
Facoltà di Medicina Veterinaria - CORE · staminali mesenchimali da annessi fetali equini ed in...
63
Facoltà di Medicina Veterinaria Dottorato in Medicina Veterinaria SSD – VET/01 Estrazione, isolamento, espansione e caratterizzazione di cellule staminali mesenchimali prelevate da annessi fetali equini Relatori Prof. Elisabetta Giannessi Università di Pisa Dott. Alessandra Coli Università di Pisa Candidata Simona Passeri 2007-2009
Transcript of Facoltà di Medicina Veterinaria - CORE · staminali mesenchimali da annessi fetali equini ed in...

Facoltà di Medicina Veterinaria
Dottorato in Medicina Veterinaria SSD – VET/01
Estrazione, isolamento, espansione e caratterizzazione di cellule staminali mesenchimali prelevate da annessi
fetali equini
Relatori
Prof. Elisabetta Giannessi Università di Pisa
Dott. Alessandra Coli Università di Pisa
Candidata Simona Passeri
2007-2009

- 2 -
I sogni sono fatti di tanta fatica. Forse, se cerchiamo di prendere delle scorciatoie, perdiamo di vista la ragione per cui abbiamo cominciato a sognare e alla fine scopriamo che il sogno non ci appartiene più. Se ascoltiamo la saggezza del cuore il tempo infallibile ci farà incontrare il nostro destino. Ricorda: “Quando stai per rinunciare, quando senti che la vita è stata troppo dura con te, ricordati chi sei. Ricorda il tuo sogno.” (Sergio Bambarén)

- 3 -
Sommario
Riassunto……………………………………………………………………5
Abstract……………………………………………………………………..6
1. Introduzione..………………………………………………………….7 1.1 Gli annessi embrionali………………………………………………..7
1.2 Le cellule staminali…………………………………………………...13
1.3 Le cellule staminali mesenchimali………………………………..14
1.4 Le cellule staminali mesenchimali nel cavallo…………………..19
1.5 Scopo della tesi………………………………………………………….20
2 Materiali e Metodi…………………………………………………...21
2.1 Campioni di cordone ombelicale e di amnios……………………21
2.2 Isolamento ed espansione delle MSCs……………………………..21
2.3 Selezione dei mezzi di coltura………………………………………..22
2.4 CFU-F test (Colony Forming Unit-Fibroblast)…………………..23
2.5 Parametri utilizzati per valutare la cinetica cellulare………….23
2.6 Protocolli di differenziazione…………………………………………24
2.7 Protocollo di citofluorimetria………………………………………...26
2.8 Protocollo di inclusione e di colorazione istologica……………...26
2.9 Protocollo di immunoistochimica……………………………………26

- 4 -
2.10Analisi statistica………………………………………………………….27
3 Risultati……………………………………………………………………...28
3.1 Estrazione ed isolamento delle MSCs………………………………..28
3.2 Cinetica di crescita delle MSCs da gelatina di Wharton………...31
3.3 Cinetica di crescita delle MSCs da amnios………………………….35
3.4 Analisi del differenziamento delle MSCs…………………………...37
3.5 Analisi citofluorimetrica delle MSCs isolate da amnios…………42
3.6 Analisi istochimica ed immunoistochimica delle MSCs………….45
4 Discussione………………………………………………………………….51
Ringraziamenti……………………………………………………………….55
Bibliografia …………………………………………………………………….56

- 5 -
Riassunto
Le cellule staminali mesenchimali sono studiate da molti autori per la capacità di differenziarsi in
cellule della linea mesenchimale (adipociti, condrociti e osteociti) e per le loro proprietà
immunogeniche che le rendono idonee per essere impiegate nella medicina rigenerativa.
Proprio per l’importanza dell’utilizzo di queste cellule a fini terapeutici, lo scopo del lavoro è stato
quello di definire un protocollo di estrazione, isolamento, espansione e caratterizzazione di cellule
staminali mesenchimali da annessi fetali equini ed in particolare da cordone ombelicale (gelatina di
Wharton) e da amnios, evidenziando al contempo la loro localizzazione nei tessuti da cui sono state
estratte.
A tale scopo abbiamo prelevato campioni di cordone ombelicale ed amnios dai quali sono state
estratte cellule mononucleate ed isolate cellule staminali mesenchimali che sono state fatte crescere
su terreni diversi quali MSCBM, αMEM ed αMEM+EGF.
I risultati ottenuti hanno evidenziato una cinetica di crescita simile per le cellule coltivate in
MSCBM e per quelle coltivate in αMEM, mentre l’utilizzo di EGF ha portato ad un incremento
notevole del numero di cellule ottenute ed un numero di passaggi più elevato fino alla senescenza.
L’analisi citofluorimetrica delle MSCs isolate da amnios ha evidenziato la presenza di antigeni di
superficie quali CD44, CD90, CD45 e CD14, mentre l’analisi immunoistochimica su campioni di
tessuto di cordone ombelicale e di amnios è risultata positiva al CD117, CD105 ed Oct-4.
Queste cellule, fatte moltiplicare in mezzi di coltura idonei, si sono differenziate nella linea
condrogenica, osteogenica ed adipogenica confermando la loro natura di cellule staminali
mesenchimali.
Le cellule da noi studiate hanno dimostrato caratteristiche di forte staminalità ed una buona resa
nell’espansione per cui possono ritenersi ottime candidate ad essere usate nella medicina
rigenerativa, con particolare riferimento alle patologie osteoarticolari del cavallo sportivo.

- 6 -
Abstract
Mesenchymal stem cells have been studied by many authors because their can differentiate into
adipocytic, chondrocytic and osteoblastic lineages and because they have immunogenic properties
that make them suitable to be used in regenerative medicine.
The aim of this work is to define a protocol for extraction, isolation, expansion and characterization
of mesenchymal stem cells from equine umbilical cord (Wharton’s jelly) and amnion. Their
localization into the tissues from which they were extracted, was reported.
The mesenchymal stem cells were grown on different media: MSCBM, αMEM and αMEM + EGF.
Our results pointed out that α-MEM and MSCBM media performed similarly in terms of growth
kinetic and EGF supplementation of α-MEM determined a significant increase of the population
doubling value.
The flow cytometric analysis of MSCs isolated from amnion showed the presence of surface
antigens such as CD44, CD90, CD45 and CD14, whereas immunohistochemical analysis on tissue
samples from umbilical cord and amnion was positive for CD117, CD105 and Oct -4 antigens.
These cells, differentiated in adipocytic, chondrocytic and osteoblastic lineages confirming the
nature of mesenchymal stem cells.
The MSCs isolated from equine extra-embryonic tissues, showed characteristics of stem cells and
therefore can be regarded as good candidates to be used in regenerative medicine, with special
reference to osteo-articular diseases of horses.

- 7 -
1. Introduzione
1.1 Gli annessi embrionali
a) GENERALITA’
Gli annessi embrionali (Fig. 1) sono organi particolari che, pur non partecipando alla
costituzione del corpo dell’embrione, svolgono funzioni importantissime quali la protezione
ed il nutrimento, atte a garantire la sopravvivenza dell’embrione stesso.
Sono costituiti dal sacco vitellino, dall’amnios, dal corion, dall’allantoide, dal cordone
ombelicale e dalla placenta; sono tutti presenti solo nei mammiferi euteri o placentati anche
se con modalità di formazione e grado di sviluppo che variano da specie a specie.
Gli annessi si formano dalla espansione extraembrionale dei tre primitivi foglietti
(ectoderma, mesoderma ed endoderma) e sono considerati organi transitori in quanto
destinati a regredire, venendo in parte riassorbiti (sacco vitellino) o abbandonati dal feto a
termine.
Quando la blastocisti è ancora allo stadio didermico e l’area embrionale è pressoché
pianeggiante, dorsalmente a questa si forma l’amnios. Al di sotto dell’area embrionale si
trova la cavità lecitocelica dalla quale prende origine il tubo intestinale primitivo ed il sacco
vitellino. Dalla parte caudale della parete lecitocelica si forma un diverticolo che darà
origine all’allantoide mentre le cellule del trofoblasto andranno a costituire il corion il quale,
nelle parti in cui si pone in contatto morfo-funzionale con la mucosa uterina, costituirà con
questa la placenta. Con il sollevamento dell’embrione e la comparsa delle pieghe craniale,
caudale e laterali si delimita un’apertura ovalare (anello ombelicale primitivo) che va via via
restringendosi ed allungandosi per dare origine al cordone ombelicale che stabilisce il
collegamento tra embrione ed annessi extraembrionali.

- 8 -
Fig. 1: gli annessi embrionali
b) ANNESSI EMBRIONALI DEGLI EQUIDI (Fig.2)
AMNIOS: è l’annesso che avvolge completamente prima l’embrione e poi il feto ed è in
continuità con il cordone ombelicale. La parete è trasparente e sottile ma molto resistente
alla trazione ed è costituita da due foglietti: ectoderma all’interno e mesoderma
somatopleurico all’esterno. Al suo interno è contenuto il liquido amniotico costituito da
acqua, proteine, acido urico, fruttosio e glucosio; quest’ultimo è accumulato nelle
cosiddette papille glicogenetiche o fibroepiteliomi che sono proliferazioni epiteliali della
parete. Le sue funzioni sono quelle di idratare e proteggere il feto, di favorirne i movimenti,
di mantenerne costante la temperatura corporea e di agevolarne la progressione nel canale
del parto. Produce inoltre enzimi proteolitici e lipolitici in grado di dissolvere gli eventuali
embrioni morti durante le prime fasi della gravidanza.
SACCO VITELLINO: è unito all’intestino medio dell’embrione tramite il peduncolo
vitellino e la sua parete è costituita da endoderma all’interno e mesoderma splancnico
all’esterno. Al suo interno è contenuto solo un po’ di liquido salino per cui nei mammiferi
non ha la funzione trofica che invece è caratteristica di altri vertebrati. Le sue funzioni sono
quelle di essere il primo organo emopoietico (splancnopleura) e di dare origine alle cellule

- 9 -
germinali primordiali (endoderma); prima della formazione dell’allantoide, la
splancnopleura contatta il corion per fornirgli i vasi sanguigni, costituendo con esso e la
mucosa uterina un’onfaloplacenta che perdura per circa 14 settimane e poi regredisce in
concomitanza con lo sviluppo dell’allantoide.
ALLANTOIDE: nasce come diverticolo dall’intestino posteriore dell’embrione e,
fuoriuscendo dall’ombelico, si espande nel celoma extraembrionale, avvolgendo
completamente il sacco amniotico, ponendosi a contatto con il corion per fornirgli i vasi
sanguigni (allantocorion) e partecipando alla formazione della placenta (allantoplacenta:
dalla 14a settimana in poi). La sua parete è costituita da endoderma e mesoderma splancnico il
quale darà origine ai vasi allantoidei od ombelicali (due arterie e due vene). Nel suo interno
è contenuto un liquido simile a quello amniotico nel quale sono presenti gli ippomani, masse
rotondeggianti di materiale amorfo e detriti cellulari contenenti mucoproteine e fosfato di
calcio di cui non è nota la funzione.
CORION: è l’annesso più esterno che stabilisce rapporti con la mucosa uterina. La sua
parete, che corrisponde inizialmente al trofoblasto, è costituita da ectoderma all’esterno e
mesoderma somatopleurico all’interno; sulla superficie esterna, inizialmente liscia, si
sviluppano i villi coriali che si diffondono su tutta la superficie del corion contraendo
rapporti con la mucosa uterina e costituendo con essa la placenta.
Fig. 2: gli annessi embrionali degli equidi

- 10 -
PLACENTA: è l’annesso embrionale proprio delle specie vivipare ed è costituito da una
componente fetale che è il corion e da una componente materna che è la mucosa uterina. Ha
molteplici funzioni:
� Protezione: impedisce il passaggio di sostanze o microrganismi nocivi nella
circolazione fetale (barriera placentale).
� Scambi materno-fetali: nutrizione, respirazione ed escrezione dei cataboliti attuata
tramite la permeabilità selettiva del filtro placentare senza che ci sia contatto tra
sangue materno e sangue fetale.
� Funzione endocrina: la placenta produce gonadotropine corioniche, estrogeni
(valutati per la diagnosi di gravidanza fin dall’8a settimana) progesterone, prolattina
e relaxina.
La placenta definitiva degli equidi è una semiplacenta adeciduata (Fig. 3), vascolarizzata
dall’allantoide, con villi distribuiti su tutta la superficie del corion (diffusa-completa) e con
un rapporto tra villi coriali e mucosa uterina di tipo epitelio-coriale. Al parto, il distacco
placentare è molto rapido per la debole aderenza del corion alla mucosa uterina. Si ha prima
la rottura dell’allantocorion che provoca la fuoriuscita del liquido allantoideo e quindi la
rottura dell’amnios con la fuoriuscita del feto.
Fig. 3: schema di semiplacenta adeciduata

- 11 -
CORDONE o FUNICOLO OMBELICALE: annesso che stabilisce il collegamento tra
embrione ed annessi extraembrionali (lunghezza di 80-100 cm e diametro di 2-3 cm). La
parete è costituita da un rivestimento amniotico in vicinanza del corpo dell’embrione (circa i
2/3 della lunghezza), e da un rivestimento allantoideo nel terzo distale. All’interno di questi
rivestimenti troviamo la gelatina di Wharton che è un tessuto connettivo mucoso maturo di
origine mesenchimale la cui funzione è quella di evitare occlusioni dovute a stiramenti o
torsioni dei vasi che percorrono il cordone ombelicale.
Le strutture presenti all’interno del cordone sono:
� Dotto o peduncolo vitellino che va incontro a regressione
� Dotto o peduncolo allantoideo
� Due arterie e due vene vitelline che regrediscono precocemente
� Due arterie e due vene allantoidee od ombelicali. Delle due vene quella sinistra in
vicinanza del corpo embrionale confluisce nella destra che quindi è l’unica ad entrare
nell’embrione.
Fig. 4: schema del cordone ombelicale equino (sx) e foto di arteria e vena ombelicale (dx)

- 12 -
Fig. 5: schema del cordone ombelicale equino (sx, sezione trasversale) ed foto dello stesso (dx)

