
Apprendimento e individualità: le basi...
6
Apprendimento e individualità: le basi biologiche Recenti scoperte hanno evidenziato che l'apprendimento si basa su semplici insiemi di regole e processi che, agendo sulle connessioni fra i neuroni, contribuiscono a rendere unico ciascun individuo di Eric R. Kandel e Robert D. Hawkins N egli ultimi decenni, due campi della scienza originariamente separati (la neurobiologia, la scienza del cervello, e la psicologia co- gnitivista, la scienza della mente) si sono andati gradualmente fondendo. In tempi recenti, il processo di unificazione è di- ventato ancora più rapido, e ne è emerso un nuovo quadro di riferimento per lo studio della percezione, del linguaggio, della memoria e della coscienza, che si basa sulla possibilità di studiare i sub- strati biologici di queste funzioni men- tali. L'apprendimento ce ne offre un esempio affascinante. Oggi è possibile studiare a livello cellulare, addirittura molecolare, gli aspetti elementari dei meccanismi importanti per molti tipi di- versi di apprendimento; l'analisi dell'ap- prendimento, quindi, può darci una pri- ma idea dei meccanismi molecolari che stanno alla base di un processo mentale e può quindi contribuire alla costruzione di un ponte fra psicologia cognitivista e biologia molecolare. L'apprendimento è il processo me- diante il quale acquisiamo nuova cono- scenza, e la memoria è il processo con il quale conserviamo nel tempo questa conoscenza. Quasi tutto ciò che sappia- mo del mondo e delle sue civiltà l'ab- biamo appreso. Apprendimento e me- moria, quindi, sono fondamentali per il nostro senso di individualità. L'appren- dimento va oltre l'individuo: è trasmis- sione di cultura di generazione in gene- razione; è un veicolo importante di adat- tamento comportamentale e una potente forza di progresso sociale. Viceversa, la perdita di memoria porta a una perdita di contatto con l'io immediato, con la propria storia e con gli altri esseri umani. Fino alla metà del ventesimo secolo, la maggior parte degli studiosi del com- portamento non pensava che la memoria fosse una funzione mentale distinta, in- dipendente da movimento, percezione, attenzione e linguaggio. Anche quando queste funzioni erano state localizzate ormai da tempo in regioni diverse del cervello, si continuava a dubitare che sa- rebbe mai stato possibile assegnare la memoria a una regione specifica. Il pri- mo che lo abbia fatto è stato Wilder G. Penfield, neurochirurgo del Montreal Neurological Institute. Negli anni quaranta Penfield comin- ciò a utilizzare la stimolazione elettrica per individuare la localizzazione di fun- zioni motorie, sensoriali e del linguaggio nella corteccia di pazienti sottoposti a interventi neurochirurgici per la cura dell'epilessia. Dato che il cervello in sé non possiede recettori del dolore, è pos- sibile intervenire chirurgicamente sul cervello in anestesia locale, con il pa- ziente quindi perfettamente cosciente e in grado di descrivere che cosa prova in risposta a stimoli elettrici applicati ad aree corticali diverse. Penfield esplorò la superficie corticale di oltre mille pazien- ti osservando talvolta che la stimolazio- ne elettrica produceva una risposta in cui il paziente descriveva un ricordo coeren- te di un'esperienza passata. Queste ri- sposte, simili a ricordi, venivano ottenu- te invariabilmente dai lobi temporali. Ulteriori dati a sostegno del ruolo dei lobi temporali nella memoria sono stati ottenuti negli anni cinquanta dallo studio di pazienti che avevano subito la rimo- zione bilaterale dell'ippocampo e delle regioni adiacenti del lobo temporale per il trattamento dell'epilessia. Nel primo caso, che è anche il meglio approfondi- to, Brenda Milner del Montreal Neuro- logical Institute ha descritto un operaio di 27 anni, H. M., addetto a una catena di montaggio, che per oltre 10 anni ave- va subito attacchi epilettici, non trattabili e debilitanti, con origine nel lobo tem- porale. Il chirurgo, William B. Scoville, aveva rimosso la porzione centrale dei lobi temporali su ambo i lati del cervello di H. M. I disturbi migliorarono sensi- bilmente, ma subito dopo l'intervento il paziente accusò una drastica perdita di memoria: aveva perso la capacità di for- mare nuovi ricordi a lungo termine. Nonostante avesse difficoltà a forma- re nuovi ricordi, H. M. conservava però tutti i ricordi di lungo termine acquisiti in precedenza. Ricordava il suo nome, conservava una buona padronanza del linguaggio e un vocabolario normale; il suo quoziente di intelligenza era ancora nella fascia del «brillante-normale». Ri- cordava bene gli eventi che avevano pre- ceduto l'intervento, per esempio il tipo di lavoro che svolgeva, e aveva un ricor- do vivo della sua fanciullezza. Inoltre, H. M. aveva ancora, perfettamente intat- ta, la facoltà della memoria a breve ter- mine. Quello che gli era venuto a man- care, e in profondità, era la capacità di trasferire ciò che imparava dalla memo- ria a breve termine a quella a lungo ter- mine. Per esempio, poteva conversare normalmente con il personale della cli- nica, ma non si ricordava dei singoli in- dividui pur vedendoli quotidianamente. Inizialmente si pensava che questo deficit di memoria a seguito di lesioni L'esperimento del disegno allo specchio nel caso di pazienti con lesioni ai lobi tem- porali indicò nel 1960 l'esistenza di due diversi sistemi di apprendimento. Uno, che non è pregiudicato da queste lesioni, riguarda compiti con un certo grado di auto- matismo, come quello qui illustrato: il soggetto, che vede la propria mano solo nello specchio, cerca di ricalcare una stella. Il secondo tipo di apprendimento dipende dalla coscienza e dai processi cognitivi ed è cancellato dalle lesioni ai lobi temporali. LE SCIENZE n. 291, novembre 1992 49
Transcript of Apprendimento e individualità: le basi...

Apprendimento e individualità:le basi biologiche
Recenti scoperte hanno evidenziato che l'apprendimento si basa susemplici insiemi di regole e processi che, agendo sulle connessionifra i neuroni, contribuiscono a rendere unico ciascun individuo
di Eric R. Kandel e Robert D. Hawkins
N
egli ultimi decenni, due campidella scienza originariamenteseparati (la neurobiologia, la
scienza del cervello, e la psicologia co-gnitivista, la scienza della mente) si sonoandati gradualmente fondendo. In tempirecenti, il processo di unificazione è di-ventato ancora più rapido, e ne è emersoun nuovo quadro di riferimento per lostudio della percezione, del linguaggio,della memoria e della coscienza, che sibasa sulla possibilità di studiare i sub-strati biologici di queste funzioni men-tali. L'apprendimento ce ne offre unesempio affascinante. Oggi è possibilestudiare a livello cellulare, addiritturamolecolare, gli aspetti elementari deimeccanismi importanti per molti tipi di-versi di apprendimento; l'analisi dell'ap-prendimento, quindi, può darci una pri-ma idea dei meccanismi molecolari chestanno alla base di un processo mentalee può quindi contribuire alla costruzionedi un ponte fra psicologia cognitivista ebiologia molecolare.
