
Ba 2003-1 Copertina - cisba.eu · Roberto Spaggiari [email protected] resp. di segreteria...
100
BIOLOGIA AMBIENTALE Centro Italiano Studi di Biologia Ambientale ISSN 1129-504X Volume 17 Numero 1 Gennaio 2003
Transcript of Ba 2003-1 Copertina - cisba.eu · Roberto Spaggiari [email protected] resp. di segreteria...

BIOLOGIAAMBIENTALE
CentroItalianoStudi diBiologiaAmbientale
ISSN 1129-504X
Volume 17
Numero 1
Gennaio 2003

Tipografia NUOVA FUTURGRAF, Via Soglia 1, REGGIO EMILIA
BIOLOGIAAMBIENTALE
PROPRIETÀ: Rossella Azzoni, Presidente del C.I.S.B.A.
DIRETTORE RESPONSABILE: Rossella AzzoniREDAZIONE:Giuseppe Sansoni [email protected] resp. di redazioneRoberto Spaggiari [email protected] resp. di segreteriaGilberto N. Baldaccini [email protected] redattorePietro Genoni [email protected] redattore
Pubblicazione del C.I.S.B.A., vol. 17, n. 1/2003Autorizzazione del Tribunale di Reggio Emilia n. 837 del 14 maggio 1993
Biologia Ambientale, viene inviata ai soci del Centro Italiano Studi di Biologia Ambientale (C.I.S.B.A).Per iscriversi o per informazioni: Centro Italiano Studi di Biologia Ambientale, via Amendola 2, 42100 Reggio EmiliaSegretario: Roberto Spaggiari, tel. 0522 336060 � 0335 7712847; fax 0522 330546; e-mail: [email protected]
www.cisba.it [email protected] annuali di iscrizione al Centro Italiano Studi di Biologia Ambientale: socio ordinario: E 40,00; socio collaboratoreE 30,00; socio sostenitore E 310,00. conto corrente postale n. 10833424 intestato a: CISBA, RE
Biologia Ambientale raccoglie e diffonde in-formazioni sulle tematiche ambientali, con parti-colare attenzione ai seguenti campi di interesse:
q Bioindicatori e biomonitoraggioq Ecotossicologiaq Depurazione delle acque reflueq Ecologia delle acque interne e dell’ambiente
marinoq Gestione dell’ambienteq Igiene ambientaleq Ecologia urbanaq Impatto ambientaleq Ingegneria naturalisticaq Rinaturazione e riqualificazione ambientaleq Conservazione della naturaq Ecologia del paesaggio
Biologia Ambientale è articolata in due sezioni:
Lavori Originali, in cui vengono pubblicati arti-coli e rassegne bibliografiche originali;
Informazione & Documentazione –sezione vol-ta a favorire la circolazione di informazioni e diidee tra i soci– in cui vengono riportate recensio-ni di libri, riviste e altre pubblicazioni nonché no-tizie e lavori già pubblicati ritenuti di particolareinteresse o attualità.
Comitato ScientificoRoberto ANTONIETTI
Dip. Scienze Ambientali, Univ. di ParmaNatale Emilio BALDACCINI
Dip. di Etologia, Ecologia, Evoluzione, Univ. di PisaRoberto BARGAGLI
Dip. Scienze Ambientali, Univ. di SienaAntonio DELL�UOMO
Dip. di Botanica ed Ecologia, Univ. di CamerinoSilvana GALASSI
Università dell�Insubria, ComoPier Francesco GHETTI
Dip. Scienze Ambientali, Univ. Cà Foscari, VeneziaStefano LOPPI
Dip. Scienze Ambientali, Univ. di SienaSergio MALCEVSCHI
Ist. Ecologia del territorio e degli ambienti terrestri,Univ. di Pavia
Maurizio G. PAOLETTIDip. di Biologia, Univ. di Padova
Luciano SANTINIDip. C.D.S.L. Sez. Entomologia agraria, Univ. di Pisa
Paolo Emilio TOMEIDip. Agronomia e gestione agroecosistema, Univ. di Pisa
Mariagrazia VALCUVIA PASSADOREDip. Ecologia del territorio e degli ambienti terrestri,Univ. di Pavia
Pierluigi VIAROLIDip. Scienze Ambientali, Univ. di Parma
Luigi VIGANÓIRSA - CNR, Brugherio MI
Sergio ZERUNIANParco Nazionale del Circeo, Sabaudia (LT)
Aldo ZULLINIDip. di Biotecnologie e Bioscienze, Univ. Milano Bicocca

Biologia Ambientale, 17 (1): 1-8, 2003
Il declino della biodiversità nella dimensione territoriale
Pignatti Sandro
Forum Plinianum, Via della Piana, 00185 Roma; fax 06 49917130; e-mail [email protected]
Pervenuto il 12.9.2002; accettato il 25.9.2002
RiassuntoNel monitoraggio degli ecosistemi naturali si rilevano frequentemente esempi di scomparsa di specie vegetali ed animali, tuttavia questieventi solo raramente risultano rilevabili attraverso dati statistici di valore globale. Infatti, attraverso la discussione di alcuni esempi, sidimostra che è facile accertare la presenza di una specie non osservata in precedenza, mentre è estremamente difficile dimostrarel’estinzione. Per questo motivo, i totali riguardanti i singoli paesi danno una visione ottimistica della realtà. Si propone pertanto dispostare l’obbiettivo del monitoraggio dalla scala globale o continentale a quella del territorio, di singoli ecosistemi e delle popolazioni. Imetodi di inventario floristico (recentemente estesi anche a singoli gruppi zoologici) permettono di rilevare una situazione drammatica; nelnostro paese sono documentabili casi nei quali l’impatto umano ha causato un grave declino della biodiversità: in qualche caso si arriva allascomparsa del 40 % della flora preesistente.
PAROLE CHIAVE: biodiversità / flora / estinzione / ecologia urbana
AbstractDecline of the biodiversity at the landscape scaleIn monitoring natural ecosystems, frequent examples of species (both animals and plants) disappearing were registered; indeed, it seemsrather difficult to give figures on a global scale. In fact, careful examination of some examples, shows that it is easy to assess the presenceof a species not previously observed, while it seems extremely difficult to demonstrate the contrary, i.e. extinction. For this reason, grosstotals of flora or fauna for large areas give an optimistic vision of reality. In consequence, the proposition is made to shift monitoring fromthe global or continental scale to the analysis of single landscapes, ecosystems and populations. The methods of floristic inventory(recently extended also to specific zoological groups) show a dramatic situation; in Italy some cases are documented in which humanimpact caused severe decline in biodiversity: sometimes up to 40 % of the original flora disappeared.
KEY WORDS: biodiversity / flora / extinction / urban ecology
1. INTRODUZIONEIl concetto di biodiversità, che per molto tempo era
rimasto nell’uso di pochi studiosi specializzati, vieneora reso accessibile al pubblico ed ai media. Dopo laconferenza di Rio (1992) si è infatti presa coscienzadel fatto che la biodiversità è una delle variabili piùsignificative per valutare lo stato della biosfera.
Oggi è opinione diffusa, almeno nel campo deglistudiosi di ecologia e biologia ambientale, che la biodi-versità sia minacciata dallo sviluppo tecnologico e
produttivo; la preoccupazione in questo senso ha por-tato alla stesura del protocollo di Rio che, appunto,impegna tutti i paesi alla salvaguardia della biodiversità.La biodiversità è minacciata dall’estinzione delle spe-cie, che si ritiene proceda con una continua accelera-zione e che, secondo stime autorevoli, potrebbe am-montare alla scomparsa di 350-1000 specie/anno, ingran parte insetti (WILSON, 1992). Molte specie anima-li, anche tra quelle di maggior significato simbolico, si

PIGNATTI S. - Il declino della biodiversità2
avvicinano al livello di estinzione, oppure lo sono statein tempi recenti: bisonte, tigre, rinoceronte, balene,scimmie antropoidi. Da questi dati si potrebbe conclu-dere che stiamo attualmente di fronte ad un esempioimponente di estinzione di massa.
In un libro recente (MYERS & SIMON, 1994) siconfrontano le opinioni di un ecologo (Myers) e di uneconomista (Simon). Il primo lancia l’allarme per labiodiversità; il secondo risponde con una puntigliosaenumerazione di dati: negli ultimi 50 anni la popola-zione mondiale è passata da 2 a 6 miliardi di umani,ma la produzione agricola è aumentata di più del triplocosì che ora la disponibilità di cibo pro capite èaumentata e, contemporaneamente, è aumentata ladurata della vita media, è calata la mortalità infantile,etc. Tutto questo viene dimostrato con dati che tuttiben conosciamo e che si possono ritenere statistica-mente affidabili. Ora, chiede l’economista, qual è ilprezzo pagato per questo innegabile progresso? Diquante specie si può documentare l’avvenuta estin-zione, mediante dati altrettanto validi sul piano stati-stico? E qui l’ecologo viene a trovarsi in difficoltà: èdifficile, spesso impossibile, dimostrare un’estinzio-ne ed eventualmente la perdita di poche specie notesoltanto allo specialista sembra un fatto trascurabiledi fronte ai bisogni dell’umanità.
Il significato della biodiversità per l’equilibrio dellabiosfera viene posto con autorevolezza da WILSON
(1992). Tra i molti esempi, questo Autore cita il casodi Centinela (Ecuador), dove su una catena montuosadi piccole dimensioni erano note 90 specie di pianteendemiche: in seguito alla trasformazione agricola,gran parte di queste non vengono ritrovate in seguito.Però non è possibile dare dati precisi e statisticamentevalidi sul numero delle specie effettivamente estinte:quante tra esse sopravvivono in popolazioni di pochiindividui, o magari in un orto botanico? Non sappiamo.
Dobbiamo dunque concludere che la temuta estin-zione di massa è solo una proiezione di una mentalitàcatastrofista?
Chi si occupa di biologia ambientale sa bene che nonè così e che il problema è reale; tuttavia sembraattualmente impossibile fornire dati numerici altrettan-to convincenti come quelli riguardanti i fenomeni so-cio-economici. Questo ha due motivazioni, tra lorostrettamente connesse: anzitutto, manca fino ad orauna definizione di biodiversità convincente e general-mente accettata; in secondo luogo, e come conse-guenza di ciò, manca un metodo per dare una valuta-zione numerica della biodiversità.
Da anni chi scrive si occupa di banche dati riguar-danti la flora: vediamo che cosa si possa ricavare daqueste per avvicinarsi ad una risposta a questo proble-ma.
2. LO STATO DELL’ARTEConsideriamo anzitutto il computo generale riguar-
dante le specie indicate per la flora italiana: ci si limitaalla flora vascolare (Tracheophyta), cioè pteridofite,gimnosperme ed angiosperme (monocotiledoni e dico-tiledoni), in quanto per gli altri gruppi (alghe, miceti,briofite, licheni) non si dispone di dati storici omoge-nei. L’ultima revisione generale effettuata in occasionedella pubblicazione della Flora d’Italia (PIGNATTI, 1982)dava i risultati esposti alla tabella I. Da questa si puòconstatare che il numero totale delle specie note per ilnostro territorio è aumentato durante 130 anni da 4309a 5599, cioè di 1290 specie, con un incremento pari acirca il 30 % rispetto alla cifra iniziale. Si tratta però diun dato poco significativo; infatti, come già osservatonel lavoro originale, non si tratta tanto di nuove specieentrate in Italia, quanto soprattutto di differenze neicriteri di valutazione del livello specifico utilizzati daivari Autori.
Consideriamo ora le differenze intercorse dopo lapubblicazione dell’opera nel 1982 e fino ad oggi. At-tualmente risultano indicate per l’Italia ben 224 speciein più, portando il totale generale a 5823; l’aumento, inpoco meno di 20 anni è del 4 %, ed il rateo risulta nonmolto diverso da quello prima calcolato su un periododi 130 anni. Va inoltre osservato che in questo caso icriteri utilizzati sono i medesimi, trattandosi dello stes-so Autore. La seconda edizione della Flora è in fase diavanzata redazione ed è verosimile che alla fine dellavoro il numero delle specie da aggiungere sarà anco-ra superiore. Da questo tuttavia non si può concludereche la biodiversità sia aumentata, con oltre 200 speciein più rispetto a venti anni fa. Infatti, se i dati vengono
Tab. I. Specie di piante vascolari segnalate per l’Italia(semplif. da PIGNATTI, 1982)
Autore data n° specie
Bertoloni 1833-54 4309Arcangeli 1894 4932Fiori 1923-29 3877Pignatti 1982 5599
Tab. II . Motivazioni per l’aggiunta di nuove specie alla Floraitaliana
numero %
Nuovo criterio tassonomico 135 64,6Ampliamento dell’areale 52 24,9Specie esotiche naturalizzate 22 10,5(non definibili: 15 specie)Totale 209 100,0

PIGNATTI S. - Il declino della biodiversità 3
analizzati e raggruppati per tipologie, si nota (Tab. II)che le nuove segnalazioni sono dovute in larga maggio-ranza (64,6 %) ai progressi degli studi biosistematici,oppure (24,9 %) all’ampliamento di areali di specie giànote: quasi sempre si citano campioni d’erbario, chedimostrano come queste specie fossero presenti già inpassato, ma non fossero state identificate corretta-mente. Le specie certamente nuove sono il residuo10,5 %, cioè le esotiche introdotte, direttamente oindirettamente, ad opera dell’uomo: 22 in tutto. Dun-que, in realtà lo stock biologico è aumentato di solo 22specie (0,4 %). È dubbio se questo possa venir consi-derato un aumento della biodiversità: si tratta di specieestranee al contesto della flora indigena, in alcuni casi(es. Arctotheca calendula) potenzialmente invasive.
L’approfondimento delle conoscenze biosistemati-che presuppone un’intensa laboriosità da parte dellacomunità scientifica: in effetti questo aspetto caratte-rizza l’attività in campo floristico dopo la pubblicazio-ne della Flora, nella quale è data l’indicazione di pre-senza/assenza a livello regionale, mettendo così inevidenza vistose lacune geografiche. È stata costituitauna banca dati comprendente le segnalazioni degliultimi venti anni di specie da ritenere nuove per la floradelle singole regioni italiane. Il numero di questi ritro-vamenti, accorpato in classi triennali, mostra una chia-ra tendenza verso l’incremento, che si deduce dai datidella tabella III. Approfondendo l’analisi di questi dati,sono stati separati i ritrovamenti di specie esotiche,rispetto a quelli dovuti all’allargamento di areali dispecie già note per altre regioni. Anche in questo casosi ha un’incidenza del 10 %, che mostra una rimarche-vole rispondenza con il dato della tabella II.
Per quanto riguarda il problema delle estinzioni, nonsi può affermare che esistano esempi accertati di estin-zioni avvenute nel periodo considerato. I casi nei qualiuna specie va esclusa dalla Flora italiana sono quasisempre riconducibili all’approfondimento di conoscen-ze biosistematiche, come sopra indicate, e per lo più laspecie eliminata viene rimpiazzata da un’altra speciecongenere, quindi il tutto si risolve in cambio di nome,ma la consistenza dello stock biologico rimane invariata.
Un discorso più ampio riguarda la validità di questidati, nei quali si ha a che fare con un totale generale(gross total): infatti essi tendono a dare un arrotonda-mento per eccesso. Quando si afferma che la florad’Italia nel 1982 era composta da 5599 specie di piantevascolari, ciò significa che tale era il numero dellespecie delle quali, in epoca antecedente, era stata ac-certata la presenza; però in moltissimi casi, soprattuttoper specie rare, non era stato possibile verificare seesse fossero ancora effettivamente presenti al momen-to della pubblicazione dei dati. Va ricordato che alcunesegnalazioni risalgono al sec. XVIII e non sono stateripetute successivamente.
Da questi dati si può giungere ad una prima conclu-sione parziale: nei vent’anni trascorsi lo stock biologi-co della flora italiana è rimasto sostanzialmente invaria-to, mentre si è avuto un aumento abbastanza cospicuodelle conoscenze scientifiche riguardanti la flora. Que-sto risultato sembra poter smentire la preoccupazioneper il deperimento della flora.
3. ESTINZIONE, EFFETTIVA O PRESUNTASi possono qui esaminare criticamente alcuni casi
ben noti e documentati di estinzione, che tuttavia sisono rivelati soltanto dei casi di morte presunta.
Cheilanthes persica – Una piccola felce ad ampiadistribuzione (Asia Occidentale, Balcani) nota in Italiaper un’unica popolazione sul M. Mauro nell’Appenni-no Romagnolo presso Faenza. Ben nota nel secoloscorso, ne esiste ricco materiale conservato in varierbari; una ricognizione effettuata negli anni ’60 hadato esito negativo e se ne è dedotto che fosse estinta,opinione accettata anche in PIGNATTI (1982). Ricerchesuccessive nello stesso posto su rupi poco accessibilihanno chiarito che ne esiste tuttora un’abbondantepopolazione. Va osservato che comunque l’estinzionesarebbe stata relativa soltanto al territorio italiano, inquanto la sopravvivenza della specie è assicurata dallepopolazioni del mediterraneo Orientale.
Genista holopetala – Arbusto nano della flora illiri-ca, molto raro; la sola popolazione italiana di M. Spac-cato presso Trieste, ben nota e documentata, è statadistrutta nel 1943 da un incendio. Dopo le modifiche alconfine orientale non ne rimanevano individui su terri-torio italiano, però in seguito è stata nuovamente os-servata nei pressi di Trieste. Anche in questo casol’estinzione eventuale sarebbe stata soltanto relativaalla flora italiana: in Slovenia e Croazia se ne conosco-no altre popolazioni.
Ionopsidium thalianum – Nota per un’unica popo-lazione di poche decine di individui, nella Maremma: èspecie annuale, dunque con ridotta capacità di soprav-vivenza, endemica, e pertanto la scomparsa di questapopolazione avrebbe il significato di una perdita irre-
Tab. III . Numero di rilevanti scoperte floristiche, per le singoleregioni italiane, in serie triennale (su 409 dati, pari a ¼ del totale)
anni n°
1978-80 321981-83 721984-86 531987-89 441990-92 661993-95 561996-98 86

PIGNATTI S. - Il declino della biodiversità4
versibile. Già prima del 1982, ricerche più estese neavevano individuato altre due popolazioni, poi seguiteda altri rinvenimenti, anche in Umbria e Lazio.
Drosera anglica – Le droseracee, piante carnivoredella flora australe, nell’emisfero boreale sono ridotte asole tre specie esclusive delle torbiere a sfagni inambiente oligotrofo. Durante l’ultima fase fredda po-stwürmiana dovevano essere frequenti sulle Alpi, nel-l’alta Pianura Padana e sparse anche nella Penisola;oggi se ne conoscono poche popolazioni, tutte a ri-schio a causa dello sfruttamento delle torbiere, deldrenaggio e dell’eutrofizzazione. Di D. anglica nel1982 erano accertate solo 18 popolazioni, dalla Carniaal Piemonte; in seguito ne sono state identificate altretre, però non sappiamo quante delle popolazioni prece-dentemente indicate siano ancora esistenti, forse due otre soltanto. Comunque si tratta di specie diffusa nellezone fredde dell’Eurasia.
Centaurea cyanus e Agrostemma githago – Fiordali-so e gittaione, due specie con fiori appariscenti che untempo (fino al 1950 ed oltre) caratterizzavano la vege-tazione commensale delle colture di frumento in Pie-monte e Lombardia, con fioriture di grande bellezza,oggi scomparse. Il fiordaliso è ancora sporadicamente
presente in ambienti di agricoltura marginale sull’Ap-pennino, mentre il gittaione è ormai una rarità. Non siconoscono le cause della scomparsa di queste duespecie, ma si può supporre che essa sia collegataall’aratura meccanica ed all’uso di fertilizzanti di sintesi.
Altri esempi di “piante redivive” sono Trifoliumlatinum e Vicia pimpinelloides della flora romana,descritte nei primi decenni del sec. XIX, non ritrovatesuccessivamente e date per estinte (però ne esistonopopolazioni nel Mediterraneo orientale!); tuttavia, me-diante accurate ricerche, negli ultimi anni sono statenuovamente accertate.
Inverso è il caso di Cistus laurifolius: in Italiasoltanto due popolazioni a Bibbiena e sui Colli Euganei;la seconda, di cui esistono campioni che risalgono al1818, non è stata ritrovata in seguito e va consideratadefinitivamente scomparsa.
Merita qui ricordare la vicenda del Krakatau, la cuiflora è stata completamente annientata dalla catastrofedel 1883, ma si è in seguito rapidamente ricostituita perintroduzione passiva dalle isole vicine ed oggi contacirca 500 specie. La ricostruzione della copertura ve-getazionale, avvenuta in maniera del tutto spontanea,dimostra le straordinarie capacità di recupero del-l’ecosistema naturale.
L’analisi di questi casi (ai quali se ne potrebberoaggiungere molti altri) porta ad un riesame critico delconcetto di estinzione. Va osservato che, se da un latoè facile dimostrare la presenza di una specie, mediantel’osservazione della stessa in popolazioni naturali, èinvece molto difficile dare la prova dell’assenza di unaspecie, e questo è il caso dell’estinzione. Infatti, unaricerca che dia risultato negativo non è sufficiente peraffermare che una specie sia effettivamente scompar-sa in maniera irreversibile: essa può essere rimastainosservata, oppure essere confinata in un’area inac-cessibile, esser presente con semi o rizomi nascosti nelsuolo, ma in grado di germinare quando le condizioniesterne siano favorevoli, ecc. La prova dell’estinzionesi può raggiungere soltanto quando una specie è notaper un’unica popolazione, strettamente legata ad undeterminato biotopo, che per cause esterne è andatocompletamente annientato. Si tratta di casi abbastanzaeccezionali. Dunque, il gross total tende a sopravaluta-re le presenze e sottovalutare le estinzioni. Questepossono venire realmente dimostrate soltanto a distanzadi decenni, quando è ormai troppo tardi per intervenire.
4. VARIAZIONI DELLA BIODIVERSITÀSU SCALA TERRITORIALE
Gli esempi esposti in precedenza riguardano casi distudio che assumono un significato se vengono esami-nati nel contesto dell’intera flora italiana; consideriamoora altri casi che appaiono rilevanti soprattutto a livello
Fig. 1. Esempi di specie ritenute estinte in Italia, ma rinvenute insuccessive indagini.
Cheilanthes persica Drosera anglica
Centaurea cyanus Agrostemma githago

PIGNATTI S. - Il declino della biodiversità 5
territoriale.Un esempio classico di diminuzione della biodiversi-
tà, relativo all’ittiofauna, è quello del Lago Victoria: ilgruppo dei ciclidi era rappresentato fino al 1980 dacirca 300 specie endemiche, un esempio unico diradiazione adattativa, basata essenzialmente sulle diffe-renti abitudini alimentari e riproduttive. Dopo l’intro-duzione nel lago della perca del Nilo, pesce predatore,un gran numero di specie è stato annientato. Ad esem-pio, su un’area di controllo “scomparvero più di 80delle 123 specie, pari al 70 %” (GOLDSCHMIDT, 1994).
Possiamo dunque chiederci se, nell’ambito dellaflora italiana, esistano casi analoghi di scomparsa dispecie. Vengono scelti esempi, nei quali una diminu-zione della biodiversità, rappresentata dalla scomparsadi specie vegetali, è un fatto evidente.
Il caso ZannoneSul nostro territorio l’unico caso che permette una
comparazione diretta è l’isola di Zannone, al largo dellacosta laziale, di 1 km2 di superficie ed alta 194 m, privadi abitanti e gestita come riserva naturale. L’isola èstata studiata in tre occasioni ottenendo i seguentirisultati numerici (MENEGONI, dati ined.):
anno del rilevamento specie n°1902 2721954 2561991 198
La diminuzione nel numero di specie è evidente(circa il 25 % in novant’anni), ma sulle sue cause sipossono soltanto fare ipotesi: potrebbe trattarsi di ungenerale inaridimento del clima, oppure delle conse-guenze del pascolo dei mufloni, introdotti da pochidecenni.
La Pianura PadanaL’intera fascia planiziare è completamente trasfor-
mata ad opera dell’uomo. La Pianura Padana avevauna vegetazione naturale con boschi e paludi fino allaconquista romana nel III sec. a. C. Da quel momento èstata in gran parte colonizzata, ma dopo il crollo del-l’Impero è stata abbandonata e nell’alto medioevo eranuovamente occupata da boschi naturali o seminatura-li. La colonizzazione successiva è stato un processo
lento, completato nel sec. XVIII per le aree con buondrenaggio naturale, mentre le aree soggette alle pienedei fiumi sono state bonificate solo dopo il 1900.Rimangono pochi lembi di bosco, conservatisi quasiper caso, che forniscono un quadro orientativo diquello che poteva essere il popolamento naturale dellapianura. Per un paragone si possono utilizzare i datirecenti (rilievi degli ultimi trent’anni) di inventario flo-ristico. La tabella IV sintetizza i dati di due esempi: ilBosco Olmé (prov. di Venezia: Fig. 2) ed il bosco diMuzzana (prov. di Udine: Fig. 3).
Nel primo caso (su dati originali inediti) le speciecensite sull’area boscata sono 357, mentre nelle areeagricole circostanti si scende ad una media di 153specie con una diminuzione di oltre il 50 %. Per ilsecondo caso (da POLDINI, 1991) sono state utilizzatearee di superficie maggiore, quindi i totali sono piùelevati, tuttavia il fenomeno è analogo: anche qui ladiminuzione nel numero di specie presenti sulle areedisboscate è prossima al 50 %. Siamo certamente difronte ad un fenomeno generale, che potrebbe venireverificato in altri distretti intensivamente coltivati d’Eu-ropa (Languedoc, Olanda, Westfalia, Valacchia).
La variazione rispetto allo stato naturale è quantitati-va, ma contemporaneamente anche qualitativa. La com-posizione della flora è nei due casi molto differente.Nella pianura coltivata in maniera intensiva si addensa-no le specie a distribuzione cosmopolita (45-57 suciascuna area) mentre le endemiche scendono a sola-mente 1-3; al contrario, sulle vicine Prealpi Carniche(alta Val Meduna) le cosmopolite scendono a meno di15 mentre le endemiche salgono a 21-27. Dunque,nella pianura coltivata non solamente si perde circa lametà della flora, ma il rimanente è costituito per un 20-25 % da specie ad ampio areale, dunque non stretta-mente collegate a questo ecosistema.
Il caso Paludi PontineLa flora di quest’area, studiata da Béguinot negli
anni ’30, prima della realizzazione del parco Nazionaledel Circeo, riflette le condizioni naturali di questoambiente (oggi indicato come Agro Pontino). Nel set-tore planiziare bonificato e messo a coltura è statoeffettuato dopo il 1990 un inventario floristico (osser-vazioni personali) su aree standard di 35 km2, con i
Tab. IV . Numero di specie registrate in boschi planiziali relitti e in aree limitrofe.
n° specie med. min. max. calo %
Bosco Olmé (1 area di 35 km2) 357Aree limitrofe (6 su 35 km2) 153 121 205 57,1
Bosco Muzzana (2 aree di 140 km2) 534Aree limitrofe (4 su 140 km2) 272 235 314 49,1

PIGNATTI S. - Il declino della biodiversità6
seguenti risultati:aree in condizioni dopo la diminuzione
naturali bonifica %Specie n° 400 180 55
Anche in questo caso il calo floristico è certamentecausato dalla distruzione dell’ecosistema naturale e suasostituzione con l’agroecosistema.
In questi casi esistono ancora le vestigia dell’ecosi-stema naturale, che è possibile utilizzare per un con-fronto; ma purtroppo abbiamo anche biotopi nei qualilo stato naturale è ormai scomparso irreversibilmente.Il Bosco Ramitelli presso Termoli oppure quello di
Policoro in Basilicata o quello di Rosarno: nel sopral-luogo del 1951 l’autore, ancora studente, constatò cheessi, in conseguenza della riforma agraria, erano statitagliati pochi anni prima e non esistevano più (datiined.). Non ne abbiamo una descrizione moderna, lastruttura e composizione di questi monumenti millenariè perduta per sempre.
5. VARIAZIONI DELLA BIODIVERSITÀALLA SCALA DI COMUNITÀ
Non sempre l’impatto umano sugli ecosistemi natu-rali ha l’effetto di ridurne la diversità. Nell’ambientemediterraneo è anzi normale avvenga il contrario, al-meno per quanto riguarda la vegetazione. Le associa-zioni climax, come il caso tipico della lecceta, sonocostituite da un numero ridotto di specie (PIGNATTI,1988), circa una sessantina (comprese quelle rare edaccidentali), mentre nella macchia e nella gariga, che siformano come conseguenza di pascolo ed incendio, siha una flora di 300-400 specie. Anche la flora sinan-tropica è molto ricca di specie: nell’area urbana diRoma, all’interno del Raccordo Anulare, sono statecensite 1285 specie (CELESTI GRAPOW, 1995), cioèoltre 1/5 della flora d’Italia. Tuttavia anche in questocaso si hanno modificazioni importanti. Vengono diseguito analizzati alcuni casi esemplificativi.
Aree archeologiche a RomaL’analisi della flora è stata effettuata su aree di
saggio di 1,6 km2 di superficie (CELESTI GRAPOW et al.,1994; CELESTI GRAPOW, 1995). Sono stati distinti treambienti principali, ciascuno individuato mediante trearee (Tab. V): aree archeologiche, aree che includonorelitti di vegetazione forestale ed aree intensamenteurbanizzate. Le aree archeologiche e quelle con vege-tazione forestale hanno circa la medesima densità flori-stica (361-374 specie in media), mentre in quelle urba-nizzate si scende a 173, con un calo del 53 %, moltovicino a quello osservato nella pianura friulano-veneta.Molto interessante però è la variazione nella concentra-zione di nutrienti nel suolo (rilevata mediante indicatorivegetali, utilizzando gli indici di ELLENBERG, 1974):l’indice medio delle specie presenti nella vegetazioneforestale è di 4,12, risulta un poco inferiore nelle aree
Tab. V. Relazione tra aumento dei nutrienti e decadimento della flora in Roma
specie presenti indicatori di nutrienti*min med max min med max
Aree archeologiche 367 374 381 3,72 3,87 3,99
Aree con ambienti forestali 265 361 450 4,0 4,12 4,19
Aree intensamente costruite 162 173 184 4,34 4,46 4,66
* media degli indici di ELLENBERG (1974) calcolata su tutte le specie presenti
Fig. 2. Numero di specie registrate in 10 quadranti di 35 km2
nella Pianura veneta (dati originali). Il quadrante grigio comprendeil bosco di Olmé
Rustigné Oderzo Motta di Annone PradipozzoLivenza Veneto
Callalta Priuli Cessalto S. Stino di BonazzaLivenza
121 127 140 141 144
171 178 357 205 non rilevato
Fig. 3. Numero di specie registrate su 6 quadranti di 150 km2 inFriuli (da POLDINI , 1996). I due quadranti grigi comprendono ilbosco di Mezzana.
Dignano Campoformido
251 314
Codroipo Talmasson
Cordovado Muzzana
235 288
448 620

PIGNATTI S. - Il declino della biodiversità 7
archeologiche (ma si tratta di differenza scarsamentesignificativa), mentre aumenta decisamente (4,46) nel-le aree urbanizzate.
La pineta di CastelporzianoLa “duna antica” è un’area pianeggiante ricoperta
dal bosco misto di caducifoglie, del quale sono statianalizzati numerosi esempi. Su circa 5 km2 è statoeffettuato attorno al 1900 un impianto con Pinus pineaper la produzione dei pinoli. La comparazione tra leflore delle aree con vegetazione naturale e delle areerimboschite mostra un calo, sia pure non molto accen-tuato, delle specie presenti.
foresta pineta di diminuz.naturale rimboschim. %
Specie n°:media 24 20 17min-max 15-35 17-23
Le differenze diventano invece cospicue se vengo-no paragonati i tipi distributivi delle specie presenti neidue ambienti: nella foresta naturale sono prevalenti lespecie eurasiatiche e le sudeuropee, mentre nel rimbo-schimento si ha un forte contingente di specie esotichee subcosmopolite.
Impianti sciistici di risalitaLa conseguenza del calpestamento sul tappeto erbo-
so nei prati di altitudine dell’Appennino Centrale (co-munità a Avenula praetutiana) è stata valutata (PI-GNATTI, 1993) mediante analisi dei pattern su 20 aree di2 m2 di superficie, lungo tre striscie rispettivamentesotto lo skilift (vegetazione calpestata) ed a 10 m didistanza sui due lati. I risultati (tab. VI) mostrano unadiminuzione del numero di specie da 17,7-18,8 (med.18,2) a 9,7, pari ad un calo del 47 %. Contemporanea-mente anche l’indice di Shannon mostra una nettadiminuzione, da 1,77-2,0 a 1,34, e ciò significa checon il calpestio la comunità diviene meno equilibrata eduna o poche specie tendono a prendere il sopravvento.
6. MONITORAGGIO DELLA BIODIVERSITÀA DIMENSIONE TERRITORIALE
I dati sopra riportati, pur nella loro apparente con-traddittorietà, permettono di avvicinarsi ad una con-clusione.
La biodiversità appare il risultato di fenomeni com-plessi che si sovrappongono e condizionano a vicenda.
Il semplice conteggio delle specie presenti su unadeterminata superficie è un dato troppo grezzo peresprimere questa complessità e può avere un significa-to – eventualmente – soltanto nel paragone tra aree consuperficie eguale o poco differente, e comunque incontesti ecologici simili tra loro. Va tenuto presenteche nella dimensione locale o territoriale il totale dellespecie presenti è fortemente determinato dalla compe-tizione e dai fattori del suolo e del microclima che laregolano, mentre nella dimensione geografica il nume-ro delle specie sembra dipendere soprattutto dai fattoriclimatici generali. Dunque, una comparazione tra datisu scala territoriale con altri su scala geografica apparepriva di senso.
I dati qui presentati e discussi permettono di giunge-re ad alcune conclusioni riguardo al problema postoall’inizio, e cioè se l’allarme per il declino della biodi-versità sia o meno giustificato. Considerando il feno-meno alla scala territoriale si notano i fatti seguenti.
1. L’analisi della flora dell’area urbana di Roma dimo-stra chiaramente che le aree più intensamente urbaniz-zate sono quelle nelle quali si registra il più vistoso calonel numero delle specie presenti, e contemporaneamen-te si verifica l’aumento degli indicatori di nutrienti. Si hal’impressione di trovarsi di fronte ad un fenomeno diportata generale, che viene confermato anche dai rilieviriguardanti la pianura friulana e veneta. Va tenuto pre-sente che nei tre casi i rilievi floristici sono stati effettuatisu quadranti di superficie diversa:
Roma (CELESTI GRAPOW, 1995) 1,6 km2
Pianura Veneta (dati originali ined.) 35,0 km2
Pianura Friulana (POLDINI, 1991) 140,0 km2
Si tratta di valori che stanno nella relazione 1 : 25 :100, ma comunque sempre al livello di unità territorialiomogenee.
2. La trasformazione dell’ambiente da uno statosemi-naturale (ma comunque compatibile con l’esi-stenza di una densa popolazione) allo sfruttamentoagricolo intensivo oppure all’urbanizzazione comportala scomparsa di una componente cospicua della flora,valutabile attorno al 50 % delle specie.
3. La flora residua viene profondamente modificata
Tab. VI. Relazione tra calpestio e decadimento della flora su piste sciistiche nell’Appennino centrale
n° specie presenti indice di Shannonnat skilift nat nat skilift nat
Scindarella (1948 m) 15,4 7,3 16,5 1,68 1,01 1,57
Valletta (2079 m) 17,7 9,7 18,8 2,0 1,34 1,77
nat: superfici in condizioni naturali; skilift: striscia centrale calpestata

PIGNATTI S. - Il declino della biodiversità8
nella sua composizione, essenzialmente per l’immis-sione di un forte contingente di specie cosmopolite,per lo più di origine esotica, che possono raggiungere il10-25 % del totale. Dunque la perdita di specie dellaflora indigena è certamente superiore al 50 %, maviene parzialmente mascherata dall’ingresso di specieestranee.
4. Queste modificazioni sono causate non tantodall’intervento diretto delle trasformazioni dell’ambientefisico, che si esplicano a scala locale, ma soprattuttodalla immissione di sostanze estranee, soprattutto nu-trienti, che avviene alla scala dell’intero sistema.
5. Le conseguenze di questa situazione sono scarsa-mente rilevabili alla scala geografica (cioè nel casonostro sui totali relativi al Sistema-Italia) e ciò per leintrinseche caratteristiche di struttura del dato; esseinvece divengono macroscopiche su scala locale. Tut-tavia in ultima analisi il Sistema-Italia risulta dalla som-ma di un gran numero di sistemi locali, dunque ilpeggioramento a scala locale non può non portare adun degrado anche su scala generale.
6. Le cause della decadenza della flora alla scalalocale continuano ad agire a livello generale, soprattut-to l’eutrofizzazione (come accumulo di composti d’azo-to e fosforo nell’ecosistema); dunque si può prevedereun ulteriore aggravarsi della situazione.
I metodi per quantificare la biodiversità rimangonoancora da precisare. Il semplice conteggio delle specieha i limiti sopra indicati. Poco diverso è il giudizio chesi può dare sull’applicazione dell’indice di Shannon.Dunque, una valutazione quantitativa della biodiversitàurta tuttora su gravi ostacoli metodologici.
Sembra chiaro che siamo di fronte ad un problemacentrale per la salvaguardia degli ecosistemi, che per-tanto non può venire trascurato, sia per motivi scienti-fici che applicativi. Sembra necessario pensare allarealizzazione di un sistema esperto in grado di acquisi-
re ed elaborare un gran numero di informazioni tra loroeterogenee, e cioè di tipo biologico, ecologico, climati-co, etc. Questo sistema esperto dovrebbe permetterela gestione di un monitoraggio concentrato sul rileva-mento alla scala territoriale, che meglio permette dievidenziare le modificazioni in atto.
7. SUL CONCETTO DI BIODIVERSITÀLe considerazioni qui sviluppate ci portano a consi-
derare la biodiversità come un processo dinamico, e nonsoltanto come una somma di organismi conviventi. Labiodiversità è il risultato del processo di auto-organizza-zione della materia, che si esplica come evoluzionebiologica e come interazione tra organismi nell’ecosiste-ma. Per quest’ultimo aspetto, la biodiversità va conside-rata un fenomeno dipendente dalla scala (scale depen-dent). Questa può apparire una formulazione più com-prensiva di quanto finora viene generalmente inteso,sebbene il principio dell’auto-organizzazione sia incluso,almeno in maniera implicita, anche nei migliori tentatividi definizione, come ad es. «…the full array of organicdiversity and the origin of that diversity, together withthe methods by which it can be maintained and used forthe benefit of humanity» (WILSON, 1992).
Il carattere di processo fa comprendere la difficoltàdi quantificare la biodiversità attraverso il semplicecomputo delle specie: infatti una specie non vieneeliminata soltanto attraverso la distruzione degli indivi-dui che la compongono, come nel caso della caccia adun vertebrato, ma soprattutto quando gli indispensabilimezzi di sussistenza vengono a mancare. Ciò avviene acausa di fenomeni che si esplicano su scala mondiale,come l’eutrofizzazione, alla quale possono essere ag-giunti ancora: precipitazioni acide, cambio climaticoed, eventualmente, l’aumento della radiazione ultravio-letta a causa del buco di ozono. Le conseguenze sirilevano anzitutto a scala locale, ma agiscono a livelloglobale, e questo fa intendere la gravità del problema.
BIBLIOGRAFIA
CELESTI GRAPOW. L., 1995. Atlante della flora di Roma. Argos ed.,Roma, 222 pp.
CELESTI GRAPOW L., PIGNATTI S., PIGNATTI WIKUS E., 1994. Analisidella flora dei siti archeologici di Roma. Allionia 32: 113-118.
ELLENBERG H., 1974. Zeigerwerte der Gefässpflanzen Mitteleu-ropas. Scripta Geobot. 9. Göttingen, 1974. 2. Aufl. (1979). 3.Aufl. (1992) in Ellenberg H. et al., Scripta Geobot. 18: 9-166.
GOLDSCHMIDT T., 1999. Lo strano caso del Lago Vittoria. Einaudi,Torino, 276 pp. (ed. nederlandese, 1994).
MYERS N. & SIMON J.L., 1995. Scarcity or abundance? W.W.Norton & Co., Padova, 255 pp.
PIGNATTI E., PIGNATTI S., HUANG C.C., DING G.Q., HUANG Z.L.,1991. ß-diversity and phytogeographical patterns in the Ding
Hu Shan Reserve forest vegetation. Rend. Fis. Accad. Lincei s.9, 2: 79-85.
PIGNATTI S., 1982. Flora d’Italia. Edagricole, Bologna, 3 voll.PIGNATTI S., 1988. The relationships between natural vegetation
and social system in the Mediterranean basin, in Miyawaki A.et al. “Vegetation ecology and creation of new environments”:35-45. Tokai University Press, Tokyo (1987).
PIGNATTI S., 1993. Impact of tourism on the mountain landscapeof Central Italy. Landscape and urban planning 24: 49-53.
POLDINI L., 1991. Atlante corologico delle piante vascolari nel Friuli-Venezia Giulia. Reg. Aut. Friuli-Venezia Giulia. Udine, 899 pp.
WILSON E. E., 1992. The diversity of life. Harvard University Press,406 pp.

Biologia Ambientale, 17 (1): 9-16, 2003
Influenza di alcuni fattori ambientali sulla composizione dellecenosi macrobentoniche dei corsi d’acqua planiziali minoriGenoni Pietro
ARPA Lombardia, Dipartimento di Parabiago, Via Spagliardi 19, 20015 Parabiago (MI)
Fax 0331 449703; e-mail: [email protected]
Pervenuto il 31.10.2001; accettato il 16.4.2002
RiassuntoSono stati indagati i principali fattori ambientali in grado di influenzare la distribuzione degli invertebrati bentonici in un gruppo di roggeappartenenti al reticolo idrografico minore della Provincia di Milano e prive di evidenti impatti dovuti a scarichi puntiformi.Nei siti d’indagine sono stati eseguiti, con cadenza bimestrale, campionamenti di macrobenthos e analisi fisiche e chimiche delle acque;inoltre, è stata definita la qualità idromorfologica mediante la procedura dell’Habitat Assessment.L’analisi delle componenti principali (PCA) condotta sui dati biologici ha individuato, quali variabili maggiormente significative, lecaratteristiche delle rive e del substrato, la conducibilità e, in misura minore, le concentrazioni di fosforo e azoto nelle acque. Tali variabiliprevalgono anche su fattori stagionali, quali la variazione di temperatura dell’acqua e sul carico organico.Gli organismi più sensibili rispetto ai gradienti ambientali individuati appartengono ad efemerotteri e tricotteri, gruppi noti per la lorovalidità quali indicatori dell’integrità dei corsi d’acqua. I risultati suggeriscono di non trascurare la presenza di altri taxa, quali molluschi,eterotteri e odonati, che potrebbero fornire ulteriori indicazioni sullo stato ecologico dell’ambiente esaminato.
PAROLE CHIAVE: roggia / macroinvertebrati bentonici / qualità idromorfologica / inquinamento diffuso
AbstractEffects of environmental factors on macroinvertebrate community composition in minor low-gradient streams.The main environmental factors structuring benthic invertebrate communities have been investigated in the province of Milan (Italy) ina group of minor low-gradient irrigation channels without relevant pollution from point sources.Samplings were performed every two months. Water samples were analysed for physical and chemical variables. The macrobenthiccommunity was analysed using the IBE (italian version of the Extended Biotic Index) approach. The idromorphological quality was alsoassessed with the Habitat Assessment protocol.Principal component analysis (PCA) of macrobenthos data identified two main groups of factors, namely, riparian and substratechracteristics and conductivity, total phosphorus and nitrogen in the water. Such variables prevailed even over seasonal factors, asvariation of water temperature, and over organic load.Ephemeroptera and Trichoptera are recognised as the most sensitive amongst the macroinvertebrates. Therefore they can be used asindicators of the aquatic environment integrity. Molluscs, dragonflies and bugs should be also considered.
KEY WORDS: irrigation channel / benthic macroinvertebrates / hydromorphological quality / non-point pollution
INTRODUZIONELa trasformazione del territorio a seguito delle attivi-
tà antropiche può incidere anche profondamente sugliambienti acquatici ad esso connessi. L’agricoltura rap-presenta spesso uno dei fattori di pressione principali,anche se lo sviluppo urbano ed industriale può causaremutamenti più drastici a scala locale (KARR et al.,
1985).L’incremento delle superfici destinate ad uso agri-
colo di solito porta alla riduzione o alla scomparsa dellafascia di vegetazione perifluviale e determina interventisulle caratteristiche idromorfologiche dei corsi d’ac-qua, causando un incremento dell’erosione e del tra-

GENONI P. - Macrobenthos nelle rogge10
sporto di sedimenti e nutrienti. L’alterazione della strut-tura degli ambienti fluviali è considerata una dellemaggiori cause di stress dei sistemi acquatici, essendola diversità biologica strettamente correlata alla con-servazione di tali ecosistemi (KARR et al., 1986; RAVENet al., 1998). L’agricoltura, infine, rappresenta la piùimportante sorgente diffusa di inquinanti per gli am-bienti acquatici (ALLAN, 1995).
Nel presente lavoro è stata studiata l’influenza dialcuni fattori ambientali legati all’utilizzo agricolo delterritorio sulla struttura delle cenosi macrobentonichedi roggia. È stata considerata l’area nord-occidentaledella provincia di Milano caratterizzata, oltre che dallapresenza del fiume Ticino e dei principali canali daesso derivati (Naviglio Grande e Canale Villoresi), dauna fitta rete di corsi d’acqua naturali ed artificiali(fontanili, rogge, cavi, fossi) con funzioni prevalente-mente applicate alle esigenze agricole (PROVINCIA DIMILANO, 2000).
METODI DI INDAGINESono state prese in considerazione 15 stazioni su
altrettante rogge nel territorio nord-occidentale dellaprovincia di Milano (Fig. 1). Volendo indagare preva-lentemente gli effetti delle turbative connesse alle atti-vità agricole, sono stati esclusi i siti posti a valledell’immissione di scarichi puntiformi, quali fognaturee depuratori.
I dati relativi alle cenosi macrobentoniche sono statiraccolti tra il dicembre 1994 ed il dicembre 1996.Campioni qualitativi sono stati raccolti con cadenzabimensile ricostruendo, per ciascun sito, la comunitàmacrobentonica rappresentativa secondo il protocollodi campionamento dell’Indice Biotico Esteso (IBE)descritto in forma aggiornata da GHETTI (1997). Ladeterminazione sistematica è stata effettuata almeno alivello di famiglia (tricotteri, coleotteri, ditteri, crosta-cei, oligocheti) o di genere (plecotteri, efemerotteri,odonati, eterotteri, gasteropodi, bivalvi, tricladi, irudi-nei, megalotteri). Inoltre, a ciascun taxon è stata attri-buita una classe di abbondanza relativa pari a 1 per 1-2individui (taxon raro), a 2 per 3-9 individui (taxonpoco abbondante), a 3 per 10-100 individui (taxoncomune o abbondante) a 4 per più di 100 individui(taxon dominante).
La qualità biologica di ciascuna stazione considerataè stata definita mediante il metodo dell’IBE (GHETTI,1997).
In occasione dei campionamenti di macroinverte-brati sono state eseguite in campo misure di tempera-tura dell’acqua, pH, ossigeno disciolto e conducibilitàa 20°C, mentre in laboratorio si è proceduto alla deter-minazione di COD, azoto totale (organico e inorgani-co) e fosforo totale (IRSA-CNR, 1994). Le analisichimiche sono state condotte presso il Dipartimento diScienze dell’Ambiente e del Territorio dell’Universitàdegli Studi di Milano Bicocca.
La caratterizzazione ambientale dei tratti indagati èstata completata utilizzando i dati raccolti dal Diparti-mento di Biologia Strutturale e Funzionale dell’Univer-sità degli Studi dell’Insubria (CROSA, com. pers.),relativi alla qualità idromorfologica valutata secondo ilmetodo dell’Habitat Assessment (BARBOUR et al.,1999).
Sull’insieme dei dati biologici disponibili (esclusi itaxa rari) è stata condotta l’analisi delle componentiprincipali (PCA), un metodo di ordinamento che per-mette di disporre le osservazioni in uno spazio bidi-mensionale, in modo che i punti tra loro vicini corri-spondano a siti con composizione in specie simili,mentre i punti tra loro lontani corrispondano a siti concomposizione in specie dissimili. L’analisi è stata ese-guita con il programma CANOCO (TER BRAAK, 1989).
Le componenti individuate dalla PCA sono statesuccessivamente interpretate in termini di gradientiambientali, correlando la posizione delle stazioni nelgrafico di ordinamento con i valori dei parametri fisici,chimici ed idromorfologici misurati.
Questo approccio in due fasi può essere definito,nel senso espresso da WHITTAKER (1967), una ”analisidi gradiente indiretta” (JONGMAN et al., 1987; TERBRAAK e PRENTICE, 1988).
0 5 10Km
Fiume Ticino
Naviglio Grande
Naviglio di Bereguardo
Milano
Fig. 1. Inquadramento territoriale del reticolo idrografico oggettodi studio con le 15 stazioni di campionamento (cerchietti).