- 13 -
1.2 Le cellule staminali
Le cellule staminali (SC) sono cellule “non specializzate” che presentano le seguenti
caratteristiche:
� sono capaci di replicarsi per lunghi periodi;
� possono dare origine, in risposta ad opportuni stimoli ed in determinate condizioni
fisiologiche o sperimentali, a progenitori in grado di differenziarsi in uno o più tipi cellulari
maturi diversi;
� si replicano attraverso divisioni cellulari asimmetriche, cioè originano due cellule figlie di
cui una è una cellula staminale uguale alla cellula madre e l’altra è una cellula
maggiormente differenziata, ossia un precursore di una cellula specializzata. (Morrison e
coll., 1997; Lovell-Badge, 2001 Reya e coll., 2001; Donovan e coll., 2001).
Tra le cellule staminali si distinguono cellule staminali embrionali, cellule staminali fetali e
cellule staminali adulte.
Le cellule staminali embrionali derivano dalla massa cellulare interna della blastocisti. Queste
cellule hanno la capacità di replicarsi mediante illimitate divisioni cellulari simmetriche senza
differenziarsi e sono cellule con caratteristiche pluripotenti, cioè in grado di differenziare in tutti i
tipi cellulari derivanti dai 3 foglietti embrionali: mesoderma, endoderma ed ectoderma.
Sono inoltre cellule clonogeniche, in quanto una singola cellula può dare origine a colonie di cellule
geneticamente uguali, ed inoltre esprimono il fattore di trascrizione Oct-4, che mantiene le cellule
staminali embrionali in uno stato di proliferazione ed indifferenziate.
Le cellule staminali adulte sono cellule che si trovano tra le cellule differenziate all’interno
di un tessuto o di un organo. Il ruolo principale delle cellule staminali adulte è quello di mantenere
l’omeostasi e riparare il tessuto in cui si trovano. Tra le cellule staminali adulte sono comprese
quelle fetali, presenti negli abbozzi degli organi, quelle neonatali, isolabili dal cordone ombelicale,
e quelle propriamente adulte. Le cellule staminali adulte sono presenti in molti o forse in tutti gli

- 14 -
organi dei mammiferi, anche se il loro numero diminuisce con l’età. Sono state isolate dalla pelle,
dal muscolo scheletrico, dal fegato, dal tessuto nervoso, dal midollo osseo e dal sangue periferico.
In particolare nel midollo osseo e nel sangue periferico sono state identificate cellule staminali
emopoietiche, precursori endoteliali e cellule staminali mesenchimali.
1.3 Le cellule staminali mesenchimali
Le prime evidenze dell'esistenza di cellule staminali non emopoietiche nel midollo osseo
risalgono al lavoro pionieristico di Friedenstein del 1961, in cui fu dimostrato che cellule derivate
dal midollo osseo potevano differenziare in osteoblasti. Allo stesso autore si deve l’isolamento,
mediante adesione alla plastica, di una popolazione di cellule morfologicamente simile ai
fibroblasti, in grado di formare colonie (CFU-F, Colony Forming Unit-Fibroblast) (Friedenstein e
coll. 1970, 1976 ). Il sistema consisteva nel piastrare il midollo osseo totale su dischi di plastica per
colture cellulari e rimuovere successivamente le cellule non aderenti. Le cellule aderenti così
ottenute erano in maggioranza cellule fusiformi, morfologicamente simili a fibroblasti. Le
osservazioni iniziali di Friedenstein furono confermate poi da altri gruppi intorno agli anni '80
(Ashton e coll. 1980; Castro-Malaspina e coll. 1980). Fu così dimostrata l’esistenza, nel midollo
osseo, di cellule staminali non-emopoietiche definite Cellule Staminali Mesenchimali
(Mesenchymal Stem Cells o MCSs), per la loro capacità di differenziare in cellule di origine
mesodermica (osteoblasti, adipociti, condrociti, cellule del muscolo scheletrico e dei tendini).
La fonte principale di MSCs è il midollo osseo, dove le cellule formano anche lo stroma che
costituisce la nicchia delle cellule staminali emopoietiche. Negli ultimi anni le MSCs sono state
isolate anche da tessuto adiposo, periosto, muscolo scheletrico, liquido amniotico, sangue
periferico, cordone ombelicale, membrana sinoviale, polpa dentale, fegato, tessuti fetali (Docheva e
coll. 2007; Pountos e coll. 2005). Anche se riconosciute tutte come staminali mesenchimali, queste

- 15 -
cellule presentano differenze nella loro capacità proliferativa e differenziativa a seconda dei tessuti
di origine.
Le MSCs non possiedono un fenotipo univoco, esprimono al contrario markers presenti anche
in altri tipi cellulari. Numerosi autori hanno isolato le MSCs da diverse specie animali e da diversi
tessuti, dimostrando nel tempo che mentre alcune molecole vengono espresse, altre sono sempre
assenti.
Uno dei primi lavori sulla fenotipo delle MSCs è stato effettuato da midollo osseo umano nel
1991 da Simmons e coll. che hanno evidenziato la presenza dell’antigene STRO-1, una proteina
della superficie cellulare espressa sulle cellule staminali stromali midollari e sui precursori eritroidi.
Altri antigeni espressi da queste cellule sono l’SH-2 o CD105 (endoglina) e SH-3/SH-4 o CD73 che
sono ecto-5’-nucleotidasi di membrana, (Barry e coll. 2001; Haynesworth e coll. 1992).
Oltre a questi antigeni indicativi di staminalità, sulle MSCs sono espressi anche altri markers
quali, il CD44 molecola di adesione cellulare, il CD90 o Thy-1, il CD 106 (Vascular Cell Adhesion
Molecule o VCAM-1), il CD166 (Activated Leukocyte Cell Adhesion Molecule o ALCAM) ed il
CD29 o β-integrina (Bruder e coll., 1997; Bruder e coll., 1998; Conget e coll., 1999; Pittenger e
coll., 1999; Colter e coll., 2001; Minguell e coll., 2001; Musina e coll., 2005; Miao e coll., 2006).
Il fenotipo delle MSCs viene influenzato da fattori secreti da altre cellule presenti nei
passaggi iniziali durante l’isolamento in vitro, dipende dal grado di differenziamento delle MSCs e
dall’ambiente in cui esse crescono: in vitro l’espressione di alcuni marker non sempre è correlata
con la loro espressione in vivo (Gronthos e coll., 2001; Bruder e coll. 1997).
Esiste, inoltre, una variabilità di espressione dei marker dovuta al tipo di tessuto dal quale le
MSCs vengono isolate, al metodo di isolamento e di coltura ed alle diverse specie animali dalle
quali vengono estratte (Javanzon e coll., 2004; Badoo e coll., 2003). Per esempio, il tessuto adiposo
umano è una valida risorsa per ottenere cellule staminali mesenchimali mediante lipo-aspirazione,
ma, a differenza delle MSCs isolate da midollo osseo, esse sono CD49(+) e CD106(-). Il CD106 nel
midollo osseo è associato al processo di emopoiesi, quindi l’assenza del CD106 sulle MSCs isolate

- 16 -
da tessuto adiposo può essere dovuta alla localizzazione di queste cellule in un tessuto non-
ematopoietico (Zuc e coll., 2002).
Per quanto riguarda variazioni fenotipiche delle MSCs dipendenti da differenze di specie, un
esempio si può ritrovare nel lavoro di Peister e collaboratori (2004) sul topo; sebbene le MSCs di
uomo (Pittenger e coll., 1999; Colter e coll., 2001; Martin e coll, 2002; Musina e coll., 2005)
abbiano mostrato di essere CD34(-) e CD90(+), le MSCs isolate da midollo osseo di topo (Peister e
coll., 2004) esprimevano il CD34(+) e il CD90(-).
Il CD44, invece è sempre espresso sulle MSCs isolate da midollo osseo di specie diverse: infatti è
risultato positivo sulle MSCs isolate da midollo osseo di pecora (Cristino e coll. 2004), di gatto
(Martin e coll, 2002), di topo e ratto (Peister e coll., 2004) e nell’uomo (Conget e coll., 1999;
Pittenger e coll., 1999; Colter e coll., 2001; Musina e coll., 2005; Miao e coll., 2006).
Nella ricerca del fenotipo delle MSCs isolate da midollo osseo, risulta di fondamentale
importanza, anche, escludere l’espressione di alcune molecole di superficie.
E’ necessario infatti escludere la presenza di molecole specifiche delle linee emolinfopoietiche
(Pittinger e coll.,1999; Colter e coll., 2001; Musina e coll., 2005; Miao e coll., 2006). In particolare
il CD34, specifico delle cellule staminali emopoietiche, il CD14, marker specifico di
monociti/macrofagi e il CD45, antigene leucocitario, non devono essere espressi sulle MSCs, per
differenziarle nettamente dalle cellule emopoietiche staminali.
Le MSCs inoltre non esprimono molecole co-stimolatorie quali il CD80, il CD86 e il CD40,
il complesso di istocompatibilità (MHC) di classe II (mentre è sempre espresso in modo più o meno
elevato il MCH di classe I (Le Blanc e coll.2005) e molecole di adesione come il CD31 (Platelet
Endothelial Cell Adhesion Molecule o PECAM-1), CD18 (Leukocyte Function - Associated
Antigen o LFA-1) ed il CD56 (Neuronal Cell Adesion Molecule-1), (Chamberlain, e coll. 2007).
Vari studi descrivono metodi per l’isolamento delle MSCs basati su selezione positiva o
negativa. Nessuno di questi però fornisce una metodica ritenuta valida in assoluto a causa della
mancanza di un marcatore specifico per le MSCs. Il sistema classico utilizzato per isolare le MSCs

- 17 -
parte dalla frazione di cellule mononucleate di midollo osseo e sfrutta la loro capacità di aderire alla
plastica. La frazione delle mononucleate contiene anche cellule emopoietiche in grado di aderire: si
tratta di monociti-macrofagi che aderiscono normalmente più debolmente e che vengono perse
durante i cambi di terreno. Si ottengono quindi cellule fusiformi, simili a fibroblasti, che iniziano a
dividersi e proliferare rapidamente dopo pochi giorni di coltura, con una velocità che dipende molto
dal donatore e dalla densità in cui sono state piastrate. In vitro, le MSCs posso essere espanse
abbastanza facilmente per diversi passaggi, fino al raggiungimento di un blocco proliferativo
dovuto a senescenza.
Una delle caratteristiche funzionali fondamentali delle MSCs è la capacità delle cellule di
differenziare in osteoblasti, adipociti e condrociti, utilizzata molto spesso in saggi in vitro per
l’identificazione delle MSCs (Pittenger e coll. 1999). La metodica classica per il differenziamento
in vitro delle MSCs in osteoblasti prevede l'utilizzo dell’acido ascorbico, ßglicerofosfato e
dexamethasone. Durante la coltura, si osservano cambiamenti dal punto di vista morfologico delle
cellule, che si orientano a formare noduli. Il differenziamento in osteoblasti è accompagnato
dall'aumento di espressione della fosfatasi alcalina, coinvolta nel processo di mineralizzazione.
La formazione dei depositi di calcio può essere osservata tramite colorazione specifica con Alizarin
Red S (ARS) e von Kossa.
Il differenziamento in adipociti richiede dexamethasone, insulina, indometacina e isobutil-
metil26 xantina. Si verifica un accumulo di vacuoli lipidici all'interno delle cellule, che possono
essere osservati tramite colorazione con Oil Red.
Per indurre il differenziamento in condrociti, le MSCs vengono coltivate sotto forma di
pellets in presenza di TGF-ß (Tumor Necrosis Factor- ß) oppure su piastra utilizzando sempre lo
stesso fattore di crescita. (Chamberlain e coll. 2007).
Le MSCs sono state descritte come cellule MHC I+, MHC II-, CD40-, CD80- e CD86-,
profilo definito non-immunogenico, che non richiede immunosoppressione in caso di trapianti
allogenici. Infatti, le molecole MHC di classe I possono attivare linfociti T, che, in mancanza di

- 18 -
molecole co-stimolatorie, restano però anergici. In molti studi le MSCs sono state descritte come
cellule immunosoppressive e immunomodulatorie, in quanto sono in grado di modulare la
proliferazione, il differenziamento e l'attivazione di linfociti T, linfociti B e cellule dendritiche.
Basandosi su questa proprietà immunosoppressiva, sembra che le MSCs possano essere utilizzate
nei trapianti allogenici di cellule staminali emopoietiche per favorire l'attecchimento e diminuire il
rischio di GvHD (Chamberlain e coll. 2007).
In base al potenziale differenziativo, alle caratteristiche immunologiche e alla relativa
facilità con cui possono essere espanse in vitro, le MSCs sono ritenute ottime candidate per diversi
tipi di applicazioni cliniche: trapianti localizzati per danni tissutali, trapianti sistemici per malattie
diffuse, terapia genica ed ingegneria tissutale (Abdallah e coll. 2008).
Vari studi su modelli animali e casi clinici hanno dimostrato l’efficacia dei trapianti di
MSCs per la cura di danni del tessuto osseo e cartilagineo e del miocardio (Orlic e coll. 2001;
Stamm e coll. 2003). Trapianti sistemici di MSCs allogeniche normali sono stati eseguiti in bambini
con osteogenesi imperfetta. In questi studi è stata dimostrata la capacità delle cellule trapiantate di
migrare nell’osso e produrre osteoblasti funzionanti, la cui presenza correla con miglioramenti della
struttura ossea e delle funzioni. In modelli di infarto, trapianti sistemici suggeriscono che le MSCs
sono in grado di migrare e infiltrarsi nel tessuto danneggiato per ripararlo.
Gussoni e coll. (1999) hanno dimostrato la capacità delle MSCs di differenziarsi, in vivo, in
tessuto muscolare scheletrico: se iniettate nel muscolo quadricipite di una cavia affetta da distrofia
muscolare, possono esprimere distrofina in associazione con il sarcolemma delle fibre muscolari.
Nell’area della medicina ortopedica ci sono diversi esempi di applicazioni che coinvolgono
l’impianto locale di cellule di midollo osseo per il trattamento nella riparazione di difetti ossei
segmentali (Quarto e coll. 2001) e da craniotomia (Frebsbach e coll. 1998).
Simili approcci sono stati inoltre descritti per la riparazione di lesioni a livello cartilagineo
(Ponticello e coll. 2000) e tendineo (Young e coll. 1998) . In un animale affetto da osteoartrite con
conseguente lesione all’articolazione del ginocchio, l’utilizzo di cellule staminali mediante