L'apprendimento è il processo me-diante il quale acquisiamo nuova cono-scenza, e la memoria è il processo conil quale conserviamo nel tempo questaconoscenza. Quasi tutto ciò che sappia-mo del mondo e delle sue civiltà l'ab-biamo appreso. Apprendimento e me-moria, quindi, sono fondamentali per ilnostro senso di individualità. L'appren-dimento va oltre l'individuo: è trasmis-sione di cultura di generazione in gene-razione; è un veicolo importante di adat-tamento comportamentale e una potente
forza di progresso sociale. Viceversa, laperdita di memoria porta a una perditadi contatto con l'io immediato, con lapropria storia e con gli altri esseri umani.
Fino alla metà del ventesimo secolo,la maggior parte degli studiosi del com-portamento non pensava che la memoriafosse una funzione mentale distinta, in-dipendente da movimento, percezione,attenzione e linguaggio. Anche quandoqueste funzioni erano state localizzateormai da tempo in regioni diverse delcervello, si continuava a dubitare che sa-rebbe mai stato possibile assegnare lamemoria a una regione specifica. Il pri-mo che lo abbia fatto è stato Wilder G.Penfield, neurochirurgo del MontrealNeurological Institute.
Negli anni quaranta Penfield comin-ciò a utilizzare la stimolazione elettricaper individuare la localizzazione di fun-zioni motorie, sensoriali e del linguaggionella corteccia di pazienti sottoposti ainterventi neurochirurgici per la curadell'epilessia. Dato che il cervello in sénon possiede recettori del dolore, è pos-sibile intervenire chirurgicamente sulcervello in anestesia locale, con il pa-ziente quindi perfettamente cosciente ein grado di descrivere che cosa prova inrisposta a stimoli elettrici applicati adaree corticali diverse. Penfield esplorò lasuperficie corticale di oltre mille pazien-ti osservando talvolta che la stimolazio-ne elettrica produceva una risposta in cuiil paziente descriveva un ricordo coeren-te di un'esperienza passata. Queste ri-sposte, simili a ricordi, venivano ottenu-
te invariabilmente dai lobi temporali.Ulteriori dati a sostegno del ruolo dei
lobi temporali nella memoria sono statiottenuti negli anni cinquanta dallo studiodi pazienti che avevano subito la rimo-zione bilaterale dell'ippocampo e delleregioni adiacenti del lobo temporale peril trattamento dell'epilessia. Nel primocaso, che è anche il meglio approfondi-to, Brenda Milner del Montreal Neuro-logical Institute ha descritto un operaiodi 27 anni, H. M., addetto a una catenadi montaggio, che per oltre 10 anni ave-va subito attacchi epilettici, non trattabilie debilitanti, con origine nel lobo tem-porale. Il chirurgo, William B. Scoville,aveva rimosso la porzione centrale deilobi temporali su ambo i lati del cervellodi H. M. I disturbi migliorarono sensi-bilmente, ma subito dopo l'intervento ilpaziente accusò una drastica perdita dimemoria: aveva perso la capacità di for-mare nuovi ricordi a lungo termine.
Nonostante avesse difficoltà a forma-re nuovi ricordi, H. M. conservava peròtutti i ricordi di lungo termine acquisitiin precedenza. Ricordava il suo nome,conservava una buona padronanza dellinguaggio e un vocabolario normale; ilsuo quoziente di intelligenza era ancoranella fascia del «brillante-normale». Ri-cordava bene gli eventi che avevano pre-ceduto l'intervento, per esempio il tipodi lavoro che svolgeva, e aveva un ricor-do vivo della sua fanciullezza. Inoltre,H. M. aveva ancora, perfettamente intat-ta, la facoltà della memoria a breve ter-mine. Quello che gli era venuto a man-care, e in profondità, era la capacità ditrasferire ciò che imparava dalla memo-ria a breve termine a quella a lungo ter-mine. Per esempio, poteva conversarenormalmente con il personale della cli-nica, ma non si ricordava dei singoli in-dividui pur vedendoli quotidianamente.
Inizialmente si pensava che questodeficit di memoria a seguito di lesioni
L'esperimento del disegno allo specchio nel caso di pazienti con lesioni ai lobi tem-porali indicò nel 1960 l'esistenza di due diversi sistemi di apprendimento. Uno, chenon è pregiudicato da queste lesioni, riguarda compiti con un certo grado di auto-matismo, come quello qui illustrato: il soggetto, che vede la propria mano solo nellospecchio, cerca di ricalcare una stella. Il secondo tipo di apprendimento dipendedalla coscienza e dai processi cognitivi ed è cancellato dalle lesioni ai lobi temporali.
LE SCIENZE n. 291, novembre 1992 49
NEURONESENSORIALE
PRESINAPTICO
FOSFOLIPASI A
LIBERAZIONEINTENSIFICATA
DI TRASMETTITORE
CANALEDEL POTASSIO------,...„.
