
carlavirus - francescofiume.altervista.org · pathway di taglio di Dicer e determina...
26
Fiume Francesco Genere Carlavirus Classificazione Gruppo IV (+ssRNA) Ordine Tymovirales Famiglia Betaflexiviridae Genere Carlavirus Le caratteristiche sintetiche del genere Carlavirus sono: • Proprietà delle particelle virali nella linfa: TIP (thermal inactivation point; la più bassa temperatura alla quale l’infettività della linfa è distrutta entro un’ora): 50-68,5-85 °C. LIV (longevity in vitro; longetività dell’infettività della linfa in vitro): 0,5-4,8-2,01 giorni. DEP(diluition end point; punto finale di diluizione dell’infettività della linfa) : log 10 -2 (4 su 21), 3 (12 su 21), 4 (11 su 21), 5 (5 su 21), 6 (3 su 21) o 7 (1 su 21). L’infettività della linfa non è cambiata dal trattamento con etere dietilico. La linfa della foglia contiene pochi virioni (in 20 casi su 33) o molti virioni (nei restanti 13 casi su 33). • Proprietà morfologiche: virioni filamentosi, senza pericapside, di solito dritti o flessuosi, con una lunghezza modale libera di 509-655,7-720 nm e larghezza di 11-13,14-18 nm, diametro di 2-2,7-3,5 nm (figura 1), canale assiale evidente o scuro, elica di base evidente o poco chiara, con un passo elicale di base di 3,3-3,4-3,5 nm. Figura 1 – Particelle virali di Carlavirus con differenti dimensioni.
Transcript of carlavirus - francescofiume.altervista.org · pathway di taglio di Dicer e determina...

Fiume Francesco
Genere Carlavirus Classificazione
Gruppo IV (+ssRNA) Ordine Tymovirales Famiglia Betaflexiviridae
Genere Carlavirus
Le caratteristiche sintetiche del genere Carlavirus sono: • Proprietà delle particelle virali nella linfa: TIP (thermal inactivation point; la più bassa
temperatura alla quale l’infettività della linfa è distrutta entro un’ora): 50-68,5-85 °C. LIV (longevity in vitro; longetività dell’infettività della linfa in vitro): 0,5-4,8-2,01 giorni. DEP(diluition end point; punto finale di diluizione dell’infettività della linfa) : log10 -2 (4 su 21), 3 (12 su 21), 4 (11 su 21), 5 (5 su 21), 6 (3 su 21) o 7 (1 su 21). L’infettività della linfa non è cambiata dal trattamento con etere dietilico. La linfa della foglia contiene pochi virioni (in 20 casi su 33) o molti virioni (nei restanti 13 casi su 33).
• Proprietà morfologiche: virioni filamentosi, senza pericapside, di solito dritti o flessuosi, con una lunghezza modale libera di 509-655,7-720 nm e larghezza di 11-13,14-18 nm, diametro di 2-2,7-3,5 nm (figura 1), canale assiale evidente o scuro, elica di base evidente o poco chiara, con un passo elicale di base di 3,3-3,4-3,5 nm.
Figura 1 – Particelle virali di Carlavirus con differenti dimensioni.

2
• Proprietà fisiche: un componente di sedimentazione in preparazioni purificate; coefficiente
di sedimentazione 147,5-163,2-176,0 S. Densità 1,28-1,31-1,32 g cm-3 in CsCl. Densità 1,29 g cm-3 in saccarosio. Punto isoelettrico pH 4,5. Rapporto A260/A280 di 1,08-1,24-1,4.
• Proprietà biochimiche: i virioni contengono 2,5-5,7-8,5% di acido nucleico; 93,75-94,52-95,00% di proteine; 0% di lipidi.
• Genoma: è costituito, nel 100% dei casi, da RNA a singolo filamento e lineare. Le dimensioni totali del genoma sono di 6,48-7,77-8,50 kb. Il genoma è unipartito, con la parte più grande di 6,48-7,76-8,53 kb. La composizione di base è 31% di guanina, 24% di adenosina, 23% di citosina, 22% di uracile. L’infettività è mantenuta quando il genoma è deproteinizzato con proteasi; conservata quando il genoma è deproteinizzato con fenolo o detergente (4 casi su 4), o persa quando è deproteinizzato mediante fenolo o detergente (1 caso su 4). Fattore aggiuntivo non richiesto per l’infettività (2 casi su 2). L’acido nucleico non genomico non è presente nei virioni. L’mRNA subgenomico non è presente nelle cellule infette. Il virus ha evidenziato specie dsRNA trovate nelle cellule infette. Gli studi citopatologici hanno dimostrato che, all’interno delle cellule infette, le particelle virali si presentano sparse nel citoplasma e in parallelo, in aggregati legati a fascio.
Figura 2 – Schema del genoma del genere Carlavirus. RdRp è il dominio RNA dependent RNA
polymerase, CP è la coat protein, TGB è un triple gene block, RBP è l’RNA binding
protein.
Con riferimento alla figura 2 bisogna precisare che: RdRp è un importante enzima presente in tutti i virus contenenti il filamento di RNA come genoma. Questo enzima catalizza:
1. la sintesi del filamento (-) complementare, che è un intermedio durante la replicazione, utilizzando il filamento (+) di RNA come calco;
2. la sintesi del filamento (+), mediante il filamento (-) di RNA, usato come calco.
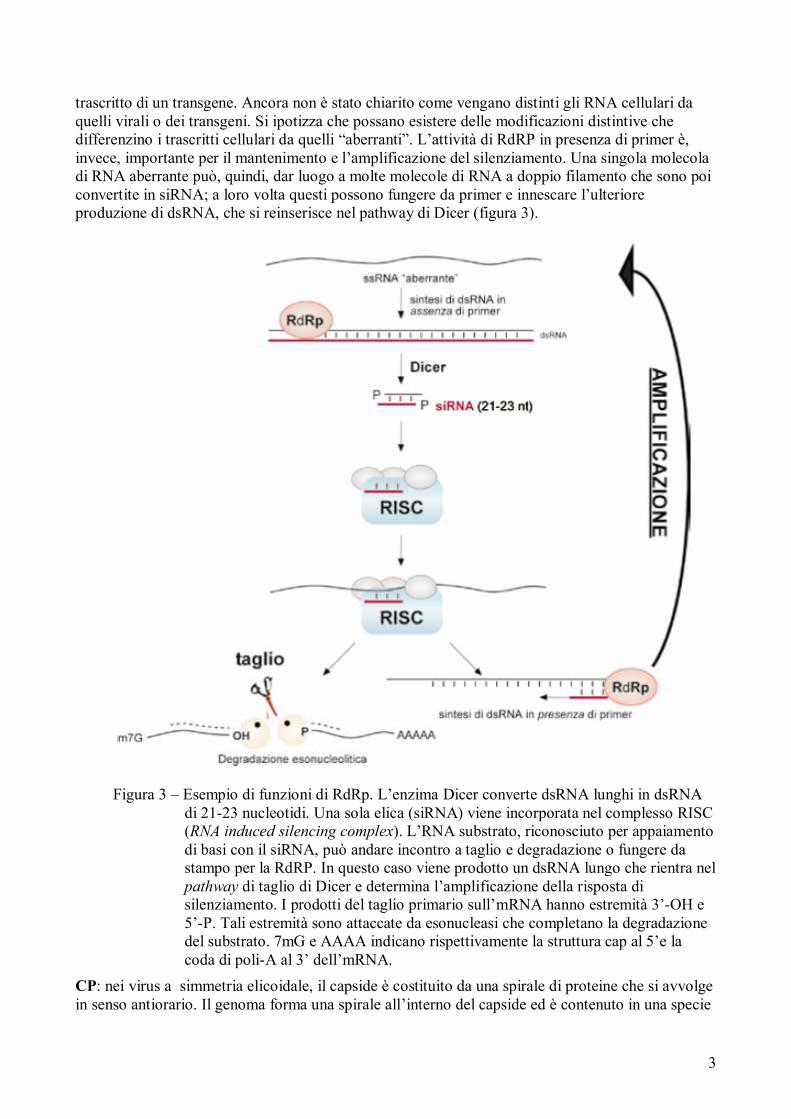
RdRp è un enzima in grado di convertire RNA a singolo filamento in dsRNA, in presenza e in assenza di un primer. L’azione di tale enzima è importante per l’inizio, per l’amplificazione del processo di silenziamento e permette la produzione di RNA a doppio filamento che innescano il ciclo dell’RNAi (RNA interference), dall’inglese interferenza dell’RNA, abbreviata comunemente come RNAi. Questo è un meccanismo epigenetico mediante il quale alcuni frammenti di RNA sono in grado di interferire e spegnere, silenziare, l’espressione genica, determinando la produzione di siRNA (small interfering RNA). L’attività polimerasica in assenza di primer è particolarmente importante per l’innesco del silenziamento successivo ad un’infezione virale o alla presenza di un
3
trascritto di un transgene. Ancora non è stato chiarito come vengano distinti gli RNA cellulari da quelli virali o dei transgeni. Si ipotizza che possano esistere delle modificazioni distintive che differenzino i trascritti cellulari da quelli “aberranti”. L’attività di RdRP in presenza di primer è, invece, importante per il mantenimento e l’amplificazione del silenziamento. Una singola molecola di RNA aberrante può, quindi, dar luogo a molte molecole di RNA a doppio filamento che sono poi convertite in siRNA; a loro volta questi possono fungere da primer e innescare l’ulteriore produzione di dsRNA, che si reinserisce nel pathway di Dicer (figura 3).
Figura 3 – Esempio di funzioni di RdRp. L’enzima Dicer converte dsRNA lunghi in dsRNA di 21-23 nucleotidi. Una sola elica (siRNA) viene incorporata nel complesso RISC (RNA induced silencing complex). L’RNA substrato, riconosciuto per appaiamento di basi con il siRNA, può andare incontro a taglio e degradazione o fungere da stampo per la RdRP. In questo caso viene prodotto un dsRNA lungo che rientra nel pathway di taglio di Dicer e determina l’amplificazione della risposta di silenziamento. I prodotti del taglio primario sull’mRNA hanno estremità 3’-OH e 5’-P. Tali estremità sono attaccate da esonucleasi che completano la degradazione del substrato. 7mG e AAAA indicano rispettivamente la struttura cap al 5’e la coda di poli-A al 3’ dell’mRNA.
CP: nei virus a simmetria elicoidale, il capside è costituito da una spirale di proteine che si avvolge in senso antiorario. Il genoma forma una spirale all’interno del capside ed è contenuto in una specie

