Le curve di legame per l’ ossigeno della Mioglobina e della Emoglobina
16
1 Le curve di legame per l’ ossigeno della Mioglobina e della Emoglobina . Sergio Barocci La matematica viene da sempre considerata un valido strumento specialmente nel campo della chimica, della fisica e della biologia allo scopo di quantificare e razionalizzare ipotesi formulate sulla base di osservazioni sperimentali. In questo modo, fenomeni reali presentati attraverso funzioni, equazioni, logaritmi ecc., relazionati sulla base di nozioni chimiche, fisiche o biologiche , possono essere presentati con utili modelli matematici. Nel ventesimo secolo , la maggior parte delle connessioni o delle interazioni tra la matematica e la chimica ma in particolar modo tra la matematica e la biochimica sono state affrontate da molti ricercatori per risolvere alcuni concetti in maniera tale che determinate interpretazioni e/o la soluzione di processi biologici potesse avvenire con configurazioni , strutture e/o modelli. Alcuni esempi di queste interazioni sono rappresentate dallo studio della combinazione reversibile tra proteine (P) (enzima, recettore, trasportatore) e ligandi (L) (substrato, ormone, molecole) come base della funzione di molte proteine; interazioni in genere governate da un equilibrio chimico mediante la legge dell’azione di massa : P + L P L [P-L] [P] [L] Ka = ------------ Kd = ----------- [P] [L] [P-L] Ka = costante di associazione e Kd = costante di dissociazione dove Kd = 1/ Ka Significato della Kd di un complesso binario [P] [L] Kd = ---------- [P-L]
-
Upload
gianluigi-filippelli -
Category
Documents
-
view
218 -
download
0
description
Articolo divulgativo di Sergio Barocci che utilizzando il legame tra chimica e matematica racconta del trasporto dell'ossigeno, sulla mioglobina e l'emoglobina
Transcript of Le curve di legame per l’ ossigeno della Mioglobina e della Emoglobina

1
Le curve di legame per l’ ossigeno della Mioglobina e
della Emoglobina .
Sergio Barocci
La matematica viene da sempre considerata un valido strumento specialmente nel campo della
chimica, della fisica e della biologia allo scopo di quantificare e razionalizzare ipotesi formulate
sulla base di osservazioni sperimentali. In questo modo, fenomeni reali presentati attraverso
funzioni, equazioni, logaritmi ecc., relazionati sulla base di nozioni chimiche, fisiche o
biologiche , possono essere presentati con utili modelli matematici.
Nel ventesimo secolo , la maggior parte delle connessioni o delle interazioni tra la matematica e la
chimica ma in particolar modo tra la matematica e la biochimica sono state affrontate da molt i
ricercatori per risolvere alcuni concetti in maniera tale che determinate interpretazioni e/o la
soluzione di processi biologici potesse avvenire con configurazioni , strutture e/o modelli.
Alcuni esempi di queste interazioni sono rappresentate dallo studio della combinazione reversibile
tra proteine (P) (enzima, recettore, trasportatore) e ligandi (L) (substrato, ormone, molecole) come
base della funzione di molte proteine; interazioni in genere governate da un equilibrio chimico
mediante la legge dell’azione di massa :
P + L P L
[P-L] [P] [L]
Ka = ------------ Kd = -----------
[P] [L] [P-L]
Ka = costante di associazione e Kd = costante di dissociazione
dove Kd = 1/ Ka
Significato della Kd di un complesso binario
[P] [L]
Kd = ----------
[P-L]

2
Da questa relazione si possono dedurre le seguenti definizioni:
[Ptot] = [P] + [P-L]
[Ltot] = [L] + [P-L]
L’equazione P + L P L può essere anche riscritta nella forma:
[P-L] / [P] = [L] / K
Combinando l’equazione [Ptot] = [P] + [P-L] con l’equazione [P-L] / [P] = [L] / K si ricava :
[P-L] [L]
Y = ----------------- = ------------------
[Ptot] Kd + [L]
Y= frazione di saturazione in funzione della concentrazione di ligando libero
Kd = “concentrazione di semisaturazione”di L libero.
La frazione di saturazione Y di P (frazione di P complessata ) varia al variare di [L], per effetto
della legge dell’azione di massa. La rappresentazione grafica dell’ultima equazione è un’iperbole
equilatera che presenta [L] in ascissa, la frazione di proteina combinata () in ordinata, l’inizio
nell’origine degli assi e asintoto = 1. Più la Kd è bassa, più l’interazione proteina-ligando forte
La stechiometria del complesso tra proteina e legante è molto variabile nelle proteine come si può
individuare nelle reazioni della mioglobina (Mb) e dell’emoglobina (Hb) con le molecole di
ossigeno:
Mb + O2 <=> MbO2
Hb + 4 O2 <=> HbO2 + 3 O2 <=> Hb(O2)2 + 2 O2 <=> Hb(O2)3 + O <=> Hb(O2)4
Le molecole che trasportano l’ossigeno
L’ossigeno riveste un ruolo fondamentale nei meccanismi chimici che sottostanno alla vita
aerobica. Infatti, il passaggio dalla vita anaerobica alla vita aerobica rappresenta una delle tappe
più importanti dell’evoluzione, perché ha reso disponibile una grande riserva di energia. L’energia
che si estrae dal glucosio è circa diciotto volte più elevata in presenza di ossigeno che in sua
assenza.
( CH2O6)6 + 6O2 6CO2 + 6H2O + Energia
L’evoluzione ha quindi dovuto affinare nei vertebrati dei meccanismi in grado di rifornire
costantemente le cellule di ossigeno o O2 e al tempo stesso di liberarle dalla anidride carbonica o
CO2. Uno di questi meccanismi è rappresentato dal sistema circolatorio che porta di continuo
ossigeno alle cellule e l’altro è invece da alcune molecole che trasportano l’ossigeno permettendo
così di superare la limitazione imposta dalla bassa solubilità dell’O2 in acqua (circa 10-4
M). Questi
trasportatori di ossigeno nei vertebrati sono rappresentati da due proteine globulari : l’emoglobina e
la mioglobina. Tali proteine possono essere in soluzione (in alcuni invertebrati) oppure

