LEGGI DI SCALA IN BIOLOGIA E DINAMICA DELLO...
12
24 < IL NUOVO SAGGIATORE Molti fenomeni in ambito fisico, fisiologico, ecologico…, sono descritti in maniera quantitativamente corretta da leggi piuttosto semplici, dette leggi di scala. Tali leggi hanno giocato un ruolo non secondario nello sviluppo di teorie fisiche ed il loro emergere in ambito biologico fa ritenere che possano assumere un ruolo di indirizzo altrettanto importante. Alcune relazioni, come quelle che regolano il rapporto tra metabolismo basale e masse corporee, sembrano ricoprire un ruolo fondamentale che va al di là della biologia stessa. Nel corso dell’articolo se ne discuterà l’importanza e si vedrà come da esse possano essere determinate leggi di crescita inclusa quella delle masse tumorali. 1 Introduzione Molti fenomeni, per quanto apparentemente complicati, sono caratterizzati da “leggi” estremamente semplici, ovvero da relazioni tra pochi parametri macroscopici, in grado di riprodurre quantitativamente la fenomenologia osservata. In passato leggi di tale tipo hanno avuto un ruolo determinante nello sviluppo delle teorie fisiche. La legge di Balmer permette, ad esempio, di riprodurre quanto verificato sperimentalmente in merito alle righe spettrali dell’idrogeno atomico, tramite il solo uso di una costante e di un rapporto tra numeri interi. Tale legge, formulata, come ben noto, prima LEGGI DI SCALA IN BIOLOGIA E DINAMICA DELLO SVILUPPO DELLE MASSE TUMORALI GIUSEPPE DATTOLI, PIER LUIGI OTTAVIANI*, SIMONETTA PAGNUTTI * ENEA, Tecnologie Fisiche e Nuovi Materiali, Centro Ricerche Frascati Frascati, Roma, Italia Le leggi di scala hanno avuto, ed hanno, un impatto notevolissimo in Fisica e non solo. In questo articolo si ricorda il significato metodologico di tali leggi e l’importanza da esse rivestita in ambito di problematiche biologiche ed ecologiche. Una delle leggi di scala, che ha suscitato maggiore interesse negli ultimi anni, è la legge di Kleiber, che lega la potenza metabolica di un determinato sistema biologico alla sua massa. Il suo utilizzo, associato a considerazioni di natura energetica, permette di ottenere curve universali di crescita delle masse dei mammiferi ed è stato recentemente esteso alla evoluzione delle masse tumorali, la cui dinamica può essere trattata tramite un insieme di equazioni molto simile a quelle applicate allo studio dei sistemi laser. L’articolo riporta alcuni recenti risultati sull’argomento, con particolare riferimento alla diffusione delle metastasi. * ENEA, Tecnologie Fisiche e Nuovi Materiali, Centro Ricerche Bologna. FISICA E...
Transcript of LEGGI DI SCALA IN BIOLOGIA E DINAMICA DELLO...

24 < il nUovo saggiatoRe
Molti fenomeni in ambito fi sico, fi siologico, ecologico…, sono descritti in maniera quantitativamente corretta da leggi piuttosto semplici, dette leggi di scala. Tali leggi hanno giocato un ruolo non secondario nello sviluppo di teorie fi siche ed il loro emergere in ambito biologico fa ritenere che possano assumere un ruolo di indirizzo altrettanto importante. Alcune relazioni, come quelle che regolano il rapporto tra metabolismo basale e masse corporee, sembrano ricoprire un ruolo fondamentale che va al di là della biologia stessa. Nel corso dell’articolo se ne discuterà l’importanza e si vedrà come da esse possano essere determinate leggi di crescita inclusa quella delle masse tumorali.
1 IntroduzioneMolti fenomeni, per quanto apparentemente complicati, sono caratterizzati da “leggi” estremamente semplici, ovvero da relazioni tra pochi parametri macroscopici, in grado di riprodurre quantitativamente la fenomenologia osservata.In passato leggi di tale tipo hanno avuto un ruolo determinante nello sviluppo delle teorie fi siche. La legge di Balmer permette, ad esempio, di riprodurre quanto verifi cato sperimentalmente in merito alle righe spettrali dell’idrogeno atomico, tramite il solo uso di una costante e di un rapporto tra numeri interi. Tale legge, formulata, come ben noto, prima
LEGGI DI SCALA IN BIOLOGIA E DINAMICA DELLO SVILUPPO DELLE MASSE TUMORALI
GIUSEPPE DATTOLI, PIER LUIGI OTTAVIANI*, SIMONETTA PAGNUTTI *ENEA, Tecnologie Fisiche e Nuovi Materiali, Centro Ricerche FrascatiFrascati, Roma, Italia
Le leggi di scala hanno avuto, ed hanno, un impatto notevolissimo in Fisica e non solo.In questo articolo si ricorda il signi� cato metodologico di tali leggi e l’importanza da esse rivestita in ambito di problematiche biologiche ed ecologiche.Una delle leggi di scala, che ha suscitato maggiore interesse negli ultimi anni, è la legge di Kleiber, che lega la potenza metabolica di un determinato sistema biologico alla sua massa. Il suo utilizzo, associato a considerazioni di natura energetica, permette di ottenere curve universali di crescita delle masse dei mammiferi ed è stato recentemente esteso alla evoluzione delle masse tumorali, la cui dinamica può essere trattata tramite un insieme di equazioni molto simile a quelle applicate allo studio dei sistemi laser.L’articolo riporta alcuni recenti risultati sull’argomento, con particolare riferimento alla di� usione delle metastasi.
* ENEA, Tecnologie Fisiche e Nuovi Materiali, Centro Ricerche Bologna.
FISICA E...

25vol25 / no1-2 / anno2009 >
della nascita della meccanica quantistica, costituì una sorta di banco di prova per le successive formulazioni teoriche [1].Più recentemente analoghe relazioni empiriche sono state dedotte per sistemi complessi, quali organismi biologici o sistemi ecologici. Come vedremo in seguito, la legge di Kleiber lega la potenza metabolica basale, ovvero quella necessaria per mantenere le funzioni vitali in condizioni di riposo, alla massa di una determinata specie, tramite la relazione [2]
(1)
dove M è la massa e B è una costante tassonomica, ovvero caratteristica di una determinata specie. La relazione di cui sopra ha una natura certamente fondamentale, essendo stata verificata su specie che differiscono in massa di 20 ordini di grandezza (batteri e virus inclusi) ma a tutt’oggi non si dispone di una interpretazione (univoca) in termini di principi primi.In questo articolo discuteremo alcune leggi di scala (note e meno note) relative a vari campi di ricerca, vedremo come esse siano legate da un fattore comune e ne analizzeremo le conseguenze in particolare per quanto riguarda il caso della biologia.Prima di entrare nello specifico, cercheremo di chiarire cosa si intenda con gli aggettivi complicato e semplice, se riferiti ad una legge che riproduca fatti relativi ad osservazioni di carattere empirico.È opportuno mettere in evidenza che una o un insieme di leggi possono avere carattere di principio come, ad esempio, le leggi del moto di Newton. In questo caso esse, all’interno dell’impianto teorico di una certa disciplina, giocano un ruolo non dissimile dai postulati in una teoria matematica e vengono accettate come i capisaldi intorno ai quali inquadrare tutti gli aspetti teorici.Leggi di natura non fondamentale sono quelle che descrivono un fenomeno nella propria interezza, ma che
possono essere derivate dai principii generali prima citati. La legge dell’isocronismo del pendolo è una di queste, ma la sua interpretazione necessita dell’analisi del moto oscillatorio in termini delle leggi di Newton, che sono inoltre in grado di specificarne i limiti di validità1.Nel seguito ci riferiremo al secondo tipo di “legge” ed in particolare a quelle che furono formulate prima che fosse sviluppato il complesso di principii e il formalismo matematico che, successivamente, ne ha permesso una spiegazione completa. In alcuni casi, tali leggi hanno costituito una sorta di punto di riferimento per gli sviluppi successivi.Il diagramma di flusso riportato in fig. 1 sintetizza quanto precedentemente discusso
Una serie di osservazioni, che si traducono in una serie di dati, caratterizzano un fenomeno (ϕαινοµενον= ciò che appare).I dati vengono organizzati sotto forma di “legge”, se questa è espressa in termini matematici si otterranno una o più relazioni tra le grandezze che la caratterizzano.È evidente che i fenomeni sono, per loro stessa definizione, quello che sono, pertanto non possono essere classificati come semplici o complicati, diversamente la “legge” ad essi riferita sarà tanto più semplice, quanto più semplici sono le strutture formali ad essa associate (minimo numero di identità, minimo numero di variabili coinvolte…).