CANALEDEL CALCIO-
VE SCICOLADI NEURO-
TRASMETTITORE
SITO DILIBERAZIONE
NEURONEMODULATORE
ADENILIL-CICLASI
CALMODULINA
CANALEDEL CALCIO
o RECETTOREDELLASEROTONINA
DIACILGLICEROLO----
AMPCICLICO
CALCIO
PROTEIN-CHINASIDELL'AMPCICLICO
PROTEIN-CHINASI C
NEURONEMOTORE
POSTSINAPTICO
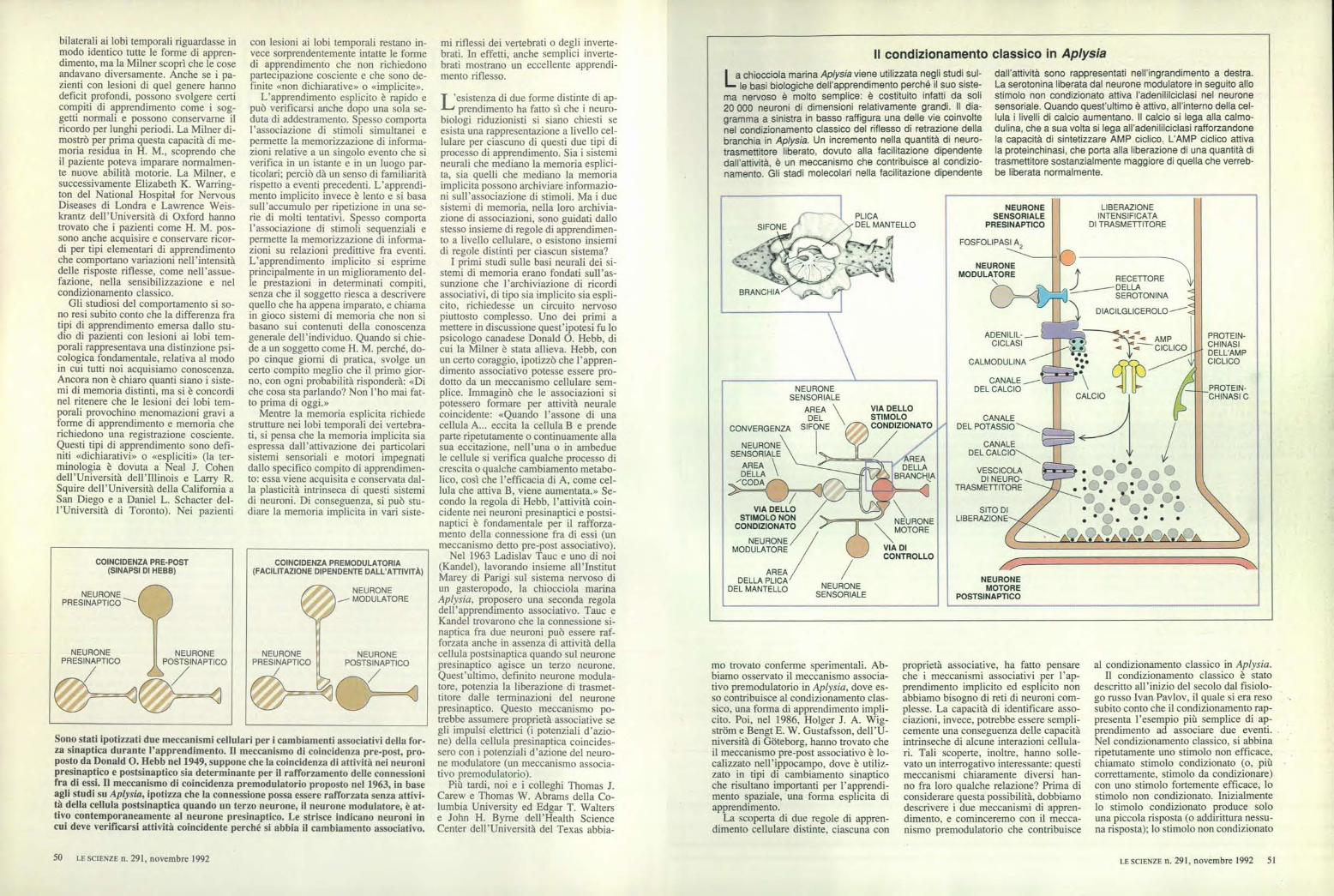
Il condizionamento classico in Aplysia
La chiocciola marina Aplysia viene utilizzata negli studi sul--le basi biologiche dell'apprendimento perché il suo siste-
ma nervoso è molto semplice: è costituito infatti da soli20 000 neuroni di dimensioni relativamente grandi. Il dia-gramma a sinistra in basso raffigura una delle vie coinvoltenel condizionamento classico del riflesso di retrazione dellabranchia in Aplysia. Un incremento nella quantità di neuro-trasmettitore liberato, dovuto alla facilitazione dipendentedall'attività, è un meccanismo che contribuisce al condizio-namento. Gli stadi molecolari nella facilitazione dipendente
dall'attività sono rappresentati nell'ingrandimento a destra.La serotonina liberata dal neurone modulatore in seguito allostimolo non condizionato attiva l'adenililciclasi nel neuronesensoriale. Quando quest'ultimo è attivo, all'interno della cel-lula i livelli di calcio aumentano. Il calcio si lega alla calmo-dulina, che a sua volta si lega all'adenililciclasi rafforzandonela capacità di sintetizzare AMP ciclico. L'AMP ciclico attivala proteinchinasi, che porta alla liberazione di una quantità ditrasmettitore sostanzialmente maggiore di quella che verreb-be liberata normalmente.
NEURONESENSORIALE
AREADEL
SIFONE
NEURONESENSORIALE
AREA \DELLA
-"CODA
VIA DELLOSTIMOLO NON
CONDIZIONATO
NEURONEMODULATORE
VIA DELLOSTIMOLOCONDIZIONATO/
READELLA
BRANCHIA
NEURONE\ MOTORE
VIA DICONTROLLO
CONVERGENZA
AREADELLA PLICA
)EL MANTELLO NEURONESENSORIALE
PLICADEL MANTELLOSIFONE
BRANCHIA
COINCIDENZA PRE-POST(SINAPSI DI HEBB)
NEURONEPRESINAPTICO
NEURONEPRESINAPTICO
NEURONEPOSTSINAPTICO
COINCIDENZA PREMODULATORIA(FACILITAZIONE DIPENDENTE DALL'ATTIVITÀ)
NEURONETPRESINAPTICO
OV-4/
NEURONEMODULATORE
NEURONEPOSTSINAPTICO
Sono stati ipotizzati due meccanismi cellulari per i cambiamenti associativi della for-za sinaptica durante l'apprendimento. Il meccanismo di coincidenza pre-post, pro-posto da Donald O. Hebb nel 1949, suppone che la coincidenza di attività nei neuronipresinaptico e postsinaptico sia determinante per il rafforzamento delle connessionifra di essi. Il meccanismo di coincidenza premodulatorio proposto nel 1963, in baseagli studi su Aplysia, ipotizza che la connessione possa essere rafforzata senza attivi-tà della cellula postsinaptica quando un terzo neurone, il neurone modulatore, è at-tivo contemporaneamente al neurone presinaptico. Le strisce indicano neuroni incui deve verificarsi attività coincidente perché si abbia il cambiamento associativo.
bilaterali ai lobi temporali riguardasse inmodo identico tutte le forme di appren-dimento, ma la Milner scoprì che le coseandavano diversamente. Anche se i pa-zienti con lesioni di quel genere hannodeficit profondi, possono svolgere certicompiti di apprendimento come i sog-getti normali e possono conservarne ilricordo per lunghi periodi. La Milner di-mostrò per prima questa capacità di me-moria residua in H. M., scoprendo cheil paziente poteva imparare normalmen-te nuove abilità motorie. La Milner, esuccessivamente Elizabeth K. Warring-ton del National Hospital for NervousDiseases di Londra e Lawrence Weis-krantz dell'Università di Oxford hannotrovato che i pazienti come H. M. pos-sono anche acquisire e conservare ricor-di per tipi elementari di apprendimentoche comportano variazioni nell'intensitàdelle risposte riflesse, come nell'assue-fazione, nella sensibilizzazione e nelcondizionamento classico.