GENONI P. - Macrobenthos nelle rogge 11
RISULTATILa caratterizzazione fisica e chimica delle acque nei
siti indagati è riassunta in tabella I. I risultati evidenzia-no una discreta omogeneità dei valori delle variabilimisurate e confermano l’assenza di evidenti alterazioniqualitative dovute a scarichi diretti. In particolare, peralmeno tre quarti delle misure effettuate l’ossigenodisciolto si è mantenuto al di sopra dell’80% di satura-zione ed il COD non ha oltrepassato gli 8 mg/L O2. Leconcentrazioni di azoto e fosforo totale mostrano unambito di variazione relativamente contenuto, distri-buendosi rispettivamente attorno a valori medi di 3,9mg/L N (dev. std. 1,5 mg/L N) e di 0,07 mg/L P (dev.std. 0,06 mg/L P).
I 90 campionamenti di macrobenthos eseguiti han-no permesso la determinazione di 90 taxa complessivi,come riassunto in tabella II. A livello di famiglia, lamaggiore ricchezza tassonomica si osserva nel gruppodei tricotteri (13 famiglie), dei ditteri (10 famiglie), deigasteropodi (9 famiglie) e degli odonati (8 famiglie).
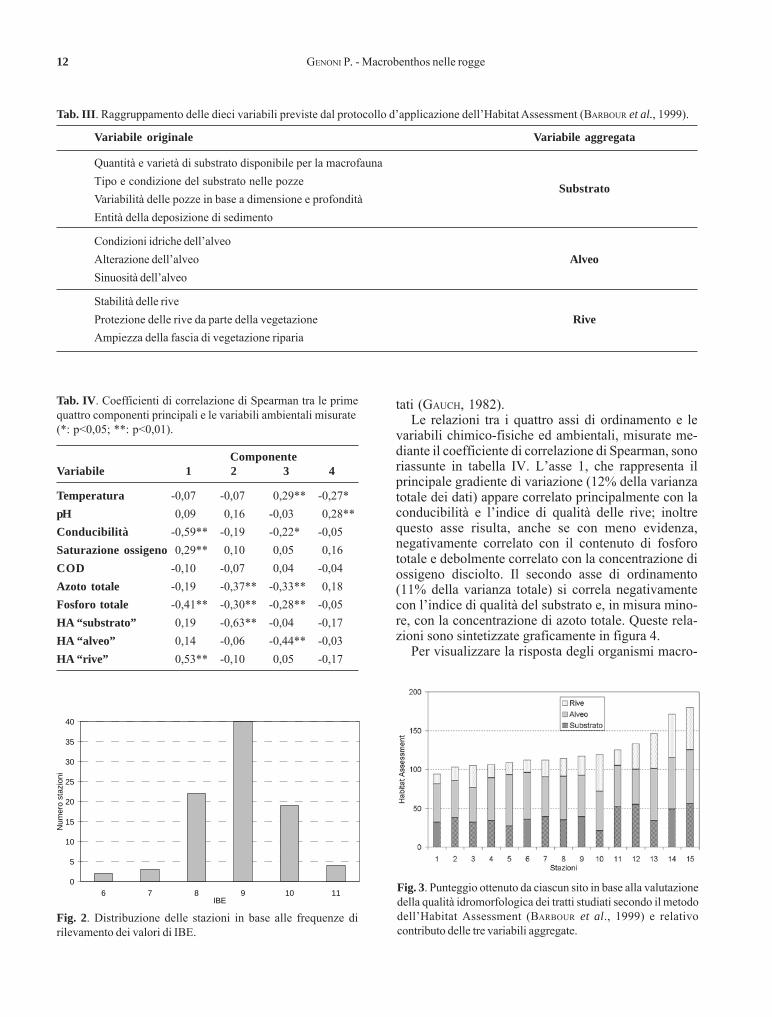
I valori dell’IBE sono compresi tra 6 e 11; quasi il70% dei dati ricade in II classe di qualità, con valoridell’indice pari a 8 e 9, e circa il 20% rientra in I classe,presentando valori di IBE pari a 10 e 11 (Fig. 2).
Per la successiva fase di elaborazione dei dati, ledieci variabili previste dal protocollo di applicazionedell’Habitat Assessment sono state aggregate in tregruppi, rinominati “Substrato”, “Alveo” e “Rive” (Tab.III). La condizione idromorfologica dei tratti selezio-nati ed il contributo di ciascuno dei tre gruppi divariabili aggregate sono riportate in figura 3. La mag-gior parte delle stazioni presenta un punteggio compre-so tra 100 e 150, che denota condizioni ambientali
Tab. I. Statistica riassuntiva delle variabili chimico-fisiche misurate.
Tem
pera
tura
°C pH Con
duci
bilit
àμS
cm-1
Oss
igen
o di
scio
ltom
g O
2 l-1
Satu
razi
one
ossi
geno
% CO
Dm
g O
2 l-1
Azo
to t
otal
em
g N
l-1
Fosf
oro
tota
lem
g P
l-1
Numero osservazioni 90 90 87 90 90 82 88 84Minimo 6,9 7,1 137 6,4 65 0,2 0,2 0,01Massimo 24,4 8,9 513 13,2 131 18,4 8,3 0,42Media 14,2 7,7 338 9,3 91 6,2 3,9 0,07Deviazione standard 3,5 0,3 90 1,4 12 3,3 1,5 0,06Coefficiente di variazione (%) 24,7 4,5 26,8 14,6 13,0 53,7 38,4 85,725° percentile 11,2 7,5 273 8,4 83 4,2 3,0 0,04Mediana 14,7 7,7 337 9,1 90 5,8 3,8 0,0675° percentile 16,2 7,9 407 10,1 99 7,9 4,9 0,09
Tab. II. Numero di taxa identificati a livello di famiglia e di generenei 90 campionamenti eseguiti.(n.d.: taxa non determinati a livello di genere).
Gruppo Numero Numerodi famiglie di generi
Turbellaria 3 3Oligochaeta 5 n.d.Hirudinea 3 8Gastropoda 9 10Bivalvia 4 4Amphipoda 1 n.d.Isopoda 1 n.d.Ephemeroptera 6 9Odonata 8 13Plecoptera 3 3Hemiptera 3 3Megaloptera 1 1Coleoptera 6 n.d.Trichoptera 13 n.d.Diptera 10 n.d.
TOTALE 76 54
definite da “marginali” a “subottimali”.Le prime quattro componenti principali spiegano
insieme il 36% della varianza totale dei dati biologici.Valori relativamente bassi (fino al 5%) della frazione divarianza spiegata sono piuttosto comuni nella ricercaecologica quando si trattano dati numerosi e comples-si; ciononostante essi non inficiano la qualità dei risul-
GENONI P. - Macrobenthos nelle rogge12
tati (GAUCH, 1982).Le relazioni tra i quattro assi di ordinamento e le
variabili chimico-fisiche ed ambientali, misurate me-diante il coefficiente di correlazione di Spearman, sonoriassunte in tabella IV. L’asse 1, che rappresenta ilprincipale gradiente di variazione (12% della varianzatotale dei dati) appare correlato principalmente con laconducibilità e l’indice di qualità delle rive; inoltrequesto asse risulta, anche se con meno evidenza,negativamente correlato con il contenuto di fosforototale e debolmente correlato con la concentrazione diossigeno disciolto. Il secondo asse di ordinamento(11% della varianza totale) si correla negativamentecon l’indice di qualità del substrato e, in misura mino-re, con la concentrazione di azoto totale. Queste rela-zioni sono sintetizzate graficamente in figura 4.
Per visualizzare la risposta degli organismi macro-
Tab. III. Raggruppamento delle dieci variabili previste dal protocollo d’applicazione dell’Habitat Assessment (BARBOUR et al., 1999).
Variabile originale Variabile aggregata
Quantità e varietà di substrato disponibile per la macrofaunaTipo e condizione del substrato nelle pozzeVariabilità delle pozze in base a dimensione e profonditàEntità della deposizione di sedimento
Condizioni idriche dell’alveoAlterazione dell’alveo AlveoSinuosità dell’alveo
Stabilità delle riveProtezione delle rive da parte della vegetazione RiveAmpiezza della fascia di vegetazione riparia
Substrato
Fig. 2. Distribuzione delle stazioni in base alle frequenze dirilevamento dei valori di IBE.
0
5
10
15
20
25
30
35
40
6 7 8 9 10 11IBE
Num
ero
staz
ioni
Tab. IV. Coefficienti di correlazione di Spearman tra le primequattro componenti principali e le variabili ambientali misurate(*: p<0,05; **: p<0,01).
ComponenteVariabile 1 2 3 4
Temperatura -0,07 -0,07 0,29** -0,27*pH 0,09 0,16 -0,03 0,28**Conducibilità -0,59** -0,19 -0,22* -0,05Saturazione ossigeno 0,29** 0,10 0,05 0,16COD -0,10 -0,07 0,04 -0,04Azoto totale -0,19 -0,37** -0,33** 0,18Fosforo totale -0,41** -0,30** -0,28** -0,05HA “substrato” 0,19 -0,63** -0,04 -0,17HA “alveo” 0,14 -0,06 -0,44** -0,03HA “rive” 0,53** -0,10 0,05 -0,17
0
50
100
150
200
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15Stazioni
Hab
itat A
sses
smen
t
RiveAlveoSubstrato
Fig. 3. Punteggio ottenuto da ciascun sito in base alla valutazionedella qualità idromorfologica dei tratti studiati secondo il metododell’Habitat Assessment (BARBOUR et al., 1999) e relativocontributo delle tre variabili aggregate.

GENONI P. - Macrobenthos nelle rogge 13
bentonici ai principali gradienti ambientali individuati,l’abbondanza di ciascun taxon è stata rappresentatagraficamente in corrispondenza delle stazioni ordinatesecondo i primi due assi della PCA. La ricerca dei taxameglio rappresentati dall’ordinamento ottenuto è stataeffettuata osservando il corrispondente valore di adat-tamento cumulativo (cumulative fit) rispetto ai primidue assi estratti. Questo parametro può essere inter-pretato come la frazione cumulativa della varianza deltaxon spiegata da tali assi (TER BRAAK, 1990).
In questa fase sono stati trascurati i taxa più comu-ni, che, anche se con valori di densità variabili, sonostati quasi sempre rinvenuti in tutte le stazioni (comeGammaridae, Asellidae, Chironomidae, alcuni tricladi eirudinei) e quindi sono meno intreressanti come orga-nismi indicatori.
In primo luogo, i risultati più significativi sono statiottenuti per tre taxa: Ecdyonurus sp. (efemerotteri),Aphelocheirus aestivalis (eterotteri) e Sphaerium sp.(bivalvi), i quali presentano preferenze ambientali traloro complementari, facilmente interpretabili medianteil confronto con le variabili rappresentate graficamentein figura 4. Il genere Ecdyonurus predilige stazioni conun ambiente ripario in buone condizioni ed una elevataeterogeneità nella composizione del substrato; inoltre,esso mostra una certa tolleranza nei confronti dellapresenza di azoto nelle acque (Fig. 5). Al contrario, A.aestivalis si rinviene nei siti con substrato più fine eduniforme, ma non compare dove vi è un elevato conte-nuto di nutrienti (Fig. 6). Infine, Sphaerium sp. èrisultato abbondante soprattutto nelle stazioni che pre-sentano elevati valori di conducibilità, condizioni alte-rate dell’ambiente ripario e dove la deposizione delsedimento crea un substrato omogeneo e le acquepresentano in genere concentrazioni di fosforo piùelevate (Fig. 7).
Inoltre, i risultati evidenziano che la distribuzione dialtri organismi, soprattutto gli odonati del genere Ca-lopteryx, gli efemerotteri dei generi Paraleptophlebia eRhithrogena ed i tricotteri della famiglia Lepidostoma-tidae, è in relazione principalmente con il primo asse diordinamento. Le famiglie Goeridae e Glossosomatidae(tricotteri) e Ancylus fluviatilis (gasteropodi), risento-no invece del gradiente ambientale rappresentato dalsecondo asse dell’analisi.
La disposizione complessiva dei 64 taxa utilizzatiper la PCA è riassunta in figura 8; in questo tipo dirappresentazione gli organismi distanti dall’origine de-gli assi sono i più significativi nell’indicare le differenzetra le stazioni, mentre i punti al centro del grafico (peri quali non viene riportato il nome del taxa corrispon-dente) sono di minore importanza (JONGMAN et al.,1987; TER BRAAK e PRENTICE, 1988).
Fig. 4. Relazioni tra le prime due componenti principali e levariabili ambientali misurate. La lunghezza dei vettori e la vicinanzaagli assi ne definiscono l’importanza rispetto ai gradienti estrattidall’analisi (T°C: temperatura dell’acqua; O2 %: percentuale disaturazione di ossigeno; Cond.: conducibilità).
Fig. 5. Distribuzione delle abbondanze relative del genereEcdyonurus nelle stazioni rappresentate secondo i primi due assidi ordinamento. La dimensione del simbolo è proporzionale allaclasse di abbondanza del taxon; il punto indica l’assenza deltaxon nella stazione corrispondente.
Fig. 6. Distribuzione delle abbondanze relative di Aphelocheirusaestivalis nelle stazioni rappresentate secondo i primi due assi diordinamento. Simboli come in figura 5.
Ecdyonurus sp.
-1,0
0,0
1,0
-1,0 0,0 1,0asse 1
asse 2
Aphelocheirus aestivalis
-1,0
0,0
1,0
-1,0 0,0 1,0asse 1
asse 2T°C
O2 %COD
Azoto
Fosforo
Substrato
Alveo
Rive
pH
Cond.
-0,2
-0,1
0
0,1
0,2
-0,2 -0,1 0 0,1 0,2asse 1
asse
2

GENONI P. - Macrobenthos nelle rogge14
Fig. 7. Distribuzione delle abbondanze relative di Sphaerium sp.nelle stazioni rappresentate secondo i primi due assi diordinamento. Simboli come in figura 5.
Fig. 8. Ordinamento dei 64 taxa secondo le prime due componentiprincipali; i taxa più significativi sono indicati con il simbolopieno ( ), quelli molto comuni con il simbolo vuoto ( ), mentrei punti al centro del grafico indicano i taxa di minore importanza(vedi testo).
DISCUSSIONELa distribuzione delle comunità di invertebrati negli
ecosistemi acquatici dipende da fattori ambientali dif-ferenti in base alla diversa scala spaziale esaminata(regionale, di bacino, di corso d’acqua) e all’interno diun’ecoregione le biocenosi sono condizionate preva-lentemente dagli effetti dell’uso del territorio (TATE eHEINY, 1995). Secondo uno studio condotto da RI-CHARDS e HOST (1993) in un bacino idrografico degliStati Uniti a prevalente attività agricola, le caratteristi-che fisiche dei corsi d’acqua, in particolare substrato eambiente ripario, sono risultate le principali variabiliche influenzano la struttura delle comunità macroben-toniche.
I risultati ottenuti nel presente lavoro sembranosostenere l’osservazione che, in assenza di cause d’im-
patto puntiformi quali gli scarichi diretti, l’alterazionedelle caratteristiche degli ambienti ripari e del substratoin alveo siano tra i principali fattori ambientali cheinfluenzano la composizione delle cenosi macrobento-niche dei corsi d’acqua minori che scorrono in conte-sti agricoli.
Oltre a ciò, particolarmente importante è risultato ilparametro conducibilità, una misura indiretta del con-tenuto di ioni disciolti nelle acque. Le differenze tra ipunti di campionamento, relativamente a questo para-metro, sono spiegate solo parzialmente dalla diversaorigine – da risorgive e fontanili piuttosto che dal fiumeTicino – delle acque presenti nelle rogge considerate.Si può ipotizzare che, nel contesto in esame, i valori diconducibilità più elevati siano in relazione anche all’usodi fertilizzanti per l’agricoltura intensiva, i quali rappre-sentano una sorgente di arricchimento antropico perdiversi ioni, come il sodio, i cloruri ed i solfati (ALLAN,1995).
Anche altri effetti conseguenti all’utilizzo agricolodel territorio, quali l’incremento di nutrienti (fosforo eazoto) appaiono significativamente legati, anche secon un ruolo di minore importanza, ai principali gra-dienti di variazione della comunità macrobentonica.
Al contrario, fattori non dipendenti dalle attivitàantropiche, nello specifico la temperatura dell’acqua,che contribuisce a condizionare i cicli biologici degliorganismi, assumono un ruolo di secondo piano. I datiraccolti evidenziano che anche il contenuto di sostanzechimicamente ossidabili appare ininfluente.
I risultati ottenuti sembrano confermare, anche perla tipologia studiata, l’elevata sensibilità alle diversecause di alterazione da parte di efemerotteri e tricotteri,gruppi comunemente considerati validi indicatori dellaqualità delle acque e delle condizioni ambientali (USSE-GLIO-POLATERA, 1989; BUFFAGNI, 1997). Tra l’altro,negli ambienti esaminati la ricchezza tassonomica diquesti gruppi è risultata piuttosto elevata (Tab. II). Laprima componente principale presenta una relazionemigliore con il numero di taxa di efemerotteri, plecot-teri e tricotteri (indice EPT; BARBOUR et al., 1999),piuttosto che con l’IBE (rispettivamente, r di Spear-man pari a 0,73 e 0,63; p < 0,01), nonostante il primoindice sia stato calcolato usando un livello tassonomi-co poco approfondito, in particolare per i tricotteri, edil gruppo dei plecotteri sia quasi assente in questatipologia (solo tre generi identificati occasionalmente).
Anche le frequenze di altri organismi bentonici ap-paiono in buona relazione con i principali gradientiambientali individuati nell’indagine; è interessante no-tare che alcuni di essi (nello specifico: Ancylidae,Sphaeridae e Aphelocheirus) sono inclusi tra i taxaindicatori nella procedura di calcolo del Belgian BioticIndex (DE PAUW e VANHOOREN, 1983). Si tratta di
Sphaerium sp.
-1,0
0,0
1,0
-1,0 0,0 1,0asse 1
asse 2
BaetisGammaridae
Elmidae
Asellidae
Erpobdella
Chironomidae
Ephemerella
Dugesia
Dendrocoelum
Glossiphonia
Ancylus
Ecdyonurus
Calopteryx
Goeridae
Lumbriculidae
Ephemera
LepidostomatidaeHelobdella
CaenisSphaerium
Aphelocheirus
Tubificidae
PolycelisOdontoceridae
LimnephilidaeLeptoceridae
Paraleptophlebia
Glossosomatidae
Platycnemis
Leuctra
Rhithrogena
-2,0
0,0
2,0
-2,0 0,0 2,0
asse 1
asse 2

GENONI P. - Macrobenthos nelle rogge 15
organismi che vivono di solito in acque ben ossigenatee sufficientemente pulite (CASTAGNOLO et al., 1980;CAMPAIOLI et al., 1994), come quelle degli ambienti inesame, ma la cui distribuzione evidentemente risente dicaratteristiche ecologiche differenti.
I risultati indicano l’utilità dell’approfondimento tas-sonomico anche di gruppi generalmente consideraticattivi indicatori della qualità delle acque, come gliodonati. Ad esempio, al genere Calopteryx, frequente-mente rinvenuto nel corso dello studio e significativa-mente correlato al primo asse di ordinamento, appar-tengono specie che, pur convivendo spesso nei mede-simi ambienti, presentano diverse esigenze dal punto divista idroqualitativo (CARCHINI, 1983); la loro distinzio-ne potrebbe pertanto migliorare la conoscenza sullostato di conservazione dei siti in esame.
CONCLUSIONINegli ultimi anni l’attenzione della ricerca nell’ambi-
to dell’ecologia fluviale si è spostata verso lo studiodegli elementi di qualità idromorfologica dei corsi d’ac-qua e l’analisi delle loro connessioni con gli elementi diqualità biologica. I risultati esposti confermano l’im-portanza della conservazione dell’ambiente fisico an-che per quei corsi d’acqua planiziali – spesso semina-turali – considerati minori, ma che mantengono tutta-via un elevato valore biologico e naturalistico. La sem-
plificazione della struttura idromorfologica di tali am-bienti si traduce di solito in una riduzione della diversitàdegli organismi più sensibili presenti all’interno dellebiocenosi bentoniche. A questi interventi diretti si ag-giunge l’effetto dei carichi diffusi generati dall’agricol-tura tradizionale, i quali assumono, nei confronti deimacroinvertebrati bentonici, un’importanza maggiorerispetto ai fattori stagionali, quali la variazione di tem-peratura dell’acqua, ed agli indicatori di carico organi-co.
Come emerso nel presente lavoro, gli indici bioticitradizionalmente usati, come l’IBE, a volte non sono ingrado di evidenziare gli effetti precoci di tali alterazionie pertanto si rende necessario il ricorso ad indicatori dimaggior dettaglio.
In ultima analisi, l’indagine suggerisce l’opportunitàdi adottare pratiche agricole compatibili con le esigen-ze di conservazione di quegli ecosistemi acquatici soloapparentemente marginali.
RingraziamentiRingrazio la dr.ssa Letizia Garibaldi (Università degli Studi diMilano Bicocca) per il contributo alla raccolta dei dati chimici edil prof. Giuseppe Crosa (Università dell’Insubria) per i datiidromorfologici, per l’utilizzo del programma di analisi multiva-riata e per alcuni suggerimenti critici. Ringrazio inoltre un reviso-re anonimo per le utili osservazioni sul testo.
BIBLIOGRAFIA
ALLAN J.D., 1995. Stream ecology. Structure and function ofrunning waters. Chapman & Hall, London, 388 pp.
BARBOUR M.T., GERRISTEN J., SNYDER B.D., STRIBLING J.B., 1999.Rapid Bioassessment Protocols for use in streams and wa-deable rivers: periphyton, benthic macroinvertebrates andfish. Second edition. EPA 841-B-99-002. U.S. EnvironmentalProtection Agency; Office of Water; Washington , D.C.
BUFFAGNI A., 1997. Mayfly community composition and thebiological quality of streams. In: Landolt P. & Sartori M.(eds.), Ephemeroptera & Plecoptera: Biology – Ecology –Systematics. MTL, Fribourg: 235-246.
CAMPAIOLI S., GHETTI P.F., MINELLI A., RUFFO S., 1994. Manualeper il riconoscimento dei macroinvertebrati delle acque dolciitaliane. Volume I, Provincia Autonoma di Trento, 357 pp.
CARCHINI G., 1983. Odonati (Odonata). Consiglio Nazionaledelle Ricerche. Collana del progetto finalizzato “Promozionedella qualità dell’ambiente”, AQ/1/198, 80 pp.
CASTAGNOLO L., FRANCHINI D., GIUSTI F., 1980. Bivalvi (Bival-
via). Consiglio Nazionale delle Ricerche. Collana del progettofinalizzato “Promozione della qualità dell’ambiente”, AQ/1/49, 64 pp.
DE PAUW N., VANHOOREN G., 1983. Method for biologicalassessment of watercourses in Belgium. Hydrobiologia, 100:153-168.
GAUCH H.G., 1982. Multivariate analysis in community ecology.Cambridge University Press, Cambridge, 298 pp.
GHETTI P.F., 1997. Manuale di applicazione. Indice BioticoEsteso (IBE). I macroinvertebrati nell’analisi di qualità deicorsi d’acqua. Provincia Autonoma di Trento, Trento, 222pp.
IRSA-CNR, 1994. Metodi analitici per le acque. Quaderni, 100.CNR – Istituto di Ricerca Sulle Acque, Roma, 342 pp.
JONGMAN R.H.G, TER BRAAK C.J.F. and VAN TONGEREN O.F.R.,1987. Data analysis in community and landscape ecology.Centre for Agricultural Publishing and Documentation, Wa-geningen, 299 pp.

GENONI P. - Macrobenthos nelle rogge16
KARR J.R., FAUSCH K.D., ANGERMEIER P.L., YANT P.R. andSCHLOSSER I.J., 1986. Assessing biological integrity in runningwaters: a method and its rationale. Special publication 5,Illinois Natural History Survey.
KARR J.R., TOTH L.A. and DUDLEY D.R., 1985. Fish communitiesof midwestern rivers: a history of degradation. Bioscience, 35:90-95.
PROVINCIA DI MILANO, 2000. Carta della vocazioni ittiche. Carat-terizzazione ambientale degli ecosistemi acquatici. Provinciadi Milano, Tutela e Sviluppo Ambientale, 340 pp.
RAVEN P.J., HOLMES N.T.H., DAWSON F.H., FOX P.J.A., EVERARD
M., FOZZARD I.R. and ROWEN K.J., 1998. River HabitatQuality: the physical character of rivers and streams in theUK and Isle of Man. Environment Agency, Bristol, England.
RICHARDS C., HOST G.E., 1993. Identification of predominantenvironmental factors structuring stream macroinvertebratecommunities within a large agricultural catchment. Freshwa-ter Biology, 29: 285-294.
TATE C.M. and HEINY J.S., 1995. The ordination of benthicinvertebrate communities in the South Platte River Basin inrelation to environmental factors. Freshwater Biology, 33:439-454.
TER BRAAK C.J.F., 1989. CANOCO – an extension of DECORA-NA to analyze species-environment relationships. Hydro-biologia, 184: 169-170.
TER BRAAK C.J.F., 1990. Update notes: CANOCO version 3.10.Agricultural Mathematics Group, Wageningen, 35 pp.
TER BRAAK C.J.F. and Prentice I.C., 1988. A theory of gradientanalysis. Advances in Ecological Research, 18: 271-317.
USSEGLIO-POLATERA P. and BOURNARD M., 1989. Trichoptera andEphemeroptera as indicators of environmental changes of theRhone River at Lyons over the last twenty-five years.Regulated Rivers, 4: 249-262.
WHITTAKER R.H., 1967. Gradient analysis of vegetation. Biol.Rev., 49: 207-264.

Biologia Ambientale, 17 (1): 17-23, 2003
Giuseppe Sansoni1*, Bruno Borghini1, Gino Camici1,Monica Casotti1, Paolo Righini2, Chiara Rustighi3
1 ARPAT, Dipartimento Provinciale di Massa Carrara, Via del Patriota, 2 – 54100 Massa.2 ARPAT, Dipartimento Provinciale di Livorno, Via Marradi, 114 – 57126 Livorno3 Centro Sub Alto Tirreno, Massa (MS)* Autore referente per la corrispondenza (Fax: 0585 47000; e-mail: [email protected])
Pervenuto il 5.11.2001; accettato il 16.4.2002
RiassuntoVengono presentati i risultati delle osservazioni su fioriture algali di Ostreopsis ovata, accompagnate da estese morie di organismiappartenenti a diverse cenosi bentoniche animali e vegetali, verificatesi nelle stagioni estive degli anni 1998, 2000 e 2001 nel mare costieroapuano (Toscana nord-occidentale). L’intensità massima del fenomeno si è manifestata in un tratto di litorale nel quale opere di difesadall’erosione circoscrivono uno specchio marino a debole ricambio idrico, soggetto quindi a maggior riscaldamento delle acque. Inconcomitanza con la fioritura algale, numerose persone che avevano soggiornato in tali zone e verosimilmente inalato aerosol marino hannoaccusato malesseri di diversa intensità alle prime vie respiratorie.
PAROLE CHIAVE: fioriture algali; tossine algali; Ostreopsis ovata; morie d’invertebrati marini.
AbstractAlgal blooms of Ostreopsis ovata (Gonyaulacales: Dinophyceae): an emerging problemThe observations of summer blooms of Ostreopsis ovata in the Apuan benthic seawaters (Tuscany, Italy) from 1998 to 2001 arepresented. Heavy consequences for benthic communities.This phenomenon has taken place in a sheltered marine area which has a slow water circulation and heating of the waters. The inhalationof marine aerosols has also caused human malaises.
KEY WORDS: algal blooms; algal toxins; Ostreopsis ovata; marine invertebrate mortality.
INTRODUZIONELe fioriture algali marine, soprattutto quelle attribui-
bili ai dinoflagellati, sono ritenute responsabili dellaproduzione di tossine che possono causare morie diorganismi marini (ANDERSON, 1989, VOLTERRA, 1989).
Alcuni dinoflagellati epibentici, principalmente Gam-bierdiscus toxicus, ma anche diverse specie di Ostreopsis(O. siamensis, O. ovata, O. lenticularis), o altre ap-partenenti ai generi Prorocentrum e Coolia, produco-no tossine del gruppo della ciguatera. Anche l’uomo
può ingerire le tossine consumando pesci o altri orga-nismi marini che le hanno preventivamente bioaccu-mulate mediante la catena alimentare (ANDERSON eLOBEL, 1987; HALLEGRAEFF, 1993; BOMBER et al., 1989;BRUSLÉ, 1997; GLAZIOU e LEGRAND, 1994; RUSSEL eEGEN, 1991).
Ostreopsis ovata, la peridinea più piccola del genere(dimensioni 47-55μm x 27-35μm) (FUKUYO, 1981),sebbene produca tossine del gruppo della ciguatera è
Fioriture algali di Ostreopsis ovata (Gonyaulacales:Dinophyceae): un problema emergente

SANSONI et al. - Ostreopsis ovata: un problema emergente18
ritenuta tossica solo per gli animali marini; in letteratu-ra non sono segnalati casi d’intossicazione alimentareumana attribuibili chiaramente ad essa. Scopo del pre-sente lavoro è segnalare le conseguenze sulle biocenosibentoniche e casi di malesseri umani concomitanti conevidenti fioriture di O. ovata, verificatisi presso illitorale apuano negli anni 1998, 2000 e 2001.
MATERIALI E METODIL’area indagata, compresa nel tratto di mare più
prossimo alla riva, è larga circa 100 m e si estende percirca 13 km sul litorale della provincia di Massa-Carrara (Fig. 1); informazioni addizionali su eventianaloghi sono state raccolte anche su altri litorali italia-ni.
Il tratto centrale del litorale apuano, a levante delporto di Marina di Carrara, è soggetto a fenomenierosivi ed è stato perciò difeso da scogliere parallele eperpendicolari alla riva. Questi interventi hanno provo-cato una compartimentazione in specchi d’acqua, conun conseguente debole ricambio idrico che sembragiocare un ruolo rilevante per la comparsa e l’intensitàdella fioritura algale nel periodo tardo-estivo.
Lo spunto per l’avvio dell’indagine che ha condottoall’accertamento della fioritura di O. ovata è statotratto dalla segnalazione, pervenuta all’inizio dell’ago-sto 1998, di malesseri sofferti da diverse decine dituristi mentre soggiornavano in un breve tratto dispiaggia. Accertata preventivamente l’assenza di trat-tamenti fitosanitari e di altre attività che avessero potu-to influire sul fenomeno segnalato, l’indagine si èconcentrata sulla qualità delle acque marine in questiparticolari tratti di mare (che in pratica sono racchiusitra barriere perpendicolari e parallele alla costa, costi-tuite da scogliere in massi ciclopici), privilegiandol’osservazione microscopica di campioni d’acqua e,soprattutto, eseguendo, mediante immersione in ap-nea, un’accurata ispezione visiva delle acque e dellebiocenosi bentoniche.
Durante tali sopralluoghi è stata constatata la pre-senza di una pellicola gelatinosa perifitica brunastrache ricopriva monotonamente i substrati sommersi;tale pellicola è stata sottoposta ad esame microscopi-co. Per gran parte degli aspetti indagati (schiumesuperficiali, opalescenza delle acque, segni d’anaero-biosi sui fondali, stato di sofferenza delle cenosi bento-niche, materiali di consistenza gelatinosa in sospensio-ne) i risultati sono stati espressi in una scala arbitrariad’intensità del fenomeno, da assente ad elevata.
Nel 1998, considerata l’intensa opalescenza bianca-stra delle acque, si è proceduto, dopo blanda centrifu-gazione dei campioni raccolti (10 minuti a 500 rpm), alconteggio in camera di Burker degli aggregati di parti-celle sospese con dimensioni comprese tra 5 e 50 μm.
Anche l’osservazione delle alghe nelle acque è statacondotta in microscopia a luce trasmessa e in contra-sto di fase, a 400 x, su campioni sottoposti a blandacentrifugazione.
RISULTATI E DISCUSSIONEIn quasi tutte le aree indagate, le acque, viste dalla
superficie, si presentavano da leggermente opalescentia torbide, riflettendo la luce in modo tale da produrreun “effetto nebbia” che riduceva la trasparenza fino asoli circa 15 cm. Sporadicamente erano presenti anche“fiocchi” di materiale sospeso di ridotte dimensioni(alcuni cm) che, in controluce, mostravano una legge-rissima trama ialina con una punteggiatura finissimarossastra (dovuta a milioni di cellule di O. ovata, comerivelato dall’esame microscopico).
In alcuni campioni d’acqua si è proceduto al con-teggio in camera di Burker degli aggregati sospesi didimensioni da 5 a 50 μm, riscontrandone concentra-zioni tali da giustificare la torbidità rilevabile ad occhionudo (fino a 8 milioni di aggregati/litro). Gli aggregatierano costituiti prevalentemente da particelle minori di1 μm, tenute assieme da materiale gelatinoso traspa-rente.
Sulla superficie degli specchi marini protetti dallescogliere erano inoltre riscontrabili chiazze galleggiantibiancastre, d’aspetto gelatinoso, costituite da migliaiad’esoscheletri di cirripedi balanidi a conferma di unamoria che aveva colpito tale popolazione. In altri casisono stati riscontrati, in sospensione, aggregati sparsidi materiali molli, grumosi, sfrangiati, di color brunocostituiti da frammenti della pellicola perifitica distac-catasi dagli scogli (Fig. 2).
È opportuno tra l’altro sottolineare che nel tratto dimare in cui la fioritura si era manifestata con la massi-ma intensità, la temperatura media delle acque dagiugno ad agosto 1998 raggiunse un massimo di circa25°C (fig. 3).
Nel 1998 l’esame microscopico dei campioni d’ac-qua di mare aveva rilevato costantemente la presenzadi O. ovata in concentrazioni variabili da circa 3.000 acirca 50.000 cellule/litro (Fig. 4); non sono state ri-scontrate altre specie di alghe tossiche. Sebbene taliconcentrazioni non fossero molto elevate, la diffusionedella peridinea in un tratto di litorale molto ampio eraindice di una intensa fioritura algale. Tale risultato èstato confermato dall’esame microscopico della pelli-cola gelatinosa brunastra che rivestiva il substrato,nella quale sono state riscontrate concentrazioni eleva-tissime di O. ovata, variabili da 250 milioni a 3 miliardidi cellule per litro di materiale.
Nel corso degli anni 2000 e 2001 l’indagine sulleacque si è limitata ad escludere la presenza di altremicroalghe tossiche mediante analisi microscopica.

SANSONI et al. - Ostreopsis ovata: un problema emergente 19
Fig. 1. Quadro d’insieme dei risultati dell’indagine ambientale. L’area d’indagine (nell’ovale tratteggiato) è la fascia d’acque marineprossima alla riva, lungo tutto il litorale della provincia di Massa-Carrara. Ogni aspetto è sinteticamente espresso in una scala arbitrariad’intensità. La freccia grigia indica lo specchio marino ove si è verificata la massima intensità della fioritura algale.
Parmignola Fossa MaestraCarrione
PortoM. di Carrara
CONFINE SP-MS
Aree indagateTorbiditàAnaerobiosi fondoEsoscheletri balaniFiocchi gelat. sosp.Sofferenza scogliere
123456789012345678901234567891234567890123456789012345678912345678901234567890123456
12345678901234567890123456
1234567890123456789012312345678901234567890123
123123
12345678901234567812345678901234567812
121212123456789012345678901234567
123456789012345678901234567
LavelloRicortola
Aree indagateTorbiditàAnaerobiosi fondoEsoscheletri balaniFiocchi gelatinosi sospesiSofferenza scogliere
Partaccia TorreFIAT
12345678901234567890
1234567890112345678901
12345671234567
12345612345612
12
123456789011234567890112345678901
Brugiano FRIGIDO
○ ○
Pontile
Ronchi
Aree indagateTorbiditàAnaerobiosi fondoEsoscheletri balaniFiocchi gelatin. sosp.Sofferenza scogliere
scogliere soffolte
12341234
12341234
12341234
12341234 123456789012
123456789012123456789012123456789012123456789012123456789012123456789012123456789012123456789012
123456123456123123123123123123
F. Poveromo VERSILIA CONFINE MS-LU
Aree indagateTorbiditàAnaerobiosi fondoEsoscheletri balaniFiocchi gelatin. sosp.Sofferenza scogliere
PoveromoCinquale
123456789012345678901234567890121234567890123456123456789012345678901234567890121234567890123456123456789012345678901234567890121234567890123456123456789012345678901234567890121234567890123456123456789012345678901234567890121234567890123456123456789012345678901234567890121234567890123456123456789012345678901234567890121234567890123456123456789012345678901234567890121234567890123456
0 200 400 600 800 1000 m
assente o quasi lieve medio
LEGENDA:12341234
12341234 consistente elevato
M S
LU
PI
LI
PT
ARFI
GR
SI
PO

SANSONI et al. - Ostreopsis ovata: un problema emergente20
Fig. 2. I frammenti flottanti della pellicola di O. ovata, distaccatisidal substrato, sono talora confusi con escrementi dispersi, a causadel colore e dell’aspetto (4 x).
Porto
T. P
arm
igno
la
Foss
a M
aest
ra
Marina diCarrara
T. C
arrio
ne
Foss
o L
avel
lo
T. R
icor
tola
Foss
o B
rugi
ano
F. F
rigid
o
Marinadi Massa
Foss
o M
aglia
no
Foss
o P
over
omo
F. V
ersi
lia
1 km
23,8
24,0
24,2
24,4
24,6
24,8
25,0
°C
Fig. 3. Andamento della temperatura delle acque (media dei mesi giugno-agosto 1998; misure bimensili) lungo il litorale apuano. Lamassima intensità della fioritura algale e della compromissione delle biocenosi bentoniche si è manifestata nello specchio acqueo con latemperatura più elevata. Le frecce indicano la localizzazione dei punti di misura della temperatura.
Fig. 4. Individui d’Ostreopsis ovata (50 x 30 μm).
L’opalescenza delle acque è apparsa invece modestanel 2000 e ancora molto contenuta nel 2001, mentreha continuato a riscontrarsi la presenza di esoscheletridi balanidi, sia pure in quantità modeste.
L’osservazione delle più comuni biocenosi, eseguitanell’estate del 1998 nel tratto di mare interessato dalfenomeno della fioritura, ha evidenziato una situazionedecisamente alterata riguardo a presenza (o assenza) econsistenza delle varie specie rinvenibili comunementenei diversi piani.
Nel Piano Sopralitorale, non si notavano alterazionidelle popolazioni di gasteropodi (Littorina neritoides) edi cirripedi (Chthamalus stellatus), come del resto nei
numerosi esemplari osservati del granchio corridore(Pachygrapsus marmoratus), evidentemente non sog-getti agli effetti diretti o indiretti della intensa fiorituradella peridinea. Sui substrati duri del Piano Mesolitora-le si osservava un gran numero d’impronte di patella(Patella caerulea) le cui conchiglie, ormai vuote, sirinvenivano in gran numero sul fondo, ai piedi dellescogliere. La situazione di “stress ambientale” è stataconfermata dalla scarsa presenza di gasteropodi tipicidi questo piano, quali Monodonta turbinata. Nell’oriz-zonte inferiore dello stesso piano risultavano inoltrepraticamente assenti alcune specie caratteristiche, trale quali il comune pomodoro di mare (Actinia equina).

SANSONI et al. - Ostreopsis ovata: un problema emergente 21
Di particolare rilievo è stato il comportamento ano-malo riscontrato nei banchi naturali di mitili (Mytilusgalloprovincialis), localizzati preferenzialmente sullescogliere semisommerse parallele alla linea di riva.Questi, infatti, anziché aderire tenacemente al substra-to, mostravano un evidente allentamento dei filamentidel bisso, così che i bivalvi oscillavano, in modoinnaturale, con il moto ondoso. Nei banchi di mitili,inoltre, si manifestava la mortalità di una frazioneelevata della popolazione; sul fondo erano infatti pre-senti numerose valve vuote.
Nel Piano Infralitorale, l’impatto della fioritura erarappresentato dalla perdita degli aculei, in percentualevariabile fino alla loro totale scomparsa, rilevata innumerosi esemplari di riccio di mare (Paracentrotuslividus) (Fig. 5). Inoltre, numerosi dermascheletri de-positati sul fondo testimoniavano l’elevata mortalitàche aveva colpito questa specie (caratterizzata, comealtre interessate al fenomeno, da una ridotta vagilità).La risposta allo stress ambientale fornita dalla popola-zione del comune asteroide Coscinasterias tenuispinasi manifestava invece nell’anomala postura delle brac-cia, riflesse verso il dorso. Sempre nello stesso pianosono risultati inoltre assenti anche le spugne e le asci-die (es. Halocynthia papillosa), solitamente frequentie comuni nel piano infralitorale.
Nelle aree sottoposte a tale fenomeno colpiva l’aspettouniforme degli scogli sommersi, in gran parte spoglidall’abituale copertura di fitobenthos e zoobenthos ericoperti invece da una pellicola marrone di consisten-za gelatinosa, i cui lembi flaccidi assecondavano ilmoto ondoso. Frammenti della pellicola, residui di unaprecedente copertura più estesa, aderivano anche allevalve dei mitili e agli aculei dei ricci.
All’esame microscopico la pellicola risultava costi-tuita da una trama gelatinosa che ricopriva i residui diun corto tallo algale e inglobava particelle mineralipolverulente, microalghe di diverse specie e organismianimali microscopici (ciliati, gastrotrichi, nematodi,larve di crostacei, ecc.). La sua superficie esternaappariva, invece, letteralmente brulicante della peridi-nea O. ovata (Fig. 6), il cui fitto rivestimento microal-gale conferiva il colore marrone-rossasto alla pellicola.
Nel 1998, nei fondali a prevalente substrato mobile,soprattutto in prossimità delle scogliere e all’internodegli specchi d’acqua a minor ricambio, si rilevavanotracce più o meno marcate d’anaerobiosi. Tali tracce,consistenti in chiazze nerastre dovute all’azione dibatteri solfito-riduttori, si rendevano particolarmenteevidenti sollevando i massi (diametro 30-70 cm) edosservandone la superficie precedentemente infossatanel substrato sabbioso. La frequenza, l’estensione el’intensità contenuta del colore nerastro riscontratedeponevano per un fenomeno pregresso molto esteso
di anaerobiosi dei fondali e ormai superato o in fase disuperamento, ad eccezione di siti localizzati. Tracceevidenti d’anaerobiosi sono state riscontrate con fre-quenza anche nei primi 20 cm di sedimenti sabbiosi,nei tratti di mare più chiusi, mentre la superficie delfondale appariva in condizioni normali. L’anaerobiosidei sedimenti sabbiosi è, invece, risultata assente neitratti di mare non racchiusi da scogliere.
Nei substrati mobili (sabbiosi) più prossimi alla rivasono stati inoltre rinvenuti dermascheletri di Spatangidi(echinidi irregolari), la cui morte recente era testimo-niata dalla persistenza in situ dei sottili aculei, a testi-monianza che anche tali aree erano state soggettedirettamente o indirettamente alle conseguenze del fe-nomeno di fioritura algale precedentemente menziona-to.
Nell’agosto 2000 è stato rilevato, nella stessa zona,un quadro analogo, ma di minor intensità: la popolazio-
Fig. 5. Sopra: esemplari sopravvissuti di Paracentrotus lividusin differenti stadi di perdita degli aculei. Sotto: esemplarisopravvissuti di Coscinasterias tenuispina con numero ridottodi braccia.
Fig. 6. Il biofilm bentonico che tappezza gli scogli sommersi èbrulicante di O. ovata.

SANSONI et al. - Ostreopsis ovata: un problema emergente22
ne di alghe macroscopiche del Piano Mesolitorale disubstrato duro, infatti, non mostrava sofferenza, men-tre il tappeto algale subiva una certa rarefazione amaggiori profondità. In ogni caso, sia i popolamentialgali sia i fondali anche sabbiosi oltre i due metri diprofondità erano ricoperti dalla pellicola di O. ovata,sotto forma di una trama reticolare di colore marrone-rossastro. Le tracce d’anaerobiosi risultavano moltocontenute.
Nell’estate 2001, a parte la rarefazione dei popola-menti di macroalghe alle maggiori profondità (ridottealla sola parte basale del tallo), l’intensità del fenomenoappariva ancora minore: i popolamenti di echinidi nonevidenziavano una mortalità di rilievo, ma alcuni esem-plari presentavano soltanto la perdita parziale o totaledegli aculei (in circa il 30% della popolazione) mentreuna apprezzabile mortalità si evidenziava nei banchinaturali di mitili. Sui fondali non si rilevavano tracceevidenti di anaerobiosi.
Nell’agosto 1998, in concomitanza con il rilevamentodella fioritura algale, un centinaio di persone che avevasoggiornato sul litorale antistante il tratto marino mag-giormente interessato dalla fioritura algale manifestòmalesseri (irritazioni congiuntivali e delle prime vie ae-ree, dolori muscolari e articolari, tosse, febbre). Lasintomatologia insorgeva dopo 2-3 ore d’esposizioneall’aerosol marino e regrediva nel giro di 12 ore, ricom-parendo a seguito di una nuova esposizione. I casi dipersone colpite dagli stessi sintomi si sono ridotti apoche decine nell’agosto 2000 e a poche unità nel 2001.
CONCLUSIONII dati raccolti e le osservazioni effettuate nelle sta-
gioni estive del 1998, 2000 e 2001 consentono diaffermare che nelle acque costiere apuane la fiorituradella microalga bentonica O. ovata si manifesta conuna certa frequenza. La fioritura sembra avere provo-cato, direttamente o indirettamente, morie dei popola-menti animali e macroalgali sommersi. L’intensità delfenomeno è risultata massima in aree marine che,essendo chiuse da “pennelli” perpendicolari alla riva e
da una doppia barriera soffolta, parallela ad essa, sonosicuramente soggette ad un ridotto ricambio idrico ead un eccessivo riscaldamento delle acque in condizio-ni meteorologiche particolari. Ciò considerato si puòragionevolmente ritenere che la caduta degli aculeidegli echinidi, uno degli aspetti più appariscenti delfenomeno, non dipenda esclusivamente dalla produ-zione di tossine ma possa essere attribuita anche acondizioni ambientali avverse quali il riscaldamentodelle acque e la conseguente diminuzione del tenore diossigeno (NICHOLS, 1966).
Altre informazioni, raccolte nel 1998 da colleghi esubacquei (ABATE M., PALMIERI F., comunicazionipersonali), consentono d’affermare che la fioritura diO. ovata e le morie di organismi marini hanno interes-sato anche le acque costiere della Liguria di levante edell’arcipelago toscano, indicando che si è trattato diun fenomeno molto esteso.
Una sintomatologia umana del tutto analoga a quellaosservata nel litorale apuano è stata segnalata nell’esta-te 2001 in quello barese, dove è stata attribuita dairicercatori del locale istituto di biologia marina allabalneazione in acque con fioritura di O. ovata(BARIWEB.NET, 2001). Nel litorale apuano, gran partedelle persone colpite non si era immersa nelle acqueper cui si può ritenere che i disturbi possano esserestati causati dall’inalazione di sostanze irritanti veicola-te dagli aerosol marini, un fenomeno finora segnalatoin letteratura solo per Gymnodinium breve (HEMMERT,1975; PIERCE et al., 1989, 1990; WATANABE et al.,1988).
Non si dispone di elementi per stabilire se responsa-bile dei malesseri sia una tossina prodotta dalla micro-alga, oppure sostanze derivanti dalla decomposizionedegli animali e vegetali marini morti. La mancata mani-festazione di un’analoga sintomatologia in altri casi dimorie di animali marini depone per la prima ipotesi.
Più motivi suggeriscono quindi la necessità d’ulte-riori indagini volte a chiarire ed approfondire le causedella fioritura algale, l’estensione geografica, la ricor-renza e le conseguenze del fenomeno.
BIBLIOGRAFIA
Pagina web del 20.10.2001.BOMBER J. W., RUBIO M. G. and NORRIS D. R., 1989. Epiphytism
of dinoflagellates associated with the disease ciguatera: sub-strate specifity and nutrition. Phycologia, 28: 360-368.
BRUSLÉ J., 1997. Ciguatera fish poisoning - a review. Sanitary andeconomic aspects. INSERM, Paris, 147 pp.
FUKUYO Y., 1981. Taxonomical study of benthic dinoflagellates,
ANDERSON, D. M., P. S. LOBEL, 1987. The continuing enigma ofciguatera. Biol. Bull., 172: 89-107.
ANDERSON, D. M. 1989. Toxic algal blooms and red tides: a globalperspective. In T. Okaichi, D. M. Anderson and T. Nemoto(eds.), Red Tides: Biology Environmental Science and Toxico-logy, Elsevier, New York: 11-16.
BARIWEB.NET, 2001. Attualità. Mare barese, tra squali e alghe killer.

SANSONI et al. - Ostreopsis ovata: un problema emergente 23
collected in coral reefs. Bull. Jap. Soc. Sci. Fish. 47 (8): 967-978.GLAZIOU P. and LEGRAND A. M., 1994. The epidemiology of
ciguatera fish poisoning. Toxicon, 31: 1151-1154.HALLEGRAEFF, G. M., 1993. A review of harmful algal blooms and
their apparent global increase. Phycologia, 32: 79-99.HEMMERT W.H., 1975. The public health implications of Gymno-
diniurn breve red tides, a review of the literature and recentevents. Proceedings of the First International Conference onToxic Dinoflagellate Blooms. Boston, MIT: 489-497.
NICHOLS D. , 1966. Echinoderms. Hutchinson & Co., LondonPIERCE, R., M. HENRY, S. BOGGESS and A. RULE, 1989. Marine toxins
in bubble-generated aerosol. In: E. Monahan and P. van Patton(eds.), The Climate and Health Implications of Bubble-MediatedSea-Air Exchange, Connecticut Sea Grant Publications: 27-42.
PIERCE R.H., M.S. HENRY, L.S. PROFFITT and P.A. HASBROUCK, 1990.Red tide toxin (brevetoxin) enrichment in marine aerosol. In: E.Graneli, S. Sundstron, L. Elder and D.M. Anderson (eds.),Toxic Marine Phytoplankton: 397-402.
RUSSEL F. E, and EGEN N.B., 1991. Ciguatera fishes, ciguatoxin(CTX) and ciguatera poisoning. J. Toxicol. - toxin review, 10(1): 37-62.
VOLTERRA L., 1989. Alghe produttrici di biotossine. BiologiaAmbientale, III (5): 5-17.
WATANABE T., LOCKEY R.F., KRZANOWSKI J.J., 1988. Airwaysmooth muscle contraction induced by Ptychodiscus brevis(red tide) toxin as related to a trigger mechanism of bronchialasthma. Immuno Allergy Pract., 10 (5):185-192.