3
concentrate in cellule specializzate come gli eritrociti dei vertebrati o ancora in cellule che
richiedono grossi apporti di O2 come le cellule muscolari . Nei vertebrati la proteina trasportatrice
di O2 nel sangue è l’emoglobina che gioca anche un ruolo importante nel trasporto di CO2 e di
ioni H+
(protoni) . Il suo legame con l ’O2 è regolato oltre che da H
+ e CO2 anche da fosfato
inorganico. Questi regolatori sono capaci di modificare profondamente le proprietà di legame
dell’O2 come l’emoglobina che può occupare anche siti disposti nella proteina in posizioni
lontane da quelle specifiche per l’O2. Queste interazioni tra siti diversi sono presenti e operanti in
molte proteine prendono il nome di interazioni allosteriche. L’emoglobina viene , quindi, definita
per queste proprietà, una proteina allosterica
La mioglobina o Mb è invece localizzata nei muscoli e la sua funzione è quella di fungere da
riserva di O2 (forma di immagazzinamento) e di facilitare il movimento di O2 nel muscolo.Nei
muscoli dei mammiferi acquatici la concentrazione di Mb è ~10 volte superiore a quella dei
mammiferi terrestri.
Mioglobina (Mb) e Emoglobina (Hb) costituiscono i più importanti membri della superfamiglia
delle globine , proteine tra loro omologhe (e strutturalmente simili). Sono state anche tra le prime
proteine di cui è stata definita anche la struttura tridimensionale.
Ma quali requisiti devono avere queste molecole trasportatrici di O2 ?
1. una sostanza deputata al trasporto di O2 deve:
a) legarlo e rilasciarlo in modo opportuno
b) impedire che reagisca con altre sostanze
Le proteine non hanno in genere gruppi funzionali adatti a fungere da siti di legame per l’O2
Alcuni ioni di metalli di transizione nei loro stati di ossidazione più bassi (Cu+, Fe
++ ) legano
fortemente l’O2 ma sono soggetti a venirne ossidati. Le proteine trasportatrici di ossigeno sono
proteine coniugate contenenti Fe eminico (gruppo eme) laddove lo ione ferroso dell’eme (Fe++
)
lega l’O2 mentre la componente proteica crea un ambiente idrofobico in cui il Fe++
non può essere
ossidato a Fe+++.
Mioglobina (Mb) e Emoglobina (Hb) rappresentano i più importanti membri
della superfamiglia delle globine, proteine tra loro omologhe (e strutturalmente simili). Sono state
anche tra le prime proteine da cui è stata definita anche la struttura tridimensionale (Fig 1 e Fig 2)

4
Figura 1 . Struttura tridimensionale della mioglobina
I chiarimenti sulla struttura tridimensionale rispettivamente della mioglobina e dell’emoglobina
sono dovuti a J. Kendrew e a M. Perutz (Fig.3)
L’emoglobina e la mioglobina si differenziano significativamente per le loro funzioni
La mioglobina o Mb è una proteina monomerica capace di legare o liberare O2 a seconda della
sua concentrazione nel citoplasma delle cellule muscolari; essa presenta una catena polipeptidica
costituita da 153 aminoacidi e un sito di legame per l’ossigeno al contrario dell’emoglobina che è
invece costituita da quattro catene globiniche ognuna contenente un sito di legame per l’O2 ed a
due a due uguali. La struttura della Mioglobina fu studiata ai raggi X dal muscolo scheletrico di
capodoglio in quanto particolarmente stabile per la formazione di ottimi cristalli. Il muscolo
scheletrico di mammiferi marini come balene, delfini e foche è infatti ricco in mioglobina per la
sua funzione di riserva di ossigeno durante le immersioni.
Figura 2. Struttura tridimensionale dell’emoglobina
John Kendrew ( 1917 – 1997) Max Perutz (1914 – 2002)
Figura 3 John Kendrew biochimico e cristallografo inglese, insignito del Premio Nobel per la
chimica nel 1962 insieme a Max Perutz biologo austriaco naturalizzato britannico,