Onde evitare errori di natura concettuale, è opportuno sottolineare che ci stiamo riferendo a questioni di carattere empirico e, quando diciamo legge, facciamo riferimento ad una sorta di riorganizzazione sintetica dei dati.Facendo di nuovo riferimento alla serie di Balmer notiamo che, quanto mostrato in fig. 2, è una delle serie spettrali
a)
b)
c)
Lunghezza d’onda (Å=10-10 m)
6562
.8Å
4861
.3Å
4340
.5Å
4101
.7Å
α β γ δ
1 La legge dell’isocronismo del pendolo vale per piccole oscillazioni; il corretto uso delle equazioni del moto derivate dal secondo principio della dinamica è in grado di inglobarla in una più generale legge oraria.
g. dattoli et al.: leggi di scala in biologia ecc.
Fig. 1 Flow chart per la formulazione di una legge empririca.
Fig. 2 Righe spettrali dell’atomo di idrogeno.

26 < il nUovo saggiatoRe
dell’atomo di idrogeno ed è quello di cui si disponeva intorno al 1885, quando J. J. Balmer, un insegnante2 con una insolita abilità nel trovare relazioni tra serie di numeri, mostrò che le lunghezze d’onda delle righe α, β, γ, and δ, delle serie spettrali mostrate in fi g. 2, possono essere riscritte come
e che pertanto possono essere sintetizzate nella semplice formula
(2)
Verso la fi ne del secolo XIX gli esperimenti condotti sulle proprietà spettrali della radiazione di corpo nero avevano portato Wien a formulare la seguente legge:L’energia irraggiata per unità di volume e per intervallo di frequenza e di energia è legata alla frequenza ν e alla temperatura assoluta T dalla relazione
(3)
In altri termini, a parte la dipendenza dal cubo della frequenza, la legge formulata da Wien asseriva che i fenomeni pertinenti alla fi sica della radiazione di corpo nero, venivano regolati da una funzione universale Φ, dipendente unicamente dal rapporto frequenza-temperatura assoluta.In un tentativo di trovare accordo tra la legge di Wien, in grado di riprodurre le proprietà spettrali del corpo nero alle
alte frequenze e quella di Jeans, appropriata per le basse frequenze3, Planck, usando una procedura di interpolazione trovò la seguente espressione:
(4)
dove c1 e c2 sono costanti di interpolazione; i dati sperimentali permisero di identifi carle come legate alle costanti di Boltzmann e a quella che poi diventò la costante di Planck.Le equazioni precedenti (2), (4) sono dunque due genuini esempi di ciò che abbiamo defi nito come leggi empiriche, ma che non rappresentano, dal punto di vista epistemologico, un grande passo in più rispetto alle osservazioni sperimentali. Non vogliamo con questa asserzione sminuire l’importanza della serie di Balmer, e della legge di corpo nero nella sua primitiva formulazione (entrambe hanno giocato un ruolo determinante nello sviluppo della moderna teoria quantistica) ciò che intendiamo dire è che esse non rappresentano lo schema teorico in cui i fenomeni possono essere inquadrati. Il passo successivo, ovvero la creazione di una teoria, ha richiesto un salto concettuale, secondo lo schema riportato in fi g. 3, in cui si fa riferimento a principi primi e ad una opportuna riformulazione in termini matematici. Nei prossimi paragrafi faremo una breve rassegna di leggi empiriche, discutendo in particolare le cosiddette leggi di scala, che emergono in ambito non solo fi sico, ma nei settori più svariati dalla planetologia alla ecologia. Vedremo come leggi di natura empirica, emergenti in biologia, possano determinare un impulso aff atto nuovo nello studio quantitativo di vari fenomeni di crescita ed in particolare per quello che concerne lo sviluppo delle masse tumorali.
2 Il fatto che Balmer fosse un insegnante di scuola media e dunque una persona senza una specializzazione in tecniche di spettroscopia è un elemento di non secondaria importanza, perché è rivelatore del fatto che è stato in grado di desumere la sua legge basandosi solo su evidenze di carattere numerico.
3 Si ricordi che nel caso di Wien , mentre nel caso di Jeans
.
FISICA E...
Fig. 3 Flow chart dalla legge empirica all’impianto teorico.

27vol25 / no1-2 / anno2009 >
maggiore di quantità fi siche e quindi di dimensioni da combinare. Prima di procedere oltre è il caso di ricordare un teorema fondamentale dell’analisi dimensionale, il cosiddetto teorema π di Buckingham, che non appare così noto come dovrebbe essere.Una formulazione, non proprio rigorosa, di detto teorema è la seguente [3]:Se un determinato fenomeno è descrivibile in termini di una equazione che contiene n variabili � siche, esprimibili in termini di k quantità � siche fondamentali, allora l’espressione originale è equivalente a n − k variabili adimensionali costruite a partire dalla relazione originale.Si potrebbe applicare il teorema precedente al problema dell’onda d’urto nelle esplosioni nucleari, notando che il processo può essere adeguatamente descritto da cinque quantità fi siche, il tempo trascorso dalla detonazione, il raggio dell’onda, la pressione atmosferica, la densità atmosferica e l’energia rilasciata in un preciso punto dello spazio al momento della detonazione. Le variabili precedenti dipendono da 3 costanti fondamentali (massa, lunghezza e tempo), per cui esisteranno 5 − 3 = 2 quantità adimensionali adatte a descrivere il problema. Non è diffi cile costruire tali variabili4, che vengono portate qui di seguito:
(10)
L’equazione che lega queste due quantità può essere scritta nella forma
(11)
La funzione g (π1) è nota da esperimenti su esplosivi convenzionali ed è molto vicina all’unità, per tempi prossimi all’istante dell’esplosione. Pertanto possiamo concludere che la relazione tra raggio ed energia rilasciata è proprio data dalla eq. (8) riportata in precedenza. Le cose non sono sempre così semplici perché le leggi di scala riguardano, come notato i fenomeni più disparati. Ad esempio, in sismologia la legge di Gutenberg-Richter stabilisce che, in una determinata zona, il numero di terremoti di magnitudo M per anno, è dato dalla relazione
(12)
con a, b costanti.Mentre in economia la cosiddetta curva di esperienza, ovvero l’abbattimento dei costi di produzione dopo la realizzazione
2 Le leggi di scalaTra le leggi empiriche quelle che assumono una forma particolarmente semplice sono le cosiddette “leggi di potenza”, costituite da relazioni del tipo [2]
(5)
dove a è una costante di proporzionalità e k è l’esponente della legge di potenza.Se le variabili, che caratterizzano una data legge di potenza, vengono riportate su un grafi co log-log il risultato è
(6)
cosicché i punti sperimentali si dispongono su una retta, di modo che l’esponente k può essere immediatamente defi nito dalla pendenza della retta.Una importante proprietà delle leggi di potenza è la cosiddetta invarianza di scala, esprimibile come segue:
(7)
secondo cui un incremento di scala della variabile produce semplicemente una variazione di scala della funzione medesima. Vedremo in seguito come tale proprietà, estremamente semplice, si traduca nel fenomeno di self-similarity, che gioca un ruolo determinante nella comprensione di fenomeni complessi come le transizioni di fase.Discuteremo nel seguito alcune di queste leggi e cercheremo di comprenderne il fi lo conduttore.Cominceremo pertanto con due esempi senza un apparente legame diretto.