4
di doccia situata sulla parte superiore della proteina. Al centro del virus è presente un tubicino con enzimi e matrice proteica. Nella figura 4 è riportata la posizione della proteina del capside in un virus privo di pericapside ed in un virus con pericapside
Figura 4 – Virus senza pericapside (A) e con (B). Si può osservare: 1) capside, 2) acidi nucleici, 3) proteina del capside, 4) nucleocapside, 5) virione, 6) proteine di membrana, 7) proteine esterne alla membrana.
Le proteine che costituiscono il capside dei virus si dispongono in modo simmetrico o quasi-simmetrico. Ci sono due possibili disposizioni (figura 5)
Figura 5 - Virus con capside elicoidale, in cui le subunità proteiche si associano a formare un bastoncello elicoidale (A). Virus con capside sferico, in cui le subunità proteiche si aggregano a formare un icosaedro (B).

5
TGB. Perché un virus possa invadere con successo una pianta e causare malattie, deve avere la capacità di muoversi tra le cellule, stabilire punti di infezione localizzati, entrare e uscire dal sistema vascolare delle piante e sviluppare infezioni sistemiche. Per realizzare queste attività, i virus dei vegetali codificano una o più proteine di movimento (MP) che facilitano il movimento cellulare e il trasporto attraverso i vasi. Queste proteine generalmente si localizzano nei plasmodesmi delle cellule ed aumentano la permeabilità plasmodesmale e, quindi, intercellulare, in maniera sufficiente per consentire il movimento delle macromolecole attraverso il desmotubulo, che attraversa la parete intercellulare. Molte di queste proteine di movimento hanno attività di legame con l’RNA e alcune agiscono in concerto con altre proteine codificate dal virus, per agevolare il movimento virale ed altre attività, come quella dell’RNA o la soppressione del silenziamento del gene. Diverse classi generali di proteine virali sono note e forniscono strumenti per indagare un’ampia gamma di interazioni tra virus e ospiti e delle funzioni cellulari. Le proteine di movimento maggiormente indagate sono quelle della superfamiglia di 30K, codificate da un gran numero di virus RNA e DNA, con diverse organizzazioni genomiche. Negli ultimi anni, gli studi sui processi realizzati dalle proteine di movimento di 30K hanno fornito una grande conoscenza dei requisiti per il trasporto localizzato ed a lunga distanza, con particolare riferimento al virus del mosaico del tabacco ed a molti altri virus (Boevink et Oparka, 2005; Heinlein et Epel, 2004; Lucas, 2006; Waigmann et al., 2005). La superfamiglia del blocco genico triplo (TGB) rappresenta un’altra classe importante di proteine di movimento, codificate da virus a RNA, in modo tassonomicamente diversificato (Morozov et Solovyev, 2003). In questa superfamiglia, i tre geni necessari per il movimento sono organizzati in sovrapposizioni di ORF e funzionano in concerto, per mediare il movimento dei virus. Numerosi studi hanno dimostrato che le proprietà molecolari delle proteine TGB di 30K sono abbastanza distinte e le analisi di sequenza suggeriscono di aver subito un’evoluzione convergente. La superfamiglia TGB può essere suddivisa in due classi principali che presentano differenze sostanziali nella struttura e nel requisito della proteina del capside (CP) per il movimento intercellulare e vascolare. Le proteine TGB di classe I o hordeivirus sono codificate da virus tubolari costituiti dai generi Benyvirus, Hordeivirus, Pecluvirus e Pomovirus, in cui la coat protein è dispensabile per il movimento intercellulare. I tipi di classe II o potexvirus sono rappresentati da virus flessuosi, classificati nei generi Allexivirus, Carlavirus, Foveavirus e Potexvirus, che richiedono la coat protein per il movimento. Alcuni studi hanno messo in evidenza che la strategia di movimento del TGB richiede l’attività coordinata di ciascuna delle proteine TGB, in particolare Barley stripe mosaic virus (BSMV) e Poa
semilatent virus (PSLV) ed il potexvirus Potato virus X (PVX) e che i loro livelli di espressione sono mediati dalla trascrizione e dalla traduzione da due RNA subgenomiali (sgRNA). Tuttavia, sono disponibili solo limitate evidenze circa le interazioni fisiche delle proteine TGB o l’importanza di queste interazioni nel movimento virale. Pertanto, sono necessarie analisi più dettagliate delle associazioni e delle funzioni biochimiche delle proteine TGB, per fornire una raffinata comprensione dei meccanismi in cui i loro processi di movimento sono mediati. Un complesso di ribonucleoproteina (RNP) da piante d’orzo, infetti da BSMV, che contiene una proteina simile a quella della proteina TGB1, è stato isolato. Anche se questi esperimenti sono stati condotti con virus di tipo selvatico, la capacità dei mutanti di BSMV, deficienti in coat protein per stabilire infezioni sistemiche, fornisce una prova evidente che il movimento del virus avviene tramite un complesso RNP. Tuttavia, le analisi biochimiche del complesso RNP non sono state condotte, quindi non si comprendono le possibili interazioni della proteina TGB1 con RNA virali o altre proteine TGB. È noto però che BSMV e le relative proteine PSDL TBD1 di hordeivirus sono in grado di legare sia RNA a singolo filamento, sia RNA a doppio filamento (dsRNA), con elevata affinità in vitro. Inoltre, le proteine TGB2 e TGB3 di BSGB sono necessarie per il movimento intercellulare, ma la loro presenza in RNP non è stata studiata, né le loro interazioni con TGB1 sono state comprese. Pertanto, è necessario rispondere a diverse domande riguardo le possibili interazioni

6
di binding delle proteine TGB e alla natura degli RNA virali associati ai complessi RNP, per comprendere i meccanismi di movimento di BSMV e di altri hordeivirus. Di conseguenza, sono state caratterizzate le interazioni BSMV/TGB più dettagliatamente e sono state analizzate le associazioni fisiche delle proteine TGB e le esigenze di espressione cis e trans per il movimento intercellulare. Barley stripe mosaic virus (BSMV) codifica tre proteine di movimento in un blocco genico triplo (TGB) sovrapposto, ma poco si sa sulle interazioni fisiche di queste proteine. Si è caratterizzato un complesso ribonucleoproteinico (RNP), costituito dalla proteina TGB1 e da RNA senso positivo di BSMV, da piante infette di orzo ed è stato identificato il complesso TGB1 nella pianta ed in vitro. Il legame omologo di TGB1 è stato interrotto da mutazioni specifiche del sito in ognuno dei primi due motivi N terminali dell’elicasi, ma non da mutazioni in due motivi C terminali dell’elicasi. Le proteine TGB2 e TGB3 non sono state rilevate in RNP, ma la cromatografia di affinità e gli esperimenti di due ibridi di lievito hanno dimostrato che TGB1 si lega a TGB3 e che TGB2 e TGB3 formano interazioni eterologhe. Queste interazioni hanno richiesto la TGB2 glicina 40 e la TGB3 isoleucina 108 e mutanti BSMV, contenenti tali sostituzione di aminoacidi, non sono stati in grado di dar luogo al movimento intercellulare. Gli esperimenti di infettività hanno indicato che TGB1, separato su un diverso RNA genomico da TGB2 e TGB3, potrebbe funzionare in un movimento limitato intercellulare, ma i tassi di movimento dipendono dai livelli di espressione delle proteine e dai contesti in cui sono espressi. Inoltre, l’elevata espressione della proteina TGB3 di tipo selvatico interferisce con il movimento intercellulare, ma il movimento non è stato influenzato dall’espressione simile di un mutante TGB3 che non interagisce con TGB2. Questi esperimenti suggeriscono che il movimento BSMV necessita di interazioni fisiche di TGB2 e TGB3 e che la deviazione sostanziale dai rapporti proteici TGB, espressa dal virus di tipo selvatico, compromette il movimento. RBP ha il compito di elaborare e modificare l’RNA; di regolazione trascrizionale; di splicing (una modifica del nascente pre-mRNA che avviene insieme o dopo la trascrizione, nella quale gli introni sono rimossi e gli esoni vengono uniti); di trasporto, localizzazione, stabilizzazione e traduzione di mRNA; di interferenza dell’RNA (RNAi). Le prime proteine che legano l’RNA (RBP) furono descritte negli anni ‘80, ma le RBP oggi conosciute sono solo una piccola parte di quelle esistenti. Basta considerare che la famiglia di geni contenenti PRNA Recognition Motif (RRM), uno dei domini di legame all’RNA, rappresenta da sola ben 1,5-2% del genoma umano. Le RBP sono implicate in tutti i processi di regolazione post-trascrizionale, dallo splicing e l’esportazione dell’mRNA dal nucleo, fino alla regolazione della localizzazione della traduzione e della degradazione dell’RNA messaggero. Le RBP possono essere classificate in base ai domini di legame all’RNA che contengono. Anche se questi domini sono molto conservati e le loro proprietà strutturali sono state ben caratterizzate, ad oggi non é ancora possibile associare ad una struttura, una determinata specificità di riconoscimento. Allo stesso modo non é possibile predire la funzione specifica di una RBP, conoscendo solo il dominio di legame all’RNA che contiene. Il motivo é che le RBP si sono evolute per combinazioni successive di questi domini e di altri moduli funzionali non implicati direttamente nel legame all’RNA. I domini ausiliari possono contribuire alla funzione della proteina, conferendo una maggiore specificità di riconoscimento, interagendo con altre proteine o svolgendo un’attività enzimatica. Infine, una stessa RBP può avere funzioni diverse, sia se regolata a livello post-traduzionale (fosforilazione, localizzazione cellulare), sia se interagisce con cofattori proteici diversi. Le RBP possono regolare la traduzione in diversi stadi (inizio, allungamento o terminazione) e possono esercitare un’attività attivatrice o inibitrice. L’inizio della traduzione è il passaggio più regolato. Nell’inizio, cap-dipendente proteine che legano il cap al 5’ dell’mRNA e proteine che legano la coda di poli(A) prendono contatto tra loro, formando una struttura detta closed-loop. La coda delle poli(A) è legata e stabilizzata da proteine dette PABP, cioè Poli(A) Binding Proteins.