5
rispettivamente per i loro studi effettuati sulla struttura delle proteine globulari quali la
mioglobina e l’emoglobina per mezzo dell'utilizzo di tecniche di diffrazione dei raggi X.
Analisi dell’emoglobina ai raggi X
La delucidazione della struttura tridimensionale dell’emoglobina o Hb ad opera di Perutz
cominciata nel 1936 viene considerata un lavoro monumentale : una fatica appassionante durata
quasi venticinque anni. Il successo arrivò nel 1959 quando Perutz ottenne una immagine della
densità ottica a bassa risoluzione dell’emoglobina di cavallo seguita alcuni anni dopo da mappe ad
alta risoluzione dei due stati due stati quaternari dell’emoglobina umana ed equina (Ossi -
Emoglobina e Deossi - Emoglobina). La molecola dell’Hb è quasi sferica. Le quattro catene sono
molto ravvicinate e disposte in forma tetraedrica. I gruppi eme (uno per subunità) sono localizzati in
cavità vicine alla superficie della molecola. I quattro siti di legame per l’ossigeno sono abbastanza
lontani tra di loro. La distanza tra i due atomi di Ferro più vicini è di 25 °A. Ogni catena è in
contatto con entrambe le catene mentre esistono poche interazioni tra le due catene o tra le due
catene . La struttura tridimensionale delle subunità dell’Hb assomiglia a quella della mioglobina.
Risulta quindi evidente che la struttura tridimensionale della Mb di capodoglio e delle catene e
dell’Hb umana racchiude un grande significato biologico. Tale struttura denominata “
ripiegamento globinico” ( le otto eliche in ciacuna catena dell’Hb sono sovrapponibili a quelle
della Mb) è comune a tutte le mioglobine ed emoglobine conosciute dei vertebrati. Il ripiegamento è
un disegno fondamentale della natura per un trasportatore di ossigeno che consente di porre il
gruppo eme in un ambiente che permette il legame reversibile dell’ossigeno. Gli aminoacidi
all’interno dell’Hb variano considerevolmente e spesso un residuo non polare viene sostituito da un
altro residuo non polare (alanina che sostituisce isoleucina). In questa maniera , il carattere non
polare dell’interno della molecola non viene alterato. L’insieme di questi residui non polari è molto
importante per legare il gruppo eme e anche per stabilizzare la struttura tridimensionale di ciascuna
subunità. Invece, i residui aminoacidici localizzati sulla superficie della molecola sono molto
variabili e alcuni hanno cariche nette positive o negative.
L’ HbA1 è la forma maggiormente rappresentata di emoglobina nell’uomo (circa il 98%) ed è
costituita da due catene globiniche α, di 141 residui aminoacidici ciascuno e due β costituite da 146
residui (α2β2). E’ anche presente l’HbA2 (circa il 3%) in cui le due catene β sono sostituite da due
catene δ (α2δ2). Sia le quattro catene globiniche dell’emoglobina che quelle della mioglobina
contengono un gruppo prostetico in grado di legare una molecola di O2 (gruppo Eme).
Il gruppo eme
La capacità dell’emoglobina e della mioglobina di legare l’ossigeno dipende infatti, come già
accennato in precedenza, dalla presenza di una struttura non polipeptidica chiamata gruppo eme
(Fig 4) . Queste unità non polipeptidiche saldamente legate alle proteine vengono dette anche
gruppi prostetici. Il gruppo eme è costituito da una parte organica e da un atomo di ferro. La parte
organica prende il nome di protoporfirina IX ( è una porfirina) ed è costituita da quattro anelli
pirrolici legati tra loro da ponti metilenici per formare un anello tetrapirrolico. All’anello
tetrapirrolico sono legati quattro gruppi metilici, due vinilici e due propionici. Nel gruppo eme,
l’atomo di ferro è centrale e presenta un numero di ossidazione 2+ . Esso è in grado di formare 5 o

