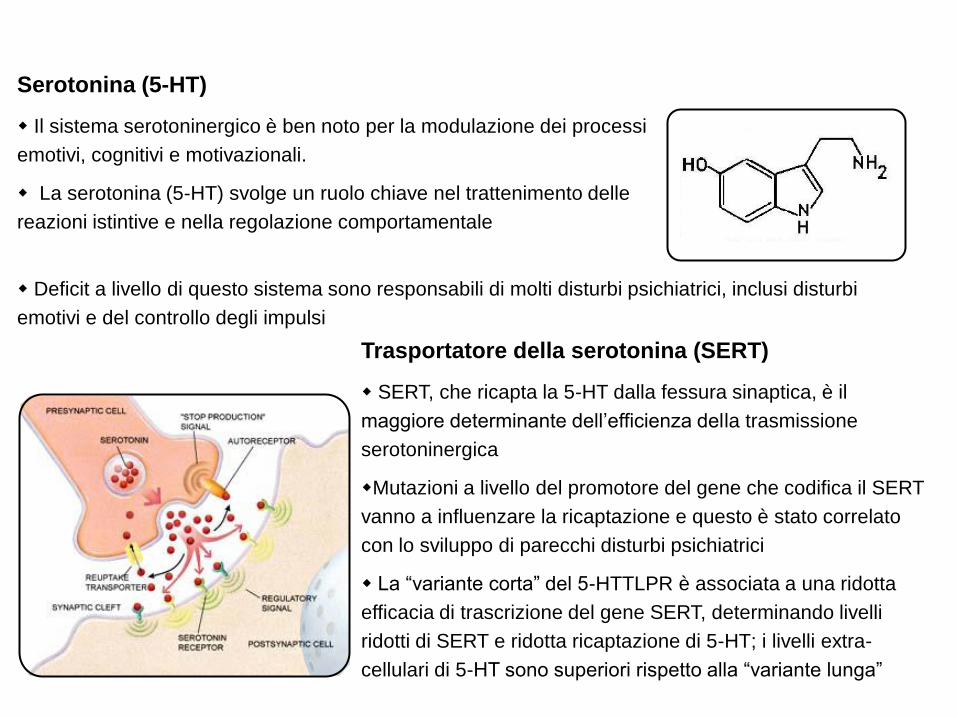
Serotonina (5-HT)
Il sistema serotoninergico è ben noto per la modulazione dei processi
emotivi, cognitivi e motivazionali.
La serotonina (5-HT) svolge un ruolo chiave nel trattenimento delle
reazioni istintive e nella regolazione comportamentale
Deficit a livello di questo sistema sono responsabili di molti disturbi psichiatrici, inclusi disturbi
emotivi e del controllo degli impulsi
Trasportatore della serotonina (SERT)
SERT, che ricapta la 5-HT dalla fessura sinaptica, è il
maggiore determinante dell‟efficienza della trasmissione
serotoninergica
Mutazioni a livello del promotore del gene che codifica il SERT
vanno a influenzare la ricaptazione e questo è stato correlato
con lo sviluppo di parecchi disturbi psichiatrici
La “variante corta” del 5-HTTLPR è associata a una ridotta
efficacia di trascrizione del gene SERT, determinando livelli
ridotti di SERT e ridotta ricaptazione di 5-HT; i livelli extra-
cellulari di 5-HT sono superiori rispetto alla “variante lunga”