7
L’assemblaggio del closed loop richiede il legame del fattore di inizio eIF4E all’estremità 5’ dell’mRNA e il legame del fattore eIF4G alle PABP e a eIF4E. Alcuni regolatori della traduzione regolano la lunghezza della coda delle poli(A): RNA messaggeri, con una coda di poli(A) corta, hanno una bassa probabilità di assumere correttamente la struttura closed loop e possono rimanere silenti o essere degradati. Altri fattori, invece, agiscono competendo con eIF4G per il legame a eIF4E. Queste RBP sono in grado di riconoscere sequenze specifiche nel 3’ UTR degli mRNA bersaglio e possono essere regolate da effettori a valle di vie di trasduzione del segnale. A differenza della regolazione trascrizionale, il controllo della traduzione ha due proprietà importanti che lo rendono cruciale in diverse fasi dello sviluppo e nel differenziamento. Queste proprietà sono la velocità e la possibilità di un controllo locale. La velocità di regolazione consente ad una cellula di rispondere rapidamente ai segnali ambientali. Ad esempio, in Aplysia, in seguito all’azione sinaptica, viene attivata una proteina CPEB, in grado di legare specifici mRNA e di promuoverne la traduzione molto prima che possano avvenire cambiamenti dell’espressione genica a livello nucleare. La possibilità del controllo locale della traduzione permette di limitare la sintesi di una proteina ad uno specifico compartimento cellulare o di distribuirla lungo un gradiente.
Figura 6 – Distribuzione degli mRNA, in relazione allo stadio, del bicoid anteriore e del nanos posteriore.
Questo é stato descritto in diversi casi, come nello stabilirsi degli assi embrionali nell’oocita di Drosophila, dove i gradienti dei morfogeni Hunchback e Caudal (figura 6) vengono stabiliti grazie ad inibitori della traduzione distribuiti nell’oocita secondo gradienti opposti. I geni ad effetto materno sono i primi ad attivarsi. Una serie di geni provenienti dalle cellule della madre determina l’asse antero-posteriore e quello dorso-ventrale dell’uovo. Le uova e le larve di

8
Drosophila, ad esempio, sono caratterizzate da una distribuzione disomogenea di specifiche molecole segnale, nel citoplasma. Queste molecole, che includono mRNA e proteine, sono codificate da alcuni geni particolari, chiamati geni a effetto materno; questi geni sono trascritti nelle cellule dell’ovario, che circondano la futura porzione anteriore dell’uovo, e determinano quelli che saranno gli assi corporei. In Drosophila due proteine, bicoid e nanos, sono concentrate, rispettivamente, all’estremità anteriore e a quella posteriore dell’embrione precoce L’asse anteroposteriore dell’embrione viene determinato dal gradiente di un morfogeno codificato dal gene bicoid. La proteina bicoid svolge la funzione di fattore di trascrizione e attiva un gene che stabilisce se la regione in questione svilupperà le strutture della testa. Altri geni ad effetto materno, presenti nella porzione posteriore dell’embrione, inibiscono l’azione di bicoid, delimitando, pertanto, la regione caudale. La proteina prodotta sullo stampo di RNA bicoid sarà disposta in un gradiente di concentrazione antero-posteriore. Questa proteina è un fattore di trascrizione, dunque è capace di attivare la sintesi di RNA da parte di geni specifici ai quali si va a legare. Essa si lega allora al DNA dei nuclei che si trovano nella parte anteriore dell’embrione e vi attiva la trascrizione (cioè la sintesi di RNA) da parte di un gene detto hunchback. Solo nella zona anteriore, dunque, si produrrà la proteina di hunchback, che a sua volta andrà solo in questa parte a stimolare la trascrizione di altri geni, ancora in via di individuazione, che consentiranno la formazione della testa. D’altro canto si è trovato che RNA prodotto da un altro gene, detto caudal, importante per la formazione di strutture posteriori, è distribuito uniformemente in tutto l’ovocita, ma la sua proteina forma pure un gradiente con massimo posteriore e minimo anteriore, perché la proteina bicoid si lega all’estremità 3′ non tradotta di RNA di caudal e ne inibisce la traduzione in proteina nella zona anteriore. È questo un meccanismo che sarà ritrovato altre volte nello sviluppo e che sottolinea ancora l’importanza di questa estremità del RNA per regolare il suo ancoraggio in territori particolari delle cellule e per la sua traducibilità in proteina. La formazione dell’addome si basa ancora infatti su questo meccanismo: l’RNA di hunchback che stimola la formazione della testa, anche se prodotto solo anteriormente nell’embrione, come visto, è presente anche nella sua parte posteriore, perché vi era già nell’ovocita prima della fecondazione; esso però non viene tradotto in proteina in quanto si è ancorato nella zona posteriore, per mezzo del proprio estremo 3′ non tradotto, al RNA prodotto dal gene nanos. Pertanto, si è formato un gradiente di concentrazione con massimo posteriore e minimo anteriore della proteina di nanos. Questa si lega all’estremità 3′ non tradotta del RNA di hunchback e ne inibisce la traduzione in proteina; nella zona posteriore, dunque, sarà presente la proteina di caudal ma non quella di hunchback; ciò porta all’attivazione solo nei nuclei della zona posteriore dell’embrione dei geni detti knirp e giant che, attraverso meccanismi ancora allo studio, stimolano la formazione dell’addome.
Pertanto: • gli mRNA bicoid sono localizzati nella parte anteriore dell’uovo non fecondato, legati ai
microtubuli anteriori; • i messaggi nanos sono legati al citoscheletro nella regione posteriore dell’uovo non fecondato; • gli mRNA hunchback e caudal sono distribuiti in tutto l’oocito.
Queste considerazioni sulla funzione delle RBP possono essere sufficienti nell’ipotesi che una RBP regoli un numero limitato di mRNA. In realtà l’applicazione dei microarray allo studio dei bersagli di RBP ha rivelato che una singola RBP è in grado di legare in vivo fino a un centinaio di mRNA. Per interpretare questi dati, è stata formulata l’ipotesi dell’operone post-trascrizionale. Secondo questo modello, gli mRNA regolati da una stessa RBP rappresenterebbero i geni del tradizionale operone batterico e le RBP sarebbero i regolatori dell’operone. I membri di uno stesso operone avrebbero in comune uno stesso elemento in cis, riconosciuto dalla RBP corrispondente. Questi elementi sarebbero presenti in combinazioni differenti negli UTR dei messaggeri così regolati (figura 7).

9
Figura 7 - Modello dell’operone post-trascrizionale. A livello trascrizionale (in alto), i fattori di trascrizione (TF) regolano l’inizio della trascrizione, legandosi a sequenze specifiche del DNA (in giallo). A livello post-trascrizionale (al centro), le RBP regolano la degradazione, la traduzione o la localizzazione degli mRNA in modo coordinato, interagendo con sequenze o strutture specifiche dell’RNA in modo coordinato, interagendo con sequenze o strutture specifiche dell’RNA, spesso presenti nel 3’UTR (in rosso). Le relazioni funzionali a livello delle proteine (in basso) possono corrispondere a geni co-regolati a livello trascrizionale e post-trascrizionale: geni che codificano per proteine funzionalmente correlate, come le subunità di complessi stechiometrici (blu) o componenti della stessa via metabolica (grigio), possono esssere regolati dagli stessi TF e i 9 loro mRNA possono essere co-regolati a livello post-trascrizionale da RPB (linee tratteggiate).

