La popolazione come livello di organizzazione
14
This article was downloaded by: [SUNY Health Science Center] On: 06 October 2014, At: 05:42 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Bolletino di zoologia Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/tizo19 La popolazione come livello di organizzazione Agostino Parise a a Istituto di Biologia Animale, Università di Padova Published online: 14 Sep 2009. To cite this article: Agostino Parise (1969) La popolazione come livello di organizzazione, Bolletino di zoologia, 36:3, 217-228, DOI: 10.1080/11250006909436918 To link to this article: http://dx.doi.org/10.1080/11250006909436918 PLEASE SCROLL DOWN FOR ARTICLE Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) contained in the publications on our platform. However, Taylor & Francis, our agents, and our licensors make no representations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of the Content. Any opinions and views expressed in this publication are the opinions and views of the authors, and are not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon and should be independently verified with primary sources of information. Taylor and Francis shall not be liable for any losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoever or howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use of the Content. This article may be used for research, teaching, and private study purposes. Any substantial or systematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form to anyone is
Transcript of La popolazione come livello di organizzazione

This article was downloaded by: [SUNY Health Science Center]On: 06 October 2014, At: 05:42Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH,UK
Bolletino di zoologiaPublication details, including instructions for authorsand subscription information:http://www.tandfonline.com/loi/tizo19
La popolazione come livello diorganizzazioneAgostino Parise aa Istituto di Biologia Animale, Università di PadovaPublished online: 14 Sep 2009.
To cite this article: Agostino Parise (1969) La popolazione come livello diorganizzazione, Bolletino di zoologia, 36:3, 217-228, DOI: 10.1080/11250006909436918
To link to this article: http://dx.doi.org/10.1080/11250006909436918
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all theinformation (the “Content”) contained in the publications on our platform.However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness,or suitability for any purpose of the Content. Any opinions and viewsexpressed in this publication are the opinions and views of the authors, andare not the views of or endorsed by Taylor & Francis. The accuracy of theContent should not be relied upon and should be independently verified withprimary sources of information. Taylor and Francis shall not be liable for anylosses, actions, claims, proceedings, demands, costs, expenses, damages,and other liabilities whatsoever or howsoever caused arising directly orindirectly in connection with, in relation to or arising out of the use of theContent.
This article may be used for research, teaching, and private study purposes.Any substantial or systematic reproduction, redistribution, reselling, loan,sub-licensing, systematic supply, or distribution in any form to anyone is

expressly forbidden. Terms & Conditions of access and use can be found athttp://www.tandfonline.com/page/terms-and-conditions
Dow
nloa
ded
by [
SUN
Y H
ealth
Sci
ence
Cen
ter]
at 0
5:42
06
Oct
ober
201
4

Boll. ZOO^., 36: 217-228, 1909 A f f i del XXX V I I I Convegno del1’U.Z.Z.
LA POPOLAZIONE COME LIVELLO D I ORGANIZZAZIONE
ACOSTINO PARISE Istituto di Biologirr Anhale, UnlversitA di Pedova
La materia viventc non costituiscc uno strato continuo nella bio- sfera del nostro pianeta, ma si presenta frammentata in unith discrete ge- neralmente, ma non sempre, separate e riconoscibili, dette individui. Que- sto mod0 di distribuzione della materia organizzata, ha attirato sull’indi- viduo un eccesso di attenzione da parte degli osservatori chc solo da un secolo ad oggi B compensato da un cresccntc intcrcssc per le popolazioni, interesse senza dubbio destato dall’affermarsi della teoria evolutiva. Que- sto mod0 pih recente di considerare la materia organizzata- & certamente all’origine delle tecniche statistiche e del conseguente sviluppo che questc hanno consentito alla genetica e alla ecologia.
L’approccio organismico e l’approccio popolazionistico sono due modi complementari di studio della materia vivente e, ben lontani dall’csclu- dersi, sono cntrambi legittimi e necessari per comprcndernc le manifesta- zioni. D’altra parte lc delimitazioni convenzionali della biologia stanno rapidamente sparendo e lo prova la rapiditl con cui il concetto di seledone si B esteso a1 livello molecolare ed i modelli ecologici sono stati applicati alla evoluzione chimica prebiologica, mentre ii concetto di flusso di energia ha introdotto nell’ecologia i modelli della chimica-fisica e l’ecosistema si delinea scmpre meglio come I’unith biologica elementare.