Biologia Ambientale, 17 (1): 25-33, 2003
Osservazioni preliminari sulla predazione della Trota fario edella Trota marmorata nei confronti dei macroinvertebratibentonici presenti nel drift del torrente But (Bacino del fiumeTagliamento, Friuli Venezia Giulia)
Elisabetta Pizzul*, Paola Bottos, Silvia Vanzo e Gloria Tosolini
Dipartimento di Biologia – Università di Trieste, Via Weiss 2 - 34127 Trieste
* Autore referente per la corrispondenza (Fax 040-5582011; e-mail: [email protected])
Pervenuto il 3.2.2002; accettato il 16.7.2002
RIASSUNTOViene analizzata la presenza di macroinvertebrati bentonici nella dieta di Salmo (trutta) trutta e Salmo (trutta) marmoratus in relazione allapresenza di questi organismi nel drift del torrente But, principale affluente montano del fiume Tagliamento.Lo studio è stato condotto mediante campionamento stagionale degli organismi presenti nel drift ed analisi dei contenuti stomacali deiSalmonidi in due stazioni fisse: Timau e Noiaris, nelle quali precedentemente era stata rilevata la presenza rispettivamente di popolazionidi Salmo (trutta) trutta e Salmo (trutta) marmoratus, quest’ultima sostenuta da un piano di reintroduzione. La dieta dei due Salmonidi èrisultata basata esclusivamente sulla predazione di macroinvertebrati bentonici, più in particolare nella dieta di Salmo (trutta) truttaprevalgono i Tricotteri ed i Plecotteri, mentre in Salmo (trutta) marmoratus i Ditteri e gli Efemerotteri.
PAROLE CHIAVE: Salmo (trutta) trutta / Salmo (trutta) marmoratus / contenuto stomacale / macroinvertebrati bentonici di deriva.
AbstractPreliminary observations on brown trout and marble trout predation of drifting benthic macroinvertebrates of River But(Tagliamento River basin, Friuli Venezia Giulia, Italy)The presence of benthic macroinvertebrates in the diet of Salmo (trutta) trutta and Salmo (trutta) marmoratus was analysed andcompared with the occurrence of these organisms in the drift of river But, main mountain tributary of Tagliamento River. Seasonalsamplings of drifting macroinvertebrates and examinations of salmonids stomach contents were carried out in two stations: Timau andNoiaris. In the first station lives Salmo (trutta) trutta, instead in the second Salmo (trutta) marmoratus, that is supported by areintroduction plain. The diet of the two Salmonids resulted to be composed entirely of benthic macroinvertebrates. In particular the dietof Salmo (trutta) trutta was composed mostly of Trichoptera and Plecoptera species, on the other hand the diet of Salmo (trutta)marmoratus was composed of Diptera and Ephemerptera species.
KEY WORDS: Salmo (trutta) trutta / Salmo (trutta) marmoratus / stomach contents / drifting macroinvertebrates.
INTRODUZIONELo studio delle comunità animali che popolano gli
ambienti acquatici riveste notevole importanza in quantopermette l’acquisizione di dati utili ad una loro correttagestione ed inoltre fornisce un giudizio sulla qualità esulla funzionalità del sistema acquatico stesso. Negliultimi anni è infatti risultato evidente che affidarsi adinterventi che non seguano corretti modelli gestionali,limitati il più delle volte alla sola semina massiccia dimateriale ittico, significa innanzitutto alterare, in modo
consistente, l’equilibrio dell’ecosistema acquatico e,da ultimo, trovarsi di fronte ad operazioni che si risol-vono anche in uno spreco economico.
Nella regione Friuli Venezia Giulia da più di diecianni vengono condotti ripopolamenti con il ceppo au-toctono dell’emispecie Salmo (trutta) marmoratus.
A questo fine vengono periodicamente monitorati itratti di fiumi e di torrenti più idonei ai ripopolamenti,essendo consapevoli che una convivenza della Trota

PIZZUL et al. - Predazione dei Salmonidi sugli organismi di deriva26
marmorata con l’altra trota presente, la Trota fario,Salmo (trutta) trutta, potrebbe rendere vano ogni sfor-zo, poiché le due forme possono incrociarsi e dar vitaad esemplari ibridi, a loro volta fertili. Nell’intraprende-re i ripopolamenti della Trota marmorata, oltre a chiu-dere alla pesca sportiva le aree di immissione e adaccertarsi che nelle stesse non vi sia la presenza dellaTrota fario, è stato ritenuto importante osservare sequesti ambienti fossero idonei anche da un punto divista trofico e quindi analizzare la comunità a macroin-vertebrati bentonici, organismi che costituiscono unelemento importante nella dieta sia della Trota fario chedella Trota marmorata (GANDOLFI et al., 1991). Inparticolare in un recente studio condotto sul torrenteAupa (bacino del Tagliamento), è stato osservato chel’alimentazione della Trota fario, in questo torrente(ma è ipotizzabile anche in altri corsi d’acqua dallecaratteristiche idrologiche simili), è basata sulla pre-senza di invertebrati derivanti più che su quelli bentoni-ci (MORGAN et al., in stampa).
Il presente studio, volto a caratterizzare lo spettroalimentare della Trota fario e della Trota marmorata inrelazione alla presenza di organismi macrobentonici dideriva, è stato condotto nel torrente But, affluentemontano del fiume Tagliamento. In base a ricerchecondotte su tutti i corsi d’acqua del Friuli VeneziaGiulia negli anni 1996-2000 (MORO et al., 1998; VANZOet al., 1998), questo torrente è risultato mantenere unatra le popolazioni più numerose e ben strutturate diTrota marmorata, la quale presenta peraltro una distri-buzione pressoché separata rispetto alla Trota farioche occupa quasi esclusivamente il tratto montano deltorrente.
MATERIALI E METODIDal novembre 2000 al settembre 2001 sono stati
condotti quattro campionamenti ittici (novembre–mag-gio–luglio–settembre) e tre campionamenti di macro-invertebrati bentonici (maggio–luglio–settembre) in duestazioni poste sul torrente But: la stazione di Timau e lastazione di Noiaris (Fig. 1). La prima è caratterizzatadalla presenza di esemplari di Trota fario, Salmo (trut-ta) trutta, la seconda da una popolazione numerosa eben strutturata di Trota marmorata, Salmo (trutta)marmoratus. É stato quindi effettuato un confronto trai campioni stagionali di macroinvertebrati bentonicirinvenuti nel drift e nei contenuti stomacali rispettiva-mente di esemplari di Trota fario nella stazione diTimau e di Trota marmorata nella stazione di Noiaris.
La raccolta dei macroinvertebrati bentonici presentinel drift è stata condotta utilizzando la rete di Surber,che può essere facilmente fissata al letto del corsod’acqua. L’ampiezza dell’imboccatura del collettore èpari a 0,5 x 0,5 m ed il retino è fissato al fondo per
mezzo di paletti di ferro sagomati. La profondità dellarete è di 1,5 m e le maglie misurano 500 μm. Al fondodella rete è applicato un raccoglitore, fissato medianteuna ghiera a vite, utilizzato in seguito per la conserva-zione del campione. I campioni sono stati raccoltilasciando la rete in posizione per un’ora.
Nella stazione di Timau l’operazione è stata svoltanelle prime ore del mattino, mentre nella stazione diNoiaris il campionamento è stato eseguito all’imbruni-re. Ciò per il fatto che la deriva dei macroinvertebratibentonici (drift) è caratterizzata da una ciclicità giorna-liera con valori massimi notturni, la quale condiziona lapredazione da parte dei Salmonidi (ELLIOTT, 1967;ALLAN, 1981; GERKING, 1994) che risulta pertanto piùintensa all’alba ed al tramonto, periodi in cui la presen-za di luce, seppure minima, aiuta la cattura. I campioniraccolti sono stati fissati in aldeide formica al 4% esuccessivamente analizzati e determinati in laboratorio.
I campionamenti ittici, condotti nelle stesse aree diraccolta dei macroinvertebrati bentonici, sono statieffettuati mediante uso di un elettrostorditore a corren-te continua pulsata e voltaggio modulabile (0,7–7 Am-père, 150–380 Volt), manovrato da personale dell’EnteTutela Pesca del Friuli Venezia Giulia.
Ad ogni Salmonide catturato è stato associato uncodice che ne consentisse il successivo riconoscimen-
R. Moscardo
T. PontalbaT. Gladegna
T. Saustri
T. Chiarsò
T. BUT
T. B
UT
Noiaris
F. Tagliamento
CERCIVENTO
PALUZZA
SUTRIO
ARTA
ZUGLIO
CEDARCHIS
CADUNEA
CANEVA
TOLMEZZO
Timau
Fig. 1. Torrente But e suoi affluenti

PIZZUL et al. - Predazione dei Salmonidi sugli organismi di deriva 27
to, quindi è stato identificato sistematicamente e ne èstato rilevato il peso e la lunghezza totale. La misura delpeso è stata effettuata con una bilancia digitale conprecisione di 1g, la lunghezza totale per mezzo di unittiometro standard con precisione di 1 mm. Lo stoma-co dei 126 Salmonidi analizzati è stato rimosso sulcampo e posto in alcool al 70%, in un contenitorecontrassegnato con il codice di identificazione del-l’esemplare. In laboratorio gli stomaci sono stati disse-zionati e ne è stato identificato il contenuto. Gli organi-smi rinvenuti sono stati determinati, separati per taxa econservati in alcool al 70%.
Al fine di definire la biomassa dei macroinvertebratibentonici rinvenuti nel drift e nei contenuti stomacalidei pesci, per ogni taxon si è proceduto alla misuradella biomassa espressa come peso secco. I campionisono stati posti in una stufa a 105 °C per ventiquattroore, lasciati raffreddare per altre ventiquattro ore edinfine pesati per determinarne il peso secco. Tutte lepesate sono state eseguite per mezzo di una bilanciaanalitica con precisione di 0,1 mg.
Ad ogni Salmonide, infine, sono stati prelevati gliotoliti, più in particolare le sagitte, al fine di determina-re l’età degli esemplari. Queste strutture, una voltapulite con acqua distillata, sono state osservate intrasparenza al microscopio ottico a luce riflessa.
L’esame della selettività alimentare, considerandogli organismi derivanti come prede disponibili, è statoeffettuato con l’indice di Shorigin (1939), nella suaforma logaritmica per le categorie di prede più fre-quenti, ovvero quelle la cui frequenza è tale da renderestatisticamente valida l’analisi.
L’indice di Shorigin è definito come:S = log10 (Fs / Fd)
dove:S= selettività;Fs= frequenza relativa di una preda nei contenuti sto-macali;Fd= frequenza relativa di una preda nell’ambiente.
RISULTATINel mese di novembre 2000 nella stazione di Timau
sono state catturate esclusivamente Trote fario, Salmo(trutta) trutta. Complessivamente sono stati analizzati15 esemplari, dei quali 7 maschi (Tab. I). L’età deglistessi variava tra 2 e 5 anni. L’esemplare più piccolopresentava un peso di 34 g ed una lunghezza totale di15,1 cm, mentre l’esemplare più grande raggiungeva304 g e 30,3 cm.
Soltanto una trota si presentava digiuna anche se, ingenerale, gli stomaci contenevano un totale di macro-invertebrati piuttosto scarso.
Per quanto concerne i valori del peso secco deidiversi taxa di macroinvertebrati bentonici rinvenuti
negli stomaci degli esemplari, i Tricotteri sono risultaticostituire la componente alimentare più rappresentati-va, in particolare le famiglie Glossosomatidae e Serico-stomatidae. Seguono gli Imenotteri, benché la loronumerosità non sia molto elevata, come pure quella deiColeotteri e dei Neurotteri. L’ordine dei Plecotteri,rappresentato dal genere Leuctra, non raggiunge i 0,01g ma è tra i taxon numericamente più abbondanti (Fig.2A).
Nella stazione di Noiaris, sempre nel mese di no-vembre 2000, sono stati pescati 15 esemplari di Salmo(trutta) marmoratus con età comprese tra 2 e 4 anni.L’esemplare più piccolo presentava un peso di 55 g eduna lunghezza di 17,6 cm, mentre le dimensioni mag-giori sono relative ad un maschio di 3 anni che rag-giungeva 233 g e 27,8 cm (Tab. I).
Tra i macroinvertebrati bentonici rinvenuti neglistomaci, gli Efemerotteri appartenenti al genere Baetissono risultati i più numerosi. Per quanto concerne idati relativi al peso secco dei diversi taxa osservati, iColeotteri, i Tricotteri ed i Plecotteri sono risultati i piùrappresentativi. Non molto frequenti invece i Ditteri sia
1
0,00
0,01
0,02
0,03
0,04
0,05
0,06
0,07
0,08
0,09
0,10
Pl Ef Trt Clt Dt Et Om Im Nr Cr Gs Mr Ol Trl Ar Clm
peso
sec
co (
g)
contenuto stomacale
19 9
60
4
1
125 1
1
Timau - novembre 2000 (Trota fario)
Noiaris - novembre 2000 (Trota marmorata)
0,00
0,01
0,01
0,02
0,02
0,03
0,03
0,04
0,04
Pl Ef Trt Clt Dt Et Om Im Nr Cr Gs Mr Ol Trl Ar Clm
peso
sec
co (
g)
contenuto stomacale
6
107
1310
10 1 1
Fig. 2. Peso secco e numero di esemplari di macroinvertebratirinvenuti nei contenuti stomacali nel novembre 2000. Su ciascunacolonna è riportato il numero di esemplari.Pl= Plecotteri, Ef= Efemerotteri, Trt= Tricotteri, Clt= Coleotteri,Dt= Ditteri, Et= Eterotteri, Om= Omotteri, Im= Imenotteri, Nr=Neurotteri, Cr= Crostacei, Gs= Gasteropodi, Mr= Mermitidi,Ol= Oligocheti, Trl= Tricladi, Ar= Aracnidi, Clm= Collemboli.
A
B

PIZZUL et al. - Predazione dei Salmonidi sugli organismi di deriva28
per quanto riguarda il loro numero che il peso secco(Fig. 2B).
In questo mese non è stato possibile condurre icampionamenti di macroinvertebrati bentonici nel drifte quindi effettuare confronti con la presenza di questiorganismi nel contenuto stomacale dei pesci.
Nel mese di maggio 2001 nella stazione di Timausono stati catturati 16 esemplari di Trota fario, deiquali 9 maschi e 7 femmine. L’età delle trote eracompresa tra 2 e 4 anni. L’esemplare di dimensionimaggiori pesava 171 g e presentava una lunghezzatotale di 25,9 cm, mentre quello di dimensioni inferioriera un femmina di 68 g e 18,2 cm (Tab. I).
Negli stomaci delle Trote fario catturate il maggiorcontributo in peso secco è dato dai Coleotteri, daiPlecotteri e dai Tricotteri. In questi tre taxa si osserva-no, in figura 3A, dei valori in peso secco che risultano,in rapporto al numero di esemplari, diversi tra drift econtenuto stomacale; ciò è in relazione alle diversefamiglie, generi o stadi di sviluppo rinvenuti nei cam-pioni. L’ordine dei Plecotteri, rappresentato dai generiNemoura, Isoperla, Leuctra e Capnia, e quello deiTricotteri, in particolare le famiglie Limnephilidae, Glos-sosomatidae e Sericostomatidae, risultano i taxa nu-mericamente più abbondanti. Nel contenuto stomacaledi qualche esemplare sono stati rinvenuti Crostacei(Fig. 3A).
L’analisi dei macroinvertebrati bentonici presentinel drift ha permesso di rilevare la prevalenza, sia inpeso secco che in numero di individui, dei Ditteri ed inparticolare di forme larvali della famiglia Chironomi-dae, seguite dalle forme pupali ed adulte della stessafamiglia che in questo periodo è in sfarfallamento.Elevato è inoltre il valore del peso secco degli Efeme-rotteri, tra i quali i generi numericamente più abbon-danti sono Baetis ed Ecdyonurus. I Plecotteri, qualiquelli appartenenti alle famiglie Nemouridae, Leuctri-dae e Perlodidae, sono numericamente più abbondanti
degli Efemerotteri ma il loro peso secco è prossimo aquello dei Tricotteri che sono invece poco numerosi.Presenti anche Oligocheti e Aracnidi terrestri caduticon molta probabilità accidentalmente in acqua (Fig.3A). Nella tabella II sono riportati i dati relativi allaselettività di predazione calcolata mediante l’indice diShorigin, i valori negativi indicano che nell’ambiente lafrequenza del taxon preso in esame è maggiore rispettoalla frequenza registrata nei contenuti stomacali, il
Tab. I. Misure biometriche ed età degli esemplari analizzati
Staz. Mese Specie n° Peso min. Peso max Media Lung. min Lung. max Media Età Sesso
Timau 11/2000 T. fario 15 34 304 136 15,1 30,3 22,6 2 - 5 8F - 7MNoiaris 11/2000 T.marmorata 15 55 233 130 17,6 27,8 22,9 2 - 4 7F - 8MTimau 5/2001 T. fario 16 68 171 111,7 18,2 25,9 21,8 2 - 4 7F - 9MNoiaris 5/2001 T. marmorata 14 42 357 180,2 16,2 34,9 25,8 2 - 5 7F - 7MTimau 7/2001 T. fario 15 47 199 104,2 17 26,8 21 2 - 4 7F - 8MNoiaris 7/2001 T. marmorata 18 29 345 160 14,5 32,2 24,7 2 - 4 11F - 7MTimau 9/2001 T. fario 16 19 136 58,2 12,3 23,3 17 2 - 4 7F - 9MNoiaris 9/2001 T. marmorata 17 18 45 30,6 12 17,6 14,2 2 - 4 7F - 10M
F= femmine; M= maschi; n°= numero esemplari
Timau - maggio 2001 (Trota fario)
0,00
0,05
0,10
0,15
0,20
Pl Ef Trt Clt Dt Et Om Im Nr Cr Gs Mr Ol Trl Ar Clm
peso
sec
co (
g)
driftcontenuto stomacale
196131
278
12
38
105
6
4
1237
223 12
6 13
39 442
Noiaris - maggio 2001 (Trota marmorata)
0,00
0,05
0,10
0,15
0,20
Pl Ef Trt Clt Dt Et Om Im Nr Cr Gs Mr Ol Trl Ar Clm
peso
sec
co (
g)
driftcontenuto stomacale
4
131
36
12 2
105
2
41504
223 29 1
6
32 119 7
A
B
Fig. 3. Peso secco e numero di esemplari di macroinvertebratirinvenuti nel drift e nei contenuti stomacali nel maggio 2001. Peril significato delle abbreviazioni sulle ascisse si veda la figura 2.Su ciascuna colonna è riportato il numero di esemplari.

PIZZUL et al. - Predazione dei Salmonidi sugli organismi di deriva 29
contrario nel caso di valori di selettività positiva. Per gliesemplari di Trota fario; si riscontra una selettivitànegativa per quanto concerne gli Efemerotteri, i Tri-cotteri della famiglia Rhyacophilidae ed i Ditteri.
Nella stazione di Noiaris, sono stati pescati 14 esem-plari di Trota marmorata, 7 dei quali maschi. L’etàvariava tra 2 e 5 anni; l’esemplare di maggiori dimen-sioni pesava 357 g ed aveva lunghezza totale pari a34,9 cm, in quello più piccolo il peso era di 42 g e lalunghezza di 16,5 cm (Tab. I). Nei contenuti stomacalidi questi esemplari l’ordine dei Ditteri è numericamen-te quello più cospicuo, benché basso sia il peso seccoregistrato, contrariamente a quanto osservato per iColeotteri. I Plecotteri ed i Tricotteri, questi ultimirappresentati dalle famiglie Limnephilidae, Glossoso-matidae e Beraeidae, rappresentano sia in peso seccoche in numero una componente importante della dieta(Fig. 3B).
Nel campione di drift della stessa stazione, moltonumerosi sono i Ditteri nelle forme adulte, larvali epupali di Chironomidi, i quali registrano anche il piùelevato valore in peso secco. Gli Efemerotteri, delgenere Baetis, sono il secondo taxon in termini diabbondanza, mentre i Plecotteri sono rappresentatiesclusivamente dai generi Leuctra e Capnia. Anche iTricotteri sono rari (Fig. 3B). In tabella II si puòosservare che i valori di selettività negativa si riferisco-no, per la Trota marmorata, ai Ditteri adulti ed allepupe di Chironomidi.
Nel mese di luglio, per quanto concerne i campiona-menti ittici, nella stazione di Timau sono stati catturati15 esemplari di Trota fario di cui 7 femmine. L’etàvariava tra 2 e 4 anni, mentre peso e lunghezza totaleerano rispettivamente compresi tra 47 e 199 g e tra 17e 26,8 cm (Tab. I).
Dall’analisi dei contenuti stomacali numeroso è ri-sultato l’ordine degli Efemerotteri rappresentato so-prattutto dal genere Baetis, il cui contributo in pesosecco è tuttavia modesto. Segue l’ordine dei Tricottericostituito da un alto numero di famiglie con e senzafodero, che registra il valore in peso secco più elevato.Seguono in numero i Ditteri, presenti in maggior partecon forme adulte e larvali della famiglia Chironomidae,gli Imenotteri ed i Plecotteri (Fig. 4A).
Nel campione di drift, relativo alla stessa stazione,prevalgono, per quanto concerne i valori del pesosecco, gli Efemerotteri del genere Baetis, quindi iPlecotteri, rappresentati dai generi Protonemura, Iso-perla e Leuctra. I Ditteri, in particolare le forme larvalidella famiglia Chironomidae, sono invece il taxon nu-mericamente più abbondante. I Coleotteri compaionosporadicamente nelle forme adulte e rari, ma presenti,sono gli Aracnidi (Fig. 4A).
In tabella III i valori di selettività, riferiti alla preda-zione della Trota fario sugli organismi di drift, riporta-no valori negativi per i Plecotteri, i Coleotteri e le larvedi Ditteri Chironomidi.
Sempre in luglio nella stazione di Noiaris sono staticatturati 18 esemplari di Salmo (trutta) marmoratus, lacui età variava tra 2 e 4 anni, il peso era compreso tra29 e 345 g e la lunghezza totale tra 14,5 e 32,2 cm(Tab. I).
Nel contenuto stomacale, gli Efemerotteri, del gene-re Baetis, ed i Ditteri, con le famiglie Simuliidae,Chironomidae ed Empididae, sono i taxa numerica-mente più cospicui. Per quanto invece concerne ilpeso secco, i Gasteropodi appartenenti ai generi Val-vata e Lymnaea, ed i Tricotteri registrano i valori piùelevati. (Fig. 4B).
Nel campione di drift della stazione prevalgono iDitteri adulti e le larve e le pupe della famiglia Chirono-midae. Seguono gli Oligocheti e gli Efemerotteri, que-
Tab. II. Selettività alimentare calcolata mediante l’indice diSHORIGIN (1939) in relazione ai gruppi predati con maggiorefrequenza nel mese di maggio 2001
TIMAU
T. fario Taxa selettività
Plecotteri Nemoura 0,68Protonemura 0,38Amphinemura 0,21
Efemerotteri Ecdyonurus -0,08Baetis -0,69
Tricotteri Rhyacophilidae -0,79Philopotamidae 0,59Sericostomatidae 0,92Beraeidae 0,89Limnephilidae 2,14
Ditteri Dittero adulto -0,74Chironomidae larva -0,05Chironomidae pupa -0,21Limoniidae larva -0,56Empididae pupa -0,01
NOIARIS
T. marmorata Taxa selettività
Efemerotteri Baetis 0,07
Tricotteri Rhyacophilidae 0,53Beraeidae 1,13
Ditteri Dittero adulto -1,50Chironomidae larva 0,08Chironomidae pupa -0,22

PIZZUL et al. - Predazione dei Salmonidi sugli organismi di deriva30
sti ultimi più frequenti rispetto alla precedente stagionee rappresentati principalmente dal genere Baetis. IPlecotteri sono molto rari, ed assenti i Tricotteri (Fig.4B).
Per quanto concerne i valori di selettività questiriportano valori negativi per i Ditteri adulti e le formelarvali e pupali di Chironomidi (Tab. III).
Infine, in settembre nella stazione di Timausono stati catturati 16 esemplari di Salmo (trutta)trutta, di cui 9 di sesso maschile. L’età variava da 2 a 4anni. L’esemplare di maggiori dimensioni presentavaun peso di 136 g ed una lunghezza totale pari a 23,3cm, l’esemplare più piccolo invece pesava 19 g eraggiungeva 12,3 cm di lunghezza (Tab. I).
Per quanto concerne i macroinvertebrati bentonicirinvenuti negli stomaci degli esemplari analizzati, piùnumerosi sono risultati i Ditteri, i Plecotteri, gli Efeme-rotteri ed i Tricotteri. Gli Efemerotteri sono rappresen-tati dal genere Baetis, i Tricotteri dalla famiglia Glosso-somatidae, i Plecotteri dal genere Leuctra ed infine iDitteri da forme larvali appartenenti alle famiglie Simu-liidae e Chironomidae. (Fig. 5A).
Nel campione di drift della stazione di Timau, relati-vo allo stesso mese, prevalgono i Ditteri con le formelarvali della famiglia Chironomidae, mentre le forme
adulte sono pressappoco dimezzate di numero rispettoagli altri campionamenti. Quindi seguono gli Efemerot-teri, rappresentati dal genere Baetis, ed i Gasteropodidel genere Valvata (Fig. 5A). Gli Efemerotteri regi-strano un valore in peso secco nel drift superiorerispetto a quello osservato nei contenuti stomacali, puressendo presenti in numero inferiore a causa delladiversa dimensione degli esemplari osservati nei duecampioni.
Anche in questo caso i valori di selettività nellapredazione della Trota fario risultano negativi per iDitteri e più in particolare per le forme larvali e le pupe
Timau - luglio 2001 (Trota fario)
0,00
0,10
0,20
0,30
0,40
0,50
0,60
0,70
0,80
Pl Ef Trt Clt Dt Et Om Im Nr Cr Gs Mr Ol Trl Ar Clm
peso
sec
co (
g)
driftcontenuto stomacale
241
6
804
195
2
132
18 2
2539
47 6 206 2 1 67 6
Noiaris - luglio 2001 (Trota marmorata)
0,00
0,05
0,10
0,15
0,20
0,25
0,30
0,35
0,40
Pl Ef Trt Clt Dt Et Om Im Nr Cr Gs Mr Ol Trl Ar Clm
peso
sec
co (
g)
driftcontenuto stomacale
41112 153
22
6 2
2700
1056 18
76
4
212
8 1 2
Fig. 4. Peso secco e numero di esemplari di macroinvertebratirinvenuti nel drift e nei contenuti stomacali nel luglio 2001. Suciascuna colonna è riportato il numero di esemplari. Per ilsignificato delle abbreviazioni sulle ascisse si veda la figura 2.
A
B
Tab. III. Selettività alimentare calcolata mediante l’indice diSHORIGIN (1939) in relazione ai gruppi predati con maggiorefrequenza nel mese di luglio 2001
TIMAU
T. fario Taxa selettività
Plecotteri Isoperla -0,45Leuctra -0,35Protonemura -1,21
Efemerotteri Efemerottero adulto 1,65Rhithrogena 0,28Baetis 0,31
Coleotteri Dryopidae -0,05
Ditteri Dittero adulto 0,01Blephariceridae larva 0,65Simuliidae larva 0,12Chironomidae larva -1,26Psychodidae larva 0,95Empididae larva 1,13
Imenotteri Imenottero adulto 1,17
Eterotteri Velia 0,17
NOIARIS
T. marmorata Taxa selettività
Efemerotteri Efemerottero adulto 1,30Baetis 0,94
Ditteri Dittero adulto -1,36Simuliidae larva 0,78Simuliidae pupa 0,78Chironomidae larva -1,32Chironomidae pupa -1,71Empididae larva 0,96
Imenotteri Formicidae 1,78Imenottero adulto 0,78
Aracnidi Ipsodidae 0,004

PIZZUL et al. - Predazione dei Salmonidi sugli organismi di deriva 31
di Chironomidi (Tab. IV).Sempre nello stesso mese, nella stazione di Noiaris
sono state pescate 17 trote marmorate, di cui 7 disesso femminile. Gli esemplari presentavano età com-prese tra 2 e 4 anni, il peso variava tra 18 e 45 g,mentre la lunghezza totale tra 12 e 17,6 cm (Tab. I).
La dieta degli esemplari è risultata costituita preva-lentemente da Ditteri, in particolare larve della famigliaChironomidae e pupe delle famiglie Simuliidae e Psycho-didae. Frequenti nella dieta anche gli Efemerotteri ap-partenenti ai generi Baetis, Ephemerella e Rhitrogena.Presenti Coleotteri e Plecotteri, questi ultimi rappre-sentati soprattutto dalla famiglia Nemouridae (Fig. 5B).Per i Coleotteri si registra un peso secco superiore neicontenuti stomacali rispetto a quello registrato neldrift, benché il numero di esemplari analizzati sia infe-riore, anche in questo caso ciò è legato alla differentenumerosità nei due campioni dei diversi stadi di svilup-po degli organismi.
Nel campione di drift di Noiaris i Ditteri sono iltaxon che registra il maggior valore in peso secco,mentre gli Efemerotteri risultano l’ordine numerica-mente più frequente. Presenti Emitteri ed Aracnidi,probabilmente per le piogge che hanno preceduto ilcampionamento e che hanno trascinato questi esem-
plari nel torrente (Fig. 5B).Selettività negativa è stata osservata nella predazio-
ne degli Efemerotteri del genere Baetis, per i Ditteriadulti e per le pupe di Chironomidi e le forme larvalidella famiglia Psychodidae (Tab. IV).
DISCUSSIONE E CONCLUSIONINel corso di questo studio i macroinvertebrati ben-
tonici rinvenuti nel drift sono risultati in tutti i campio-namenti numerosi e con un alto livello di diversità inentrambe le stazioni. I campioni hanno denunciato unadiversa composizione in taxa, dovuta anche alla diver-sa propensione degli stessi a subire il trasporto versovalle, nonché all’avvicendamento dei periodi di sfarfal-lamento e, quindi, di abbandono dell’ambiente acquati-co.
Dall’analisi del contenuto stomacale dei Salmonidipresenti sono state rilevate differenze e similitudininella dieta delle due semispecie.
Più in particolare, nella dieta della Trota fario unacomponente numericamente frequente in tutte le sta-
Tab. IV. Selettività alimentare calcolata mediante l’indice diSHORIGIN (1939) in relazione ai gruppi predati con maggiorefrequenza nel mese di settembre 2001
TIMAU
T. fario Taxa selettività
Plecotteri Leuctra 1,63Efemerotteri Baetis 0,30Tricotteri Glossosomatidae 1,50Ditteri Simuliidae larva 1,35
Chironomidae larva -0,56Chironomidae pupa -1,39
NOIARIS
T. marmorata Taxa selettività
Plecotteri Leuctra 0,68
Efemerotteri Efemerottero adulto 0,24Baetis -0,56
Coleotteri Dytiscidae larva 0,70
Ditteri Dittero adulto -0,40Simuliidae larva 0,27Simuliidae pupa 1,39Chironomidae larva 0,98Chironomidae pupa -0,27Psychodidae larva -0,04Psychodidae pupa 1,18Tipulidae larva 0,48Empididae larva 0,77Empididae pupa 1,41
Timau - settembre 2001 (Trota fario)
0,00
0,01
0,02
0,03
0,04
0,05
0,06
Pl Ef Trt Clt Dt Et Om Im Nr Cr Gs Mr Ol Trl Ar Clm
peso
sec
co (
g)
driftcontenuto stomacale
1
106
60
92
1
30
360
1086
4 311
Noiaris - settembre 2001 (Trota marmorata)
0,00
0,05
0,10
0,15
0,20
0,25
0,30
Pl Ef Trt Clt Dt Et Om Im Nr Cr Gs Mr Ol Trl Ar Clm
peso
sec
co (
g)
driftcontenuto stomacale
10
12
3654
177
28
25
2472
806
6 1 4 62
Fig. 5. Peso secco e numero di esemplari di macroinvertebratirinvenuti nel drift e nei contenuti stomacali nel settembre 2001.Su ciascuna colonna è riportato il numero di esemplari. Per ilsignificato delle abbreviazioni sulle ascisse si veda la figura 2.
A
B

PIZZUL et al. - Predazione dei Salmonidi sugli organismi di deriva32
gioni è rappresentata dai Tricotteri i quali, al contrario,non risultano tra le prede preferenziali della Trotamarmorata, benché il loro peso secco sia in alcunicampioni elevato. I Tricotteri sono stati rinvenuti rara-mente nel drift poiché non mostrano una grande attitu-dine alla deriva ed il retino di Surber non è risultatoadatto al campionamento: i Tricotteri accidentalmentecaduti nel retino, possono infatti arrampicarsi sullemaglie della rete, vincere la corrente ed uscire dall’im-boccatura. Gli esemplari catturati appartenevano so-prattutto alla famiglia Rhyacophilidae, che dimostrauna scarsa attitudine alla deriva accidentale poiché gliesemplari non sono buoni nuotatori (MORETTI, 1983).Con probabilità gli esemplari di questa famiglia, vengo-no strappati dalla corrente quando si trovano sullasuperficie di pietre sommerse. La predazione quindi acarico di questi organismi è collegabile alla loro pre-senza sul fondo piuttosto che nel drift.
Frequenti in numero nei contenuti stomacali dellaTrota fario sono poi risultati i Plecotteri e gli Efemerot-teri. Di questi il genere Rhithrogena, presente in en-trambe le stazioni, mostra nella stazione di Timau il suomassimo di deriva nel mese di luglio in corrispondenzadel periodo di emersione degli adulti. Con molta proba-bilità l’attitudine alla deriva del genere è maggiore nelperiodo di sfarfallamento. Trattandosi di un Heptageni-dae, è un macroinvertebrato che ha un elevata resi-stenza alla corrente per cui la deriva, come riportato daBELFIORE (1983), è accidentale e legata al momentodell’emersione. Gli Efemerotteri del genere Baetis han-no un massimo di frequenza nel drift nel mese di luglio,in periodo di sfarfallamento, anche se la propensionealla deriva fa sì che gli esemplari siano presenti in tuttele stagioni. La deriva di Baetis è dunque sia accidentaleche comportamentale.
Nella dieta della Trota marmorata, in tutti i campio-namenti ad eccezione del mese di novembre, elevato èrisultato invece il numero di Ditteri, soprattutto dellafamiglia Chironomidae. La particolare abbondanza diDitteri adulti fra gli organismi di deriva è attribuibile alloro trascinamento accidentale, da parte della corrente,nella fase di deposizione delle uova in ambiente acqua-tico. Questa tesi è supportata dal fatto che non siosserva un numero elevato di pupe nel drift, comesarebbe da attendersi qualora la cattura da parte dellacorrente avvenisse nella fase di risalita e sfarfallamen-to. Gli individui della famiglia Chironomidae, per esem-pio, sono soliti risalire alla superficie per lo sfarfalla-mento nuotando ed utilizzando l’esuvia come base perspiccare il volo. In questo caso, quindi, i Ditteri adultidevono essere considerati come organismi derivanticomportamentali, non volontari; apparterrebbero dun-que alla categoria dell’adult drift secondo la definizio-ne di RADER (1997). L’elevata frequenza di Ditteri nei
contenuti stomacali delle trote, benché queste tenden-zialmente predino in modo più selettivo nei confronti diorganismi di grandi dimensioni poiché più facilmentelocalizzabili, è legata all’elevata numerosità di questoordine nel drift; sovente, infatti, si osservano per iDitteri, sia nella Trota fario che nella Trota marmoratavalori negativi di selettività.
Nei contenuti stomacali della Trota marmorata purefrequenti sono risultati gli Efemerotteri, osservati intutti i campionamenti e sempre presenti in numeroconsistente con l’unica eccezione del mese di maggio.A Noiaris il massimo numero di esemplari nel drift èstato registrato nel mese di settembre.
Esclusivamente negli esemplari di Trota marmorataè stata rilevata la presenza di Gasteropodi.
Nell’analisi dei diversi taxa presenti nel drift e neicontenuti stomacali ed anche nell’analisi dei valori diselettività registrati nella predazione della Trota fario edella Trota marmorata, bisogna tuttavia ricordare ladifficoltà di ottenere nel campionamento di drift cam-pioni che rappresentino in modo oggettivo la composi-zione in taxa degli organismi di deriva. La rete da driftopera infatti una selezione negativa nei confronti degliorganismi che sono in grado di spostarsi contro cor-rente sottraendosi così alla cattura.
In entrambe le stazioni, nei Salmonidi analizzati nonsono stati riscontrati casi di ittiofagia. Rappresentandoquindi gli invertebrati l’unica componente alimentare,si può ritenere che la disponibilità di questi ultimi possarappresentare un elemento determinante per la valuta-zione della capacità di sostentamento della Trota farioe della Trota marmorata in torrenti quali quello studia-to.
Sia per la Trota fario che per la Trota marmoratanon sono state evidenziate differenze significative nellacomposizione della dieta e nel numero di prede inrelazione alla taglia ed all’età degli esemplari.
Bisogna tuttavia sottolineare che, principalmente acausa del ridotto numero di esemplari analizzati, condi-zionato dal piano di ripopolamento della Trota marmo-rata in atto nel torrente, i dati fin qui ottenuti, purrisultando interessanti, richiedono ulteriori approfondi-menti e verifiche. La ricerca, tuttora in corso, prevedecampionamenti qualitativi e quantitativi della fauna stret-tamente bentonica; i dati verranno messi in relazionecon lo spettro alimentare di questi Salmonidi, al fine diaccertare l’effettiva propensione alla predazione sugliorganismi di deriva.
Ringraziamenti
Si ringrazia l’Ente Tutela Pesca del Friuli Venezia Giulia per averfornito i mezzi ed il personale qualificato a condurre i campiona-menti ittici. Un ringraziamento inoltre al dott. Moro per i prezio-si consigli forniti.

PIZZUL et al. - Predazione dei Salmonidi sugli organismi di deriva 33
BIBLIOGRAFIA
ALLAN J.D., 1981. Determinants of diet of brook trout (Salvelinusfontinalis) in a mountain stream. Can. J. Fish. Aquat. Sci., 38:184-192.
BELFIORE C., 1983. Efemerotteri (Ephemeroptera). Guide per ilriconoscimento delle specie animali delle acque interne italia-ne, C.N.R. AQ/1/201, 24, 63 pp.
ELLIOTT J.M., 1967. The food of trout (Salmo trutta) in a Dartmoorstream. J. Appl. Ecol., 4: 59-71.
GANDOLFI G., ZERUNIAN S., TORRICELLI P., MARCONATO A., 1991.I pesci delle acque interne italiane. Istituto Poligrafico e Zeccadello Stato, Roma, 616 pp.
GERKING S.D., 1994. Feeding Ecology of Fish. Academic Press,Inc, San Diego, California, 385 pp.
MORETTI G.P., 1983. Tricotteri (Trichoptera). Guide per il rico-noscimento delle specie animali delle acque interne italiane,C.N.R. AQ/1/196, 19, 149 pp.
MORGAN B., MORO G.A., PIZZUL E., SPECCHI M., in stampa.Macroinvertebrati ed alimentazione della Trota fario nel tor-rente Aupa (Alpi Carniche, Friuli-Venezia Giulia). In: Atti VIIIConv. A.I.I.A.D. Codroipo (Udine), giugno 2000. QuaderniEnte Tutela Pesca del Friuli Venezia Giulia, 30.
MORO G.A., PIZZUL E., VANZO S., SPECCHI M., 1998. Studio sullecomunità macrozoobentoniche ed ittiche del torrente But(bacino del Tagliamento, Friuli Venezia Giulia). QuaderniEnte Tutela Pesca del Friuli Venezia Giulia, 27: 37-60.
RADER R.B., 1997. A functional classification of the drift: traitsthat influence invertebrate availability to Salmonids. CanadianJournal of Fisheries and Acquatic Sciences, 54 (6): 1211-1234.
SHORIGIN A.A., 1939. Food and food preference in some Gobiidaeof the Caspian Sea. Zool. Zh., 18: 27-53.
Vanzo S., Specchi M., Pizzul E., 1998. Studio sulle comunitàittiche del bacino dell’Alto Tagliamento (Nord-Est Italia).Quaderni Ente Tutela Pesca del Friuli Venezia Giulia, 27: 1-13.


Biologia Ambientale, 17 (1): 35-44, 2003
Le rane verdi del complesso Rana esculenta come bioindicatoridella qualità degli ambienti fluviali italiani
Paolo Andreani1 *, Fiammetta Santucci2, Giuseppe Nascetti1.1 Dipartimento di Scienze Ambientali, Università degli Studi della Tuscia, Via S.C. de Lellis - 01100 Viterbo
2 Smithsonian Tropical Research Institute. APO 2072 Aucon, Balboa, Panama, Republic of Panama
* Autore referente per la corrispondenza: fax 0761 357123; e-mail [email protected]
Pervenuto il 15.4.2002; accettato l’8.9.2002
RiassuntoLo studio si propone di verificare le capacità bioindicatrici delle due specie di rane verdi (Rana lessonae e Rana esculenta) appartenential complesso Rana esculenta negli ambienti fluviali italiani. Tali capacità, avvalorate da varie evidenze sperimentali, trovano riscontronella risposta fornita da popolazioni di anfibi alle variazioni della qualità degli ambienti colonizzati. In particolare, per le due specie di raneverdi, si evidenzia come in habitat integri prevalga R. lessonae, in quelli disturbati competa meglio R. esculenta grazie al vantaggio eteroticoderivante dalla sua origine ibrida. L’elevato grado di eterozigosi fornisce a R. esculenta una più ampia capacità di risposta alle variazionidei parametri ecologici.Le frequenze relative delle due specie sono state messe a confronto con le risposte di indici biotici e di naturalità ambientale (I.B.E.,BMWP’ e I.F.F.) in quattro transetti: Oasi di Punte Alberete (RA), Riserva Naturale del Lago di Penne (PE), Tenuta Presidenziale diCastelporziano (RM), Riserva Naturale Orientata Bosco Pantano di Policoro (MT).I risultati mostrano come ad una buona qualità delle acque e ad una elevata funzionalità fluviale corrispondano una dominanza di R.lessonae e viceversa.Le capacità bioindicatrici delle rane verdi del complesso Rana esculenta sembrano confermate e, grazie alle loro caratteristichecomplementari alla comunità dei macroinvertebrati, dovrebbero permettere di valutare il disturbo antropico anche in habitat ove non sonoapplicabili gli indici biotici maggiormente usati: i tratti potamali dei fiumi, i laghi, gli stagni e gli ambienti di transizione.
PAROLE CHIAVE: complesso Rana esculenta / bioindicatori / specie ibride / eterozigosi / competizione.
AbstractFrogs of Rana esculenta complex as Italian river quality bioindicatorsIn this paper we tested as bioindicators two species of pond frogs belonging to the Rana esculenta complex. Their value as biondicators,comproved by several studies, is based on the particular physiological and ecological adaptations of amphibians, which are generallyhighly sensitive to the presence of pollutants. In particular the two species of pond frogs considered here respond differently tovariations of environmental parameters: Rana lessonae prevails in natural habitats, while the hybrid species Rana esculenta competesbetter in polluted habitats. The latter prevails in polluted habitats thanks to the heterotic advantage deriving by its hybrid origin: the highlevel of heterozigosity seems to confer to this species the ability to respond to a wider spectrum of environmental parameters.The relative frequencies of these two species were compared with biotic indexes and indexes of environmental integrity in four areas: Oasidi Punte Alberete (RA), Riserva Naturale del Lago di Penne (PE); Tenuta Presidenziale di Castelporziano (RM), Riserva NaturaleOrientata Bosco Pantano di Policoro (MT).Our results show a prevalence of R. lessonae in natural habitats and viceversa.The importance as bioindicators of the pond frogs belonging to the Rana esculenta complex appears to be confirmed, and thanks to theircharacteristics complementary to the macrobenthos community, their relative frequencies should prove useful in evaluating the anthropicpollution in habitats where it is not possible to use the biotic indexes most widely used: river ponds, lakes, marshes, and transitionalhabitats.
KEY WORDS: Rana esculenta complex / bioindicators / hybrid species / heterozigosity / competition

ANDREANI et al. - Le rane verdi bioindicatrici in ambienti fluviali36
INTRODUZIONEL’uso dei bioindicatori, oramai sancito anche nella
normativa nazionale ed internazionale, si basa sullecaratteristiche intrinseche delle specie indicatrici. Tragli organismi scelti come indicatori gli anfibi rivestonoun ruolo di rilievo, rafforzato dall’ampio utilizzo cheessi hanno anche in ambiti di analisi diversi: ecotossi-cologico, biomolecolare, istologico, fisiologico, ecc.
Le caratteristiche che esaltano la capacità bioindica-trice degli anfibi sono diverse: citiamo ad esempio lasensibilità nei confronti di fenomeni di bioaccumulo(BURY, 1988; VITT et al., 1990; DUNSON et al., 1992;WALLAS et al., 1992; COHN, 1994; NCER, 1998) o inrisposta agli impatti antropici responsabili del declinogeneralizzato delle popolazioni (BARINAGA, 1990; WAKE,1991; HALL e HENRY, 1992).
Tra le diverse linee di ricerca numerose sono quelleche hanno considerato specificatamente gli anuri, perevidenziare la sensibilità all’inquinamento da metallipesanti (LEFCORT et al., 1998; LEONTYEVA et al., 1997),da sostanze azotate (OLDHAM et al., 1997) e da pestici-di (SCHUYTEMA et al., 1991, 1993).
In modo particolare, le rane del complesso Ranaesculenta sono state utilizzate come biosensori (KISE-LEVA, 1997) o come indicatori, correlando i cambia-menti nella loro distribuzione geografica con la presen-za di stress antropici (CHUBINISHVILI, 1998a), di speci-fiche sostanze chimiche (MISYURA et al., 1996; TOK-TAMYSOVA e KAIDAULOVA, 1996; CHUBINISHVILI, 1998b)o di isotopi radioattivi (CHERNYOSHOVA e STAROSTIN,1994).
Nel campo della bioindicazione un ruolo di un certointeresse lo rivestono quindi le specie appartenenti alcomplesso Rana esculenta, geneticamente affini traloro ma con evidenti elementi discriminanti (il genotipoeterozigote di R. esculenta ai loci differenziati tra R.lessonae e Rana ridibunda). La specie ibrida R. escu-lenta si origina dall’incrocio tra i due parentali R.ridibunda e R. lessonae ed, essendo autosterile, riescea riprodursi solo sfruttando dal punto di vista sessualela specie parentale con la quale convive. Il particolaremeccanismo che permette la formazione di R. esculen-ta viene definito ibridogenesi (HOTZ et al., 1992.)
Questa caratteristica ha fatto sì che tali specie sianostate oggetto di numerosi studi scientifici di naturamolto diversa, come l’analisi del meccanismo di esclu-sione del genoma parentale sintopico da parte di R.esculenta (GUERRINI et al., 1997), la riduzione delladiversità genomica in aree contaminate da radiazioni(VINOGRADOV e CHUBINISHVILI, 1999) o l’influenza delmaggiore grado di eterosi del genoma ibrido sullacapacità di adattamento (TUNNER e NOPP, 1979).
In particolare, diversi sono i lavori che prendono inconsiderazione la capacità di adattamento delle larve
delle specie parentali R. lessonae e R. ridibunda, edell’ibrido R. esculenta. In questi studi si evidenzia ilfenomeno della competizione asimmetrica che permet-te a R. esculenta di prevalere in presenza di parametriambientali mutevoli, in contrasto con la migliore capa-cità adattativa delle specie parentali in condizioni disviluppo stabili (SEMLITSCH e REYER, 1992; HOTZ et al.,1993; SEMLITSCH, 1993a, 1993b; SEMLITSCH et al.,1993; FIORAMONTI et al., 1997; RIST et al., 1997;SEMLITSCH et al., 1997; HOTZ et al., 1999; PLENET etal., 2000).
La maggiore capacità competitiva mostrata dallespecie ibride in determinate condizioni ambientali, ri-spetto alle specie parentali, è stata ampiamente studiatae viene riassunta dalla teoria del vantaggio eterotico.Questa ipotizza l’esistenza di una stretta correlazionetra tale capacità e l’elevato grado di eterozigosi presen-te nelle specie ibride (BULLINI, 1985). Tale caratteristi-ca genetica fornirebbe un più ampio spettro di adatta-menti fisiologici ad ambienti con parametri ecologicimutevoli. Vari autori, confrontando coppie di specieaffini ma con valori di variabilità genetica differenti,osservano come la maggiore capacità competitiva inambienti con un’ampia oscillazione dei fattori di distur-bo venga proprio influenzata da un alto valore dieterozigosi (UZZEL e DAREWSKY, 1973, 1975; NEVO,1976; NASCETTI e BULLINI, 1980; LAVIE e NEVO, 1986;NEWMAN, 1988; BULLINI e NASCETTI, 1990; MCALPINE,1993).
Sulla base di questa teoria e per identificare i fattoriambientali che influenzano la distribuzione delle speciedel complesso Rana esculenta, nel presente lavoro lapercentuale delle due specie R. lessonae e R. esculentaè stata comparata con le risultanze di indici biotici e dinaturalità ambientale.
La necessità di avere questi parametri di riscontro,pur costruiti su dati di origine differenti, nasce dall’evi-denza che le rane sono selezionate sia dalla qualità delleacque, poiché la mortalità differenziale dovrebbe avve-nire soprattutto nella fase larvale trascorsa interamentein acqua, sia dall’integrità dell’ambiente terrestre, poi-ché gli adulti trascorrono parte della loro vita fuoridall’acqua.
MATERIALI E METODI
Le stazioni di campionamentoLe rilevazioni effettuate nell’ambito di questo lavoro
riguardano alcune località dell’Italia peninsulare (Fig.1) caratterizzate dall’essere diverse dal punto di vistageografico ed ecologico, al fine di saggiare le rispostedelle due specie di rane in condizioni il più possibiledifferenziate.
Sono stati analizzati transetti nelle seguenti aree:

ANDREANI et al. - Le rane verdi bioindicatrici in ambienti fluviali 37
l’Oasi di Punte Alberete (RA) e le aree circostanti; ilterritorio del bacino idrografico del Fiume Tavo e delTorrente Gallero e la Riserva Naturale del Lago diPenne (PE); la Tenuta Presidenziale di Castelporziano(RM) e la Riserva Naturale Orientata Bosco Pantano diPolicoro con gli immediati dintorni (MT).
La distribuzione delle stazioni è stata effettuata inmodo da ottenere dei transetti ecologicamente diffe-renziati all’interno delle singole aree considerate.
I vari siti di indagine differiscono anche per l’estensio-ne dell’area investigata: dalle varie decine di chilometriper quello in Abruzzo, alle poche centinaia di metri perquello in provincia di Ravenna.
Distribuzione delle stazioniL’Oasi di Punte Alberete, sita sulla costa adriatica in
provincia di Ravenna, è costituita da una foresta plani-ziale percorsa da un fitto reticolo di canali oramainaturalizzati ricchi di vegetazione che permette unsensibile abbattimento della sostanza organica. L’areaprotetta è circondata da una zona dove si svolgeattività agricola intensiva; l’oasi e i suoi dintorni sonostati studiati con cinque siti di monitoraggio.
Nei bacini idrografici del Fiume Tavo e del TorrenteGallero e all’interno dell’Oasi del Lago di Penne (pro-
vincia di Pescara) sono state considerate dieci stazionidi campionamento. L’intero bacino idrografico è ca-ratterizzato dalla presenza di uno stress antropico cre-scente da monte a valle. L’inquinamento presente nel-l’area è prevalentemente di tipo agricolo-zootecnico,anche se non mancano insediamenti urbani. In ottostazioni di rilevamento è stato possibile utilizzare tuttigli indici; altre due sono state considerate con gliindicatori non riferiti alle rane verdi.
All’interno della Tenuta Presidenziale di Castelpor-ziano (RM), dichiarata riserva naturale fin dal secondodopoguerra, sono stati scelti quattro siti di studio: duedelle stazioni di campionamento sono poste in pozzeperenni situate nella zona di riserva integrale e inun’area a stress antropico medio (taglio boschi, pa-scolo brado, viabilità interna). Gli altri due siti di studiosono stati individuati in altrettanti corsi d’acqua cheattraversano coltivazioni semi intensive di cereali.
L’area della Riserva Naturale Orientata Bosco Pan-tano di Policoro (MT) è stata studiata attraverso cin-que stazioni di rilevamento. Anche in questo caso lazona a più alta naturalità è caratterizzata da un boscoplaniziale litoraneo percorso da canali artificiali in parterinaturalizzati. L’acqua defluisce all’interno della riser-va entrando da una zona utilizzata per l’agricolturaintensiva dove sono state poste tre stazioni, e nefuoriesce dopo aver attraversato la parte più internadell’area protetta, studiata con due siti di rilevamento.
Il complesso Rana esculentaLe rane verdi presenti in Italia sono rappresentate da
due delle specie che compongono il complesso Ranaesculenta: R. lessonae e R. esculenta, (SANTUCCI et al.,1996; SANTUCCI et al., 2000).
Il metodo analitico che ha permesso il riconosci-mento tra le due specie è stato l’analisi elettroforeticadei sistemi gene-enzima. Tale strumento di indagine,come confermato da PAGANO e JOLY (1998), è l’unicoche consenta un’attribuzione certa degli individui al-l’una o all’altra specie.
Nel complesso sono stati analizzati circa 1100 indi-vidui, la cattura è stata effettuata a mano o con l’ausiliodi un retino principalmente nelle ore notturne; gli indi-vidui catturati sono stati rilasciati dopo avere prelevatoda ognuno di essi, previa anestesia, porzioni di falangedel secondo dito dell’arto posteriore. I campioni sonostati conservati a - 80 °C sino al momento delle analisielettroforetiche.
Il riconoscimento degli individui di rana verde èstato condotto attraverso lo studio dei loci genici dia-gnostici, mediante elettroforesi dei sistemi gene-enzi-ma su gel d’amido.
Per la nomenclatura dei loci e degli alleli sono statiadottati i seguenti criteri:
Fig. 1. Distribuzione geografica delle aree investigate (1 = Oasi diPunte Alberete; 2 = Bacino idrografico del Lago di Penne; 3 =Tenuta Presidenziale di Castel Porziano; 4 = Oasi del BoscoPantano di Policoro).