Gli studiosi del comportamento si so-no resi subito conto che la differenza fratipi di apprendimento emersa dallo stu-dio di pazienti con lesioni ai lobi tem-porali rappresentava una distinzione psi-cologica fondamentale, relativa al modoin cui tutti noi acquisiamo conoscenza.Ancora non è chiaro quanti siano i siste-mi di memoria distinti, ma si è concordinel ritenere che le lesioni dei lobi tem-porali provochino menomazioni gravi aforme di apprendimento e memoria cherichiedono una registrazione cosciente.Questi tipi di apprendimento sono defi-niti «dichiarativi» o «espliciti» (la ter-minologia è dovuta a Neal J. Cohendell'Università dell'Illinois e Larry R.Squire dell'Università della California aSan Diego e a Daniel L. Schacter del-l'Università di Toronto). Nei pazienti
con lesioni ai lobi temporali restano in-vece sorprendentemente intatte le formedi apprendimento che non richiedonopartecipazione cosciente e che sono de-finite «non dichiarative» o «implicite».
L'apprendimento esplicito è rapido epuò verificarsi anche dopo una sola se-duta di addestramento. Spesso comportal'associazione di stimoli simultanei epermette la memorizzazione di informa-zioni relative a un singolo evento che siverifica in un istante e in un luogo par-ticolari; perciò dà un senso di familiaritàrispetto a eventi precedenti. L'apprendi-mento implicito invece è lento e si basasull'accumulo per ripetizione in una se-rie di molti tentativi. Spesso comportal'associazione di stimoli sequenziali epermette la memorizzazione di informa-zioni su relazioni predittive fra eventi.L'apprendimento implicito si esprimeprincipalmente in un miglioramento del-le prestazioni in determinati compiti,senza che il soggetto riesca a descriverequello che ha appena imparato, e chiamain gioco sistemi di memoria che non sibasano sui contenuti della conoscenzagenerale dell'individuo. Quando si chie-de a un soggetto come H. M. perché, do-po cinque giorni di pratica, svolge uncerto compito meglio che il primo gior-no, con ogni probabilità risponderà: «Diche cosa sta parlando? Non l'ho mai fat-to prima di oggi.»
Mentre la memoria esplicita richiedestrutture nei lobi temporali dei vertebra-ti, si pensa che la memoria implicita siaespressa dall'attivazione dei particolarisistemi sensoriali e motori impegnatidallo specifico compito di apprendimen-to: essa viene acquisita e conservata dal-la plasticità intrinseca di questi sistemidi neuroni. Di conseguenza, si può stu-diare la memoria implicita in vari siste-
mi riflessi dei vertebrati o degli inverte-brati. In effetti, anche semplici inverte-brati mostrano un eccellente apprendi-mento riflesso.
I 'esistenza di due forme distinte di ap-1-2 prendimento ha fatto sì che i neuro-biologi riduzionisti si siano chiesti seesista una rappresentazione a livello cel-lulare per ciascuno di questi due tipi diprocesso di apprendimento. Sia i sistemineurali che mediano la memoria esplici-ta, sia quelli che mediano la memoriaimplicita possono archiviare informazio-ni sull'associazione di stimoli. Ma i duesistemi di memoria, nella loro archivia-zione di associazioni, sono guidati dallostesso insieme di regole di apprendimen-to a livello cellulare, o esistono insiemidi regole distinti per ciascun sistema?
I primi studi sulle basi neurali dei si-stemi di memoria erano fondati sull'as-sunzione che l'archiviazione di ricordiassociativi, di tipo sia implicito sia espli-cito, richiedesse un circuito nervosopiuttosto complesso. Uno dei primi amettere in discussione quest'ipotesi fu lopsicologo canadese Donald O. Hebb, dicui la Milner è stata allieva. Hebb, conun certo coraggio, ipotizzò che l'appren-dimento associativo potesse essere pro-dotto da un meccanismo cellulare sem-plice. Immaginò che le associazioni sipotessero formare per attività neuralecoincidente: «Quando l'assone di unacellula A... eccita la cellula B e prendeparte ripetutamente o continuamente allasua eccitazione, nell'una o in ambeduele cellule si verifica qualche processo dicrescita o qualche cambiamento metabo-lico, così che l'efficacia di A, come cel-lula che attiva B, viene aumentata.» Se-condo la regola di Hebb, l'attività coin-cidente nei neuroni presinaptici e postsi-naptici è fondamentale per il rafforza-mento della connessione fra di essi (unmeccanismo detto pre-post associativo).
Nel 1963 Ladislav Tauc e uno di noi(Kandel), lavorando insieme all'InstitutMarey di Parigi sul sistema nervoso diun gasteropodo, la chiocciola marinaAplysia, proposero una seconda regoladell'apprendimento associativo. Tauc eKandel trovarono che la connessione si-naptica fra due neuroni può essere raf-forzata anche in assenza di attività dellacellula postsinaptica quando sul neuronepresinaptico agisce un terzo neurone.Quest'ultimo, definito neurone modula-tore, potenzia la liberazione di trasmet-titore dalle terminazioni del neuronepresinaptico. Questo meccanismo po-trebbe assumere proprietà associative segli impulsi elettrici (i potenziali d'azio-ne) della cellula presinaptica coincides-sero con i potenziali d'azione del neuro-ne modulatore (un meccanismo associa-tivo premodulatorio).
Più tardi, noi e i colleghi Thomas J.Carew e Thomas W. Abrams della Co-lumbia University ed Edgar T. Walterse John H. Byrne dell'Health ScienceCenter dell'Università del Texas abbia-
mo trovato conferme sperimentali. Ab-biamo osservato il meccanismo associa-tivo premodulatorio in Aplysia, dove es-so contribuisce al condizionamento clas-sico, una forma di apprendimento impli-cito. Poi, nel 1986, Holger J. A. Wig-strtim e Bengt E. W. Gustafsson, dell'U-niversità di Góteborg, hanno trovato cheil meccanismo pre-post associativo è lo-calizzato nell'ippocampo, dove è utiliz-zato in tipi di cambiamento sinapticoche risultano importanti per l'apprendi-mento spaziale, una forma esplicita diapprendimento.