I1 concetto di popolaziom, che si tratti di molecolc, cellule o orga- nismi, B intuitivamente molto semplice ; una sun eventuale definizione rigorosa prcsenta invece alcune dificoltl, esso & owiamente in relazione con le proprieth degli individui cd il luogo in cui essi si trovano. A livello organismico, nella pratica gcnetica ed ecologicn, si & impost0 un concetto convenzionale detto u popolazione mendeliana u di indubbin utilita. In laboratorio lc dificolth concettuali sono ovvinmente sormontntc in pratica con l’esplicito rifcrimento ai ceppi in colturn dei quali B indicata la loca- lith di provcnienza e, per quanto & possibile, l’identith sistemntica.
Gih nel 1924 LOTKA osservava che a livello molecolare si studiano
Dow
nloa
ded
by [
SUN
Y H
ealth
Sci
ence
Cen
ter]
at 0
5:42
06
Oct
ober
201
4

218
pi t facilmcntc lc propricth di insicmc chc quelle individuali, mcntrc a livello degli organismi si notnno pih facilmente le proprietb degli individui che I’cffctto collcttivo. Lc propricth degli individui in termini di popola- zione sono relativamente semplici rispetto a tuttc le caratteristiche dell’in- dividuo, I’effetto globale della popolazione riguards solo tale livello e seb- bene esso possa essere posto in relazione con caratteri individuali tale rela- zione non i: semprc orvia e va di volta in volta stabilita.
Lo studio di una popolazione h volto a rispondere ad una sola do- manda, l’unica d’altm partc rilevante per tale livello, e cioE come varia nel tempo la sun dimensione. A questo scopo si utilizzano per gli organismi due quantith, il tasso intrinseco di accrescimento ed il tasso netto di ripro- duzione, indicate tradizionalmente con le lettere r ed Ro. Esse sono state individuate da LOTPA (1925) per la demografia umana ed applicate ad altri organismi per la prima volta da BIRCII. (1948). Non & detto che queste quantith sinno di facile rilevamento in ogni tip0 di organism0 ; talvolta infatti h necessnria una cospicua dose di immaginazione per applicarle ad organismi a ciclo biologic0 complesso ed una ancor maggior capacith ana- litica per individuarne gli strumenti matematici di calcolo.
I1 tasso intrinseco di accrescimento r & definito dalla relazione d N / dt = rh’, in cui N h il numero degli individui e t il tempo, e rappresenta la velocith di accrescimento della popolazione, esso h perfettamente analogo, a1 livello molecolare, alla costante di velocith dclle reazioni chimiche. In-
tegrando e risolvendo rispetto ad r si ottiene r = -In NtIN,, dove No
& il numero di individui, o la concentrazione, iniziale. La sua determinazione sarcbbe semplice se le concentrazioni iniziali e finali fossero molto elevate, come avviene nel caso delle molecole. Per la misura della r in pratica con- viene prima definire la quantitb R,.
I1 tasso netto di riproduzione R, non h definibile facilmente per ogni organismo. Per quelli che sono utilizzati nel presente lavoro i: possibile misurare una quantith T, detta tempo medio di generazione, tale che R, = NT /No. Ro rappresenta quindi la quantith per cui si moltiplicano gli orga- nismi da una generazione a118 successiva, essa C comprensiva della soprav- vivenza e della fecondith specifica per etb. Sostituendo il valore di Ro
1 nella precedente equazione si otticne T = - l n R, che definisce la rela- T zione tra r cd R,. I1 problema della validith del calcolo di r i: quindi rin- viato ad una opportuna determinazione di T in funzione della lunghczza del periodo riproduttivo (cfr. LAUORLINO, 11165), per le considerazioni che seguono, l’errore di T h praticamente costante e quindi irrilevante ; que-
1 t
Dow
nloa
ded
by [
SUN
Y H
ealth
Sci
ence
Cen
ter]
at 0
5:42
06
Oct
ober
201
4

21Q
sta semplificazionc sarcbbc insostenibile se si dovcsscro confrontare popo- lazioni s ciclo biologico molto diverso.
Le due quantith r ed Ro variano a1 variare dellc condizioni ambientali second0 funzioni non nncora completamente note, ed uno dei pro$rammi dcl nostro laboratorio i: nppunto lo studio s u bnsi sperimcntali di tali funzioni.