10
Verifiche sperimentali hanno dimostrato che gli operoni post-trascrizioali esistono ed hanno una plasticità elevata. In Saccharomyces cerevisiae i cinque omologhi (Puflp-Puf5p) legano set specifici di mRNA, ad esempio Puf3p lega mRNA di proteine mitocondriali. In Drosophila, si riconosce mRNA con la stessa sequenza consensus riconosciuta da Puf3p, ma questi mRNA appartengono a gruppi funzionali completamente diversi da quelli individuati nel lievito. In altri termini, gli operoni post-trascrizionali possono evolvere molto velocemente, semplicemente per aggiunta o perdita dei siti di regolazione in cis che li identificano. Considerato che il repertorio di RBP dei genomi degli eucarioti superiori è confrontabile a quello dei fattori di trascrizione, ciò espande enormemente le possibilità di regolazione di genomi "più piccoli di quanto atteso". La coordinazione veloce e simultanea dell’espressione di decine di geni correlati funzionalmente é alla base dei meccanismi di switch molecolare. Il modo più efficiente per ottenere questo tipo di coordinazione dell’espressione genica é il controllo della traduzione da parte di RBP. Esistono diversi esempi: in Caenorhabditis elegans il passaggio da uno stadio larvale al successivo o lo switch da spermatogenesi a oogenesi richiedono l’attività di diverse RBP. Questo scenario é ulteriormente complicato dalla possibilità di regolazione della traduzione tramite i miRNA. Agli miRNA é stato attribuita la funzione di regolatori fini dell’espressione genica e di responsabili della "robustezza" dei processi di sviluppo. Anche gli miRNA regolano simultaneamente molti messaggeri, che hanno in comune lo stesso seed
match. Tuttavia la regolazione traduzionale da parte di RBP mostra una maggiore plasticità, sia per il tipo di regolazione (non necessariamente inibitoria), sia per la possibilità di risposta diretta alle vie di trasduzione del segnale. Indagini bioinformatiche hanno mostrato che esiste una correlazione positiva tra la distribuzione di siti di legame di RBP note e quella dei siti di legame di miRNA (George et Tenenbaum, 2006). Ciò suggerisce che miRNA e RBP possono competere per siti di legame adiacenti e che legandosi ai siti bersaglio potrebbero indurre la formazione di strutture secondarie mutualnente esclusive, oppure cooperare in maniera positiva per la regolazione dello stesso messaggero. Dato che le RBP possono rispondere a segnali ambientali, queste potrebbero funzionare come veri e propri modulatori dell’attività delle miRNA. Grazie a queste proprietà, i meccanismi di regolazione della traduzione potrebbero essere alla base del "patterning temporale" nei processi di differenziamento. In altri termini, il controllo della traduzione potrebbe essere usato per stabilire una serie discreta di stati distinti, che si susseguono in un ordine definito, tramite meccanismi di switch molecolare. La successione di stati di competenza distinti nei progenitori multipotenti del sistema nervoso è alla base della generazione della diversità neuronale.
• Caratteristiche delle proteine: una proteina virale nel 95% dei casi o due, nel restante 5% dei casi; Nel caso di un’unica proteina virale, il valore di Mr è 23.200-32.530-40.000, nel caso di due proteine virale, il valore Mr di quella più grande è 24.000.
• Replicazione: la replicazione non dipende da un helper virus nel 100% dei casi. • Citopatologia: i virioni sono stati trovati nel citoplasma (23 casi su 23), nei cloroplasti (1 caso
su 23) o nei mitocondri (1 su 23). Le inclusioni sono presenti nelle cellule infette (12 casi su 20), o sono assenti da cellule infette (8 casi su 20). Nel citoplasma vi sono cristalli (1 casi su 12), o corpi X-amorfi (2 casi su 12) o corpi membranosi (3 casi su 12) o viroplasmi (3 casi su 12) o di forma inusuale (6 casi su 12). Contengono virioni (7 casi su 8), o non contengono virioni (1 caso su 8).
Il genere Carlavirus è stato proposto per la prima volta nel primo rapporto dell’International
Committee on Taxonomy of Viruses (ICTV) nel 1971 come Carnation latent virus group, ma è stato

11
rinominato nel 1975 come gruppo Carlavirus e come genere Carlavirus nel 1995 (VI rapporto ICTV). Nel 2005 (IIX rapporto) è stato inserito nella famiglia Flexiviridae, precedentemente non assegnato. La posizione attuale nel IX rapporto ICTV (2009) è indicato come un genere della famiglia Betaflexiviridae derivata dalla successiva suddivisione dei Flexiviridae. Nalla versione 2014 del database ICTV sono riportate 52 virus inclusi nel genere Carlavirus. Attualmente sono contate 55.
Ciclo vitale
La replicazione virale è citoplasmatica e lisogena. L’entrata nella cellula ospite è ottenuta mediante la penetrazione nella cellula ospite. La replica segue il modello di reazione dei virus RNA a filamento positivo. La trascrizione del virus RNA a filamento positivo è il metodo della trascrizione. Il virus esce dalla cellula ospitante mediante movimento tripartito non tubulare guidato virale. Le piante servono come ospiti naturali.
Prima di procedere occorre ricordare alcuni concetti sulla di replicazione dei virus a RNA. I virus a RNA seguono 4 principali strategie, due delle quali con significative variazioni a seconda dei gruppi di virus (figura 8).
Figura 8 – Strategie di replicazione e traduzione dei virus a ssRNA positivo e ssRNA negatico
Virus con RNA monocatenario, monofilamento, a polarità positiva non hanno inizialmente bisogno di trascrivere il proprio genoma perché il loro RNA può essere subito tradotto dai ribosomi della cellula. Nei Picornavirus e nei Flavivirus, il prodotto della traduzione è un’unica poliproteina che viene successivamente scissa nelle varie proteine codificate per opera di proteasi virali.
12
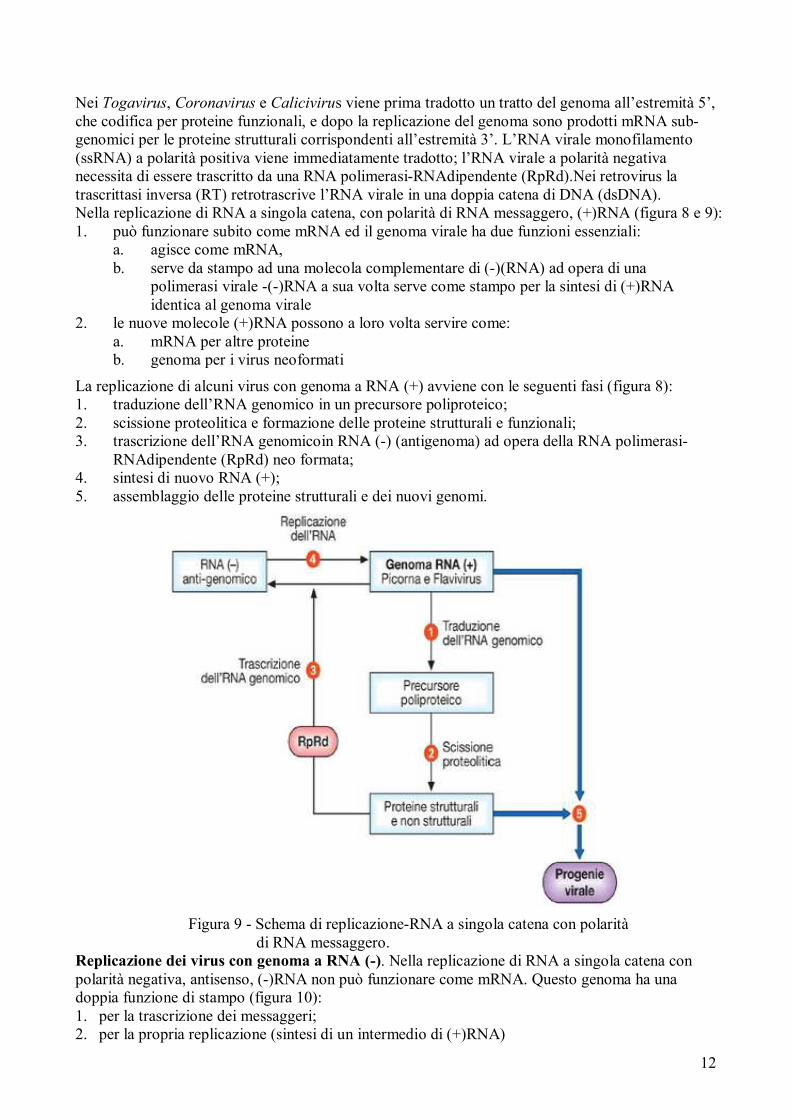
Nei Togavirus, Coronavirus e Calicivirus viene prima tradotto un tratto del genoma all’estremità 5’, che codifica per proteine funzionali, e dopo la replicazione del genoma sono prodotti mRNA sub-genomici per le proteine strutturali corrispondenti all’estremità 3’. L’RNA virale monofilamento (ssRNA) a polarità positiva viene immediatamente tradotto; l’RNA virale a polarità negativa necessita di essere trascritto da una RNA polimerasi-RNAdipendente (RpRd).Nei retrovirus la trascrittasi inversa (RT) retrotrascrive l’RNA virale in una doppia catena di DNA (dsDNA). Nella replicazione di RNA a singola catena, con polarità di RNA messaggero, (+)RNA (figura 8 e 9): 1. può funzionare subito come mRNA ed il genoma virale ha due funzioni essenziali:
a. agisce come mRNA, b. serve da stampo ad una molecola complementare di (-)(RNA) ad opera di una
polimerasi virale -(-)RNA a sua volta serve come stampo per la sintesi di (+)RNA identica al genoma virale
2. le nuove molecole (+)RNA possono a loro volta servire come: a. mRNA per altre proteine b. genoma per i virus neoformati
La replicazione di alcuni virus con genoma a RNA (+) avviene con le seguenti fasi (figura 8): 1. traduzione dell’RNA genomico in un precursore poliproteico; 2. scissione proteolitica e formazione delle proteine strutturali e funzionali; 3. trascrizione dell’RNA genomicoin RNA (-) (antigenoma) ad opera della RNA polimerasi-
RNAdipendente (RpRd) neo formata; 4. sintesi di nuovo RNA (+); 5. assemblaggio delle proteine strutturali e dei nuovi genomi.
Figura 9 - Schema di replicazione-RNA a singola catena con polarità di RNA messaggero.
Replicazione dei virus con genoma a RNA (-). Nella replicazione di RNA a singola catena con polarità negativa, antisenso, (-)RNA non può funzionare come mRNA. Questo genoma ha una doppia funzione di stampo (figura 10): 1. per la trascrizione dei messaggeri; 2. per la propria replicazione (sintesi di un intermedio di (+)RNA)

