Bioermi a Rudiste nel Cretacico del Matese Orientale. · Sphaerucaprina, Neocaprina, Mitrocaprina,...
14
73 Bioermi a Rudiste nel Cretacico del Matese Orientale. Le Rudiste Singolari bivalvi sessili che vissero tra la fine del Giurassico e la fine del Cretacico, le Rudiste sono descritte esaurientemente in M.Mainelli, 2000, cui si riman- da per le opportune conoscenze. In questo contesto se ne considerano soltanto alcuni aspetti paleoecologici ed in parte filogenetici al fine di una più completa comprensione delle formazioni bioer- mali a Rudiste che si rinvengono nel Cretacico del Matese Orientale, scopo del libro. Sull’origine delle Rudiste ci sono ancora tanti aspetti non completamente accertati per la mancanza di testimonianze fossili che possano chiarirne la filogene- si. Gli attuali orientamenti fanno risalire le Rudiste al Giurassico superiore col genere Diceras che si diversifica in una ampia varietà di forme per tutto il Cretacico. Sono state formulate due ipotesi circa l’origine delle Rudiste. La prima, che si deve a H.Douvillè (1886; 1887; 1935), sostiene che l’origine delle Rudiste è attribuita al genere Pterocardium del Giurassico superiore, bivalve fisso al substrato per mezzo di una valva, la destra o la sinistra, per resistere, dice Douvillè, al moto violento delle correnti d’acqua che investivano gli individui cercando di rimuoverli dal loro ambiente di vita. La fissilità al substrato determina nel mollusco essenzialmente : - la crescita sproporzionata della valva fissa al substrato, - lo sviluppo abnorme dell’apparato mio-cardinale. Queste modificazioni, tra l’altro, si rilevano nel genere Diceras da cui derivano per evoluzione tutte le diversificazioni nelle famiglie delle Rudiste. La sistematica delle Rudiste desunta, seguita fino agli anni 50 del secolo scorso, espri- me le vedute del Douvillè. La seconda ipotesi si deve a C.Dechaseaux in J.Piveteau,1952, secondo cui l’origine delle Rudiste risale al genere Megalodon del Devoniano, da cui deriva Protodiceras del Lias e quindi Diceras del Giurassico superiore. Le modificazioni riguardano sempre, innanzitutto: - la crescita abnorme della valva fissa; - gli adattamenti dell’apparato mio-cardinale. La sistematica proposta da Dechaseaux esprime le vedute che si leggono in R.C.Moore, 1969, secondo cui le Rudiste, comprese nell’ordine Hippuritoida , si clas- sificano in 7 famiglie. Poiché le sistematiche suindicate non convincono pienamente nel 1959 V.Pchelintsev ne elabora una nuova classificando le Rudiste in 2 sottordini, Dextrodonta e
Transcript of Bioermi a Rudiste nel Cretacico del Matese Orientale. · Sphaerucaprina, Neocaprina, Mitrocaprina,...

