
I Potenziali d’azione - omero.farm.unipi.itomero.farm.unipi.it/matdidFarm/14/08-03-12.pdf ·...
25
I Potenziali d’azione I potenziali d’azione sono variazioni transitorie del potenziale di riposo (Vm), tipici delle cellule eccitabili (neuroni, muscolo) ma anche di cellule secernenti e protozoi. E’ un evento del tipo “tutto o nulla”, una volta innescato, il potenziale d’azione produrrà una variazione del potenziale di membrana della stessa ampiezza e con lo stesso andamento temporale. La genesi di un potenziale d’azione dipende da tre fattori: Trasporto attivo degli ioni operato da proteine specifiche di membrana in grado di mantenere una differente concentrazione ionica ai lati della membrana citoplasmatica. Presenza di un gradiente elettrochimico che costituisce la fonte di energia potenziale Apertura di canali ionici selettivi che permette il fluire degli ioni secondo i relativi gradienti di concentrazione.
Transcript of I Potenziali d’azione - omero.farm.unipi.itomero.farm.unipi.it/matdidFarm/14/08-03-12.pdf ·...

I Potenziali d’azione
I potenziali d’azione sono variazioni transitorie del potenziale di riposo (Vm), tipici delle cellule eccitabili (neuroni, muscolo) ma anche di cellule secernenti e protozoi.
E’ un evento del tipo “tutto o nulla”, una volta innescato, il potenziale d’azione produrrà una variazione del potenziale di membrana della stessa ampiezza e con lo stesso andamento temporale.
La genesi di un potenziale d’azione dipende da tre fattori:
Trasporto attivo degli ioni operato da proteine specifiche di membrana in grado di mantenere una differente concentrazione ionica ai lati della membrana citoplasmatica.
Presenza di un gradiente elettrochimico che costituisce la fonte di energia potenziale
Apertura di canali ionici selettivi che permette il fluire degli ioni secondo i relativi gradienti di concentrazione.

TUTTO O NULLA
1) Tutto o nulla
2) Soglia
3) Propagazione
POTENZIALE D’AZIONE

SOGLIA
La soglia per il potenziale
d’azione non é costante
nel tempo, ma é
modificabile dall’attività
pregressa (refrattarietà
assoluta e relativa).
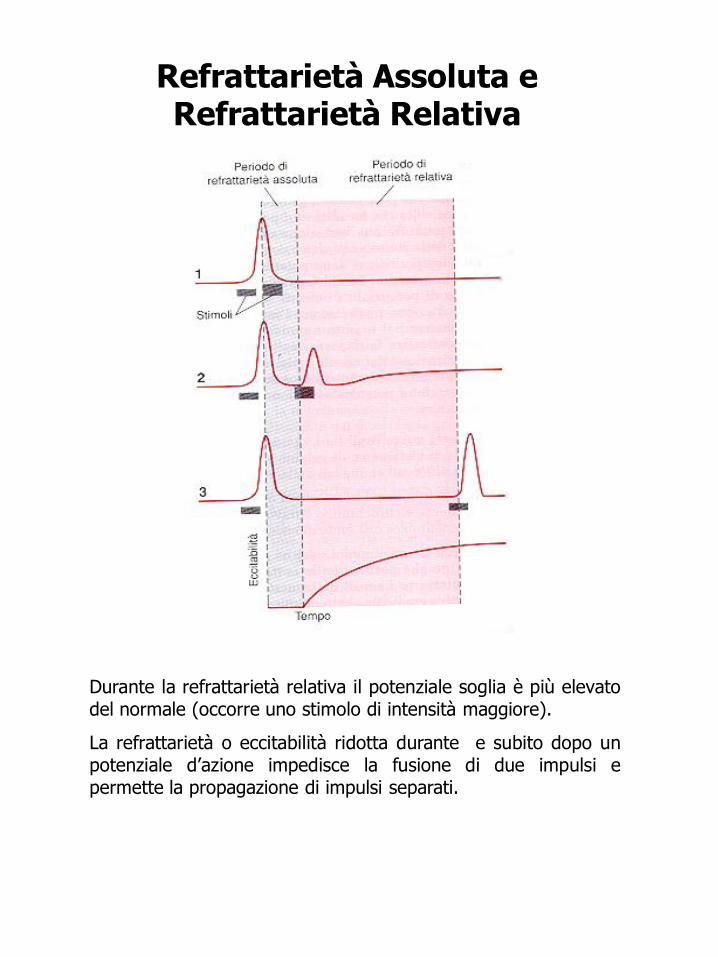
Refrattarietà Assoluta e Refrattarietà Relativa
Durante la refrattarietà relativa il potenziale soglia è più elevato del normale (occorre uno stimolo di intensità maggiore).
La refrattarietà o eccitabilità ridotta durante e subito dopo un potenziale d’azione impedisce la fusione di due impulsi e permette la propagazione di impulsi separati.