L’equazione di Taylor che fornisce l’andamento temporale del raggio dell’onda di shock di una esplosione atomica in termini dell’energia sprigionata dall’esplosione stessa, ovvero
(8)
dove ρ è la densità del mezzo in cui l’onda d’urto dell’esplosione si propaga.
La legge dell’isocronismo del pendolo, secondo cui il periodo delle piccole oscillazioni è legato alla lunghezza l del pendolo e alla accelerazione di gravità, dalla relazione
(9) .
Ci si potrebbe ora chiedere cosa abbiano in comune le due fenomenologie, tenuto conto che la prima si riferisce ad un fenomeno che coinvolge un numero signifi cativamente
a)
b)
4 Buckingham indicò con π tali quantità non dimensionali, da cui il nome del teorema.
g. dattoli et al.: leggi di scala in biologia ecc.
28 < il nUovo saggiatoRe
dell’unità prototipale, è data da
(13)
dove n indica il numero delle unità prodotte e C1, C(n) sono i costi della prima e della n-esima unità rispettivamente.È il caso di sottolineare che ci sono stati tentativi di riconciliare le leggi di cui sopra, con il teorema di Buckingham, introducendo, ad hoc, nuove “unità fondamentali”, specifi che della disciplina cui la legge di scala fa riferimento. Nel caso di problemi economici, ad esempio, è risultato conveniente introdurre grandezze come quantità e costo (Q, $) come quantità fondamentali. Forse la prima legge di scala, ancora abbastanza controversa e che non ha trovato una sua completa spiegazione teorica, è la cosiddetta legge di Titius-Bode (formulata nel XVIII secolo), che fornisce la distanza dei pianeti dal sole secondo la seguente relazione5:
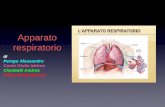
(14)
dove i parametri A, B sono dati da 44 e 1,73 rispettivamente e la misura è data in termini di raggi solari. In fi g. 4 abbiamo riportato il confronto tra le distanze predette dalla (14) e quelle osservate. Il confronto è decisamente soddisfacente.Torneremo sul problema di un approccio unifi cato alle leggi di scala, nel paragrafo conclusivo, dopo aver discusso la
log 1
0d
4.5
4.0
3.0
2.0
3.5
2.5
1.50 2 4 6 8 10
n
Den
sità
di p
opol
azio
ne (n
/km
2 )
1016
1014
1012
1010
108
106
104
102
100
10-6 10-4 10-2 100 102 104
Lunghezza (m)
Batteri
Ameba
Piccoli invertebrati
Grandiinvertebrati
Invertebrati,piccoliroditori e piccolimammiferi
GrandimammiferiUccelli, rettili, anfibi
5 La relazione (14) non è del tutto completa, l’espressione dovrebbe essere moltiplicata per una funzione φ(n) che tiene conto degli eff etti caotici, durante l’evoluzione primordiale del Sistema Solare, tale correzione pesa comunque in maniera non signifi cativa (meno del 10%).
rilevanza e le conseguenza di relazioni di tipo allometrico in biologia, ovvero una relazione tra due attributi di un sistema vivente.
3 Leggi di scala in ecologia e biologia e relazioni di carattere allometricoIn fi g. 5 abbiamo riportato un grafi co relativo all’andamento della densità della popolazione di varie specie in funzione della loro dimensione caratteristica. Poiché il grafi co viene riportato su scala logaritmica possiamo concludere, in base a quanto discusso in precedenza, che si tratta di una legge di scala, nota come legge di Damouth [4].La legge, la cui validità è stata provata su 8 ordini di grandezza, recita come segue:La densità di popolazione di una specie, all’interno del proprio ecosistema, segue una legge del tipo
(15)
dove δ rappresenta la densità e l le dimensioni caratteristiche della specie in questione. Come mostrato dalla fi g. 5 la validità della legge di scala si estende dai mammiferi ai batteri. Una relazione viene detta allometrica quando descrive la relazione tra due attributi di sistemi viventi; di solito queste relazioni vengono espresse come leggi di potenza, in cui il termine all’esponente viene detto coeffi ciente allometrico. Un esempio di relazione di tale tipo è quella che riguarda ad esempio la frequenza cardiaca dei mammiferi, che viene legata alla massa corporea dalla relazione6
(16) .
Fig. 4 Legge di Titius-Bode, distanze osservate (cerchi vuoti) e predette (cerchi pieni) dei pianeti dal Sole (n = 1 Mercurio, n = 2 Venere, n = 3 Terra, n = 4 Marte, n = 5 fascia degli asteroidi, n = 6 Saturno, n = 7 Giove, n = 8 Urano, n = 9 Nettuno, n = 10 Plutone).
Fig. 5 Densità di popolazione di varie specie in funzione della loro lunghezza.
FISICA E...