13
La trascrizione avviene per una trascrittasi virale presente nel virione. La molecola di (+)RNA serve come stampo per la sintesi di (-)RNA che costituirà il genoma delle particelle virali.
Figura 10 – Schema di replicazione dei virus con genoma a RNA (-).
Replicazione di alcuni virus con genoma a RNA (-). Avviene secondo le seguenti fasi:
1. trascrizione dell’RNA (-) in RNA (+) ad opera della RNA polimerasi-RNAdipendente (RpRd) associata al virione;
2. traduzione dell’RNA (+) neoformato; 3. trascrizione dell’RNA genomico in RNA (+), antigenoma, ad opera della
RpRd virale (figura 9); 4. sintesi del nuovo RNA di progenie; 5. assemblaggio delle proteine strutturali e dei nuovi genomi.
Ribovirus con RNA bicatenario (figura 9). É necessaria una trascrittasi virale (RNA-polimerasiRNA-dipendente) per iniziare il ciclo replicativo. Solo la catena (-)RNA del genoma viene trascritta per formare molecole di (+)RNA che hanno duplice funzione: 1. quella di messaggero; 2. quella di stampo per la sintesi di molecole di (-)RNA complementari; 3. (+)RNA e (-)RNA rimangono uniti generando RNA a doppio filamento del genoma virale.
Trasmissione dei Carlavirus nell’infezione L’infezione virale è talvolta diffusa dagli afidi in modo semi-persistente, vale a dire che il vettore è infetto per un certo numero di ore. Alcune specie vengono trasmesse da Bemisia tabaci in modalità semi-persistente o attraverso il seme. La maggior parte delle specie infetta solo pochi ospiti e causa infezioni con pochi o nessun sintomo, ad esempio l’American hop latent virus e il Lily symptomless
virus. Alcuni, come il Blueberry scorch virus e il Poplar mosaic virus, causano gravi malattie. Con riferimento alla trasmissione virale per seme, questa è oggi nota per circa un quinto dei fitovirus descritti. Per la precisione, facendo riferimento al più recente aggiornamento dell’International Committee on Taxonomy of Viruses (ICTV), essa risulta segnalata per membri di

14
26 dei 66 generi virali appartenenti a 8 delle 16 famiglie che includono virus dei vegetali, per un totale di 130 patogeni. Si ritiene, tuttavia, che il fenomeno sia alquanto più comune di quanto oggi non risulti, interessando, forse, un terzo o più dei fitovirus. Stima che appare fondata se si considera che l’incremento delle ricerche portò il numero dei virus trasmessi per seme dagli 8 conosciuti all’inizio degli anni Cinquanta a ben 156 verso la fine degli anni Ottanta. Successive verifiche hanno ridimensionato questo dato mettendo in evidenza sia segnalazioni improprie per svariati motivi, (identificazione dei patogeni approssimativa e procedure sperimentali poco attendibili. ad esempio), sia l’esistenza di sinonimie tra alcuni dei virus menzionati. Poiché il seme risulta dalla fusione di due gameti, uno della pianta madre (ovulo) ed uno della pianta fecondatrice (polline) — che è la stessa pianta nelle specie autogame — la trasmissione per seme può avvenire attraverso l’uno, l’altro o entrambi i gameti, o le piante progenitrici. Pertanto, la trasmissione per seme e quella per polline sono così strettamente correlate che l’argomento può essere trattato in modo esauriente solo riferendosi ad entrambe. Vi sono tuttavia. problematiche specificamente pertinenti ciascuna delle due: 1. la trasmissione per seme sensu stricto considera in dettaglio, ad esempio, i fattori che
influenzano i movimenti dei virus sia dai tessuti della pianta madre a quelli del seme che da quelli dell’embrione alla plantula che ne deriva,
2. mentre la trasmissione per polline esamina argomenti come gli effetti dei virus sulla germinazione del granulo pollinico, la possibilità di replicazione virale all’interno di questo, l’eventualità. che il polline infetti — oltre al seme — la pianta stessa che viene fecondata.
La trasmissione per seme dei virus può avvenire secondo diversi meccanismi: a. per contaminazione della superficie esterna del seme o di parti del seme di origine materna,
come il tegumento esterno e interno (trasmissione non embrionale). b. per infezione dell’embrione (trasmissione embrionale) che può aver luogo mediante:
• infezione indiretta, per invasione di tessuti e organi di riproduzione (ovario cellule madri delle megaspore, cellule madri del polline) prima dell’embriogenesi,
• infezione dietta. per contaminazione dell’embrione nel corso dell’embriogenesi. Trasmissione non embrionale. Se ne conoscono pochi casi. tutti riguardanti virus con particelle molto stabili, resistenti alla degradazione e trasmissibili per inoculazione meccanica. Si tratta dei tobamovirus TMV, ToMV), che possono ritrovarsi sia sulla superficie che nel tegumento e nell’endosperma dei semi di pomodoro e peperone, e CGMMV). che può contaminare il tegumento dei semi di cetriolo. La trasmissione ai giovani semenzali si verifica per inoculazione meccanica. attraverso microlesioni provocate principalmente dalla rottura di peli fogliari durante la manipolazione delle piante (soprattutto con le operazioni di trapianto o ripiccaggio). Altro caso. unico nel suo genere. è rappresentato dal necrovirus MNSV che sopravvive nel tegumento interno dei semi di cetriolo e di qui può essere trasmesso ai giovani semenzali dal fungo chitridiaceo Olpidium bornavanus. Trasmissione embrionale indiretta. Studi ultrastrutturafi hanno dimostrato che alcuni virus sono capaci di contaminare l’embrione indirettamente (ossia prima della sua formazione), mettendo in evidenza la presenza di particelle virali nelle cellule madri delle megaspore, in quelle del polline e nel polline stesso. Particelle di TRV, ad esempio, sono state osservate in cellule madri premeiotiche del polline di pomodoro e, in stadio più avanzato, nel polline stesso, mentre la presenza di AMV è stata accertata nel citoplasma del polline di erba medica. Entrambi i virus sono trasmessi per seme contaminato da polline infetto. TRSV è stato osservato sia nel megagametofito che nel polline di soia, specie nella quale il virus si trasmette in elevata percentuale verosimilmente perché — in base a quanto accertato — è capace di invadere il tessuto meristematico e quindi di infettare la cellula madre della megaspora. Sono stati condotti studi molto approfonditi sulla trasmissione per seme di BSMV nell’orzo, rilevando le modificazioni citologiche che si verificano nei meristemi fiorali durante la meiosi e la formazione dell’embrione in relazione alla distribuzione di un ceppo del virus trasmissibile (MI-1) e di uno non trasmissibile (NSP) per seme nei tessuti di riproduzione. Si è osservato che:

15
la formazione dello strato di callosio intorno alle megaspore ed alle cellule madri del polline ha luogo negli stadi premeiotici; non si rileva la presenza di plasmodesmi che mettano in comunicazione la cellula madre della megaspora con le circostanti cellule nucellari; dopo la fecondazione non si osservano plasmodesmi che connettano gli embrioni in via di sviluppo con i tessuti circostanti. Dei due ceppi virali studiati. quello trasmesso per seme è risultato presente nei tessuti delle cellule madri delle megaspore e del polline, oltrechè nell’ovario e nel polline,. mentre quello non trasmissibile non è stato mai osservato in questi tessuti. La conclusione è stata che la trasmissibilità per seme di BSMV è determinata dalla capacità del virus di invadere i meristemi riproduttivi maschili e femminili in stadi molto precoci del loro sviluppo, contaminando successivamente l’embrione per via indiretta. La marcata influenza dello stadio di sviluppo della pianta al momento dell’inoculazione sull’’entità della trasmissione per seme è stata più volte citata come elemento a sostegno della contaminazione indiretta dell’embrione (ossia presenza del virus nei tessuti riproduttivi prima dell’embriogenesi). In effetti, l’inoculazione della pianta in stadi fenologici avanzati o molto avanzati induce generalmente livelli di trasmissione per seme assai modesti, annullandosi anche del tutto se la pianta viene infettata durante o dopo la fioritura. Al contrario, l’infezione del seme per introduzione del virus nel sacco embrionale ad opera del polline si verifica, ovviamente, nel corso della fioritura. Nel caso specifico, semi infetti possono essere prodotti anche da piante madri esenti dal virus in questione. I casi di trasmissione per polline più noti in letteratura sono quelli di Prune dtwarf virus (PDV) e di Prunus necrotic ringspot virus (PNRSV) in diverse prunoidee (Prunus spp.), e di BCMV in fagiolo, oltre a quello già citato di BSMV in orzo. L’eventualità che il virus possa diffondersi dagli embrioni contaminati via polline ai tessuti adiacenti della pianta madre e infettarla, è stata presa in considerazione e studiata. Ricerche recenti hanno chiarito che la trasmissione da polline a pianta è effettivamente possibile, ad esempio per ilarvirus di specie ortensi e da frutto, ma sembra aver luogo soltanto con la mediazione di tripidi (e forse anche di altri insetti, soprattutto pronubi), i quali trasportano sui loro corpo granuli pollinici ricoperti in superficie di particelle virali, che vengono poi inoculate meccanicamente nelle piante visitate. Questo tipo di trasmissione è stato riscontrato anche ne. diffusione di KSV da piante spontanee a coltivazioni di pomodoro, sempre ad opera di tripidi. Trasmissione embrionale diretta. L’infezione diretta del seme si verifica in seguito al trasferimento del virus direttamente dalla pianta madre all’embrione, ma i meccanismi di questo processo sono ancora poco conosciuti. Prime informazioni sono state ottenute con ricerche sulla combinazione pisello cv “Vedette”/Pea seedbome mosaic virus (PSbMV), nella quale non si riscontra trasmissione per polline. I movimenti di PSbMV nei baccelli e nei semi in via di sviluppo sono stati rilevati mediante ELISA, saggi di immuno-citochimica e di ibridazione in situ. Il virus è stato rinvenuto nel funicolo prima della fecondazione, ma è risultato presente soltanto sporadicamente negli ovuli non fecondati. Dopo la fecondazione, esso è stato facilmente individuato nei tessuti sia della testa e dell’endosperma, sia del sospensore embrionale. Tuttavia. una vasta diffusione del virus nella testa e nell’endosperma è stata osservata soltanto in una cv di pisello nella quale PSbMV è trasmesso per seme ma non in un’altra, non soggetta a trasmissione per seme, nella quale la diffusione del virus nei tessuti citati è risultata molto limitata. In qual modo il virus penetri nell’embrione non è ancora chiarito, ma è stata formulata l’ipotesi che ciò si realizzi al punto di contatto ta la testa e il sospensore dell’embrione). La parete cellulare al punto di contatto è alquanto convoluta, ma priva di plasmodesmi, per cui si è suggerito che PSbMV potrebbe attraversarla con un meccanismo non meglio specificato che non richiede la presenza di plasmodesmi o, in alternativa,. che il virus stesso possa indurre la formazione di plasmodesmi e in tal modo invadere l’embrione .direttamente. PSbMV. contrariamente a quanto riscontrato nella cv “Vedette”. è trasmissibile per polline nella cv di pisello “Dual”, in percentuale del 4% circa. Le indagini condotte sulla combinazione PSbMV/pisello “Dual” hanno evidenziato che 4 di 96 semi ottenuti da fiori di piante sane sottoposti

16
a castrazione e fecondati con polline di piante affette dal virus hanno dato origine a semenzali infetti. La fecondazione di fiori di piante infette da PSbMV con polline proveniente da piante sane ha indotto la trasmissione per seme del virus in misura del 38%. Questi risultati indicano che nella combinazione virus/pianta ospite presa in esame, l’infezione dell’embrione può verificarsi sia direttamente che indirettamente. L’ostacolo principale all’invasione diretta dell’embrione da parte dei virus sembra essere l’apparente separazione tra polline, ovario e sacco embrionale, da un lato, e tessuti parentali dall’altro. Tale separazione deriva dalla scomparsa dei plasmodesmi tra le cellule madri delle megaspore e il tessuto nucellare e tra le cellule madri del polline e le cellule del cosiddetto tappeto, nel corso della meiosi. Inoltre, durante la meiosi, si forma intorno alle megaspore ed alle cellule madri del polline, uno strato di callosio, tipicamente interrotto intorno al sacco embrionale neoformato. Successivamente, lo strato di callosio attorno al polline ed al sacco embrionale viene riassorbito e rimpiazzato da altri tessuti, mentre una spessa parete di pectocellulosa, priva di plasmodesmi, si sviluppa attorno al sacco embrionale. Durante l’impollinazione i gametofiti maschile e femminile restano separati, poiché la parete del budello pollinico in via di sviluppo produce uno strato di callosio - assente soltanto alla sua estremità. terminale - via via che attraversa i tessuti dello stilo. In considerazione di quanto sopra, sembrerebbe ovvio concludere che la contaminazione del seme dalla pianta madre possa aver luogo più facilmente mediante penetrazione del virus nei tessuti riproduttivi, prima della scomparsa dei plasmodesmi e della formazione delle barriere protettive nominate, e quindi per infezione indiretta dell’embrione. In conclusione, fino a qualche decina di anni fa. la trasmissione per seme dei fitovirus è stata considerata un evento relativamente poco comune e la sua importanza sottovalutata. Prima che se ne comprendesse l’effettiva portata, essa ha causato difficoltà non indifferenti anche alla ricerca virologica, oltre che alla lotta contro le virosi in pieno campo. Le piante nate infette da seme, infatti, spesso non presentano sintomi evidenti (infezione latente), hanno aspetto normale e sono pertanto scambiate per sane. Questo fenomeno, verificandosi in alcune combinazioni virus/piante indicatrici – Sowbane mosaic virus (SwMV)/Chenopodium spp., ad esempio - ha indotto talora a ritenere di aver isolato questi virus da piante (fonte dell’ inoculo) che ne erano esenti mentre, in realtà, erano contaminate da seme le piante utilizzate nel saggio di inoculazione. Per quanto concerne la diffusione delle virosi in pieno campo, la trasmissione per seme - anche in bassa percentuale - può svolgere diverse funzioni cruciali per il ciclo epidemiologico di virus fitopatogeni, quali: • garantire la sopravvivenza dei patogeni in assenza di coltivazioni e da un ciclo colturale
all’altro, attraverso il seme di specie sia coltivate (BCMV in fagiolo) che spontanee (CMV in Stellaria media);
• costituire sorgenti di infezione primarie dalle quali prende avvio annualmente la diffusione secondaria dei virus trasmessi da vettori, quali afidi (AMV, CMV, LMV) e nematodi (TRV);
• consentire la disseminazione dei patogeni a lunga distanza, in seguito all’introduzione di semi infetti in aree agricole, anche molto lontane dal luogo di produzione. L’importanza di questa via di diffusione è provata dal fatto che tutti i virus trasmessi per seme in specie coltivate, soggette a commercializzazione, sono ubiquitari;
• causare la contaminazione di specie da frutto moltiplicate agamicamente, innescando l’infezione attraverso portinnesti riprodotti per seme. È il caso di PNRSV e PDV che si trasmettono per seme in varie specie di Prunus sp. utilizzate come portinnesti di fruttiferi;
• costituire nel terreno una duratura riserva di inoculo, consistente di semi infetti che conservano la germinabilità per lunghi periodi di tempo.
La trasmissione per seme non embrionale è propria di virus resistenti alla degradazione, molto infettivi e facilmente trasmissibili per contatto. Per questo motivo, essendo limitata a pochi casi, riveste rilevante importanza economica, soprattutto in specie come pomodoro e peperone in coltura protetta (TMV, ToMV). La trasmissione embrionale, diretta e indiretta, dei virus che invadono estesamente i tessuti parenchimatici e sono trasalissi. , inoculazione meccanica. Non sono cono.. invece casi di trasmissione , seme di virus localizzati esclusivamente nel tessuto floematico

17
(htteovirus, fitoreovirus, fijivirus, ecc.), i quali sono trasmessi da insetti in raodo persistente - circolativo o propagativo - ma non mediante inocul.one meccanica. Lo stesso vale , i fitoplasmi, agenti "virus-sir.i", floematici, . malattie delle piante, che sono trasmessi in modo propagativo da cicaline e psille. La presenza di un virus nel seme, anche nell’embrione, non sempre risulta nell’infezione del semenzale che ne deriva: in molti casi, infatti, nel corso d. processi metabolici connessi cm la germinazime, il virus .ene degradato e ,de linfettività. Ciò non pregiudica tuttavia l’utilità del controllo sanitario delle semedi attraverso le qudi determinati virus possono trasmettersi, , accertare che ne siano esenti. . raetodologie diagnostiche consistono in saggi di gerrainazione e infettività, sierologici, di immuno-microscopia elettronica e nell’utilizzazione di me. molecolari sempre più sensi. e affidabili. Il genere Carlavirus, la cui specie tipo è Carnation latent virus (CLV), comprende le seguenti specie definitive, le specie non ancora definitive.
Specie definitive
1. Alfalfa latent virus (ALV) 2. American hop latent virus (AHLV) 3. Blueberry scorch virus (BlScV) 4. Cactus virus 2 (CaV2) 5. Caper latent virus (CapLV) 6. Carnation latent virus (CLV) – specie tipo, figura 11
7. Chrysanthemum virus B (CVB) 8. Dandelion latent virus (DaLV) 9. Elderberry symptomless virus (ElSLV) 10. Fig virus S (FVS) 11. Helenium virus S (HVS) 12. Honeysuckle latent virus (HnLV) 13. Hop latent virus (HpLV) 14. Hop mosaic virus (HpMV) 15. Kalanchoe latent virus (KLV) 16. Lilac mottle virus (LiMV) 17. Lily symptomless virus (LSLV) 18. Mulberry latent virus (MLV) 19. Muskmelon vein necrosis virus (MuVNV) 20. Nerine latent virus (NeLV) 21. Passiflora latent virus (PLV) 22. Pea streak virus (PeSV) 23. Poplar mosaic virus (PopMV) 24. Potato virus M (PVM), figura 12
25. Potato virus S (PVS) 26. Red clover vein mosaic virus (RCVMV) 27. Shallot latent virus (SLV) 28. Strawberry pseudo mild yellow edge virus (SPMYEV)
Per le seguenti specie ci si trova ancora nella fase propositiva di inclusione nei Carlavirus: 29. Anthriscus virus (AntV) 30. Arracacha latent virus (ALV) 31. Artichoke latent virus M (ArLVM) 32. Artichoke latent virus S (ArLVS) 33. Aster chlorotic stunt virus (AsCSV) 34. Butterbur mosaic virus (ButMV) 35. Cardamine latent virus (CaLV) 36. Cassava brown streak-associated virus (CBSAV)