I1 principale presupposto da cui muovono queste ricerche i: che Ic funzioni r ed Ro di tutte le variabili ambientali, descrivano completamente una popolazione dal punto di vista dinamico ed idcntificliino percib univo- camente un detcrminato assetto genetico.
Lo scopo dclla presentc esposizione 6 di riferire i primi risultati di queste indggini e di delinearnc lc prospettive future.
* * *
I1 materiale di ricerca 6 costituito da specie e razze del genere Tisbe (Copepoda, Harpacticoida), il metodo di alllevamento B quell0 mess0 a punto da BATTAGLIA (1958).
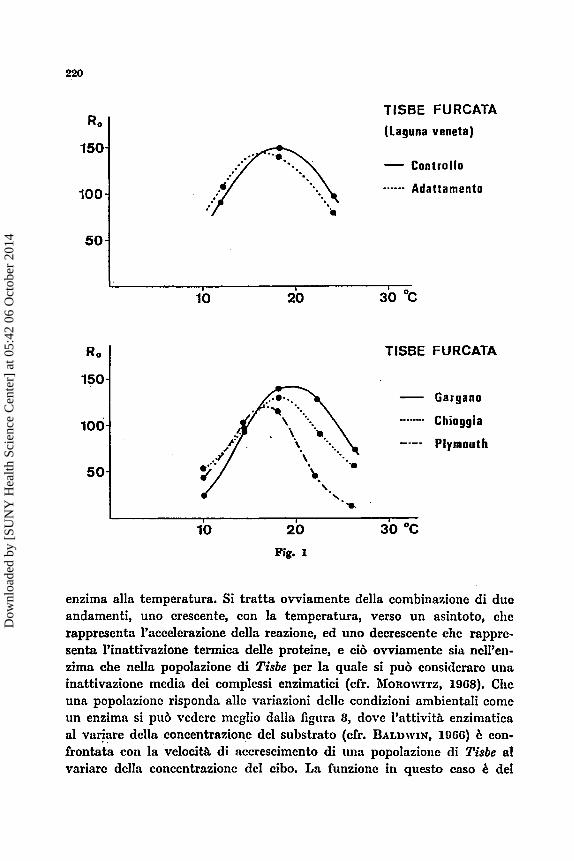
Nella figura 1, diagramma in basso, 6 rappresentata la-feconditb Ro di popolazioni di Tisbe ftcrcata rispcttivamcnte del Gargano, di Chioggia e di Plymouth a1 variare della temperatura. L’andamento della curva i: analog0 per le tre popolazioni, ma si pub notare che le tre curve presentano mas- simi diversi non solo per entita ma anche per posizione rispetto alla tem- pcratura. evidente in questo caso che sebbcne la risposta alla temperatura sia espressa dnllo stesso tip0 di funzione le costanti della funzione sono diverse per le trc popolazioni, e gizl la norma di risposta ad un solo para- metro ambientale serve a differcnziarc le popolnzioni. Cii, d’altra partc era atteso, da un punto di vista genetico, in quanto le tre popolazioni pro- vcngono ds tre latitudini diverse, ed era quindi presumibile che avessero un adattamento termico diverso. 2 da ribadire il fatto che non I? la fecon- dith, ma la norma di risposta della feconditii che caratterizza le popolazioni, infatti una eventuale misura di fecondita attorno ai 16OC non avrcbbe permcsso di rilevare alcuna differcnza, come pure una lettura tra 100 c 160 per le popolazioni di Chioggia e Plumouth.