29vol25 / no1-2 / anno2009 >
log
(pot
enza
met
abol
ica)
log (massa)
5
0
-5
-10
-15
-20
-20 -15 -10 -5 0 5 10
Toporagno
Elefanti
Mammiferi
Numero medio delle celluledi mammifero in cultura
Mitocondrio (miocite di mammifero)Cellula a riposo
In vitro
CcORC
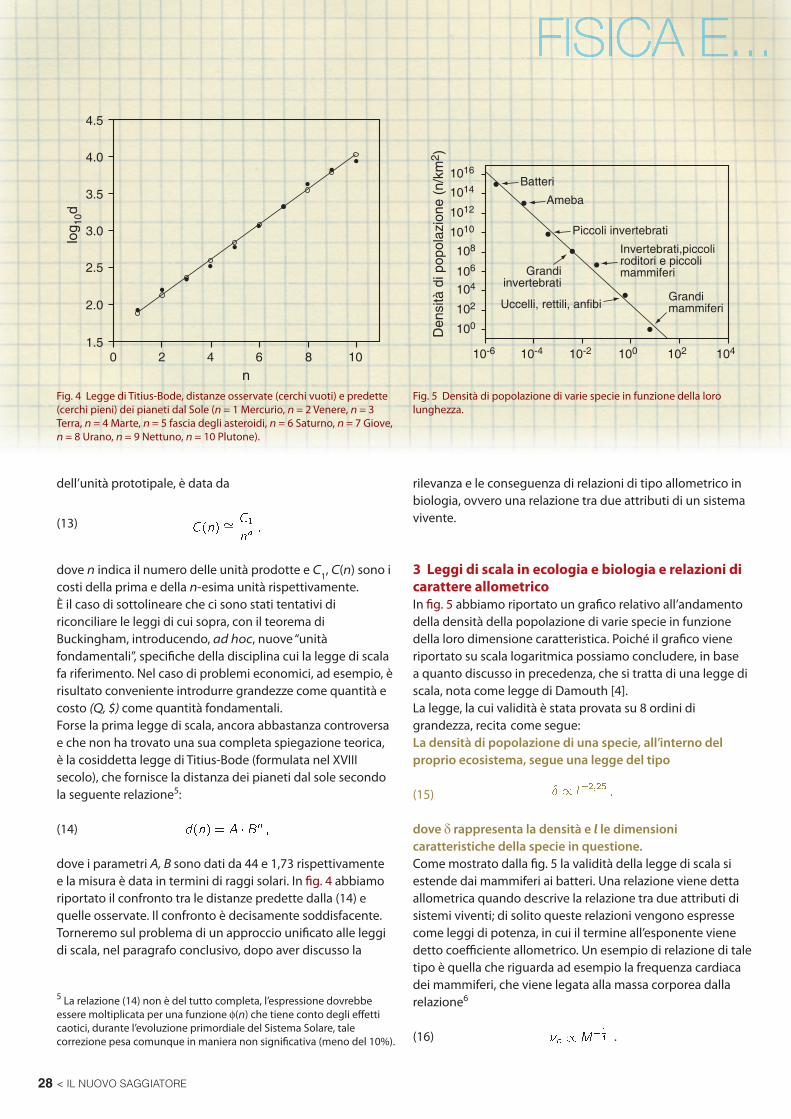
Analogamente la durata della vita è legata alla massa da
(17)
Dalle relazioni precedenti nasce immediatamente una considerazione relativa al tempo biologico, che non va misurato in anni, ma più propriamente in numero di battiti cardiaci che risulta essere una costante, come segue dal fatto che νcTL = k.La già citata legge di Kleiber mette in relazione la massa di una specie al suo consumo metabolico (fi g. 6).Prima di cercare di dare una giustifi cazione alla relazione riportata in eq. (1), cerchiamo di capire se esista un fi lo conduttore tra la leggi (1), (15), (16), (17). Supponiamo che la (1) sia vera. Poiché il volume è legato alle dimensioni da l = V 1/3 e poiché V ∝ M potremo riscrivere la (15) come
(18)
che potrebbe essere vista come una conseguenza della ipotesi di Kleiber in termini di competizione di risorse (si veda il paragrafo conclusivo).La dipendenza dalla massa della frequenza cardiaca può essere facilmente spiegata notando che essendo l’energia metabolica proporzionale alla massa, si ha quanto segue:
(19)
6 Si noti che tali relazioni valgono per specie e non per individuo appartenente ad una specie. All’interno di una stessa specie valgono relazioni diff erenti, nella specie umana gli individui con massa minore, in particolare di bassa statura, vivono, in media, un tempo maggiore di quelli di alta statura.
Un analogo ragionamento può essere utilizzato per derivare la relazione (17). Esiste dunque una sorta fi lo conduttore. Assunta come vera la (1) siamo riusciti ad ottenere, in maniera costruttiva le altre. Ovviamente avremmo potuto procedere partendo da una a caso delle quattro leggi.Come già detto le leggi di scala riguardano le cose più disparate, ad esempio la velocità ottimale di crociera per uccelli di massa M è data da
(20)
La relazione precedente, indipendente dalla legge di Kleiber, può essere derivata utilizzando solo argomenti di natura meccanica.Infatti la potenza necessaria ad un sistema di massa M, per volare ad una velocità V è data da
(21)
Il primo termine è il lavoro necessario per tenere la massa in quota nell’intervallo di tempo √h/g che è il tempo di caduta libera7, il secondo termine è quello di Bernouilli, ρa è la densità dell’aria e Σ è la superfi cie del corpo in volo8. L’energia dissipata per coprire una distanza L a velocità costante sarà data da
(22)
che una volta minimizzata rispetto alla velocità, fornisce l’espressione
(23)
Avendo assunto che M = ρbV, V 1/3 ≅ h, utilizzando valori tipici per la densità dell’aria e per la densità del corpo di un uccello, assunta essere comparabile a quella dell’aria, si ottiene la legge di scala riportata nell’eq. (20).È dunque evidente che, data la vastità delle problematiche coinvolte e il livello di validità di tali leggi, sarebbe opportuno tentare di inquadrarle in un contesto che vada al di là della semplice legge empirica. Sforzi in tal senso sono stati compiuti e si comincia a delineare un complesso di ragioni teoriche che potrebbero inquadrare tale eterogenea fenomenologia. Nel paragrafo conclusivo vedremo che direzione stanno prendendo tali ricerche.
7 Abbiamo pertanto ipotizzato che la frequenza di battito d’ala sia √g/h. 8 Si ricordi che ρaV 2 è una pressione, pertanto ρaV 2Σ è una forza, che moltiplicata per una velocità, dà una potenza. Dall’uguaglianza ρaV 2
mΣ = mg segue la minima velocità per il mantenimento della massa in volo che risulta essere legata alla velocità ottimale dalla relazione
.
g. dattoli et al.: leggi di scala in biologia ecc.
Fig. 6 Potenza metabolica in funzione della massa per diverse specie (scale logaritmiche).
30 < il nUovo saggiatoRe
Nel prossimo paragrafo assumeremo come vera la legge di Kleiber e vedremo come da questa possano essere dedotte una serie di leggi di crescita per quanto riguarda gli organismi di esseri viventi, incluse le masse cancerose.
4 Leggi di crescita ontogenetiche9
L’evoluzione temporale o ontogenetica della massa di un organismo è regolata da un sistema di equazioni che deve essere in grado di rendere conto di
una crescita di tipo esponenziale,un meccanismo che determini un eff etto di decelerazione del processo di crescita fi no alla completa saturazione.