18
37. Cassia mild mosaic virus (CaMMV) 38. Chicory yellow blotch virus (ChYBV) 39. Chinese yam necrotic mosaic virus (CYNMV) 40. Cole latent virus (CoLV) 41. Cowpea mild mottle virus (CPMMV) 42. Cynodon mosaic virus (CynMV) 43. Dandelion virus (DaV) 44. Daphne virus S (DVS) 45. Eggplant mild mottle virus (EMMV) 46. Fuchsia latent virus (FLV) 47. Garlic common latent virus (GCLV) 48. Gentiana virus (GenV) 49. Helleborus mosaic virus (HeMV) 50. Hydrangea latent virus (HdLV) 51. Lilac ringspot virus (LiRSV) 52. Plantain 8 virus (PlV8) 53. Prunus S virus (PruVS) 54. Sint-Jan’s onion latent virus (SJOLV) 55. Southern potato latent virus (SoPLV) 56. White bryony mosaic virus (WBMV)
Specie ospiti suscettibili
1. Adansonia digitata
2. Allium ampeloprasum var. holmense
3. Allium ampeloprasum var. sectivum
4. Allium cepa
5. Allium cepa var. ascalonicum
6. Allium cepa var. cepa
7. Allium cepa var. perutile
8. Allium chinense
9. Allium fistulosum
10. Allium porrum
11. Allium sativum
12. Allium vineale
13. Alstroemeria
14. Amaranthus caudatus
15. Ammi majus
16. Angelica sylvestris
17. Anthriscus cerefolium
18. Anthriscus sylvestris
19. Antirrhinum majus
20. Apium graveolens
21. Arachis hypogaea
22. Arracacia xanthorrhiza
23. Aster amellus
24. Aster novae-angliae
25. Aster novi-belgii
26. Atriplex hortensis
27. Beta patellaris
28. Beta vulgaris
29. Beta vulgaris ssp. cicla

19
30. Brassica campestris ssp. rapa
31. Brassica juncea
32. Brassica oleracea
33. Brassica oleracea var. acephala
34. Brassica oleracea var. botrytis
35. Brassica oleracea var. capitata
36. Bryonia dioica
37. Cactaceae
38. Cajanus cajan
39. Calendula officinalis
40. Canavalia ensiformis
41. Capparis spinosa
42. Capparis spinosa var. rupestris
43. Capsicum annuum
44. Cardamine
45. Cardamine lilacina
46. Carum carvi
47. Cassia coluteoides
48. Cassia macranthera
49. Cassia obtusifolia
50. Cassia occidentalis
51. Cassia sylvestris
52. Catharanthus roseus
53. Celosia argentea
54. Chenopodium album
55. Chenopodium amaranticolor
56. Chenopodium capitatum
57. Chenopodium foetidum
58. Chenopodium giganteum
59. Chenopodium hybridum
60. Chenopodium murale
61. Chenopodium quinoa
62. Chrysanthemum carinatum
63. Chrysanthemum morifolium
64. Cicer arietinum
65. Cichorium endiva
66. Cichorium intybus
67. Citrullus lanatus
68. Crotalaria juncea
69. Crotalaria spectabilis
70. Cucumis melo
71. Cucumis sativus
72. Cucurbita maxima
73. Cucurbita pepo
74. Cyamopsis tetragonoloba
75. Cynara scolymus
76. Cynodon dactylon
77. Daphne
78. Daphne cneorum
79. Daphne genkwa
80. Daphne giraldii

20
81. Daphne laureola
82. Daphne odora
83. Daphne retusa
84. Daphne × burkwoodii
85. Daphne × dauphinii
86. Daphne × napolitana
87. Datura metel
88. Datura stramonium
89. Datura tatula
90. Daucus carota
91. Dianthus barbatus
92. Dianthus caryophyllus
93. Dioscorea batatas
94. Dioscorea batatas f. typica
95. Fagopyrum esculentum
96. Ficus carica
97. Fragaria chiloensis
98. Fragaria vesca
99. Fragaria virginiana
100. Fragaria × ananassa
101. Fuchsia speciosa × Fuchsia hybrida
102. Gentiana sino-ornata
103. Glycine max
104. Gomphrena globosa
105. Gynura aurantiaca
106. Helenium amarum
107. Helianthus annuus
108. Helleborus niger
109. Hippeastrum hybridum
110. Humulus japonicus
111. Humulus lupulus
112. Hydrangea macrophylla
113. Impatiens holstii
114. Kalanchoe blossfeldiana
115. Lactuca sativa
116. Lathyrus odoratus
117. Lavatera trimestris
118. Lens culinaris
119. Lilium longiflorum
120. Lonicera americana
121. Lonicera brownii
122. Lonicera caprifolium
123. Lonicera heckottii
124. Lonicera henryi
125. Lonicera japonica
126. Lonicera periclymenum
127. Lonicera telmanniana
128. Lupinus albus
129. Lupinus angustifolius
130. Lycopersicon esculentum
131. Malva parviflora

21
132. Manihot esculenta
133. Medicago alba
134. Medicago hispida
135. Medicago sativa
136. Melilotus albus
137. Melilotus officinalis
138. Morus alba
139. Nerine bowdenii
140. Nicandra physalodes
141. Nicotiana affinis
142. Nicotiana benthamiana
143. Nicotiana clevelandii
144. Nicotiana debneyi
145. Nicotiana glutinosa
146. Nicotiana glutinosa × Nicotiana clevelandii
147. Nicotiana megalosiphon
148. Nicotiana occidentalis
149. Nicotiana pavicola
150. Nicotiana rustica
151. Nicotiana sylvestris
152. Nicotiana tabacum
153. Nicotiana × edwardsonii
154. Ocimum basilicum
155. Passiflora caerulea
156. Pastinaca sativa
157. Pennisetum pedicellatum
158. Petasites officinalis
159. Petunia × hybrida
160. Phaseolus lunatus
161. Phaseolus vulgaris
162. Phlox drummondii
163. Physalis floridana
164. Physalis franchetii
165. Pisum sativum
166. Plantago lanceolata
167. Populus balsamifera
168. Populus candicans
169. Populus deltoides subspecies angulata, monilifera, missouriensis
170. Populus maximowiczii
171. Populus nigra
172. Populus trichocarpa
173. Populus × canadensis
174. Populus × interamericana
175. Prunus
176. Raphanus sativus
177. Rubus idaeus
178. Sambucus canadensis
179. Sambucus nigra
180. Sambucus racemosa
181. Senecio cruentus
182. Sesamum indicum

22
183. Sinapis alba
184. Solanum demissum
185. Solanum demissum × Solanum tuberosum
186. Solanum melongena
187. Solanum rostratum
188. Solanum tuberosum
189. Sonchus oleraceus
190. Sorghum bicolor
191. Spinacia oleracea
192. Syringa oblata var. affinis
193. Syringa vulgaris
194. Taraxacum officinale
195. Tetragonia tetragonioides
196. Theobroma cacao
197. Trifolium hybridum
198. Trifolium incarnatum
199. Trifolium pratense
200. Trifolium repens
201. Trifolium subterraneum
202. Tropaeolum majus
203. Tulipa gesneriana cultivars
204. Urtica urens
205. Vaccinium corymbosum
206. Vicia faba
207. Vicia sativa
208. Vicia villosa
209. Vigna radiata
210. Vigna subterranea
211. Vigna unguiculata
212. Zea mays
213. Zinnia elegans

23
Per concludere, sono riportate alcune immagini riguardanti la sintomatologia rilevata su alcune piante con infezioni da Carlavirus.
Figura 11 - Carnation latent virus (CLV) sul luppolo comune (Humulus
lupulus). In generale, CLV di solito si manifesta senza sintomi nelle piante infette. Tuttavia, queste sono meno vigorose e producono meno fiori di migliore qualità. Il virus può anche incrementare il danno causato da altri virus con cui coesiste nel complesso.
Figura 12 – Potato virus M (PVM). Il sintomo più comune, che si riscontra sulle piante sviluppate, preferibilmente sulle foglie più alte, consiste nell’aspetto a forma di cucchiaio di foglie, corrispondente ad un leggero arricciamento, mentre in Potato leaf roll virus si osservano rotture e fessurazioni fogliari. Viene trasmesso dagli afidi con modalità non persistente ed ha una gamma limitata di ospiti.