6
6 legami di coordinazione. Quattro legami di coordinazione sono formati con i quattro atomi di
azoto degli anelli pirrolici della protoporfirina e giacciono sullo stesso piano degli anelli pirrolici
quando l’Hb è ossigenata, mentre quando l’O2 è assente il ferro si trova ad una distanza dal piano di
0.6 Å. Il quinto legame di coordinazione lo ione Fe++
lo forma con un atomo di azoto presente
nell’anello imidazolico di un residuo di istidina (istidina prossimale o His F8) della catena
polipeptidica. Il sesto legame può essere libero o impegnare il ferro con la molecola di ossigeno.
Questi due ultimi legami sono perpendicolari al piano dell’anello tetrapirrolico dell’eme. Quando
l’ossigeno è legato allo ione Fe++
si verrà a trovare in prossimità dell’anello imidazolico di un
secondo residuo di istidina della catena polipeptidica (istidina distale o His E7).
Figura 4. Struttura del gruppo eme
Quando l’eme non è legato a proteine, la reazione dell’ossigeno con uno dei due siti di
coordinazione del ferro (perpendicolari al piano dell’anello porfirinico) genera l’ossidazione
irreversibile del Fe++
a Fe+++
. Quando l’eme è inserito in una proteina, questa reazione non avviene
in quanto il gruppo eme è immerso in profondità nella struttura proteica e l’accessibilità ai siti di
coordinazione è limitata. L’importanza dell’impedimento sterico presentato dall’ambiente in cui si
trova il sito di legame dell’eme, sia nell’emoglobina che nella mioglobina, è dimostrata dall’affinità
di queste due proteine per il monossido di carbonio o CO , che compete con l’ossigeno per tale sito.
In soluzione acquosa l’affinità del CO per l’eme libero è circa 25.000 volte maggiore di quella
dell’ossigeno, mentre l’affinità del CO nei confronti dell’eme legato alla proteina è solo 250 volte
superiore a quella dell’ossigeno. Questo a causa dell’impedimento sterico determinato dall’istidina
E7 (His E7) , che causa un’inclinazione nella conformazione del complesso globina-CO con
conseguente riduzione dell’affinità di legame.
Anche il monossido di azoto o NO, può coordinarsi al ferro dell’eme con una affinità in questo
caso superiore anche a quella dell’ossigeno. Quando si lega l’ossigeno, le proprietà elettroniche del
ferro si modificano. Questo spiega il diverso colore che ha il sangue venoso povero di ossigeno
(rosso scuro) rispetto al sangue arterioso ricco di ossigeno (rosso brillante).
Il gruppo eme si trova alloggiato all’interno di una tasca idrofobica formata dal ripiegamento di
ciascuna catena polipeptidica all’interno del quale le catene laterali di circa 18 aminoacidi
stabiliscono all’incirca una ottantina di interazioni non covalenti con gli atomi di carbonio

7
dell’eme. Le due catene di acido propionico sono invece stabilizzate dalla interazione con la
catena laterale priva di carica di un residuo di istidina e con molecole d’acqua, entrambe presenti
sulla superficie esterna della molecola. La diffrazione ai raggi X ha mostrato che le catene
globiniche dell’emoglobina cosi come quelle della mioglobina si presentano costituite da numerose
regioni ad α-elica interrotte per la presenza di residui di prolina, che permettono il ripiegamento
della catena a cui si accompagna spesso una inversione della direzione della stessa catena. Questo
tipo di andamento fa assumere a queste catene globiniche una struttura grossolanamente sferoidale
(proteine globulari) .
Le catene sono caratterizzate da sette zone ad α-elica, mentre le β da otto. Esiste una notevole
somiglianza tra la struttura della catena globinica della mioglobina e la subunità β
dell’emoglobina. Queste si differenziano solo per la disposizione del residuo carbossilico e
amminico terminale. Il posizionamento verso l’esterno di tali gruppi, nella catena β
dell’emoglobina permetterà l’interazione tra le due catene beta. La funzione fisiologica della
mioglobina e della emoglobina dipende dalla reversibilità del legame fra ossigeno e gruppo eme,
processo il cui andamento è descritto dalle curve di saturazione con l’O2 .
La cinetica di legame O2 per Mb
La solubilizzazione dei gas nei liquidi è governata dalla legge di Henry (Fig.5) : un gas che esercita
una pressione sulla superficie di un liquido, può entrare in soluzione finché non avrà raggiunto in
quel liquido la stessa pressione che esercita sopra di esso. A temperatura costante, la solubilità di
un gas è direttamente proporzionale alla pressione che il gas esercita sulla soluzione. Quando viene
raggiunto l'equilibrio, il liquido si definisce saturo di quel gas a quella pressione. Tale stato di
equilibrio può permanenere fino a quando la pressione esterna del gas resterà inalterata, altrimenti,
se essa aumenta, altro gas entrerà in soluzione. Se diminuisce, il liquido si troverà in una situazione
di sovrasaturazione ed il gas si libererà tornando all'esterno fino a quando le pressioni saranno
nuovamente equilibrate. L’espressione matematica della legge è la seguente : P = k C dove P è la
pressione del gas sulla soluzione , C è la concentrazione del gas nella soluzione e k è una costante
tipica di ciascun gas che correla la pressione del gas sulla soluzione e la sua concentrazione, ad
esempio: aumentando la temperatura , che provoca un aumento dell'energia cinetica del gas, si
procura una diminuzione della solubilità per l'effetto dell’ allontanamento delle molecole gassose
dalla fase liquida.