New Scientist (5 March 2005)
Age differences in humans

Sex differences in humans: anatomical MRI
Fully processed images from one 8-year-old
female participant (Sowell et al., 2002).
Total cerebral volume peaks at 14.5 years in
males and 11.5 years in females.
Male brains are approx. 9% larger than those of
females. This difference is statistically significant,
even when controlling for height and weight
(Lenroot & Giedd, 2006).
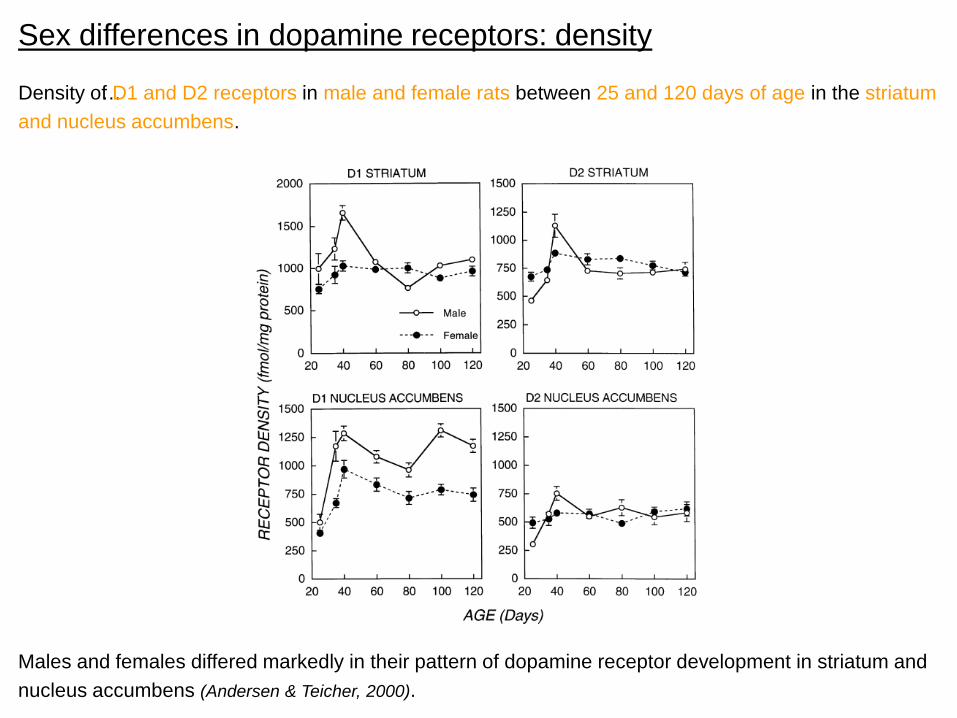
Sex differences in dopamine receptors: density
Density o…f D1 and D2 receptors in male and female rats between 25 and 120 days of age in the striatum
and nucleus accumbens.
Males and females differed markedly in their pattern of dopamine receptor development in striatum and
nucleus accumbens (Andersen & Teicher, 2000).

Sex differences in dopamine receptors: laterality
These data show the pronounced lateralization of D1 dopamine receptors for females, and the transient
expression of D2 receptor lateralization in male striatum at 40 days of age (Andersen & Teicher, 2000).
Laterality indices …for D1 and D2 receptors in male and female rats between 25 and 120 days of age in
the striatum and nucleus accumbens.

Age differences in dopamine receptors: density
Developmental changes in the density of D1 and D2 receptors from weanling (25 days of age) to the
onset of puberty (40 days of age) to full adulthood (120 days) in male rats.
Marked reduction in the density of D1 and
D2 receptors in the prefrontal cortex and
striatum between 40 and 120 days of age.
The pattern of receptor elimination is
similar, although more extensive and
protracted in prefrontal cortex.
The nucleus accumbens shows minimal
evidence of receptor elimination.
(Andersen & Teicher, 2000)


The three group-averaged functional maps of percentage signal change illustrate differences in both the
magnitude and extent of activation.
Children showed strongest activation bilaterally in the caudate nucleus, the thalamus and the anterior
insula. Adolescents showed strongest activation in the right dorsolateral prefrontal cortex (DLPFC), and
adults showed concentrated activation in the left prefrontal and posterior parietal regions (Bunge & Wright,
2007).
Developmental shifts in the location of active voxels during
performance of a visuospatial working memory

Depletion of forebrain 5-HT levels by means of ICV infusions of the
serotonergic neurotoxin 5,7-dihydroxytryptamine

Lack of effect of the same manipulation on delay-discounting (Winstanley et al., 2004).
(a) Effects of reducing 5-HT levels in the central nervous system on the acquisition of conditioned
locomotor activity and (b) effects of prefeeding prior to conditioning sessions (Winstanley et al., 2004).

Neonatal serotonin depletion


Dietary serotonin depletion

The relationship between 5-HT and impulsivity
Experimental studies with different serotonergic
manipulations have demonstrated an inverse relationship
between 5-HT levels and impulsivity, with a reduction in the
neurotransmitter causing an increase in impulsivity and vice
versa.
However, in humans, an increase in impulsivity appears to be
associated with the short (s) allelic variant of 5-HTTLPR,
leading to reduced SERT gene transcription.
We aimed to determine whether a partial silencing of the
SERT-encoding gene within the hippocampus could induce
alterations relevant to the modelling of ADHD, in particular
symptoms of hyperactivity and impulsivity. Lentiviral delivery of shRNAs and the
mechanism of RNAi interference in
mammalian cells (Cojocari D., 2010)