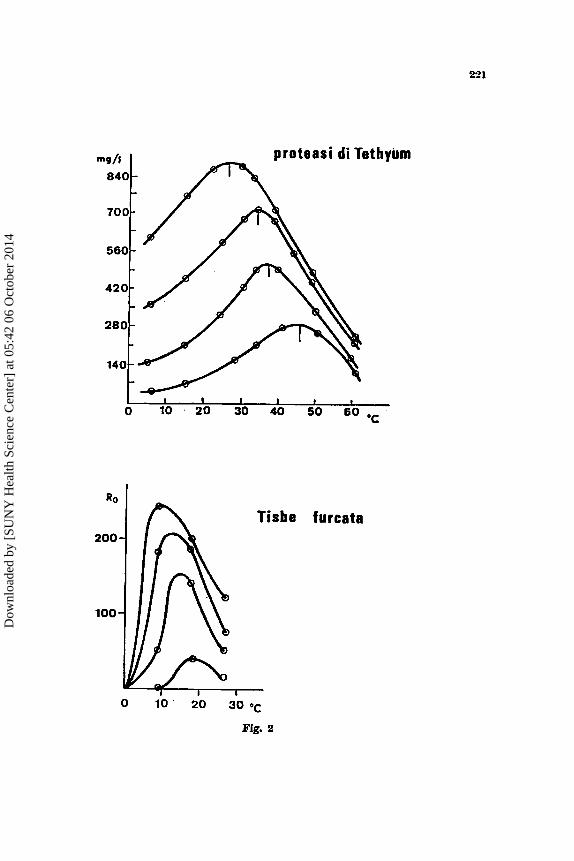
Quale sia il tipo di funzione che collega la fccondith alla tempera- tura b suggerito dalla figura 2 dove la norma di risposta della fecondit& di un’altra popolazione di Tisbe furcata B confrontata con la norma di rispo- sta dell’attivitii enzimatica di una protcasi d variare della temperatura. Le analogie di andamento sono evidenti, sfortunatamente non ho potuto trovare in letteratura nessun trattamento analitico della risposta di un
Dow
nloa
ded
by [
SUN
Y H
ealth
Sci
ence
Cen
ter]
at 0
5:42
06
Oct
ober
201
4
220
R O
150
100
50
TlSBE FURCATA (Laguna veneta)
I 1
10 20 30 "c
TlSBE FURCATA Ro I
150-
100-
50-
'x I
- Gargano
Chioggia -...... -.-- Plymouth
i o 20 30 "C
Fig. 1
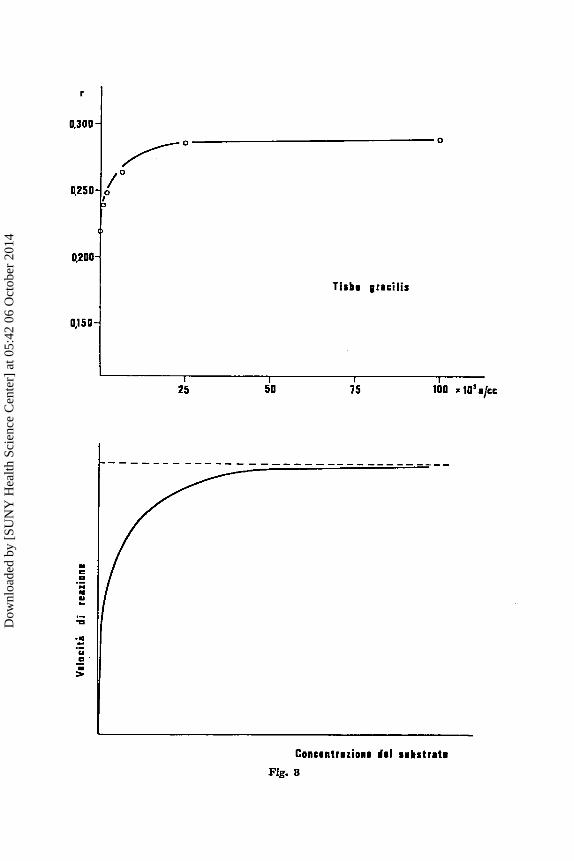
enzima alla temperatura. Si tratta ovviamente de11a combinazione di due andamenti, uno crescente, con la tempcratura, verso un asintoto, clie rappresenta I'accelerazione della redone, ed uno decrescente che rappre- senta l'inattivazione termica delle proteine, e cib ovviamente sia nell'en- zima che nella popolazione di Tisbe per la quale si pub considcrare una inattivazione media dei complessi enzirnatici (cfr. MOROWITZ, 1908). Chc unn popolnzione risponda alle variazioni delle condizioni ambientali come un enzima si pub vedere mcglio dalla figura 3, dove I'attivitg enzimatics a1 variare - . della concentrazione del substrato (cfr. BALDWIN, 1900) B con- frontata con la velocita di accrescimento di una popolnzione di Tisbe a1 variare della concentrazione del cibo. La funzione in questo cnso C del
Dow
nloa
ded
by [
SUN
Y H
ealth
Sci
ence
Cen
ter]
at 0
5:42
06
Oct
ober
201
4
21 1
mo/l proteasi di Tethyum 840 -
700- -
560 - -
420- -
200 - - 140 - -
I I I I 1 1
0 10 20 30 40 50 60 oc
Tisbe furcata
Dow
nloa
ded
by [
SUN
Y H
ealth
Sci
ence
Cen
ter]
at 0
5:42
06
Oct
ober
201
4
r
0.300
0,250
0,200
O,l50.