- 19 -
un’iniezione intra-articolare ha avuto come risultato l’adesione di tali cellule sul menisco, sul grasso
patellare e sulla sinovia con rigenerazione del menisco e con miglioramento della cartilagine
(Murphy e coll. 2003).
1.4 Le cellule staminali mesenchimali nel cavallo
Le cellule staminali mesenchimali nel cavallo sono state isolate da midollo osseo, tessuto adiposo,
sangue periferico e sangue da cordone ombelicale (Vidal e coll. 2006, 2007). Cellule staminali sono
state isolate anche dalla gelatina di Wharton da Hoynowski e coll. nel 2007 che le hanno anche
caratterizzate. Gli autori hanno studiato l’espressione fenotipica di queste cellule determinando la
presenza di antigeni propri delle cellule embrionali, come Oct-4, SSEA-4 e c-Kit, e di antigeni tipici
di cellule staminali adulte, come il CD54, CD90, CD105 e CD146; queste cellule possono inoltre
differenziarsi nelle linee condrogenica, adipogenica ed osteogenica.
Una questione rilevante, nella terapia con cellule staminali, è ottenere un prelievo non
invasivo di tessuti insieme ad una fonte abbondante di cellule e un basso livello di contaminazione.
I risultati riportati nel cavallo (Hoynowsky e coll. 2007) e l'uomo (Wang e coll. 2004; Fong e coll.
2007; Secco e coll. 2008; Qiao e coll. 2008) indicano la gelatina di Wharton come una ricca fonte di
cellule staminali mesenchimali la cui raccolta è ovviamente non invasiva. D'altra parte il canale del
parto e ambiente in cui avviene il parto rappresentano una condizione non sterile, soprattutto se
facciamo riferimento alla specie equina.
Il fattore di crescita epidermico (EGF) interviene nel modulare la crescita e la riparazione di
vari tessuti. Dopo il legame con il suo recettore (EGFR), è attiva una pathway intracellulare in
grado di promuovere la migrazione, adesione, proliferazione e la sopravvivenza in vari tipi cellulari
(Lembach e coll. 1976; Wells e coll. 1999; Roux e Blenis 2004). Sia l’ espressione di EGFR e la
sensibilità ad EGF sono stati segnalati in MSCs derivate dal midollo (Gronthos e Simmons, 1995;
Satomura e coll., 1998). Fan e coll. (2008) ha recentemente riportato che EGF è in grado di indurre

- 20 -
l’espressione di EGFR e di promuovere la proliferazione e la migrazione delle MSCs senza effetti
collaterali sulla pluripotenza (Tamama coll., 2006).
1.5 Scopo della tesi
Data l’importanza dell’utilizzo a fini terapeutici delle cellule staminali mesenchimali, lo scopo della
tesi è definire un protocollo di estrazione, isolamento, espansione e caratterizzazione di cellule
staminali mesenchimali da annessi fetali equini ed in particolare da amnios e da cordone
ombelicale, evidenziando al contempo la loro localizzazione nei tessuti da cui sono state estratte.

- 21 -
3 Materiali e Metodi
2.1 Campioni di cordone ombelicale e di amnios
I campioni sono stati prelevati, nel Centro Militare Veterinario di Grosseto e
nell’allevamento Poggetti di Altopascio, da 5 cavalle di età compresa tra i 10 ed i 13 anni che hanno
partorito al termine della gestazione.
Le cellule sono state estratte dalla gelatina di Wharton e dall’amnios ed isolate presso il
laboratorio di Biologia Cellulare della Banca di Cellule e Tessuti dell’UO Immunoematologia II
dell’Azienda Ospedaliera Universitaria pisana (responsabile Dott. Simone Lapi)
Per effettuare un buon prelievo dei campioni, in modo da mantenere vitali le cellule, il
materiale è stato prelevato in breve tempo dalla nascita del puledro, ben lavato con soluzione
fisiologica e posto in un contenitore contenente soluzione fisiologica, 2% di una soluzione di
streptomicina/penicillina e 2% di amfotericina B.
I campioni sono stati successivamente trasferiti in laboratorio per l’estrazione delle cellule.
2.2 Isolamento ed espansione delle MSCs
Prima di estrarre le cellule abbiamo cercato di ridurre al minimo la contaminazione
microbica del materiale conservandolo in una soluzione di RPMI contenente il 5% di streptomicina
e penicillina ed il 2% di amfotericina B a 4°C overnight.
L’amnios, ridotto in piccoli frammenti, e la gelatina di Wharton, ottenuta dopo asportazione
dei vasi dal cordone, sono stati posti separatamente in una soluzione contenente 1 mg/ml di
collagenasi ed incubati per 30 minuti a 37°C. Successivamente il materiale è stato filtrato,

- 22 -
centrifugato, il surnatante scartato e dopo almeno due lavaggi in PBS le cellule estratte sono state
contate con trypan blue 0,4% e seminate.
Il mezzo di coltura utilizzato è stato Alfa MEM con 20% di FCS (Foetal Calf Serum) più
antibiotici e glutammina; le cellule sono state seminate ad una densità di 105 cellule/cm2 in fiasche
da 25 cm2 e poste in incubatore umido a CO2 a 37°C.
Dopo 24 h il mezzo è stato eliminato insieme alle cellule non adese alla fiasca; è stato
aggiunto mezzo fresco e le fiasche sono state riposte nell’incubatore nelle stesse condizioni.
Le cellule mesenchimali estratte dalla gelatina di Wharton e dall’amnios, dal passaggio 1,
sono state seminate ad una densità di 5.000 cellule/cm2 in fiasche da 25 cm2 per determinare la loro
cinetica di crescita. Al 90% di confluenza le cellule sono state tripsinizzate, contate con
l’emocitometro utilizzando il colorante vitale trypan blue e riseminate per un successivo passaggio,
fino alla senescenza.
2.3 Selezione dei mezzi di coltura
Le cellule estratte dalla gelatina di Wharton sono state espanse utilizzando due mezzi di
coltura diversi, α-MEM e MSCSBM.
Le cellule isolate dall’amnios sono state espanse utilizzando α-MEM addizionato di 10%
FCS (FCS e 10% HS (Horse Serum).
α-MEM-medium: mezzo di coltura cellulare tipico composto da α-MEM (Alpha Modified Minimum
Essential Medium ), 100 U/mL penicillina, 100 µg/mL streptomicina , 2 mM L-glutammina.
MSCSBM-medium: Mesenchymal Stem Cells Basal Medium, terreno selettivo per cellule staminali
contenente diversi fattori di crescita non specificati dalla casa produttrice.

- 23 -
Sia per le cellule della gelatina di Wharton che per quelle dell’amnios, al mezzo di coltura α-
MEM è stato aggiunto il fattore di crescita EGF (Epidermal Growth Factor) ad una concentrazione
di 10 ng/mL, per valutare l’efficacia di tale fattore sull’espansione. Le cellule sono state coltivate
sia con α-MEM che con α-MEM più EGF.
2.4 CFU-F test (Colony Forming Unit-Fibroblast)
Per valutare il numero delle cellule mesenchimali nella coltura primaria, le cellule isolate
dalla gelatine di Wharton sono state seminate a 105 cells/cm2 in piastre da 6 pozzetti ed incubate per
8 giorni.
Successivamente le cellule sono state lavate con PBS e colorate con Crystal Violet (0.5%) in
metanolo a temperatura ambiente per 10 minuti, sciacquate due volte con PBS e contate usando un
microscopio a contrasto di fase.
La frequenza delle MSCS sulle cellule totali estratte dalla gelatina di Wharton è stata stimata
dividendo il numero totale di cellule nucleate seminate a passaggio 0 con il numero di colonie
CFU-F contate nella coltura primaria.
2.5 Parametri utilizzati per valutare la cinetica cellulare
I parametri utilizzati per valutare la cinetica delle MSCS sono stati:
• Population Doubling
• Fold Increase
• Doubling Time
Population Doubling: (logNh-logN0)/log2 dove Nh è il numero di cellule ottenute alla fine di
ciascun passaggio e N0 è il numero di cellule iniziali seminate.
Fold Increase: si calcola dividendo il numero di cellule ottenute al 90% di confluenza con il numero
di cellule seminate.

- 24 -
Doubling Time: è la quantità di tempo, misurato in ore, intercorso tra la semina iniziale e la
confluenza del 90% per ciascun passaggio.
2.6 Protocolli di differenziazione
Tre indipendenti esperimenti sono stati condotti con MSCs prese dal passaggio 1 (P1) fino a
passaggio 3 (P3). Le cellule sono state seminate alla densità di 5000 cellule/cm2 in chambers slide
da 4 pozzetti e incubate in α-MEM in modo da permettere l’adesione delle cellule ed il loro
adattamento; successivamente sono stati aggiunti i vari mezzi di differenziamento.
Per la differenziazione osteogenica, le colture cellulari sono state incubate in α-MEM
addizionato con 20% di FCS, 100 U/ml penicillina, 100 µg/ml streptomicina, 2 mM L-glutammina,
20 mM β-glicerolo fosfato, 100 nM desametasone e 250 µM ascorbato 2-fosfato. Il mezzo è stato
cambiato due volte a settimana per tre settimane.
Per la differenziazione adipogenica, le colture cellulari sono state incubate con α-MEM
addizionato con 20% FCS, 100 U/ml penicillina, 100 µg/ml streptomicina, 12 mM L-glutammina,
5 µg/ml insulina, 50 µM indometacina, 1x10-6 M desametasone e 0,5 µM 3-isobutil-1-metilxantina.
Il mezzo è stato cambiato due volte la settimana per tre settimane.
Per la differenziazione condrogenica, le colture sono state incubate per tre settimane in
Chondrocyte Basal Medium addizionato con R3-IGF-1 1mL, bFGF 2.5 ML, transferring 0.5 mL,
insulina bovin 1 M, FBS 25 mL e gentamicina/amfotericina-B 0.5mL.
Per testare l’effetto dell’ EGF sull’abilità differenziativa, le cellule cresciute in α-MEM +
EGF sono state incubate ad una densità di 5000 cell/cm2 in chamber slides da 4 wells utilizzando i
tre mezzi di differenziazione ciascuno addizionato con EGF (10 ng/ml).
L’avvenuta differenziazione cellulare è stata valutata utilizzando opportune colorazioni.

- 25 -
La differenziazione osteoblastica è stata valutata con la colorazione Alizarin Red S.
L’Alizarina determina la formazione di lacche calciche insolubili ed intensamente colorate,
permettendo di mettere in evidenzia la matrice ossea.
Il monostrato di cellule è stato lavato con PBS e fissato con formalina al 10% per 15 minuti. Dopo
lavaggio con PBS, il monostrato è stato coperto con la soluzione di Alizarina, corrispondente a 2g
di Alizarin Red S in 90 ml di acqua. La soluzione dopo 5 minuti è stata rimossa con lavaggi di
acqua deionizzata; in seguito le cellule sono state colorate con ematossilina. Il colorante è stato
rimosso dopo 1 minuto con acqua corrente. La matrice ossea si colora in rosso.
La differenziazione condrogenica è stata valutata mediante colorazione Alcian Blue che
forma ponti salini tra i polianioni dei mucopolisaccaridi acidi.
Il monostrato cellulare è stato lavato con PBS, fissato con alcool per 6 minuti e lavato con PBS. Le
cellule sono state coperte con la soluzione di Alcina Blu (1g di Alcian Blu, 97 ml di acqua distillata,
3 ml di acido acetico, cristalli di Timolo). Il monostrato è stato lavato con acqua corrente e poi con
acqua distillata, coperto con rosso neutro per 1 minuto e lavato con acqua deionizzata. I
glicosaminoglicani si colorano in blu.
La differenziazione adipocitaria è stata evidenziata con colorazione Oil Red O che mette in
evidenza i lipidi solubilizzati dal solvente presente nella soluzione colorante.
Il monostrato di cellule è stato lavato con PBS, fissato con formalina 10% e rimossa con PBS e
successivamente con acqua distillata. Rimossa l’acqua distillata, le cellule sono state lavate con
Isopropanolo 60% per 5 minuti. Il monostrato è stato coperto con la soluzione di lavoro dell’Oil red
O (6:4 Oil red e acqua distillata). I depositi di grasso risultano di colore rosso.

- 26 -
2.7 Protocollo di citofluorimetria
I dati sono stati acquisiti con un citofluorimetro a flusso FACScan (Becton Dickinson)
fornito di laser ad argon con lunghezza d’onda di 488nm, con potenza di emissione di 15mW con i
canali settati per l’acquisizione in scala logaritmica.
Le cellule a P3 sono state staccate, centrifugate e risospese in PBS a una densità di 1x106 cellule/ml
e sono state incubate con anticorpi monoclonali specifici per molecole di superficie nei confronti
di:
� CD14 (Serotec, Oxford, UK)
� CD44 (Chemicon, Temecula, CA, U.S.A.)
� CD45 (Serotec, Oxford, UK)
� CD90 (Santa Cruz Biotechnology, Santa Cruz, CA, U.S.A.)
E’ stato utilizzato come fluorocromo l’isotiocianato di fluresceina (FITC) legato ad
anticorpi secondari di capra anti-IgG di topo. Per ogni campione sono stati raccolti un minimo di
10.000 eventi.
2.8 Protocollo di inclusione e di colorazione istologica
Campioni prelevati da cordone ombelicale e da amnios sono stati disidratati e fissati in
formalina ed inclusi in paraffina.
Dai campioni sono state ottenute sezioni di 10µ successivamente colorate con ematossilina eosina e
Azan Mallory.
2.9 Protocollo di immunoistochimica
Dagli stessi campioni sono state ottenute sezioni di 7µ incubate per 60 minuti con anticorpi
diretti contro i seguenti antigeni:
� CD 105 (Abcam, Cambridge, UK)

- 27 -
� CD 117 (BioLegend, San Diego)
� Oct-4 (Abcam, Cambridge, UK)
Successivamente le sezioni sono stati incubate con anticorpo secondario biotinilato (Vectastain,
Vector Laboratories Inc.) per 10 minuti a temperatura ambiente e con il complesso streptavidina-
perossidasi diluito a 1:250 in PBS per 10 minuti a temperatura ambiente. Per ogni passaggio le
sezioni sono state lavate tre volte con PBS.
NovaRED è stato usato come sistema di rilevazione cromogeno; la reazione enzimatica è stata
monitorata e verificata al microscopio ed i vetrini sono stati infine montate con Vectamount.
2.10 Analisi statistica
L’analisi statistica è stata condotta tramite Graph-Pad Prism software (GraphPad San Diego,
CA, U.S.A.). Sono riportati i valori medi e la deviazione standard. E’ stata effettuato un test
ANOVA a due vie ed il T-Test. Le differenze sono state considerate statisticamente significative
con p<0.05