73
Bioermi a Rudiste nel Cretacico del Matese Orientale.
Le Rudiste
Singolari bivalvi sessili che vissero tra la fine del Giurassico e la fine delCretacico, le Rudiste sono descritte esaurientemente in M.Mainelli, 2000, cui si riman-da per le opportune conoscenze. In questo contesto se ne considerano soltanto alcuni aspetti paleoecologici ed inparte filogenetici al fine di una più completa comprensione delle formazioni bioer-mali a Rudiste che si rinvengono nel Cretacico del Matese Orientale, scopo del libro.
Sull’origine delle Rudiste ci sono ancora tanti aspetti non completamenteaccertati per la mancanza di testimonianze fossili che possano chiarirne la filogene-si.Gli attuali orientamenti fanno risalire le Rudiste al Giurassico superiore col genereDiceras che si diversifica in una ampia varietà di forme per tutto il Cretacico.
Sono state formulate due ipotesi circa l’origine delle Rudiste.
La prima, che si deve a H.Douvillè (1886; 1887; 1935), sostiene che l’origine delleRudiste è attribuita al genere Pterocardium del Giurassico superiore, bivalve fisso alsubstrato per mezzo di una valva, la destra o la sinistra, per resistere, dice Douvillè,al moto violento delle correnti d’acqua che investivano gli individui cercando dirimuoverli dal loro ambiente di vita.La fissilità al substrato determina nel mollusco essenzialmente :
- la crescita sproporzionata della valva fissa al substrato,- lo sviluppo abnorme dell’apparato mio-cardinale.
Queste modificazioni, tra l’altro, si rilevano nel genere Diceras da cui derivano perevoluzione tutte le diversificazioni nelle famiglie delle Rudiste.La sistematica delle Rudiste desunta, seguita fino agli anni 50 del secolo scorso, espri-me le vedute del Douvillè.
La seconda ipotesi si deve a C.Dechaseaux in J.Piveteau,1952, secondo cui l’originedelle Rudiste risale al genere Megalodon del Devoniano, da cui deriva Protodicerasdel Lias e quindi Diceras del Giurassico superiore.Le modificazioni riguardano sempre, innanzitutto:
- la crescita abnorme della valva fissa;- gli adattamenti dell’apparato mio-cardinale.
La sistematica proposta da Dechaseaux esprime le vedute che si leggono inR.C.Moore, 1969, secondo cui le Rudiste, comprese nell’ordine Hippuritoida , si clas-sificano in 7 famiglie.
Poiché le sistematiche suindicate non convincono pienamente nel 1959 V.Pchelintsevne elabora una nuova classificando le Rudiste in 2 sottordini, Dextrodonta e

74
La sistematica delle Rudiste proposta da M.Mainelli, 1990, tenendo conto di tutte levedute formulate, rettificando alcune inesattezze tassonomiche, emendandoPchelintsev, 1959, distingue l’ordine Rudistae nei sottordini Dicerasodonta,Dextrodonta e Sinistrodonta, nei quali si classificano 10 famiglie.La sistematica proposta da M.Mainelli, 1990, non affronta il problema delle originirimandandone la soluzione a quando si potrà disporre di campionature più comple-te adatte per una ricostruzione filogenetica. La classificazione delle Rudiste, elaborata secondo le vedute di M.Mainelli, 1990, èriportata in M.Mainelli, 2000.
Le Rudiste nel Cretacico del Matese Orientale.
Secondo le conoscenze attuali nel Matese Orientale le Rudiste sono presenti con 6famiglie distinte in 31 generi, come da seguente schema:
Famiglie Sottofamiglie Generi
Requienidae Requienia, Apricardia, Toucasia
Monopleuridae Monopleura, Himerelites.
Caprotinidae Caprotina, Sellaea
Caprinidae Caprina, Caprinula,Ichthyosarcolites,
Mathesia, Mitrocaprina, Neocaprina,Offneria, Schiosia, Orthoptychus,Plagioptychus, Sphaerucaprina
Radiolitidae Radiolitinae Radiolites, Eoradiolites,Gorjanovicia,
Praeradiolites, Sphaerulites
Biradiolitinae Biradiolites, Bournonia, Distefanella
Sauvagesinae Sauvagesia, Durania
Lapeirousinae Lapeirousia, Katzeria
Hippuritidae Hippurites, Hippuritella, Vaccinites
La loro distribuzione stratigrafica nella Piattaforma Carbonatica del Matese Orientale