0
5
10
15
20
25
control depleted
co
ncen
trati
on
(p
g/m
g)
**
0
4
8
control depleted
co
ncen
trati
on
(p
g/m
g)
**
0
10
20
30
control depleted
co
ncen
trati
on
(p
g/m
g)
*
** P < 0.01; * P < 0.05 when comparing depleted vs control rats in post-hoc test
Experimental subjects
adult male rats tested in classical operant
chambers during the dark phase of the cycle
mild food restriction
testing: 11 days, 1 session per day (25
min), timeout 15 s
Experimental groups
depleted (n=12): food without TRP (0.0 g/kg)
control (n=12): food with a standard content
of TRP (2.8 g/kg)
from 2 weeks before and throughout the
duration of the experiment (until sacrifices)
5-HT in prefrontal cortex 5-HT in striatum 5-HT in hippocampus

** P < 0.01; * P < 0.05; # 0.05 < P < 0.1 when comparing depleted vs control rats in post-hoc test
0
20
40
60
80
100
99 80 66 50 33 25 20 17 14 11 9
depleted
control
gambling part
chance level
indifferent point
**
***
#*%
ch
oic
e o
f th
e l
arg
e r
ew
ard
set level of probability (%)
Levels of gambling proneness

# 0.05 < P < 0.1 when comparing depleted vs control rats in post-hoc test
0
,5
1
99 80 66 50 33 25 20 17 14 11 9
depleted
control
##
gambling part
n i
na
de
qu
ate
no
se
-po
kes x
tri
al
set level of probability (%)
Frequency of inadequate responding, an index of frustration

Punishment insensitivity and/or enhanced secondary reinforcement
In the clinical literature, it has been reported that:
gamblers may be insensitive to losses (Rachlin, 1990);
despite repeated losses, feelings of “near win” and the occasional large payoff seem to provide
sufficient reinforcement (Walker, 1992).
In our preclinical model, the nose-poking for large reward was constantly turning on the reward-
associated lights while occasionally triggering binge reward delivery: this may perhaps reproduce the
addictive features of slot machines.

PHARMACOLOGICAL TOOLS
Methylphenidate (MPH, Ritalin) is currentlyprescribed to ADHD children, since it decreasesimpulsivity and increases sustained attention (Ward et
al., 1997).
However, MPH interacts with the same brain pathways activated
by drugs of abuse, producing striatal dopamine (DA) overflow
similarly to cocaine (Marsteller et al., 2002). Its use raises
concerns for public health (Rapport & Moffitt, 2002;
Klein-Schwartz, 2002; Carlezon & Konradi, 2004).
Great effort is devoted to the identification ofnovel non-psychostimulant agents.

Interestingly, total creatine and taurine, reputed to be involved
in neural activation, were up-regulated in the STRIATUM and
conversely down-regulated in the NUCLEUS ACCUMBENS
of MPH-exposed rats. We suggest that the influence of these
two areas on behavioral output was modulated accordingly.
Based on the functional role played by these two brain areas,
MPH-exposed animals may be suggested to be more prone to
elaborate novel behavioral habits, in order to cope with actual
reinforcement delay (STRIATUM), and to be less instinctive,
i.e. less affected by the immediacy of reward (ACCUMBENS).
MAGNETIC RESONANCE SPECTROSCOPY
in: striatum, nucleus accumbens, and prefrontal
cortex