- 28 -
3 Risultati
3.1 Estrazione ed isolamento delle MSCs
Dalla gelatina di Wharton sono state estratte in media 1.6±0.6 x 107 cellule nucleate da cui
sono state isolate in media 4.1±1.8 x 106 cellule staminali mesenchimali alla fine della coltura
primaria (P0), ottenendo un numero di CFU di 136.7±39.4.
Quindi la frequenza delle MSCs nelle cellule nucleate estratte dalla gelatina di Wharton del cordone
ombelicale equino che è stata di 1:118.
Dall’amnios sono state estratte in media 1.2±0.6 x 107 cellule nucleate da cui sono state
isolate in media 8±3 x 105 cellule staminali mesenchimali a P0. All’inizio della coltura sono stati
evidenziati due tipi cellulari, uno di forma fibroblastoide, riconducibile alle MSCs, e l’altro di
forma rotondeggiante, che scompariva già nei primi passaggi. Nei giorni seguenti sono state
evidenziati cloni di cellule ciascuno dei quali identifica una CFU (unità formanti colonie) (fig 6,7,8)
All’interno delle fiasche le cellule hanno raggiunto la confluenza (fig. 9) in media dopo circa
8 giorni nella coltura primaria (P0).
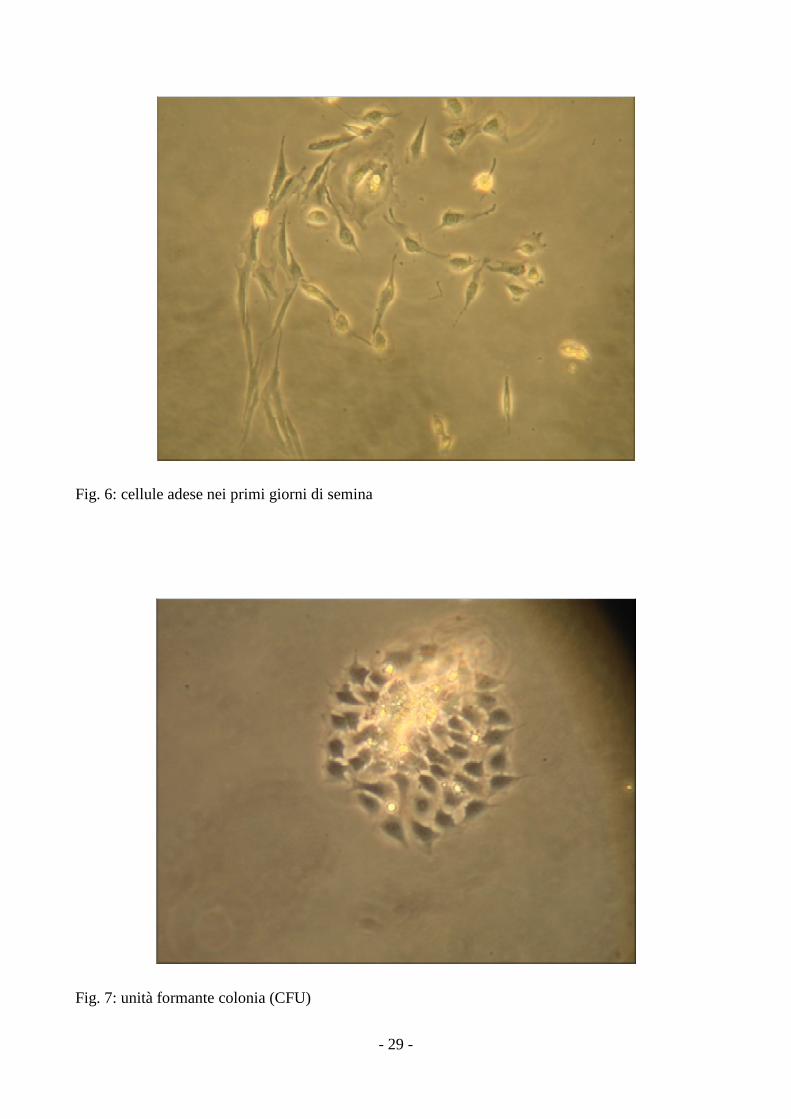
- 29 -
Fig. 6: cellule adese nei primi giorni di semina
Fig. 7: unità formante colonia (CFU)

- 30 -
Fig. 8: CFU fissata e colorata con cristal violetto
Fig. 9: cellule a confluenza

- 31 -
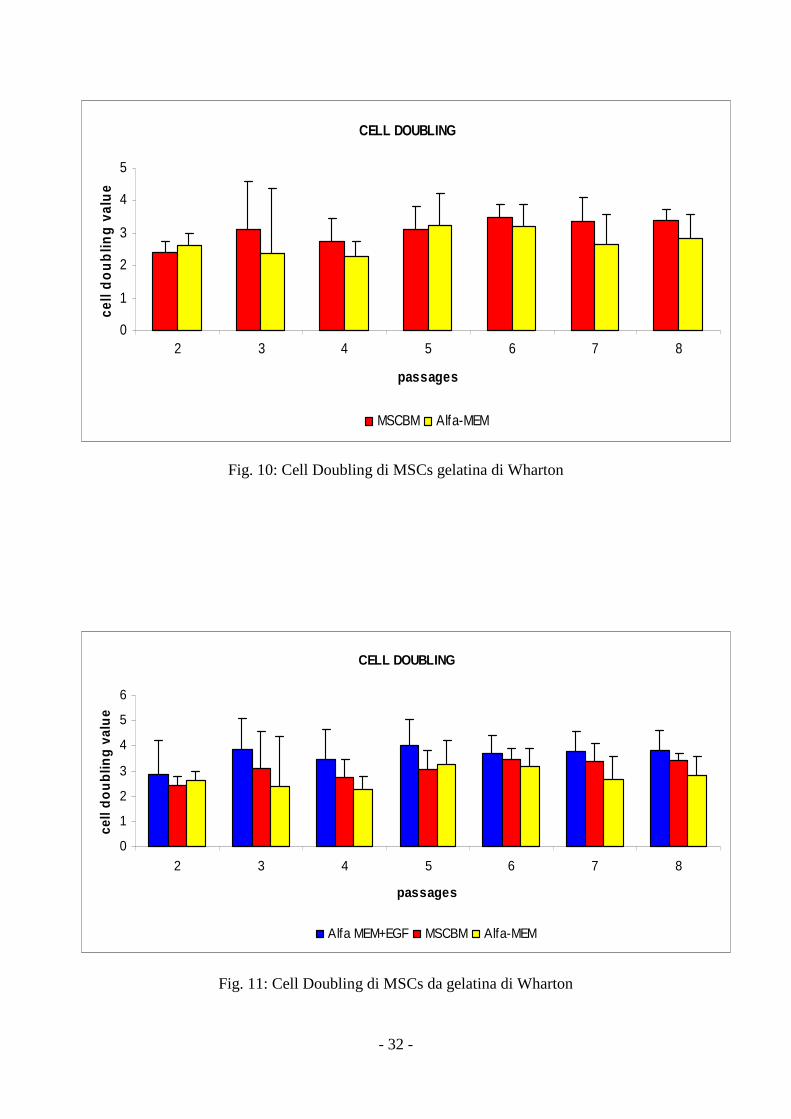
3.2 Cinetica di crescita delle MSCs da gelatina di Wharton Population doubling
L’utilizzo di α-MEM o di MSCBM non mostra differenze statisticamente significative di
questo parametro relativamente a ciascun passaggio, mentre l’aggiunta di EGF ad α-MEM
determina un significativo incremento del valore di Population Doubling (p = 0,007) per ciascun
passaggio (fig 10, 11). Il valore del Population Doubling Cumulativo (PDC) è stato di 36.5±3.4.
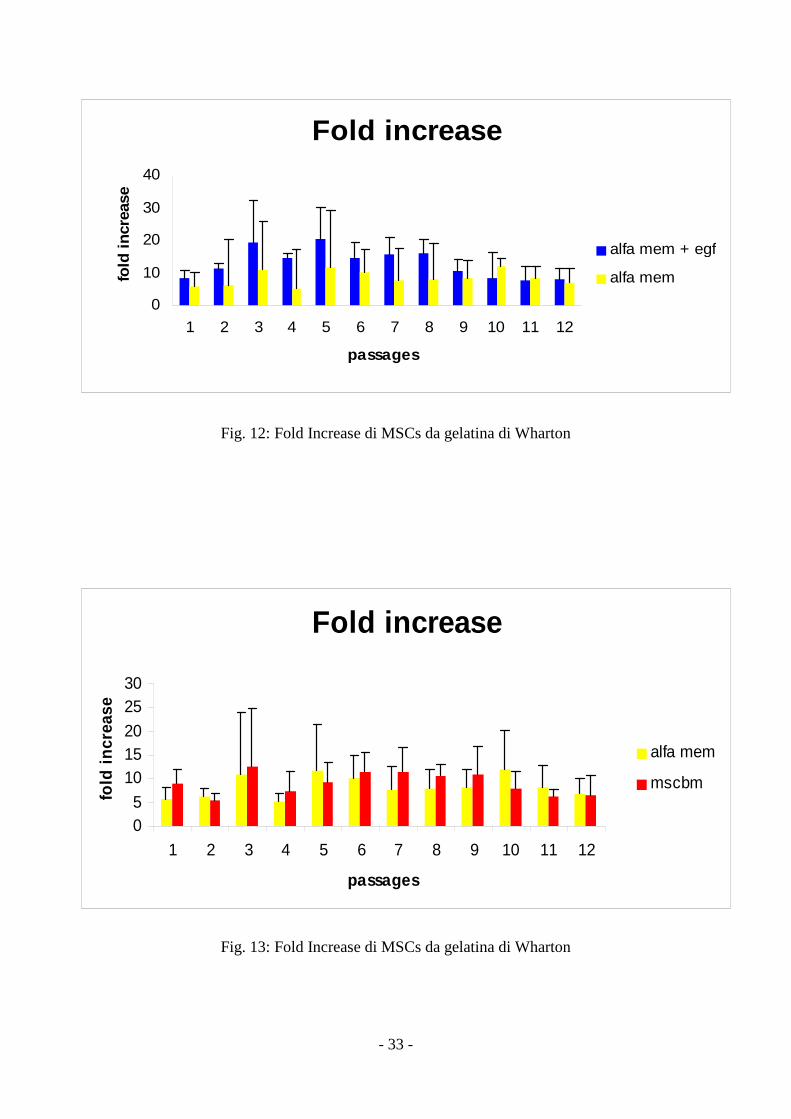
Fold increase
Questo parametro calcolato risulta essere statisticamente più alto (p<0,01) per le cellule
coltivate con α-MEM+EGF rispetto a quelle coltivate solo con α-MEM. Non sono state evidenziate
differenze significative per le cellule coltivate con α-MEM o MSCBM (fig 12, 13).
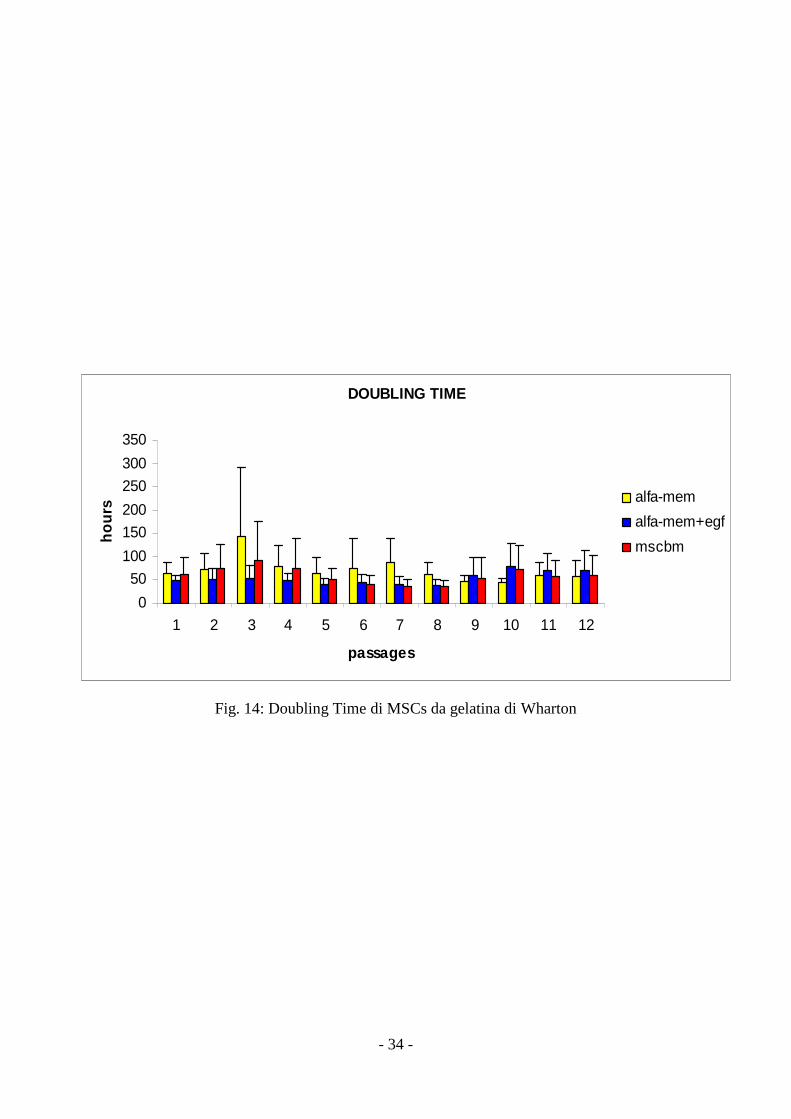
Doubling time
Il parametro, calcolato per ogni mezzo e per ciascun passaggio, mostra variazioni di velocità
di crescita sia al variare del mezzo di coltura utilizzato che al variare del numero di passaggio
analizzato. Gli alti valori di deviazione standard non consentono comunque un’analisi statistica
corretta (fig, 14).
- 32 -
Fig. 10: Cell Doubling di MSCs gelatina di Wharton
Fig. 11: Cell Doubling di MSCs da gelatina di Wharton
CELL DOUBLING
0
1
2
3
4
5
2 3 4 5 6 7 8
passages
cell
do
ub
ling
val
ue
MSCBM Alfa-MEM
CELL DOUBLING
0
1
2
3
4
5
6
2 3 4 5 6 7 8
passages
cell
do
ub
ling
val
ue
Alfa MEM+EGF MSCBM Alfa-MEM
- 33 -
Fig. 12: Fold Increase di MSCs da gelatina di Wharton
Fig. 13: Fold Increase di MSCs da gelatina di Wharton
Fold increase
0
10
20
30
40
1 2 3 4 5 6 7 8 9 10 11 12
passages
fold
in
crea
se
alfa mem + egf
alfa mem
Fold increase
05
101520
2530
1 2 3 4 5 6 7 8 9 10 11 12
passages
fold
in
crea
se
alfa mem
mscbm
- 34 -
Fig. 14: Doubling Time di MSCs da gelatina di Wharton
DOUBLING TIME
0
50
100
150
200
250
300
350
1 2 3 4 5 6 7 8 9 10 11 12
passages
ho
urs
alfa-mem
alfa-mem+egf
mscbm