La scoperta di due regole di appren-dimento cellulare distinte, ciascuna con
proprietà associative, ha fatto pensareche i meccanismi associativi per l'ap-prendimento implicito ed esplicito nonabbiamo bisogno di reti di neuroni com-plesse. La capacità di identificare asso-ciazioni, invece, potrebbe essere sempli-cemente una conseguenza delle capacitàintrinseche di alcune interazioni cellula-ri. Tali scoperte, inoltre, hanno solle-vato un interrogativo interessante: questimeccanismi chiaramente diversi han-no fra loro qualche relazione? Prima diconsiderare questa possibilità, dobbiamodescrivere i due meccanismi di appren-dimento, e cominceremo con il mecca-nismo premodulatorio che contribuisce
al condizionamento classico in Aplysia.Il condizionamento classico è stato
descritto all'inizio del secolo dal fisiolo-go russo Ivan Pavlov, il quale si era resosubito conto che il condizionamento rap-presenta l'esempio più semplice di ap-prendimento ad associare due eventi.Nel condizionamento classico, si abbinaripetutamente uno stimolo non efficace,chiamato stimolo condizionato (o, piùcorrettamente, stimolo da condizionare)con uno stimolo fortemente efficace, lostimolo non condizionato. Inizialmentelo stimolo condizionato produce solouna piccola risposta (o addirittura nessu-na risposta); lo stimolo non condizionato
50 LE SCIENZE n. 291, novembre 1992 LE SCIENZE n. 291, novembre 1992 51
CORTECCIAVISIVA
LOBO TEMPORALE
VIA COLLATERALEDI SCHAFFER
IPPOCAMPO
VIA DELLE FIBREPERFORANTI
VIA DELLE FIBREMUSCOIDI
REGIONE DENTATA
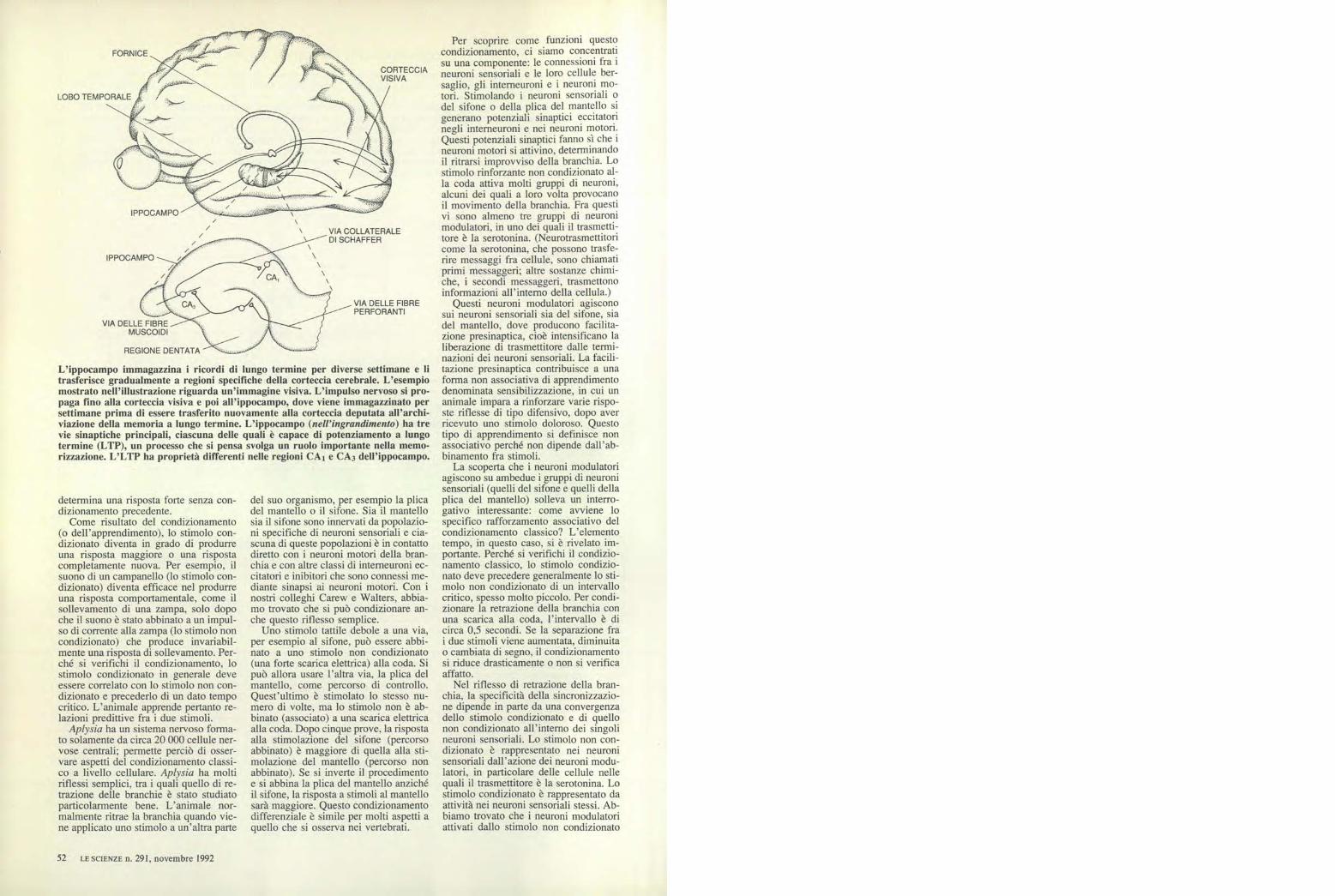
L'ippocampo immagazzina i ricordi di lungo termine per diverse settimane e litrasferisce gradualmente a regioni specifiche della corteccia cerebrale. L'esempiomostrato nell'illustrazione riguarda un'immagine visiva. L'impulso nervoso si pro-paga fino alla corteccia visiva e poi all'ippocampo, dove viene immagazzinato persettimane prima di essere trasferito nuovamente alla corteccia deputata all'archi-viazione della memoria a lungo termine. L'ippocampo (nell'ingrandimento) ha trevie sinaptiche principali, ciascuna delle quali è capace di potenziamento a lungotermine (LTP), un processo che si pensa svolga un ruolo importante nella memo-rizzazione. L'LTP ha proprietà differenti nelle regioni CA I e CA 3 dell'ippocampo.
determina una risposta forte senza con-dizionamento precedente.
Come risultato del condizionamento(o dell'apprendimento), lo stimolo con-dizionato diventa in grado di produrreuna risposta maggiore o una rispostacompletamente nuova. Per esempio, ilsuono di un campanello (lo stimolo con-dizionato) diventa efficace nel produrreuna risposta comportamentale, come ilsollevamento di una zampa, solo dopoche il suono è stato abbinato a un impul-so di corrente alla zampa (lo stimolo noncondizionato) che produce invariabil-mente una risposta di sollevamento. Per-ché si verifichi il condizionamento, lostimolo condizionato in generale deveessere correlato con lo stimolo non con-dizionato e precederlo di un dato tempocritico. L'animale apprende pertanto re-lazioni predittive fra i due stimoli.