0
To /"
Tisba urrcilis
I I I I
25 50 75 100 .10'r/cc
Concrntrrzionr drl rrbrtrrto
Fig. 8
Dow
nloa
ded
by [
SUN
Y H
ealth
Sci
ence
Cen
ter]
at 0
5:42
06
Oct
ober
201
4
229
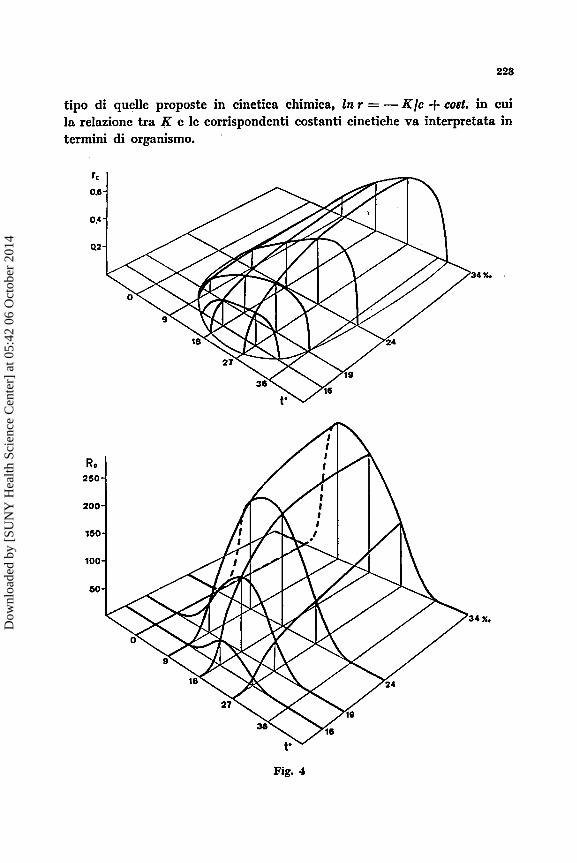
tipo di quelle proposte in cinetica chimica, In r = - K / c + cost. in cui la relazionc tra R e le corrispondenti costanti cinetiche va interpretata in termini di organismo.
rc 0.6
0.4
0.2
34%. .
Fig. 4
Dow
nloa
ded
by [
SUN
Y H
ealth
Sci
ence
Cen
ter]
at 0
5:42
06
Oct
ober
201
4

Se si esamina ora la risposta di una popolazionc alla varinzione con- temporanen di duc condizioni ambientali si ottiene un andamento conic qucllo rapprescntato nella figura 4. Ncllo stereogramma inferiorc la fecon- dits Ro ha lo stesso andamento, per la temperatura, a diverse salinith ; le singolc curve, rappresentate sullo stesso piano nella figura 2, hanno perb owiamcnte costanti divcrsc. I massimi di Ro retrocedono sulla tempera- tura a1 crescerc della salinith c ncl complcsso il tasso netto di riproduzione descrive una superficie a collina la cui curva di intcrsezionc con il piano orizzontale rappresenta il limitc di esistenza della popolazionc. Ricordando la relazione NT = NORo, sc Ro = 0 la popolazionc si estinguer8 comple- tamcntc in un tempo T, per la persistenza della popolazione dcve esserc almeno Ro = 1, cio& Nr = No. Esiste quindi una fascia inferiore della collina, sotto il valore 1 dell’ordinata verticale, per i valori della quale la popolazione h destinata a sconiparire, la superficie sopra la fascia rapprc- senta invece il campo, pcr assortimento dei valori delle condizioni ambienta- li, di csistcnza della popolazione. Analoghe considerazioni valgono per il tasso di accrescimcnto r ; la supcrficic dello stereogramma, in alto nella figura 4, sopra il piano orizzontale indica la capacith di accrcscimento della popo- lazione, la curva di intersezione rappresenta i valori delle ordinate per cui la popolazione 6 stazionaria ; per valori sotto il piano orizzontale la popo- lazione & in estinzione.