ANDREANI et al. - Le rane verdi bioindicatrici in ambienti fluviali38
• gli isozimi (enzimi codificati da loci diversi, ma chepresentano stessa funzione enzimatica) sono statinumerati in ordine di mobilità decrescente a partire daquello più anodico (per esempio Ldh-1, Ldh-2, ecc.);
• gli allozimi (forme enzimatiche prodotte dagli alleli diciascun locus) sono stati invece indicati con unnumero che esprime la loro mobilità in mm (peresempio Ldh-1100, Ldh-1106, ecc.). Il valore 100 èstato attribuito all’allele più frequente nella popolazio-ne di R. lessonae di Morozzo, Piemonte (regionetipica di R. lessonae), presa come riferimento (alleli> 100 = mobilità maggiore, alleli < 100 = mobilitàminore) (SANTUCCI, 1993).I loci analizzati geneticamente sono i seguenti: G3pdh,
Ldh-1, 6Pgdh, Pep-2, Pgi-2. Tali loci (UZZELL e BER-GER 1975) sono differenziati nelle specie parentali di R.esculenta, R. lessonae e R. ridibunda e sono stati usatisu vasta scala per il riconoscimento di queste trespecie (BERGER, 1983; SEMLITSCH et al., 1996, SEMLIT-SCH et al., 1997, PAGANO et al., 1997).
Sulla base del genotipo ai loci diagnostici tutti gliindividui saggiati sono stati attribuiti a R. lessonae o aR. esculenta.
La qualità delle acque è stata definita attraverso lostudio dei macroinvertebrati delle acque dolci, analiz-zati secondo le disposizioni date dalle metodiche diapplicazione dell’I.B.E. (GHETTI, 1997) e del BMWP’(ALBA TERCEDOR e SANCHEZ-ORTEGA, 1988).
Inoltre è stato utilizzato l’Indice di Funzionalità Flu-viale (I.F.F.) (SILIGARDI et al., 2000) per verificare seun disturbo ambientale generalizzato possa essere re-sponsabile della variazione della frequenza relativa dellerane verdi. L’indice I.F.F. è stato applicato e calcolatoattraverso l’apposita scheda di rilevamento (SILIGARDIet al., 2000).
RISULTATII risultati che illustrano le frequenze relative delle
due specie di rane verdi, e i valori dei diversi indicisono riassunti nella tabella I. Le stazioni di monitorag-gio sono ordinate come di seguito descritto: nei tran-setti dell’Oasi di Punte Alberete, della Tenuta Presiden-ziale di Castel Porziano e dell’Oasi del Bosco Pantanodi Policoro, da quelle caratterizzate da un sensibilestress antropico a quelle poste nelle zone più internedelle diverse aree protette; per quanto riguarda il tran-setto del bacino idrografico del Lago di Penne, questesono ordinate da monte a valle. Il valore dell’I.F.F. èstato mediato tra le due sponde.
Tutti i dati ottenuti riguardanti la frequenza delle duespecie nei popolamenti di rana verde sono stati sotto-posti al test del χ2 che ne ha confermato la significati-vità statistica (p < 0,005).
Il livello di eterozigosi media osservata in R. escu-
lenta é notevolmente superiore ai valori misurati in R.lessonae (Ho rispettivamente 0,45 e 0,09).
L’oasi di Punte AlbereteLa frequenza relativa delle specie appartenenti al
complesso Rana esculenta si differenzia sensibilmentenelle tre stazioni all’esterno dell’oasi, rispetto a quelleposte all’interno. Nella zona agricola la specie ibridainfatti è sempre preponderante con una presenza me-dia del 70%; la comunità macrobentonica (IV CQ) el’analisi dei parametri che influenzano la funzionalitàdei corsi d’acqua, presentano valori relativamente bas-si (IV/V LF).
All’interno dell’oasi dal lato Sud (PFA-2), la percen-tuale della specie sensibile R. lessonae è del 90%, lacomunità macrobentonica mostra una lieve alterazione(II CQ) con l’I.F.F. tra il I ed il II LF. Nella zona piùinterna (PFA-1) della riserva la comunità macrobento-nica presenta una struttura relativamente integra (I/IICQ) mentre l’I.F.F. e la percentuale di R. lessonae(86%) non mostrano significative variazioni.
Il bacino idrografico del Lago di Penne(fiume Tavo e torrente Gallero)
L’analisi del bacino idrografico del Fiume Tavo èstata effettuata con quattro stazioni di campionamen-to, determinando sia i valori degli indici biotici che lefrequenze relative di R. lessonae e R. esculenta; inoltresono state aggiunte altre due stazioni monitorate esclu-sivamente attraverso la comunità del macrobenthos el’I.F.F.
Nella prima stazione del Fiume Tavo R. esculenta ècompletamente assente, mentre I.B.E. e BMWP’ mo-strano una I CQ; l’I.F.F. presenta un II LF e valorepari a 212.
Nella stazione TVA l’I.B.E. ed il BMWP’ sono aduna II CQ, anche l’I.F.F. presenta un II LF. Nelsuccessivo sito, dove sono presenti le rane verdi (TVO),la specie ibrida più resistente è al 57%. Gli altri indicimostrano valori che vanno da una III CQ dell’I.B.E.(valore 6), ad una II CQ per il BMWP’ ad un II LF perl’I.F.F.
Scendendo l’ungo l’asta fluviale, in prossimità di uncentro abitato (TFA), gli indicatori mostrano l’I.B.E.in III CQ, il BMWP’ in II CQ, e l’I.F.F. in II/III LF, lafrequenza relativa di R. esculenta arriva all’84%.
Nella stazione successiva, dove il corso d’acquariemerge dopo aver percorso diversi chilometri in su-balveo (TMA), la percentuale di R. lessonae sale al31%, le CQ e il LF sono rispettivamente II, I e II/III.
Nella stazione a chiusura di bacino (TCA) investiga-ta con gli indici di riferimento, l’I.B.E. presenta una IVCQ, il BMWP’ una II CQ e l’I.F.F. un III Livello diFunzionalità.
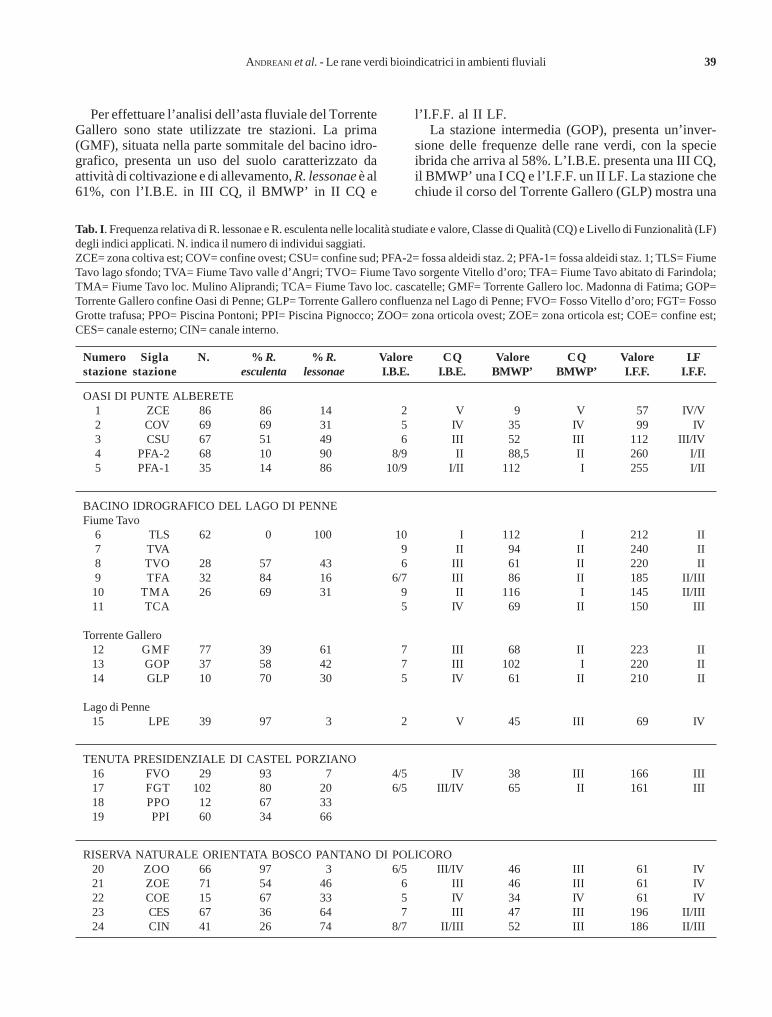
ANDREANI et al. - Le rane verdi bioindicatrici in ambienti fluviali 39
Per effettuare l’analisi dell’asta fluviale del TorrenteGallero sono state utilizzate tre stazioni. La prima(GMF), situata nella parte sommitale del bacino idro-grafico, presenta un uso del suolo caratterizzato daattività di coltivazione e di allevamento, R. lessonae è al61%, con l’I.B.E. in III CQ, il BMWP’ in II CQ e
l’I.F.F. al II LF.La stazione intermedia (GOP), presenta un’inver-
sione delle frequenze delle rane verdi, con la specieibrida che arriva al 58%. L’I.B.E. presenta una III CQ,il BMWP’ una I CQ e l’I.F.F. un II LF. La stazione chechiude il corso del Torrente Gallero (GLP) mostra una
Tab. I. Frequenza relativa di R. lessonae e R. esculenta nelle località studiate e valore, Classe di Qualità (CQ) e Livello di Funzionalità (LF)degli indici applicati. N. indica il numero di individui saggiati.ZCE= zona coltiva est; COV= confine ovest; CSU= confine sud; PFA-2= fossa aldeidi staz. 2; PFA-1= fossa aldeidi staz. 1; TLS= FiumeTavo lago sfondo; TVA= Fiume Tavo valle d’Angri; TVO= Fiume Tavo sorgente Vitello d’oro; TFA= Fiume Tavo abitato di Farindola;TMA= Fiume Tavo loc. Mulino Aliprandi; TCA= Fiume Tavo loc. cascatelle; GMF= Torrente Gallero loc. Madonna di Fatima; GOP=Torrente Gallero confine Oasi di Penne; GLP= Torrente Gallero confluenza nel Lago di Penne; FVO= Fosso Vitello d’oro; FGT= FossoGrotte trafusa; PPO= Piscina Pontoni; PPI= Piscina Pignocco; ZOO= zona orticola ovest; ZOE= zona orticola est; COE= confine est;CES= canale esterno; CIN= canale interno.
Numero Sigla N. % R. % R. Valore CQ Valore CQ Valore LFstazione stazione esculenta lessonae I.B.E. I.B.E. BMWP’ BMWP’ I.F.F. I.F.F.
OASI DI PUNTE ALBERETE1 ZCE 86 86 14 2 V 9 V 57 IV/V2 COV 69 69 31 5 IV 35 IV 99 IV3 CSU 67 51 49 6 III 52 III 112 III/IV4 PFA-2 68 10 90 8/9 II 88,5 II 260 I/II5 PFA-1 35 14 86 10/9 I/II 112 I 255 I/II
BACINO IDROGRAFICO DEL LAGO DI PENNEFiume Tavo
6 TLS 62 0 100 10 I 112 I 212 II7 TVA 9 II 94 II 240 II8 TVO 28 57 43 6 III 61 II 220 II9 TFA 32 84 16 6/7 III 86 II 185 II/III10 TMA 26 69 31 9 II 116 I 145 II/III11 TCA 5 IV 69 II 150 III
Torrente Gallero12 GMF 77 39 61 7 III 68 II 223 II13 GOP 37 58 42 7 III 102 I 220 II14 GLP 10 70 30 5 IV 61 II 210 II
Lago di Penne15 LPE 39 97 3 2 V 45 III 69 IV
TENUTA PRESIDENZIALE DI CASTEL PORZIANO16 FVO 29 93 7 4/5 IV 38 III 166 III17 FGT 102 80 20 6/5 III/IV 65 II 161 III18 PPO 12 67 3319 PPI 60 34 66
RISERVA NATURALE ORIENTATA BOSCO PANTANO DI POLICORO20 ZOO 66 97 3 6/5 III/IV 46 III 61 IV21 ZOE 71 54 46 6 III 46 III 61 IV22 COE 15 67 33 5 IV 34 IV 61 IV23 CES 67 36 64 7 III 47 III 196 II/III24 CIN 41 26 74 8/7 II/III 52 III 186 II/III

ANDREANI et al. - Le rane verdi bioindicatrici in ambienti fluviali40
frequenza di R. esculenta pari al 70%. Tra gli altriindicatori l’I.B.E. arriva ad una IV CQ, mentre ilBMWP’ ed l’I.F.F. presentano una II CQ ed un II LF.
Per quanto concerne l’analisi del Lago di Penne(LPE), poiché l’applicazione degli indici di confrontonon è possibile in ambienti lentici, si è ovviato effet-tuando i campionamenti nel rio che si origina da unbacino artificiale alimentato dall’acqua del lago. Inquesto piccolo corso d’acqua si è potuta stabilire unacomunità bentonica stabile viste le caratteristiche diregime idrologico costante.
La percentuale di R. esculenta, campionata nel me-desimo rio, raggiunge valori pari al 97%. Gli altri indicipresentano i seguenti valori: I.B.E. V CQ, BMWP’ IIICQ e I.F.F. IV LF.
La tenuta presidenziale di Castel PorzianoNella Tenuta Presidenziale di Castel Porziano il pri-
mo sito di campionamento è posto su un piccolo fossoche corre in una zona adibita a coltivazione di cereali.In questa stazione (FVO) R. esculenta è dominante(93%), con i seguenti valori per gli indici di confronto:I.B.E. valore 4/5, IV CQ; BMWP’ valore 38, III CQ;I.F.F. valore 166, III LF. Anche nel secondo sito(FGT) R. lessonae è presente in basse percentuali(20%); il valore di I.B.E. è 6/5, (III/IV CQ), valore 65e II CQ per il BMWP’; valore 161 e III LF per l’I.F.F..
Procedendo verso la parte più interna della riserva,sono localizzati due siti di rilevamento in piccoli spec-chi d’acqua perenni e quindi monitorabili esclusiva-mente con le rane verdi. La terza stazione (PPO), sitrova in un’area sensibilmente influenzata dall’uomo;la frequenza di R. lessonae si attesta al 33%. Nella zonaa protezione integrale (PPI), la frequenza relativa di R.lessonae diviene maggioritaria ma non dominante (66%).
Riserva naturale orientataBosco Pantano di Policoro
Le aree esterne alla zona protetta presentano fre-quenze di R. esculenta sempre superiori a quelle dellaspecie più sensibile con frequenze che vanno dal 97%(ZOO), al 67% (ZOE) al 54% (COE) i valori medi degliindici di riferimento sono i seguenti: I.B.E. III/IV CQ;BMWP’ III/IV CQ; I.F.F. IV LF.
All’interno dell’area protetta le frequenze si inverto-no e diventa preponderante R. lessonae: 64% nellastazione CES; 74% in quella situata nella zona piùinterna della riserva (CIN). I valori degli altri indicimostrano rispettivamente una III CQ per quelli basatisulla comunità macrobentonica ed un II/III LF perl’I.F.F. nella prima stazione; il valore dell’I.B.E. (8/7,II/III CQ) nella stazione più interna; presenta un lieverecupero, il BMWP’ e I.F.F. confermano i valori dellastazione precedente.
DISCUSSIONE E CONCLUSIONILe evidenze sperimentali fin qui raccolte conferma-
no che alla base delle capacità bioindicatrici delle raneverdi esiste una mortalità differenziale tra le due specieappartenenti al complesso Rana esculenta. Infatti R.lessonae, tende a prevalere in ambienti non disturbatifino ad escludere del tutto R. esculenta. Per contro, inambienti molto stressati, la specie ibrida R. esculentapresenta frequenze crescenti, mentre R. lessonae siriduce notevolmente.
Le capacità adattative di R. esculenta sono presumi-bilmente attribuibili all’elevato grado di eterozigosi delsuo genoma, che ne aumenta la fitness potenzialeampliando gli intervalli di variazione dei parametri am-bientali che questa specie può tollerare.
Che la diversa distribuzione non sia influenzata dafattori di tipo strettamente ecologico (altitudine, mor-fologia dell’alveo, velocità della corrente, ecc.) vieneconfermato dal fatto che stazioni di rilevamento checondividono le stesse caratteristiche ambientali mo-strano frequenze differenziate a causa proprio dei fat-tori di alterazione esterna.
Questa distribuzione viene ben evidenziata nel casodi stazioni di campionamento che presentano frequen-ze relative molto differenziate anche se con parametriecologici paragonabili, come nel caso delle stazione deltransetto di Punte Alberete o quello della Riserva diPolicoro. I risultati ottenuti in questi transetti rivestonoun particolare interesse poiché la drastica inversionedelle frequenze delle due specie avviene nel brevespazio di alcune centinaia di metri, rafforzando l’ipote-si che responsabili della distribuzione delle rane verdisiano proprio le variazioni della qualità dell’ambiente.
Ulteriori indicazioni si hanno dallo studio di altripopolamenti del centro Italia, che mostrano come lafrequenza relativa di R. esculenta decresca con ildiminuire dell’altitudine se questo corrisponde anchead una diminuzione delle attività antropiche (bacinoidrografico del Lago di Mezzano-Fosso Olpeta), oppu-re presentino frequenze simili pur trovandosi a quotemolto diverse (bacino idrografico del Lago di Bolsena-Fiume Marta) (ANDREANI et al 1997; ANDREANI 1999a;1999b).
La competizione differenziale tra una specie di origi-ne ibrida e le parentali evidenziata in questo lavoro èanche supportata da numerose evidenze sperimentali(UZZEL e DAREVSKY, 1975; NEVO, 1987; QUATTRO etal., 1992). La possibilità delle specie di origine ibrida diadattarsi meglio delle parentali agli ambienti variabili èstata recentemente confermata nei pesci del generePoeciliopsis, confrontando l’ibridogeno P. monacha-lucida con le specie parentali P. monacha e P. lucida(VRIJENHOEK e PFEILER, 1997). L’analisi di questo siste-ma a tre specie conferma come al successo delle

ANDREANI et al. - Le rane verdi bioindicatrici in ambienti fluviali 41
forme ibridogenetiche concorrano sia l’eterosi che lavariabilità clonale.
Queste caratteristiche, che consentono di ampliarel’intervallo di condizioni tollerabili dagli ibridogeni (me-diante l’elevato grado di eterozigosi) e di diversificarele nicchie ottimali dei vari emicloni (mediante la diver-sità clonale) (BULGER e SCHULTZ, 1979), sono stateevidenziate anche nei popolamenti italiani di rane verdie possono concorrere sensibilmente alla variazionedelle frequenze relative nei popolamenti naturali delledue specie di rane (SANTUCCI, 1993).
L’analisi dei fattori determinanti la distribuzione dif-ferenziale delle due specie facenti parte del complessoRana esculenta, formulata nel presente lavoro, vienerafforzata anche da precedenti studi che mostranocome le attività umane in genere, o più specificatamen-te quelle agricole, tendano a selezionare le popolazionidi anfibi (SEMLITSCH e REYER, 1992; BAUER DIAL eDIAL, 1995; NORRIS e QUEVEDO, 1996; FIORAMONTI etal., 1997; OLDHAM et al., 1997; CHUBINISVILI, 1998a,1998b; LEFCORT et al., 1998).
A questo scopo è importante notare come sia rile-vante la somiglianza nelle risposte delle rane verdi conla comunità del macrobentos investigata attraversol’I.B.E. Questa similarità riveste una certa importanzapoiché evidenzia come l’insieme delle caratteristichequalitative e quantitative delle sostanze disciolte nel-l’acqua e bioaccumulabili influenzino le risposte fisio-logiche ed adattative delle due specie di anfibi.
Per contro la concordanza alle risposte dell’indiceI.F.F., che considera sia l’alveo bagnato che le carat-teristiche dell’ambiente di contorno ad una determinatasezione fluviale, indica come tali anfibi anuri possanoin qualche modo risentire dell’insieme dei fattori cheinfluenzano negativamente la funzionalità del corsod’acqua. È inoltre di rilievo notare come non ci siano lestesse relazioni con l’indice BMWP’ che d’altronde,come osservato in studi precedenti, sembra avererisposte meno concordanti anche rispetto all’I.B.E. eagli indicatori chimico-fisici (ANDREANI, 1999a).
Le corrispondenze evidenziate si possono spiegarepensando proprio alla natura anfibia di questi organi-smi, che trascorrono parte del ciclo vitale in acqua(tutta la fase larvale e quella riproduttiva) e parte sulterreno, specificatamente nella fascia riparia (da adul-ti). In base ai dati raccolti la distribuzione di R. lesso-nae e R. esculenta appare quindi influenzata in mododeterminante dai parametri ecologici che generalmentevengono utilizzati per la valutazione dello stress diorigine antropica.
I dati ottenuti su un transetto studiato nel centroItalia (Fiume Marta – VT), confermano i risultati deri-vanti dal presente lavoro. Attraverso l’analisi dei para-metri chimico-fisici, microbiologici e della composi-
zione della comunità del macrobentos, infatti, si èevidenziato come la presenza di R. esculenta sia stati-sticamente correlata con le alte quantità di nutrientidisciolti, con le cariche batteriche di coliformi e diclostridi e con la presenza di quelle Unità Sistematichedi macroinvertebrati che notoriamente sono ritenutemaggiormente resistenti all’inquinamento (oligocheti,chironomidi, ecc.). Inoltre si è anche potuta verificareuna relativa sensibilità del sistema che, in presenza dilievi incrementi dei parametri di confronto, vede unamarcata crescita di R. esculenta rispetto a R. lessonae(ANDREANI, 1999a).
Inoltre, analisi ecotossicologiche, come la determi-nazione della LC50
96h per il CuSO4, sembrano confer-mare una maggiore resistenza di R. esculenta rispetto aR. lessonae, almeno per quanto concerne l’esposizionea questo inquinante (SANTUCCI, 1993).
A scopo riepilogativo, si riportano i risultati dellediverse stazioni di campionamento ordinati in base allapercentuale crescente della specie ibrida (Tab. II).
Da questa tabella si rileva una discreta correlazionetra gli intervalli di variazione delle frequenze relativedelle due specie e i valori di I.B.E. Naturalmente esisto-no delle discrepanze che, in alcuni casi andranno ulte-
Tab. II. Intervallo di variazione della frequenza relativa di R.esculenta –ordinata in modo crescente– e del valore dell’indiceI.B.E. (Con l’asterisco sono evidenziate le stazioni nelle quali lerisposte dei due indici si discostano maggiormente dall’intervallodi riferimento). Per il significato delle sigle delle stazioni si vedala Tab. I.
Intervallo % Intervallo del % di Valore SiglaR. esculenta valore I.B.E. R. esculenta I.B.E. stazione
1 10 TLS0 - 20 % 9/8-10 10 9/8 PFA-2
14 10/9 PFA-126 8/7 CIN
20 - 40 % 7 - 7/8 36 7 CES39 7 GMF51 6 CSU54 6 FRA
40 - 60 % 6 57 6 TVO58 7 * GOP67 5 COE69 5 COV
60 - 80 % 5 - 5/6 69 9 * TMA70 5 GLP80 5/6 FGT84 6/7 * TFA86 2 FRU
80 - 100 % 2 - 5/6 93 2 FVO97 2 LPE97 5/6 * AGR

ANDREANI et al. - Le rane verdi bioindicatrici in ambienti fluviali42
riormente indagate (stazioni TFA e GOP), mentre inaltri possono ben evidenziare come tali anfibi sianoanche sensibili ad uno stress generalizzato come mo-strato dai Livelli di Funzionalità dell’I.F.F. nelle stazio-ni TMA e AGR. Inoltre è interessante notare come, neicasi di discrepanza, sia sempre la frequenza deglianfibi a mostrare una qualità dell’ambiente peggiorerispetto a quella evidenziata dall’I.B.E. Questo datopotrebbe essere a possibile conferma della sensibilitàdel bioindicatore rana verde già evidenziata in prece-denti studi (ANDREANI, 1999a).
A conforto dell’ipotesi formulata nel presente lavo-ro per spiegare la distribuzione differenziale delle duespecie di rane verdi, un recente studio (NEGOVETIC etal., 2001) evidenzia come esistano specifiche rispostealla temperatura ambientale da parte di R. esculenta eR. lessonae. In questo studio viene dimostrato che, aparità di condizioni di sviluppo, le larve di R. esculentacompetono meglio di quelle di R. lessonae quando letemperature sono più basse e viceversa.
Questo dato potrebbe confermare come le più altefrequenze di R. esculenta da noi rilevate nelle stazioniposte a quote più basse (dove la temperatura è maggio-re), siano quindi proprio da imputare al grado di stress,che generalmente tende ad aumentare nelle zone dipianura maggiormente antropizzate.
L’associazione tra le frequenze delle due specie dirane e specifici livelli di inquinamento è stata ampia-mente studiata ma, al fine di costruire un vero eproprio indice biotico è necessario correlare, con ulte-riori studi, i valori delle frequenze relative con le Classidi Qualità ambientali. La possibilità di utilizzare le ri-
sposte del complesso Rana esculenta per piani dimonitoraggio ambientale a larga scala appare comun-que non distante dagli altri modelli costruiti su specie dianfibi (NCER, 1998). Inoltre la possibilità di studiarequesti bioindicatori in ambienti lentici, già comprovatain precedenti studi (ANDREANI et al., 1997; ANDREANI,1999a, 1999b; SCIALANCA et al., 1999) li rende parti-colarmente adatti ad essere affiancati ai sistemi piùtradizionali, potendo produrre dati alternativi e comple-mentari a questi, anche ribadendo che le rane in quantovertebrati sono relativamente sensibili ai preoccupantifenomeni di bioaccumulo.
Questo lavoro rappresenta un primo contributo pervalidare la capacità bioindicatrice delle rane verdi. Ben-ché i dati analizzati rappresentino diverse condizionigeografiche ed ambientali, sarebbe comunque auspi-cabile aumentare il numero dei transetti da studiare,per verificare le risposte del complesso Rana esculen-ta in ambienti diversi. Inoltre incrementare i dati diconfronto permetterebbe di ampliare la base statistica,per associare le risposte di questi anfibi ai diversistress ambientali e costruire una scala delle frequenzerelative rispetto all’entità ed alla tipologia degli impattiantropici.
BIBLIOGRAFIA
ALBA TERCEDOR J., SANCHEZ-ORTEGA A., 1988. Un metodo rapidoy simple para evaluar le calidad biologica de las aguas corrientesbasado en el de Hellawell (1978). Limnetica, 4: 51-56.
ANDREANI P., MOTAWI A., NASCETTI G., 1997. Valutazione dellaqualità delle acque del bacino idrografico del fiume Marta (AltoLazio): applicazione dell’Indice Biotico Esteso e analisi dellafrequenza relativa di rane verdi del complesso Rana esculenta.In: Atti Conv. Scient. “VIII Congresso Nazionale della SocietàItaliana di Ecologia”, Parma, 10-12 settembre 1997.
ANDREANI P., 1999a. Le rane verdi del complesso Rana esculentacome indicatori di disturbo ambientale: analisi delle frequenzerelative e confronto con indici biotici e parametri ecologici. Tesidi Dottorato in Scienze Ambientali (XII ciclo), Università degliStudi della Tuscia, Viterbo.
ANDREANI P., 1999b. Rana lessonae e Rana esculenta: un test peril valore adattivo dell’eterozigosi. In: Atti Conv. Scient. “IXCongresso Nazionale della Società Italiana di Ecologia”,Lecce, 14-17 settembre 1999.
BARINAGA M., 1990. Where have all the frogs is gone? Sciences,247: 1033-1034.
BAUER DIAL C.A., DIal N.A., 1995. Lethal effects of the consump-tion of fields level of paraquat-contaminated plants of frogtadpoles. Bulletin of Enviromental Contamination and Toxico-logy, 55, 6: 870-877.
BERGER L., 1983. Western Paleartic water frogs (Amphibia,Ranidae): Systematic, genetic and population composition.Experientia, 39: 127-130.
BULGER A.J., SChultz R.J., 1979. Heterosis and interclonal varia-tion in thermal tolerance in unisexual fishes. Evolution, 33: 848-859.
BULLINI L., 1985. Speciation by hybridization in animals. Bol.Zool., 52: 121-137.
BULLINI L., NASCETTI G. 1990. Speciation by hybridization inphasmids and other insects. Can. J. Zool., 68: 1747-1760.
BURY R.B., 1988. Habitat relationship and ecological importanceof amphibians and reptiles in streamside management. In:
RINGRAZIAMENTIGli autori desiderano ringraziare Laura Mancini, per la preziosacollaborazione alla revisione del testo, Maurizio Battegazzore,per le preziose informazioni sugli indici biotici e Gaetano Aloise,Fabrizio Baldini, Osvaldo Locasciulli, il personale dell’Oasi delLago di Penne (WWF) e della Tenuta Presidenziale di CastelPorziano per la collaborazione alla raccolta dei campioni.

ANDREANI et al. - Le rane verdi bioindicatrici in ambienti fluviali 43
Raedeke K.J., (ed.). Riparian wildlife and forestry interactions.Seattle, University of Whashington Press, 61-76.
CHERNYOSHOVA E.V., STAROSTIN V.I., 1994. The peripheral bloodof frogs in the genus Rana as a test system for assessingenvironmental pollution. Izv. Akad. Nauk. Ser. Biol., 4: 656-660.
CHON J.P., 1994. Salamanders slip-sliding away or too surreptiousin count? Biosciences.44: 219-223.
CHUBINISHVILI A.T., 1998a. The status of the natural populationof the Rana esculenta complex in response to anthropogenicinfluences: a morfological approach. In: Pensoft Publishers(ed.). Advance in amphibian research in the former SovietUnion. Sofia, 2: 45-70.
CHUBINISHVILI A.T., 1998b. Development in omeostasis in popu-lation of the marsh Frog (Rana ridibunda Pall.) living under thecondition of chemical pollution in the middle Volga region.Russian J. Ecol., 29, 1: 63-65.
DUNSON W.A., WYMAN R.L., CORBETT E.S., 1992. A symposiumon amphibian declines and habitat acidification. J. Herpetol, 26:349-342.
FIORAMONTI E., SEMLITSCH R.D., REYER H.U., FENT K., 1997.Effects of triphenyltin and pH on the growth and developmentof R. lessonae e R. esculenta tadpoles. Environmental Toxico-logy and Chemistry, 16, 9: 1940-1947.
GHETTI P.F., 1997. Manuale di applicazione. Indice Biotico Esteso(I.B.E.). I macroinvertebrati nell’analisi dei corsi d’acqua. Ed.Provincia autonoma di Trento, Agenzia Provinciale per laprotezione dell’ambiente.
GUERRINI F., BUCCI S., RAGGHIANTI M., MANCINI G., HOTZ H.,UZZELL T., BERGER L., 1997: Genomes of two water frog speciesresist germ line exclusion in interspecies hybrids. J. Exp. Zool.,279, 2: 163-176.
HALL R.J., HENRY P.F.P., 1992. Review: assessing effects ofpesticides on amphibian and reptiles: status and needs. J.Herpetol, 2: 65-71.
HOTZ H., BEERLI P., SPOLSKY C., 1992. Mitodrial DNA revealsformation of nonhybrid frogs by natural mating bet hemiclonalhybrids. Molecular Biology Evolotuion, 9, 4: 610-620.
HOTZ H., SEMLITSCH R.D., SCHMIEDEHAUSEN S., BEERLI P., GUEX
G.D., 1993. Differential performance of LDH-B genotypes inwater frogs tadpoles. In: Catzeflis, Gautier (ed). Evolution 93.Fourth Congress of the European Society for EvolutionaryBiology. Montpellier, August, 182: 22-28.
HOTZ H., SEMLITSCH R.D., GUTMANN E., GUEX G.D., BEERLI P.,1999. Spontaneus heterosis in larval life-history traits ofhemiclonal frog hybrids. Proceeding of National Academy ofSciences U.S.A., 96: 2171-2176.
KISELEVA E.I., 1997. Aspect of the chemical ecology and che-mosensory guided behavioral reaction in anuran tadpole. In:Pensoft Publishers (ed.). Advance in amphibian research in theformer Soviet Union. Sofia, 2: 95-101.
LAVIE B., NEVO E., 1986. Genetic diversity of marine gastropods:contrasting strategies of Cerithium rupestre and C. scabridumin the Mediterranean Sea. Mar. Ecol. Prog. Ser. 28: 99-103.
LEFCORT H., MEGUIRE R.A., WILSON L.H., ETTINGER W.F., 1998.Heavy metals alter the survival, growth methamorphosis andantipredatory behaviour of Columbia spotted frog (Ranaluteiventris) tadpoles. Arch. Environ. Contam. Toxicol. 35, 3:
447-456.LEONTYEVA O.A., SEMENOV D.V., ESENIN A.V., 1997. Brown frogs
as bioindicator of haevy metal contamination: prelminaryresults on the common frog (Rana temporaria) in MoscowProvince. In: Pensoft Publishers (ed.). Advance in amphibianresearch in the former Soviet Union. Sofia, 2: 125-131.
MCALPINE S.,1993. Genetic Heterozygosity and reproductivesuccess in the green treefrog, Hila cinerea. Heredity, 70: 553-558.
MISYURA A.N., GASSO V.Y., SUKHANOVA V.N., 1996. Comparativeecology and biochemistry of amphibian and reptiles of pollu-ted habitats. In: Pensoft Publishers (ed.). Advance in amphi-bian research in the former Soviet Union. Sofia, 1: 111-116.
NASCETTI G., BULLINI L., 1980. Differenziamento genetico especiazione in fasmidi dei generi Bacillus e Clonopsis (Cheleu-toptera, Bacillidae). In: Atti Con. Scient.. “XII Congr. Naz. ital.Ent.” (Roma, 1980), 215-223.
NCER (NACIONAL CENTER FOR ENVIRONMENTAL RESEARCH) ResearchProject, 1998. Environmental factors that influence amphibiancommunity structure and healt as indicators of ecosystems.Investigator: Val Beasley, Carl Richards, Rebecca Cole, RogerBrannian, Lucinda Johnson, Pat Schoff, Jack Cochran, Chri-stopher Phillips, and Martin Ouellet. Institution: Universityof Illinois, University of Minnesota-Duluth, United StatesGeological Survey; Illinois Natural History Survey; and Mc-Gill University. EPA Project Officer: Amanda Senft. ProjectPeriod: June 1, 1998 - May 31, 2001. Research Category:Ecosystem Indicators.
NEGOVETIC S., ANHOLT B., SEMLITSCH R.D., REYER H.U., 2001.Specific response of sexual and hybridogenetic europeanwaterfrog tadpoles to temperature. Ecology, 82, 3: 766-774.
NEVO E., 1976. Adaptive strategies of genetic systems in constantand varing environments. In: Karlin S., Nevo E. (ed.). Popula-tion genetics and ecology. Academic Press, New York, 141-158.
NEWMAN R.A., 1988. Genetic variation for larval anuran (Sca-phiopus couchii) development time in uncertain environment.Evolution, 42, 4: 763-773.
NORRIS B., QUEVEDO L., 1996. Adverse effects of polluted conti-nental water bodies in Chile on frog adrenergic synapse.Bulletin of Enviromental Contamination and Toxicology, 57, 4:640-647.
OLDHAM R.S., LATHAM D.M., HILTON-BROWN D., TOWNS M.,COOKE A.S., BURN A., 1997. The effect of ammonium nitratefertiliser on frog (Rana temporaria) survival. Agriculture,Ecosystems and Environment, 61: 69-74.
PAGANO A., JOLY P., HOTZ H., 1997. Taxon competition andgenetic variation of water frogs in the Mid-Rhone floodplain.Ecology/Ecologie, 320: 759-766.
PAGANO A., JOLY P., 1998. Limits of the morphometric methodfor taxonomic field identification of water frogs. Alytes, 16:3-4.
PLENET S., HERVANT F., JOLY P., 2000. Ecology of the hybridoge-netic Rana esculenta complex: differential oxigen requirementsof tadpoles. Evolutionary Ecology, 14: 13-23.
QUATTRO J.M., AVISE J.C., VRIJENHOEK, R.G., 1992. An ancientclonal linage in the fish genus Poeciliopsis (Atheriniformes:Poeciliidae). Proceeding of National Academy of SciencesU.S.A., 89: 348-352.

ANDREANI et al. - Le rane verdi bioindicatrici in ambienti fluviali44
RIST L., SEMLITSCH R.D., HOTZ H., REYER H.U., 1997. Feedingbeehaviour, food consumption, and growth efficiency of hemi-clonal and parental tadpoles of the Rana esculenta complex.Funcional Ecology, 11: 735-742.
SANTUCCI F., 1993. Ricerche di genetica ecologica su una speciedi origine ibrida: Rana esculenta. Tesi di Dottorato, Università“La Sapienza” di Roma.
SANTUCCI F., NASCETTI G., BULLINI L., 1996. Hybrid zonesbetween two genetically differentiated forms of the pond frogRana lessonae in southern Italy. Journal of EvolutionaryBiology, 9: 429-450.
SANTUCCI F., ANDREANI P., NASCETTI G., BULLINI L., 2000. Geneticidentification and geographic distribution of the water frogsRana lessonae and Rana esculenta in Italy, Sicily and Corsica(amphibia, anura). Mus. Reg. Sci. Nat.. Torino, 359-367.
SCHUYTEMA G.S., NEBEKER A.V., GRIFFIS W.L., WILSON K.L., 1991.Teratogenesis, toxicity, and bioconcentration in frogs exposedto dieldrin. Arch. Environ. Contam. Toxicol., 21, 3: 332-350.
SCHUYTEMA G.S., NEBEKER A.V., PETERSON J.A., GRIFFIS W.L., 1993.Effects of pentaclhorophenol – contaminated food organismson toxicity and bioaccumulation in the frog Xenopus leavis.Arch. Environ. Contam. Toxicol., 24, 3: 359-364.
SCIALANCA F., ANDREANI P., MUCCIOLO L., NASCETTI G., 1999.Valutazione della qualità delle acque litorali del Lago di Vico(Centro Italia.): analisi chimica, studio della comunità macro-benthonica e analisi delle rane verdi del complesso Ranaesculenta. In: Atti Conv. Scient. “IX Congresso Nazionale dellaSocietà Italiana di Ecologia”, Lecce, 14-17 settembre 1999.
SEMLITSCH R., REYER H.U., 1992. Performance of tadpoles from thehybridogenetic Rana esculenta complex: interaction with ponddrying and interspecific competition. Evolution, 46: 665-676.
SEMLITSCH R.D., 1993a. Asymmetric competition in mixed popu-lation of tadpoles of the hybridogenetic Rana esculenta com-plex. Evolution 47: 510-519.
SEMLITSCH R.D., 1993b. Effects of different predators on thesurvival and development of tadpoles from hybridogeneticRana esculenta complex. Oikos, 67: 4-46.
SEMLITSCH R.D., SCHMIEDEHAUSEN S., HOTZ H., BEERLI P., GUEX
G.D., 1993. Differences in larval performance among coexi-sting hemiclones of hybridogenetic water frogs are of similarmagnitude us those between hybrids and parental species. In:Atti Conv. Scient. “Fourth Congress of the European Society
for Evolutionary Biology”. Montpellier, August, 22-28: 409.SEMLITSCH R.D., Schmiedehausen S., Hotz H., Beerli P., 1996.
Genetic compatibility between sexual and clonal genomes inlocal populations of the hybridogenetic Rana esculenta com-plex. Evolutionary Ecology, 10: 531-543.
SEMLITSCH R.D., HOTZ H., GUEX G.D., 1997. Competition amongtadpoles of coexisting hemiclones of hybridogenetic Ranaesculenta: support for the frozen niche variation model. Evo-lution, 51, 4: 1249-1261.
SILIGARDI et al., 2000. I.F.F. Indice di Funzionalità Fluviale.ANPA (Agenzia Nazionale per la Protezione dell’Ambiente),Roma, 224 pp.
TUNNER H.G., NOPP H., 1979. Heterosis in the common Europeanwaterfrog. Naturwissenschaften, 66: 268-269.
TOKTAMYSOVA Z.S., KAIDAULOVA E.V., 1996. Growth, develop-ment, and respiration rate of Rana ridibunda Pall. larvaeexposed to chemicals. Russian Journal of Ecology, 27, 6: 450-451.
UZZELL T., DAREVSKY I.S., 1973. The relationships of Lacertaportschinskii and Lacerta raddei (Sauria, Lacertidae). Herpe-tologica, 29: 1-6.
UZZELL T., BERGER L., 1975. Electrophoretic phenotypes of R.rididunba, R. lessonae and their hybridogenetic associate, R.esculenta. Proc. Acad. Nat. Sci. Philadelphia, 127: 13-24.
UZZELL T., DAREVSKY I.S., 1975. Biochemical evidence for thehybrid origin of the parthenogenetic species of the Lacertasaxicola complex (Sauria: Lacertidae), with a discussion ofsome ecological and evolutionary implications. Copeia, 204-222.
VINOGRADOV A.E., CHUBINISHVILI A.T., 1999. Genome reductionin a hemiclonal frog Rana esculenta from radioactively conta-minated areas. Genetics, 151, 3: 1123-1125.
VITT L.J., CALDWELL J.P., WLBUR H.M., SMITH D.C., 1990. Amphi-bian as Harbingers of decay. Biosciences, 40: 418.
VRIJENHOEK R.C., PFEILER E., 1997. Differential survival of sexualand asexual poeciliopsis during environmental stress. Evolu-tion, 51, 5: 1593-1600.
WAKE D.B., 1991. Declining amphibian populations. Sciences,253: 860.
WALLAS S.C., BLAUSTAIN A.R. BEATTY J.J., 1992. Amphibianbiodiversity of the Pacific Northwest with special reference toold-growth stands. Northwest Environ. J., 8: 53-69.

Biologia Ambientale, 17 (1): 45-53, 2003
Effetti delle asciutte sulle cenosi macrobentoniche:taxa sensibili, resistenti e resilienti
Rossi Simone*, Simone Vanessa, Nardi Pietro Angelo, Montagna Silvia
Università degli Studi di Pavia, Dip. di Biologia Animale, Laboratorio di Ecologia delle Acque Interne, Piazza Botta, 9 - 27100 Pavia
* Autore referente per corrispondenza (e-mail: [email protected])
Pervenuto il 12.6.2002; accettato il 19.10.2002
RiassuntoIl lavoro si propone di analizzare, mediante analisi qualitative e quantitative delle biocenosi macrobentoniche effettuate successivamentea due periodi di asciutta, le relazioni che si instaurano tra gli organismi acquatici al fine di comprendere quali possano essere i taxa sensibilie quali viceversa traggano vantaggio, in termini sia di resistenza che di resilienza, da tale situazione. Obiettivo finale è quello di individuareuna lista di organismi tipici dei tratti soggetti ad asciutta che possano fungere da indicatori nei confronti di questa tipologia di alterazione.I risultati del lavoro mettono in luce la resistenza alle asciutte di un numero relativamente elevato di taxa, tra cui Elmidae e Goeridaerisultano i più abbondanti dal punto di vista quantitativo. Tra i taxa sensibili si rinviene la famiglia Gammaridae, dominante in termini didensità nei tratti con presenza continua di acqua e, viceversa, assente o rara nei tratti soggetti ad asciutte periodiche. I taxa Chironomidaee Baetis, pur relativamente sensibili alle asciutte, traggono vantaggio dalle stesse grazie ad una strategia di ricolonizzazione estremamenterapida, favorita dall’assenza di competitori e/o predatori.
PAROLE CHIAVE: asciutte / macrobenthos / ricolonizzazione
AbstractEffect of droughts to freshwater invertebrates: sensitive, resistant and resilient taxaThe purpose of this work is to analyse the ecological relationships among freshwater invertebrates after an environmental alteration dueto the absence of water. We wish to find which of these organisms are sensitive and which are resistant and/or resilient to this condition.Ultimately, we would create a check list containing taxa typical to these limiting conditions. The obtained results indicate that a lot of taxaare resistant to droughts (Tab. I). In particular, Elmidae and Goeridae (Fig. 3) are abundant. Viceverse, if we consider the sensitive taxa,the most important name is Gammaridae, which has a lot of densities (Tab. II, Fig. 3 and 4) in reaches with presence of water. Howeverit is absent or sporadic in dry reaches. The situation of Chironomidae and Baetis is special, because these taxa are sensitive to drought butthanking for their extremely fast recolonization strategy, they take advantage from dry situations.
KEY WORDS: droughts / freshwater invertebrates / recolonization
INTRODUZIONELe alterazioni delle caratteristiche fisiche dei corpi
idrici a seguito di eventi “catastrofici” quali piene(ANDERSON, 1992; MATTHAEI et al., 1996; DOLE-OLI-VIER et al., 1997; MATTHAEI et al., 1997), incendi(MINSHALL et al., 2001), eruzioni vulcaniche (ANDER-SON, 1992) ed asciutte (NECKLES et al., 1990; MACKAY ,1992) si riflettono inevitabilmente sulle biocenosi ma-crobentoniche. L’esame di tali situazioni, sia istanta-
neo che prolungato nel tempo, può essere molto utile alfine di comprendere le dinamiche di ricolonizzazione(recovery) dei tratti soggetti a impatto. I macroinverte-brati acquatici possono ricolonizzare un tratto alteratoin diversi modi: drift, nuoto attivo, movimenti adiacen-ti al fondo e deposizione delle uova da parte di adultialati. Velocità e modalità di ricolonizzazione dipendonodalla motilità degli organismi, dalla tessitura del sub-

ROSSI et al. - Effetti delle asciutte: taxa sensibili, resistenti e resilienti46
strato, dalle disponibilità alimentari nonché da fenome-ni di competizione e/o predazione (MACKAY , 1992). Ilpresente lavoro si propone di comprendere le rispostedelle comunità macrobentoniche all’impatto delle asciut-te totali. Tale situazione è stata già analizzata mediantericorso ad indici ecologici (ROSSI et al., 2002); tuttaviasi è ritenuto necessario approfondire l’analisi focaliz-zando l’attenzione sui rapporti qualitativi e quantitativitra le singole unità sistematiche al fine di comprenderese le biocenosi preesistenti scompaiano o, viceversa,vi siano organismi resistenti e/o taxa resilienti capaci dicostituire, anche a distanza di mesi, comunità differen-ti rispetto a quelle di tratti che mantengono sempreacqua all’interno dell’alveo. A tal proposito TOWNSEND
(1989) ha postulato che corsi d’acqua con regimiidrici estremamente variabili tendono ad essere domi-nati da specie a selezione r (MACARTHUR e WILSON,1967).
L’obiettivo finale è quello di individuare associazionidi organismi che potrebbero risultare indicatrici di“mancanza periodica d’acqua”. Ciò agevolerebbe, adesempio, la comprensione delle cause di alterazione dialcuni corpi idrici, anche senza sapere a priori se iltratto in esame possa essere stato o meno soggetto adasciutte o a forti riduzioni dell’alveo bagnato, dovute asiccità oppure a pratiche di ritenzione e/o captazioneidrica.
MATERIALI E METODI
Inquadramento territorialeLa ricerca è stata condotta sulla Roggia Roggione
(rif. B6a4-Trezzano Sul Naviglio CTR Lombardia),che scorre in Provincia di Milano e che fa parte delgruppo di corpi idrici connesso al sistema dei NavigliMilanesi. La Roggia Roggione riceve le acque delNaviglio Grande in Comune di Gaggiano ed è soggettaa due asciutte complete, una autunnale e una verso lafine del periodo invernale, rispettivamente della duratadi due mesi e un mese circa. Il tratto di corso monito-rato è quello che scorre in prossimità del Lago Boscac-cio in località Bonirola di Gaggiano (Fig. 1). Sonopresenti nell’area in esame tre differenti situazioni cuicorrispondono altrettante stazioni (Fig. 2). La stazionepiù a monte, denominata MON, durante i periodi diritenzione idrica non presenta acqua all’interno dell’al-veo. Il disalveo del tratto comprendente la stazione,durante il periodo di asciutta febbraio-marzo 2000, haimposto la successiva suddivisione dell’area MONnelle stazioni MONa, non intaccata dalle attività diescavazione e MONb.
La stazione a valle, chiamata VAL, pur mantenendoportate prossime allo zero presenta sempre volumi diacqua all’interno dell’alveo grazie probabilmente a fe-
nomeni di infiltrazione di acqua dal Lago Boscaccio.Nel tratto compreso tra le stazioni MON e VAL è
presente un salto artificiale che costituisce un ostacoloinvalicabile per la risalita controcorrente dei macroin-vertebrati acquatici. La stazione individuata a valle delsalto (MED) comprende un tratto che, pur asciutto neiperiodi di ritenzione idrica, non presenta ostacoli allarisalita degli organismi acquatici per cui potrebbe esse-re soggetto a fenomeni di ricolonizzazione controcor-rente durante i periodi di rilascio.
Corso idrico di riferimento per gli aspetti ecologici èla Roggia Rabica che presenta una situazione idroqua-litativa relativamente simile a quella della Roggia Rog-gione senza i problemi connessi alle fluttuazioni diportata. Il punto di campionamento (RAB) presceltosulla Rabica è quello posto in prossimità della CascinaLasso in Comune di Morimondo (Rif. A7e1-MottaVisconti Nord CTR Lombardia).
Metodiche utilizzateLe attività di campionamento sono state eseguite
mediante retino immanicato (indagini qualitative) e cam-pionatore di Surber con superficie campionabile pari a0,1 m2 (indagini quantitative). Per i dettagli relativi alletecniche di campionamento si rimanda a Ghetti (1997)per le indagini qualitative e a ROSSI et al. (2002) perquelle quantitative.
Il periodo di lavoro va dal 12 novembre 1999 (resti-tuzione tardo autunnale dell’acqua alla Roggia Roggio-ne) al 19 settembre 2000. Nel periodo seguente la
Fig. 1. Localizzazione delle stazioni di monitoraggio sulla RoggiaRoggione.