24
Bibliografia
1. Bayne EH., Rakitina D.V, Morozov S.Y, Baulcombe DC, 2005. Cell-to-cell movement of potato potexvirus X is dependent on suppression of RNA silencing. Plant J. 44:471-482.
2. Becker DM, Fikes JD, Guarente L, 1991. A cDNA encoding a human CCAAT-binding protein cloned by functional complementation in yeast. Proc. Natl. Acad. Sci. USA 88:1968-1972.
3. Bleykasten-Grosshans C, Guilley H, Bouzoubaa S, Richards KE, Jonard G, 1997. Independent expression of the first two triple gene block proteins of beet necrotic yellow vein virus complements virus defective in the corresponding gene but expression of the third protein inhibits viral cell-to-cell movement. Mol. Plant-Microbe Interact. 10:240-246.
4. Boevink P, Oparka KJ, 2005. Virus-host interactions during movement processes. Plant Physiol. 138:1815-1821.
5. Bragg JN, Jackson AO, 2004. The C-terminal region of the barley stripe mosaic virus γb protein participates in homologous interactions and is required for suppression of RNA silencing. Mol. Plant Pathol. 5:465-481.
6. Bragg JN, Lim HS, Jackson AO, 2008. Hordeiviruses. In B. Mahy and M. van Regenmortel (ed.), Encyclopedia of virology, 3rd ed., in press. Elsevier Ltd., Oxford, United Kingdom.Google Scholar
7. Brakke MK, Ball EM, Langenberg WG, 1988. A non-capsid protein associated with unencapsidated virus RNA in barley infected with barley stripe mosaic virus. J. Gen. Virol. 69:481-491.
8. Chapman S, Hills G, Watts J, Baulcombe D, 1992. Mutational analysis of the coat protein gene of potato virus X: effects on virion morphology and viral pathogenicity. Virology 191:223-230.r
9. Cowan GH, Lioliopoulou F, Ziegle A, Torrance L, 2002. Subcellular localisation, protein interactions, and RNA binding of potato mop-top virus triple gene block proteins. Virology 298:106-115.r
10. Donald RGK, Lawrence DM, Jackson AO. 1997. The barley stripe mosaic virus 58-kilodalton βb protein is a multifunctional RNA binding protein. J. Virol. 71:1538-1546.
11. Donald RGK, Zhou H, Jackson AO, 1993. Serological analysis of barley stripe mosaic virus-encoded proteins in infected barley. Virology 195:659-668.r
12. Fancy DA, Melcher K, Johnston SA, Kodadek T, 1996. New chemistry for the study of multiprotein complexes: the six-histidine tag as a receptor for a protein crosslinking reagent. Chem. Biol. 3:551-559.
13. George AD, Tenenbaum SA, 2006. MicroRNA modulation of RNA-binding protein regulatory elements. RNA Biol. 3, 57–59.
14. Gerber AP, Herschlag D, Brown PO, 2004. Extensive association of functionally and cytotopically related mRNAs with Puf family RNA-binding proteins in yeast. PLoS Biol. 2:E79.
15. Gerber AP, Luschnig S, Krasnow MA, Brown P O, Herschlag D, 2006. Genome-wide identification of mRNAs associated with the translational regulator PUMILIO in Drosophila melanogaster. Proc. Natl. Acad. Sci. U.S.A. 103, 4487–4492.
16. Goodin MM, Austin J, Tobias R, Fujita M, Morales C, Jackson AO. 2001. Interactions and nuclear import of the N and P proteins of sonchus yellow net virus, a plant nucleorhabdovirus. J. Virol. 75:9393-9406.
17. Gorbalenya AE, Koonin EV, 1993. Helicases: amino acid sequence comparisons and structure-function relationships. Curr. Opin. Struct. Biol. 3:419-429.
18. Heinlein M, Epel BL, 2004. Macromolecular transport and signaling through plasmodesmata. Int. Rev. Cytol. 235:93-164.MedlineGoogle Scholar
19. James P, Halladay J, Craig EA. 1996. Genomic libraries and a host strain designed for highly efficient two-hybrid selection in yeast. Genetics 144:1425-1436.

25
20. Johnson JA, Bragg JN, Lawrence DM, Jackson AO, 2003. Sequence elements controlling expression of barley stripe mosaic virus subgenomic RNAs in vivo. Virology 313:66-80.r
21. Kalinina NO, Rakitina DV, Solovyev AG, Schiemann J, Morozov SY, 2002. RNA helicase activity of the plant virus movement proteins encoded by the first gene of the triple gene block. Virology 296:321-329.r
22. Karpova OV, Zayakina V, Arkhipenko MV, Sheval EV, Kiselyova OI, Poljakov VY, Yaminsky IV, Rodionova NP, Atabekov JG, 2006. Potato virus X RNA-mediated assembly of single-tailed ternary ‘coat protein-RNA-movement protein’ complexes. J. Gen. Virol. 87:2731-2740.
23. Keene JD, Tenenbaum SA, 2002. Eukaryotic mRNPs may represent posttranscriptional operons. Mol. Cell 9, 1161–1167.
24. Lauber E, Jonard G, K. Richards KE, Guilley H. 2005. Nonregulated expression of TGBp3 of hordei-like viruses but not of potex-like viruses inhibits beet necrotic yellow vein virus cell-to-cell movement. Arch. Virol. 17:1459-1467.Google Scholar
25. Lawrence DM, Jackson AO, 2001. Interactions of the TGB1 protein during cell-to-cell movement of Barley stripe mosaic virus. J. Virol. 75:8712-8723.
26. Lawrence DM, Jackson AO, 2001. Requirements for cell-to-cell movement of barley stripe mosaic virus in monocot and dicot hosts. Mol. Plant Pathol. 2:65-75.
27. Leshchiner AD, Solovyev AG, Morozov SY, Kalinina NO, 2006. A minimal region in the NTPase/helicase domain of the TGBp1 plant virus movement protein is responsible for ATPase activity and cooperative RNA binding. J. Gen. Virol. 87:3087-3095.
28. Li F, Ding S, 2006. Virus counterdefense: diverse strategies for evading the RNA silencing immunity. Annu. Rev. Microbiol. 60:503-531.r
29. Lucas WJ, 2006. Plant viral movement proteins: agents for cell-to-cell trafficking of viral genomes. Virology 344:169-184.r
30. MacDiarmid R, 2005. RNA silencing in productive virus infections. Annu. Rev. Phytopathol. 43:523-544.r
31. Melcher U, 2000. The ‘30K’ superfamily of viral movement proteins. J. Gen. Virol. 81:257-266.
32. Miller C A, Martinat MA, Hyman LE, 1998. Assessment of aryl hydrocarbon receptor complex interactions using pBEVY plasmids: expression vectors with bi-directional promoters for use in Saccharomyces cerevisiae. Nucleic Acids Res. 26:3577-3583.
33. Morozov, S. Y., and A. G. Solovyev. 2003. Triple gene block: modular design of a multifunctional machine for plant virus movement. J. Gen. Virol. 84:1351-1366.
34. Petty ITD, Jackson AO, 1990. Mutational analysis of barley stripe mosaic virus RNAβ. Virology 179:712-718.r
35. Petty ITD, French R, Jones RW, Jackson AO, 1990. Identification of barley stripe mosaic virus genes involved in viral RNA replication and systemic movement. EMBO J. 9:3453-3457.MedlineGoogle Scholar
36. Petty ITD, Hunter BG, Jackson AO, 1988. A novel strategy for one-step cloning of full-length cDNA and its application to the genome of barley stripe mosaic virus. Gene 74, 423–432.
37. Petty IT, Hunter BG, Wei N, Jackson AO, 1989. Infectious barley stripe mosaic virus RNA transcribed in vitro from full-length genomic cDNA clones. Virology 171:342-349.r
38. Samuels TD, Ju HJ, Ye CM, Motes CM, Blancaflor EB, Verchot-Lubicz J, 2007. Subcellular targeting and interactions among the potato virus X TGB proteins. Virology 367:375-389.r
39. Verchot J, Angell SM, Baulcombe DC. 1998. In vivo translation of the triple gene block of potato virus X requires two subgenomic mRNAs. J. Virol. 72:8316-8320.
40. Verchot-Lubicz J, 2005. A new cell-to-cell transport model for potexviruses. Mol. Plant-Microbe Interact. 18:283-290.MedlineGoogle Scholar
41. Verchot-Lubicz J, Ye CM, Bamunusinghe D, 2007. Molecular biology of potexviruses: recent advances. J. Gen. Virol. 88:1643-1655.

26
42. Waigmann E, Ueki S, Trutnyeva K, Citovsky V, 2004. The ins and outs of nondestructive cell-to-cell and systemic movement of plant viruses. Crit. Rev. Plant Sci. 23:195-250.
43. Wong SM, Lee KC, Yu HH, Leong WF, 1998. Phylogenetic analysis of triple gene block viruses based on the TGB 1 homolog gene indicates a convergent evolution. Virus Genes 16:295-302.
44. Wurch T, Lestienne F, Pauwels P. 1998. A modified overlap extension PCR method to create chimeric genes in the absence of restriction enzymes. Biotechnol. Tech. 12:653-657.
45. Zhou H, Jackson AO, 1996. Expression of the barley stripe mosaic virus RNA β “triple gene block”. Virology 216:367-379.













![340]) - Cochrane cochrane/Lavalle.pdf · 3) Clinical pathway con Care Manager specialistico in TI Generale 4) Clinical Pathway con Stroke Team Care (Care+Case Manager) Recidiva di](https://static.fdocumenti.com/doc/165x107/5e34198b33e43241b0647e1a/340-cochranelavallepdf-3-clinical-pathway-con-care-manager-specialistico.jpg)