Aplysia ha un sistema nervoso forma-to solamente da circa 20 000 cellule ner-vose centrali; permette perciò di osser-vare aspetti del condizionamento classi-co a livello cellulare. Aplysia ha moltiriflessi semplici, tra i quali quello di re-trazione delle branchie è stato studiatoparticolarmente bene. L'animale nor-malmente ritrae la branchia quando vie-ne applicato uno stimolo a un'altra parte
del suo organismo, per esempio la plicadel mantello o il sifone. Sia il mantellosia il sifone sono innervati da popolazio-ni specifiche di neuroni sensoriali e cia-scuna di queste popolazioni è in contattodiretto con i neuroni motori della bran-chia e con altre classi di intemeuroni ec-citatori e inibitori che sono connessi me-diante sinapsi ai neuroni motori. Con inostri colleghi Carew e Walters, abbia-mo trovato che si può condizionare an-che questo riflesso semplice.
Uno stimolo tattile debole a una via,per esempio al sifone, può essere abbi-nato a uno stimolo non condizionato(una forte scarica elettrica) alla coda. Sipuò allora usare l'altra via, la plica delmantello, come percorso di controllo.Quest'ultimo è stimolato lo stesso nu-mero di volte, ma lo stimolo non è ab-binato (associato) a una scarica elettricaalla coda. Dopo cinque prove, la rispostaalla stimolazione del sifone (percorsoabbinato) è maggiore di quella alla sti-molazione del mantello (percorso nonabbinato). Se si inverte il procedimentoe si abbina la plica del mantello anzichéil sifone, la risposta a stimoli al mantellosarà maggiore. Questo condizionamentodifferenziale è simile per molti aspetti aquello che si osserva nei vertebrati.
Per scoprire come funzioni questocondizionamento, ci siamo concentratisu una componente: le connessioni fra ineuroni sensoriali e le loro cellule ber-saglio, gli intemeuroni e i neuroni mo-tori. Stimolando i neuroni sensoriali odel sifone o della plica del mantello sigenerano potenziali sinaptici eccitatorinegli intemeuroni e nei neuroni motori.Questi potenziali sinaptici fanno sì che ineuroni motori si attivino, determinandoil ritrarsi improvviso della branchia. Lostimolo rinforzante non condizionato al-la coda attiva molti gruppi di neuroni,alcuni dei quali a loro volta provocanoil movimento della branchia. Fra questivi sono almeno tre gruppi di neuronimodulatori, in uno dei quali il trasmetti-tore è la serotonina. (Neurotrasmettitoricome la serotonina, che possono trasfe-rire messaggi fra cellule, sono chiamatiprimi messaggeri; altre sostanze chimi-che, i secondi messaggeri, trasmettonoinformazioni all'interno della cellula.)
Questi neuroni modulatori agisconosui neuroni sensoriali sia del sifone, siadel mantello, dove producono facilita-zione presinaptica, cioè intensificano laliberazione di trasmettitore dalle termi-nazioni dei neuroni sensoriali. La facili-tazione presinaptica contribuisce a unaforma non associativa di apprendimentodenominata sensibilizzazione, in cui unanimale impara a rinforzare varie rispo-ste riflesse di tipo difensivo, dopo averricevuto uno stimolo doloroso. Questotipo di apprendimento si definisce nonassociativo perché non dipende dall'ab-binamento fra stimoli.
La scoperta che i neuroni modulatoriagiscono su ambedue i gruppi di neuronisensoriali (quelli del sifone e quelli dellaplica del mantello) solleva un interro-gativo interessante: come avviene lospecifico rafforzamento associativo delcondizionamento classico? L'elementotempo, in questo caso, si è rivelato im-portante. Perché si verifichi il condizio-namento classico, lo stimolo condizio-nato deve precedere generalmente lo sti-molo non condizionato di un intervallocritico, spesso molto piccolo. Per condi-zionare la retrazione della branchia conuna scarica alla coda, l'intervallo è dicirca 0,5 secondi. Se la separazione frai due stimoli viene aumentata, diminuitao cambiata di segno, il condizionamentosi riduce drasticamente o non si verificaaffatto.
Nel riflesso di retrazione della bran-chia, la specificità della sincronizzazio-ne dipende in parte da una convergenzadello stimolo condizionato e di quellonon condizionato all'interno dei singolineuroni sensoriali. Lo stimolo non con-dizionato è rappresentato nei neuronisensoriali dall'azione dei neuroni modu-latori, in particolare delle cellule nellequali il trasmettitore è la serotonina. Lostimolo condizionato è rappresentato daattività nei neuroni sensoriali stessi. Ab-biamo trovato che i neuroni modulatoriattivati dallo stimolo non condizionato
52 LE SCIENZE n. 291, novembre 1992

LIBERAZIONEINTENSIFICATA
DI TRASMETTITORE
NEURONEPRESINAPTICO
CALCIO
VESCICOLADI NEURO.
TRASMETTITORE
SITODI LIBERA-
ZIONE • •A 4,,OACDA0A0A0A0A
GMP GUANILIL-• CICLICO CICLASI
ADP-RIBOSIL-TRANSFERASI
• • •• • • •• • •• • •
•• „CALCIO
• • •
GLUTAMMATO
CANALENMDA
NEURONEPOSTSINAPTICO CALMODULINA
vCALCIO-
CALMODULINA VCHINASI PROTEIN-
CHINASI C
TIROSIN-CHINASI
vOSSIDO D'AZOTO
SINTETASI
CANALENON-NMDA
alla coda producono una maggiore faci-litazione presinaptica dei neuroni senso-riali se questi hanno appena attivato po-tenziali d'azione in risposta allo stimolocondizionato. Nei neuroni sensoriali ipotenziali d'azione che si verificano su-bito dopo la scarica alla coda non hannoalcun effetto.
Questa particolare proprietà della fa-cilitazione presinaptica viene definita di-pendenza dall'attività. La facilitazionedipendente dall'attività richiede la stessasincronizzazione a livello cellulare che èrichiesta nel condizionamento a livellocomportamentale e può spiegare talecondizionamento. Questi risultati fannopensare che un meccanismo cellulare delcondizionamento classico del riflesso diretrazione sia un'elaborazione della fa-cilitazione presinaptica, meccanismo u-tilizzato per la sensibilizzazione del ri-flesso. Questi esperimenti hanno indica-
CALCIO •• MAGNESIO
•
to che potrebbe esistere un alfabeto cel-lulare per l'apprendimento, in cui i mec-canismi dei tipi più complessi di appren-dimento possono essere elaborazioni ocombinazioni dei meccanismi di tipi diapprendimento più semplici.