ROSSI et al. - Effetti delle asciutte: taxa sensibili, resistenti e resilienti 47
Fig. 2. Condizioni idriche delle stazioni della Roggia Roggione nei rispettivi periodi di rilascio (a sinistra) e di ritenzione (a destra).A = staz. MONa; B = staz. MONb; C = staz. MED; D = staz. VAL.
A
B
C
D

ROSSI et al. - Effetti delle asciutte: taxa sensibili, resistenti e resilienti48
prima asciutta (durata da metà settembre a metà no-vembre ’99) i campionamenti si sono susseguiti concadenza trisettimanale. Dopo la seconda asciutta (du-rata da fine febbraio a fine marzo 2000) i rilevamentisono stati effettuati ogni sei settimane circa. Nell’ela-borazione dei dati, al fine di mettere in luce particolaricontenuti informativi relativi all’evoluzione nel tempodelle biocenosi macrobentoniche, sono state utilizzatetecniche di statistica descrittiva, che hanno previstol’utilizzo della mediana come parametro principale inquanto i dati quantitativi relativi ai macroinvertebratinon seguono in genere distribuzioni normali di fre-quenza anche dopo trasformazioni logaritmiche (POTVIN
e ROFF, 1993). Inoltre si è fatto ricorso alla analisi diraggruppamento non gerarchica con il metodo delle kmedie (FABBRIS, 1997) applicata ai dati non standardiz-zati contenuti in due matrici di presenza. Per la costru-zione delle matrici è stato sommato per taxon, perperiodo (novembre ’99-febbraio ’00 e marzo ’00-settembre ’00) e per stazione, il numero di campiona-menti in cui è risultata certa la presenza di organismiappartenenti ad ogni gruppo sistematico in esame.
RISULTATI E DISCUSSIONE
Comunità presenti al momento del rilascio idrico(Tab. I: taxa resistenti e sensibili)
Nella tabella I è riportato un elenco inerente lapresunta resistenza o sensibilità di alcune unità siste-matiche nei confronti della mancanza d’acqua, stilatofacendo riferimento ai dati raccolti durante i monito-raggi (12/11/99 e 25/03/00) contemporanei al primorilascio dell’acqua dopo i periodi secchi. Per contem-poraneità del monitoraggio si intende il campionamen-to nelle ore immediatamente successive all’aperturadelle chiuse che regolano il flusso della Roggia Roggio-ne. Gli organismi considerati resistenti sono in generequelli rinvenuti in almeno una delle stazioni asciutte(MON, MONa, MONb, MED) in entrambi i campio-namenti (12/11/99 e 25/03/00). Quelli considerati sen-sibili risultano presenti in entrambe le occasioni (ecce-zionalmente in una sola) esclusivamente nella stazioneVAL. Da un’attenta interpretazione della tabella I emergeche gli organismi resistenti alle asciutte possiedonoalcune peculiari caratteristiche:• la famiglia Goeridae ha un fodero trasportabile ap-
piattito, di grani di sabbia tenacemente saldati tra lorocon pietruzze di zavorra (CAMPAIOLI et al., 1999) chepotrebbe trattenere acqua o comunque mantenereumida la zona adiacente l’animale;
• i gasteropodi, sia polmonati e quindi in grado direspirare ossigeno atmosferico (Physidae e Lymna-eidae) che prosobranchi (Bythiniidae), si comporta-no probabilmente in maniera analoga a Goeridae. La
capacità dei molluschi di conservare acqua all’inter-no della conchiglia come strategia contro l’essicca-mento è confermata da GIROD et al. (1980) e daCAMPAIOLI et al. (1994);
• la famiglia Tipulidae ha una piastra stigmatica ingrado di assumere ossigeno atmosferico mediantesifoni respiratori (CAMPAIOLI et al., 1999). La strate-gia contro la perdita d’acqua operata da questiorganismi è probabilmente quella di infossarsi nel
Tab. I. Taxa resistenti (in grado di sopravvivere alle asciutte),resilienti (favoriti, nel medio periodo, dalle asciutte) e sensibili(incapaci di sopravvivere e svantaggiati dalle asciutte).❍: risultati dell’analisi al rilascio dell’acqua, immediatamente dopoun periodo di asciutta. ●: risultati dell’analisi dei cluster applica-ta alle matrici di presenza nei periodi con flusso idrico (novembre’99-febbraio’00 e marzo’00-settembre’00).
Taxa Resistenti Resilienti Sensibili
Baetis ●
Ephemerella ❍
Goeridae ❍ ●
Hydropsychidae ●
Lepidostomatidae ❍●
Elmidae ❍ ●
Dryopidae ❍
Dytiscidae ❍
Haliplidae ❍
Onychogomphus ❍●
Ceratopogonidae ●
Chironomidae ● ❍
Tabanidae ❍ ●
Tipulidae ❍ ●
Naucoridae ❍●
Asellidae ❍ ●
Gammaridae ❍●
Bythiniidae ❍ ●
Lymnaeidae ❍ ●
Physidae ❍ ●
Dugesia ❍
Haemopis ❍
Lumbricidae ❍ ●
Lumbriculidae ❍ ●

ROSSI et al. - Effetti delle asciutte: taxa sensibili, resistenti e resilienti 49
substrato sabbioso;• la famiglia Tabanidae, le cui larve sono semiacqua-
tiche e possono vivere nei terreni umidi prossimi allerive (CAMPAIOLI et al., 1999), sembra comportarsi inmaniera del tutto simile a Tipulidae;
• Haemopis è un genere di irudinei capace di soprav-vivere a lungo in terra umida, in cui si porta non dirado. L’accoppiamento fra esemplari appartenenti aquesto genere avviene per lo più fuori dall’acqua e ladeposizione delle uova è effettuata in ambiente terre-stre (MINELLI, 1977);
• gli oligocheti, in particolare la famiglia Lumbricidae,hanno abitudini fossorie e molti generi appartenenti algruppo sono terrestri (SANSONI, 1988);
• il genere Dugesia sembra sfruttare le caratteristichedel proprio corpo, appiattito, per rifugiarsi nel subal-veo;
• la famiglia degli Elmidae è indicata come prettamenteacquatica, raramente a vita anfibia (RUFFO e SIMONET-TO in CAMPAIOLI et al., 1999) o incapace di vivere fuoridall’acqua (OLMI, 1978). I risultati del presente lavo-ro non confermano tali citazioni, poiché risulta moltoevidente la resistenza di questo taxon nei confrontidelle asciutte, testimoniata dal rinvenimento di uncospicuo numero di esemplari allo stato larvale intratti privi d’acqua. La chiave della resistenza diquesti coleotteri nei confronti delle asciutte potrebbeessere la struttura del tegumento larvale, che sipresenta complesso e costituito da setole, spine, pelie rigonfiamenti mammellonari, con un opercolo chericopre gli stiletti anali, nei pressi delle tracheobran-chie rettali. Particolare interessante dell’autoecologiadegli Elmidae è la fase di impupamento, che sembraavvenire nel bagnasciuga (OLMI , 1978). SANSONI
(1988) afferma che le pupe sono con poche eccezio-ni terrestri, per lo più ripicole;
• i coleotteri appartenenti alle famiglie Dryopidae,Dytiscidae e Haliplidae, presenti nel campionamentoprimaverile in alcune stazioni asciutte, sono organi-smi che, almeno nella forma adulta, sono in grado direspirare ossigeno atmosferico e di spostarsi in voloda biotopi con condizioni ambientali sfavorevoli (OLMI ,1978; RUFFO e SIMONETTO in CAMPAIOLI et al., 1999);
• la famiglia Asellidae nel monitoraggio contempora-neo al secondo rilascio idrico è presente anche neitratti asciutti con un discreto numero di esemplari.SANSONI (comun. pers.) afferma a proposito che perquesti crostacei è possibile dimostrare una fortecapacità d’adattamento a situazioni di asciutta. Sono,infatti, organismi che frequentano spesso ambientiastatici e quindi di provata resistenza all’estivazione.
Gli organismi sensibili, viceversa, non sono infossa-tori (eccetto Onychogomphus) e non hanno strutture
minerali di protezione. Tra di essi la famiglia Lepido-stomatidae presenta fodero che è in gran parte vegetalee che probabilmente non funge da adeguata strutturacontro la perdita d’acqua, mentre la presunta sensibili-tà del genere Ephemerella è ipotizzabile dalla conside-razione che, pur non essendo stati rinvenuti in data 12/11/99 esemplari in nessun tratto, il gruppo comparenel corso nel mese di gennaio 2000 e permane dopol’asciutta di febbraio-marzo 2000 solo nella stazioneVAL.
È opportuno rilevare a questo punto che per gliorganismi considerati come resistenti è da escludere ilfenomeno del drift, poichè è improbabile che la ricolo-nizzazione dei substrati avvenga in poche ore. Inoltre,ROSSI e LUCHELLI (dati non pubblicati), nell’ottobre2001, dopo circa tre settimane dall’inizio di un periodoasciutto, hanno constatato il rinvenimento di esemplarivivi appartenenti ai gruppi Goeridae, Elmidae, Lumbri-cidae e Lumbriculidae in un piccolo campione dighiaia raccolto negli strati superficiali del substrato eposto successivamente in acqua.
Un’altra considerazione va fatta sulla persistenza dimicrohabitat umidi che, durante secche di durata noneccessiva (che eliminano comunque la quasi totalitàdegli organismi) non permettono la completa eradica-
Fig. 3. Organismi presenti nelle stazioni della Roggia Roggione almomento del rilascio dell’acqua. I valori riportati corrispondonoalla media dei 3 sottocampioni raccolti su substrato dominante esono espressi come numero di individui per 0,1 m2.

ROSSI et al. - Effetti delle asciutte: taxa sensibili, resistenti e resilienti50
zione di una unità tassonomica sensibile alle asciutte. Èprobabilmente riconducibile a questo fenomeno il rin-venimento occasionale, al momento del rilascio idricoprimaverile, di Naucoridae e Gammaridae, che le inda-gini quantitative condotte lungo tutto il periodo dilavoro hanno dato come estremamente intolleranti neiconfronti della mancanza d’acqua. ROSSI (oss. pers.,dicembre 2001), a proposito, segnala il ritrovamentosul Fiume Adda di esemplari vivi appartenenti ai taxaBaetis, Hydropsychidae e Gammaridae in alveo asciut-to, dimostrando la loro capacità di resistere, almenoper brevi periodi, in ambiente umido.
Dal punto di vista quantitativo (Fig. 3) è possibileosservare come, al momento del rilascio dell’acqua, itratti asciutti siano dominati numericamente dagli El-midae cui si affiancano con minori densità Goeridae eBythiniidae. Viceversa in VAL (non interessata daasciutte totali) si osserva anche la presenza, con carat-tere dominante nel mese di marzo, di Gammaridae,nonché di Chironomidae.
Evoluzione qualitativa delle biocenosi(Tab. I: taxa resilienti e sensibili)
L’analisi dei cluster con il metodo delle k medie (kpari a 3) applicata alle matrici di presenza ha messo inluce, per il periodo novembre ’99-febbraio ’00, i se-
guenti raggruppamenti:• taxa resilienti: Goeridae, Elmidae, Chironomidae,
Tabanidae, Tipulidae, Asellidae, Bythiniidae, Lymna-eidae, Lumbricidae, Lumbriculidae;
• taxa saltuariamente presenti: Baetis, Ephemerel-la, Dryopidae, Haliplidae, Empididae, Simuliidae,Physidae, Dugesia, Haemopis;
• taxa sensibili alle asciutte: Hydropsychidae, Lepi-dostomatidae, Onychogomphus, Naucoridae, Gam-maridae.Tra i taxa resilienti si osservano sia organismi che
sopravvivono alle asciutte (resistenti) sia gruppi pio-nieri quali Chironomidae che, pur assenti nelle stazioni“asciutte” durante il campionamento contemporaneoal rilascio dell’acqua, non mancano mai nei prelievisuccessivi. Tra i taxa saltuariamente presenti si rinven-gono sia generi come Baetis ed Ephemerella checompaiono a gennaio, sia altri gruppi sistematici rileva-ti in maniera saltuaria. Tra i taxa sensibili si osservanogruppi intolleranti verso le asciutte totali e che risultanoquindi presenti quasi esclusivamente nella stazione VAL.
L’applicazione dell’analisi al periodo marzo ’00-settembre ’00 rileva una situazione analoga:• taxa resilienti: Goeridae, Baetis, Elmidae, Chirono-
midae, Bythiniidae, Lymnaeidae, Physidae, Lumbri-cidae, Lumbriculidae;
Tab. II . Comunità tipiche della Roggia Roggione. I valori riportati corrispondono alle mediane (n° individui per 0,1 m2) per taxon relativeai periodi novembre 1999-febbraio 2000 e marzo-settembre 2000.
taxa novembre 1999-febbraio 2000 marzo 2000-settembre 2000MON MED VAL MONa MONb MED VAL
Baetis 0,7 0,2 0,4 31,3 104,3 39,7 5,3Caenis 0,0 0,0 0,0 0,7 1,0 0,0 0,3Ephemerella 0,0 0,0 0,0 3,3 0,7 0,7 4,0Goeridae 9,7 8,0 11,7 2,7 2,3 3,0 10,3Hydropsychidae 0,2 0,0 1,2 3,0 1,3 2,3 1,7Lepidostomatidae 0,2 0,9 6,5 0,0 0,0 0,0 0,0Dryopidae 0,2 0,0 0,0 1,0 0,3 0,3 0,0Elmidae 83,0 96,2 116,0 68,3 21,3 52,0 183,7Onychogomphus 0,0 0,0 0,2 0,0 0,0 0,0 0,3Ceratopogonidae 0,0 0,0 0,0 0,0 0,0 0,3 0,7Chironomidae 20,0 16,7 15,9 17,7 3,7 3,3 9,0Simuliidae 1,0 0,0 0,5 0,3 1,0 0,7 0,3Tabanidae 0,0 0,3 0,0 0,0 0,3 0,3 0,0Tipulidae 0,0 2,8 0,5 0,0 0,0 0,3 0,0Naucoridae 0,0 0,0 1,5 0,0 0,0 0,7 4,0Asellidae 0,7 0,9 0,3 1,0 0,0 0,0 0,0Gammaridae 0,0 0,0 41,5 1,7 0,7 9,7 399,7Bythiniidae 2,0 1,5 0,0 1,7 0,3 3,0 0,0Lymnaeidae 1,0 2,0 1,0 2,0 0,3 5,0 0,3Physidae 0,2 0,7 0,0 1,0 0,0 0,3 0,0Planorbidae 0,0 0,2 0,0 0,7 0,0 0,0 0,0Dugesia 0,2 0,2 0,5 0,0 0,0 0,0 0,0

ROSSI et al. - Effetti delle asciutte: taxa sensibili, resistenti e resilienti 51
• taxa saltuariamente presenti: Hydropsychidae,Caenis, Ephemerella, Dryopidae, Dytiscidae, Hali-plidae, Simuliidae, Tabanidae, Tipulidae, Asellidae,Planorbidae, Dugesia;
• taxa sensibili alle asciutte: Lepidostomatidae,Onychogomphus, Ceratopogonidae, Naucoridae,Gammaridae.Si noti come Baetis sia entrato a far parte dei taxa
resilienti. Nonostante tali efemerotteri manchino dalcorso d’acqua al momento del rilascio primaveriledell’acqua, grazie alla strategia di ricolonizzazioneadottata (r), essi riescono in seguito a diffondersiovunque.
Evoluzione quantitativa delle biocenosiNella tabella II è possibile osservare le biocenosi
tipiche di ogni tratto campionato nei periodi novembre’99-febbraio ’00 e marzo ’00-settembre ’00, con lerelative numerosità. I valori di numerosità sono parialle mediane di ogni periodo. Si noti come nel periodofreddo le comunità dei tratti MON e MED siano moltosemplificate, con Elmidae dominante e Chironomidae,Goeridae, Bythiniidae e Lymnaeidae presenti. Vicever-sa nel tratto a valle si osserva, oltre all’assenza o allasporadica presenza di gasteropodi, anche l’abbondan-za di Gammaridae nonché un sufficiente numero diLepidostomatidae, Naucoridae e Hydropsychidae. Nel
Fig. 4. Distribuzioni di frequenza delle unità sistematiche principali nel periodo di monitoraggio. I valori di numerosità sono riportati inscala logaritmica. I risultati contemporanei al rilascio idrico (12/11/99 e 25/03/00) sono indicati, per le stazioni MONa, MONb, MED eVAL, con un marcatore di maggiori dimensioni.
Gammaridae
0,0
1,0
2,0
3,0
4,0
5,0
6,0
7,0
12/1
1/99
10/1
2/99
7/1/
00
4/2/
00
3/3/
00
31/3
/00
28/4
/00
26/5
/00
23/6
/00
21/7
/00
18/8
/00
15/9
/00
data
ln (
num
eros
ità+1
)
MONaMONbMEDVALRAB
Elmidae
0,0
1,0
2,0
3,0
4,0
5,0
6,0
7,0
12/1
1/99
10/1
2/99
7/1/
00
4/2/
00
3/3/
00
31/3
/00
28/4
/00
26/5
/00
23/6
/00
21/7
/00
18/8
/00
15/9
/00 data
ln (
num
eros
ità+1
)
MONaMONbMEDVALRAB
Goeridae
0,0
1,0
2,0
3,0
4,0
5,0
6,0
7,0
12/1
1/99
10/1
2/99
7/1/
00
4/2/
00
3/3/
00
31/3
/00
28/4
/00
26/5
/00
23/6
/00
21/7
/00
18/8
/00
15/9
/00 data
ln (
num
eros
ità+1
)
MONaMONbMEDVALRAB
Chironomidae
0,0
1,0
2,0
3,0
4,0
5,0
6,0
7,0
12/1
1/99
10/1
2/99
7/1/
00
4/2/
00
3/3/
00
31/3
/00
28/4
/00
26/5
/00
23/6
/00
21/7
/00
18/8
/00
15/9
/00 data
ln (
num
eros
ità+1
)
MONaMONbMEDVALRAB
Ephemerella
0,0
1,0
2,0
3,0
4,0
5,0
6,0
7,0
12/1
1/99
10/1
2/99
7/1/
00
4/2/
00
3/3/
00
31/3
/00
28/4
/00
26/5
/00
23/6
/00
21/7
/00
18/8
/00
15/9
/00 data
ln (
num
eros
ità+1
)
MONaMONbMEDVALRAB
Baetis
0,0
1,0
2,0
3,0
4,0
5,0
6,0
7,0
12/11/99
10/12/99
7/1/00
4/2/00
3/3/00
31/3/00
28/4/00
26/5/00
23/6/00
21/7/00
18/8/00
15/9/00
data
ln (
nu
me
ros
ità+
1)
MONaMONbMEDVALRAB

ROSSI et al. - Effetti delle asciutte: taxa sensibili, resistenti e resilienti52
periodo primaverile ed estivo la situazione ecosistemi-ca dei tratti asciutti, non dissimile dalla precedente, sifa leggermente più varia a seguito della presenza diBaetis, Ephemerella e Hydropsychidae. Da notare chele comunità di MONb, pur azzerate a seguito deldisalveo, risultano alla fine del periodo estivo analoghea quelle di MONa. La presenza di Gammaridae nellastazione MED è dovuta alla ricolonizzazione controcorrente dal tratto VAL. Quest’ultimo è caratterizzato,oltre che dagli organismi di cui sopra, da Naucoridae. IGammaridae in particolare caratterizzano il tratto VALcon esplosioni demografiche rilevanti. Tale crescita, inmisura meno accentuata, si riscontra negli Elmidae.
Analisi delle unità sistematiche principaliL’andamento delle popolazioni delle unità sistemati-
che che caratterizzano la comunità macrobentonicadella Roggia Roggione è illustrato in figura 4. Perconfronto sono indicate nei grafici anche le popolazio-ni rilevate sulla Roggia Rabica (stazione di riferimen-to).
Il gruppo di maggior peso informativo è Gammari-dae, la cui comunità, analizzata da un punto di vistaquantitativo, può fornire utili indicazioni sulla presenzadi acqua all’interno di un corso. In particolare si notacome tali organismi siano estremamente sensibili neiconfronti delle asciutte complete. La particolare dina-mica di ricolonizzazione, che sembra avvenire princi-palmente controcorrente (DAHL e GREENBERG, 1996)ed esclusivamente nel periodo primaverile può essereimpedita da salti d’acqua e sbarramenti, il che ostacolaulteriormente il ripristino degli equilibri ecologici post-asciutta. La scomparsa del taxon dalla Roggia Roggio-ne nel mese di settembre, come accennato nel lavorogià citato (ROSSI et al., 2002), è con tutta probabilitàdovuta ad alterazioni di tipo chimico tossico (pesticidie/o metalli pesanti).
La popolazione di Elmidae, ben strutturata e relati-vamente costante in termini di densità, caratterizza ilcorso del Roggione e non risente del problema asciut-te. Si osserva viceversa che il disalveo distrugge quasicompletamente gli individui di MONb e che il processodi ripristino delle condizioni originarie non è poi cosìbreve.
La famiglia Goeridae è anch’essa ampiamente diffu-sa nel Roggione. La popolazione è sempre presentecon un discreto numero di individui e sembra nonrisentire del problema asciutte. Tale affermazione èvera soprattutto per i soggetti più piccoli, che si rin-vengono con maggior frequenza dopo le secche men-tre quelli di maggiori dimensioni sembrano in partevulnerabili.
La famiglia Chironomidae, pur sensibile all’asciuttache ne annienta gli esemplari, grazie alla strategia
pioniera r (ANDERSON, 1992; MACKAY , 1992; MINSHALL
et al., 2001) si reintroduce in breve tempo all’internodel corso d’acqua. Paradossalmente la secca, chenuoce sicuramente ai singoli individui (assenza di resi-stenza), tende viceversa a favorire selettivamente iltaxon (elevata resilienza) mediante eliminazione perio-dica di predatori e/o competitori di nicchia e a permet-tere quindi le esplosioni demografiche trascorso unbreve periodo dal ripristino del flusso idrico.
Il genere Ephemerella soffre particolarmente leasciutte. Più precisamente scompare completamentenei casi di mancanza d’acqua e si riduce in numerodurante le asciutte parziali. Tale fenomeno è particolar-mente evidente a fine marzo. La presenza di popolazio-ni molto dense nel mese di maggio non sembra indicedi euriecia del taxon nei confronti della problematica inesame ma appare dovuta a drift o a soggetti adulti dicorsi limitrofi che, una volta sfarfallati, depongono leuova nella Roggia Roggione.
Relativamente al genere Baetis, mentre ogni esem-plare appare molto sensibile nei confronti delle asciuttesia totali che parziali e sembra scomparire in assenza diflusso d’acqua, indipendentemente dalla presenza dellastessa, il gruppo nel complesso tende a trarre vantag-gio dalle secche periodiche, grazie ad una strategiapioniera simile a quella di Chironomidae. La caratteri-stica abilità pioniera è confermata da ANDERSON (1992)e MACKAY (1992) nonché da MATTHAEI et al. (1996) eMINSHALL et al. (2001). Nei tratti con alveo periodica-mente asciutto possono verificarsi esplosioni demo-grafiche ostacolate invece, nelle zone perennementebagnate, da fenomeni di competizione interspecifica.
CONCLUSIONII dati del presente lavoro forniscono indicazioni
relative sia all’ecologia applicata che di base. In primoluogo è possibile identificare le comunità tipiche degliambienti di roggia soggetti ad asciutta periodica. Inparticolare la presenza di una biocenosi costituita inprevalenza da Elmidae, Goeridae, da gasteropodi dellefamiglie Physidae, Lymnaeidae e Bythiniidae, da Baetise Chironomidae e la contemporanea assenza, in am-bienti potenzialmente idonei ad ospitarli, di Gammari-dae può fungere da segnale che una delle possibilicause di alterazione degli ambienti in esame potrebbeessere un’asciutta antecedente la data di campiona-mento. Tale considerazione è particolarmente impor-tante nel caso di indagini sulle biocenosi macrobento-niche preliminari ad attività di reintroduzione di specieittiche e/o di crostacei di particolare pregio ecologico(come Austrapotamobius pallipes).
Richiamando alcune nozioni dell’ecologia di base sinota come, almeno parzialmente, i tratti soggetti adasciutta possano essere assimilati, con alcune limita-

ROSSI et al. - Effetti delle asciutte: taxa sensibili, resistenti e resilienti 53
zioni (MACKAY , 1992) alle isole indicate da MACAR-THUR e WILSON (1967), all’interno delle quali risultapossibile osservare (ed eventualmente analizzare neidettagli) la dinamica di colonizzazione delle speciepioniere (a strategia r). In appendice a quanto detto si
nota come gli ambienti con comunità macrobentoni-che sufficientemente strutturate, mentre ostacolano leesplosioni demografiche di taxa quali Chironomidae eBaetis, permettano viceversa quelle di gruppi come iGammaridae.
BIBLIOGRAFIA
ANDERSON N. H., 1992. Influence of disturbance on insect commu-nities in Pacific Northwest streams. Hydrobiologia 248: 79-92.
CAMPAIOLI S., GHETTI P.F., MINELLI A., RUFFO S., 1994. Manualeper il riconoscimento dei macroinvertebrati delle acque dolciitaliane, Vol. I. Provincia Autonoma di Trento, 357 pp.
CAMPAIOLI S., GHETTI P.F., MINELLI A., RUFFO S., 1999. Manualeper il riconoscimento dei macroinvertebrati delle acque dolciitaliane, Vol. II. Provincia Autonoma di Trento, 126 pp.
DAHL J., GREENBERG L., 1996. Effects of habitat structure onhabitat use by Gammarus pulex in artificial streams. Freshwa-ter biology 36: 487-495.
DOLE-OLIVIER M. J., MARMONIER P., BEFFY L., 1997. Response ofinvertebrates to lotic disturbance: is the hyporheic zone apatchy refugium? Freshwater Biology 37: 257-276.
GHETTI P. F., 1997. Indice Biotico Esteso (I.B.E.). I macroinver-tebrati nel controllo della qualità degli ambienti di acquecorrenti. Manuale di applicazione. Provincia Autonoma diTrento, Agenzia Provinciale per la Protezione dell’Ambiente,222 pp.
GIROD A., BIANCHI I., MARIANI M., 1980. Gasteropodi 1 (Gastro-poda: Polmonata. Prosobranchia: Neritidae, Viviparidae,Bythiniidae, Valvatidae). Guida CNR n. 7 – AQ/1/44, 86 pp.
FABBRIS L., 1997. Statistica multivariata, analisi esplorativa deidati. McGraw-Hill, 437 pp.
MACARTHUR R. H., WILSON E. O., 1967. The theory of islandbiogeography. Princeton University Press, Princeton NJ, 203pp.
MACKAY R. J., 1992. Colonization by Lotic Macroinvertebrates:A Review of Processes and Patterns. Canadian Journal ofFisheries and Acquatic Sciences 49: 617-628.
MATTHAEI C. D., UEHLINGER U., MEYER E. I., FRUTIger A., 1996.Recolonization by benthic invertebrates after experimentaldisturbance in a Swiss prealpine river. Freshwater Biology 35:233-248.
MATTHAEI C. D., UEHLINGER U., FRUTIGER A., 1997. Response ofbenthic invertebrates to natural versus experimental disturban-ce in a Swiss prealpine river. Freshwater Biology 37: 61-77.
MINELLI A., 1977. Irudinei. Guida CNR n. 1. AQ/1/2, 43 pp.MINSHALL G. W., ROYER T. V., ROBINSON C. T., 2001. Response
of the Cache Creek macroinvertebrates during the first 10 yearsfollowing disturbance by the 1988 Yellowstone wildfires.Canadian Journal of Fisheries and Acquatic Sciences 58: 1077-1088.
NECKLES H. A., MURKIN H. R., COOPER J. A, 1990. Influences ofseasonal flooding on macroinvertebrate abundance in wetlandshabitats. Freshwater Biology 23, 311-322.
OLMI M., 1978. Driopidi, Elmintidi. Guida CNR n. 2. AQ /1/6, 72pp.
POTVIN C., ROFF D. A., 1993. Distribution-free and robuststatistical methods: viable alternatives to parametric statisti-cs? Ecology 74: 1617-1628.
ROSSI S., MONTAGNA S., NARDI P. A., SIMONE V., 2002. Effetti delleasciutte sulle biocenosi macrobentoniche: confronto tra indiciecologici. Biologia Ambientale 16 (1): 1-6
SANSONI G., 1988. Atlante per il riconoscimento dei macroinver-tebrati dei corsi d’acqua italiani. Provincia autonoma di Tren-to, Agenzia Provinciale per la Protezione dell’Ambiente, 191pp.
TOWNSEND C. R., 1989. The patch dynamic concept of streamcommunity ecology. Journal of the North American Bentholo-gical Society 8: 36-50.


Biologia Ambientale, 17 (1): 55-65, 2003
Analisi della struttura del paesaggio mediante dati telerilevatidel sensore MIVIS: il bacino del torrente Baganza (Parma)
Pierfrancesca Rossi * e Alessandro Ferrarini, Orazio Rossi, Giovanni ZurliniDipartimento di Scienze Ambientali, Università degli Studi di Parma, Viale delle Scienze 11/A - 43100 Parma
* Autore referente per la corrispondenza ([email protected])
Pervenuto l’1.7.2002; accettato il 25.11.2002
RiassuntoÈ stata condotta un’analisi ad elevata risoluzione spaziale del mosaico di habitat del bacino idrografico del torrente Baganza (Prov. Parma).Le caratteristiche strutturali del territorio influenzano i processi che avvengono entro e tra gli habitat: il monitoraggio dei processi è lungoe costoso, quello delle strutture è più agevole, soprattutto se si utilizza uno strumento idoneo per questo tipo di analisi quale iltelerilevamento.Allo scopo di riconoscere con elevata accuratezza gli habitat presenti all’interno dell’area di studio, sono state classificate mediante unalgoritmo a reti neurali dodici immagini telerilevate del sensore iperspettrale MIVIS (Multispectral Infrared and Visible ImagingSpectrometer). Le classificazioni territoriali ottenute sono state georeferite e mosaicate tra loro. Sulla mappatura risultante sono staticalcolati diversi indici del paesaggio utilizzando il programma informatico FRAGSTAT. Tali indici sono stati poi analizzati allo scopo diricavare la massima informazione possibile sulle caratteristiche del paesaggio. La classificazione dell’area di studio è stata quindisovrapposta al modello digitale del terreno (DEM) allo scopo di suddividere la valle in fasce altimetriche (planiziale, collinare e montana).In tali fasce sono stati nuovamente calcolati gli indici del paesaggio allo scopo di individuarne le caratteristiche strutturali.Lo studio ha evidenziato l’utilità e la potenzialità del telerilevamento ad alta risoluzione per l’analisi delle caratteristiche strutturalidell’ecomosaico ambientale.
PAROLE CHIAVE: mosaico ambientale / analisi strutturale / sensore iperspettrale MIVIS / telerilevamento
AbstractLand-mosaic structural analysis through MIVIS sensor remotely sensed dataThis work aims to analyse land mosaic within the Val Baganza stream watershed (Parma, Italy) using a fine spatial resolution. Theanalysis of landscape structural properties is important because habitat spatial configuration influences ecological processes within andamong habitats. Furthermore while processes monitoring is difficult and expensive, the analysis of landscape structure is much easier.The actual scientific literature enhances the importance of remote sensing for this kind of analyses. Twelve remotely sensedhyperspectral MIVIS (Multispectral Infrared and Visible Imaging Spectrometer) images of the study area have been classified using aneural network algorithm. The aim of this operation was the high accurated recognition of habitats inside the Val Baganza streamwatershed. MIVIS classifications have been georeferenced and conjoined. The resulting map has been used to calculate some landscapeindices using FRAGSTAT software. These indices have been analysed aiming to get as much information as possible about structurallandscape properties. The map of the study area has been overlayed to the digital terrain model (DEM). Then, the whole watershed hasbeen divided into three altitudinal belts (lowland, hill land and mountain) and landscape indices have been calculated again to enhancestructural characteristics of these altitudinal regions.The study enhanced the utility and potential of high resolution remote sensing for the analysis of land mosaic structural properties.
KEY WORDS: land mosaic / structural analysis / hyperspectral MIVIS scanner / remote sensing

ROSSI P. et al. - Analisi del paesaggio mediante dati telerilevati56
INTRODUZIONEL’ecologia del paesaggio è stata definita in modi
diversi dagli studiosi nel corso del tempo. Troll (SCHREI-BER, 1990) la definisce come “lo studio delle comples-se relazioni di causa-effetto esistenti tra le biocenosi ele condizioni ambientali in una data porzione di territo-rio”. La definizione corrente più nota è quella di Risser(SCHREIBER, 1990), secondo cui l’ecologia del paesag-gio è “la disciplina che studia lo sviluppo e le dinamichedell’eterogeneità spaziale, la gestione di tale eterogenei-tà e la sua influenza sui processi biotici e abiotici”.
Nell’ecologia del paesaggio il landscape viene de-scritto attraverso i concetti di forma, funzione e cam-biamento nel tempo. La forma (struttura) del paesag-gio è generata dai flussi di materia, energia ed informa-zione e, viceversa, i flussi sono determinati dalla formadel landscape, nonché ne determinano il cambiamentonel tempo. Gli ecologi utilizzano vari strumenti per lostudio del paesaggio tra i quali gli esperimenti allamicroscala, la modellistica matematica (TURNER e GAR-DNER, 1991), la geostatistica (autocorrelazione spazia-le, semivariogrammi, analisi tessiturale) (RIPLEY, 1981)e le foto aeree, ma ciò che ha effettivamente rivoluzio-nato la percezione e l’approccio alla conoscenza delpaesaggio è stato l’utilizzo del telerilevamento (HALL etal., 1988; JOHNSTON e BONDE, 1989). Il remote sensingcostituisce un approccio sinottico alla mappatura delterritorio, che permette la mappatura e l’analisi dellecaratteristiche strutturali dell’ecomosaico ambientale elo studio dei suoi cambiamenti nel corso del tempo(SINGH, 1989). Gli ecosistemi della regione mediterra-nea presentano un elevato grado di frammentazione(NAVEH e LIEBERMAN, 1994; PIGNATTI, 1994) dovutoad un incessante impatto antropico; ne consegue lanecessità di operare a grana fine nel riconoscimento dihabitat a diverso grado di naturalità. Per gli scopi delpresente lavoro è stato utilizzato il sensore MIVIS, lacui elevata risoluzione spaziale e spettrale è idoneaall’analisi di un mosaico ambientale estremamente fram-mentato come quello tipico del nostro paese.
AREA DI STUDIO, MATERIALI E METODIL’area di studio è rappresentata dal bacino idrogra-
fico del Torrente Baganza, posto nel versante emilianodell’Appennino settentrionale in provincia di Parma(Fig. 1).
Chiusa fra la valle del torrente Parma a est e quelladel torrente Taro a ovest, la Val Baganza si estende perun’area compresa fra i 57 m sul livello del mare e il suopunto più alto, la cima del monte Cervellino, 1493 m.La forma del bacino risulta molto allungata nel sensodella linea di drenaggio con un rapporto di allungamen-to di 0,35 ed un’orientazione SO-NE. La lunghezzacomplessiva longitudinale dell’asta fluviale del torrente
Baganza è di 57,37 km, mentre in senso trasversale sipassa da un minimo di 3 km ad un massimo di 6.Rilevante è anche la differenza fra i due versanti cherisultano asimmetrici nel rapporto di 1:3 fra sinistro edestro. Ne consegue che i maggiori affluenti, perportata e per lunghezza, scorrono sul versante di de-stra che presenta una maggiore densità di drenaggio euna conseguente concentrazione dei deflussi nel reti-colo idrografico. Sul versante sinistro prevale un ru-scellamento superficiale con brevi e ripidi rii ad altacapacità erosiva per effetto delle pendenze e di unsubstrato a terreni argillosi, spesso caotici per frane.Nel versante destro del bacino si trova una strutturaidrografica dendritica, discretamente gerarchizzata astruttura elementare con bacini di primo e secondoordine. L’attività umana si concentra nei nuclei urbanilimitrofi al capoluogo di provincia. L’area planiziale edi prima collina è dominata da un’agricoltura intensivalegata alle necessità produttive della filiera agro-ali-mentare del parmense. La zona montana ed in partequella collinare risentono, al contrario, di un decennaleprocesso di spopolamento ed invecchiamento con unprogressivo abbandono della presenza umana sul terri-torio. La valle del torrente Baganza si configura quindicome una tipica valle appenninica, sia dal punto di vistageomorfologico che dell’influenza antropica. Questoaspetto rende paradigmatico lo studio della strutturaassunta dal suo mosaico ambientale.
Le immagini dell’area di studio sono state acquisitedallo scanner MIVIS (Daedalus AA5000 MultispectralInfrared and Visible Imaging Spectrometer del Consi-glio Nazionale Ricerche, Progetto L.A.R.A.-Laborato-rio Aereo per Ricerche Ambientali) che opera con 102bande spettrali ripartite su quattro sensori, il primo con20 bande nel visibile (0,43-0,83 mm), il secondo con 8bande nel vicino infrarosso (1,15-1,55 mm), il terzocon 64 bande nell’infrarosso medio (2,00-2,50 mm) el’ultimo con 10 bande nel termico (8,20-12,70 mm).L’intero bacino idrografico del Torrente Baganza èstato ripreso in dodici immagini MIVIS, disposte pa-rallelamente all’asse longitudinale della valle e realizzatein data 10 luglio 1998 (con inizio alle ore 13,35) allaquota di volo di 3000 m assoluti. A tale quota, larisoluzione spaziale del sensore (dimensione di un pixel)è mediamente di 4 metri (corrispondente ad una scalapari circa a 1:7500).
In un precedente lavoro (FERRARINI et al., 2001) laclassificazione dell’area di studio è stata realizzatainserendo l’informazione spettrale fornita dal sensoreMIVIS all’interno di una rete neurale di tipo MLP(multiperceptrone; RUMELHART e MCCLELLAND, 1986).I notevoli livelli di accuratezza raggiunti (intorno al98%) nel riconoscimento degli habitat dell’area di stu-dio assicurano grande precisione nel modo in cui sono

ROSSI P. et al. - Analisi del paesaggio mediante dati telerilevati 57
Fig. 1. Limite vettoriale (in nero) del bacino idrografico del torrente Baganza sovrapposto al modello digitale del terreno (DEM). In altoa sinistra, la posizione della Val Baganza rispetto all’Italia. A destra, il reticolo idrografico del bacino e la suddivisione in fasce altitudinali.
m

ROSSI P. et al. - Analisi del paesaggio mediante dati telerilevati58
state colte le caratteristiche dell’arrangiamento spazia-le del paesaggio. Il sistema di classificazione deglihabitat adottato è quello proposto nel progetto Biotopidel programma CORINE della UE (C.E.C., 1991),finalizzato a riconoscere ed elencare i siti di maggioreimportanza per la conservazione della natura nel terri-torio comunitario. Gli habitat, individuati sulla base divariabili ecologiche, vengono identificati da un codicenumerico gerarchico in cui la prima cifra definisce unadelle sette categorie di base (comunità costiere e alofi-tiche, comunità di acque dolci, praterie e arbusteti,foreste, acquitrini e paludi, spiagge e pareti rocciose,aree agricole e antropizzate) e, insieme alla seconda,denota il tipo generico di habitat. Le prime due cifresono seguite da un numero di decimali, fino ad unmassimo di cinque, utilizzati per descrivere con mag-giore precisione i singoli tipi di habitat. In tabella 1 èriassunta la corrispondenza tra le tipologie di habitatriconosciute mediante il sensore MIVIS ed il sistema diclassificazione CORINE.
Per gli scopi del presente lavoro, le dodici classifi-cazioni delle immagini MIVIS sono state georiferitesulla base di ortofoto digitali dell’area di studio esuccessivamente mosaicate per ottenere la mappa fi-nale degli habitat. Tale mappatura è stata vettorializzata
all’interno del GIS ArcView e quindi inserita all’internodel software FRAGSTAT (MCGARIGAL e MARKS, 1995)per procedere all’analisi degli aspetti strutturali delmosaico ambientale. Gli indici del pattern del paesag-gio possono essere calcolati a tre livelli gerarchici:dell’intero paesaggio (livello A), di tipologie di habitat(livello B) e di singoli habitat. Il terzo livello non è statoconsiderato per gli scopi del presente lavoro. Nellascelta degli indici da utilizzare sono stati seguiti duecriteri (FROHN, 1998): il loro contenuto doveva a)fornire informazioni sulle caratteristiche di area, for-ma, frammentazione e densità degli habitat e b) pre-sentare un basso grado di ridondanza.
Successivamente la mappa degli habitat dell’area distudio è stata sovrapposta al modello digitale del terre-no (DEM) allo scopo di suddividerla in tre fascealtitudinali (planiziale, collinare, montana). Sono statescelte le quote altimetriche di 400 e 800 metri, persuddividere rispettivamente la zona di pianura da quelladi collina e quest’ultima dalla regione montana. Talesuddivisione risponde ad una ben precisa zonazionefitoclimatica (ROSSI, 1999): oltre i 400 m è presente lafascia dei querceti a roverella e dei boschi a carpinonero, mentre oltre gli 800 m dominano incontrastate lefaggete. Dal DEM sono state ricavate anche la mappadelle pendenze e quella dei versanti, allo scopo diottenere ulteriori informazioni sulla distribuzione spa-ziale dei biotopi CORINE mappati. Per ognuna delletre fasce altimetriche considerate sono stati calcolatigli indici descritti in tabella 2. L’indice di distanzamedia dal vicino più prossimo (DMVP) fornisce infor-mazioni sul grado di frammentazione a livello A o B.Bassi valori di DMVP indicano che gli habitat tendonoad avere nell’intorno più prossimo un vicino apparte-nente alla stessa classe (basso grado di frammentazio-ne); all’opposto se i valori assunti dall’indice sonoelevati. La DMVP influenza molti importanti processidel paesaggio e gioca un ruolo fondamentale nellateoria della biogeografia delle isole (MACARTHUR eWILSON, 1967) e delle metapopolazioni (LEVINS, 1970).La distanza tra gli habitat ha avuto un ruolo fondamen-tale anche nei recenti sforzi di conservazione di specieminacciate (LAMBERSON et al., 1992). L’indice di di-spersione (ID) descrive il grado di “mescolanza” tra glihabitat: assume valori bassi se esiste una polarizzazio-ne dei confini, ossia se una certa tipologia di habitattende a confinare in modo preferenziale con un’altra;in caso opposto assume valori prossimi a 100.
L’indice di uguaglianza di Shannon (IS) descrive ilgrado di dominanza di una certa tipologia di habitat:assume valori prossimi a 0 quando esiste una classedominante (in termini di area occupata) e valori prossi-mi a 1 se l’area di studio è distribuita in modo presso-ché uguale tra le diverse tipologie. La densità di habitat
ClasseMIVIS
CodiceCORINE
Tipologia di Habitat
abete bianco 42.1B1 Rimboschimenti ad abete bianco
22.1 Corpi d'acqua (laghi, bacini)acqua
24.1 Corsi d'acqua
carpino nero 41.813 Boschi di carpino nero submontani
cerro 41.74 Boschi di cerro
faggio 41.1744 Faggete neutrofile
pino nero 42.67 Rimboschimenti di pino nero
pioppo 44.614 Pioppeti ripariali
robinia 83.3234 Formazioni di Robinia pseudoacacia
roverella 41.7312 Boschi di roverella
34.3266 Praterie semiaride appenniniche
38.13 Prati abbandonati
38.2 Prati da sfalcio submontani e montani
36.334 Praterie a brachipodio
prato
82.11 Seminativi di foraggere
86.2 Aree urbanizzate, piccoli agglomerati
62.4 Affioramenti litoidi e suolo nudolitoide
82.11 Seminativi arati di foraggere
Tab. 1 Corrispondenza tra le classi di habitat riconosciute nelbacino idrografico del torrente Baganza mediante il sensore MIVISe i codici delle tipologie di habitat CORINE Biotopes dell’UnioneEuropea.

ROSSI P. et al. - Analisi del paesaggio mediante dati telerilevati 59
(DH) quantifica l’abbondanza di habitat per unità disuperficie a livello A o B. Un paesaggio con unamaggiore densità di habitat presenta una maggioreeterogeneità spaziale, in grado di influenzare un nume-ro elevato di processi ecologici tra cui le interazioni trasub-popolazioni di una certa metapopolazione (GILPINe HANSKI, 1991) e la propagazione del disturbo (fuoco,malattie) nel paesaggio: ad un grado maggiore di DH,corrisponde una minore probabilità di propagazione deldisturbo (FRANKLIN e FORMAN, 1987). L’indice HMD(habitat di maggiori dimensioni) evidenzia la presenzao meno di un habitat di grandi dimensioni, a livello A oB. Valori elevati di HMD evidenziano grande omoge-neità spaziale con un habitat che tende ad occupareuna percentuale elevata dell’area di studio. Questo tipodi situazione tende a facilitare i flussi di materia edenergia (vettori biotici ed abiotici) nel paesaggio, per-chè all’aumentare di HMD diminuiscono le aree ditransizione tra gli habitat. L’indice frattale (IF) tiene
conto della complessità della forma degli habitat cheinfluenza alcuni processi ecologici come la migrazionedei mammiferi (BUECHNER, 1989), la colonizzazione daparte di piante (HARDT e FORMAN, 1989) e le strategiedi foraggiamento degli erbivori (FORMAN e GODRON,1986). L’indice IF è stato calcolato (a livello B) comevalore medio di IF di tutti gli habitat appartenenti aduna certa tipologia. Il grado di complessità della formaviene spesso usato nella letteratura scientifica comeindicatore del livello di pressione antropica agente suglihabitat: la presenza dell’uomo tende a semplificare leforme abbassando il valore di IF verso il valore unita-rio, all’opposto un alto grado di naturalità è evidenziatoda valori di IF prossimi a 2.
RISULTATIL’intera area di studio occupa una superficie di
17705 ettari di cui il 35,09% a prato (prati naturali eagricoli), il 20,57% a bosco di carpino nero, il 16,59%
livello: paesaggio
Nome Sigla Range divalori
Unità dimisura Descrizione
1 Habitat di maggiori dimensioni HMD da 0 a 100 percentuale rappresenta il rapporto tra l'area dell'habitat di maggiori dimensionie l'area totale di studio
2 Indice di dispersione ID da 0 a 100 percentualeassume valori prossimi a 100 quando ogni tipologia di habitat tendea confinare in modo uguale con ogni altra tipologia. Tende a 0 incaso opposto
3 Indice di uguaglianza diShannon IS da 0 a 1 nessuna
è pari a 0 quando il paesaggio contiene un solo habitat e cresceverso 1 all'aumentare del numero di habitat e di una ugualedistribuzione degli habitat dentro ogni tipologia
4 Densità di habitat DH da 0 in poi nessuna rappresenta il numero di habitat per ogni ettaro di area di studio
5 Distanza media dal vicinoprossimo DMVP da 0 in poi metri
rappresenta la distanza media di un habitat dal vicino più prossimoappartenente alla stessa classe. Il valore viene mediato su tutti glihabitat del paesaggio
livello: tipologie di habitat (classe)
Nome Sigla Range divalori
Unità dimisura Descrizione
1 Habitat di maggiori dimensioni HMD da 0 a 100 percentuale rappresenta il rapporto tra l'area dell'habitat di maggiori dimensionidi una certa classe e l'area totale di studio
2 Indice di dispersione ID da 0 a 100 percentualeassume valori prossimi a 100 quando una classe tende a confinarein modo uguale con ogni altra tipologia. Tende a 0 nel casoopposto
3 Densità di habitat DH da 0 in poi nessunarappresenta il numero di habitat appartenenti ad
una certa classe per ogni ettaro di area di studio
4 Indice frattale IF da 1 a 2 nessunaassume valori prossimi a 1 per habitat con forme semplici (circolario quadrate) e cresce fino a 2 per habitat con forme complesse. Ilvalore viene mediato per tutti gli habitat di una certa classe
5 Distanza media dal vicinoprossimo DMVP da 0 in poi metri
rappresenta la distanza media di un habitat dal vicino più prossimoappartenente alla stessa classe. Il valore viene mediato su tutti glihabitat di una certa classe
Tab. 2. Sono elencati gli indici, a livello di paesaggio e di singole tipologie di habitat, calcolati utilizzando il software FRAGSTAT sulleclassificazioni realizzate con il sensore MIVIS.