- 35 -
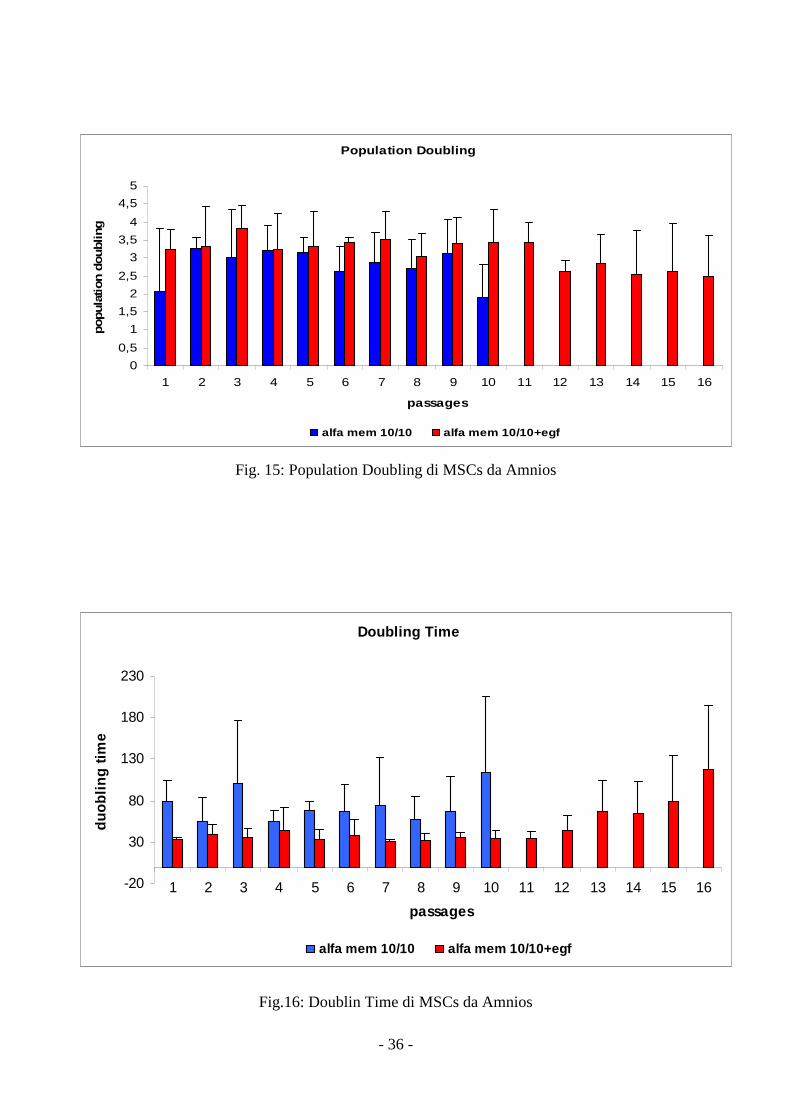
3.3 Cinetica di crescita delle MSCs da amnios
Analizzando le cellule staminali mesenchimali estratte dall’ amnios, abbiamo evidenziato
come EGF addizionato ad α-MEM determini un significativo incremento del valore del parametro
Population Doubling per ciascun passaggio (p=0.005) (fig 15).
Inoltre, l’utilizzo di α-MEM addizionato con 10% di FCS e 10% di HS risulta essere un
ottimo mezzo di coltura in termini di espansione cellulare e di numero di passaggi effettuati.
Il parametro Doubling Time, calcolato per ogni mezzo e per ciascun passaggio, mostra
variazioni di velocità di crescita sia per mezzo di coltura che per numero di passaggio analizzato ma
anche in questo caso gli alti valori di deviazione standard non consentono un’analisi statistica
corretta (fig 16).
- 36 -
Fig. 15: Population Doubling di MSCs da Amnios
Fig.16: Doublin Time di MSCs da Amnios
Population Doubling
0
0,5
1
1,5
2
2,5
3
3,5
4
4,5
5
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
passages
popula
tion d
oubling
alfa mem 10/10 alfa mem 10/10+egf
Doubling Time
-20
30
80
130
180
230
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
passages
du
ob
lin
g t
ime
alfa mem 10/10 alfa mem 10/10+egf

- 37 -
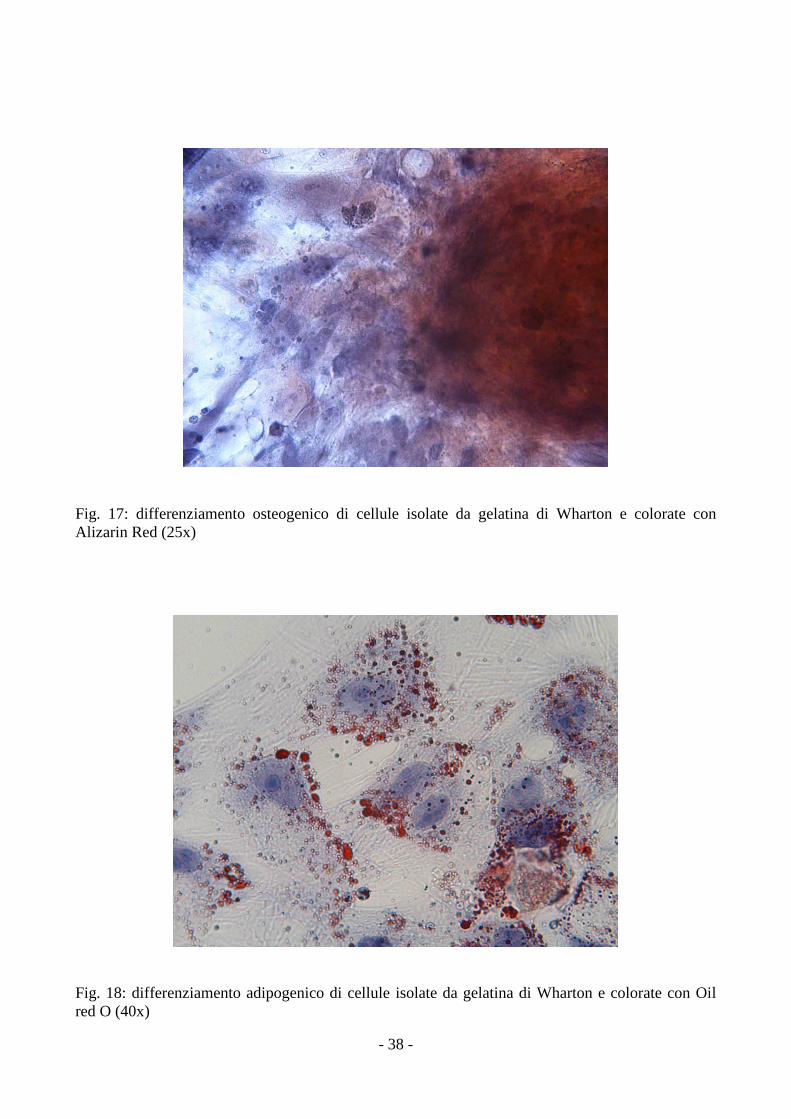
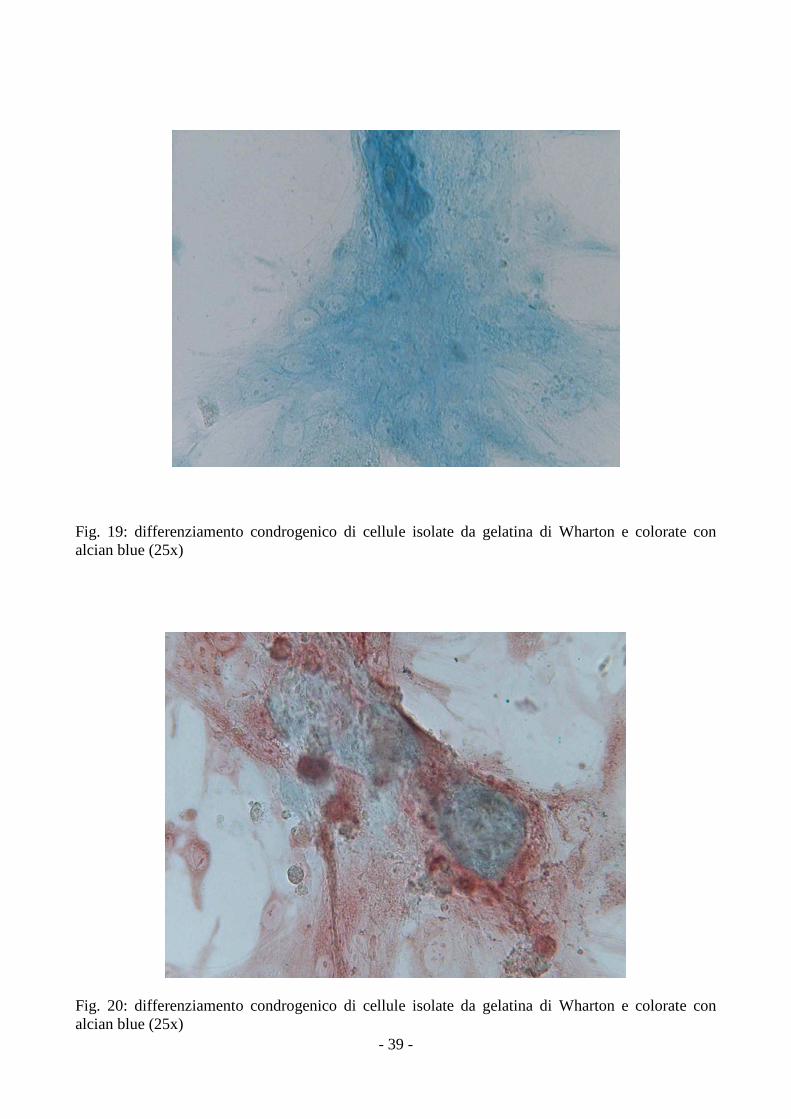
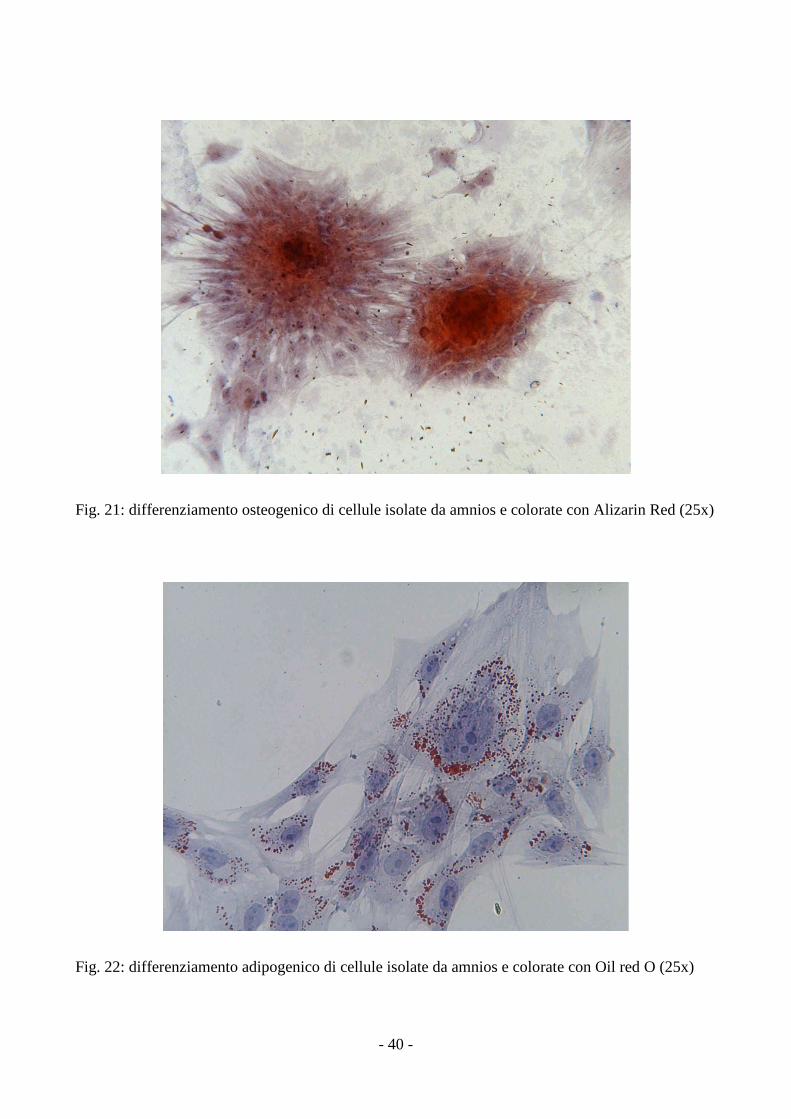
3.4 Analisi del differenziamento delle MSCs
Le MSCs isolate dalla gelatina di Wharton e dall’amnios, coltivate in terreni specifici, si
sono differenziate in senso condrogenico, adipogenico e osteogenico, come dimostrato dalla
positività alle colorazioni con Alcian Blue, Oil Red O e Alizarin Red.
Nel differenziamento adipogenico sono state osservate cellule rotondeggianti che presentavano nel
citoplasma vescicole lipidiche positive alla colorazione con Oil Red O (fig.18, 22); questi granuli
apparivano dopo circa 2 giorni e diventavano molto più numerosi con il tempo.
Nel differenziamento osteogenico è stato osservata una rapida mineralizzazione della matrice con
formazione di noduli ossei, che appaiono di colore rosso con la colorazione Alizarin Red (fig.19,
20, 23, 24) a partire dal quinto giorno, associati ad un accumulo di cellule fibroblastoidi in stretto
contatto l’una con l’altra.
Nel differenziamento condrogenico la presenza di mucopolisaccaridi acidi è stata rivelata dalla
positività all’alcian blue (fig.18, 19).
I risultati ottenuti mostrano che EGF addizionato ai mezzi di differenziazione non produce
nessun effetto sul differenziamento condrogenico ed adipogenico mentre si osserva un incremento
delle dimensioni dei noduli ossei nel differenziamento osteogenico.
- 38 -
Fig. 17: differenziamento osteogenico di cellule isolate da gelatina di Wharton e colorate con Alizarin Red (25x)
Fig. 18: differenziamento adipogenico di cellule isolate da gelatina di Wharton e colorate con Oil red O (40x)
- 39 -
Fig. 19: differenziamento condrogenico di cellule isolate da gelatina di Wharton e colorate con alcian blue (25x)
Fig. 20: differenziamento condrogenico di cellule isolate da gelatina di Wharton e colorate con alcian blue (25x)
- 40 -
Fig. 21: differenziamento osteogenico di cellule isolate da amnios e colorate con Alizarin Red (25x)
Fig. 22: differenziamento adipogenico di cellule isolate da amnios e colorate con Oil red O (25x)