75
Per il Matese Orientale
- la famiglia Requienidae è presente dal Barremiano al Santoniano con i generiRequienia (Barremiano-Santoniano) a Civita e al Palumbaro di Pietraroja, a Piscina Culdi Bove, Sbregavitelli, su Monte La Costa e Colle Salva Signore, Toucasia (Barremiano-Aptiano) a Civita di Pietraroja e su Monte La Costa, Apricardia (Cenomaniano-Turoniano) su Monte La Costa e su q.916;
- la famiglia Monopleuridae è presente dal Barremiano al Turoniano con i generiMonopleura (Barremiano-Turoniano) estesa in tutto il Matese Orientale, Himerelites(Cenomaniano) su Monte La Costa, Aria dell’Orso, La Gallinola e a Campo Puzzo;
- la famiglia Caprotinidae è presente dal Barremiano al Coniaciano con i generiCaprotina (Barremiano-Coniaciano), Sellaea (Cenomaniano) che si rinvengono inparticolare su Monte La Costa, Aria dell’Orso, Campo Puzzo-La Costa Alta, LaGallinola;
- la famiglia Caprinidae presente dall’Aptiano al Campaniano con i generi Caprina(Aptiano-Campaniano) estesa in tutto il Matese Orientale, Caprinula, Ichthyosarcolites,Sphaerucaprina, Neocaprina, Mitrocaprina, Orthoptychus (Cenomaniano) che si rin-vengono su Monte La Costa, Colle Salva Signore, Aria dell’Orso, Campo Puzzo-CostaAlta, Mathesia (Aptiano superiore) a Sbregavitelli e su La Gallinola, Schiosia, Offneria(Aptiano superiore- Cenomaniano) su Monte La Costa;
- la famiglia Radiolitidae presente dal Barremiano al Campaniano nelle sottofami-glie
Radiolitinae con i generi
Radiolites (Cenomaniano-Campaniano) in particolare su Monte La Costa, Monte S.Polo, Colle Salva Signore, Colle Morotti, Quota 916, Colle Antona, La Torretta, CostaChianetta e Costa S.Angelo, Campo Puzzo-Costa Alta, La Gallinola, Eoradiolites(Albiano-Turoniano) su Monte La Costa, Aria dell’Orso dell’Orso, La Gallinola,Gorjanovicia (Santoniano-Campaniano) su Costa Chianetta e Costa S.Angelo,Praeradiolites (Albiano-Campaniano) su Monte La Costa, Aria dell’Orso, Sphaerulites(Aptiano-Turoniano) su Colle Salva Signore;
Biradiolitinae con i generi
Biradiolites (Turoniano-Campaniano) su Monte S. Polo, Colle Salva Signore, ColleMorotti, Colle Antona, Monte La Torretta, Bournonia (Turoniano-Campaniano) su

76
Colle Salva Signore, Distefanella (Turoniano-Campaniano) su Monte S. Polo, ColleSalva Signore, Quota 916,
Sauvagesinae con i generi
Sauvagesia e Durania (Albiano-Campaniano) su Monte La Costa, Monte S.Polo, ColleSalva Signore, Colle Morotti, Quota 916, Campo Puzzo-Costa Alta, La Gallinola, ColleAntona, Monte La Torretta, Costa Chiavetta- Costa S.Angelo,
Lapeirousinae con i generi
Lapeirousia e Katzeria (Santoniano-Campaniano) su Colle Salva Signore, CostaChianetta;
la famiglia Huippuritidae con i generi
Hippurites, Hippuritella e Vaccinites (Turoniano-Campaniano) in tutto il Cretacicosuperiore del Matese Orientale e in particolare su Monte S.Polo, Colle Salva Signore,Colle Morotti, Quota 916, Costa Alta, Colle Antona, Monte La Torretta, area diGuardiaregia-S.Nicola-Ponte Arcichiaro.
Paleoecologia.
Le Rudiste, in quanto molluschi bivalvi, erano organismi colonizzatori del ben-thos neritico delle piattaforme carbonatiche tropicali della Tetide cretacica.La loro notevole diversità morfologica può considerarsi, inoltre, il fenomeno biologi-co evolutivo unico per plasticità genotipica e fenotipica nell’ambito della classe deiBivalvi, con adattamenti a tutte le zonazioni e relative unità morfo-ecologiche dellepiattaforme carbonatiche Tetisiane cretaciche.
Dal punto di vista dell’evoluzione le Rudiste possono essere distinte in due gruppi.
L’uno, quello primitivo, comprende le famiglie Diceratidae, Epidiceratidae,Plesiodiceratidae, Heterodiceratidae, Requienidae, Monopleuridae, Caprotinidae, lequali si riconoscono essenzialmente
- per la tendenza alla conservazione dei caratteri, specialmente quelli dellaforma generale;
- per le piccole variazioni nell’apparato mio-cardinale;- per la scarsa adattabilità a tutti gli ambienti neritici.
L’altro gruppo, cui appartengono le famiglie Caprinidae, Radiolitidae e Hippuritidae,
Genotipo = complesso dei geni che costituiscono il patrimonio ereditario di ciascun individuo nel-l’ambito del patrimonio genetico della specie cui appartiene.Fenotipo = complesso dei caratteri di un individuo determinato dalla interazione del genotipo conl’ambiente.