& possibile ora immaginare come si configura la popolazione pren- dendo in considerazione contemporaneamente tutte le variabili ambientali. La popolazionc descrive nello spazio ambientale di n variabili un ipervo- lumc di n + 1 dimensioni che definisce interamentc la popolazione. In particolare I’ipervolume definito dai valori positivi di r rappresenta il com- plesso dcllc condizioni ambicntali nelle quali la popolazione pub pagarc un tributo in termini energetici all’ecosistcma per mantenersi in equili- brio con esso. Tale tributo deve essere proporzionato, per ogni assorti- mento di variabili ambientali, a1 valore di r per qucll’assortimento; se l’ecosistema esige con continuits un contributo maggiore la popolazione devc modificarsi oppure si estingue.
Da qunnto 6 stato accennato risulta owio che per ogni variabile ambientale esiste un vdore massimo dei parametri dinamici, come pure B possibile immaginare un valore massimo assoluto, cio6 massimo in rela- zione a tutte le variabili ambientali ; tale valore massimo 6 costante per una dcterminata popolazione e la descrive interamente in termini dinamici. L’assortimento di valori delle variabili ambientali che definiscono il mas- simo assoluto dei parametri dinnmici, & ovviamente l’ambiente ottimale della popolazione nel quale B da ritenere che la pressionc di selezione sia
Dow
nloa
ded
by [
SUN
Y H
ealth
Sci
ence
Cen
ter]
at 0
5:42
06
Oct
ober
201
4

225
nulla e quindi il. suo assetto gcnctico sia stabile. Una popolazionc in qucstc condizioni descriverh scmpre nelle successive genernzioni lo stesso ipervo- lume nello spazio delle variabili ambientali ; per ogni altro assortimento di rariabili ambientali invece le gcnerazioni successive non prescnteranno neccssariamentc le stesse norme di reazione.
Un escnipio, a chiarimento dcll’affermazione precedcntc, 2: rapprc- sentato nella figura 1 in alto. Di una popolazione di Tisbe, dn tempo’accli- mata alle condizioni standard di allcraniento dei nostri lavoratori, 2: stata dcterminata la norma di reazionc dclla fccondith alla temperaturn cd tin ccppo di essa b stato conservato a lungo a parith dellc altrc condizioni, a tcmpcratura pih bassa. Si sono quindi di nuovo determinate le norme di reazione dcl ceppo alln tcniperatura originaria (180 C) e di quello acclima- tato a tcmpcratura pih bassa (120 C). Si pub notarc chc le normc di rea- zione dei due ceppi sono diverse c che il massimo dclla curvn di adatta- mcnto 6 spostato rispctto a quello dclla curva originaria. D a notarsi soprat- tutto la misura di Ro a 120 nei due ceppi: & evidcntc chc la fecondith a quella tcmpcratura b numentata rispetto alla prima lettura ; b da supporrc chc un piL anipio abbassamento o una pih lunga permnnenza a bassa tcm- peratura avrcbbero consentito una mnggiorc resa nella fecondith a 120.
Questa circostanza pub csscrc utilizzata per m a indaginc sulla fitncss globalc di una popolazione. Per scmplicitit consideriamo la norma di rca- zione dclln fecondith R, s u unh soln varinbile ambicntalc, ad cscmpio la tempcratura. Se una popolazionc chc prcsenta iin massimo vnlore di Ro per un dato valore di tcmpcratura, vicne portata n tempcratura diversa da quclla corrispondcntc a1 massimo si possono avcrc nellc gcncrazioni SUC-