ROSSI P. et al. - Analisi del paesaggio mediante dati telerilevati60
a litoide (urbanizzato, suolo nudo e affioramenti roc-ciosi), il 15,02% a bosco di faggio, il 5,08% a roverellaed il 4,09% a cerro. Percentuali minori della valle sonooccupate da pinete (1,71%), pioppeti (1,07%), boschiad abete (0,61%), robinieti (0,15%) e acque lentiche olotiche (0,01%). Le acque lentiche sono rappresentateda alcuni laghetti appenninici di modeste dimensioni,mentre il basso valore di superficie occupata dalleacque lotiche dipende dalla scarsità d’acqua del gretodel Torrente Baganza (e dei suoi affluenti) durante imesi estivi (periodo della ripresa con il sensore MI-VIS).
Le aree boscate ammontano quindi al 48,3% dell’in-tera area di studio (8551 ettari) di cui circa la metà acarpino nero. L’elevatissima risoluzione spaziale delsensore MIVIS ha permesso di riconoscere un nume-ro molto elevato di habitat (47088) con una densitàcomplessiva di 2,65 habitat per ettaro. L’intero bacinopresenta un indice di dispersione pari 63,26, ovvero letipologie di habitat (classi) risultano tra loro ben “me-scolate” con rari casi di isolamento geografico rappre-sentati dagli habitat a pioppo (ID = 42,44). Il valoreelevato dell’indice di uguaglianza di Shannon (0,72)indica una ripartizione non polarizzata dell’area di stu-dio con 4 classi (prato, carpino, faggio e litoide),ognuna delle quali occupa più del 15% del bacino. Inmedia ogni habitat della valle dista 99,1 m dal suovicino più prossimo appartenente alla stessa classe, matale valore risulta poco significativo poiché varia mol-tissimo a seconda della classe che si considera: assu-me valore di 70,76 m per gli habitat a prato, di 77,79 mper i carpineti, di 81,07 m per le faggete e sale fino a827,31 m per i boschi a robinia. Il valore risultante di2742,63 m per gli habitat acquatici è scarsamentesignificativo per via delle già citate condizioni di scarsi-tà d’acqua del bacino idrografico al momento dellaripresa. Le classi che presentano habitat ininterrotti dimaggiori dimensioni sono quella dei boschi di faggionella fascia montana (6% dell’intera superficie dellavalle), dei boschi di carpino (3% dell’intero bacino)nella fascia collinare e dei prati (3%) nella zona dipianura.
La zona di pianura è costituita per più del 53% daprati (quasi esclusivamente di tipo agricolo) e per il30% circa da aree a litoide (urbanizzato e seminativiarati). Le aree boschive occupano circa il 17% dellafascia planiziale con dominanza di roverella (6,17%),di carpino (5,3%) e pioppo (4,23%). In misura minoresono presenti boschi di origine antropogenica qualipinete (0,51%) e robinieti (0,49%). La zona collinare èancora dominata dalle aree prative (36,68%, principal-mente agricole, ma con presenza anche di numerosiprati abbandonati o adibiti a pascolo). Seguono, intermini di importanza, la classe dei boschi di carpino
(30,26%) e quella del litoide (15,56%). Ben rappresen-tate sono anche le classi dei boschi di roverella (8,94%)e di cerro (5,83%). In misura esigua sono presentiboschi di pino nero (1,38%), faggio (0,92%), abete(0,27%), robinia (0,08%) e pioppo (0,08%). La fasciamontana è dominata dalle faggete (36,55%) e dallearee a prato (22,19%) prevalentemente di tipo natura-le. I boschi a carpino rappresentano il 21% dellasuperficie totale di questa fascia, mentre il litoide (pre-valentemente sotto forma di affioramenti rocciosi)occupa poco meno del 9%. Sono presenti, in misurainferiore, boschi a cerro (5%), pino nero (2,79%),roverella (2,18%) e abete (1,31%). La classe acqua(rappresentata da piccoli laghetti appenninici e pre-appenninici) occupa appena lo 0,03% della fascia mon-tana.
Il confronto tra gli indici calcolati a livello di paesag-gio e delle tre fasce di pianura (4337 ettari), collina(6533 ettari) e montagna (6835 ettari) è riassunto infigura 2. L’indice HMD assume il valore dell’11% perla fascia di pianura, dell’8% per la fascia di collina e del16% per la fascia di montagna. I valori delle zone dipianura e di collina sono attribuibili a due habitat adibitiad uso agricolo, mentre quello della fascia montana adun habitat di faggio di quasi 1100 ettari, situato oltre i1000 metri. La densità di habitat è massima in collina(2,92 habitat/ettaro) e minima nella zona di pianura(DH pari a 2,49) dove sono presenti campi agricoli digrandi dimensioni. L’indice ID aumenta dalla pianura(48,64) in direzione della montagna (64,78), all’oppo-sto la DMVP è massima in pianura (134,8 m) e similenelle fasce di collina (114,3 m) e di montagna (114,6m). L’indice di Shannon mostra un andamento chiara-mente crescente nel passare dalla pianura (0,55) allamontagna (0,68). I cinque indici considerati risultanoben differenziati per fasce altimetriche supportando lasuddivisione dell’intero bacino secondo il gradientealtimetrico. Inoltre tre (IS, ID, DMVP) dei cinqueindicatori hanno un andamento marcato crescente odecrescente passando dalla pianura alla montagna.
Gli indicatori calcolati a livello di classe sono statianalizzati mediante la tecnica delle componenti princi-pali (PCA; Fig. 3, 4, 5 e 6) per facilitarne l’interpreta-zione. Se si considera l’intera area di studio (Fig. 3), leprime due componenti riassumono più del 90% dellavarianza iniziale. Sul primo asse (correlato positiva-mente con gli indici ID e DMVP e negativamente congli indici DH, IF e HMD) vengono distinte le classifaggio (valori negativi) e acqua (valori positivi). Lefaggete presentano, in effetti, valori elevati dell’IF(1,53) e dell’indice HMD (6%), mentre la classe acquaha un valore molto elevato di DMVP (2742 m) cherappresenta la distanza tra i laghetti nella fascia monta-na e pedemontana. Le classi cerro e roverella hanno

ROSSI P. et al. - Analisi del paesaggio mediante dati telerilevati 61
valori quasi uguali su entrambe le componenti a indica-re una forte somiglianza di quasi tutti gli indici presi inconsiderazione. Il punteggio elevato della classe robi-nia sulla prima componente è ascrivibile principalmen-te al valore elevato di DMVP (827 m), mentre ilpunteggio basso della classe a prato sulla seconda ècausato sia da valori elevati sia di DH (88,69) che di ID(69,78).
La PCA delle classi di pianura (Fig. 4) riassume
sulle prime due componenti l’88,4% dell’informazionecontenuta negli indici calcolati. Sul primo asse vieneseparata la classe robinia che, pur avendo un IF basso(1,17), presenta valori molto elevati di ID (67,29) e diDMVP (785,3 m). Le classi prato e litoide si distinguo-no per punteggi molto bassi sul primo asse dovuti avalori molto elevati sia di DH (107,12 per i prati e 75,6per il litoide) che di HMD (11% per i prati e 7% per illitoide). Le posizioni ravvicinate nel piano delle compo-
0,04
0,06
0,08
0,10
0,12
0,14
0,16
0,18
pianura collina montagna bacino
HM
D
2,4
2,5
2,6
2,7
2,8
2,9
3
pianura collina montagna bacino
DH
46
48
5052
54
56
58
6062
64
66
pianura collina montagna bacino
ID
0,540,560,58
0,60
0,62
0,640,66
0,68
0,700,72
0,74
pianura collina montagna bacino
IS
90
100
110
120
130
140
pianura collina montagna bacino
DM
VP
HMD = habitat di maggiori dimensioni
DH = densità di habitat
ID = indice di dispersione
DMVP = distanza minima dal vicino più
prossimo
IS = indice di Shannon
Fig. 2. Grafici relativi ai 5 indici del paesaggio calcolati sia a livello dell’intero bacino idrografico del Torrente Baganza che delle tre fascealtitudinali (pianura, collina, montagna) considerate.
ROSSI P. et al. - Analisi del paesaggio mediante dati telerilevati62
nenti delle classi carpino, roverella e pioppo evidenzia-no la forte somiglianza per i 5 indici considerati. Laclasse a pino si stacca da queste tre classi per i valoribassi di DH (2,31) e di HMD (40,88).
Le classi della fascia di collina risultano ben distintesia sulla prima che sulla seconda PCA (Fig. 5). Pioppe-ti e robinieti assumono punteggi elevati sulla primacomponente dovuti a valori molto alti di DMVP(1831,15 m e 1443,27 m rispettivamente), mentre i
punteggi negativi di carpineti e prati sono dovuti avalori elevati di IF (1,42 e 1,4 rispettivamente) e di DH(78,37 e 100,38 rispettivamente).
L’analisi delle PCA per la fascia di montagna (Fig.6) mette in evidenza le caratteristiche peculiari delleclassi acqua e faggio, che assumono valori estremisulla prima componente. Il valore elevato di DMVP(2742,6 m) per la classe acqua giustifica il valoreelevato positivo sul primo asse, mentre i valori elevatidi IF (1,53) e di HMD (16%) della classe faggio nedeterminano il valore negativo. La classe prato si ca-ratterizza per il punteggio elevato sulla seconda com-ponente, risultato che è determinato principalmente dalvalore elevato di ID (70,07). I prati della fascia dimontagna sono quasi esclusivamente prati naturali con
Fig. 3. Analisi delle componenti principali (calcolate sulla matricedi correlazione) degli indici relativi alle tipologie di habitat ditutto il bacino del torrente Baganza. Sono rappresentate la prima(59,4% della varianza iniziale) e la seconda componente (30,2%della varianza iniziale). Di ogni componente vengono fornite lecorrelazioni (solo quelle significative) con le variabili di partenzaall’interno dei box rettangolari. Per esempio, valori alti sulla primacomponente sono attribuibili alle classi con valori elevati dell’indiceDMVP e dell’indice ID mentre punteggi bassi sono dovuti avalori elevati degli indici DH, IF e HMD.
Fig. 5. Analisi delle componenti principali (calcolate sulla matricedi correlazione) degli indici relativi alle tipologie di habitat dellazona di collina. Sono rappresentate la prima (58,3% della varianzainiziale) e la seconda componente (26,8% della varianza iniziale).
Fig. 4. Analisi delle componenti principali (calcolate sulla matricedi correlazione) degli indici relativi alle tipologie di habitat dellazona di pianura. Sono rappresentate la prima (63,5% della varianzainiziale) e la seconda componente (24,9% della varianza iniziale).
Fig. 6. Analisi delle componenti principali (calcolate sulla matricedi correlazione) degli indici relativi alle tipologie di habitat dellazona di montagna. Sono rappresentate la prima (57,3% dellavarianza iniziale) e la seconda componente (27,8% della varianzainiziale).

ROSSI P. et al. - Analisi del paesaggio mediante dati telerilevati 63
un valore di IF (1,42) superiore a quello della classecarpino (1,37). Queste due classi presentano valoripraticamente uguali sul primo asse poiché molto similiper tutti e cinque gli indicatori considerati. La classeprato presenta una complessità media delle forme (in-dice IF) nettamente crescente andando dalla pianura(1,15), alla collina (1,38) fino alla fascia montana(1,42). Il trend è giustificato dal fatto che i prati inpianura sono fondamentalmente superfici agricole conforme semplificate tipiche degli habitat antropogenici,mentre in collina e montagna sono fondamentalmenteabbandonati o adibiti al pascolo.
DISCUSSIONELe emergenze vegetazionali nell’area di studio sono
espressione del marcato gradiente altitudinale e morfo-logico che caratterizza la valle. La sua forma stretta edallungata determina la presenza di precise zonazionivegetazionali:− area planiziale, da Parma fino alle prime colline;− area submediterranea, che comprende la fascia col-
linare dai 400 agli 800 m;− area submontana e montana al di sopra degli 800 m.
La zona pianeggiante è dominata dall’attività agrico-la e la vegetazione naturale è limitata alle aree soggettea minor pressione antropica, in prossimità del greto delTorrente Baganza, anche se, per questo, non menodisturbate da altre sorgenti di stress, come i periodi dimagra e di piena dei corsi d’acqua. Le limitate zoneboscate sono sostanzialmente rappresentate da boschidi pioppo nero (Populus nigra) e robinia (Robiniapseudoacacia). Nell’area collinare la vegetazione fore-stale è costituita dai boschi misti dove le specie arboreepiù abbondanti sono le querce e i carpini. Tra le quercedomina la roverella (Quercus pubescens), che peròperde il suo valore di indicatore di “versante caldo earido”, trovandosi sia in condizioni xerofile che se-mixerofile. Condizioni di carattere più mesofilo favori-scono il carpino nero (Ostrya carpinifolia). L’areasubmontana vede la forte prevalenza di cerro (Quercuscerris) soprattutto sui versanti meno acclivi. La rove-rella è quasi scomparsa e le aree prative sono inprevalenza prati permanenti ancora sfalciati (arrenate-reti). Infine, la fisionomia dell’area montana è caratte-rizzata dal bosco neutrofilo a faggio (Fagus sylvatica),da praterie arbustate e pascoli a brachipodio. La pre-senza di rimboschimenti è legata ad essenze comel’abete bianco (Abies alba) e il pino nero (Pinus ni-gra).
Nel suo complesso, il bacino idrografico del torren-te Baganza presenta una struttura paesistica con tresottostrutture annidate e ben distinte. Il gradiente alti-metrico origina e mantiene questo pattern strutturalesia perché influenza la presenza dell’uomo sia perché
determina differenze climatiche e geomorfologiche. Lafascia planiziale presenta una struttura molto omoge-nea, dominata da habitat antropizzati di tipo agricolo eurbano che si interfacciamo quasi esclusivamente traloro, determinando un grado molto basso di mescolan-za tra gli habitat (ID = 48,64). La copertura boschivaplaniziale ha natura prevalentamente antropogenica cometestimoniato dai bassi valori di IF delle classi a robinia,pioppo e pino nero (1,17, 1,25 e 1,28 rispettivamente).Le tipologie a carpino nero e a roverella presentanovalori frattali superiori (1,33 e 1,38 rispettivamente)dovuti al loro posizionamento ad altitutidini intorno ai300 m dove il disturbo antropico è inferiore. Le tipolo-gie a pioppo e pino nero risultano estremamente confi-nate (ID pari a 45,75 e 40,88), la prima lungo il gretodel torrente e la seconda sotto forma di rimboschimen-ti isolati di natura artificiale. La fascia planiziale puòessere quindi definita come un paesaggio “culturale”dove le strutture sono semplici, i confini tra gli habitattendenzialmente lineari e il mosaico ambientale, lungidall’essere caotico, è completamente determinato dallapresenza dell’uomo.
La fascia di collina è la più eterogenea delle tre fascedal punto di vista strutturale. Essa presenta la più altadensità di habitat e il più basso valore di habitat dimaggiori dimensioni, causati dalla mancanza di unatipologia dominante (come invece accade per i prati inpianura e le faggete in montagna). Tale fascia presentaanche il maggior numero di tipologie di uso del suolo(10, rispetto a 9 della zona di montagna e 7 della fasciaplaniziale). Le tipologie di habitat sono maggiormenteframmiste (ID pari a 57,93) e la superficie totale èmeglio ripartita tra le classi (IS = 0,66) rispetto allafascia di pianura. I boschi hanno mediamente formepiù complesse, indice di un maggior grado di naturalitàrispetto alla pianura, se si eccettuano naturalmente irimboschimenti antropogenici ad abete (IF = 1,17) e lepiantagioni a robinia (IF =1,17). Nel suo complesso, lafascia collinare ha una tessitura a grana fine, risultantedal concorso di forze antropiche e naturali entrambemotrici del cambiamento del mosaico ambientale. Pernumero e tipologia di classi presenti, la fascia collinaretende ad assomigliare alla zona montana e i processisocio-demografici lasciano prevedere una tendenzadirezionale verso la fascia montana piuttosto che versoquella planiziale. Questa ipotesi è supportata anchedalle similarità delle caratteristiche geomorfologiche.
La fascia montana ha carattere di forte naturalitàcon coperture prevalentemente boschive scarsamenteinfluenzate dalla presenza dell’uomo. L’intervento an-tropico nell’area delle faggete si realizza quasi esclusi-vamente nella fruizione turistica del bosco. La produ-zione di legname è limitata all’uso privato e comunquenormato sopra i 1200 metri (L.R. 8 agosto 1985).

ROSSI P. et al. - Analisi del paesaggio mediante dati telerilevati64
Questo favorisce una situazione di alto fusto e unacondizione di naturalità del bosco. Le tipologie di habi-tat sono molto frammiste (ID = 64,78), soprattutto iprati, costituiti principalmente da coperture erbaceenaturali. Le tipologie di habitat che sono in comunecon la fascia di collina presentano maggiore comples-sità di forma e maggiori valori di dispersione. La fasciamontana è costituita quindi da un ecomosaico com-plesso, con strutture e forme risultanti dal concorso dipiù cause (clima, pedologia, geomorfologia) in cui,probabilmente, ogni struttura è il risultato di micro-strutture a scale ancora più fini rispetto a quella utiliz-zata per gli scopi del presente lavoro. Le forme deglihabitat sono più convolute e più allungate (soprattuttose in concomitanza con le maggiori pendenze) e anchela variabilità di forme è maggiore. Le tendenze demo-grafiche all’invecchiamento e allo spopolamento diquesta parte della valle favoriscono i processi di rina-turalizzazione e lasciano prevedere un’evoluzione futu-ra, su tempi lunghi, verso pattern spaziali ancora piùcomplessi.
L’analisi delle PCA condotta sulle singole fascealtitudinali ha messo ulteriormente in evidenza la diffe-rente complessità del mosaico ambientale. Il metodo diFrontier (LEGENDRE e LEGENDRE, 1998) per le PCAindividua due soglie di significatività per la prima e laseconda componente, rispettivamente 45,67% e25,68% della varianza complessiva. Per la fascia plani-ziale, la prima componente spiega il 63,5% della va-rianza iniziale e la seconda (non significativa) solo il24,9%. Nelle altre due fasce, entrambe le componentirisultano invece significative. Questo risultato indicauna maggiore semplicità strutturale del mosaico dipianura, le cui caratteristiche sono riconducibili ad unsolo asse significativo.
La mappatura degli habitat utilizzata per il presentelavoro proviene dalla classificazione di immagini teleri-levate del sensore MIVIS. Le caratteristiche del senso-re sono di notevole utilità per il riconoscimento dibiotopi CORINE: l’elevata risoluzione spaziale (4 me-tri) ha permesso di riconoscere habitat anche di di-mensioni minime, mentre la risoluzione spettrale (102canali) è talmente elevata da consentire la discrimina-zione molto accurata delle tipologie di habitat presentinell’area di studio. Sono stati fatti tentativi per ricono-scere gli habitat dell’area di studio anche mediante ilsatellite Landsat, ma ci si è scontrati con due difficoltà:a) i bassi livelli di accuratezza raggiunti (intorno al76%) e b) l’impossibilità di cogliere in modo appro-priato ed efficace la notevole eterogeneità del patterndel mosaico ambientale della fascia di collina.
L’accorpamento dei codici CORINE relativi alla
tipologia prato, in un’unica classe, è dipeso dalla diffi-coltà di distinguere alcuni biotopi erbacei sulla solabase dell’informazione spettrale. La scelta di accorpa-re i codici CORINE 62.4, 86.2 e 82.11 nell’unicaclasse del litoide è motivabile sia alla luce del compor-tamento spettrale simile di queste superfici che delloscarso interesse nell’analizzarli come singole tipologiedi habitat. L’utilizzo del sistema CORINE Biotopes hapermesso un approccio standardizzato alla classifica-zione del paesaggio, con un’attenzione particolare neiconfronti degli habitat di tipo naturale e semi-naturale.
CONCLUSIONIUtilizzando i dati telerilevati del sensore MIVIS sono
stati riconosciuti 47088 habitat presenti nel bacinoidrografico del torrente Baganza. Sulla mappatura ri-sultante sono stati calcolati diversi indici del paesaggio,la cui analisi ha permesso di trarre conclusioni suiprincipali aspetti strutturali dell’ecomosaico ambienta-le del bacino idrografico del torrente Baganza. Utiliz-zando il modello digitale del terreno, l’area di studio èstata divisa in tre fasce altitudinali (pianura, collina emontagna) ognuna delle quali è stata analizzata sia alivello di paesaggio che di singole tipologie di habitat.La Val Baganza ha mostrato un marcato gradientealtitudinale, spiegabile sia alla luce della variazione delgrado di pressione antropica che delle condizioni cli-matiche e geomorfologiche. Lungo tale gradiente sisostituiscono classi di habitat, mentre le tipologie incomune a tutte le fasce altitudinali modificano le lorocaratteristiche in base all’uso reale che ne viene fatto; ilcaso tipico riguarda la classe prato, di natura principal-mente antropica in pianura, naturale in montagna. Lafascia planiziale e quella montana mostrano attributiquasi opposti in termini di densità, forma, connettivitàe distribuzione degli habitat. La fascia collinare mostracaratteristiche strutturali più simili a quelle della mon-tagna che della pianura, sia per motivi legati ad aspettigeomorfologici che per i processi di spopolamento edinvecchiamento della popolazione nei centri abitati.
Il sensore iperspettrale MIVIS ha permesso di co-gliere con accuratezza la grande eterogeneità delladistribuzione spaziale degli habitat (differentemente daquanto ottenuto con il satellite Landsat). L’utilizzo delsistema di classificazione CORINE Biotopes, infine,ha assicurato un approccio standard alla classificazio-ne del mosaico ambientale oggetto di studio.
RINGRAZIAMENTISi ringraziano i referee anonimi che hanno contribuito al migliora-mento della forma e del contenuto del presente lavoro.

ROSSI P. et al. - Analisi del paesaggio mediante dati telerilevati 65
BUECHNER M., 1989. Are small-scale landscape features impor-tant factors for field studies of small mammal dispersal sinks?Landscape Ecology, 2: 191-199.
C.E.C. (Commission of European Community), 1991. CORINEBiotopes Manual, habitats of the European Community. Amethod to identify and describe consistently sites of majorimportance for nature conservation. EUR 12587/3, Brux-elles, 324 pp.
FERRARINI A., ROSSI P., ZACCARELLI N., 2001. Classificazionivegetazionali ad elevata accuratezza: la metodologia delle retineurali applicata ai dati iperspettrali MIVIS. Rivista Italianadi Telerilevamento, 22: 13-22.
FORMAN, R.T.T., GODRON M., 1986. Landscape ecology. NewYork, John Wiley & Sons, 619 pp.
FRANKLIN, J.F., FORMAN R.T.T., 1987. Creating landscape pat-tern by forest cutting: ecological consequences and princi-ples. Landscape Ecology, 1: 5-18.
FROHN R.C., 1998. Remote sensing for landscape ecology, newmetric indicators for monitoring, modeling and assessment ofecosystems. Lewis Publishers, Boca Raton, Boston, London,New York, Washington D.C., 287 pp.
GILPIN M.E., HANSKI I. (eds.), 1991. Metapopulation dynamics:empirical and theoretical investigations, San Diego, Academ-ic Press, 336 pp.
HALL F.G., STREBEL D.E. e SELLERS P.J., 1988. Linking knowledgeamong spatial and temporal scales: vegetation, atmosphere,climate and remote sensing. Landscape Ecology, 2: 3-22.
HARDT R.A., FORMAN, R.T.T., 1989. Boundary form effects onwoody colonization of reclaimed surface mines. Ecology, 70:1252-1260.
JOHNSTON C.A. & BONDE J., 1989. Quantitative analysis of eco-tones using a GIS. Photogrammetric Engineering and RemoteSensing, 55: 1643-1647.
LAMBERSON R.H., MCKELVEY R., NOON B.R., VOSS C., 1992. Adynamic analysis of northern spotted owl viability in afragmented forest landscape. Conservation Biology, 6: 1-8.
LEGENDRE P. & LEGENDRE L., 1998. Numerical ecology. 2ndEnglish edition. Elsevier Science BV, Amsterdam, 853 pp.
LEVINS, R., 1970. Extinctions. In: Gertenhaber M. (eds.), Somemathematical questions in biology. Vol. 2. Providence, RI:American Mathematical Society: 77-107.
MACARTHUR R.H., WILSON E.O., 1967. The theory of islandbiogeography. Princeton, Princeton University Press, 203pp.
MCGARIGAL K., MARKS B. J., 1995. FRAGSTATS: spatial pat-tern analysis program for quantifying landscape structure.Gen. Tech. Rep. PNW-GTR-351. Portland, OR: U.S. De-partment of Agriculture, Forest Service, Pacific NorthwestResearch Station, 122 pp.
NAVEH Z., LIEBERMAN A., 1994. Landscape ecology theory andapplication. Springer Verlag, New York, 256 pp.
PIGNATTI S., 1994. Ecologia del paesaggio. UTET, Milano,Italia, 236 pp.
RUMELHART D.E., MCCLELLAND J.L., 1986. Parallel distributedprocessing. MIT Press, Cambridge, MA, 458 pp.
RIPLEY B. D., 1981. Spatial statistics, John Wiley, New York,428 pp.
ROSSI P., 1999. Analisi della diversità vegetazionale mediante laclassificazione CORINE Biotopes. Tesi di Laurea in ScienzeBiologiche, Facoltà di Scienze Matematiche Fisiche e Natura-li, Università di Parma.
SCHREIBER K.F., 1990. The history of landscape ecology in Eu-rope, in: Zonneveld I.S. e Forman R.T.T. (eds.), Changinglandscapes: an ecological perspective, Springer-Verlag, NewYork, 314 pp.
SINGH A., 1989. Digital change detection techniques using Re-motely sensed Data. In: Eden M.J. and Parry J.T. (eds.),Remote Sensing and Tropical Land Management. John Wiley& Sons, London, 237-254.
TURNER M.G. e GARDNER R.H., 1991. Quantitative methods inlandscape ecology: the analysis and interpretation of land-scape heterogeneity, Springer-Verlag, New York, 528 pp.
BIBLIOGRAFIA


Biologia Ambientale, 17 (1): 67-75, 2003
Analisi dell’evoluzione del paesaggio mediante l’approcciogeostatistico ai dati telerilevati: il bacino del torrente Baganza(Parma)
Alessandro Ferrarini* e Pierfrancesca Rossi, Orazio RossiDipartimento di Scienze Ambientali, Università degli Studi di Parma, Viale delle Scienze 11/A - 43100 Parma
* Autore referente per la corrispondenza ([email protected])
Pervenuto il 26.7.2002; accettato il 25.11.2002
RiassuntoIl paesaggio è un mosaico complesso in continuo divenire, i cui cambiamenti originano dalla interazione tra i fenomeni naturali e le azionipianificate dall’uomo. Il territorio è formato da numerose componenti strutturali, ognuna con dinamiche proprie; l’analisi della suaevoluzione temporale deve considerare la natura di tutte le parti coinvolte, ma anche le loro specifiche frequenze, velocità e intensità dicambiamento nel tempo. Per tale motivo, la metodologia scientifica è importante almeno quanto la disponibilità di una informazionegeografica adeguata riferita a periodi temporali differenti.Nel presente lavoro, sono state utilizzate immagini satellitari Landsat riguardanti il bacino idrografico del torrente Baganza, riferite aperiodi di tempo differenti, per l’acquisizione di informazioni sui cambiamenti intercorsi in tale area. Per mezzo del modello digitale delterreno (DEM), il bacino idrografico del Baganza è stato suddiviso in tre fasce altitudinali per ognuna delle quali, utilizzando l’approcciogeostatistico, è stato possibile analizzare: a) le direzioni preferenziali del cambiamento; b) la scala spaziale alla quale è avvenuto; c) il gradodi omogeneità/eterogeneità spaziale delle trasformazioni; d) l’informazione persa per l’inadeguatezza della risoluzione spaziale delsatellite Landsat.Tale approccio è in grado di cogliere efficacemente gli aspetti principali delle dinamiche del territorio e di fornire ipotesi approfondite sulleforze che le determinano, rappresentando nel contempo una metodologia scientifica a basso costo e di notevole utilità, per tutti coloro chesi occupano di pianificazione e gestione territoriale.
PAROLE CHIAVE: paesaggio / dimensione temporale / geostatistica / satellite Landsat / telerilevamento
AbstractLandscape changes analysis through the geostatistic approach to remotely sensed dataLandscape is a dynamic phenomenon that almost continuously changes. The overall change of a landscape is the result of complex andinteracting natural and spontaneous processes and planned actions by man. Landscapes are formed by many components which all havetheir own dynamics of change. Therefore, studying change must consider the nature of all the components as well as their specificfrequency, pace and magnitude of change. To detect changes, the method to describe or measure the landscape features is important aswell as the availability of adequate and comparable information in map-like format for different time periods.In this work, two Landsat images covering the Baganza stream watershed have been used. As they are related to different dates, theyprovide information about changes that happened inside the study area. Using the digital terrain model (DEM), the whole watershed hasbeen divided into three altitudinal belts. Using the geostatistic approach a) the main directions of changes, b) their spatial scale, c) thedegree of spatial omogeneity/eterogeneity of landscape modifications, d) the degree of information loosed because of the insufficientsatellite resolution, have been analyzed. Our approach can enhance the main causes of landscape changes and it provides deep hypothesesabout their driving forces. In addition, it represents a low-cost scientific methodology, that is very useful to people planning andmanaging landscape.
KEY WORDS: landscape / temporal dimension / geostatistic / Landsat data / remote sensing

FERRARINI et al. - Analisi dell’evoluzione temporale del paesaggio68
INTRODUZIONEL’informazione sull’uso reale del territorio è fonda-
mentale per progetti di pianificazione e gestione am-bientale. Il paesaggio si modifica rapidamente nel tem-po in relazione a fattori economici, sociali e naturali.Per monitorare tali cambiamenti è necessario disporredi informazioni sinottiche sul territorio, rilevate a tempidifferenti, e il telerilevamento satellitare rappresenta lametodologia più adatta e più economica per acquisirle.Nello stesso tempo sono necessari anche algoritmiscientifici che sappiano cogliere in profondità i cam-biamenti intercorsi nel paesaggio, utilizzando al massi-mo l’informazione contenuta nelle immagini telerileva-te. Sono riportati in letteratura numerosi approcci al-l’analisi delle modifiche del territorio mediante il remo-te sensing. Confronti tra classificazioni del territoriorealizzate a tempi diversi, composizioni in falso colore,uso di tecniche statistiche quali le componenti princi-pali e algebra matriciale, sono solo alcune delle meto-dologie sperimentate (JENSEN, 1996; SINGH, 1989).Nessuna delle precedenti metodologie è però in gradodi rendere esplicito l’andamento spaziale del cambia-mento intercorso nel territorio, ovvero la scala spazialee le direzioni preferenziali alle quali è avvenuto. L’ana-lisi del trend spaziale delle modifiche del territorio puòanche suggerire ipotesi approfondite sulle cause, lavelocità e le direzioni preferenziali lungo cui i cambia-
menti intercorrono. Per tale motivo, nel presente lavo-ro si propone l’approccio geostatistico ai dati telerile-vati. La geostatistica è una branca della statistica cheanalizza la variazione spaziale dei dati da analizzare erisulta particolarmente utile quando la continuità/di-scontinuità spaziale di un fenomeno rappresenta unelemento determinante.
Lo scopo del lavoro è di proporre un algoritmoscientifico di validità generale in grado di cogliere inprofondità le proprietà emergenti delle dinamiche delterritorio.
AREA DI STUDIO, MATERIALE E METODIL’area di studio è il bacino idrografico del Torrente
Baganza (Prov. Parma) situato nel versante emilianodell’Appennino settentrionale. Il bacino occupa unasuperficie di 177,5 kmq, si estende da Parma (60 ms.l.m.) al monte Cervellino (1493 m s.l.m) e presentaun asse longitudinale con direzione 45 gradi (Fig. 1). Ilprofilo complessivo della Val Baganza è determinatoessenzialmente dal gradiente altitudinale (ROSSI P., 1999)che dà luogo alla formazione di differenti popolamentivegetali e ad una diversa presenza dell’uomo. Nellazona pianeggiante la vegetazione è limitata alle zone aminore impatto antropico, mentre nell’area collinare lavegetazione forestale è rappresentata da boschi di ca-ducifoglie a roverella e carpino nero e, solo in minor
Fig. 1. A sinistra, il bacino idrografico del torrente Baganza con, in evidenza, i principali centri abitati. A destra, una sintesi dellametodologia utilizzata nel presente lavoro.

FERRARINI et al. - Analisi dell’evoluzione temporale del paesaggio 69
parte, da cenosi boschive igrofile a pioppo. Diffusisono i rimboschimenti di pino nero, espressione del-l’antropizzazione del paesaggio. Nella zona submonta-na i boschi a cerro diventano i complessi forestali piùestesi, mentre in quella montana, a quote superiori ai900-1000 m s.l.m., dominano quelli a faggio.
Le due scene Landsat TM 5 utilizzate nel presentelavoro sono state riprese alle ore 9 del mattino dell’11agosto 1990 e del 24 luglio 1995 rispettivamente egeoreferenziate secondo la proiezione UTM (fuso 32Nord). La corrispondenza degli orari delle riprese e delperiodo dell’anno rivestono grande importanza nelleanalisi di variazione temporale. L’area occupata dalbacino del torrente Baganza è stata individuata sovrap-ponendo il file vettoriale dei confini della valle alle dueimmagini satellitari.
Le modifiche subite dal territorio nel corso dei 5anni in esame sono state studiate analizzando nelle 2scene Landsat le variazioni dell’indice NDVI (Norma-lized Difference Vegetation Index, ROUSE et al., 1974).Tale indice viene calcolato sulle immagini Landsatsecondo la formula:NDVI = (banda 4 – banda 3) / (banda 4 + banda 3).
Esso utilizza la risposta nei canali del rosso centratointorno a 0,66 μm (banda 3, regione di forte assorbi-mento della clorofilla) e del vicino infrarosso, intorno a0,86 μm (banda 4, regione di forte riflettanza dellostrato vegetativo). La risposta nei due canali è funzionedella specie vegetale, delle condizioni di salute o distress (disponibilità idrica e di nutrienti) (GUYOT, 1989),ed è fortemente correlata a variabili di fisiologia vege-tale come il fipar (frazione della radiazione fotosintetica-mente attiva intercettata dalla vegetazione) e il L.A.I.(Leaf Area Index) (LAW e WARING, 1994). In lineateorica, l’NDVI può assumere valori compresi tra -1 e+1. In pratica, l’intervallo si colloca tra +0,7 (vegeta-zione molto densa a latifoglie) e -0,4 (corpi d’acqua).Valori compresi tra 0,3 e 0,4 sono tipici delle aree aprato, mentre il litoide assume valori prossimi a zero. Irimboschimenti ad aghifoglie assumono valori pari acirca 0,5. Le variazioni dell’indice NDVI possonoessere causate da modifiche dell’uso reale del suolo(per esempio, le azioni di rimboschimento determinanoun forte aumento di NDVI, mentre la cementificazionederivante dall’espansione degli abitati ne causa unaforte diminuzione) o da variazioni dello stato fenologi-co della vegetazione. L’importanza dell’indice NDVIrisiede anche nella sua proprietà di minimizzare glieffetti atmosferici. Per i suoi caratteri di completezzapuò essere quindi utilizzato come vettore sintetico diinformazione per studi di modifiche temporali del pae-saggio (GRIFFITH et al., 2002; REMMEL e PERERA,2001; WALSH et al., 2001).
In un precedente lavoro (FERRARINI et al., 2000),
dopo le necessarie correzioni geometriche e spettrali siè proceduto a calcolare l’indice NDVI per entrambe leimmagini Landsat e quindi si è operata una differenzapixel per pixel tra le due. L’immagine risultante (DEL-TA) ha fornito l’informazione sui cambiamenti inter-corsi nell’area di studio tra le due date prese in esame.Tale informazione ha permesso di rilevare alcuni aspettifondamentali del dinamismo a medio e breve terminedel bacino come, per esempio, la grande stabilità dellazona a faggeta oltre i mille metri e il dinamismo antro-pico (agricoltura ed espansione dell’urbano) della parteplaniziale e collinare della valle.
Nel presente lavoro, si procede ad un’analisi moltopiù approfondita di DELTA, allo scopo di evidenziarela presenza di eventuali trend spaziali, nonché la scala el’intensità dei cambiamenti intercorsi tra le due date. Atale scopo, è stato scelto l’approccio geostatistico aidati telerilevati. DELTA è stata inserita all’interno delGIS ArcView e sovrapposta al modello digitale delterreno (DEM) dell’area di studio, allo scopo di suddi-viderla in tre fasce altitudinali (pianura, collina, monta-gna). Tali fasce sono state scelte perché soggette aduna differente pressione antropica (decrescente dallapianura alla montagna) ed in accordo alla loro diversacomposizione strutturale (geomorfologica e biologica)(ROSSI P., 1999). Per ogni fascia, DELTA è stataanalizzata mediante un semivariogramma (SV) isotro-po (che media le caratteristiche del cambiamento sututte le direzioni geografiche) e 8 semivariogrammi(SV) anisotropi che tengono conto della direzionalitàdei cambiamenti intercorsi. Il SV (Fig. 2) viene defini-to come:
γ(h) = 0,5*(Var(Z(u+h) - Z(h)))dove Z(u) è il valore assunto dalla variabile Z nellalocalizzazione u e Z(u+h) è il valore assunto nellalocalizzazione u+h, la funzione Var calcola la varianzadegli incrementi tra le coppie di osservazioni che dista-no h (valore detto lag). In accordo con MATHERON(1963), la formula del SV è:
γ(h) = )(*2))()(( 2
)(
hnuZuZ jihn∑ −
dove n(h) è l’insieme delle coppie di dati (ui, uj) chedistano h, mentre |n(h)| rappresenta il loro numero.Essendo h un vettore, il SV può essere calcolato lungodiverse direzioni dello spazio. Per un valore prefissatodell’angolo α, il SV misura la diversità dei valoriassunti dalla variabile Z al crescere della distanza tra lecoppie di punti. Al variare dell’angolo α, i SV corri-spondenti individuano le caratteristiche direzionali dellavariabile Z.
Per dati distribuiti irregolarmente può accadere cheesistano poche coppie di osservazioni distanti tra loro

FERRARINI et al. - Analisi dell’evoluzione temporale del paesaggio70
esattamente h, rendendo necessario l’utilizzo di unvalore di tolleranza sia per h che per l’angolo α. Per gliscopi del presente lavoro si è posta la tolleranza di αpari a 0, lag pari a 5 pixel e la tolleranza del lag pari a2,5 pixel: in questo modo vengono considerate nelcalcolo della semivarianza tutte e solo le coppie diosservazioni disposte lungo le 8 direzioni indagate.
Se il SV raggiunge un valore limite (detto sella)allora esiste una distanza (detta range) oltre la qualeZ(u) e Z(u+h) non sono più correlate, ovvero all’au-mentare di h la semivarianza non cambia. Per distanzeminori al valore di range, le coppie di valori tendono adessere tanto più simili quanto più sono spazialmenteravvicinate. L’informazione spaziale fornita dal rangepermette la comprensione della scala alla quale avvienela variazione della variabile Z. Valori bassi del rangeindicano che la varianza totale viene “spesa” in spazibrevi ovvero, preso un punto qualsiasi dell’area distudio, muovendosi nella direzione indagata di unadistanza pari al range si incontra tutta la variabilitàpresente nell’intera area in esame. All’opposto, valorialti indicano che le modifiche a livello locale non sonocomparabili con quelle a livello più globale. La sellarappresenta il totale di variazione (varianza) presentenei dati analizzati (CRESSIE, 1993). Per gli scopi delpresente lavoro, la sella evidenzia il grado di omogenei-tà/eterogeneità dei cambiamenti dell’indice NDVI nellediverse direzioni indagate (SV anisotropi). Bassi valoridi sella indicano che i valori di DELTA tendono adessere molto simili lungo una certa direzione (omoge-neità), all’opposto valori elevati indicano grandi diffe-renze nella variazione dell’indice NDVI (eterogeneità).
L’effetto nugget descrive il comportamento del SVvicino all’origine. Al tendere di h a 0, il valore assuntodal SV rappresenta l’informazione persa per variazioniche avvengono ad una scala inferiore a quella utilizzataper il campionamento (ISAAKS e SRIVASTAVA, 1989).Se infatti definiamo, secondo i precetti della teoriadell’informazione, la varianza (o semivarianza) dei daticampionati come l’informazione ad essi associata (avarianza nulla corrisponde informazione nulla), il nug-get rappresenta la varianza (o semivarianza) non coltaperché presente a scale inferiori a quelle campionabili.Risulta di particolare interesse il valore di nugget relati-vo (NR), definito come il rapporto tra il nugget e lasella. Il NR rappresenta il rapporto tra l’informazionepersa e quella effettivamente colta e, quindi, l’informa-zione spaziale persa per inadeguatezza del sensore. Pergli scopi del presente lavoro, valori elevati di NRevidenziano modifiche dell’indice NDVI avvenute pre-valentemente a scale sub-pixel (inferiore cioè a 30 m).All’opposto, valori bassi di NR indicano cambiamentidel valore di NDVI avvenute a scale comparabili osuperiori a quelle del satellite Landsat.
In accordo con JOURNEL e HUIJBREGTS (1978), sonostate rispettate le seguenti due regole: a) per ognivalore di h sono presenti almeno 30 coppie di punti; b)la distanza massima considerata per il calcolo del SV èminore della metà della massima distanza possibile tracoppie di punti. Le analisi esplorative condotte sui datidi partenza hanno evidenziato la presenza di alcuni,sebbene limitati, outliers spaziali, messi in evidenza daun coefficiente elevato di scostamento dalla normalitàgaussiana della distribuzione dei valori di NDVI. Per
Fig. 2. A sinistra sono schematizzate le principali caratteristiche di un semivariogramma. La sella rappresenta il valore massimo disemivarianza (asse delle ordinate). Il range è la distanza (asse delle ascisse) alla quale viene raggiunta la sella. Il nugget rapresenta il valoredi semivarianza quando il range tende a 0. Gli asterischi rappresentano il semivariogramma sperimentale ovvero quello calcolato sui datia disposizione. La linea curva rappresenta l’interpolazione (con il metodo dei minimi quadrati) del semivariogramma sperimentale. Adestra, sono indicate le 4 direzioni principali lungo cui possono essere calcolati i semivariogammi: Nord-Sud (NS, direzione 0 gradi), NordEst - Sud Ovest (NE-SO, 45 gradi), Est-Ovest (EO, 90 gradi) e Sud Est-Nord Ovest (SE-NO, 135 gradi).

FERRARINI et al. - Analisi dell’evoluzione temporale del paesaggio 71
tale motivo, si è fatto uso del SV calcolato secondo ilmetodo robusto proposto da CRESSIE e HAWKINS (1980):
))(
494.0457.0(*2
1)(
|)()(|)(
4
)(
5,0
hnhn
uZuZh hn
ji
+∗
⎟⎟⎟
⎠
⎞
⎜⎜⎜
⎝
⎛ −=∑
γ
Il metodo è particolarmente efficace quando, comenel presente caso di studio, la distribuzione dei dati èvicina alla normalità gaussiana nella parte centrale elontana da essa nelle code. Tale metodo, senza elimina-re alcuna delle osservazioni utilizzate per il calcolo delSV, rimuove quasi completamente l’effetto degli ou-tliers presenti nell’analisi ed è risultato dunque indi-spensabile per gli scopi del presente lavoro. L’alterna-tiva sarebbe stata la rimozione manuale degli outlierspresenti, lavoro oneroso e che avrebbe richiesto moltotempo per la loro individuazione tra decine di migliaiadi altre osservazioni.
La modellizzazione (fitting) dei SV isotropi delle trefasce altitudinali è avvenuta mediante il modello sferico:
( )rhperrhper
srh
rhsnh
>≤
⎪⎭
⎪⎬⎫
⎪⎩
⎪⎨⎧
−+= 3
3
*5.0*5.1*)(γ
dove s è il valore di sella, n il valore di nugget ed rquello di range. Tale equazione ha permesso di interpo-lare il SV sperimentale con precisione maggiore rispet-to ai modelli gaussiano ed esponenziale. Non è statonecessario interpolare i SV anisotropi poiché la lorofunzione si limita ad evidenziare la presenza di trenddirezionali nelle variazioni dell’indice NDVI.
RISULTATII SV anisotropi della fascia planiziale (0-400 m
s.l.m.) sono riportati in Fig. 3. Si nota che il loroandamento è complesso con una semivarianza chetende, in linea di massima, ad un valore limite (sella).Essi mostrano un andamento generalmente crescenteall’aumentare della distanza, con presenza di micro-strutture simil-sinusoidali con periodo di circa 50 pixel.Tali micro-strutture sono molto probabilmente ricon-ducibili a “oggetti” presenti nella fascia planiziale “inmodo periodico” e aventi dimensioni dell’ordine all’in-circa dei 25 pixel: presumibilmente si tratta di campicoltivati.
I SV riferiti alla direzione N-S (Nord-Sud, direzione0 gradi), N NE-S SO (Nord Nord Est-Sud Sud Ovest,22,5 gradi), NE-SO (45 gradi) e S SE-N NO (157,5gradi) sono piuttosto simili tra loro. I SV delle direzioni
Fig. 3. Semivariogrammi anisotropi sperimentali della fascia planiziale. Sull’asse delle ascisse la distanza è misurata in pixels Landsat (1pixel = 30 m). L’angolo varia di 22,5 gradi, comprendendo l’asse Nord-Sud (0 gradi, riquadro in basso a sinistra), quello Est-Ovest (90gradi, riquadro in alto a sinistra) e tutti quelli intermedi.
FERRARINI et al. - Analisi dell’evoluzione temporale del paesaggio72
Fig. 4. Semivariogrammi anisotropi sperimentali della fascia collinare. Sull’asse delle ascisse la distanza è misurata in pixels Landsat (1pixel = 30 m). L’angolo varia di 22,5 gradi, comprendendo l’asse Nord-Sud (0 gradi, riquadro in basso a sinistra), quello Est-Ovest (90gradi, riquadro in alto a sinistra) e tutti quelli intermedi.
Fig. 5. Semivariogrammi anisotropi sperimentali della fascia montana. Sull’asse delle ascisse la distanza è misurata in pixels Landsat (1pixel = 30 m). L’angolo varia di 22,5 gradi, comprendendo l’asse Nord-Sud (0 gradi, riquadro in basso a sinistra), quello Est-Ovest (90gradi, riquadro in alto a sinistra) e tutti quelli intermedi.

FERRARINI et al. - Analisi dell’evoluzione temporale del paesaggio 73
E NE-O SO (67,5 gradi), E-O (90 gradi) e SE-NO(135 gradi) sono simili tra loro con la distanza e lasemivarianza che sono correlate positivamente fino acirca 150 pixel, negativamente da 150 fino a 200 pixele di nuovo positivamente da 200 pixel in poi. Il SVdella direzione E SE-O NO (112,5 gradi) è simile ai treprecedenti con l’eccezione che manca della fase dirisalita della semivarianza oltre i 200 pixel. Questocomportamento è dovuto al fatto che, lungo quelladirezione per la particolare conformazione geograficadella valle, mancano coppie di punti che distano più di6 km. Il SV isotropo della fascia planiziale (Fig. 6)rappresenta l’andamento mediato sui singoli SV aniso-tropi. L’andamento è generalmente crescente con pre-senza di micro-strutture ad andamento simil-sinusoi-dale. Il range viene raggiunto rapidamente (60 pixelLandsat), la sella ha valore pari a 0,40 ed il nugget a0,12. Il NR è uguale a 0,3 (30%). Poiché i SV aniso-tropi mostrano 2 tipologie di andamenti ben distinti, ilSV isotropo non può essere considerato pienamenterappresentativo dell’andamento della variazione del-
l’indice NDVI della fascia planiziale. Esso riproduce ilvalore mediato di andamenti direzionali distinti. I SVanisotropi della fascia collinare (400-800 m s.l.m.; Fig.4) presentano andamenti regolari, con semivarianzageneralmente crescente all’aumentare della distanzatra le coppie di punti, presenza di sella e valori dinugget molto simili tra loro. I SV riferiti alla direzioneN-S (0 gradi), N NE-S SO (22,5 gradi), NE-SO (45gradi) e E NE - O SO (67,5 gradi) sono simili tra lorocon una correlazione tra la distanza e la semivarianzache passa da positiva a negativa verso i 200 pixel. Ledirezioni 22,5 e 45 gradi presentano valori di sellaevidentemente più elevati. I restanti SV hanno unacorrelazione sempre positiva tra distanza e semivarian-za, valori di sella ed effetto nugget praticamente uguali.Il SV isotropo della fascia collinare (Fig. 6) ha unandamento sostanzialmente crescente. Il range vieneraggiunto a 165 pixel, la sella ha valore pari a 0.16 ed ilnugget pari a 0,03. Il NR è uguale a 0,187 (18,7%). Inquesto caso, esso è rappresentativo della variazionedell’indice NDVI nella fascia collinare. I SV anisotropi
Fig. 6. Semivariogrammi isotropi sperimentali delle tre fasce (pianura, collina, montagna). La distanza sulle ascisse è espressa in pixelsLandsat (1 pixel = 30 metri). Nel riquadro in basso a destra sono mostrati i modelli teorici di tipo sferico che interpolano i semivariogrammisperimentali.

FERRARINI et al. - Analisi dell’evoluzione temporale del paesaggio74
della fascia montana (800-1500 metri; Fig. 5) hannoandamento regolare, con bassi valori sia di nugget chedi sella. La direzione 45 gradi fa eccezione per il valoreelevato di sella. Il range ha invece valori particolar-mente elevati in tutte le direzioni. Il relativo SV isotro-po (Fig. 6) ha un andamento generalmente crescente.Il range è raggiunto a 195 pixel, la sella ha valore pari a0,08 ed il nugget a 0,01. Il NR è uguale a 0,125(12,5%). Esso rappresenta molto bene la reale varia-zione dell’indice NDVI avvenuta nei 5 anni indagati.
DISCUSSIONENella fascia di pianura il motore del cambiamento
(identificabile con l’uomo) agisce in modo intenso intutte le direzioni, dando luogo ad una elevata eteroge-neità (alto valore di varianza totale) e producendovariazioni a scala locale (basso valore del range) tali darichiedere un campionamento più puntuale rispetto aquello del Landsat (alto valore del nugget). Le modifi-che dell’indice NDVI sono causate dalle rotazioni col-turali e dall’espansione/contrazione dell’urbanizzato edelle aree agricole. Tali modifiche vengono colte daiSV come un segnale caotico senza direzioni preferen-ziali. I cambiamenti avvengono ad una scala spazialeche richiederebbe un sensore con una migliore risolu-zione geometrica per essere apprezzata come, peresempio, lo scanner MIVIS (Daedalus AA5000 Multi-spectral Infrared and Visible Imaging Spectrometerdel Consiglio Nazionale Ricerche, Progetto L.A.R.A.-Laboratorio Aereo per Ricerche Ambientali).
Nella fascia di collina l’informazione spaziale persaper inadeguatezza del sensore è molto minore (NR 1,5volte inferiore rispetto a quello di pianura). Le modifi-che avvengono ad una scala molto più ampia rispetto aquella dell’area planiziale (valore di range 2,75 voltemaggiore). La diminuzione della pressione antropica èinfatti responsabile di cambiamenti che operano a scalameno locale. La presenza di una ampia superficieboscata (48% circa della fascia collinare) determinauna variazione dell’indice NDVI molto più omogeneadal punto di vista spaziale rispetto a quella della pianura(valore di sella 2,5 volte inferiore). L’analisi della dire-zionalità dei cambiamenti mostra caratteristiche so-stanzialmente uguali in tutte le direzioni, se si eccettuaquella a 45 gradi lungo la quale si ha una forte eteroge-neità. Su tale direzione si colloca l’asse longitudinaledella valle (Fig. 1) lungo il quale si concentra la granparte del disturbo antropico (costruzione di strade,espansione/contrazione degli abitati, pratiche agricole)che si traduce in una maggiore eterogeneità spazialedelle modifiche dell’NDVI.
Nella fascia montana, le modifiche dell’indice NDVIhanno una varianza totale notevolmente bassa lungotutte le direzioni, eccetto quella a 45 gradi che identi-
fica l’asse longitudinale della valle, lungo il quale sidispongono i pochi centri urbani presenti. In questocaso, il satellite Landsat è più che sufficiente percogliere le variazioni che avvengono su scala nonlocale. Il sistema è prevedibile, i suoi cambiamentisono polarizzati in una direzione preferenziale che èquella determinata dall’uomo. Su tempi brevi è im-probabile che altre direzioni vengano coinvolte damodifiche eclatanti.
L’approccio classico all’analisi delle dinamiche delterritorio richiede la mappatura delle immagini telerile-vate di periodi diversi e il loro successivo confrontoper stabilire, per ogni pixel, la classe iniziale e quellafinale (operazione detta di cross-tabulation). Questametodologia si scontra con numerose difficoltà, tra cuiper esempio: a) errori di classificazione in una o inentrambe le immagini, b) incapacità di rendere esplicitii trend spaziali del cambiamento, c) inosservanza dellascala spaziale a cui avvengono le dinamiche, d) impos-sibilità di prescindere dalla risoluzione spaziale del sen-sore utilizzato, e) dipendenza dei risultati dal sistema diclassificazione utilizzato per mappare l’area di studio.L’approccio proposto nel presente lavoro supera lesuddette difficoltà poiché non richiede la classificazio-ne delle immagini telerilevate e permette di evidenziare,oltre ai trend, l’eventuale inadeguatezza di risoluzioneda parte del sensore (analisi sub-pixel). Permette inol-tre di analizzare contemporaneamente le dimensionispaziale e temporale.
Sebbene le due riprese Landsat utilizzate non sianorecentissime, è necessario considerare due aspetti.Mano a mano che si sale nella gerarchia strutturaledell’analisi ambientale (dai microsistemi al paesaggio) icambiamenti tendono ad avvenire a scale temporalimaggiori (WIENS, 1989): è dunque ragionevole pensareche quanto osservato sette anni fa nell’area di studiosia a tutt’oggi verificabile. Inoltre, il presente lavoro hauno scopo prevalentemente metodologico. Come emer-ge dalla letteratura scientifica corrente, l’indice NDVIsi dimostra un ottimo vettore informazionale in gradodi trattenere molta della informazione necessaria peranalizzare le dinamiche del territorio. Le sue variazionisono correlabili sia a variazioni dell’uso reale del suoloche a modifiche strutturali e funzionali della vegetazio-ne.
L’approccio geostatistico ai dati telerilevati rappre-senta un metodo poco dispendioso, a livello economi-co e di tempo, per analizzare in modo sinottico ledinamiche evolutive del territorio. Esso analizza lemodificazioni che avvengono ad un livello gerarchicoelevato (paesaggio) per inferire ipotesi sulle forze che,agendo a livelli gerarchici più bassi, le determinano.Uno svantaggio del metodo proposto risiede nella pe-santezza computazionale. Il calcolo dei SV anisotropi

FERRARINI et al. - Analisi dell’evoluzione temporale del paesaggio 75
ha richiesto, ad un processore di frequenza 600 Mhz,quasi 30 minuti di tempo per ognuna delle tre fasce.Difficilmente si potrebbe fare uso di un calcolatoreavente prestazioni inferiori. Si ritiene comunque chetale inconveniente sia di lieve entità rispetto ai vantaggiche il metodo proposto può fornire.
CONCLUSIONIIl territorio è un mosaico di molteplici componenti,
ognuna delle quali ha una sua dinamica specifica.L’interazione tra le dinamiche delle singole parti general’evoluzione del territorio nel suo complesso. Com-prendere le forze che generano tali modificazioni signi-fica poter avanzare ipotesi sugli scenari evolutivi piùprobabili e, quindi, poter mettere in atto azioni pianifi-catorie efficaci.
Nel presente lavoro si propone l’approccio geostati-stico ai dati telerilevati come una metodologia a bassocosto e di grande efficacia per l’analisi delle caratteri-stiche evolutive del paesaggio. La geostatistica includeuna serie di tecniche aventi lo scopo di ricercare unmodello che descriva la dipendenza spaziale tra i valoriosservati. L’utilizzo congiunto della geostatistica e diun indice altamente informativo come l’NDVI rendepossibile comprendere molte delle proprietà spazio-temporali dell’evoluzione dell’area di studio. È statopossibile mettere in evidenza le modifiche caotiche, su
scala locale, spazialmente eterogenee della zona plani-ziale, dovute all’espansione edilizia e ai cicli colturali.Tali caratteristiche descrivono un sistema in evoluzio-ne non direzionale, variabile su micro-scale e per ilquale è assai difficile produrre scenari evolutivi anchesu tempi brevi. All’opposto, la fascia montana rappre-senta un sistema prevedibile, che evolve su macro-scale, facilmente monitorabile mediante telerilevamen-to satellitare e con una sola direzione evolutiva. Lafascia collinare rappresenta una situazione intermedia,ma più simile alla fascia montana poiché la direzioneNE-SO risulta più interessata a fenomeni evolutivirispetto alle altre.
Poiché le variazioni dell’indice NDVI sono stateanalizzate su tempi brevi (5 anni), era prevedibile chevenissero messe in luce specialmente le dinamicheevolutive di cui l’uomo è il principale artefice. Utiliz-zando immagini Landsat relative a periodi più distanti(dati non disponibili per il presente lavoro) si potrebbe-ro analizzare anche le dinamiche evolutive su tempilunghi, in cui i fenomeni naturali agiscono con maggio-re intensità.
La metodologia proposta ha validità generale, poten-dosi applicare a qualunque area di studio e a qualunquesensore satellitare. Per quanto esposto in precedenza,tale metodo ha grande valenza scientifica e applicativanel campo della gestione e pianificazione territoriale.
BIBLIOGRAFIA
CRESSIE N., 1993. Statistics for spatial data. Wiley, New York, 326pp.
CRESSIE N. e HAWKINS D.M., 1980. Robust estimation of thevariogram. Mathematical Geology, 12: 115-125.
FERRARINI A., ROSSI P., ZACCARELLI N., 2000. Change detectiondegli Habitat CORINE nel bacino del Torrente Baganza (Prov.Parma) tramite l’indice NDVI. Rivista Italiana di Telerileva-mento, 19: 29-35.
GUYOT G., 1989. Signatures spectrales des surfaces naturelles,Collection Télédétection Satellitaire n. 5. Paradigme, 178 pp.
GRIFFITH, J. A, MARTINKO E. A, WHISTLER J. L., PRICE K. P., 2002.Preliminary comparison of landscape pattern-normalized dif-ference vegetation index (NDVI) relationships to CentralPlains stream conditions. Journal of Environmental Quality,31: 846-859.
ISAAKS E.H. e SRIVASTAVA R.M., 1989. An introduction to appliedgeostatistic. Oxford University Press, New York, 284 pp.
JENSEN J. R., 1996. Introductory digital image processing. Aremote sensing perspective, second edition. Prentice Hall seriesin Geographic Information Science, 246 pp.
JOURNEL A.G. e HUIJBREGTS C.J., 1978. Mining Geostatistics.Academic Press, London, 254 pp.
LAW B. E., WARING R. H., 1994. Remote Sensing of Leaf AreaIndex and Radiation intercepted by understory vegetation,
Ecological Applications, 4: 272-279.MATHERON G., 1963. Principles of geostatistics. Economic Geo-
logy, 58: 1246-1266.REMMEL T.K., PERERA A.H., 2001. Fire mapping in a northern
boreal forest: assessing AVHRR/NDVI methods of changedetection. Forest Ecology and Mangament, 152: 119-129.
ROSSI P., 1999. Analisi della diversità vegetazionale mediante laclassificazione CORINE Biotopes. Tesi di Laurea in ScienzeBiologiche, Facoltà di Scienze Matematiche Fisiche e Naturali,Università di Parma.
ROUSE J.W., HAAS R.H., SHELL J.A., DEERING D.W., HARLAN J.C.,1974. Monitoring the vernal advancement of retrogradationof natural vegetation. Final Report, Type III, NASA/GSFC,Greenbelt, MD, 371 pp.
SINGH A., 1989. Digital change detection techniques using Remo-tely sensed Data. In: Eden M.J. and Parry J.T. (eds.), RemoteSensing and Tropical Land Management. John Wiley & Sons,London, 254 pp.
WALSH S.J., CRAWFORD T.W., WELSH W.F. e CREWS-MEYER K.A.,2001. A multiscale analysis of LULC and NDVI variation inNang Rong district, northeast Thailand. Agriculture Ecosy-stems & Environment, 85: 47-64.
WIENS J.A, 1989. Spatial scaling in ecology. Functional Ecology,3: 385-397.