77
manifesta un grande adattamento a tutti gli ambienti neritici e una grande variabilità- nella forma generale;- nell’apparato mio-cardinale;- negli elementi relativi alla cavità ventrale, l’ornamentazione, l’apparato sifona-
le, il ligamento;- nella chiusura delle valve;- nella grandezza.
Alcuni individui di Rudiste vivevano solitari, altri amavano essere gregari.I gregari formavano comunità, pseudocolonie secondo alcuni AA., ovvero popola-zioni viventi in un biotopo, oppure in una sinusia.
Le Rudiste solitarie che vivevano in un’area potevano essere o non essere della stessa spe-cie, quelle gregarie potevano convivere anche con pochi individui di altre specie.Nelle comunità gli individui potevano essere
- l’uno affiancato all’altro, in aderenza per mezzo della valva fissa con alcuni spazicomuni limitati;
- l’uno separato dall’altro nelle valve fisse con spazi ampi tra gli individui.Nell’uno e nell’altro caso gli spazi erano occupati da sedimento sindeposizionale,ovvero che si depositava mentre gli individui erano in vita.La comunità era monospecifica se costituita da individui di una sola specie, multi-specifica se costituita da individui appartenenti a più specie.Nella comunità multispecifiche una specie era sempre dominante sulle altre pernumero di individui.
I resti delle Rudiste gregarie si rinvengono oggi in formazioni bioermali.
Come in tutti i Bivalvi, anche nelle Rudiste l’apparato filtratore, che non si ritrova fossiliz-zato, doveva essere sviluppato con un sistema di ciglie vibratili, collocate nella parte supe-riore della parete interna della cavità palliale.
A valve aperte le ciglia, con il loro continuo movimento vibratile, producevano lecorrenti d’acqua che portavano i nutrienti all’apparato boccale.
L’evoluzione è il fenomeno del divenire nel tempo della materia vivente e non vivente.La biosfera, sottile involucro della Terra dove si svolge la vita, è soggetta al fenomeno dell’evoluzione.L’evoluzione chimico-fisica si fonda sul concetto che la Terra è un sistema dinamico i cui componenti, inte-ragenti, mutano nel tempo.L’evoluzione biologica studia l’origine e la diversificazione della vita che interagisce con la crosta terrestre.
Biotopo = parte fisica di un ecosistema .Ecosistema = ambiente fisico e vivente di un’area.Habitat = biotopi nei quali un organismo può vivere.Sinusia = popolazione di un habitat dentro il biotopo.
Autoecologia = studio dei rapporti che intercorrono tra gli organismi nel loro ambiente di vita.Sinecologia = studio del rapporto che intercorre tra l’organismo e l’ambiente di vita.Demoecologia = studio delle popolazioni di organismi.