cesivc clue casi possibili ; la norma di reazione della popolazionc lion varia oppure varia in qualchc niisura. I1 primo cnso rapprescntn iinn popola- zione chc dispone di m a fortc capacith di autoregolazionc termica, per cui qucsta variazionc ambicntalc, almeno nci limiti indagati, praticamentc non ha influenza nclla popolazionc stcssa. I1 sccondo cuso iiivcce B piii complcsso : i vari valori della fccondita a divcrsc temperature clic dcfini- scono la norma di risposta sono ovviamcntc da niisurarsi contcniporanca- mentc pcr una data gencrazionc ; Ic gcncrazioni siicccssivc lasciate allc temperature diverse dal massimo inizialc di fecondith, coniinceranno ad adattarsi allc nuovc temperature c tcndcranno ad innalzarc la loro fccon- ditA rispctto alla norma fino a stabilizzarsi ad un nuoro valorc, cvcntunl- mcntc diverso da quello clic inizialniente costituiva il massinio. L’insicme di qucsti nuovi vulori allc vnrie temperature descrive una curva, che chia- meremo dci massimi, rappresentata ipoteticamente nella figura 5 (linca punteggiata) ; I’area comprcsa tra la curva dei massimi c la curva della
Dow
nloa
ded
by [
SUN
Y H
ealth
Sci
ence
Cen
ter]
at 0
5:42
06
Oct
ober
201
4
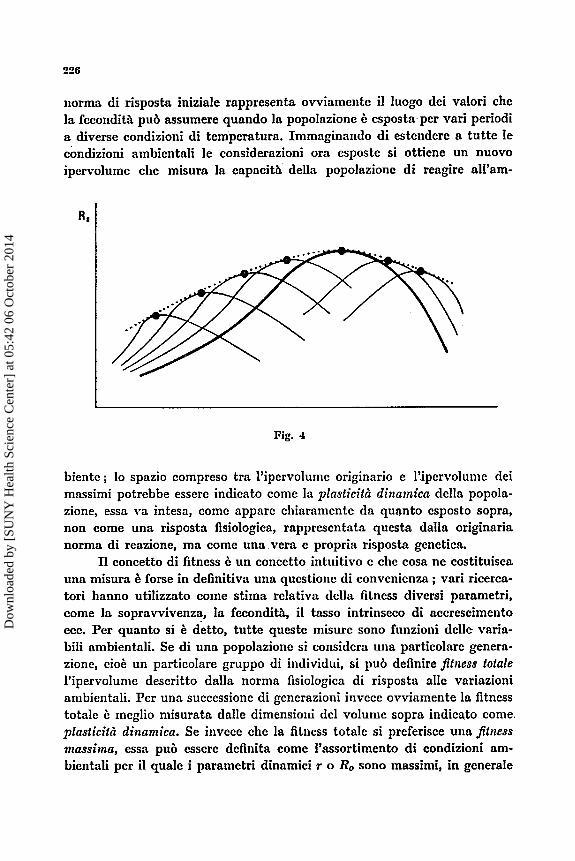
norma di risposta iniziale rappresenta ovviamente il luogo dei valori che la feeondith pub assumere quando la popolazione I: esposta,per vari periodi a diverse eondizioni di temperatura. Immaginando di estendere a tutte le condizioni ambientali le considerazioni ora esposte si ottiene un nuovo ipervolume ehe misura la capacith della popolazione di reagire all’am-
Fig. 4
biente ; lo spazio compreso tra I’ipervolume originario e l’ipervolume dei massimi potrebbe essere indieato come la plasticit2 diizninicn della popola- zione, essa va intesa, come appare cliiaramente da quanto esposto sopra, non come una risposta fisiologica, rappresentata questa dalla originaria norma di reazione, ma come una Vera e propria risposta genetica.
I1 concetto di fitness B un concetto intuitivo e che cosa ne costituisca una misura I: forse in definitiva una questione di convenienza ; vari ricerca- tori lianno utilizzato come stima relativa della fitness diversi parametri, come la sopravvivenza, la fecondith, il tasso intrinseco di accrescimento ecc. Per quanto si i? ditto, tutte queste misure sono funzioni dclle varia- bili ambientali. Se di una popolazione si considera una particolare genera- zione, ciob un particolare gruppo di individui, si pub definire fitness totale l’ipervolunie deseritto dalla norma fisiologica di risposta alle variazioni ambientali. Per una successione di generazioni invece ovviamente la fitness totale i: meglio niisurata dalle dimensioni del volume sopra indicato come. plasticit2 dinarnica. Se invece che la fitness totale si preferisce una fitiicss mmsiina, essa pub esserc definita come I’assortimento di condizioni am- bientali per il quale i parametri dinamici r o R, sono massimi, in generale
Dow
nloa
ded
by [
SUN
Y H
ealth
Sci
ence
Cen
ter]
at 0
5:42
06
Oct
ober
201
4

227
allora la fitticss rclativa per qualsiasi assortiniento di condizioni sarA la mi- sura del tnsso intrinscco di accrcscimcnto o del tasso netto di riproduzione per quell'assortimento di condizioni ambientali.