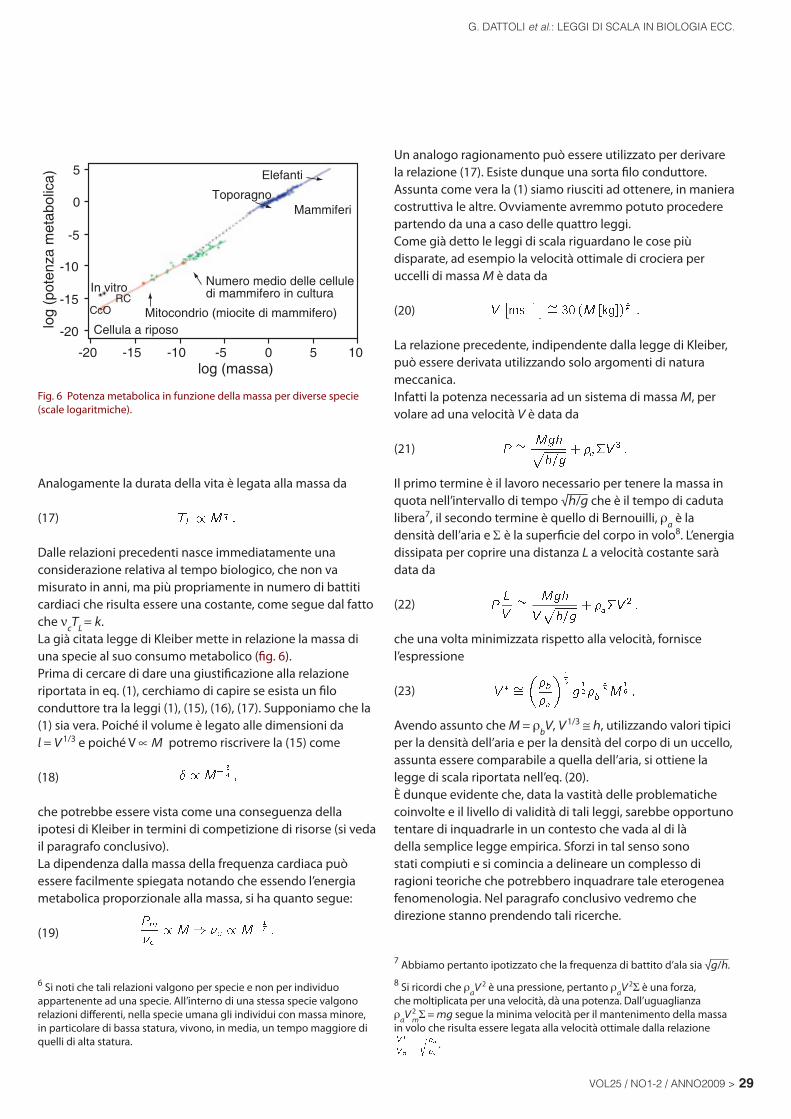
Quanto abbiamo descritto è comune ad una infi nità di processi fi sici, regolati da equazioni non lineari. Uno dei casi più semplici è quello relativo all’evoluzione di un sistema coerente come un oscillatore laser. In questo caso il campo elettromagnetico coerente cresce all’interno di una cavità ottica, seguendo una curva di tipo logistico non dissimile da quella di fi g. 7, caratterizzata da tre fasi
una fase iniziale in cui il sistema tende ad organizzarsi,una fase intermedia con crescita esponenziale,la saturazione, determinata da un eff etto di riduzione del guadagno, indotto dalla intensità del campo stesso.
Come è ben noto, l’equazione diff erenziale che regola
a)b)
i)ii)iii)
l’evoluzione di questo tipo è data da
(24)
nella quale la semplice non linearità quadratica riesce a tenere conto degli eff etti di saturazione del guadagno, la cui parte lineare è rappresentata dal coeffi ciente α. Non a caso Von Bartalanff y suggerì, verso la metà dello scorso secolo, che la crescita delle masse di un organismo vivente possa essere descritta dalla seguente famiglia di equazioni [5]:
(25)
I coeffi cienti che compaiono nell’equazione sono indeterminati perché l’eq. (25) è una ipotesi e non è derivata, come nel caso dei laser, da una teoria generale, e vengono derivati a posteriori come parametri di fi t. Assumendo come vera la legge di Kleiber possiamo ragionare come segue: la potenza P necessaria per determinare la crescita della massa di un organismo consta di due contributi, la potenza Pb necessaria per gestire l’intero complesso di cellule esistenti e quella necessaria per determinare la creazione di nuove cellule, dunque
(26)
dove Ec rappresenta l’energia necessaria per la creazione di una singola cellula di massa mc e N indica il numero totale di
9 Per ontogenesi si intende la storia evolutiva di un organismo dalla sua nascita.
FISICA E...
Mas
sa n
orm
aliz
zata
Tempo adimensionale
1.25
0.25
0.75
1.00
0.50
00 2 4 6 8 10
PigStrew
RabbitCodFlat
Guinea PigShrimpSalmonGuppy
HenRobinHeron
Cow
Fig. 7 Curva di crescita in termini dei parametri universali.

31vol25 / no1-2 / anno2009 >
cellule, costituente la massa totale m.Tenuto conto che con P si è indicata la potenza metabolica legata alla legge di Kleiber, si perviene dalla relazione di rate (26) alla seguente equazione di evoluzione:
(27) ,
che è un caso particolare della famiglia di Von Bartalanff y.L’equazione precedente fornisce immediatamente la massa di saturazione M, ovvero il valore per cui :
(28)
Mentre l’evoluzione nel tempo della massa sarà data da
(29)
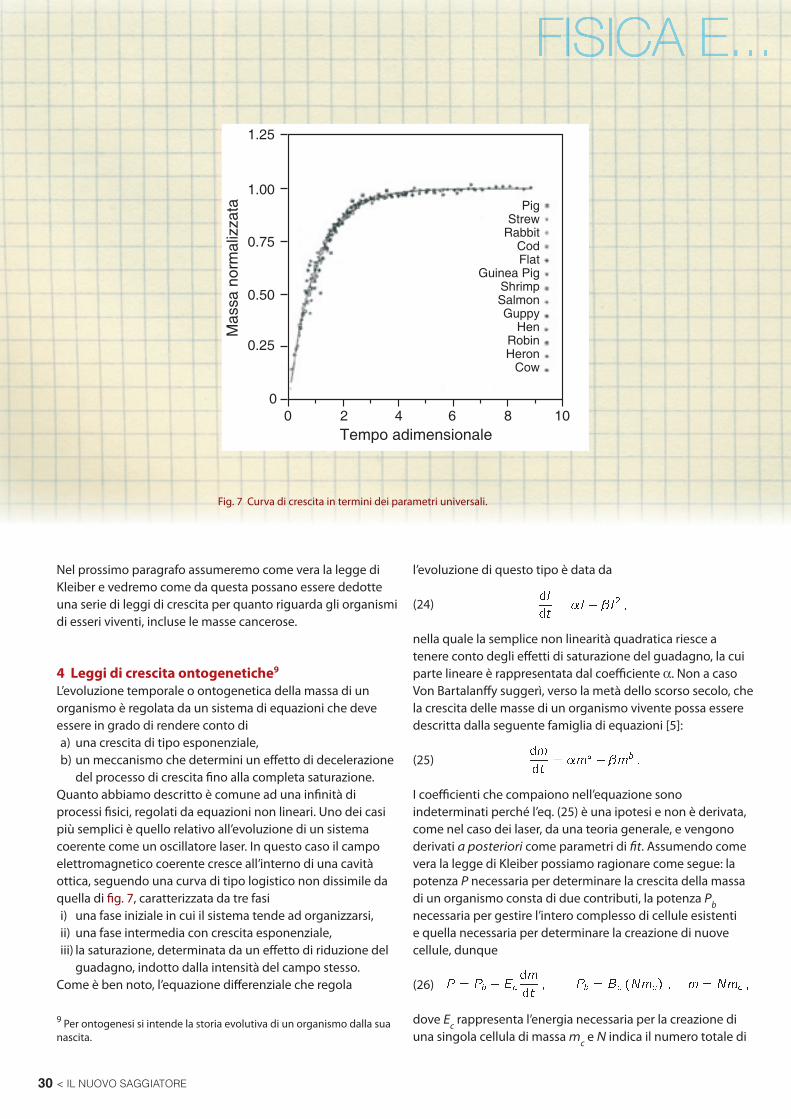
Nelle fi g. 7 e 8 riportiamo le curve di crescita per alcune specie animali, l’analisi statistica fornisce i seguenti dati per quanto riguarda i mammiferi:
(30) ,
qualora fosse noto il valore di Bc si potrebbe stimare il valore della massa asintotica, che viceversa può essere inferito dai dati sperimentali. È comunque interessante osservare, che esprimendo la curva in termini non dimensionali avremmo la curva universale, in termini di
.
g. dattoli et al.: leggi di scala in biologia ecc.