Il tassello successivo nel puzzle delcondizionamento classico è andato alsuo posto quando si è scoperto perchél'attivazione dei potenziali d'azione neineuroni sensoriali subito prima dello sti-molo non condizionato alla coda facciaaumentare la facilitazione presinaptica.In precedenza avevamo scoperto chequando i neuroni modulatori liberano se-rotonina in risposta a una scarica elettri-ca alla coda, nei neuroni sensoriali iniziauna serie di trasformazioni biochimiche(si veda la finestra a pagina 51). La se-rotonina si lega a un recettore che attival'adenililciclasi. Questo enzima a suavolta trasforma l'ATP, una delle mole-
MESSAGGERORETROGRADO
(OSSIDO D'AZOTO)
cole che forniscono l'energia necessariaad alimentare varie attività della cellula,in AMP ciclico. L'AMP ciclico agiscepoi nella cellula come secondo messag-gero (il primo messaggero è la serotoni-na) e attiva un altro enzima, una protein-chinasi. Le chinasi sono proteine cheprovocano la fosforilazione (l'aggiuntadi un gruppo fosfato) di altre proteine,aumentando così l'attività di alcune e fa-cendo diminuire quella di altre.
L'attivazione della proteinchinasi neineuroni sensoriali ha diverse conseguen-ze importanti sul breve periodo. La pro-teinchinasi fosforila le proteine dei ca-nali del potassio. La fosforilazione diquesti canali (o delle proteine che agi-scono su di essi) riduce una componentedella corrente del potassio che normal-mente ripolarizza il potenziale d'azione.La riduzione della corrente del potassioprolunga il potenziale d'azione e con-sente così l'attivazione più prolungatadei canali del calcio, e di conseguenzal'ingresso nella terminazione presinapti-ca di una maggiore quantità di calcio.All'interno della cellula il calcio ha mol-te funzioni, una delle quali è la libera-zione di vescicole di trasmettitore dallaterminazione. Quando nella terminazio-ne entra più calcio a seguito di un au-mento della durata dei potenziali d'azio-ne, si libera una maggior quantità di tra-smettitore. In secondo luogo, a causadell'attività della proteinchinasi, la sero-tonina fa sì che le vescicole di trasmet-titore si spostino dal sito di deposito aisiti di liberazione sulla membrana; que-sto facilita la liberazione di trasmettitoreindipendentemente da un aumento dellaquantità di calcio in ingresso. In questaattività, l'AMP ciclico agisce in paralle-lo con un altro secondo messaggero, laproteinchinasi C, anche questa attivatadalla serotonina.
Per quale motivo l'attivazione dei po-tenziali d'azione nei neuroni sensorialisubito prima dello stimolo non condizio-nato rafforza l'azione della serotonina?I potenziali d'azione determinano molticambiamenti nei neuroni sensoriali: per-mettono l'ingresso di sodio e calcio el'uscita di potassio, e modificano il po-tenziale di membrana. Abrams e Kandelhanno scoperto che la funzione criticadei potenziali d'azione per la dipenden-za dall'attività è il passaggio del calcionei neuroni sensoriali. Una volta che sitrova nella cellula, il calcio si lega allacalmodulina, una proteina che amplifical'attivazione dell'enzima adenililciclasida parte della serotonina. Quando lacoppia calcio-calmodulina si lega all'a-denililciclasi, l'enzima genera ulterioreAMP ciclico. Grazie a questa capacitàl'adenililciclasi diventa un sito di con-vergenza importante per gli stimoli con-dizionati e non condizionati.
I due tipi di stimolo sono quindi rap-presentati all'interno della cellula dallaconvergenza di due segnali diversi (cal-cio e serotonina) sullo stesso enzima.L'intervallo di 0,5 secondi fra stimolo
OSSIDO D'AZOTO
Nell'LTP la membrana postsinaptica è depolarizzata dall'azione dei canali nonNMDA. La depolarizzazione annulla il blocco esercitato dal magnesio sul canale as-sociato ai recettori NMDA, consentendo l'ingresso del calcio; questo attiva le chinasicalcio-dipendenti che portano all'induzione di LTP. Si pensa che il neurone postsi-naptico liberi un messaggero retrogrado che penetra la membrana del neurone pre-sinaptico. Questo messaggero, che si ritiene sia l'ossido di azoto, agirebbe nella ter-minazione presinaptica in modo da intensificare la liberazione di trasmettitore (glu-tammato), forse per attivazione della guanililciclasi o dell'ADP-ribosiltransferasi.
54 LE sciENzE n. 291, novembre 1992

NEURONESENSORIALE
NEURONEMOTORE
)
FACILITAZIONE DIPENDENTE DALL'ATTIVITÀ IN APLYSIA
NEURONEMODULATORE
POTENZIAMENTO A LUNGO TERMINENELL'IPPOCAMPO
NEURONE PRESINAPTICO NEURONE POSTSINAPTICO
I processi associativi che si pensa contribuiscano all'apprendimento in Aplysia e nel-l'ippocampo dei mammiferi potrebbero avere meccanismi simili e coinvolgere unasostanza modulatrice che produce un'intensificazione dipendente dall'attività dellaliberazione di trasmettitore dal neurone presinaptico. I neuroni a strisce sono quelliin cui deve aversi attività coincidente perché si produca il cambiamento associativo.
condizionato e non condizionato, essen-ziale per l'apprendimento nel riflesso diretrazione della branchia, può corrispon-dere all'intervallo di tempo nel corso delquale il calcio entra nella terminazionepresinaptica e va a legarsi alla calmodu-lina così da indurre l'adenililciclasi aprodurre una maggiore quantità di AMPciclico in risposta alla serotonina.
L'amplificazione dipendente dall'atti-vità della via dell'AMP ciclico non è fe-nomeno esclusivo dei riflessi di retrazio-ne della branchia o della coda di Aplysia.Studi genetici su Drosophila, il mosce-rino della frutta, hanno chiamato in cau-sa per il condizionamento un meccani-smo molecolare simile. Drosophila puòessere condizionata, e sono stati identi-ficati mutanti per un solo gene che han-no difficoltà di apprendimento. Uno diquesti mutanti, chiamato rutabaga, èstato studiato da William G. Quinn delMassachusetts Institute of Technology eMargaret Livingstone della Harvard U-niversity e da Yadin Dudai del Weiz-mann Institute in Israele. Oggi sappiamoche la proteina difettosa in questo mu-tante è una adenililciclasi dipendentedalla coppia calcio-calmodulina. Comeconseguenza della mutazione, la ciclasiha perso in rutabaga la sua capacità diessere stimolata dalla coppia calcio-cal-modulina. Inoltre, Ronald L. Davis ecolleghi del Cold Spring Harbor Labo-ratory hanno scoperto che questa forma
di adenililciclasi è abbondante nei corpifungiformi (o peduncolati), una parte delcervello del moscerino che è determi-nante per molti tipi di apprendimento as-sociativo. Sia gli studi di biologia cellu-lare su Aplysia, sia gli studi di geneticasu Drosophila evidenziano dunque l'im-portanza del sistema del secondo mes-saggero dell'AMP ciclico in alcuni tipielementari di apprendimento implicito edi memorizzazione.