Biol. Amb., 17 (n. 1, 2003) Corso “Metazoi delle acque sorgive e sotterranee” 77
Informazione&Documentazione
Resoconto sul Corso di Formazione“Metazoi delle acque sorgive e sotterranee.Biologia, campionamento e riconoscimento,aspetti ambientali ed applicativi”Genova, 18-22 settembre 2000
Daniela Rocca¹*, Marco Bodon ¹, Silvio Gaiter ¹, Elena Casarino ²
¹ Agenzia Regionale per la Protezione dell’Ambiente Ligure, Dipartimento di Genova, Via Gropallo 5 - 16121 Genova
² Agenzia Regionale per la Protezione dell’Ambiente Ligure, Direzione Scientifica, Piazza della Vittoria 5/C - 16121 Genova
* Autore referente per la corrispondenza ([email protected])
INTRODUZIONEL’entrata in vigore del decre-
to legislativo n. 152/99 ha costitui-to un’importante innovazione del-la normativa nel campo delle risor-se idriche, per il peso dato al para-metro “biologico-biocenotico” nel-la determinazione della qualitàambientale dei corpi idrici superfi-ciali. Sulle acque sorgive e sotterra-nee il testo unico non prende espres-samente in considerazione l’aspet-to ecologico fra i criteri di valuta-zione ambientale; tuttavia, nell’al-legato 3 del suddetto decreto è ac-cennato, per la prima volta: “Do-vranno inoltre essere valutate, seesistenti, le indagini relative allebiocenosi degli ambienti sotterra-nei”.
Anche se la valutazione qua-litativa delle acque sotterranee e ladeterminazione dello stato ambien-tale si basano solo sui parametriabiotici o microbiologici, la consi-derazione dell’esistenza della com-ponente vivente è da ritenere, daun punto di vista concettuale, unimportante punto di partenza e unaspinta per approfondire le cono-scenze biologiche e gli aspetti cheregolano il complesso ecosistemadelle acque sotterranee. La finalità
principale del corso –tenutosi a Ge-nova dal 18 al 22 settembre 2000–è stata quella di diffondere le cono-scenze acquisite fino ad oggi suqueste biocenosi, proponendo alcu-ne tecniche investigative messe apunto tra gli operatori del settore ecercando di fornire alcune basi espunti di lavoro per futuri studiapplicativi.
PRESUPPOSTI PER LAREALIZZAZIONE DEL CORSO
Dalla fine degli anni ’80 aGenova sono state intraprese inda-gini e studi sul mondo biologicosotterraneo e sorgivo, svolti primanell’ambito del Presidio Multizo-nale di Prevenzione (PMP), poicome Agenzia Regionale per la Pro-tezione dell’Ambiente Ligure (AR-PAL), anche in collaborazione conl’Istituto di Zoologia dell’Universi-tà di Genova (attualmente Diparti-mento per lo Studio del Territorio edelle sue Risorse: DIP.TE.RIS.). Leesperienze maturate da questo grup-po di lavoro, prevalentemente orien-tate alla conoscenza delle bioceno-si e alla lettura degli aspetti di alte-razione ambientale e di rischio igie-nico delle acque, hanno portato
alla collaborazione a diverse ini-ziative di studio promosse dal-l’Agenzia Nazionale per la Prote-zione dell’Ambiente (ANPA) e dal-l’Istituto Superiore di Sanità (ISS).
Proprio nell’ambito di un Pro-gramma triennale ISS “Le Acque difalda: nuovi indicatori di qualità edi rischio ambientale”, è nata l’esi-genza di promuovere questa espe-rienza in altre realtà italiane e quin-di la necessità di estendere le cono-scenze generali e metodologiche adaltri operatori. Nel 1999 l’ISS pro-pose ad ARPAL l’organizzazione diun corso di formazione teorico-pra-tico relativo allo studio della faunastigo-crenobionte; la finalità eraquella di colmare le lacune tecni-co-scientifiche e metodologiche,mettendo a disposizione l’esperien-za maturata in questo settore e, alcontempo, promuovere la conoscen-za del mondo vivente sotterraneo ele molteplici informazioni di carat-tere ecologico e igienico che posso-no essere tratte.
La Direzione ARPAL ha ade-rito al progetto anche grazie all’in-tervento di ANPA (che ha provve-duto a sostenere la copertura fi-nanziaria necessaria alla realizza-

Corso “Metazoi delle acque sorgive e sotterranee” Biol. Amb., 17 (n. 1, 2003)78
Informazione&Documentazione
zione del corso) e alla fattiva colla-borazione di DIP.TE.RIS., che si èoccupato, grazie a proprie specifi-che competenze, dell’organizzazio-ne della parte dedicata agli appro-fondimenti tassonomici. Gli espertiintervenuti hanno collaborato allaredazione di dispense sui gruppi dimetazoi considerati. L’Ufficio Co-municazione, Informazione, Edu-cazione della Direzione Generalesi è occupato delle molteplici pro-blematiche relative all’organizzazio-ne generale. La strutturazione el’organizzazione tecnica del corsosono state demandate allo staffscientifico, composto quasi esclusi-vamente da personale strutturato ocomunque operante in ARPAL, inquanto coincidente con il gruppodi lavoro (Tab. I).
CONTENUTI SCIENTIFICIL’argomento principale del
corso ha riguardato le comunitàbiologiche che popolano i diversiambienti di acque sotterranee (fal-de alluvionali, carsiche e fessura-te), ampliando la trattazione anchealle situazioni ambientali di pas-saggio o connessione tra i sistemiipogeo ed epigeo, più propriamentedefiniti ambienti ecotonali (vediTab. II). In quest’ultima tipologiarientrano le sorgenti, i fontanili egli ambienti iporreici. È stata inol-tre affrontata la tematica relativa
Tab. I. Elenco degli esperti intervenuti al corso.
DOCENTI DELL’UNIVERSITÀ DI GENOVA ED ESPERTI TASSONOMI
••••• DIP.TE.RIS. (Dipartimento per lo Studio del Territorio e delle sue Risorsedell’Università di Genova)– Prof. Attilio Arillo– Prof. Andrea Balduzzi– Dott. Mario Mori– Dott. Sebastiano Salvidio– Prof. Silvio Spanò
••••• Esperti tassonomi - gruppi trattati– Dott. Marco Bodon (ARPAL di Genova) –Molluschi– Dott. Antonio Di Sabatino (Università de L’Aquila) – Acari– Dott. Uberto Ferrarese (Museo St. Nat. Verona) – Chironomidi– Prof.ssa Diana Galassi (Università de L’Aquila) – Copepodi– Dott.ssa Beatrice Sambugar (Museo St. Nat. Verona) – Oligocheti– Dott. Fabio Stoch (Museo St. Nat. Verona) – Isopodi e Anfipodi– Prof. Aldo Zullini (Università di Milano Bicocca) – Nematodi
STAFF TECNICO-SCIENTIFICO ED ESPERTI INTERVENUTI
••••• Staff tecnico-scientifico– Dott. Marco Bodon (Dip. Genova ARPAL)– Dott. Silvio Gaiter (Dip. Genova ARPAL)– Dott.ssa Mirvana Feletti (collaboratrice borsista ARPAL)– Dott.ssa Laura Volterra (Lab. di Microbiologia Ambientale ISS)– Dott.ssa Daniela Rocca (Tirocinante ARPAL)– Dott.ssa Elena Casarino (Consulente ARPAL)
••••• Altri esperti– Dott. Fabio Decet (ARPA Veneto) per Idrochimica– Dott.ssa Nicoletta Dotti (Direzione Scientifica ARPAL) per Idrogeologia
Tab. II. Alcuni esempi di ambienti sotterranei o di connessione con le acque superficiali e relativa caratterizzazione fisicaed ecologica.
Siti di indagine Caratterizzazione fisica (acquifero) Caratterizzazione ecologica
Scorrimento in cavità carsica Falda carsica Ambiente stigaleScorrimento in cavità artificiale “Carsico” Ambiente stigalePozzo in falda fessurata Falda in roccia fessurata Ambiente stigalePozzo in piana alluvionale Falda freatica (interstiziale) Ambiente freaticoPozzo profondo in piana alluvionale Falda artesiana (interstiziale) Ambiente freatico profondoMaterasso alluvionale del corso d’acqua Falda freatica (interstiziale) Ecotono: ambiente iporreicoSorgente in roccia fessurata Falda in roccia fessurata Ecotono: (ambiente stigale)-crenalSorgente carsica Falda carsica Ecotono: (ambiente stigale)-crenalSorgente captata in roccia fessurata Falda in roccia fessurata Ecotono: ambiente stigale-(crenal)Sorgente in piana alluvionale (fontanile) Falda freatica (interstiziale) Ecotono: ambiente freatico-crenalVasche lungo la rete idrica “Carsico” Varia in dipendenza dell’origine
delle acque
ai siti di potenziale colonizzazionepresenti in sistemi artificiali cheattingono da acque sotterranee osorgive (captazioni per acquedotti,manufatti lungo la rete idrica, etc.).
Per ogni tipo di habitat sonostate prese in esame le classificazio-ni ecologiche degli organismi ap-partenenti al micro- meio- macro-
benthos e quindi la tipologia dellabiocenosi, le caratteristiche degliorganismi (morfo-fisiologiche) equelle del popolamento presente nelbiotopo (catene trofiche, correlazio-ni, etc.), con particolare attenzionealla biodiversità e alle variazionidovute a fattori fisico-chimici e an-tropici. Si è trattato, inoltre, della

Biol. Amb., 17 (n. 1, 2003) Corso “Metazoi delle acque sorgive e sotterranee” 79
Informazione&Documentazione
geonemia e origine della fauna sti-gobionte, distribuzione attuale edevoluzione, biogeografia ed ende-mismi.
Alla parte di sistematica gene-rale è seguita una trattazione piùapprofondita sui gruppi di metazoibentonici che costituiscono la qua-si totalità dei taxa presenti negliambienti, in particolare sono staticonsiderati: Oligocheti, Molluschi,Copepodi, Anfipodi e Isopodi.
Il primo obiettivo concreto eraquindi quello di consentire ai cor-sisti di essere in grado di compilare
correttamente la lista faunistica edi fornire la caratterizzazione, dalpunto di vista sia fisico che biologi-co, del sito in esame.
Gli organismi viventi costitui-scono gli indicatori più validi dellostato di salute di un ambiente, per-ché sono in grado di interagire congli stimoli provenienti dalla com-ponente biotica e abiotica. Anchenegli habitat sotterranei, infatti, labiocenosi instaurata è la risultantedell’interazione tra fattori tipici delsito di colonizzazione, e caratteriz-
za il singolo biotopo e le sue evolu-zioni temporali. L’esame attentodella tanatocenosi, ricavabile dallacomponente organica del sedimen-to, può fornire utili elementi percompletare e approfondire il qua-dro conoscitivo della risorsa idrica.
La limitata conoscenza e l’ete-rogeneità di questi ambienti e dellecomunità ivi presenti non ha anco-ra portato alla stesura di un vero eproprio manuale di applicazione;non sono state ancora messe a pun-to, infatti, metodiche di indaginesufficientemente sperimentate da
Tab. III. Elenco degli indicatori e degli indici biologici basati sulla composizione tassonomica del popolamento di invertebratie loro tipologia: aspecifico (indice di diversità o indice comparativo) o specifico (come l’indice saprobico o biotico).
Tipo di indice Sull’intera comunità Indice / Indicatore/indicatore / su singoli gruppi
Aspecifico Sull’intera comunità Indici di diversitàIndice di diversità o su singoli gruppi
Aspecifico Sull’intera comunità Indice di equitabilitàIndice di diversità o su singoli gruppi
Specifico Su singoli gruppi Rapporto tra Secernentea sul totale della nematofaunaIndice saprobico
Specifico Su singoli gruppi Rapporto tra specie di Copepodi stigobi e stigofili o stigosseniIndice saprobico
Specifico Sull’intera comunità Indice freatobiologico di qualità delle acque sotterranee (IPB)Indice saprobico
Specifico Sull’intera comunità Struttura della biocenosi in relazione all’inquinamento per acquedi falda interstiziale
Specifico Sull’intera comunità Struttura della biocenosi in relazione all’inquinamento per acquedi falda alluvionale
Specifico Sull’intera comunità Struttura della biocenosi in relazione all’inquinamento per acquedi falda carsica
Specifico Sull’intera comunità Struttura della biocenosi in relazione all’inquinamento peracque sorgive
Specifico Sull’intera comunità Valutazione delle variazioni ambientali su lunghi periodi nellesorgenti, basata sull’accumulo di resti animali e vegetali nei sedimenti
Specifico Sull’intera comunità Struttura della biocenosi come descrittore idrologico per acquedi falda interstiziale
Specifico Sull’intera comunità Struttura della biocenosi come descrittore idrologico per acquedi falda carsica
Specifico Sull’intera comunità Struttura della biocenosi e della tanatocenosi come descrittoreidrologico per acque sorgive
Specifico Sull’intera comunità Struttura della biocenosi e della tanatocenosi come descrittoredel grado di isolamento per captazioni di acque sorgive

Corso “Metazoi delle acque sorgive e sotterranee” Biol. Amb., 17 (n. 1, 2003)80
Informazione&Documentazione
poter essere standardizzate. A diffe-renza delle acque superficiali, nel-le acque sotterranee e sorgive man-cano consolidate possibilità appli-cative finalizzate all’elaborazionedi un vero e proprio indice biologi-co, basato sulla diversa valenza eco-logica degli organismi stigo- e cre-nobionti. Sulla base delle attualiconoscenze tassonomiche è impen-sabile poter standardizzare un in-dice valido per tutto il territorioitaliano, spesso improponibile an-che per territori limitrofi. Se la crea-zione di un indice resta uno deiprincipali obiettivi di ricerca sullecomunità bentoniche di acque sor-give e sotterranee, a tutt’oggi lo sfor-zo per ottenere tale risultato appa-re estremamente ambizioso.
Attraverso lo studio della bio-
cenosi e della sua tanatocenosi, va-lutando le caratteristiche ecologi-che dei taxa, si ha la possibilità ditrarre diverse indicazioni che per-mettono di formulare ipotesi sul-l’idrogeologia (origine dell’acquife-ro alimentante), grado di vulnera-bilità del sito d’indagine (in parti-colare per risorse captate) e gradodi inquinamento o alterazione del-le acque (vedi Tab. III). A titolo diesempio, fra i vari indicatori/de-scrittori, si riporta la tabella riguar-dante il descrittore idrologico (Tab.IV).
ASPETTIDIDATTICO-ORGANIZZATIVI
La partecipazione è stata ri-volta principalmente a laureati inScienze Biologiche e Naturali delle
ARPA italiane ed estesa altresì adoperatori di altri enti pubblici (Di-partimenti di Prevenzione delle ASLe PMP ove non ancora costituite leARPA), di aziende private e a liberiprofessionisti, proprio per l’ampiointeresse applicativo di questo cor-so. Per motivi legati in buona partealle possibilità operative della di-dattica sul campo, il numero dipartecipanti è stato limitato ad unmassimo di una ventina.
Non essendo stato stabilito perl’iscrizione alcun requisito relativoalle conoscenze della materia, illivello dei partecipanti è risultatopiuttosto eterogeneo e ciò ha com-portato difficoltà ad alcuni corsisti(completamente digiuni in materia)e, conseguentemente, ai relatori eallo staff scientifico. Solo alcuni
Tab. IV. Esempio di indicatore biologico come descrittore idrologico per acque sorgive e cenosi corrispondente.
STRUTTURA DELLA BIOCENOSI E DELLA TANATOCENOSI COME DESCRITTORE IDROLOGICO PER ACQUE SORGIVE
Descrizione dell’indicatore Analisi della struttura della comunità bentonica e della tanatocenosi dell’ambiente disorgente
Metodi di misura Riconoscimento a livello di taxa (specie o genere). Per il giudizio vedi la seconda partedella tabella
Scopo dell’indicatore Valutazione della provenienza dell’acqua sorgiva in relazione agli apporti idrici (dallafalda o dalle acque superficiali)
Unità di misura Analisi qualitativa della composizione della biocenosi
Limite dell’indicatore Valutazione qualitativa
Comparto ambientale Acque sorgive
Copertura geografica dei dati Liguria
Periodo di riferimento dei dati 1988-2000
Documenti di riferimento Bodon & Gaiter, 1989, 1995
BIOCENOSI E TANATOCENOSI DI RIFERIMENTO
Sorgente alimentata solo Biocenosi: presenza di organismi crenobionti o stigobionti (Bythinella, Hydrobiidae, daacque sotterranee Niphargus, Proasellus).
Tanatocenosi: presenza di nicchi di molluschi Hydrobiidae stigobionti.
Sorgente alimentata sia da Biocenosi: presenza di organismi crenobionti o stigobionti (Hydrobiidae: Avenionia,acque sotterranee che da Bythinella; Crostacei: Niphargus, Proasellus) e presenza di organismi di acque superficialiacque superficiali (Leuctra, Dytiscidae, Philopotamidae, Chironomidae, Ceratopogonidae).
Tanatocenosi: abbondante detrito vegetale, soprattutto di foglie.
Sorgente alimentata solo da Biocenosi: assenza di organismi stigobionti o tipici crenobionti; presenza quasi esclusivaacque superficiali di larve di insetti.
Tanatocenosi: assenza di nicchi di molluschi Hydrobiidae crenobionti o stigobionti.

Biol. Amb., 17 (n. 1, 2003) Corso “Metazoi delle acque sorgive e sotterranee” 81
Informazione&Documentazione
partecipanti avevano avuto, in pre-cedenza, esperienze connesse conle problematiche dell’analisi dellecomunità biotiche e della sistema-tica del benthos di acque sotterra-nee. Per questo motivo si è ritenutoutile distribuire una cospicua quan-tità di materiale didattico (Tab. V)comprendente una guida tassono-mica di facile lettura per il ricono-scimento e la classificazione degliorganismi, una serie di dispensespecifiche per i principali gruppisistematici oggetto di approfondi-mento, e altra documentazione suidiversi argomenti trattati.
Il corso è stato impostato trat-tando soprattutto gli argomenti ine-
renti la biologia e l’ecologia delleacque ipogee, avendo cura di deli-neare in parallelo i diversi ambien-ti fisici oggetto d’indagine. Fra lemolteplici metodiche per la raccol-ta della biocenosi e della sua tana-tocenosi, sono state scelte e propo-ste quelle ritenute maggiormenteapplicabili nei tre diversi siti di cam-pionamento: una sorgente, un poz-zo che attinge da falda freatica eun ambiente iporreico (connessio-ne fra un corso d’acqua superficia-le e la falda sotterranea). Ad ognicampionamento e fase analitica dicampo sono seguiti il trattamentodel campione e l’analisi di labora-torio.
PROGRAMMA ESTRUTTURAZIONE DEL CORSO
Per poter sviluppare tutti gliargomenti e i momenti di lavorodurante cinque giornate, le diverseparti sono state così strutturate: par-te teorica, parte pratica con fase insitu (raccolta dati di campo e cam-pionamento), parte pratica in labo-ratorio (separazione e classificazio-ne), parte concernente gli appro-fondimenti tassonomici e parte con-clusiva (analisi dei risultati, appli-cazioni, prospettive; Tab. VI). Lecinque fasi sono dettagliate di se-guito.• Parte teorica, propedeutica edi inquadramento della materia incui sono stati affrontati alcuni ar-gomenti generali riguardanti i pos-sibili siti investigabili e le intercon-nessioni con gli altri ecosistemi esistemi artificiali (ambienti fisici ecaratterizzazioni biologiche relati-ve); elementi fisici di idrogeologiadelle acque sotterranee; elementi diidrogeochimica per la valutazionedelle acque; microbiologia (batte-riologia ambientale con particola-re risvolto all’importanza igienico-sanitaria dei metazoi di questi am-bienti); ecologia e biologia della fau-na bentonica. Sono state introdottele metodiche di campionamento, diconservazione e di studio del mate-riale; l’analisi della componenteorganica, vegetale ed animale, delsedimento (tanatocenosi) e, infine,è stata trattata la parte sistematicadei principali gruppi di metazoi sti-go- e crenobionti.• Parte pratica di indagine am-bientale in situ: sono stati esaminatitre differenti habitat. Durante l’in-dagine sono state rilevate le carat-teristiche ambientali del sito in esa-me, i parametri di campo e quellifisico-chimici. Si è proceduto conl’applicazione della metodica dicampionamento specifica per cia-scun sito: retino immanicato per lasorgente; sonda collegata a pompa
Tab. V. Materiale didattico distribuito durante il corso.
Guide Autore
Guida al riconoscimento degli organismi rinvenibili M. Felettinelle acque sorgive e sotterranee
Dispense
Inquadramento ecologico e finalità del corso A. BalduzziLegislazione ed altri contributi normativi d’indirizzoInquadramento dei siti (ambienti) investigabili e intercon- S. Gaiternessioni con gli altri ecosistemi e sistemi artificialiApplicazioni dell’idrogeochimica nelle valutazioni della F. Decetqualità delle acque sotterranee: limiti, successi e frontiereElementi biologici ed ecologici (con particolare risvolto alla M. Bodonfauna micro- meio- e macrobentonica)Analisi della componente organica del sedimento (tanatocenosi)Metodiche di campionamento, conservazione e studio del materialeModulistica: significato delle voci e compilazione delle S. Gaiterschede Sorg, Poz, Ipor e BiolCriteri di valutazione sull’origine, grado di protezione e M. Bodonqualità delle acque (autoecologia ed analisi della comunità)Indici e Indicatori M. Bodon,
S. Gaiter,A. Risso
Altra documentazione
I Molluschi M. BodonCopepodi sotterranei: pattern di diversità su scala D. Galassiecologica ed evolutivaClasse: Malacostraca - Isopodi, Anfipodi e ordini minori F. Stochcon rappresentanti stigobiGli Oligocheti acquatici italiani B. SambugarNematodi delle acque sorgive e sotterranee A. ZulliniI Chironomidi delle sorgenti fredde europee U. FerrareseThe biology and ecology of lotic water mites (Hydrachnidia) A. Di Sabatino,
R. Gerecke, P.Martini (trad.M. Feletti)

Corso “Metazoi delle acque sorgive e sotterranee” Biol. Amb., 17 (n. 1, 2003)82
Informazione&Documentazione
a vuoto per il pozzo; pompa Nortone metodo Karaman-Chappuis perl’ambiente iporreico. La filtrazionedei campioni è stata effettuata conretini a maglie di 500 μm e 100 μm.I dati sono stati registrati in apposi-ti moduli (Sorg, Poz, Ipor e Biol).• Parte teorico-pratica di labo-ratorio: dapprima è stato fornitoun quadro sistematico di riferimen-to: interpretazione e lettura dellechiavi tassonomiche per la defini-zione del gruppo di appartenenzacon riscontro diretto al microsco-pio, grazie all’esame di alcuni pre-parati di laboratorio. Si è procedu-to con il trattamento dei campioniraccolti sul campo: setacciatura,lavaggio e separazione delle frazio-ni, misurazione volumetrica, sepa-razione degli organismi e delle al-tre componenti del sedimento. Suc-
cessivamente, con l’ausilio dello ste-reomicroscopio, i taxa sono staticlassificati fino al livello tassono-mico previsto in questa sede, è stataaltresì determinata e valutata lacomponente organica del sedimen-to.• Parte di approfondimenti: èstato dedicato ampio spazio agliapprofondimenti tassonomici deiprincipali taxa di metazoi di acquesorgive e sotterranee; gli esperti han-no fornito, per ciascun gruppo si-stematico, brevi note sulle caratte-ristiche morfologiche ed anatomi-che; è stata sviluppata la parte rela-tiva all’ecologia e alla distribuzio-ne geografica.• Parte conclusiva: è stata im-postata una discussione sull’anali-si dei dati risultanti dai campiona-menti effettuati e sull’elaborazione
delle informazioni ottenute, ponen-do in risalto ciò che è emerso dallalettura della componente metazoi-ca. Con la spiegazione e l’applica-zione di specifici indicatori biologi-ci sono stati affrontati i criteri divalutazione sull’origine, sul gradodi protezione e sulla qualità delleacque (autoecologia ed analisi del-la comunità); inoltre è stata propo-sta una prova pratica su questotema.
I corsisti, suddivisi in 11 grup-pi di lavoro, hanno sostenuto unaprova mirata all’individuazionedella risposta corretta riguardantela caratterizzazione biologica (bio-cenosi e tanatocenosi) del sito dicampionamento. A ciascun grup-po, costituito da due persone, sonostate consegnate cinque schede (suun totale di 14 schede preparate);ogni scheda è stata strutturata contre possibili risposte di cui soltantouna corretta. Nella tabella VII èriportato un esempio di scheda (li-sta faunistica ed esame del sedi-mento) la cui risposta corretta è laB. Infine, sono state valutate ipote-si e prospettive di lavoro applicati-vo nei settori trattati nel corso, inconsiderazione dell’interesse dimo-strato, garantendo la disponibilitàfutura dello staff scientifico.
BILANCIO DEL CORSO• Valutazione del rispetto dei tem-pi di lavoro: complessivamente i tem-pi di svolgimento del corso sonostati perfettamente rispettati. Il pro-gramma, anche se apparso troppocorposo, è stato interamente com-pletato; grazie anche alla vicinan-za ed accessibilità dei siti presceltiè stato inoltre possibile prevederenel dettaglio sia i tempi di sposta-mento sul territorio, che i tempirelativi al campionamento stesso.• Valutazione qualitativa dei cor-sisti (prova finale): la figura 1 mostraun grafico che sintetizza i risultatidella prova pratica eseguita dai cor-
Tab. VI. Schematizzazione del programma svolto.
AMBIENTI E FAUNA DELLE ACQUE SOTTERRANEEAula didattica CREA - Via al Porto Antico - Palazzina Millo– Registrazione partecipanti– Saluto delle Autorità e presentazione del corso– Introduzione: scopi e finalità del corso– Elementi fisici (idrogeologia)– Elementi chimici (idrochimica)– Elementi microbiologici (batteriologia ambientale)– Inquadramento dei siti investigabili ed interconnessioni con gli altri ecosistemi e sistemi
artificiali– Elementi biologici ed ecologici– Sistematica dei principali gruppi di metazoi stigo- e crenobionti– Analisi della componente organica del sedimento (tanatocenosi)– Metodiche di campionamento, conservazione e studio del materiale– ModulisticaCAMPIONAMENTO - Comune di Casella– Indagine ambientale presso un pozzo, una sorgente, una falda iporreicaSMISTAMENTO E CLASSIFICAZIONE - Aula DIP.TE.RIS. – Corso Europa, 26– Separazione degli organismi dal sedimento e riconoscimento dei taxa– Valutazione della componente organica del sedimento– Compilazione dei modelli relativiAPPROFONDIMENTI TASSONOMICI - Aula DIP.TE.RIS. – Corso Europa, 26– Tassonomia, caratteristiche morfologico-anatomiche– Classificazione principali gruppi di metazoi stigo- e crenobionti, con approfondimenti
sull’ecologia e sulla distribuzione geografica (in particolare: Nematodi, Molluschi,Oligocheti, Acari, Copepodi, Isopodi, Anfipodi, Ditteri Chironomidi)
DISCUSSIONEARPAL, Dipartimento Provinciale di Genova – Aula Didattica – Via Montesano, 5– Criteri di valutazione sull’origine, grado di protezione e qualità delle acque– Analisi dei risultati dei campionamenti ed elaborazione delle informazioni ottenute

Biol. Amb., 17 (n. 1, 2003) Corso “Metazoi delle acque sorgive e sotterranee” 83
Informazione&Documentazione
sisti. Visibilmente il giudizio com-plessivo delle risposte è discreto; lamaggior parte dei gruppi ha valuta-to correttamente quasi tutti i que-stionari proposti.• Giudizio dei partecipanti sul cor-so: al fine di valutare il gradimentodal punto di vista organizzativo edel contenuto, i partecipanti sonostati invitati a compilare un “que-stionario di fine corso” e a fornireun commento finale relativo ad ungiudizio personale, nonché suggeri-
menti, accorgimenti e proposte pereventuali corsi futuri. Il grafico infigura 2 riporta il valore medio, persingola voce, delle risposte fornitedai partecipanti.
Dall’insieme dei commenti, isuggerimenti maggiormente eviden-ziati sono:– Durante la fase di campiona-mento, per ogni metodologia, ren-dere più attiva la partecipazione,ad esempio con la suddivisione erotazione di limitati gruppi di cor-sisti.– Organizzare la fase di labora-torio in piccoli gruppi di parteci-panti, dove ogni gruppo è seguitoda un proprio docente; dedicare aquesta fase un numero di ore mag-giore.– Raggruppare i partecipanti ingruppi di lavoro omogenei per livel-lo di conoscenza della materia, inparticolare per il riconoscimentodei taxa. Riguardo quest’ultima os-servazione è stato proposto di sud-dividere il corso in due o più livellidiversi, in funzione del grado diconoscenza ed esperienza del par-tecipante stesso.
Tra gli altri suggerimenti: di-sporre di una maggiore quantità di“preparati” in laboratorio, allo sco-po di non limitare l’osservazionesolo a taxa raccolti durante le le-zioni sul campo ma estenderla an-che a metazoi reperibili con minorfrequenza o rari; ridurre la partededicata alla teoria, considerata trop-po vasta, eterogenea e concentratanella prima giornata; ampliare laparte relativa ai gruppi faunistici.
Il bilancio dell’esperienza de-scritta è quindi da ritenersi moltopositivo, poiché le finalità eranoabbastanza chiare e condivise datutti i componenti organizzatori delcorso. I risultati ottenuti durante lasettimana, sia in termini di appren-dimento che di interesse suscitato,dovranno naturalmente essere veri-ficati con ulteriori esperienze di
applicazione della metodica di in-dagine da parte dei corsisti.
PROPOSTE PERUN CORSO FUTURO
In base alle richieste di ade-sione ottenute e constatato l’inte-resse suscitato prima e dopo il cor-so, si ritiene possibile la realizza-zione di un secondo corso a cuiverranno apportate tutti i migliora-menti necessari. Questa prima espe-rienza ha dimostrato che l’interes-se per l’argomento è maggiore ri-spetto a quanto previsto e sicura-mente la realizzazione di un even-tuale secondo corso, se da una par-te potrà fare tesoro degli indubbivantaggi dell’esperienza acquisita,dall’altra dovrà prevedere una mi-gliore fluidità organizzativa, alla
Tab. VII. Esempio di scheda propostanella prova pratica.
Sito di campionamento: emergenzaidrica non captata
ESAME DEL MACROBENTHOSFreq.*
RSSRFFRSR
Taxa:HaplotaxidaeLumbriculidaeAvenioniaOstracodaNiphargusProasellus (sp. anoftalma)LeuctraPhilopotamidaeChironomidae
ESAME DEL SEDIMENTOResti di origine vegetale:detrito di radicidetrito carbonizzato
Resti di origine animale:resti Artropodi terrestrifoderi di Tricotterigusci di Ostracodiresidui bozzoli Araneidi
Freq.*AR
Freq.*RSRR
* R raro, S scarso, F frequente, A abbondante.
POSSIBILI RISPOSTEA - Acque di origine superficiale(riemergenza di un corso d’acqua);B - Acque prevalentemente di originesotterranea con contributi di acquesuperficiali;C - Acque di origine sotterranea mafortemente inquinate (scarichi dinatura chimica che intercettano lafalda).
Fig. 1. Rappresentazione graficarelativa all’andamento dei risultatidella prova pratica sostenuta daicorsisti.
0
1
2
3
4
5
6
1 2 3 4 5N. Risposte e satte
N. G
rupp
i di l
avor
o
0
1
2
3
4
5 organizzazione
contenuti
metodi diinsegnamentoadeguatezza ai mieibisogniutilità per la miaprofessioneil mio coinvolgimento
il mio divertimento
Fig. 2. Giudizi medi di valutazione delcorso espressi dai partecipanti(punteggi da 1= insufficiente a 5=ottimo).

Corso “Metazoi delle acque sorgive e sotterranee” Biol. Amb., 17 (n. 1, 2003)84
Informazione&Documentazione
quale si è sopperito con l’impegnodei singoli. Schematicamente è ri-portato nella tabella VIII quantoattualmente costituisce l’ossaturadel secondo corso.
Occorre comunque dedicareparticolare cura nella scelta dei par-tecipanti, eventualmente organiz-zando il corso su diversi livelli di
approfondimento. Il corso può quin-di diventare un momento di scam-bio delle conoscenze e permetteredi assimilare esperienze e contribu-ti delle differenti situazioni ambien-tali e biologiche italiane, apportan-do miglioramenti alle metodiche eal quadro applicativo degli indica-tori.
Tab. VIII. Struttura ideata per il corso futuro.
• Selezione dei partecipanti in base alla conoscenza della materia – requisiti richiesti:– laurea in Scienze Biologiche o Naturali (o titolo equipollente);– conoscenze specifiche nel campo idrobiologico;– conoscenze approfondite sul benthos di acque superficiali (eventualmente, prima dello svolgimento del corso, si
tengono lezioni formative sul macrobenthos per gli operatori con minore esperienza su questo tema).• Organizzazione dei corsisti in gruppi di lavoro: definizione di gruppi di lavoro formati da un limitato numero di
corsisti (n° 3 max) con la disponibilità di un istruttore. Ogni singola parte del corso, in questo modo, può essereaccuratamente seguita dal partecipante sia durante il campionamento in situ, sia per la preparazione del campione inlaboratorio e l’analisi, al microscopio binoculare, del campione preventivamente preparato.
• Lezioni teoriche:– ridurre la parte dei saluti delle autorità e delle introduzioni;– migliorare le parti generali e quelle relative alle tematiche di supporto, quali microbiologia, idrogeologia e idrochimica,
attenendole concretamente alle necessità d’interpretazione del mondo vivente e limitandosi pertanto a lezionistrettamente attinenti all’argomento principale del corso: i metazoi, i fattori ecologici che contraddistinguono lebiocenosi di acque sotterranee, le caratteristiche peculiari dei diversi ambienti e l’autoecologia dei vari taxa. Anche lelezioni cardine devono essere strutturate in maniera più coinvolgente, ad esempio, mediante la proiezione di diaposi-tive integrate con altri sussidi didattici.
• Lezioni pratiche:– di campionamento: è necessario che la fase sul campo sia maggiormente partecipativa. Ogni corsista deve trovarsi
direttamente a contatto con le diverse attrezzature, pertanto ciascun gruppo di lavoro deve essere impegnato sulcampionamento di ogni sito oggetto di indagine.
– di laboratorio: il trattamento dei diversi campioni (di pozzo, di sorgente, dell’ambiente iporreico) viene eseguito daldocente una prima volta con l’ausilio di una videocamera, successivamente ciascun gruppo di lavoro, con l’aiutodell’istruttore, si deve occupare del proprio campione mediante l’utilizzo dello stereomicrosopio. Non è strettamentenecessario che il materiale da esaminare sia solo quello raccolto sul campo (la biocenosi campionata non fornisce quasimai un’elevata biodiversità tale da garantire una conoscenza sufficientemente ampia, applicabile alle diverse realtà);è possibile infatti esaminare dei preparati preventivamente raccolti, con materiale faunistico e componente organica,tanto da poter inizialmente compilare i modelli specifici per ciascun sito. Infine si conclude con un riassunto e unconfronto generale dei taxa reperiti in ogni ambiente investigato, per procedere poi alla compilazione definitiva delleschede biologiche, nelle quali si registrano la lista faunistica e la componente organica del sedimento. La valutazionesarà effettuata nella fase conclusiva, prendendo in considerazione anche i dati fisico-chimici e quelli ispettivo-ambientalirilevati.
• Lezioni di approfondimento: lezioni specifiche, approfondite dagli esperti, relative ai singoli gruppi tassonomicitrattati durante il corso. Per ciascun gruppo di metazoi deve essere rispettata la medesima linea d’esame: morfologia edanatomia, tassonomia (mediante l’utilizzo di specifiche chiavi analitiche), aspetti ecologici e distribuzione geografica. Igruppi trattati dovranno essere meglio selezionati per includere solo quelli effettivamente utili come indicatori.
• Lezioni conclusive: valutazione finale, complessiva, di ciò che è emerso dal corso, con particolare risalto ai risultatiottenuti dai campionamenti; applicazione degli indicatori biologici, esecuzione di una prova pratica, considerazioni sugliaspetti applicativi delle metodiche affrontate, eventuali prospettive di collaborazione in applicazioni di lavoro nei settoriproposti dal corso, etc.
• Materiale didattico:– Chiavi sistematiche: migliorare le chiavi generali in modo da fornire uno strumento utile e più semplice per
l’inquadramento dei taxa a livello tassonomico richiesto.– Informazioni ecologiche e biogeografiche sui taxa: fornire un quadro aggiornato e dettagliato sugli organismi di acque
sotterranee italiane.– Bibliografia: completare e aggiornare la lista bibliografica sulla materia.
RingraziamentiSi ringrazia per la collaborazione la Sig.raInes Para e la Sig.ra Fiorella Sgorbini (Uff.Com. Inf. Ed., Dir. Gen. ARPAL) che sisono occupate delle molteplici proble-matiche correlate all’organizzazionegenerale; inoltre, un particolare ringra-ziamento è rivolto alla Prof.ssa LauraVolterra (Istituto Superiore di Sanità)fautrice dell’iniziativa per la realizzazionedel corso.

Biol. Amb., 17 (n. 1, 2003) 85
Informazione&Documentazione
Il 20 maggio 2002 a New Yorkmoriva Stephen Jay Gould, docentedi paleontologia, geologia, biologiae storia della scienza alla HarvardUniversity. La maggior parte deilettori lo ricorderà soprattutto per isuoi brillanti saggi di storia natura-le, per le sue stimolanti idee sul-l�evoluzione biologica o per il suovivace impegno contro il determini-smo genetico. Molti dei suoi testisono stati tradotti e pubblicati inItalia a partire dai primi anni �80.Il presente intervento vuole essereun contributo al ricordo della figu-ra e dell�opera di questo straordi-nario scienziato ed un invito, perchi ancora non li conoscesse, allalettura dei suoi scritti.
Le prime passioni e la carrieraS.J. Gould nasce a New York
il 10 settembre 1941. Le sue piùgrandi passioni, la paleontologiaed il baseball, risalgono all�infan-zia: la prima esplode a cinque annidopo che il padre lo aveva accom-pagnato a visitare l�American Mu-
UN’IDEA DELLA VITA:RICORDO DI STEPHEN JAY GOULD
seum of Natural History (�I drea-med of becoming a scientist, in gene-ral, and a paleontologist, in particu-lar, ever since the Tyrannosaurus ske-leton awed and scared me�); la se-conda si manifesta con l�appassio-nata ammirazione per la squadradei New York Yankees (�Joe DiMag-gio activated the greatest and mostunattainable dream of all humanity,the hope and chimera of all sages andshamans: he cheated death, at leastfor a while�).
Nel 1963 Gould si laurea ingeologia e nel 1967 consegue ilPh.D. in paleontologia presso laColumbia University. Nello stessoanno inizia la sua carriera accade-mica ad Harvard come AssistantProfessor of Geology; nel 1971 di-viene Associate Professor e, due annidopo, Professor of Geology and Zo-ology. Dal 1982 è Alexander Agas-siz Professor of Zoology and of Geo-logy. Tra gli incarichi che ricoprevi sono quelli di Curator of Inverte-brate Paleontology nell�HarvardMuseum of Comparative Zoology edi membro aggiunto del Departmentof the History of Science. Dal 1996è Vincent Astor Visiting ResearchProfessor of Biology alla New YorkUniversity.
Le opere principaliNel 1974 Gould comincia a
collaborare con la rivista mensiledell�American Museum of NaturalHistory, dove tiene una rubrica in-titolata �This View of Life�, con laquale raggiunge un�enorme popo-larità. Gran parte dei suoi 300 sag-gi, pubblicati ininterrottamente per27 anni1 , sono raccolti in 10 volu-
mi, alcuni dei quali tradotti anchein Italia. La prima di queste raccol-te, pubblicata nel 1977, si intitola�Ever since Darwin: Reflections inNatural History� (trad. it.: �Questaidea della vita�, Editori Riuniti,Roma, 1984). Con la seconda rac-colta, �The Panda�s Thumb: MoreReflections in Natural History�(trad. it.: �Il pollice del panda�, Edi-tori Riuniti, Roma, 1983), Gouldvince, nel 1981, l�American BookAward for Science. Comincia adottenere una certa notorietà anchenel nostro Paese, ulteriormente ac-cresciuta dal libro successivo,�Hen�s Teeth and Horse�s Toes�(trad. it.: �Quando i cavalli aveva-no le dita�, Feltrinelli, Milano,1984), i cui saggi sono incentratisull�opposizione al creazionismo�scientifico� e sulle diverse impli-cazioni, per la società e per la cul-tura, dell�evoluzione biologica. Laquarta raccolta di saggi è �The Fla-mingo�s Smile: Reflections in Natu-ral History� (trad. it.: �Il sorriso delfenicottero�, Feltrinelli, Milano,1987), cui segue �Bully for Bronto-saurus: Reflections in Natural Hi-story� (trad. it.: �Bravo brontosau-ro�, Feltrinelli, Milano, 1992), che
1 Il primo saggio di Gould per la rubrica
�This View of Life�, intitolato �Size andshape�, può essere letto in: www.amnh. org/n a t u r a l h i s t o r y / e d i t o r s _ p i c k /november99_pick.html. L�ultimo suo sag-gio, �I have landed�, pubblicato sul nume-ro di dicembre/gennaio 2000/2001 delNatural History Magazine, si trova in:www.amnh.org/naturalhistory/features/1200_feature.html. Molti di questi saggiscritti da Gould, insieme a diversi altri suoiarticoli, sono dipsonibili in: www.stephenjaygould .org.
Pietro GenoniARPA Lombardia, Dipartimento subprovinciale di Parabiago, Via Spagliardi 19, 20015 Parabiago (MI)

Ricordo di S.J. Gould Biol. Amb., 17 (n. 1, 2003)86
raccoglie 35 saggi pubblicati nel-l�arco di sei anni. La sesta raccoltaè �Eight Little Piggies: Reflectionsin Natural History� (trad. it.: �Ottopiccoli porcellini�, Rizzoli, Milano,1995). Con �Dinosaur in a Hay-stack: Reflections in Natural Hi-story� (trad. it.: �Come un dinosau-ro nel pagliaio�, Mondadori, Mila-no, 1998), Gould supera il traguar-do di due decenni di collaborazio-ne continua con il Natural HistoryMagazine. Il libro comprende, trale altre, riflessioni sulle teorie evo-lutive, sulla trasformazione deimusei di storia naturale in parchitematici, su un testo di Edgar AllanPoe riguardo le conchiglie, sull�ope-ra �In Memoriam� di Tennyson, su�Jurassic Park�, sul Protocollo Wan-nsee, che documenta la �soluzionefinale� di Hitler per lo sterminiodegli ebrei.
Ricordiamo altre due raccol-te di saggi scritti per il NaturalHistory, �Leonardo�s Mountain ofClams and the Diet of Worms�(1998) e �The Lying Stones of Mar-rakech� (2000), nonché l�ultima econclusiva raccolta, che si intitola�I have landed: The End of a Be-ginning in Natural History� (2002).
Si tratta di scritti che testimo-niano l�ecletticità e la vastità degliinteressi di Gould per la storia, l�ar-te, la letteratura, sempre viste nelleloro connessioni, anche solo evoca-tive, con la storia naturale e labiologia. Sono lavori destinati, conenorme successo, ad un gruppo dilettori molto più vasto rispetto allostretto ambito scientifico degli �ad-detti ai lavori�. In questa operazio-ne delicata, che potrebbe essere fat-ta facilmente bersaglio di critiche,Gould non rinuncia ad un perso-nale rigoroso principio: �i concettiscientifici, in tutta la loro ricchez-za ed ambiguità, possono esserespiegati senza alcun compromessonè semplificazione fuorviante, inun linguaggio accessibile a tutte le
persone intelligenti�.Il rigore scientifico, affianca-
to ad un�accattivante abilità espo-sitiva, pervade anche tutte le altresue opere, di cui richiamiamo alcu-ne tra le più significative.
Risale al 1977 il primo impor-tante testo monografico, �Ontogenyand Phylogeny�, in cui Gould esa-mina le dimensioni storiche, filoso-fiche e scientifiche della biologiadello sviluppo. Punto di partenzadei ragionamenti dell�autore èl�enunciato di Ernst Haeckel se-condo cui �l�ontogenesi ricapitolala filogenesi�.
Un secondo fondamentale la-voro è rappresentato da �The Mi-smeasure of Man� (trad. it.: �Intelli-genza e pregiudizio: le pretese scien-tifiche del razzismo�, Editori Riu-niti, Roma, 1985), pubblicato nel1981. Gould critica il convincimen-to che l�intelligenza possa esseredescritta quantitativamente da unsingolo valore misurabile e che l�in-telligenza degli individui (e dei grup-pi di individui) possa essere ordina-ta in base a tale valore. Nel 1996,come risposta all�uscita negli USAdi un libro che pretende di provarescientificamente le differenze raz-ziali dei Q.I. (�The Bell Curve: TheReshaping of American Life by Dif-ference in Intelligence�, di R.J. Herr-nstein e C. Murray, 1994), Gouldripubblica �The Mismeasure ofMan� in forma rivista ed ampliatacon l�aggiunta di cinque saggi2 .
Con �Time�s Arrow, Time�sCircle: Myth and Metaphor in theDiscovery of Geological Time� (trad.it.: �La freccia del tempo, il ciclodel tempo�, Feltrinelli, Milano,1989) Gould ripercorre la storia del-la geologia da Burnet a Lyell ed ilsuo impatto sulla nozione cultura-le di tempo geologico, giungendoall�elaborazione del concetto di�tempo profondo�, indispensabileper la comprensione dei meccani-smi evolutivi e geologici.
Nel 1990, Wonderful Life: TheBurgess Shale and the Nature ofHistory (trad. it.: �La vita meravi-gliosa. I fossili di Burgess e la natu-ra della storia�, Feltrinelli, Milano,1990), è premiato con il ScienceBook Prize. Viene descritta la sto-ria, l�interpretazione ed il significa-to dei fossili scoperti nel 1909 daCharles Walcott a Burgess, nellaBritish Columbia. Il titolo fa riferi-mento ad un film del 1946 di FrankCapra, in cui un angelo mostra alprotagonista, che intende togliersila vita, come sarebbe stato il mon-do se non fosse mai nato. Gouldsostiene che l�intera storia biologi-ca può essere vista allo stesso modo:eventi accidentali hanno determi-nato, in modo del tutto casuale, ilcorso della vita sulla terra e se ilfilm della vita potesse essere riav-volto e riproiettato, ne risulterebbequalcosa di completamente diffe-rente da quello che attualmente cicirconda.
Nel 1982 a Gould viene dia-gnosticato un mesotelioma addo-minale, una rara e pericolosa for-ma di cancro. La storia della suamalattia diventa un saggio sull�uti-lizzo e sull�interpretazione della sta-tistica3 , pubblicato insieme ad altriin un successo letterario del 1996:�Full House: The Spread of Excel-lence from Plato to Darwin� (trad.it.: �Gli alberi non crescono fino alcielo�, Mondadori, Milano, 1997).
La sua ultima importante ope-ra, di oltre 1300 pagine, si intitola�The Structure of Evolutionary The-ory� ed è stata pubblicata due mesiprima della morte.
2 Uno di questi è l�articolo �Curveball�,
pubblicato su The New Yorker del 28 no-vembre 1994, che si può leggere in:www.dartmouth.edu/~chance/course/topics/curveball.html.3 Il saggio, che si intitola �The Median isn�t
the Message�, può essere letto in: /cancerguide.org/median_not_ msg.html.