- 41 -
Fig. 23: differenziamento condrogenico di cellule isolate da amnios e colorate con alcian blue (25x)
Fig. 24: differenziamento condrogenico di cellule isolate da amnios e colorate con alcian blue (25x)

- 42 -
3.5 Analisi citofluorimetrica delle MSCs isolate da amnios
L’analisi citofluorimetrica sulle cellule mesenchimali staminali isolate da amnios ha rilevato
la positività agli antigeni di superficie CD 90 e CD 44 ma anche agli antigeni CD 14 ed il CD
45,tipici delle cellule staminali emopoietiche (fig. 26).
L’analisi citofluorimetrica della porzione linfo/monocitaria estratta da un campione di
sangue periferico equino ha confermato la validità degli anticorpi da noi utilizzati (fig. 25).
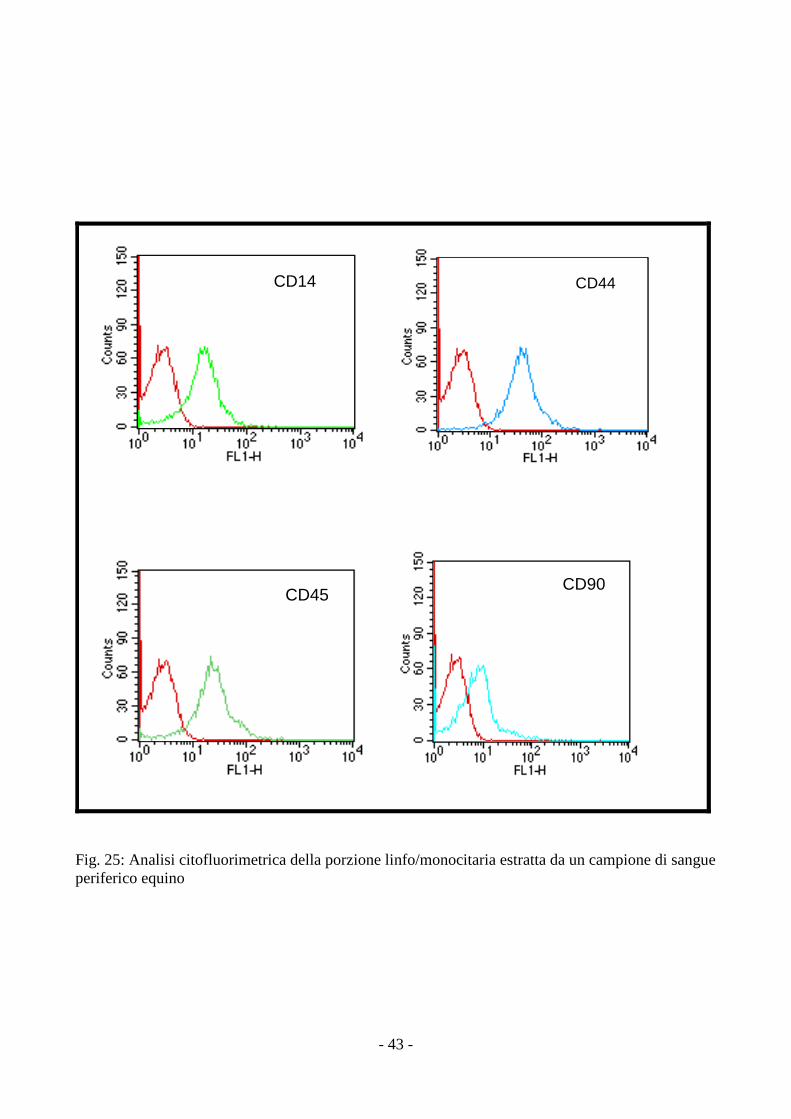
- 43 -
CD14 CD44
CD45CD90
Fig. 25: Analisi citofluorimetrica della porzione linfo/monocitaria estratta da un campione di sangue periferico equino
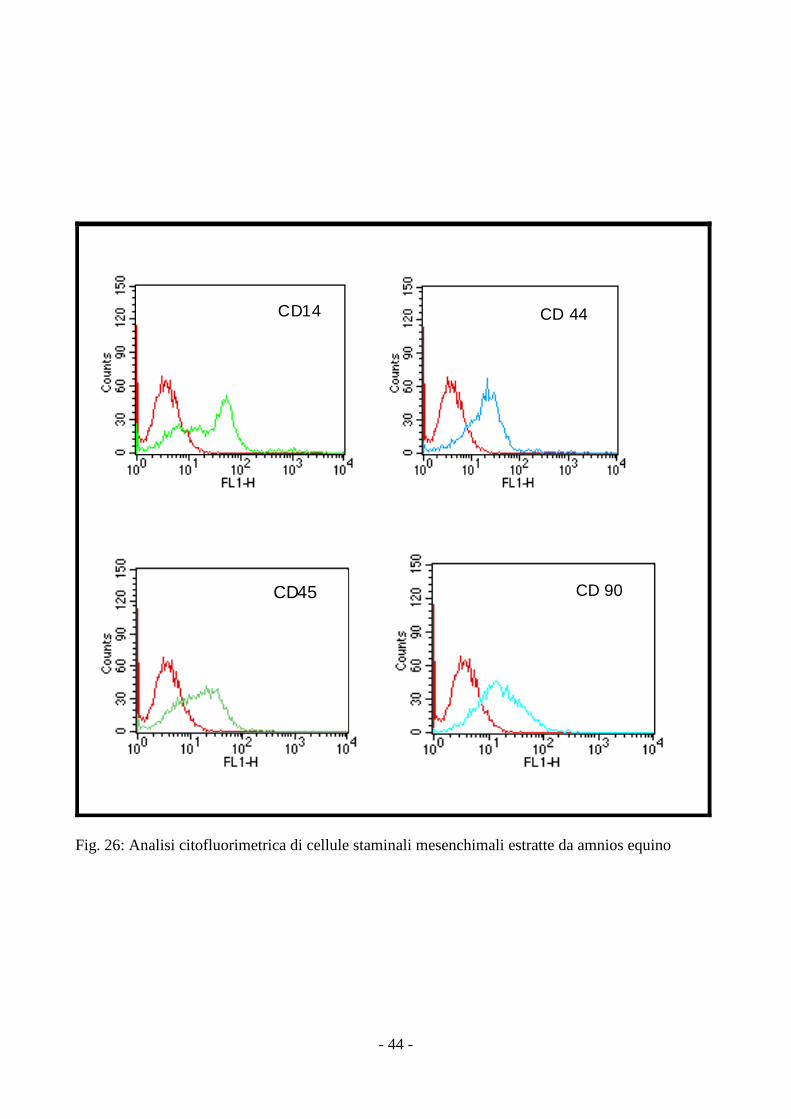
- 44 -
CD14 CD 44
CD45 CD 90
Fig. 26: Analisi citofluorimetrica di cellule staminali mesenchimali estratte da amnios equino

- 45 -
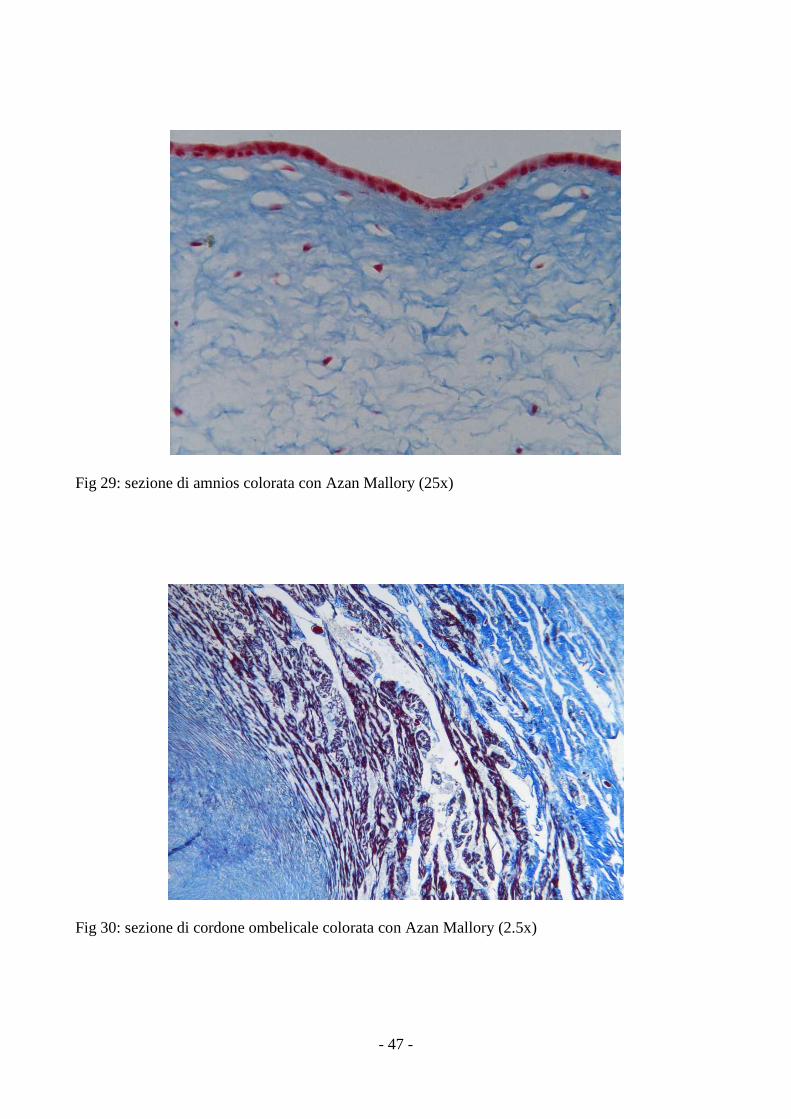
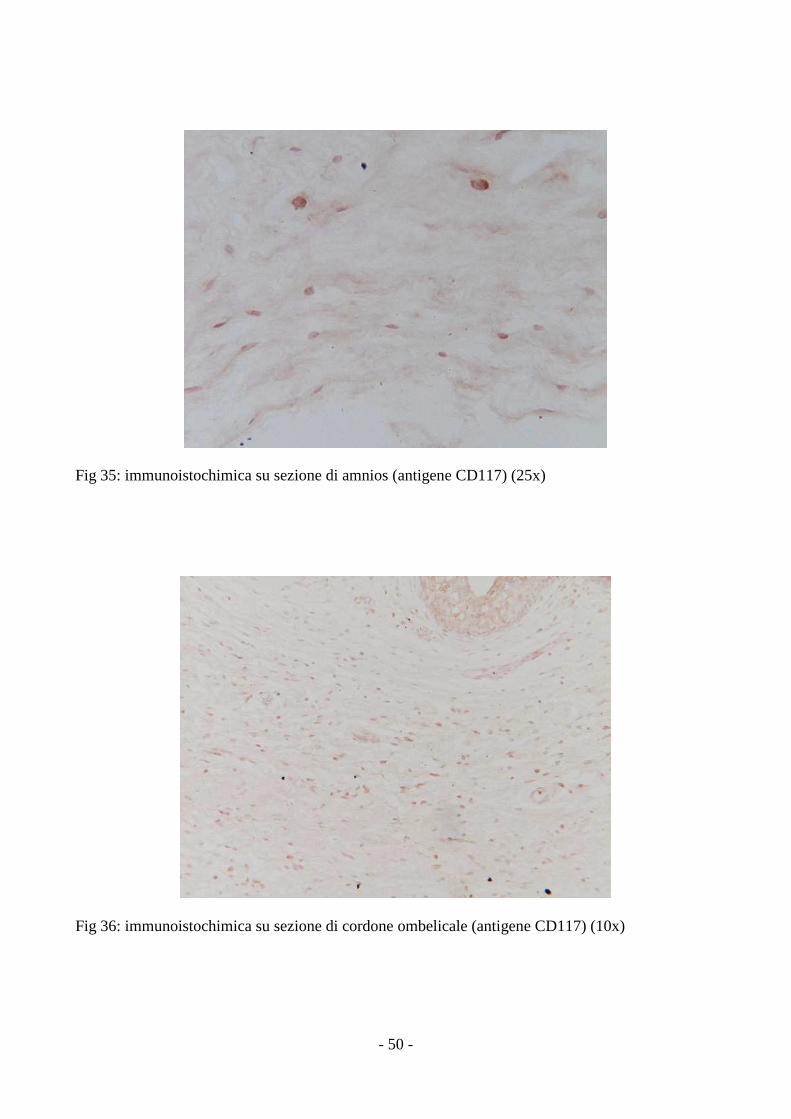
3.6 Analisi istochimica ed immunoistochimica delle MSCs
L’analisi delle sezioni ottenute da cordone ombelicale ed amnios ha rilevato la presenza di
cellule di forma fibroblastoide, nel contesto della componente connettivale, molto più numerose
nella gelatina di Wharton rispetto all’amnios (fig 27, 28, 29, 30).
L’analisi immunoistochimica ha mostrato la positività delle cellule isolate dalla gelatina di
Wharton e dall’amnios a CD117 (antigene importante nella sopravvivenza, proliferazione e
differenziazione cellulare), a CD105 (antigene cellulare associato alla proliferazione endoteliale e
coinvolto nell’angiogenesi) ed a Oct-4 (antigene coinvolto nell’auto-rinnovamento delle cellule
staminali embrionali ovvero nel mantenimento della staminalità) (fig. da 31 a 36).
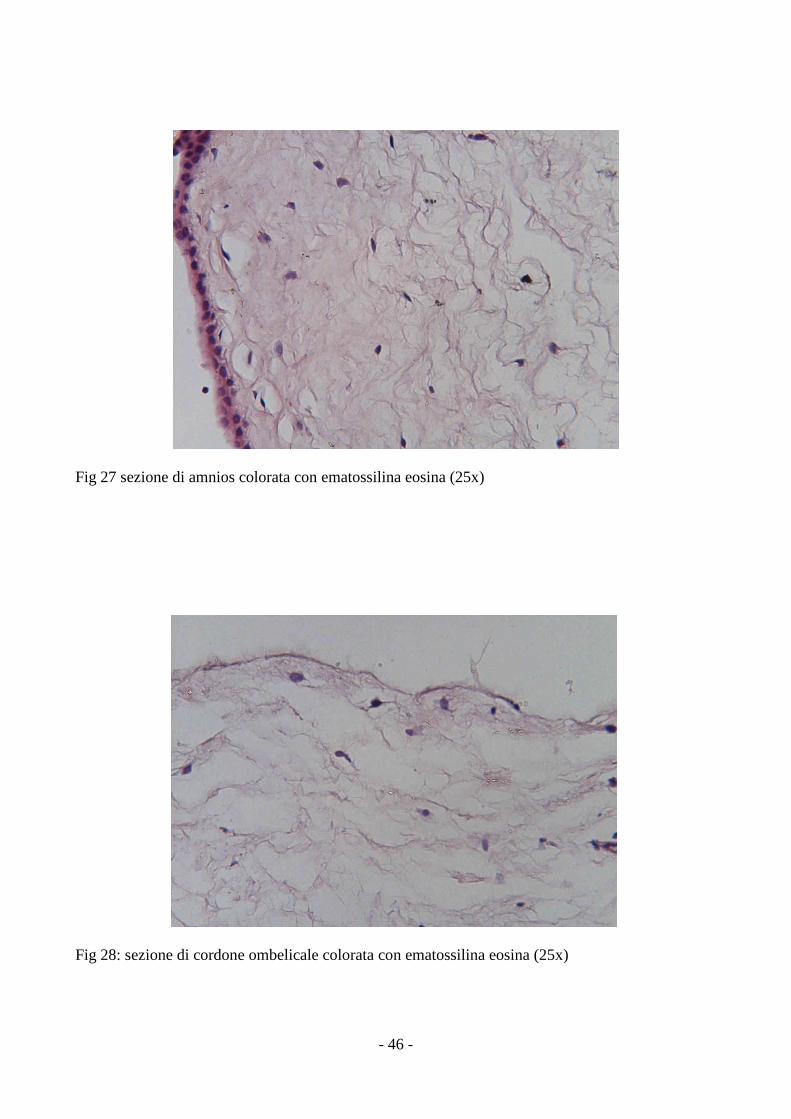
- 46 -
Fig 27 sezione di amnios colorata con ematossilina eosina (25x)
Fig 28: sezione di cordone ombelicale colorata con ematossilina eosina (25x)
- 47 -
Fig 29: sezione di amnios colorata con Azan Mallory (25x)
Fig 30: sezione di cordone ombelicale colorata con Azan Mallory (2.5x)