In population dynamics tlic population may be considcrcd as a set of elements reacting in a changing environment just like a solution of a chemical substance. As in chemical kinetics indeed we may define for a population a reaction constant
r = - In A'/N0 or t = -- in A'(I<Aro - l)/iVo(KN - 1). whose mathematical expres-
sion is formally identical with the related constant of the first order and second order autocatalytic reactions.
In tlie case of a population thc constant r is referred to as the tnallhusian para-
nieter and its actual calculation is often carried through the formuln r = - I n Ro
(about the error involved in using this expression see IY~UGHLIN~ 1906). in which T is thc mean gcneration time and Ro is the net rcproduction ratc DS dcviscd by LOTKA (1025).
I f it is now taken into account how both the constants r and Ro behave in a changing environment, a striking relation may bc dctected bctwccn the r and I20 func- tions and tlie kinetics of the enzymes.
To point out this fact laborntory populations of different species or races of the Gcnus Tisbe (Copepoda, Harpacticoido) have been raised a t diffcrcnt conditions of temperature, salinity and food supply. As it can be seen the Ro does vary a t changing temperaturc (fig. 2) and food supply (fig. 3) as an enzyme a t dinercnt temperatures and substratc concentrations. It should be noted in this connection that i t is the shape of the curve which is rclevnnt, while the curve constants arc obviously different in diffe- rent populations, as can be seen in three dinerent races of Tisbe (fig. 1, bclow).
If wc let two environmental conditions cliange simultaneously, sue11 as tempe- rature and salinity, r and Ro will describe a curved surface as in fig. 4. Taking now into account nll the relevant environmental variables (mitli thc csccption of predators and parasites) the population shall be tlicn completely described by an hypervolume. In particular the hypervolume described by the positivc values of the riialtliusian para- meter r will definc every set of environmental conditions in which the population can pays an cncrgy toll to predators and parasites, proportional to tlic value of t for the given set, to mantain herself in balance wliitin tlic ecosystem.
The r and Ro values so fa r considercd liowcvcr, have been mcasurcd in cohorts of coeval newborns coming from the samc strain raised at the standard laboratory condi- tions, and the r's and no's curves should bc considered as pliysiological response curves. I f we let the following generation of a cohort to develop in conditions, say of tempera- ture, far from the standard ones, after several gencrations (fig. 1, abovc) n dincrent phy- siological response curve (ndattamcnto) as compared with the previous onc (controllo) may occur. W e may tlicn suppose that a population may develop in the following ge- nerations a complete set of physiological response curves whose maxima will describe a new curve (fig. 5, dotted line) which shall account for a dynamic pZasficify of the popu- lation. Taking into account, as before, all tlic relevant environmental conditions wc may
1 1 t t
1 . T
Dow
nloa
ded
by [
SUN
Y H
ealth
Sci
ence
Cen
ter]
at 0
5:42
06
Oct
ober
201
4

modcl a new hypervolume which will define the dynamic plasticity of a population and which represents n truly genetic response volume.
The word 5tness has a n intuitive meaning and its definition may be a matter of convenience. We may state, from n population dynamics point of view, that n total fitness may be measured, in n slowly changing environment. by the dynnmie plasticity hypervolume or, in n swiftly (in relation t o thc population 'generation time) changing environment, b y the physiological response hypervolume. In this sense n maximum fitness could be the maximum value of r or 22, of a given generation taking into account every possible set of conditions,' and the relatiue fitness the r or Ro values as occur in any undefined set of environmental conditions.
BIBLIOGRAFIA
BALDWIN E., 1966 - Biochirnica dinarnica. Boringhieri, Torino. BAITAQUA B., 1958 - 11 polinwrJsmo adanioo e i fattori della selerione nel Coepode Tisbc
BIRCX L. C., 1908 - The infrinsic rate of natural increase of an insect population. J. Anim.
LAWQIILING R., 1965 - Capacify for increme a useJulpopuIafion statistic. J . h i m . Ecol.,34. LOTKA A. J. e DUBLIN L. I., 1925 - On fhe true r& of natural increase. J. Am. Statist.
MoRowrTz XI. J., 1068 - Energy floro in biology. Academic Press. h'ew York-London.
reticulatn Boquef. Arch. Oceanogr. Limnol., 11.
Eco., 17.
Ass., 20.
Dow
nloa
ded
by [
SUN
Y H
ealth
Sci
ence
Cen
ter]
at 0
5:42
06
Oct
ober
201
4