IMPULSIVITY IN THE SHR ADOLESCENT
A validated animal model for ADHD is the SHR(Spontaneously Hypertensive Rat) strain.
Aim of the work was to compare SHRs to theirWistar-Kyoto controls in a test for intolerance-to-delay during adolescence (PND 30 to 45), inorder to further validate the adolescent SHR as asuitable animal model for ADHD.
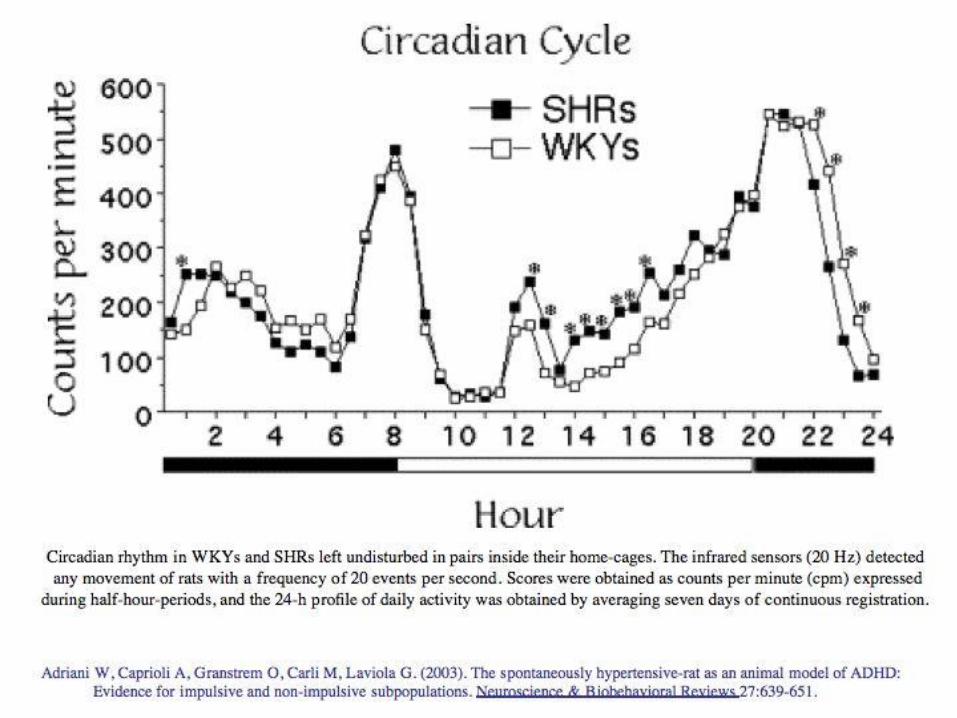
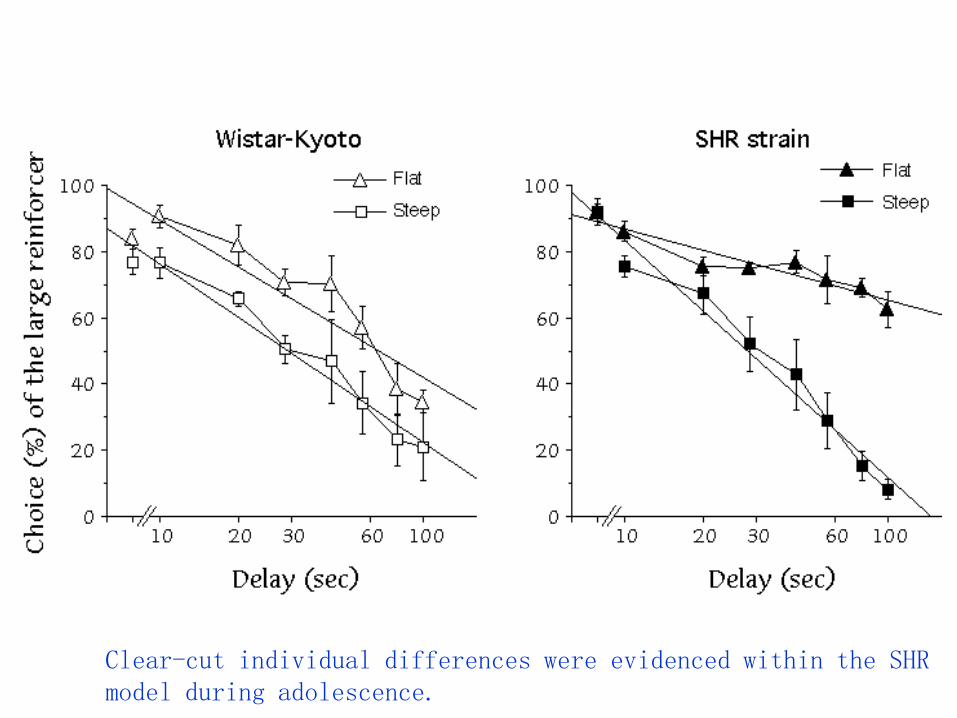
Clear-cut individual differences were evidenced within the SHR model during adolescence.