- 48 -
Fig 31: immunoistochimica su sezione di amnios (antigene CD105) (25x)
Fig 32: immunoistochimica su sezione di cordone ombelicale (antigene CD105) (25x)

- 49 -
Fig 33: immunoistochimica su sezione di amnios (antigene Oct-4) (40x)
Fig 34: immunoistochimica su sezione di cordone ombelicale (antigene Oct-4) (25x)
- 50 -
Fig 35: immunoistochimica su sezione di amnios (antigene CD117) (25x)
Fig 36: immunoistochimica su sezione di cordone ombelicale (antigene CD117) (10x)

- 51 -
4 Discussione
Per ottimizzare l’estrazione delle MSCs dagli annessi fetali, in modo da ridurre al minimo la
contaminazione batterica e funginea, non avendo riferimenti bibliografici in merito, abbiamo messo
a punto un protocollo di campionamento utilizzando una soluzione di RPMI contenente il 5% di
streptomicina e penicillina ed il 2% di amfotericina B a 4°C in cui abbiamo immerso i campioni
overnight.
Lo studio della cinetica di crescita delle MSCs isolate da gelatina di Wharton’s ha
evidenziato una correlazione positiva tra il numero di cellule mononucleate (1.6± 0.6 x 107 ) ed il
numero di CFU (136.7±39.4) che è in accordo con quanto precedentemente riportato da altri autori
(Da Silva Meirelles e Nardi, 2003).
Il Cumulative Population Doubling (CPD) da noi ottenuto (36) risulta inferiore al valore
(50) riportato nello studio di Sarugaser e coll. (2005) da cellule derivate da cordone ombelicale
umano ed a quello osservato da Mitchell e coll. (2003) che riportavano un valore di 80 nella specie
suina. Una possibile spiegazione di queste differenze tra i valori di CPD potrebbe essere in
relazione alle differenti specie animali studiate ed ai mezzi di coltura utilizzati per l’espansione
delle MSCs.
Sono stati testati due differenti mezzi di coltura per la crescita delle MSCs isolate da gelatina
di Wharton: MSCBM, un terreno commerciale addizionato di fattori di crescita tipico per le cellule
mesenchimali staminali, ma molto costoso, e α-MEM, comunemente usato per espandere le MSCs
(Javazon e coll., 2001; Fukuchi e coll., 2004; Smith e coll., 2004).
Dai risultati ottenuti, abbiamo rilevato che la proliferazione cellulare non è influenzata dal
tipo di mezzo di coltura utilizzato (α-MEM vs MSCBM) infatti entrambi i mezzi permettono
performance simili in termini di numero di CFU, population doubling e fold increase.

- 52 -
Questo dato può essere importante per poter utilizzare, durante l’espansione delle MSCs, α-
MEM che è molto meno costoso ma altrettanto efficace di MSCBM.
Per tale ragione per l’espansione delle MSCs isolate dall’amnios abbiamo utilizzato α-
MEM come mezzo di coltura ed i valori relativi all’estrazione delle cellule mononucleate sono stati
di 1.2±0.6 x 107 e quelli relativi all’isolamento delle MSCs sono stati di 8±3 x 105; risultati che
concordano con quelli riportati in letteratura nella specie umana (Toda e coll. 2007).
Per l’espansione delle MSCs estratte sia da gelatina di Wharton che da amnios, l’aggiunta
di EGF ad α-MEM ha portato ad un forte incremento del numero di cellule e dei passaggi ottenuti
fino alla senescenza. Questo risultato è supportato anche dalla letteratura: diversi studi effettuati su
cellule mesenchimali staminali isolate da altre specie (uomo, topo e suino) dimostrano che le MSCs
presentano il recettore per l’EGF. Sarebbe quindi il legame EGF - recettore a stimolare la
migrazione, la proliferazione e la sopravvivenza delle cellule (Krampera e coll. 2005; Tamama e
coll. 2006).
Quando fatte crescere in mezzi di coltura specifici, le MSCs isolate da gelatina di Wharton e
da amnios si sono differenziate nelle linee adipogenica, condrogenica e osteogenica, capacità tipica
delle cellule staminali mesenchimali, indipendentemente dalla fonte di estrazione, come descritto da
diversi autori (Bruder e coll. 1998; Pittenger e coll., 1999; Muraglia e coll. 2000).
Gli effetti dell’aggiunta di EGF nel mezzo di coltura non ha influito sulla loro capacità
differenziativa come riportano anche Tamama e coll. (2006), mentre nel differenziamento
osteogenico l’aggiunta di EGF ha indotto formazione di noduli ossei di maggiori dimensioni.
Per giustificare l’incremento nelle dimensioni dei noduli ossei, dobbiamo ricordare che,
come riportano Sarugaser e coll. (2005), le MSCs se lasciate crescere in un mezzo di coltura non
differenziante, dopo diversi passaggi vanno verso una spontanea differenziazione in senso
osteogenico.

- 53 -
Poiché il fattore EGF stimola la proliferazione cellulare, la sua aggiunta al mezzo di coltura
potrebbe determinare una più veloce differenziazione delle MSCs nella linea cellulare a loro più
favorevole, con conseguente formazione di noduli ossei di dimensioni maggiori.
Lo studio citofluorimetrico del fenotipo delle MSCs isolate da amnios ha evidenziato
l’espressione degli antigeni di superficie CD90, CD44, CD45 e CD14, mentre l’analisi
immunoistochimica su sezioni di cordone ombelicale e di amnios ha dato risultati positivi verso
CD105, CD117(c-Kit) e Oct-4.
Hoyonowski e coll. (2007) hanno isolato e caratterizzato una popolazione di cellule staminali
equine dalla matrice del cordone ombelicale che esprimevano Oct-4, SSEA-4, c-Kit, CD54, CD90,
CD105, CD146. I dati da noi ottenuti sono il linea con quelli ottenuti da questi autori e mettono in
evidenza come queste cellule esprimano antigeni tipici di cellule staminali adulte (CD54, CD90,
CD105, CD146) ma anche di cellule staminali embrionali (Oct-4, SSEA-4, c-Kit).
Lo studio da noi condotto sulle cellule staminali mesenchimali equine vuole essere un
contributo alla possibilità del loro utilizzo nella terapia cellulare in relazione alle loro proprietà
differenziative e caratteristiche immunologiche, come mostrato da altri autori (Wakitani e
coll.1995; Hanada e coll.1997; Mackay e coll.1998; Kopen e coll.1999; Pittinger e coll.1999;
Woodbury e coll.2000; Kohyama e coll.2001; Reyes e coll.2001;Hofstetter e coll.2002;
Knippenberg e coll.2006; Chamberlain e coll. 2007).
In particolare, le cellule staminali mesenchimali isolate da amnios rivestono un ruolo importante per
la loro capacità di differenziarsi in vitro in cellule con caratteristiche simili alle neurogliali, ai
cardiomiociti, osteoblasti, adipociti e condroblasti e di esprimere markers specifici di cellule
staminali embrionali (Oct-4). Queste cellule amniotiche mesenchimali possono essere quindi
definite cellule mesenchimali staminali primitive perché capaci di differenziare in cellule del tessuto
mesodermico ed ectodermico (Kim e coll. 2007).

- 54 -
Le cellule staminali di derivazione amniotica sono inoltre più efficaci nella terapia dei trapianti
allogenici rispetto alle altre cellule staminali adulte per le loro caratteristiche anti-infiammatorie e di
bassa immunogenicità (Toda e coll. 2007; Sakuragawa e coll.1992).
Kobayashi e coll. (2008) hanno isolato dalla componente mesenchimale dell’amnios, una
sottopopolazione negativa sia per MHC I che per MHC II, indicando queste cellule come buone
candidate negli allotrapianti e nella medicina rigenerativa.
Un problema, però, nell’utilizzo terapeutico delle cellule staminali, è la possibilità di ottenerle con
tecniche non invasive per l’estrazione associate ad un’alta resa di cellule isolate.
Attualmente le cellule staminali mesenchimali del cavallo vengono estratte principalmente dal
midollo osseo, dal tessuto adiposo o da sangue periferico; nel primo caso l’estrazione richiede un
intervento invasivo mentre negli altri due casi il numero delle cellule estratte è molto basso.
Questo è stato il motivo per cui nel nostro lavoro abbiamo preso in considerazione i tessuti
extraembrionali del cavallo al momento del parto e dai quali abbiamo isolato cellule con
caratteristiche di forte staminalità e quindi ottime candidate ad essere usate nella medicina
rigenerativa, con particolare riferimento alle patologie osteoarticolari del cavallo sportivo.

- 55 -
Ringraziamenti
Ringrazio sinceramente e con affetto, la dottoressa Francesca Nocchi che mi ha insegnato la
biologia cellulare in laboratorio e mi è stata vicino in questi anni, la dottoressa Roberta Lamanna
che ha lavorato con me anche in condizioni difficili, il dottor Simone Lapi e la dottoressa Patrizia
Urciuoli che hanno dato un contributo rilevante nella stesura ed organizzazione del lavoro ed il
Dottor Fabrizio Scatena , primario dell’unità di Immunoematologia II, che mi ha permesso di
utilizzare le strutture della Banca di Cellule e Tessuti.
Ringrazio, inoltre, il dottor Vincenzo Miragliotta, compagno di stanza, e fondamentale
collaboratore nel lavoro di immunoistochimica e nella stesura dell’articolo pubblicato su Cell
Biology International, la dottoressa Maria Rita Stornelli per l’aiuto nella comprensione dello studio
degli annessi fetali e la dottoressa Maria Carla Iorio per il contributo nello studio citofluorimetrico.
Un ringraziamento speciale va a Stefania Mazzanti fedele compagna nella pausa caffè ed
importante supporto nelle questioni amministrative.
Infine, ma non per ultimi, ringrazio mio marito per la pazienza e l’amore con i quali ha vissuto
questi anni accanto a me e mio figlio per essere nato ed avere portato infinita gioia nella mia vita.

- 56 -
Bibliografia
1. Abdallah BM, Kassem M. Human mesenchymal stem cells: from basic biology to clinical
applications. Gene Ther 2008;15:109-116.
2. Ashton BA, Allen TD, Howlett CR, Eaglesom CC, Hattori A, Owen M. Formation of bone
and cartilage by marrow stromal cells in diffusion chambers in vivo. Clin Orthop Relat Res
1980;151:294–307.
3. Baddoo M., Hill K., Wilkinson R., (2003): Characterization of mesenchymal stem cells
isolated from murine bone marrow by negative selection. J Cell Biochem, 89, 1235-1249.
4. Barry F.P., Murphy M.J. Mesenchymal stem cells: clinical applications and biological
characterization. The International Journal of Biochemistry & Cell Biology 2004;36, 568-
584.
5. Bruder S., Jaiswal N., Hayneswoerth S.E. Growth kinetics, self-renewal and the osteogenic
potential of purified human mesenchymal stem cells during extensive subcultivation
following cryopreservation. Journal of Cellular Biochemistry 1997;64, 278-294.
6. Bruder S., Kurth A., Shea M., Hayes W.C., Jaiswal N., Kadiyala S. Bone regeneration by
implantation of purified, culture-expanded human mesenchymal stem cells. Journal of
Orthopaedic Researche, 1998;16, 155-162.
7. Castro-Malaspina H, Gay RE, Resnick G, Kapoor N, Meyers P, Chiarieri D, McKenzie S,
Broxmeyer HE, Moore MA. Characterization of human bone marrow fibroblast colony-
forming cells (CFU-F) and their progeny. Blood 1980;56:289–301.
8. Chamberlain G, Fox J, Ashton B, Middleton J. concise review: mesenchymal stem cells:
their phenotype, differentiation capacity, immunological features, and potential for homing.
Stem Cells 2007;25:2739–2749.