8
Figura 5 . William Henry (1775 – 1836) è stato un chimico inglese. Egli formulò nel 1803 una
legge che regolava la solubilità dei gas nei liquidi.
Ogni gas, così come viene specificato nella legge delle pressioni parziali di Dalton
(la pressione totale esercitata da una miscela ideale di gas ideali, è uguale alla somma delle
pressioni parziali che sarebbero esercitate dai gas se fossero presenti da soli in un eguale volume.
La pressione parziale P di un componente di una miscela di gas è la pressione che questo avrebbe
qualora occupasse, da solo, il volume a disposizione dell'intera miscela), può entrare in soluzione o
liberarsi indipendentemente da ciò che fanno gli altri gas presenti. Quindi, una definizione
alternativa della costante di Henry è la seguente: P = kx , dove P è la pressione parziale del gas
sopra la soluzione e x è la frazione molare del gas disciolto. Secondo questa definizione, a 298°
K la costante di Henry assume i valori: O2 : k = 4,34×104 l·atm/mol ; CO2 : k = 1,64×10
3
l·atm/mol; H2 : k = 7,04×104 l·atm/mol. Il legame dell’O2 alla Mb o alla Hb può essere
studiato con metodi spettroscopici (spettrofotometria UV -visibile) (Fig. 6)
Figura 6. Determinazione dell’emoglobina con metodi spettrofotometrici. Il colore della proteina e
diverso a seconda del fatto che l’ossigeno sia o no legato al Fe++.
Il legame di Mb all’O2 è un legame reversibile. Si tratta di una reazione all’equilibrio
dove Mb + O2 MbO2 la cui costante di dissociazione o di equilibrio K è rappresentata da
[Mb][O2]
K = ------------------
[MbO2]
Dove [MbO2] = concentrazione della ossimioglobina e [Mb] = concentrazione della
deossimioglobina e [O2] = concentrazione dell’ossigeno non legato. Tutte le concentrazioni sono
espresse in moli/litro. La dissociazione dell’O2 da Mb può essere caratterizzata mediante la sua
“saturazione frazionale Y O2 ” cioè la frazione di siti di ossigeno legato

9
[MbO2] [O2]
YO2 = ---------------------------- = ------------------------- cioè frazione di siti di legame di O2 occupati
[Mb] + [MbO2] K + [O2]
Essendo O2 un gas la [O2] si può scrivere in funzione della pressione parziale pO2 o tensione di
ossigeno. Quindi si avrà
pO2
YO2 = ---------------- dove K può essere anche sostituito con il termine P50
K + pO2
pO2
L’equazione YO2 = ------------------------ rappresenta graficamente una curva iperbolica (Fig. 7)
P50 + pO2
Figura 7. Curva di legame dell’O2 alla Mb. In ascisse la pO2 (pressione parziale di O2) e in
ordinata la % di saturazione della Mb ( YO2) o la frazione dei siti di legame di una proteina
quando viene occupata dal ligando. La Mb è per metà saturata dall’O2 ( YO2 = 0,5 : P50) alla
pressione parziale pO2 di circa 1 Torr. La pO2 va da 100 torr dei distretti arteriosi ai 30 torr dei
distretti venosi. In entrambe queste condizioni, la mioglobina che è nel muscolo si satura quasi
completamente di O2
I punti essenziali del grafico sono :
a) a bassi valori di pO2 si lega poco O2 alla Mb
b) all’aumentare dei valori di pO2 aumentano i siti occupati da O2
c) la pendenza della curva aumenta al diminuire di K
d) quando K è uguale a pO2 si ha YO2 = 0,5 cioè la Mb è saturata al 50% da O2
e) operativamente la costante di dissociazione K può essere definita come: K = P50
f) la Mb ha una P50 molto bassa (circa 1 torr) e quindi un’ alta affinità per l’O2