PROPAGAZIONE
La propagazione del
potenziale d’azione
avviene grazie a circuiti
locali
L’area della membrana interessata
dal potenziale d’azione ha potenziale
differente dalle regioni limitrofe:
durante il potenziale d’azione si
genera un dipolo elettrico.
La formazione dei dipoli é alla base
della rilevazione dell’attività elettrica
dei tessuti eccitabili con tecniche
non invasive, quali
elettrocardiogramma (EKG),
elettroencefalogramma (EEG),
elettromiogramma (EMG) ed
elettroretinogramma (ERG).


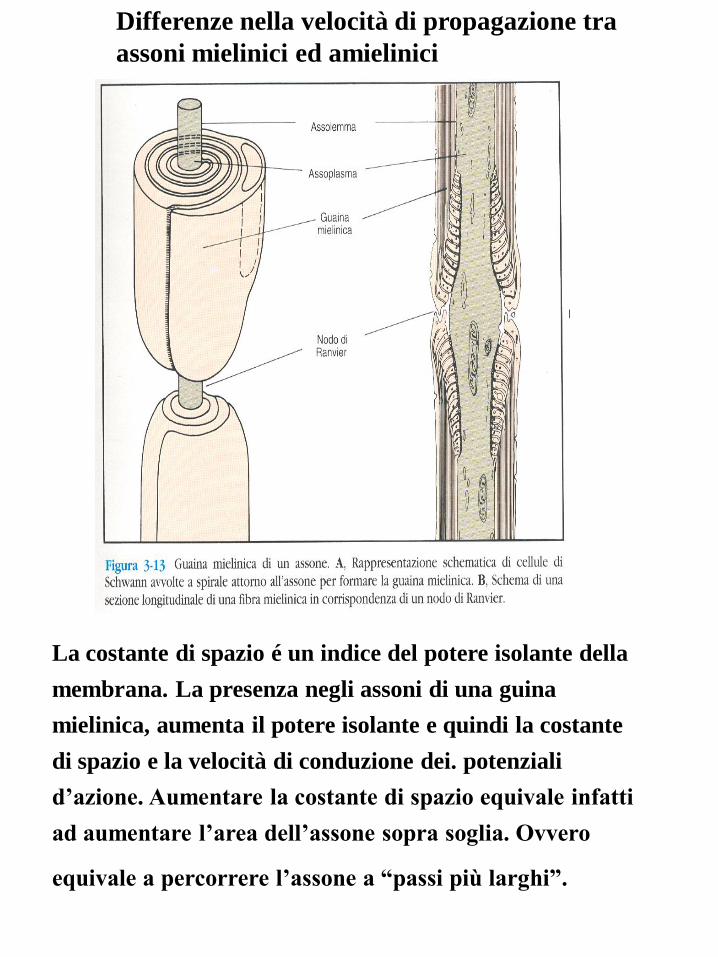
Differenze nella velocità di propagazione tra
assoni mielinici ed amielinici
La costante di spazio é un indice del potere isolante della
membrana. La presenza negli assoni di una guina
mielinica, aumenta il potere isolante e quindi la costante
di spazio e la velocità di conduzione dei. potenziali
d’azione. Aumentare la costante di spazio equivale infatti
ad aumentare l’area dell’assone sopra soglia. Ovvero
equivale a percorrere l’assone a “passi più larghi”.

La presenza di mielina permette di
compensare vistose differenze nei
fattori geometrici (calibro
dell’assone) Nella figura 3-14 si
nota che la velocità di conduzione
delle fibre mieliniche di 7 mm di
diametro (circa 60 m s-1) é oltre 3
volte maggiore delle fibre
amieliniche di 700 mm di diametro.
E’ come se la fibra mielinica avesse
reso il diametro della fibra 9 volte
maggiore (3 = 18/2) della fibra
amielinica (che é invece 100 volte
inferiore).
Differenze nella velocità di conduzione del potenziale
d’azione tra fibre mieliniche ed amieliniche

Basi ioniche del Potenziale d’Azione
Concentrazioni e Potenziali di equilibrio per i principali ioni
La conduttanza della membrana plasmatica per un determinato ione (gx) è una misura della sua permeabilità a quello ione.