Fig. 8 Curve di crescita per alcuni mammiferi.
Mas
sa (g
)M
assa
(g)
1000
750
250
0
500
2500
2000
1000
500
0
1500
0 200 400 600 800
0 100 200 300 400
0.14
0.12
0.10
0.08
0.06
0.04
0.02
0
Guppy
MuccaGallina
Maiale della Guinea
0 20 40 60 80
Giorni Giorni
500000
400000
300000
200000
100000
00 1000 2000
32 < il nUovo saggiatoRe
1×1012
1×1010
1×108
1×106
1×104
100
1
T (giorni)
Num
ero
di c
ellu
le tu
mor
ali
0.01 1 100 10000
FISICA E...
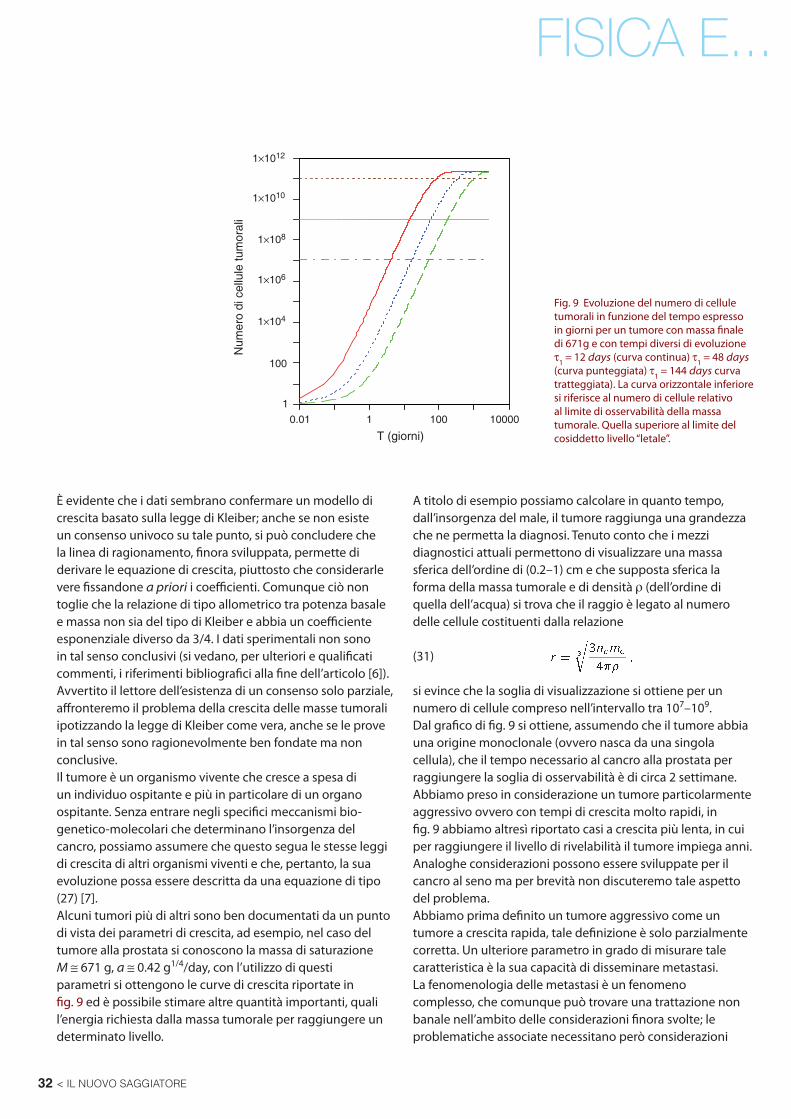
Fig. 9 Evoluzione del numero di cellule tumorali in funzione del tempo espresso in giorni per un tumore con massa fi nale di 671g e con tempi diversi di evoluzione τ1 = 12 days (curva continua) τ1 = 48 days (curva punteggiata) τ1 = 144 days curva tratteggiata). La curva orizzontale inferiore si riferisce al numero di cellule relativo al limite di osservabilità della massa tumorale. Quella superiore al limite del cosiddetto livello “letale”.
È evidente che i dati sembrano confermare un modello di crescita basato sulla legge di Kleiber; anche se non esiste un consenso univoco su tale punto, si può concludere che la linea di ragionamento, fi nora sviluppata, permette di derivare le equazione di crescita, piuttosto che considerarle vere fi ssandone a priori i coeffi cienti. Comunque ciò non toglie che la relazione di tipo allometrico tra potenza basale e massa non sia del tipo di Kleiber e abbia un coeffi ciente esponenziale diverso da 3/4. I dati sperimentali non sono in tal senso conclusivi (si vedano, per ulteriori e qualifi cati commenti, i riferimenti bibliografi ci alla fi ne dell’articolo [6]).Avvertito il lettore dell’esistenza di un consenso solo parziale, aff ronteremo il problema della crescita delle masse tumorali ipotizzando la legge di Kleiber come vera, anche se le prove in tal senso sono ragionevolmente ben fondate ma non conclusive.Il tumore è un organismo vivente che cresce a spesa di un individuo ospitante e più in particolare di un organo ospitante. Senza entrare negli specifi ci meccanismi bio-genetico-molecolari che determinano l’insorgenza del cancro, possiamo assumere che questo segua le stesse leggi di crescita di altri organismi viventi e che, pertanto, la sua evoluzione possa essere descritta da una equazione di tipo (27) [7].Alcuni tumori più di altri sono ben documentati da un punto di vista dei parametri di crescita, ad esempio, nel caso del tumore alla prostata si conoscono la massa di saturazione M ≅ 671 g, a ≅ 0.42 g1/4/day, con l’utilizzo di questi parametri si ottengono le curve di crescita riportate in fi g. 9 ed è possibile stimare altre quantità importanti, quali l’energia richiesta dalla massa tumorale per raggiungere un determinato livello.
A titolo di esempio possiamo calcolare in quanto tempo, dall’insorgenza del male, il tumore raggiunga una grandezza che ne permetta la diagnosi. Tenuto conto che i mezzi diagnostici attuali permettono di visualizzare una massa sferica dell’ordine di (0.2–1) cm e che supposta sferica la forma della massa tumorale e di densità ρ (dell’ordine di quella dell’acqua) si trova che il raggio è legato al numero delle cellule costituenti dalla relazione
(31)
si evince che la soglia di visualizzazione si ottiene per un numero di cellule compreso nell’intervallo tra 107–109.Dal grafi co di fi g. 9 si ottiene, assumendo che il tumore abbia una origine monoclonale (ovvero nasca da una singola cellula), che il tempo necessario al cancro alla prostata per raggiungere la soglia di osservabilità è di circa 2 settimane. Abbiamo preso in considerazione un tumore particolarmente aggressivo ovvero con tempi di crescita molto rapidi, in fi g. 9 abbiamo altresì riportato casi a crescita più lenta, in cui per raggiungere il livello di rivelabilità il tumore impiega anni.Analoghe considerazioni possono essere sviluppate per il cancro al seno ma per brevità non discuteremo tale aspetto del problema.Abbiamo prima defi nito un tumore aggressivo come un tumore a crescita rapida, tale defi nizione è solo parzialmente corretta. Un ulteriore parametro in grado di misurare tale caratteristica è la sua capacità di disseminare metastasi. La fenomenologia delle metastasi è un fenomeno complesso, che comunque può trovare una trattazione non banale nell’ambito delle considerazioni fi nora svolte; le problematiche associate necessitano però considerazioni

33vol25 / no1-2 / anno2009 >
di natura più specialistica e, pertanto, rimandiamo il lettore interessato alla letteratura riportata alla fine dell’articolo.