10
g) la Mb deve essere in grado di catturare, nel citoplasma di cellule metabolicamente
molto attive, l’O2 proveniente dal torrente circolatorio (pO2 circa 30 torr) ed essere
in grado di rilasciarlo ai mitocondri (pO2 <4 torr)
h) alle pO2 fisiologiche nel sangue (100 torr arterie, 30 torr vene) la Mb è sempre
saturata (quando pO2 = 100 torr ⇒ YO2 = 0.97; quando pO2 = 30 torr ⇒ YO2
= 0.89)
i) in aggiunta, la Mb è molto efficace nel favorire il passaggio di O2 dai capillari
alle cellule muscolari.
Emoglobina
Le subunità e dell’emoglobina hanno lo stesso disegno strutturale della mioglobina ma
l’emoglobina è una molecola molto più complicata della mioglobina. Oltre all’O2 l’Hb è in grado
di trasportare anche H+ e CO2 . In aggiunta, le proprietà di legame con l’ossigeno nell’Hb sono
controllate da interazioni tra siti lontani nella molecola. Queste proprietà rendono l’Hb una proteina
allosterica mentre la Mb non lo è. Questa differenza può essere espressa in tre modi :
a) il legame di O2 all’Hb favorisce il legame di altro O2 alla stessa molecola; ciò
significa che l’O2 si lega in maniera cooperativa all’Hb mentre il legame
dell’O2 alla Mb non è cooperativo (curva sigmoide).
b) L’affinità dell’Hb per l’O2 dipende dal pH mentre nella Mb è indipendente
dal pH .Anche la CO2 altera le proprietà di legame dell’O2 per l’Hb. Sia H+
che CO2 promuovono il rilascio di O2 legato. Viceversa, l’O2 promuove il
rilascio di H+ e della CO2 legata (Effetto Bohr).
c) L’affinità dell’Hb per l’O2 è inoltre regolata da composti organici con gruppi
fosforici ( 2,3 bifosfoglicerato o BPG) (adattamento ad alte quote).
Come risultato finale, è che l’Hb presenta un’affinità per l’O2 più bassa della Mb.
La saturazione definita come la frazione dei siti leganti ossigeno occupata da molecole di ossigeno
in soluzione viene indicata con la lettera Y. Il valore di Y varia da 0 (tutti i siti sono vuoti) a 1 (tutti
i siti sono occupati). La rappresentazione grafica in un asse cartesiano di Y (in ordinata) in
funzione di pO2 o pressione parziale di O2 (in ascisse) viene chiamata curva di dissociazione
dell’ossigeno. (Fig. 8). Se si disegna un grafico della curva di dissociazione dell’ossigeno al
variare della pressione parziale dell’ossigeno (pO2) per la Mb e per l’Hb si noteranno tra queste
due proteine delle sostanziali differenze (Fig. 8).
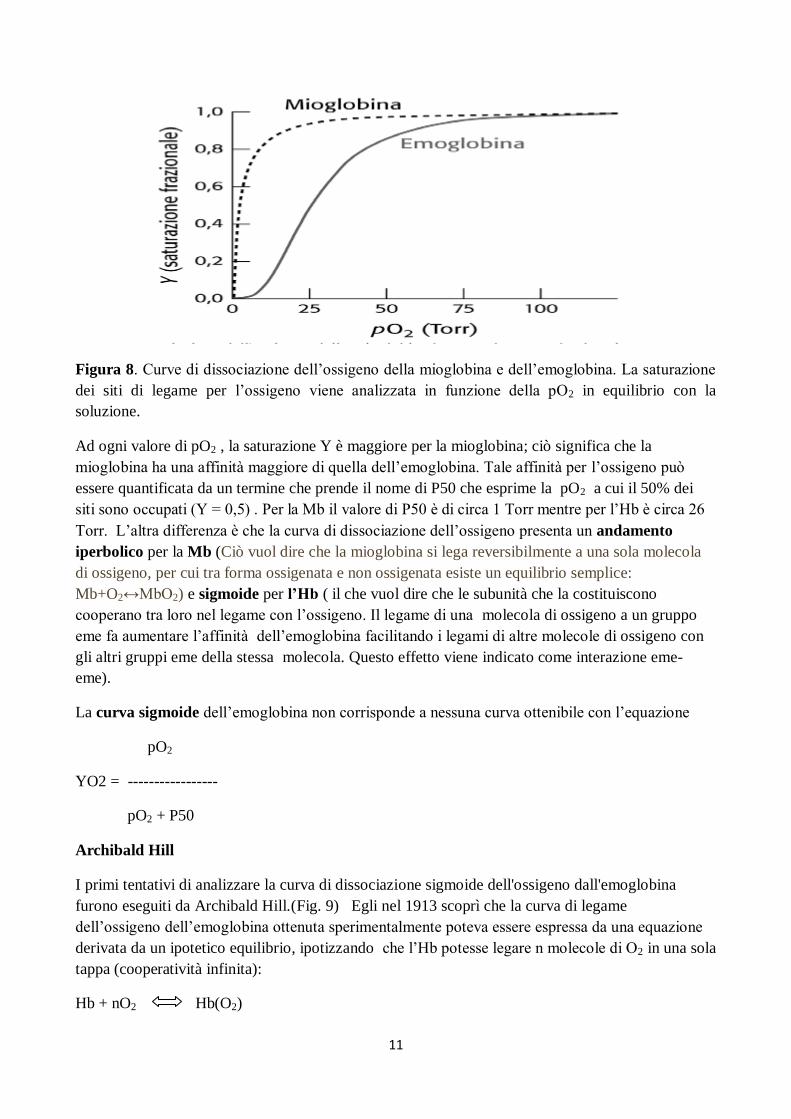
11
Figura 8. Curve di dissociazione dell’ossigeno della mioglobina e dell’emoglobina. La saturazione
dei siti di legame per l’ossigeno viene analizzata in funzione della pO2 in equilibrio con la
soluzione.
Ad ogni valore di pO2 , la saturazione Y è maggiore per la mioglobina; ciò significa che la
mioglobina ha una affinità maggiore di quella dell’emoglobina. Tale affinità per l’ossigeno può
essere quantificata da un termine che prende il nome di P50 che esprime la pO2 a cui il 50% dei
siti sono occupati (Y = 0,5) . Per la Mb il valore di P50 è di circa 1 Torr mentre per l’Hb è circa 26
Torr. L’altra differenza è che la curva di dissociazione dell’ossigeno presenta un andamento
iperbolico per la Mb (Ciò vuol dire che la mioglobina si lega reversibilmente a una sola molecola
di ossigeno, per cui tra forma ossigenata e non ossigenata esiste un equilibrio semplice:
Mb+O2↔MbO2) e sigmoide per l’Hb ( il che vuol dire che le subunità che la costituiscono
cooperano tra loro nel legame con l’ossigeno. Il legame di una molecola di ossigeno a un gruppo
eme fa aumentare l’affinità dell’emoglobina facilitando i legami di altre molecole di ossigeno con
gli altri gruppi eme della stessa molecola. Questo effetto viene indicato come interazione eme-
eme).
La curva sigmoide dell’emoglobina non corrisponde a nessuna curva ottenibile con l’equazione
pO2
YO2 = -----------------
pO2 + P50
Archibald Hill
I primi tentativi di analizzare la curva di dissociazione sigmoide dell'ossigeno dall'emoglobina
furono eseguiti da Archibald Hill.(Fig. 9) Egli nel 1913 scoprì che la curva di legame
dell’ossigeno dell’emoglobina ottenuta sperimentalmente poteva essere espressa da una equazione
derivata da un ipotetico equilibrio, ipotizzando che l’Hb potesse legare n molecole di O2 in una sola
tappa (cooperatività infinita):
Hb + nO2 Hb(O2)