Meccanismi di gating per il canale del sodio
Il Modello teorico di Hodgkin e Huxely del potenziale d’azione trova la sua spiegazione nei meccanismi di gating dei canali voltaggio dipendenti

Modalità di Inattivazione dei Canali
I canali inattivati entrano in uno stato refrattario (chiuso) che rende impossibile la loro ulteriore attivazione.

L’ampiezza del potenziale d’azione dipende dal gradiente del Na
Tratto da:
Kuffler, Nicholls & Martin,
“From neuron to brain”
Sinauer 2nd Edition

BASI IONICHE DEL POTENZIALE D’AZIONE
Il citoplasma non é necessario per la genesi del potenziale di azione
Tratto da:
Kuffler, Nicholls & Martin,
“From neuron to brain”
Sinauer 2nd Edition


Vogliamo analizzare la genesi del potenziale d’azione
(PA), cioè misurare la natura, la intensità e la durata
dei flussi ionici durante la sua formazione.
Due problemi da risolvere:
1 - il PA raggiunge il picco in modo rapido
ed autorigenerativo, non dando la possibilità di
determinare le conduttanze ioniche ( V- e T-
dipendenti) ai livelli di potenziale compresi tra il
potenziale di riposo ed il picco del PA,
2 – la continua evoluzione spontanea del PA
impedisce di misurare e isolare i flussi ionici (IR
=V.R) dalle correnti capacitive (IC= Cm . dV/dt).
Quindi è necessario bloccare il meccanismo
autorigenerativo e portare il potenziale di membrana
per il tempo desiderato, ad un qualsiasi livello di
voltaggio (soprasoglia) compreso nell’ambito del PA.
Tutto questo è stato risolto da Hodgkin & Huxley
(1950 ca) con il metodo del blocco di voltaggio.
Utilizzando il circuito che analizzeremo, è infatti
possibile portare e mantenere nel tempo il voltaggio
di membrana ad un valore desiderato mediante il
passaggio di correnti che, appunto, rappresentano
esse stesse la conduttanza ionica (1/Rm) della
membrana a quel determinato voltaggio.
Questo metodo, inoltre, consente di comprimere in
un istante iniziale le correnti capacitive, dando così
la possibilità di osservare il successivo sviluppo delle
correnti resistive, tempo dipendenti.




LA CORRENTE
RAPIDA INIZIALE E’
DOVUTA AD UN
AUMENTO DELLA
PERMEABILITA’ AL
Na+
Tratto da:
Kuffler, Nicholls & Martin,
“From neuron to brain”
Sinauer 2nd Edition





CLASSIFICAZIONE CANALI
I CANALI IONICI SONO CLASSIFICATI IN BASE ALLA:
1) SELETTIVITA’ IONICA (ES.: CANALI SODIO,
POTASSIO, CALCIO, CLORO)
2) DIPENDENZA DAL VOLTAGGIO (ES.: CANALI
SODIO DIPENDENTI DAL VOLTAGGIO E CANALI
SODIO NON DIPENDENTI DAL VOLTAGGIO)
3) PROPRIETA’ FUNZIONALI (Es.: DIFFERENTI
CANALI CALCIO HANNO DIFFERENTE INATTIVAZIO-
NE
3) CONTROLLATI DA TRASMETTITORI E/O
SECONDI MESSAGGERI (ES.: CANALI DIPENDENTI
DA cGMP, cAMP, ATP; CANALI DIPENDENTI DA
GABA, GLICINA, GLUTAMMATO, ACETILCOLINA)
4) CANALI MODULATI DA SECONDI MESSAGGERI (Es.:
CANALI MODULATI DA cAMP, PROTEINE LEGANTI IL
GTP, ETC)
UN CANALE PUO’ APPARTENERE A PIU’ DI UNA
CATEGORIA (Es.: I CANALI CALCIO DIPENDENTI DAL
VOLTAGGIO SONO MODULATI DAL cAMP)
LA CLASSIFICAZIONE DEI CANALI E’ OGGI BASATA
SU DATI MOLECOLARI


