5 Considerazioni conclusiveAbbiamo visto nei paragrafi precedenti come leggi di natura empirica possano emergere da una opportuna riorganizzazione di dati “sperimentali” e come queste possano influenzare lo sviluppo di una determinata disciplina. Abbiamo altresì messo in evidenza che molte relazioni, note come leggi di potenza, stanno giocando un ruolo sempre più decisivo nell’ambito di scienze quali l’ecologia e la biologia. Per quanto riguarda quest’ultima si è potuto apprezzare il fatto che, assunta come vera l’ipotesi di Kleiber, sia possibile derivare l’equazione di rate che determina lo sviluppo di una data specie biologica, incluse le masse tumorali.Come abbiamo già fatto notare, non esiste un consenso univoco sul coefficiente allometrico 3/4 nella legge di Kleber, molti autori riportano stime diverse (si veda la parte conclusiva di questo paragrafo e la bibliografia alla fine dell’articolo) ma, a quanto pare, esiste un vasto accordo sul fatto che il metabolismo basale delle specie viventi possa essere riprodotto da una legge del tipo
(32)
È pertanto evidente che in tale caso le considerazioni generali fino ad ora svolte non cambiano da un punto di vista concettuale. L’equazione di evoluzione (27) dovrà essere modificata, tenendo conto della diversa dipendenza funzionale di B(M) e dovremo risolvere una equazione differenziale con un coefficiente esponenziale diverso10.I processi di evoluzione in biologia sono basati su meccanismi complessi, pertanto la possibilità di fare luce su tali problematiche utilizzando un apparato analitico estremamente modesto è certamente interessante. In merito all’evoluzione delle masse cancerose il modello cui abbiamo accennato in precedenza ha offerto, ad esempio, la possibilità di ipotizzare che alla base dei processi di metastatizzazione vi siano meccanismi di competizione energetici tra cancro e organo ospitante, la quantificazione di tale processo ha permesso di calcolare quantità critiche come la carrying capacity dell’organo ospitante, ovvero il livello di massa tumorale che l’organo può tollerare, prima che il tumore senta la necessità di migrare in cerca di zone energeticamente più appetibili.Vogliamo di nuovo sottolineare che la complessità dei
fenomeni non va sottovalutata e che quindi la validità di leggi di scala in biologia e in particolare la legge di Kleiber va considerata come approssimazioni di leggi più generali.Come è ben noto l’evoluzione biologica è fortemente influenzata da processi di natura chimica che sono a loro volta fortemente dipendenti dalla temperatura.Si è pertanto ipotizzato che la legge di Kleiber e l’effetto della temperatura possano essere combinate di modo che il metabolismo basale possa essere in generale espresso come [8]
(33)
dove il termine esponenziale contiene una sorta di energia di attivazione Ei e la temperatura assoluta T.L’accelerazione dei processi biologici al crescere della temperatura è infatti ben nota e pertanto possiamo ipotizzare che la durata media di vita sia data da una relazione del tipo
(34)
dove per E si utilizza un valore medio di 6 eV, tipico di reazioni biochimiche.Questo aspetto del problema necessiterebbe ulteriori approfondimenti che non possono essere affrontati in questa sede; vogliamo comunque far notare che
un eccesso calorico, determinato ad esempio dall’eccesso di cibo, determina un diminuzione della vita media, anche in assenza di patologie specifiche legate al cibo, la produzione di calore interno determinata da eccessivo esercizio fisico produce analoghi effetti negativi.
Gli effetti di cui sopra, associati ad un aumento dell’entropia interna, producono un invecchiamento precoce insieme a tutte le relative conseguenze.È dunque evidente che una corretta analisi del problema dell’evoluzione implica la soluzione dell’eq. (27) con l’inclusione degli effetti di dipendenza dalla temperatura e di una possibile variazione nel tempo della medesima.Per dare una idea dell’utilità delle considerazioni precedenti consideriamo la soluzione (29) a piccoli tempi per m0 = 0, in questo caso avremo
(35)
dove T0 è una temperatura di riferimento scelta uguale a quella del punto di congelamento dell’acqua, in corrispondenza della quale si ha un significativo rallentamento dei processi biologici, inoltre α (T0) = α (si veda la seconda delle equazioni (27)). La relazione (35) fornisce il legame tra masse embrionali, temperatura e tempo di
a)
b)
10 È il caso di notare che tutte le equazioni di forma sono equivalenti e possono essere risolte in termini analitici; il caso in cui entrambi i coefficienti a, b nell’eq. (25) sono coefficienti generici diversi da 1 presenta qualche difficoltà.
g. dattoli et al.: leggi di scala in biologia ecc.

34 < il nUovo saggiatoRe
È evidente che tale quantità varia da specie a specie e in particolare, per quanto riguarda la specie umana, dipende anche da fattori di tipo culturale, tecniche di coltura più effi cienti forniscono, ad esempio, valori di k più elevati.È forse il caso di sottolineare che nel derivare l’eq. (36) abbiamo fatto implicitamente l’assunzione che ogni singolo individuo sfrutti il territorio in maniera incoerente dagli altri, abbiamo infatti semplicemente moltiplicato il numero di costituenti la popolazione per il metabolismo di base del singolo individuo. Diversamente si sarebbe potuto assumere una sorta di eff etto coerente, defi nendo una massa totale della popolazione, secondo la relazione NcM, in modo da avere
(38)
Ovvero
(39)
dove N—
è dato dall’eq. (37).È dunque evidente che, se organizzata secondo meccanismi defi niti coerenti, una determinata popolazione potrà allocare, a parità di risorse, un maggiore numero di individui sulla stessa superfi cie, in modo da violare la legge di Damouth.Un argomento di questo tipo può essere adottato per tipi particolari di popolazioni, come quelli cellulari o per specie, quali gli imenotteri, per i quali il ruolo del formicaio o dell’alveare è dominante rispetto a quello del singolo individuo. Argomenti basati sulla carrying capacity sono estremamente
rK
LKπrK
2LK
FISICA E...ln
(t/m
1/4 )
Tc/(1+(Tc/273))
6
3
00 20 40
ectodermi acquaticiUccelli
Fig. 10 Dipendenza dalla temperatura del tempo di evoluzione degli embrioni di pesci e di uccelli.
evoluzione, confermato dall’analisi dei dati sperimentali, si vedano i riferimenti bibliografi ci e la fi g. 10 dove vengono riportati i dati relativi all’evoluzione degli embrioni di uccelli.È evidente che un decremento della temperatura determina un aumento del tempo di crescita, fatto che certamente potrebbe avere conseguenze per la crescita di masse tumorali. Questo aspetto necessita però di ulteriori approfondimenti e non può essere trattato in questo ambito.Torniamo ora alla legge di Damouth, che deriveremo utilizzando il seguente argomento. Un determinato territorio potrà essere caratterizzato in base alla sua capacità di sostentamento, che è un termine vago per defi nire la quantità di calorie per unità di superfi cie che una determinata specie può trarre dal territorio. Indicando con C
— tale densità
calorica, assumendo come vera la legge di Kleiber, potremo legare la superfi cie di territorio e il numero di individui che la occupano secondo la relazione
(36)
da cui segue che la densità di popolazione è data da
(37)
Poiché M ∝ L3 ricaviamo dalla relazione precedente la legge di Damouth.Vogliamo però attirare l’attenzione sul coeffi ciente k, che gioca un ruolo importante e che potrebbe essere inteso come una sorta di carrying capacity di un determinato territorio.