12
(pO2)n
Y pO2 n
Dove Y = ------------------ che può essere arrangiata per dare ---------- = ----------
(pO2)n + (P50)
n 1 - Y P50
Questa equazione stabilisce che il rapporto tra ossiemoglobina (Y) e deossiemoglobina (1 – Y) è
uguale alla n potenza del rapporto tra pO2 e P50 . Facendo il logaritmo di entrambi i termini
dell’equazione si avrà :
Y
log ----------- = n log pO2 - n log P50
1 - Y
L’ equazione che permette di comprendere il grado di saturazione dell’emoglobina in funzione
della pressione parziale dell’ ossigeno (pO2)
Il grafico di log [Y/ 1 – Y] in funzione di log pO2 rappresenta una retta e viene chiamato grafico di
Hill (Fig. 10). La quantità n, cioè il coefficiente di Hill, è una misura della cooperatività del sistema.
Quando:
a. n = 1: il sistema non è cooperativo. Il binding del legante è semplice
(comportamento iperbolico)
b. n > 1: il binding presenta cooperatività positiva (curva sigmoide)
c. n < 1: il binding presenta cooperatività negativa (curva a scalino)
La cooperatività positiva (n>1) del legame dell’ossigeno nell’Hb deriva dalle modificazioni
strutturali che il binding di una molecola di O2 ad un gruppo eme può indurre sull’affinità per l’O2
di un altro gruppo eme.
Quale è il significato biologico del legame cooperativo dell’ossigeno all’emoglobina ?
Il legame cooperativo dell’ossigeno rende l’emoglobina un trasportore di ossigeno più efficiente
rispetto ad un legame non cooperativo.

13
Figura 9. Il grafico di Hill
Figura 10. Archibald Vivian Hill (1886 – 1977) è stato un fisiologo britannico, insignito del
Premio Nobel per la medicina nel 1922, insieme a Otto Meyerhof per i meccanismi di produzione
del calore nei muscoli scheletrici. Fra i suoi primi lavori spiccano la caratterizzazione delle
cinetiche enzimatiche, oggi note come equazioni di Michaelis-Menten, e lo sviluppo
del coefficiente di Hill. La sua prima pubblicazione avvenuta nel 1909, ha infatti fornito le basi per
la comprensione dei meccanismi allosterici e del legame cooperativo delle proteine.
Effetto Bohr
Mentre nella Mb non si hanno differenze nel legame dell’ossigeno in un largo range di pH così
come per la CO2, nell ’Hb una acidità stimola il rilascio di ossigeno. Infatti, un abbassamento del
valore del pH sposta la curva di dissociazione dell’ossigeno verso destra, riducendo in questo modo
l’affinità dell’Hb per l’O2 . Anche concentrazioni crescenti di CO2 seppure a valori di pH costanti
abbassano l’affinità per l’ossigeno. In tessuti metabolicamente attivi come il tessuto muscolare in
contrazione, vengono prodotte grandi quantità di CO2 e acidi (H+). Quindi, la presenza di elevati
livelli di CO2 e di ioni H+ nei capillari nel tessuto muscolare determina il rilascio di O2 dall’Hb. La
scoperta del meccanismo adatto a rifornire di grandi quantità i tessuti metabolicamente attivi come
i muscoli , si deve a C. Bohr nel 1904 (Fig.11)

14
Figura 11. Christian Bohr (1855-1911) medico danese a cui si deve la descrizione del fenomeno
ora chiamato effetto Bohr.
L’effetto reciproco spiegato dieci anni dopo da J.S. Haldane (Fig.12) avviene nei capillari dei
polmoni. L’elevata concentrazione di O2 nel tessuto polmonare induce il distacco di CO2 e di H+
dall’Hb esattamente come CO2 e H+ provocano il rilascio di O2 nei tessuti metabolicamente attivi.
Le interconnessioni tra gli effetti causati da O2, H+ e CO2 sono noti come effetto Bohr.
Figura 12. John Scott Haldane (1860 – 1936) fisiologo britannico a cui si deve la scoperta
dell’effetto Haldane , una proprietà dell’ emoglobina
Adattamento ad alte quote
L’affinità dell’Hb per l’O2 nei globuli rossi è più bassa di quella dell’emoglobina in soluzione. Si
deve a due biochimici americani Reinhold Benesch e Ruth Benesh nel 1967 la scoperta del 2,3
bifosfoglicerato o BPG chiamato anche 2,3 difosfoglicerato o 2,3 DPG. Il 2,3 bifosfoglicerato e un
sottoprodotto della glicolisi e si lega all’Hb provocando profonde modificazioni nella sua affinità
per l’ossigeno. Questo composto organico contenente fosfato è presente nei globuli rossi umani alla
stessa concentrazione molare dell’Hb. In assenza di BPG , il P50 dell’Hb è come quello della Mb (
circa 1 Torr) e in presenza di BP G , il P50 dell’Hb aumenta a 26 Torr. Pertanto il BPG è essenziale
per poter scaricare ossigeno nei capillari tissutali.In poche parole, la sua presenza e un segnale di
necessita di O2.Il BPG diminuisce l’affinità per l’ossigeno legandosi alla deossiemoglobina ma non
alla ossiemoglobina.