- 57 -
9. Colter D.C., Sekiya I., Prockop D.J. Identification of a subpopulation of rapidilly self-
renewing and multipotential adult stem cells in colonies of human marrow stromal cells.
Proc Natl Acad Sci (USA) 2001;98, 7841-7845.
10. Conget P.A., Minguell J.J. Phenotypical and functional properties of human bone marrow
mesenchymal progenitor cells. J Cell Physiol, 1999;181, 63-73.
11. Cristino S., Grassi., Toneguzzi S., Piacentini A., Grigiolo B. Analysis of mesenchymal stem
cells grown on a three-dimensional HYAFF 11-based prototype ligament scaffold.
Interscience 2004; 276-282.
12. Docheva D, Popov C, Mutschler W, Schieker M. Human mesenchymal stem cells in contact
with their environment:surface characteristics and the intergin system. J Cell Mol Med
2007;11:21 38.
13. Donovan P.J., Gearhart J. The end of the beginning for pluripotent stem cells. Nature
2001;41, 492-97.
14. Fan H, Hu Y, Qin L et al. Porous gelatin-chondroitin-hyaluronate tri-copolymer scaffold
containing microspheres loaded with TGF-beta1 induces differentiation of mesenchymal
stem cells in vivo for enhancing cartilage repair. J. Biomed. Mater. Res. A. 2006; 77,785-94.
15. Fong CY, Richards M, Manasi N, Biswas A, Bongso A. Comparative growth behaviour and
characterization of stem cells from human Wharton's jelly. Reprod Biomed Online. 2007;15:
708-718.
16. Friedenstein AJ, Chailakhajan RK, Lalykina KS. The development of fibroblast colonies in
monolayer cultures of guinea-pig bone marrow and spleen cells. Cell Tissue Kinet
1970;3:393-403.
17. Friedenstein AJ, Gorskaja JF, Kulagina NN. Fibroblast precursors in normal and irradiated
mouse hematopoietic organs. Exp Hematol 1976;4:267–274.
18. Friedenstein AJ. Osteogenic activity of transplanted transitional epithelium. Acta Anat
(Basel) 1961;45:31-59.

- 58 -
19. Fukuchi Y, Nakajima H, Sugiyama D, Hirose I, Kitamura T, Tsuji K. Human placenta-
derived cells have mesenchymal stem/progenitor cell potential. Stem Cells 2004;22:649-
658.
20. Gronthos S, Simmons PJ. The growth factor requirements of STRO-1- positive human bone
marrow stromal precursors under serum-deprived conditions in vitro. Blood 1995;85:929 –
940.
21. Gronthos S., Franklin D.M., Leddy H.A., Robey P.G. Surface protein characterization of
human adipose tissue-derived strom cells,Journal of Cellular Physiology 2001;189, 54-63.
22. Gussoni E., Soneoka Y., Strickland C.D., Buzney E.A., Khan M.K., Flint A.F., Kunkel
L.M., Mulligan R.C. Dystrophin expression in the mdx mouse restored by stem cell
transplantation, Nature1999;401, 390-394.
23. Hanada K, Dennis JE, Caplan AI. Stimulatory effects of basic fibroblast growth factor and
bone morphogenetic protein-2 on osteogenic differentiation of rat bone marrow-derived
mesenchymal stem cells. J Bone Miner Res 1997; 12: 1606-1614.
24. Haynesworth S.E., Baber M.A., Caplan A. Cell surface antigens on juman marrow-derived
mesenchymal cells are detected by monoclonal antibodies. Bone, 1992;13, 69-80.
25. Hofstetter CP, Schwarz EJ, Hess D et al. Marrow stromal cells form guiding strands in the
injured spinal cord and promote recovery. Proc Natl Acad Sci U S A 2002; 99: 2199-2204.
26. Hoynowski S M, Fry M M, Gardner B M, Leming M T, Tucker J R, Black L, Sand T,
Mitchell KE. Characterization and differentiation of equine umbilical cord-derived matrix
cells. Biochem Biophys Res Commun. 2007;362:347–353.
27. Javazon EH, Colter DC, Schwarz EJ, Prockop DJ. Rat marrow stromal cells are more
sensitive to plating density and expand more rapidly from single-cell-derived colonies than
human marrow stromal cells. Stem Cells 2001;19:219-225.
28. Kim J, Kang HM, Kim H et al. Ex vivo characteristics of human amniotic membrane-
derived stem cells. Cloning and stem cells 2007; 9: 581-594.

- 59 -
29. Knippenberg M, Helder MN, Zandieh DB et al. Osteogenesis versus chondrogenesis by
BMP-2 and BMP-7 in adipose stem cells. Biochem Biophys Res Commun 2006; 342: 902-
908.
30. Kobayashi M, Yakuwa T, Sasaki K, Sato K, Kikuchi A, Kamo I, Yokoyama Y, Sakuragawa
N. Multilineage potential of side population cells from human amnion mesenchymal layer.
Cell Transplant. 2008;17(3):291-301.
31. Kohyama J, Abe H, Shimazaki T et al. Brain from bone: Efficient meta-differentiation of
marrow stroma-derived mature osteoblasts to neurons with Noggin or a demethylating
agent. Differentiation 2001; 68: 235-244.
32. Kopen GC, Prockop DJ, Phinney DG. Marrow stromal cells migrate throughout forebrain
and cerebellum, and they differentiate into astrocytes after injection into neonatal mouse
brains. Proc Natl Acad Sci U S A 1999; 96: 10711-10716.
33. Krampera M, Pasini A, Rigo A, Scrupoli MT, Tecchio C, Malpeli G, Scarpa A, Dazzi F,
Zizzolo G, Vinante F. HB-EGF/HER-1 signaling in bone marrow mesenchymal stem cells:
inducing cell expansion and reversibly preventing multilineage differentiation. Blood
2005;106:59-66.
34. Le Blanc K., Trammik C., Rosendahl K., Zetterberg E., Ringden O. HLA expression and
immunologic properties of differentiated and undifferentiated mesenchymal stem cells,
Experimental Hematology, 2003;31, 890-896.
35. Lembach KJ. Induction of human fibroblast proliferation by epidermal growth factor (EGF):
Enhancement by an EGF-binding arginine esterase and by ascorbate. Proc Natl Acad Sci U
S A 1976;73:183–187.
36. Lovell-Badge R. The future for stem cell research. Nature 2001;414, 88-90.
37. Mackay AM, Beck SC, Murphy JM et al. Chondrogenic differentiation of cultured human
mesenchymal stem cells from marrow. Tissue Eng 1998; 4: 415-428.

- 60 -
38. Martin D.R., Cox N.R., Hathcock T.L., Niemeyer G.P., Backer H.J. Isolation and
characterization of multipotential mesenchymal stem cells from feline bone marrow, Exp.
Hematol. 2002;30, 879-886.
39. Miao Z., Jin J., Chen L., Zhu J., Huang W., Zhao J., Quain X., Zhang X., (2006): Isolation
of mesenchymal stem cells rfom human placenta: comparison with human bone marrow
mesenchymal stem cells. Cell Biology International, 30, 681-687.
40. Miguell J.J., Erices A., Conget P. Mesenchymal stem cells. Exp. Biol. Med. 2001;226(6),
507-520.
41. Mitchell KE, Weiss ML, Mitchell BM, Martin P, Davis D, Morales L, Helwig B,
Beerenstrauch M, Abou-Easa K, Hildreth T, Troyer D. Matrix cells from wharton’s jelly
form neurons and glia, Stem Cells 2003;21:50–60.
42. Morrison S.J., Shah N. M., Anderson D.J. Regulatory mechanisms in stem cell biology. Cell
1997;88, 287-298.
43. Muraglia A, Cancedda R, Quarto R. Clonal mesenchymal progenitors from human bone
marrow differentiate in vitro according to a hierarchical model. J Cell Sci. 2000;113:1161-
1166.
44. Murphy M., Fink D.J., Hunziker E.B., Barry F.P. Stem cell therapy in a caprine model of
osteoartheritis, Arthritis Rheumatism, 2003;48, 3464-3474.
45. Musina R.A., Bekcchanova E.S., Belyavskii A.V., Sukhikh G.T. Differentiation potential of
mesenchymal stem cells of different origin. Cell Technologies in biology and
medecine,2005; 2, 147-151.
46. Orlic D., Kajstura J., Chimenti S., Jakoniuk I., Anderson S.M., Li B., Pickel J., McKay R.,
Nadal-Ginard B., Bodine D.M., Leri A., Anversa P. Bone marrow cells regenerate infarcted
myocardium. Nature 2001;410, 701-705.

- 61 -
47. Peister A., Mellad J.A., Larson B.L., Hall B.M., Gibson L.F., Prockop D. Adult stem cells
from bone marrow (MSCs) isolated from different strains of inbred mice vary in surface
epitopes, rates of proliferation, and differentiation potential. Blood, 2004;103, 1662-1668.
48. Pittenger M.F., Mackay A.M., Beck S.C., Jaiswal R.K., Douglas R., Mosca J.D.,. Moorman
M.A,. Simonetti D.W, Craig S., Marshak D.R. Multilineage potential of adult mesenchymal
stem cells, Science 1999; 284,143-147.
49. Ponticello S.M., Schinagl R.M., Kadiyala S, Barry F.P. Gelatin-based resorbable sponge as
a carrier matrix for juman mesenchymal stem cells in cartilage regeneration therapy, Journal
of Biomedical Materials Research, 2000;52, 246-255.
50. Pountos I, Giannoudis PV. Biology of mesenchymal stem cells. Injury 2005;36S:S8-S12.
51. Qiao C, Xu W, Zhu W, Hu J, Qian H, Yin Q, Jiang R, Yan Y, Mao F, Yang H, Wang X,
Chen Y. Human mesenchymal stem cells isolated from the umbilical cord. Cell Biol Int.
2008;32:8-15.
52. Quarto R., Mastrogiacomo M., Cancedda R., Kutepov S.M., Mukhachev V., Lavroukov A.,
Kon E., Marcacci M., (2001): Repair of large bone defects with the use of autologous bone
marrow stromal cells, The New England Journal of Medecine, 344, 385-386.
53. Reya T., Morrison S.J., Clarke M.F, Weissman I. Stem cells, cancer, and cancer stem cells.
Nature 2001;414, 105-111.
54. Reyes M, Lund T, Lenvik T et al. Purification and ex vivo expansion of postnatal human
marrow mesodermal progenitor cells. Blood 2001; 98: 2615-2625.
55. Roux PP, Blenis J. ERK and p38 MAPK-activated protein kinases: A family of protein
kinases with diverse biological functions. Microbiol Mol Biol Rev 2004;68:320 –344.
56. Sarugaser R, Lickorish D, Baksh D, Hosseini M Morris, Davies J E. Human Umbilical Cord
Perivascular (HUCPV) Cells:A Source of Mesenchymal Progenitors. Stem Cells 2005;23:
220–229.

- 62 -
57. Secco M, Zucconi E, Vieira NM, Fogaça LL, Cerqueira A, Carvalho MD, Jazedje T,
Okamoto OK, Muotri AR, Zatz M. Multipotent stem cells from umbilical cord: cord is
richer than blood! Stem Cells 2008;26:146-150.
58. Smith JR, Pochampally R, Perry A, Hsu S, Prockop DJ. Isolation of a highly clonogenic
and multipotential subfraction and adult stem cells from bone marrow stroma. Stem Cells
2004;22: 823-31.
59. Stamm C., Westphal B., Klein H.D., Petzsch M., Kittner C., Klinge H., Schumichen C.,
Nienaber C.A., Freund M., Steinhoff G. Autologous bone-marrow stem cell transplantation
for myocardial regeneration, Lancet 2003;361, 45-46.
60. Tamama K, Fan VH, Griffith LG, Blair HC, Wells A. Epidermal Growth Factor as a
Candidate for Ex Vivo Expansion of Bone Marrow – Derived Mesenchymal Stem Cells.
Stem Cells 2006;24:686-695.
61. Toda A, Okabe M, Yoshida T, Nikaido T. The potential of amniotic membrane/amnion-
derived cells for regeneration of various tissues. J Pharmacol Sci. 2007 Nov;105(3):215-28.
62. Vidal MA, Kilroy GE, Johnson JR, Lopez MJ, Moore RM, Gimble JM. Cell growth
characteristics and differentiation frequency of adherent equine bone marrow-derived
mesenchymal stromal cells: adipogenic and osteogenic capacity. Vet Surg. 2006;35:601-
610.
63. Vidal MA, Kilroy GE, Lopez MJ, Johnson JR, Moore RM, Gimble JM. Characterization of
equine adipose tissue-derived stromal cells: adipogenic and osteogenic capacity and
comparison with bone marrow-derived mesenchymal stromal cells. Vet Surg. 2007;36:613-
22.
64. Wakitani S, Saito T, Caplan AI. Myogenic cells derived from rat bone marrow
mesenchymal stem cells exposed to 5-azacytidine. Muscle Nerve 1995; 18: 1417-1426.

- 63 -
65. Wang HS, Hung SC, Peng ST, Huang CC, Wie HM, Guo YJ, Fu YS, Lai MC, Chen CC.
Mesenchymal stem cells in the Wharton's jelly of the human umbilical cord. Stem Cells
2004;22:1330-1337.
66. Wells A. EGF receptor. Int J Biochem Cell Biol 1999;31:637– 643.
67. Woodbury D, Schwarz EJ, Prockop DJ et al. Adult rat and human bone marrow stromal
cells differentiate into neurons. J Neurosci Res 2000; 61: 364-370.
68. Young R.G., Butler D.L. Weber W., Caplan A.I., Gordon S.L., Fink D.J. Use of
mesenchymal stem cells in a collagen matrix for Achilles tendon repair, Journal of
Orthopaedic Research ,1998;16, 406-413.


