ONGOING EFFECTS of SUBCHRONIC MPH
DURING ADOLESCENCE
The ongoing treatment with MPH (3 mg/kg i.p.) reduced impulsivity
levels
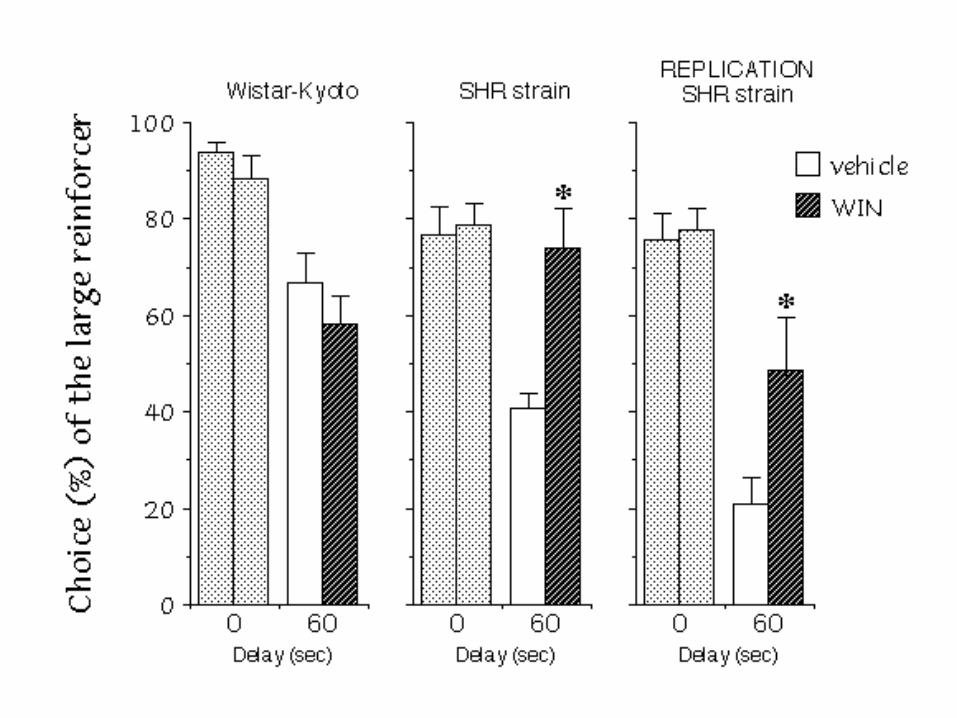

CONCLUSION - THE ADOLESCENT “SHR”
Clear-cut individual differences were evidenced within the SHR model during adolescence.
« STEEP » subpopulation showed elevated impulsivity. The ongoing treatment with MPH (3 mg/kg
i.p.) reduced impulsivity levels. Acute administration of WIN 55,212 (2 mg/kg s.c.) alsoreduced impulsivity in young-adult SHRs, without effect in WKYs.
FLAT subpopulation kept on choosing H5 even at higher delays
a) animals do not pay attention to the experimental contingency (see Sagvolden et al., 1998;Sagvolden & Sergeant, 1998; Sagvolden, 2000), as expected in an animal model of ADHD;
b) animals are unable to modulate response patterns according to changes in the experimentalcontingency (perseverative behavior).

CONCLUSIONS
This study combined behavioural analysis and genome-
wide approach, to investigate the molecular and
neurobehavioural changes resulting from subchronic MPH
exposure during adolescence.
In summary, such early treatment was able to modulate
striatal gene expression of Post Synaptic Density family in
the adolescent brain. These changes underlie enduring
plastic rearrangements in reward-related brain circuits, from
both structural and functional (dendritic spines,
neurotransmitter receptors) viewpoint.
The latter may in turn account for a complex picture of
increased self-control abilities at adulthood, associated with
an altered processing of incentive values.

* Abnormal Htr7 and/or DAT in adolescents or in ADHD patients is perhaps associated with patterns of impulsive decision-making and with risk-prone behaviours;
• The impact of stimulant drugs during such important developmental rearrangements received a fast-growing attention in preclinical experimental investigation (i.e. to model cases of illicit use for increased performance);
• It is thus emphasized the importance of characterizing novelty-seeking and risk-prone behaviour in the rodent, during adolescence and/or using new specific models.
CONCLUSIONS

Intolerance to Delay (ID) task
impulsivity
Probabilistic Delivery (PD) task
gambling proneness
Paradigms for the measure of impulsivity and gambling proneness
nose-poking
in “Small &
Sure” hole
(SS)
certain
delivery of
1-2 pellets
nose-poking in
“Large & Luck-
Linked” hole
(LLL)
delivery (or not) of 5-6 pellets,
according to the level of
probability “p”,
which is decreased
progressively each day
nose-poking
in “Small &
Soon” hole
(SS)
immediate
delivery of
1-2 pellets
nose-poking
in “Large &
Late” hole
(LL)
delivery of 5-6 pellets
after a delay,
which is increased
progressively each day
8 testing sessions, from delay 0 s to 90 s
(preceded by 3 training sessions at delay 0 s)
11 testing sessions, from probability 99% to 9%
(preceded by 2 training sessions at probability 99%)
under conditions of mild food restriction
(to increased motivation to work for food delivery)

odds = 1 odds = 2
il delay è uguale al mITI il delay è il doppio del mITI
Delay-equivalent odds = delay / mITI = delay / (TO + RT)
“Large & Luck-Linked” hole (LLL)
“Small & Sure” hole (SS) certain delivery of 2 pellets
delivery (or not) of 6 pellets, according
to the level of probability “p”
“Large & Late” hole (LL)
“Small & Soon” hole (SS) immediate delivery of 1 pellets
delivery of 5 pellets, after a delay

EXPERIMENT OUTLINE
TRP
5-HT