15
Come passa l’emoglobina dalla struttura deossi a quella ossi mentre lega di seguito molecole
di ossigeno ? E quale è il reale meccanismo allosterico dell’emoglobina ?
Le proprietà allosteriche dell’emoglobina derivano da interazioni tra le subunità e . Come
avviene la transizione T (struttura tesa, rigida) – R (struttura rilassata)? Quando si lega
all’ossigeno, i contatti tra le subunità 11 cambiano poco mentre un grande cambiamento si ha
nei contatti tra le subunità 1 2. L’analisi della struttura ai raggi X ha rivelato due conformazioni
per la emoglobina: lo stato R e quello T. Anche se l’ossigeno si lega ad ambedue gli stati si ha una
considerevole più alta affinità per lo stato R. Quando l’ossigeno è assente sperimentalmente , lo
stato T è più stabile e così lo stato T è lo stato predominante per la desossiemoglobina. T e R
stanno per “teso” e “rilassato”, questo perché lo stato T è stabilizzato da un più grande numero di
coppie ioniche nelle interfacce 12, 2 1.
L’ossigeno, legandosi all’emoglobina nello stato T, provoca un cambiamento conformazionale e la
porta nello stato R. Quando l’intera struttura subisce questo cambiamento, la struttura delle singole
subunità cambia poco ma le coppie di subunità ab scivolano e ruotano, stringendo la tasca tra le
subunità b. In questo processo alcune delle coppie ioniche che stabilizzano la forma T vengono rotte
e se ne formano delle nuove.
Lo stato desossigenato è detto T (tense) (Fig.13) : ha bassa affinità per l’O2 e il Fe+ +
è spiazzato
0.6 Å in direzione prossimale. L’eme ha una forma a cupola. Lo stato ossigenato (Fig.14) è detto R
(relaxed): ha alta affinità per l’O2 e il Fe++
giace nel piano dell’eme (che assume una forma
planare)
Figura 13. Forma deossigenata Figura 14. Forma ossigenata
Il BPG stabilizza la forma T ( deossigenata) legandosi a gruppi carichi positivamente nella cavità
centrale dell’Hb.
La CO2 un altro effettore allosterico si lega reversibilmente ai gruppi NH2-terminali di tutte le
quattro catene sotto forma di carbamato . Questa reazione produce H+ e contribuisce all’effetto
Bohr.
O-
CO2 + --NH2 ==== C—NH-- + H+
O

16
Gli ioni H+ si legano invece ad alcune coppie di siti che hanno un ambiente immediatamente
circostante più negativo nella forma deossigenata che nella forma ossigenata della molecola
Sono stati sviluppati due modelli limite : a) modello sequenziale nella forma più semplice
sviluppata da D. Koshland Jr. e b) (Fig. 15) il modello concertato o MWC model (da J. Monod -
J. Wyman-J.P. Changeux model) (Fig.15) con nuove visioni delle interazioni allosteriche. Nel
modello sequenziale semplice il legame del ligando induce un cambiamento conformazionale in
una subunità. Questo produce cambiamenti simili nelle subunità adiacenti . Possono esistere quindi,
un gran numero di conformazioni. Nel modello concertato dove viene conservata la simmetria, si
assume che tutte le subunità in una particolare proteina devono essere o nella forma R o nella
forma T. Il legame di un ligando aumenta la proporzione di molecole che si trovano nella forma a
R. I due modelli non si escludono a vicenda In realtà, Il meccanismo allosterico dell’emoglobina è
più complesso di quello proposto da ciascin modello.
Figura 15. Daniel E. Koshland, Jr. (30 marzo 1920 - 23 luglio 2007) biochimico statunitense e
Jacques Lucien Monod ( 1910 – 1976) biologo francese, vincitore del Premio Nobel per la
medicina nel 1965.per la teoria dell'operone
Bibliografia essenziale
Kendrew J.C. “The three dimensional structure of a protein molecule” Sci.Amer 1961,205,96.
Perutz M. “Hemoglobin structure and respiratory transport “ Sci Amer 1978, 239, 92.
Monod J. Wyman J and Changeux J.P. “ On the nature of allosteric transitions . A plausible
model”. J Mol Biol 1965, 12, 88.