78
Quando le valve si chiudevano e si arrestava il movimento delle ciglia l’acqua nonentrava più.Funzionava allora l’apparato sifonale formato dal sifone branchiale, E, (dal franceseentrèe = entrata), per immettere nella cavità palliale l’acqua di respirazione, e dal sifo-ne anale, S (dal francese sortiè= uscita) per emettere dalla cavità palleale l’acqua dirifiuto.
Nelle comunità a Rudiste in ogni individuo l’apparato sifonale era disposto in modotale da non interferire con la funzione respiratoria di altri individui che gli vivevanoaccanto, aderenti nella maggior parte dei casi.Nei bioermi si può constatare fossilizzata la collocazione dell’apparato sifonale di cia-scun individuo non interferente con quello degli altri.
Le Rudiste della Piattaforma Carbonatica del Matese Orientale presentano la seguen-te distribuzione paleoecologica.
I Requienidi e i Monopleuridi formavano in genere comunità monospecifi-che in ambienti subtidali lagunari aperti, protetti, costituiti da fondali fangosi o sab-biosi a energia delle acque da moderata a bassa ma a buon ricambio d’acqua.
I Requienidi raramente vivevano solitari e si associavano spesso a Radiolitidi e aCaprinidi in ambienti di scogliera ecologica.In genere Requienidi e Monopleuridi costruivano estese formazioni biostromali, conresti per la maggior parte in posizione di vita e con resti in tanatocenosi sedimenta-ti caoticamente.
I Caprotinidi vivevano in comunità monospecifiche e multispecifiche di lagu-na subtidale aperta, prossima alla scogliera ecologica, su fondali sabbiosi a buonricambio di acqua, ovvero ad energia moderata delle acque; costituivano estese for-mazioni biostromali le quali si differenziavano dai depositi con resti in tanatocenosi.
I Caprinidi vivevano prevalentemente in ambienti subtidali di scogliera eco-logica, con acque ad energia medio-alta; erano Rudiste solitarie o aggregate in comu-nità in genere multispecifiche.I Caprinidi formavano bioermi.I loro resti oggi si rinvengono per la maggior parte in depositi di scogliera.
I Radiolitidi colonizzavano in genere gli ambienti subtidali di piattaforma aper-ta o ristretta, costituiti da fondali con sedimenti prevalentemente fangosi , con acquead energia da moderata a bassa e con una adattabilità notevole a fondali fangosi,ipersalini, a basso ricambio di acque.In fondali ristretti, ovvero in condizioni ambientali estreme, i Radiolitidi erano sog-getti al fenomeno del nanismo.
Apparato filtratore= apparato boccale, apparato palliale, ciglia vibratili, apparato sifonale.

Rudiste isolate
Individui solitari, monotipici, di Radiolites sp.
Individui solitari, politipici, di1) Hippurites nabresinensis2) Vaccinites sp.
79
1
2

80
Comunità a Rudiste
Individui di Hippulites colliciatus in aderenza
Individui di Distefanella salmoiraghii
non in aderenza

81
Ricostruzione di semplari di Rudiste della famiglia Radiolitidae.
Gli esemplari, in posizione di vita, presentano le ciglia vibratili in funzione perassumere nutrienti.
Le ciglia sono estroflessioni del mantello e fuoriescono attraverso la commessuradelle due valve solo quando sono in funzione.
Quando non sono in funzione le ciglia si ritraggono nella cavità palleale.
A ciglia ritratte funziona l’apparato sifonale costituita dal sifone branchiale E perassumere nutrienti e dal sifone anale S per espellere i rifuiti.
I sifoni quando sono in funzione fuoriescono attraverso la commessura dellevalve.
valva destra fissa ciglia vibratili valva sinistra libera

82
La funzione dell’apparato sifonale di ogni individuo non interferisce conquella degli altri individui della comunità (Esempio di Hippurites colliciatus).
Aggregzione semplice negli Hippuritidi (sempio di bouquet)
Base di attacco dell’aggreagazione
E
E
E
EE
E
E
E
E
S
S
S
S S
S
S
SS
S

83
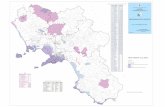
Siti dei bioermi a Rudiste considerati nella Piattaforma Carbonaticadel Matese Orientale.
Sbregavitelli Monte La Costa
Cappella diruta Anime del PurgatorioMonte S. Polo,Colle Salva Signore