Biol. Amb., 17 (n. 1, 2003) Ricordo di S.J. Gould 87
Le idee sull�evoluzionebiologica
Gould è stato una figura diprimo piano nel dibattito scientifi-co sull�evoluzione biologica e sulleteorie che tentano di spiegarne imeccanismi. Il suo pensiero si èspesso scontrato con le idee preva-lenti nel mondo accademico deineo- e degli ultra-darwinisti.Nel1972, insieme a Niels Eldredge espo-ne la famosa teoria degli equilibripunteggiati (punctuated equilibria)4,secondo cui l�evoluzione delle for-me viventi avviene durante rapidefasi di transizione (punteggiature)intervallate da lunghi periodi distasi (equilibri). La teoria si opponead una tradizione darwinista orto-dossa che vedeva l�evoluzione comeun processo graduale, lento e conti-nuo, di trasformazioni. Secondoquest�ultima idea, la testimonian-za delle forme fossili giunte fino anoi non potrebbero spiegare talegradualismo solo a causa dell�in-completezza del materiale paleon-tologico, aspetto che Darwin ed ineodarwinisti avevano attribuitoalle carenze del processo di fossiliz-zazione. Eldredge e Gould suggeri-scono che la comparsa improvvisadi nuove specie nelle forme fossilirifletta la realtà del meccanismoevolutivo: la maggior parte dellespecie per lunghi periodi di tempo,anche milioni di anni, non evolvein maniera apprezzabile, ma variacasualmente all�interno di intervallistatisticamente prevedibili5 .
Alla fine degli anni �70, un�al-tra battaglia contro il pensiero evo-luzionistico dominante soprattuttoin Inghilterra e negli Stati Unitiviene ingaggiata da Gould insiemeal biologo Richard Lewontin. Si trat-ta della critica al programma adat-tazionista6 , il quale si basa sulla�fede nel potere della selezione na-turale come agente ottimizzante�. Idue ricercatori chiamano questanozione �paradigma di Pangloss�,
riferendosi ad una celebre figuradel �Candido� di Voltaire, il qualeamava ripetere che �è impossibileche le cose non siano dove sono,poiché ogni cosa è per il meglio�.Essi contestano l�abitudine di scom-porre gli organismi in singole parti,caratteri o geni, e di cercare succes-sivamente una motivazione adatta-tiva per ciascuno di essi, perdendocosì di vista il fatto che gli organi-smi sono delle unità integrate, conpiani costruttivi limitati da costri-zioni evolutive più importanti delleforze della selezione7 .
Al di là delle differenti opinio-ni che dividono il mondo scientifi-co riguardo i meccanismi evolutivi,Gould è comunque stato uno stre-nuo oppositore del creazionismocosiddetto �scientifico�. I creazio-nisti negli Stati Uniti hanno tenta-to ripetutamente, anche in tempimolto recenti, di far attribuire lastessa dignità delle materie scienti-fiche all�insegnamento scolasticodel libro della Genesi, subendo for-tunatamente una serie di sconfittegiudiziarie. Gould, testimone innumerosi processi, avverte un forteinteresse politico dietro queste posi-zioni, poiché il creazionismo rap-presenta un tema caro �alla risor-gente destra evangelica�. Egli so-stiene che finché esisteranno gruppidi interesse con molto denaro, lalotta contro il creazionismo nonpotrà mai essere formalmente vinta.
EpilogoStephen Jay Gould è stato un
brillante scienziato che ha saputoconiugare l�amore per la storia na-turale con i suoi molteplici interes-si, riuscendo a coinvolgere nei suoiappassionati ragionamenti un enor-me numero di lettori, spesso prividi una formazione scientifica. Daisuoi scritti traspare una personali-tà caratterizzata da una limpidaonestà intellettuale, un difensoreostinato delle proprie convinzioni
�sempre scrupolosamente documen-tate�, un intellettuale pronto a lot-tare contro ogni oscurantismo scien-tifico e determinato nel demolire ilmito della neutralità della scienza.
Colpito da un cancro diffuso,una forma non correlata con quel-la diagnosticata vent�anni prima,confidò ad un amico che gli chie-deva notizie sulla sua malattia:�we�re still in the data-collection sta-ge, no conclusions yet�.
Il messaggio che ci lascia èdestinato ad aumentare i nostridubbi sul significato dell�esistenza:la comparsa di Homo sapiens sullaterra non è la conseguenza di uninevitabile aumento della comples-sità del sistema nervoso, bensì diuna rete di innumerevoli eventi con-tingenti e fortuiti che sarebbero po-tuti accadere diversamente, portan-do altrove il corso dell�evoluzione.Ma, negando risolutamente l�esisten-za di conflitti tra gli ambiti in cuimolti di noi cercano le proprie ri-sposte, ci teneva a sottolineare che:�religion is about ethics and values,and science is about facts�.
4 Eldredge N. and Gould S. J., 1972.
Punctuated equilibria: an alternative tophyletic gradualism. In: T.J.M. Schopf (ed.),Models in Paleobiology. Freeman, Cooperand Company, San Francisco: 82-115.5 Alcuni approfondimenti sulla teoria degli
equilibri punteggiati si trovano in: www.talkorigins.org/faqs/punc-eq.html, ed in:www.skeptic.com/01.3.prothero-punc-eq.html.6 Gould S.J. and Lewontin R.C., 1979. The
Spandrels of San Marco and the Panglos-sian Paradigm. Proc. R. Soc. Lond., B 205(1161): 581-598. L�articolo può essere let-to, nella versione tradotta in italiano (�Ipennacchi di San Marco e il paradigma diPangloss�), in: www.einaudi.it/einaudi/ita/pdf/gould-lewontin.pdf.7 Interessanti documenti di approfondi-
mento del dibattito tra Gould ed i suoi op-positori sui temi dell�evoluzione biologicasi possono trovare in: www.world-of-dawkins.com/Catalano/the_g_files.htm.

Biol. Amb., 17 (n. 1, 2003)88
Informazione&Documentazione
RECENSIONI
Pier Francesco Ghetti
Elementi di Ecologia
CLEUP, Padova, 2002, 333pag., 22 euro
Il testo, pensato per risponde-re alle esigenze didattiche dellanuova riforma universitaria, pre-senta i principi dell�ecologia comeconcetti di valore universale, utiliad una vasta gamma di profili for-mativi. I temi trattati sono quelliclassici dell�ecologia: ecosistemi,individui, specie, popolazioni, rela-zioni trofiche, cicli astronomici, geo-logici, biogeochimici, processi agrande scala, comunità, bioindica-tori, paesaggi, biomi, ecologia uma-na, ecc.
Colpisce, invece, la singolareassociazione tra la semplicità dellinguaggio, volutamente privo diformalizzazione matematico-statisti-ca, la sua minuziosa appropriatez-za (ad es. nel distinguere gli ap-procci multidisciplinare, pluridisci-plinare, interdisciplinare, transdi-sciplinare), la costante cura etimo-logica, l�acutezza delle considera-zioni, i lucidi e sempre pertinentirichiami filosofici ed epistemologi-
ci, l�approccio critico ai metodi d�in-dagine, le avvertenze sulle possibi-lità d�errore (tra le quali è inclusala disonestà del ricercatore, menoinfrequente di quanto si creda).
Si tratta di un testo, in �con-trotendenza� rispetto agli abituali,voluminosi e costosi, che rischianodi diluire e far perdere di efficaciaai principi fondamentali dell�eco-logia. Per la piacevolezza e scorre-volezza della lettura, l�efficacia co-municativa, il lucido e aggiornatoinquadramento delle problematiche,sempre attento ad evitare ogni sem-plificazione concettuale, la profon-da cultura e lo spirito critico, ilvolume travalica gli obiettivi pro-postisi, risultando utile ad una �ri-messa a punto� delle conoscenzeanche per coloro già dotati di unaformazione ecologica.
Per ordinazioni: LibreriaCLEUP, Via G. Prati 19 - 35122 Pado-va (tel. 049 8753496; www.cleup.it).
Giuseppe Sansoni
Hydro-ecology: linking hydrologyand aquatic ecology. Mike Acre-man (Ed.). IAHS, 2001, IAHS Pro-ceedings and Reports 266. 162 pa-ges, figs, tabs, maps.
Hydrological models for environ-mental management. Edited by MVBolgov, L Gottschalk, I Krasovskaiaand RJ Moore (Eds.). Kluwer,Netherlands, 2002, 272 pages.
Rivers, technology and society.Learning the Lessons of Water Ma-
nagement in Nepal. Dipak Gyawa-li. Zed Books, 2003, 304 pages.
Water. Local-level management.David B Brooks. IDRC, Canada,2002, 80 pages.
Guidelines for the construction ofecological river networks. Councilof Europe, France, 2002, Nature andEnvironment Series 129. 40 pages.
Managing for healthy ecosystems.DJ Rapport, BL Lasley, DE Rolston,
CO Qualset, AB Damania and NONielsen. CRC Press, USA, 2002,1184 pages, 338 b/w illus.
Advances in water monitoring re-search. Tamim Younos (Ed.). Wa-ter Resources, USA, 2002, 244 pa-ges.
Fundamentals of ecotoxicology.Michael C Newman. CRC Press,USA, 2002, 402 pages, b/w photos,illus, figs, tabs.
LIBRI DI SCIENZE AMBIENTALI(informazioni e ordini on-line sul sito www.nhbs.com)

Biol. Amb., 17 (n. 1, 2003) 89
ABSTRACTS
1
2
3
4
5
6
7
8
9
10
11
12
13
Habitat integrity of running waters � assessment criteria andtheir biological relevance
Towards the assessment of �ecological integrity� in runningwaters of the United Kingdom
Scientific base and modular concept for comprehensive assess-ment of streams in Switzerland
The Austrian way of assessing the ecological integrity of run-ning waters: a contribution to the EU Water Framework Direc-tive
Sustaining living rivers
Assessing the ecological integrity of rivers: walking the lineamong ecological, political and administrative interests
Evaluating physical habitat integrity in relation to the biologi-cal potential of streams
Integrated ecological assessment methods as a basis for sustai-nable catchment management
Measuring the attainment of biological integrity in the USA: acritical element of ecological integrity
Fish as indicators for the assessment of the ecological integrityof large rivers
Fundamentals of fish ecological integrity and their relation tothe extended serial discontinuity concept
A multi-level concept for fish-based, river-type-specific assess-ment of ecological integrity
Evaluation of the use of landscape classification for the predic-tion of freshwater biota: synthesis and recommendations
Ecologia fluvialea cura di Pietro Genoni

Abstracts Biol. Amb., 17 (n. 1, 2003)90
S. MUHAR & M. JUNGWIRTH, 1998Habitat integrity of running waters � assessment criteria and their biologicalrelevanceHydrobiologia, 386: 195-202
L�habitat assessment, che sibasa su relazioni ben note tra lecomponenti abiotiche e biotiche diun sistema fluviale, può servire qua-le approccio indiretto nella valuta-zione dell�integrità ecologica, rive-lando il livello di alterazione del-l�ambiente fisico indotto dall�uo-mo. Un�analisi dettagliata dell�am-biente ha sempre rappresentato unaparte integrante delle ricerche auto-o sinecologiche negli ecosistemi ac-quatici. In numerosi Paesi, tutta-via, la valutazione dell�ambiente
fisico su una scala spaziale più am-pia è diventata una fase regolaredei programmi di gestione dei baci-ni idrografici.
Un aspetto metodologico diprimaria importanza nell�habitatassessment su ampia scala è rappre-sentato dalla scelta e dalla defini-zione dei criteri di valutazione, se-lezionati specificamente per rappre-sentare i processi e le funzioni chia-ve dei sistemi fluviali non alterati.Pertanto, un�enfasi particolare deveessere attribuita ai parametri inte-
grativi, che riflettono le dinamichespaziali e temporali dei corsi d�ac-qua, nonchè le interazioni tra ilfiume ed il territorio circostante.Nell�articolo sono discussi quattrogruppi di criteri di valutazione nelquadro della loro importanza perle biocenosi acquatiche e della lorovalidità nell�identificare gli impattiantropici sugli ecosistemi di acquecorrenti: (1) il regime idraulico, (2)il carattere morfologico, (3) la con-nettività laterale e (4) il corridoiolongitudinale.
1
Lo sviluppo di metodi per lavalutazione dell�integrità ecologicadelle acque correnti richiede l�inte-grazione dei parametri fisici e chi-mici, nonché dei loro effetti su strut-tura, diversità e processi biologici.La gestione dei fiumi nel RegnoUnito ha visto lo sviluppo di unaserie di metodi per la valutazione
D.M. HARPER, J.L. KEMP, B. VOGEL� & M.D. NEWSON, 2000Towards the assessment of �ecological integrity� in running waters of theUnited KingdomHydrobiologia, 422/423: 133-142
qualitativa, però scarsamente inte-grati tra loro. I metodi che valuta-no la struttura dell�habitat in ma-niera significativa dal punto di vi-sta biologico rappresentano l�approc-cio migliore per integrare le misuredegli effetti dei processi fisici e chi-mici sugli ambienti fluviali. Misuresimultanee dei tipi di flusso super-
ficiale (�biotopi di flusso�) e dellastruttura dell�habitat (�habitat fun-zionali�) hanno mostrato legamievidenti. Tali legami potrebbero rap-presentare la prima fase nello svi-luppo di un metodo rapido e pococostoso per la valutazione dell�inte-grità ecologica dei fiumi nel RegnoUnito.
2
In Svizzera esiste una dicoto-mia difficilmente conciliabile trala realtà e gli obiettivi ecologica-mente auspicabili nella gestionedegli ecosistemi fluviali. La mag-
U. BUNDI, A. PETER, A. FRUTIGER, M. HÜTTE, P. LIECHTI & U. SIEBER, 2000Scientific base and modular concept for comprehensive assessment of stre-ams in SwitzerlandHydrobiologia, 422/423: 477-487
gior parte dei corsi d�acqua è sotto-posta a una varietà di impatti. Acausa dell�attività antropica moltointensa si verifica un�elevata pres-sione sull�utilizzo del suolo e del-
l�acqua.Se da un lato gli usi essenzia-
li dell�acqua devono essere garanti-ti, dall�altra parte c�è anche unforte bisogno di conservare o ripri-
3

Biol. Amb., 17 (n. 1, 2003) Abstracts 91
stinare (riportare all�integrità eco-logica) o riabilitare (riportare�un�integrità ecologica relativa�) icorsi d�acqua. Pertanto, la valuta-zione dei corsi d�acqua dovrebbeprodurre dati idonei per la caratte-rizzazione della loro condizioneecologica e per supportare una lorogestione sostenibile. I metodi do-vrebbero prevedere un approcciosistemico e principi scientifici diintegrità ecologica appropriati, en-fatizzando le connettività tra glihabitat.
I metodi dovrebbero permette-re: (1) una descrizione ed un giudi-
zio razionale sulla condizione delcorso d�acqua, (2) l�identificazionedi diversi tipi di impatto sul corsod�acqua, (3) la verifica degli effettidelle misure di tutela, (4) l�identifi-cazione di idonee azioni future nelcontesto di un intero sistema flu-viale.
Al fine di soddisfare i diversirequisiti, è stato messo a punto unconcetto modulare per l�analisi deisistemi fluviali. Sono stati elaboraticontemporaneamente nove modu-li. Ciascun modulo affronta unospecifico aspetto (due moduli idro-dinamici ed ecomorfologici, cinque
moduli biologici, due moduli chi-mici ed ecotossicologici). I modulipossono essere applicati singolar-mente, in gruppo o tutti contempo-raneamente, in base allo scopo del-l�analisi. Per ciascun modulo ven-gono suggeriti una procedura dimonitoraggio economica (metodorapido) ed un metodo più rigoroso,in grado di fornire informazioni dimaggior dettaglio. In base ai risul-tati delle analisi, i concetti inerentiil ripristino fluviale possono esseresuccessivamente elaborati con lacollaborazione dei gestori, degli in-gegneri e dei biologi.
Il termine �OecologischeFunktionsfaehigkeit�, usato comesinonimo di integrità ecologica, èdiventato una parola chiave sia neiprocessi di gestione delle acque sianell�ambito della ricerca limnolo-gica in Austria. Incluso nelle prin-cipali normative riguardanti la ge-stione delle acque, il termine inte-grità ecologica rispecchia la neces-sità di considerare i corpi idricicome sistemi ecologici.
Sebbene l�integrità ecologicadei corsi d�acqua sia e debba rima-nere un concetto olistico, la meto-dologia attualmente adottata in
A. CHOVANEC, P. JÄGER, M. JUNGWIRTH, V. KOLLER-KREIMEL, O. MOOG, S. MUHAR & ST.SCHMUTZ, 2000The Austrian way of assessing the ecological integrity of running waters: acontribution to the EU Water Framework DirectiveHydrobiologia, 422/423: 445-452
Austria tende a valutarla attraversoun certo numero di componentichiave: gli aspetti idromorfologici(in particolare le strutture degli ha-bitat, il regime idrologico, la con-nettività), i parametri chimico-fisi-ci, le comunità dei macroinverte-brati e dei pesci, la stima della qua-lità biologica e le valutazioni eco-tossicologiche. La classificazione sibasa sulla valutazione di questi sin-goli criteri, per mezzo di un con-fronto tra una condizione di riferi-mento specifica per tipo di corsod�acqua e la condizione rilevata.Questo approccio, che è stato for-
mulato nell�Austrian Standard M6232 �Linee guida per lo studioecologico e la valutazione dei fiu-mi�, soddisfa anche le richieste ge-nerali per la classificazione dellostato ecologico dei corsi d�acquasecondo la Direttiva quadro euro-pea sulle acque.
Gli autori, oltre a descrivere ilmetodo adottato in Austria per va-lutare l�integrità ecologica delle ac-que correnti, presentano brevemen-te alcuni programmi nazionali cheriguardano la qualità delle acque,la valutazione degli habitat e letipologie fluviali.
4
J.R. KARR & E.W. CHU, 2000Sustaining living riversHydrobiologia, 422/423: 1-14
I fiumi non possono continua-re a soddisfare i bisogni della socie-tà, o degli esseri viventi in generale,
se l�uomo continua a considerarela gestione dei fiumi una questionepuramente politica od ingegneristi-
ca. Il flusso dei fiumi è parte di unflusso più grande, il ciclo planeta-rio dell�acqua, che sostiene non solo
5

Abstracts Biol. Amb., 17 (n. 1, 2003)92
il ciclo dell�acqua ma anche l�inte-ra rete della vita. In ultima analisi,la condizione, o la salute, del biotaacquatico rappresenta il modo mi-gliore di comprendere e controllarel�impatto umano sui corsi d�acquae sull�intero ciclo dell�acqua.
Il monitoraggio biologico, spe-cialmente gli approcci multimetriciquali l�indice di integrità biologica,riconosce l�importanza dell�integri-tà biotica dei fiumi ed offre uno deipiù potenti strumenti disponibili perla diagnosi, minimizzando e preve-
nendo la loro degradazione. Leampie prospettive offerte dalle va-lutazioni biologiche offrono miglioriopportunità di sostenere �fiumi vi-venti� rispetto ai limitati criteri chi-mici o alle misure convenzionalidello sviluppo urbano.
L�articolo prende in conside-razione le interazioni tra scienza,politica, amministrazione pubblicae opinione pubblica nel contestodella valutazione dello stato biolo-gico per quanto concerne il concet-to di integrità ecologica. La forteattenzione dell�opinione pubblicarispetto alla protezione dell�ambien-te e alla salute aiuta ad indirizzarele scelte politiche. Il mandato legi-slativo di �mantenere e ripristinarel�integrità ecologica delle acque� ècontemporaneamente un�afferma-zione di volontà politica ed un im-pegno. Sull�amministrazione pub-blica grava la responsabilità di unagestione efficiente delle risorse idri-
O. MOOG, A. CHOVANEC, 2000Assessing the ecological integrity of rivers: walking the line among ecological,political and administrative interestsHydrobiologia, 422/423: 99-109
che, con l�obiettivo di mantenereun�elevata qualità dello stato di sa-lute dell�ambiente. La crescentecomplessità delle prospettive suitemi ambientali aumenta il pesodelle scienze ecologiche sui proces-si amministrativi.
Gli ecologi giocano un ruoloessenziale nel progresso della cono-scenza scientifica, sviluppando me-todi per attuare i mandati legislati-vi ed educando l�opinione pubbli-ca (ed i funzionari pubblici) riguar-do i processi naturali e le interazio-ni, nonché le conseguenze ecologi-che delle attività umane. Una va-rietà di domande poste a livelloamministrativo riguardo l�inquina-
mento o l�alterazione fisica può tro-vare risposta nella valutazione bio-logica.
Con un�enfasi particolare suimacroinvertebrati bentonici ed illivello di identificazione tassono-mica, sono riportati esempi dellarealtà austriaca al fine di eviden-ziare gli sforzi necessari per mante-nere l�accuratezza scientifica sen-za perdere in praticità. Sono pre-sentati alcuni esempi di parametribasati sulle specie e sulle comunitàutilizzati in Austria per valutarel�integrità ecologica dei fiumi (es.aspetti saprobiologici, gruppi fun-zionali, analisi della zonazione lon-gitudinale).
6
C.F. RABENI, 2000Evaluating physical habitat integrity in relation to the biological potential ofstreamsHydrobiologia, 422/423: 245-256
7
L�integrità dell�ambiente fisi-co, sebbene sia una condizione ne-cessaria per l�integrità ecologica,non è definita con precisione e ra-ramente viene esaminata in rela-zione alla potenzialità biologica diun corso d�acqua. Le relazioni trahabitat fisico e biota necessitano di
essere quantificate in modo da defi-nire meglio le condizioni di riferi-mento e documentare quelle alte-razioni dell�habitat fisico che han-no ripercussioni sul biota.
L�autore ha valutato le rela-zioni tra l�ambiente fisico e le co-munità di invertebrati bentonici nei
corsi d�acqua del Missouri (USA),in siti di riferimento ed in siti alte-rati. Partendo da 45 corsi d�acquadi riferimento in tre ecoregioni, sonostati individuati sei tipi di habitatcomuni all�interno dell�alveo. A cia-scun tipo di habitat è stato associa-to un distinto raggruppamento di

Biol. Amb., 17 (n. 1, 2003) Abstracts 93
macroinvertebrati. È stata eviden-ziata una gerarchia spaziale del-l�influenza del tipo di habitat. Aduna scala più ampia, gli attributifisici propri delle ecoregioni si sonomostrati più influenti delle condi-zioni locali nel determinare la strut-tura delle comunità di invertebrati.All�interno di un�ecoregione, tutta-via, le condizioni dell�habitat supiccola scala sono più importantidella localizzazione del corso d�ac-
qua, poiché le comunità campio-nate nello stesso tipo di habitat incorsi d�acqua differenti sono risul-tate tra loro più simili rispetto aquella raccolte da differenti tipi dihabitat nello stesso corso d�acqua.
L�autore ha utilizzato questeinformazioni per definire le condi-zioni ambientali di riferimento peri corsi d�acqua di ciascuna ecore-gione. Successivamente sono statevalutate le comunità di 20 corsi
d�acqua con condizioni variabilidell�habitat per ottenere un model-lo della variazione e della sensibili-tà dei parametri biologici, al fine divalutare l�entità delle risposte delbiota alle alterazioni dell�habitat.Queste risposte biologiche sono sta-te documentate in modo rapido,tenendo conto sia della presenza odell�assenza di habitat particolarisia della qualità di ciascun tipo dihabitat.
P.F.M. VERDONSCHOT, 2000Integrated ecological assessment methods as a basis for sustainable catch-ment managementHydrobiologia, 422/423: 389-412
8
Nel corso del ventesimo seco-lo le problematiche ambientali sonocresciute, passando da quelle degliscarichi fognari dei primi decennia quelle conseguenti ai cambiamenticlimatici attuali. Un aumento del-la scala dei problemi implica unincremento di scala della loro ge-stione e valutazione. L�autore so-stiene che le valutazioni fisico-chi-miche, biologiche ed ecologiche sisono succedute, fallendo nel tenta-tivo di fermare il deterioramentoambientale.
Lo sviluppo delle tecniche cor-re parallelo all�evoluzione dei siste-mi di valutazione; si possono di-stinguere otto gruppi principali ditecniche di valutazione: gli indici(saprobici, di diversità, biotici), letecniche multimetriche e quelle spe-ditive, le tecniche fisico-ecologiche,le valutazioni a scala di bacino,delle componenti degli ecosistemi,delle comunità, dei processi e quel-le non tassonomiche. Se da un latosi osserva un aumento di raffina-tezza negli obiettivi, nelle misure enella complessità, dall�altro vi è unminor dettaglio dei livelli di scala,di tipologia di corso d�acqua e, mol-
to spesso, di livello tassonomico.In passato gli algoritmi utiliz-
zati erano tipicamente limitati asingole misure di sintesi, mentrel�analisi multivariata è stata intro-dotta solo di recente.
Le valutazioni possono essereeseguite senza una condizione diriferimento; tuttavia, tale condizio-ne è spesso utilizzata nei processivalutativi. Pertanto, nell�articoloviene discusso il ruolo della condi-zione di riferimento, concludendoche, qualora venga utilizzata, deveessere definita con precisione.
Viene studiato il ruolo nellavalutazione di nove concetti di eco-logia degli ambienti lotici, distin-guendone due gruppi principali:uno correlato al funzionamento deicorsi d�acqua a scala di bacino el�altro ai processi collegati agli ha-bitat fluviali e alla biodiversità. Unprimo tentativo di utilizzare questeconoscenze nei processi di gestioneè rappresentato dal modello delle5-S (5-S-Model), secondo cui l�ecosi-stema fluviale può essere suddivisoin cinque maggiori componenti: lecondizioni del sistema (system con-ditions), l�idrologia fluviale (stream
hydrology), le strutture (structures),le sostanze (substances) e le specie(species) [Verdonschot et al., 1998:The 5-S-Model, an integrated ap-proach for stream rehabilitation.In H.O. Hansen & B.L. Madsen,River Restoration �96, Session lec-tures proceedings. National Envi-ronmental Research Institute, Den-mark, International Conference ar-ranged by the European Centre forRiver Restoration: 36-44].
Infine, viene definita la valu-tazione ecologica integrata, basatasu tre principali approcci: di tipolo-gia ecologica, di bacino ecologico edi società. La tipologia ecologicaimplica un approccio a parametrimultipli aggregati, basato sulla ti-pologia ecologica regionale, dove letipologe sono messe in ordine discala incorporando differenti grup-pi tassonomici. La tipologia ecolo-gica pone le domande necessarie arendere sostenibile la gestione. Illivello di bacino ecologico implicaun approccio a scale multiple ag-gregate che accoppia le caratteristi-che naturali e antropiche con ledinamiche del bacino, sia nello spa-zio che nel tempo. L�approccio a

Abstracts Biol. Amb., 17 (n. 1, 2003)94
livello di società aggiunge le attivi-tà umane ai primi due approcci.Esso accoppia le domande dell�eco-
logia e gli utilizzi antropici e favo-risce le scelte che rendono sosteni-bile la gestione. Secondo l�autore,
l�applicazione di questi tre approccipuò rendere possibile una gestionesostenibile del bacino idrografico.
Il concetto di integrità ecolo-gica si è diffuso a livello mondialeed è fortemente radicato nella strut-tura normativa delle leggi ambien-tali degli USA. Il raggiungimentodell�integrità ecologica richiede ilraggiungimento dei suoi tre elementicostitutivi: l�integrità fisica, chimi-ca e biologica.
Negli USA, misure di integritàchimica furono introdotte per pri-me nei programmi di monitoraggioed ebbero successo nel ridurre icarichi inquinanti nelle acque su-perficiali. Poiché le comunità bio-logiche integrano gli effetti di diffe-renti cause di turbativa �come lariduzione di ossigeno, l�eccesso dinutrienti, le sostanze tossiche, gliincrementi di temperatura, l�ecces-so di sedimento e la degradazionedegli habitat� l�introduzione dellavalutazione biologica nei program-mi normativi ha fornito una strate-gia di monitoraggio ed una valuta-zione più completa ed efficace.
Le misure di integrità biologi-ca sono diventate una priorità ne-gli USA. Lo sviluppo di criteri bio-logici (biocriteria) all�interno dei
M.T. BARBOUR, S.K. JACKSON, D.L. COURTEMANCH, S.P. DAVIES, C.O.YODER, 2000Measuring the attainment of biological integrity in the USA: a critical elementof ecological integrityHydrobiologia, 422/423: 453-464
9
programmi normativi, al fine di ser-vire quali limiti con cui confronta-re il raggiungimento di condizioniprestabilite per la vita acquatica,rappresenta un punto focale im-portante negli USA. L�individuazio-ne di condizioni di riferimento perle acque superficiali (torrenti, fiu-mi, laghi, zone umide, estuari edambienti marini) attraverso diffe-renti regioni fisiografiche rappre-senta un elemento critico nella de-finizione dei criteri biologici ed èattualmente un�iniziativa di prima-ria importanza. Quasi tutte le agen-zie statali per le risorse idriche han-no sviluppato approcci per la valu-tazione biologica dei corsi d�acqua;in ciascun stato, da 1.600 a 75.000km di corsi d�acqua necessitanouna valutazione.
Lo sviluppo della valutazionebiologica per altri corpi idrici nonè attualmente ad uno stadio cosìavanzato. La US-EPA ha prodottoguide tecniche per attuare efficaciprogrammi di biomonitoraggio; esseincludono gli elementi cruciali,quali: la definizione degli obiettivi,la classificazione dei corpi idrici
rispetto alle caratteristiche biologi-che attese, la definizione della con-dizione di riferimento per le diversetipologie di sito, lo sviluppo di pro-tocolli standardizzati per il cam-pionamento e l�analisi dei dati, lamessa a punto di un piano perl�assicurazione di qualità. Gli ap-procci alla valutazione biologicanegli USA seguono uno schema dibase che incorpora vari attributipropri degli elementi e dei processidella comunità acquatica; tale sche-ma può essere un�aggregazione inun indice multimetrico, ovvero unaserie di analisi multivariate che uti-lizzano gli attributi quali variabilidi input.
Il Clean Water Act del 1972ed i suoi successivi emendamentimiravano a mantenere, ripristinaree proteggere l�integrità ecologicadelle acque superficiali. Attraversol�impiego di metodi robusti per lavalutazione biologica e di altri me-todi di misura dell�integrità ecolo-gica, gli USA hanno sviluppato unpiano strategico per stabilire le prio-rità che permettono di raggiungerequesto obiettivo.
F. SCHIEMER, 2000Fish as indicators for the assessment of the ecological integrity of large riversHydrobiologia, 422/423: 271-278
Le comunità ittiche nei gran-di fiumi sono caratterizzate daun�elevata diversità, la quale riflet-
te la diversità strutturale e la ric-chezza di habitat delle zone ripa-rie e delle zone golenali ad esse
connesse. La connettività dei diffe-renti elementi ambientali in unampio contesto spazio-temporale �
10

Biol. Amb., 17 (n. 1, 2003) Abstracts 95
ossia a varie scale, dal bacino almicrohabitat� ed il loro raggruppa-mento, definiscono l�adattamentodelle specie ittiche a livello sia diindividuo (ad esempio, la perfor-mance di crescita) sia di popolazio-ne (struttura di popolazione, mor-talità, ecc.). Le scale spaziali impor-tanti possono essere l�intero corsodel fiume nel caso dei migratorianadromi, oppure la disponibilitàdi microhabitat complementari, adesempio di quelli necessari durantela fase iniziale dello sviluppo diuna specie.
Il significato della connettivi-tà a differenti scale, dall�intero fiu-me al tratto locale, deve essere va-lutato sulla base delle esigenze, deicomportamenti e della flessibilità
ecologica delle singole specie.L�integrità deve essere valuta-
ta in numerosi aspetti: (1) per ciòche riguarda la genetica di popola-zione in estese aree biogeografichee per lunghe scale temporali; (2)per ciò che riguarda gli habitat sup-plementari nel ciclo vitale di singo-le specie che diversificano gli habi-tat durante lo sviluppo e mostranoesigenze specifiche durante la faseriproduttiva: (3) per ciò che riguar-da il trasporto longitudinale e late-rale ed i processi di scambio chedeterminano le condizioni locali el�apporto di cibo per i pesci.
A causa di queste interdipen-denze, la condizione della faunaittica è un sensore critico dell�inte-grità alle differenti scale di detta-
glio e quindi un buon strumento dimonitoraggio, specialmente nei con-fronti dell�impatto di interventi diingegneria fluviale.
La struttura dei popolamentiittici �in particolare l�abbondanzarelativa delle specie adattate allezone golenali� fornisce informazio-ni circa le condizioni complessive.Per una valutazione più dettaglia-ta, i modelli di distribuzione localedella fauna ittica, la struttura dipopolazione, la stagionalità ed ilsuccesso di crescita degli stadi gio-vanili possono indicare la qualità el�estensione della connettività aduna scala più fine e possono essereutilizzati per stabilire i criteri ge-stionali ed ingegneristici.
Le teorie ed i concetti ecologi-ci attuali descrivono i corsi d�ac-qua come sistemi a quattro dimen-sioni, con legami, interazioni e pro-cessi di scambio longitudinali, late-rali e verticali che variano nel tem-po ed alle differenti scale. Secondoil concetto della discontinuità se-riale estesa (ESDC) formulato daWard e Stanford (1983), in un mo-dello di fiume a tre tratti, la poten-za relativa della dimensione longi-tudinale è massima in prossimitàdelle sorgenti, le interazioni verti-cali raggiungono il massimo doveil medio corso si anastomizza, men-tre la connettività laterale gioca unruolo di maggiore importanza neitratti di pianura.
Nell�articolo, gli autori esa-minano i principi generali del con-cetto ESDC dal punto di vista del-
M. JUNGWIRTH, S. MUHAR, S. SCHMUTZ, 2000Fundamentals of fish ecological integrity and their relation to the extendedserial discontinuity conceptHydrobiologia, 422/423: 85-97
l�ecologia della fauna ittica. In par-ticolare, essi indagano fino a chepunto le esigenze di connettivitàspaziale/temporale delle comunitàittiche, o di specie ittiche chiave,lungo il percorso longitudinale sche-matico di un fiume, siano compati-bili con i principi del concettoESDC, e se queste esigenze possanofornire i criteri di base per valutarel�integrità ecologica dei corsi d�ac-qua dal punto di vista dell�ecologiadell�ittiofauna.
Gli esempi riportati nell�arti-colo mostrano, sul piano teorico,un accordo con il concetto dellanatura a quattro dimensioni deicorsi d�acqua, i cui elementi fun-zionali e strutturali sono rappre-sentati dalle dinamiche fluviali spa-ziali/temporali, dai disturbi, dallaconnettività, dalle successioni e da-
gli ecotoni. In particolare, i pescisono utili indicatori delle connetti-vità che variano nel tempo nell�in-tero ambito dei diversi dettagli discala e nelle tre dimensioni spazia-li.
Due modifiche principali delconcetto ESDC sono necessariequando si considerano le esigenzedi habitat di specie tipiche o deidiversi stadi di sviluppo. Nelle sor-genti ad alveo incanalato, oltre allaconnettività longitudinale, deve es-sere enfatizzata l�importanza cru-ciale del rapporto verticale tra ilfiume ed i sedimenti ai fini ripro-duttivi. Inoltre, in aggiunta alla con-nettività laterale, la connettività lon-gitudinale rappresenta una base vi-tale per le comunità ittiche pota-mali nelle zone anastomizzate emeandriformi.
11

Abstracts Biol. Amb., 17 (n. 1, 2003)96
S. SCHMUTZ, M. KAUFMANN, B. VOGEL, M. JUNGWIRTH, S. MUHAR, 2000A multi-level concept for fish-based, river-type-specific assessment of ecologi-cal integrityHydrobiologia, 422/423: 279-289
12
Gli Autori propongono un con-cetto multi-livello per la valutazio-ne dell�integrità ecologica dei corsid�acqua basata sui pesci (MuLFA).Questo concetto è adatto ai pro-grammi di monitoraggio su ampiascala, come quelli previsti dalla Di-rettiva quadro europea sulle acque.Su cinque livelli di organizzazionebiologica �fauna, comunità, gilda(cioè gruppo funzionale), popola-zione, individuo�, sono proposti settecriteri: specie specifiche per tipolo-gia fluviale, specie con popolazioni
auto-mantenute, regioni ittiche,numero di consorzi, composizionedel consorzio, densità di popolazio-ne e struttura di età della popola-zione. Il principio del MuLFA èbasato sulla stima dello scostamen-to da condizioni di riferimento in-disturbate. Le condizioni di riferi-mento devono essere descritte perciascuna distinta tipologia fluvialeutilizzando dati storici sulla faunaittica e sulle caratteristiche abioti-che, siti di riferimento attuali speci-fici per fiume e modelli di riferi-
mento. La procedura di valutazio-ne finale è realizzata confrontandoil tratto esaminato con le condizio-ni di riferimento, utilizzando unaschema di cinque passaggi ed asse-gnando quel tratto al livello di con-cordanza più elevato. Il vantaggiodel MuLFA risiede nella sua poten-zialità nel discriminare in manierasensibile le alterazioni antropichea bassa ed elevata intensità, e gra-zie al suo carattere generale, nellasua adattabilità a tutte le tipologiefluviali.
C.P. HAWKINS, R.H. NORRIS, J. GERRITSEN, R.M. HUGHES, S.K. JACKSON, R.K. JOHNSON,R.J. STEVENSON, 2000Evaluation of the use of landscape classification for the prediction of freshwa-ter biota: synthesis and recommendationsJournal of the North American Benthological Society, 19 (3): 541-556
L�articolo sintetizza i risultaticollettivi che emergono dalla seriedi lavori pubblicati nel volume 19(3) del J-NABS, e li colloca nel con-testo della letteratura sulle varia-zioni delle biocenosi acquatiche inrapporto al paesaggio. Le classifica-zioni basate sulle scale spaziali delpaesaggio sono utilizzate, o sonostate valutate, per l�impiego nei pro-grammi di monitoraggio biologicoin numerosi Paesi. La valutazionedella robustezza della classificazio-ne secondo i differenti approcci do-vrebbe fornire elementi per il mi-glioramento dei programmi di bio-monitoraggio esistenti ed accelera-re lo sviluppo di nuovi programmi.Gli articoli pubblicati nel volumediscutono a quale livello le descri-zioni e la classificazione delle ca-ratteristiche del paesaggio permet-tono di spiegare, e quindi prevede-
re, la variazione nella composizio-ne delle biocenosi tra singoli siti.In generale, gli autori riscontranoche, sebbene le classificazioni delpaesaggio spieghino la variazionebiologica più di quanto si sarebbecasualmente atteso, l�entità di va-riazione correlata alle caratteristi-che del paesaggio non è grande.Pertanto, le regionalizzazioni suampia scala, se utilizzate da soleper definire le condizioni biologi-che attese, avranno probabilmenteun uso limitato nel biomonitorag-gio dei corsi d�acqua, dove risultacruciale specificare le condizioniattese nel modo più accurato possi-bile. Le classificazioni del paesag-gio possono comunque giocare unruolo aggiuntivo fornendo un�ini-ziale stratificazione dei siti in ma-niera da assicurarsi che differenticaratteristiche del paesaggio risulti-
no adeguatamente rappresentate inun programma di campionamento.In generale, gli autori ritengononecessaria una classificazione ge-rarchica, basata sia sul singolo trat-to sia su una scala più ampia dipaesaggio, al fine di predire accura-tamente la composizione della fau-na acquatica. Un potenziale ap-proccio implica l�uso delle classifi-cazioni del paesaggio quale stru-mento per rifinire o incrementarele classificazioni basate sulle carat-teristiche locali degli habitat, lequali sembrano spiegare sostanzial-mente meglio le variazioni biologi-che rispetto alle caratteristiche am-bientali di più ampia scala. Questirisultati hanno implicazioni signi-ficative per quanto concerne le mo-dalità di pianificazione dei progettidi valutazione e monitoraggio a li-vello locale, regionale e nazionale.
13

BIOLOGIAAMBIENTALE ISTRUZIONI PER GLI AUTORI
Manoscritti. I lavori proposti per la pubblicazione devono esserescritti su un unico lato del foglio e, compatibilmente con il lorocontenuto, devono essere suddivisi in: introduzione, materiali emetodi, risultati, discussione, eventuali ringraziamenti, bibliografia,tabelle, figure. Qualora il lavoro sia già stato pubblicato o sottopo-sto all�attenzione di altri editori, la circostanza deve essere chiara-mente segnalata: in tal caso il lavoro potrà essere preso in conside-razione solo per la recensione nella sezione Informazione & Docu-mentazione.
Titolo e Autori. Il titolo deve essere informativo e, se possibile, con-ciso; deve essere indicato anche un titolo breve (massimo cinquantacaratteri) da utilizzare come intestazione delle pagine successive allaprima. Il titolo deve essere seguito dal cognome e dal nome (peresteso) di tutti gli autori. Di ogni autore (contrassegnato da un ri-chiamo numerico) deve essere riportato l�indirizzo postale completodell�istituto nel quale è stato svolto lo studio. Il nome dell�autorereferente per la corrispondenza con la redazione e con i lettori deveessere contrassegnato anche da un asterisco; il suo indirizzo di po-sta ordinaria deve essere seguito anche dal numero di telefono, difax e dall�indirizzo di posta elettronica; soltanto tramite quest�ultimosaranno inviate le bozze per la correzione.
Riassunto, abstract e parole chiave . Il riassunto (lunghezza massi-ma 250 parole) deve sintetizzare lo scopo dello studio, descrivere gliesperimenti, i principali risultati e le conclusioni; deve essere segui-to dalle parole chiave, separate da una barra obliqua. Devono esse-re altresì riportati in lingua inglese il titolo e un abstract (massimo250 parole), seguiti dalle key words separate da una barra obliqua.
Figure e tabelle. Le figure, con la loro didascalia al piede e nume-rate con numeri arabi, non devono essere inserite nel testo, ma infogli separati alla fine del testo. È gradita l�indicazione, a penna,della posizione preferita per l�inserzione di ciascuna figura neldattiloscritto. Anche le tabelle devono essere riportate in fogli sepa-rati, alla fine del dattiloscritto; devono essere complete di titolo enumerate con numeri romani. Occorre curare titoli, legende edidascalie in modo da rendere le tabelle e le figure autosufficienti,comprensibili cioè anche senza consultare il testo. Per le figure (gra-fici, disegni o fotografie di buona qualità), si raccomanda agli auto-ri di verificare con opportune riduzioni l�aspetto finale e la leggibilitàdelle scritte, tenendo conto che verranno stampate riducendone labase a 80 mm (una colonna) o 170 mm (due colonne). Non inviarefotografie o grafici a colori senza essersi accertati che la loro stampain bianco e nero assicuri comunque l�agevole riconoscibilità dellediverse sfumature o retinature. Nella scelta degli accorgimenti grafi-ci privilegiare sempre la facilità e immediatezza di lettura agli effet-ti estetici.
Bibliografia. Al termine del testo deve essere riportata la bibliografiain ordine alfabetico. Ad ogni voce riportata nell�elenco bibliograficodeve necessariamente corrispondere il riferimento nel testo e vice-versa. Per il formato tipografico e la punteggiatura, attenersi stretta-mente ai seguenti esempi:Dutton I.M., Saenger P., Perry T., Luker G., Worboys G.L., 1994. An
integrated approach to management of coastal aquatic resources.A case study from Jervis Bay, Australia. Aquatic Conservation:marine and freshwater ecosystems, 4: 57-73.
Hellawell J.M., 1986. Biological indicators of freshwater pollution andenvironmental management. Elsevier Applied Science Publishers,London and New York, 546 pp.
Pulliam H.R., 1996. Sources and sinks: empirical evidence and popu-lation consequences. In: Rhodes O.E., Chesser R.K., Smith M.H.(eds.), Population dynamics in ecological space and time. The Uni-versity of Chicago Press, Chicago: 45-69.
Corbetta F., Pirone G., (1986-1987) 1988. I fiumi d�Abruzzo: aspettidella vegetazione. In: Atti Conv. Scient. �I corsi d�acqua minoridell�Italia appenninica. Aspetti ecologici e gestionali�, Aulla (MS),22-24 giugno 1987. Boll. Mus. St. Nat. Lunigiana 6-7: 95-98.
Proposte di pubblicazione. Due copie del manoscritto su supportocartaceo devono essere inviate a:
Redazione di Biologia Ambientale,ARPAT, Via del Patriota 2 � 54100 Massa (MS) � I
c.a. Giuseppe SansoniIl manoscritto deve essere accompagnato da una copia su supportomagnetico; in alternativa, quest�ultima può essere inviata all�indi-rizzo di posta elettronica [email protected] manoscritti saranno sottoposti alla lettura di revisori scientifici; entrodue mesi l�autore indicato come referente per la corrispondenza saràinformato delle decisioni della redazione. Per evitare ritardi nellapubblicazione e revisioni del testo, si raccomanda vivamente agliautori di prestare la massima cura anche alla forma espositiva chedeve essere concisa, chiara, scorrevole e in buon italiano, evitandoneologismi superflui. Tutte le abbreviazioni e gli acronimi devonoessere definiti per esteso alla loro prima occorrenza nel testo. I nomiscientifici delle specie devono essere sottolineati (saranno convertiti incorsivo prima della stampa).I dattiloscritti, compreso il materiale illustrativo, non verranno restitu-iti, salvo esplicita richiesta dell�autore all�atto dell�invio del materiale.La redazione si riserva il diritto di apportare ritocchi linguistici egrafici e di respingere i manoscritti che non rispettano i requisiti dellepresenti istruzioni per gli autori. Le opinioni espresse dagli autorinegli articoli firmati non rispecchiano necessariamente le posizionidel C.I.S.B.A.
Bozze ed estratti. Le bozze di stampa verranno inviate all�autoreindicato come referente per la corrispondenza, che deve impegnarsiad una correzione molto accurata e al nuovo invio alla redazioneentro 5 giorni; trascorso tale periodo, il lavoro può essere pubblicatocon le sole correzioni dell�editore. All�autore referente per la corri-spondenza verrà inviato il numero della rivista e, tramite e-mail, ilfile dell�estratto in formato *.PDF, utilizzabile per riprodurre il nu-mero desiderato di estratti.
Formato dei file. Oltre al manoscritto vanno inviati, su supportomagnetico, i relativi file. Per assicurare la compatibilità con tutti iprogrammi di videoscrittura e di impaginazione il file contenente iltesto va inviato in triplice versione: formato solo testo (*.txt), richtext format (*.rtf) e WinWord (*.doc, preferibilmente salvato nel for-mato della sua penultima versione commerciale). I grafici devonoessere in bianco e nero ed essere sempre accompagnati dalla tabelladei dati di origine; per quelli realizzati con fogli elettronici inviare ilfile contenente i grafici e i dati di origine (preferibilmente salvatonella penultima versione commerciale di Excel) al fine di consenti-re eventuali modifiche al formato volte a migliorarne la leggibilità. Ifile delle figure al tratto vanno inviati preferibilmente in formato*.tif; quelli delle fotografie preferibilmente in formato *.jpg. Per for-mati di file diversi da quelli sopra indicati, precisare il software uti-lizzato. Per ogni chiarimento tecnico contattare Giuseppe Sansoni(tel. 0585 899409, fax 0585 47000, e-mail: [email protected]).

BIOLOGIAAMBIENTALE
Spediz. in abbon. post, art. 2, comma 20/c, L. 662/96, filiale Reggio EmiliaTassa pagata - Taxe perçue
Volume 17Numero 1Gennaio 2003
Info
rmaz
ione
&do
cum
enta
zion
e
1 PIGNATTI S. - Il declino della biodiversità nelladimensione territoriale
9 GENONI P. - Influenza di alcuni fattori ambientalisulla composizione delle cenosi macrobentonichedei corsi d’acqua planiziali minori
17 SANSONI G., BORGHINI B., CAMICI G., CASOTTI
M., RIGHINI P., RUSTIGHI C. - Fioriture algali diOstreopsis ovata (Gonyaulacales: Dinophy-ceae): un problema emergente
25 PIZZUL E., BOTTOS P., VANZO S., TOSOLINI G. -Osservazioni preliminari sulla predazione dellaTrota fario e della Trota marmorata nei confrontidei macroinvertebrati bentonici presenti nel driftdel torrente But (Bacino del fiume Tagliamento,Friuli Venezia Giulia)
35 ANDREANI P., SANTUCCI F., NASCETTI G. - Lerane verdi del complesso Rana esculenta comebioindicatori della qualità degli ambienti fluvialiitaliani
45 ROSSI S., SIMONE V., NARDI P.A., MONTAGNA S.- Effetti delle asciutte sulle cenosi macrobento-niche: taxa sensibili, resistenti e resilienti
55 ROSSI P. e FERRARINI A., ROSSI O., ZURLINI G. -Analisi della struttura del paesaggio mediante datitelerilevati del sensore MIVIS: il bacino del tor-rente Baganza (Parma)
67 FERRARINI A. e ROSSI P., ROSSI O. - Analisi del-l’evoluzione del paesaggio mediante l’approcciogeostatistico ai dati telerilevati: il bacino del tor-rente Baganza (Parma)
SOMMARIO
RECENSIONIABSTRACTS
Lavo
ri or
igin
ali
RESOCONTI 77 ROCCA D., BODON M., GAITER S., CASARINO E. -Resoconto sul Corso di Formazione “Metazoidelle acque sorgive e sotterranee. Biologia,campionamento e riconoscimento, aspetti am-bientali ed applicativi”
85 GENONI P. - Un’idea della vita: ricordo di StephenJay Gould
88 Recensioni89 Ecologia fluviale
ATTUALITÀ


