Fig. 11 Esempio di creazione di struttura frattale auto-replicante da parte dei vasi capillari.

35vol25 / no1-2 / anno2009 >
utili per studiare effetti di competizione energetica tra una massa tumorale e l’organo ospitante, in modo da determinare le condizioni per l’insorgenza delle metastasi.In questo lavoro abbiamo toccato varie problematiche cercando di cogliere un filo conduttore tra varie leggi di scala che rendono conto di svariati effetti, noi ne abbiamo toccato solo una parte cercando di spiegare come, assunta vera una di queste, si possano giustificare le altre.La legge di Kleiber, lo ribadiamo, gode di un largo credito, ma osservazioni autorevoli tendono a mettere in dubbio non l’esistenza di una legge di scala, quanto il coefficiente allometrico, che avrebbe una giustificazione più semplice se, come alcuni ritengono, fosse 2/3. Il calore irraggiato è infatti proporzionale alla superficie corporea e pertanto proporzionale a M 2/3, tale spiegazione ha certamente un suo fondamento, ma trascura (come notato da West, Brown e Enquist) effetti legati al trasporto interno e alla struttura geometrica frattale della distribuzione dei vasi capillari (si veda la fig. 11). La dimensione frattale aumenta le dimensioni geometriche ordinarie di una unità, pertanto assumendo che il calore sia emesso tramite una ipersuperficie e che la massa sia legata ad un ipervolume ritroviamo la legge di Kleiber tramite la sostituzione M 2/3 → M 3/4.Rimane però un fatto misterioso ed affascinante: se leggi di tale tipo sono vere e se il coefficiente allometrico è in grado di giustificare il metabolismo basale di specie che differiscono per oltre 20 ordini di grandezza, significa che siamo di fronte ad una legge assolutamente fondamentale, che probabilmente esula dall’ambito semplicemente biologico11. Il lavoro teorico di West, Brown e Enquist fornisce certamente spunti interessanti, l’estensione dei metodi di crescita ontogenetica alle masse tumorali potrebbe avere conseguenze importanti di natura pratica. In questo articolo abbiamo toccato alcuni aspetti di un campo vasto ed interessante, forse non noto nell’ambito della comunità dei fisici come dovrebbe essere. Ci auguriamo, pertanto, di aver suscitato un interesse, che possa determinare letture più approfondite.
RingraziamentiGli autori esprimono la loro sincera riconoscenza ai Professori P. P. Del Santo, C. Guiot e al Dr. T. S. Deisboeck per numerose ed illuminanti discussioni sulla teoria della crescita delle masse tumorali.
11 Nella ricerca di una causa comune alle leggi allometriche di tipo Kleiber, ricerche recenti hanno utilizzato metodi basati sulla teoria dell’informazione di Fisher. I risultati ottenuti suggeriscono che i sistemi biologici, che soddisfano relazioni con coefficienti allometrici di tipo n/4 (n = 0, ± 1 ,± 2,…) sono controllati da due sole sorgenti di informazione. Si veda [9].
g. dattoli et al.: leggi di scala in biologia ecc.
Giuseppe Dattoli Laureato in Fisica presso l’Università La Sapienza di Roma nel 1976, si è occupato di fisica delle alte energie, di acceleratori e di Laser ad elettroni liberi, che è attualmente il suo principale campo di interesse. In tale campo ha svolto numerose ricerche, che gli hanno valso numerosi riconoscimenti internazionali e in particolare ha ricevuto, presso l’Università di Stanford, nel 1994 il premio annuale istituito per ricerche fondamentali nell’ambito della fisica dei Laser ad elettroni liberi.Attualmente è il Direttore dell’Unità di Fisica Teorica e Matematica Applicata presso il Centro ENEA di Frascati, dove abitualmente svolge la propria attività di ricerca.Ha insegnato presso istituzioni accademiche nazionali ed estere e attualmente svolge il Corso di Metodi Matematici della Fisica presso l’Università di Roma Tre.
Bibliografia
[1] Un interessante e penetrante resoconto del ruolo giocato dale leggi empiriche nello sviluppo della meccanica quantistica è offerto da: P. Stesele, “Order, Chaos, Order: The transitions from Classical to Quantum Physics“ (Oxford University Press) 1994. Per una lettura più approfondita e dettagliata si consiglia l’opera di
J. Mehera e H. Rechenberg, “The Historical Development of Quantum Theory” (Springer-Verlag, New York) 1982.
[2] M. Kleiber, “Body Size and metabolism”, Hilgardia, 6 (1932) 315. [3] Una lettura appropriata per una esposizione rigorosa del Teorema
di Buckingam è George W. Hart, “Multidimensional Analysis: Algebras and Systems for Science and Engineering” (Springer-Verlag) 1995.
[4] Per una rassegna recente sulle leggi di scala in biologia e ecologia si veda: P. Hunter, “The Power of the Power Laws”, The Scientist, 17 (2003) 22; P. A. Parquet et al., “Scaling and Power Laws in Ecological Systems”, J. Exp. Biol., 208 (2005) 1749.
[5] Si vedano inoltre gli articoli originali: L. Von Bertalanffy, “Quantitative laws in metabolism and growth”, Q. Rev. Biol., 32 (1957) 217; G. B. West, J. H. Brown e J. B. Enquist, “A General Model for the Origin of Allometric Scaling Laws in Biology”, Science, 276 (1997) 122; G. B. West, J. H. Brown e J. B. Enquist, “Allometric scaling of metabolic rate from molecules and mitochondria to cells and mammals”, Proc. Natl. Acad. Sci. U.S.A., 99, Suppl. 1 (1999) 2473.
[6] Per le critiche alla legge di Kleiber si consultino gli articoli: P. S. Dodds et al., “Re-examination of the 3/4-law of Metabolism”, J. Theor. Biol, 209 (2001) 9; J. R. Banavar, J. Damouth, A. Maritan e
A. Rinaldo, “Modelling Universality and Scaling”, Nature, 420 (2002) 626.
[7] L’utilizzo della legge di Kleiber per lo studio dell’evoluzione delle masse tumorali è stato suggerito in: C. Guiot, P. Degiorgis,
P. P. Delsanto, P. Gabriele e T. Delsboeck, “Does Tumour Growth Follow a Universal Scaling Law?”, J. Theor. Biol., 225 (2003) 147; C. Guiot, P. P. Delsanto, A. Carpentieri, Y. Mansary e T. S. Delsboeck, “The Dynamic Evolution of Power exponent in Universal Growth Model of Tumours”, J. Theor. Biol., 240 (2006) 459; G. Dattoli,
P. L. Ottaviani, e S. Pagnutti, “Ontogenetic tumor growth and evolution equations”, J. Appl. Phys., 104 (2008) 024701.
[8] Per quanto concerne effetti di variazione di temperature nello sviluppo di masse biologiche si consulti: J. M. Gilloby,
E. L. Charnov, G. B. West, V. M. Savage e J. H. Brown, “Effects of size and temperature on development time”, Nature, 417 (2002) 70, e i riferimenti in essa contenuti.
[9] B. R. Friedmann e R. A. Gateaby, “Power Laws systems from extreme physical information”, Phys. Rev. E, 72 (2005) 036101.