84
I Radiolitidi pertanto possedevano- una elevata adattabilità in ambienti estremi,- una elevata variabilità morfologica sia genotipica, sia fenotipica, selezionata
dalla variabilità ambientale.Alcuni generi di Radiolitidi (Sauvagesia, Durania) colonizzarono anche ambienti discogliera ecologica, come ad esempio nel Cenomaniano di Monte La Costa dove sirilevano comunità a Durania montislacostae Mainelli.
Gli Ippuritidi sono presenti nel Cretacico superiore, ovvero dal Turoniano alCampaniano, in comunità estese anche per centinaia di metri, riferibili adambienti di scogliera ecologica ed aree adiacenti, ad alto idrodinamismo.
Essenzialmente le comunità ad Hippuritidi occuparono tutto il NeriticoAperto che si determinò con l’evoluzione tettono-sedimentaria della piatta-forma.
La scogliera ecologica ed aree limitrofe, scarpata da una parte e laguna pros-simale dall’altra, si presentano colonizzate da dense comunità monospecifi-che innanzitutto e poi da comunità multispecifiche in secondaria importan-za. Le comunità costituirono estese formazioni bioermali.
Bioermi a Rudiste.
Le formazioni bioermali a Rudiste sono distribuite stratigraficamen-te in tutto il Cretacico; si riconoscono estesamente in quasi tutte le zonazio-ni morfo-ecologiche della piattaforma; interessano essenzialmente iCaprinidi, i Radiolitidi e gli Ippuritidi.
Quelle considerate sono distinte per siti.
Monte La Costa:
- Bioerma a Schiosia nicolinae Mainelli dell’ Aptiano superiore,- Bioerma multispecies del Cenomaniano superiore con le seguenti spe-
cie dominanti
. Durania montislacostae Mainelli,
. Caprina carinata Boehm,
. Caprina schiosensis Boehm,
. Caprinula distefanoi Boehm,
. Ichthyosarcolites bicarinatus Gemmellaro,
. “ tricarinatus Parona,

85
. “ cfr ensis Pocta,
. Schiosia paolae Mainelli,
. Toucasia danielae Mainelli.Cappella diruta Anime del Purgatorio:
- Bioerma a Hippurites colliciatus Woodward del Santoniano-Campaniano,
- Bioerma a Durania samnitica Parona del Coniaciano-Santoniano,- Bioerma a Biradiolites sp.Santoniano;
Monte S.Polo:
- Bioerma a Distefanella salmoiraghii Parona del Turoniano,- Bioerma a Distefanella bassanii Parona del Turoniano;
Colle Salva Signore:
- Bioerma a Sauvagesia nicaisei Coquand del Turoniano,- Bioerma a Vaccinites gosaviensis Douvillè del Santoniano,
Sbregavitelli:
- Bioerma a Mathesia tertiicolloquiirudistarum Mainelli dell’Aptianosuperiore

86
Distretto neritico: distinzioni
livello del mare durante l’alta marea
livello del mare durante la bassa marea
supratidale
intertidale
subtidale
circatidale
• neritico, da neritè = conchiglia marina
• distretto neritico = mare delimitato dalla riva fino a m. 200 di profondità, dove vivono i molluschi
• benthos neritico, da benthos = fondo, fondale neritico
• tidal (inglese) = zona marina influenzata dal mare
• supra, inter, sub, circa = prefissi per distinguere le sottozone.













![CARTE PENTRU PROFESIONI{TII CARE LUCREAZ~ CU P~RIN}II · 2018. 3. 31. · Via]a de zi cu zi ofer` p`rin]ilor [ansa de a se dezvolta \n rolul lor, s` fac` gre[eli din care s` \nve]e,](https://static.fdocumenti.com/doc/165x107/60ab3d26b445bd45646f0858/carte-pentru-profesionitii-care-lucreaz-cu-prinii-2018-3-31-viaa-de-zi.jpg)