Che cosa si può dire delle forme espli-cite di apprendimento? Anche que-
sti tipi più complessi di apprendimentoassociativo hanno rappresentazioni cel-lulari per l'associatività? Se la risposta èaffermativa, questi meccanismi debbonoessere diversi da quelli dell'apprendi-mento implicito perché, a differenza delcondizionamento classico, l'apprendi-mento esplicito ha sovente più successoquando i due eventi associati si verifica-no simultaneamente. Per esempio, rico-nosciamo più facilmente il volto di unapersona che ci è nota quando la vediamoin uno specifico contesto. Gli stimoli delvolto e dell'ambiente agiscono simulta-neamente e contribuiscono a farci rico-noscere quella persona.
Nell'uomo, come abbiamo visto, l'ap-prendimento esplicito richiede il lobotemporale. Tuttavia inizialmente non erachiaro quanto dovesse essere estesa lalesione bilaterale nel lobo temporale per
interferire con la memorizzazione. Studisuccessivi su esseri umani e su animali,condotti da Mortimer Mishkin dei Na-tional Institutes of Health e da Squire,David G. Amaral e Stuart Zola-Morgandell'Università della California a SanDiego ci aiutano a dare una risposta.Questi studi, infatti, fanno pensare cheuna struttura del lobo temporale partico-larmente importante per l'archiviazionedei ricordi sia l'ippocampo. Tuttaviale lesioni dell'ippocampo interferisconosolo con l'archiviazione di nuovi ricordi:pazienti come H. M. conservano unamemoria abbastanza buona di eventiprecedenti alla lesione. Sembra che l'ip-pocampo sia solo un deposito tempora-neo per la memoria a lungo termine.L'ippocampo elabora le informazioniapprese, per un periodo di qualche setti-mana o di qualche mese, poi trasferiscequelle informazioni alle aree pertinentidella corteccia cerebrale, dove troveran-no una collocazione più permanente (siveda l'articolo Cervello e linguaggio diAntonio R. Damasio e Hanna Damasioa pagina 64). Secondo l'analisi di Patri-cia S. Goldman-Rakic, i ricordi imma-gazzinati in questi diversi siti corticalisono poi espressi attraverso la memoriadi lavoro della corteccia prefrontale (siveda l'articolo La memoria di lavoro apagina 76).
Nel 1973 Timothy Bliss e TerjeLomo, allora nel laboratorio di Per An-dersen a Oslo, in Norvegia, hanno perprimi dimostrato che i neuroni dell'ip-pocampo hanno proprietà plastiche no-tevoli, del tipo che sarebbe necessarioper l'apprendimento. Hanno trovato cheun breve treno ad alta frequenza di po-tenziali d'azione in una delle vie nervosedell'ippocampo produce un aumentodella forza delle sinapsi in quella via. Sipuò vedere che l'aumento dura per orein un animale anestetizzato e per giornie addirittura settimane in un animale vi-gile e libero di muoversi.
Bliss e LOrno hanno dato a questo raf-forzamento il nome di potenziamento alungo termine (LTP). Studi successivihanno dimostrato che l'LTP ha proprietàdiverse in tipi diversi di sinapsi all'inter-no dell'ippocampo. Qui ci concentrere-mo sul tipo associativo di potenziamen-to, che ha due caratteristiche in relazionefra loro. Innanzitutto, l'associatività èdella forma pre-post di Hebb: perché siverifichi la facilitazione, i neuroni presi-naptico e postsinaptico in causa debbonoessere attivi simultaneamente. In secon-do luogo (e di conseguenza) il potenzia-mento a lungo termine è specifico: la suaazione è ristretta alla via che vienestimolata.
Perché per il potenziamento a lungotermine è necessaria l'attivazione simul-tanea dei neuroni presinaptico e postsi-naptico? Le principali vie nervose del-l'ippocampo utilizzano come trasmetti-tore un amminoacido, il glutammato.Esso genera un potenziamento a lungotermine legandosi ai recettori del glu-
56 LE SCIENZE n. 291, novembre 1992

La rappresentazione della superficie del corpo nella corteccia
Lhomunculus è un metodo tradizio-nale per illustrare come la superfi-
cie del corpo sia rappresentata nellacorteccia somatosensoriale. Aree piùgrandi della corteccia sono dedicate aparti del corpo che hanno una maggio-re sensibilità, come le dita e le labbra.
Gli effetti dell'addestramento dellasensibilità sono stati recentemente di-mostrati nell'aoto (una scimmia nottur-na del Sud America settentrionale, del-la famiglia dei cebidi). Le dita dell'aotosono rappresentate nelle aree 3b e 1
della corteccia somatosensoriale (a). I diagrammi b e d delineano le regioni cheriproducono la superficie delle dita di una scimmia adulta (c) prima e dopo l'ad-destramento. Nel corso dell'addestramento la scimmia faceva ruotare un discoper un'ora al giorno, usando solo il secondo, il terzo e talvolta il quarto dito. Dopotre mesi di questa attività, l'area che rappresenta nel cervello le dita stimolate siè ingrandita sostanzialmente.
DOPOLA STIMOLAZIONE
DIFFERENZIALE
PRIMADELLA STIMOLAZIONE
DIFFERENZIALE
aAREA 3b AREA 1
tammato sulle cellule bersaglio. Risultache esistono due tipi importanti di recet-tori del glutammato: i recettori NMDA(che prendono il nome dallo N-metil-D--aspartato, che si lega a sua volta a que-sti recettori) e i recettori non NMDA.Questi ultimi sono prevalenti nella mag-gior parte delle trasmissioni sinaptiche,perché il canale ionico associato ai re-cettori NMDA è di solito bloccato dalmagnesio e si sblocca solo quando ilneurone postsinaptico è depolarizzato.Inoltre l'attivazione ottimale del canaleassociato al recettore NMDA richiedeche i due segnali (il legame del glutam-mato al recettore e la depolarizzazionedel neurone postsinaptico) si verifichinosimultaneamente. Il recettore NMDA haquindi proprietà associative, ovvero diidentificazione della coincidenza, pro-prio come l 'adenililciclasi. Le sue carat-teristiche temporali, indispensabili perl'attivazione simultanea, sono però piùadatte alle forme esplicite di apprendi-mento, anziché a quelle implicite.
L'ingresso del calcio nel neuronepostsinaptico attraverso il canale asso-ciato al recettore NMDA non bloccato èdeterminante per il potenziamento a lun-go termine, come hanno dimostratoGary Lynch dell'Università della Cali-
fomia a Irvine e Roger A. Nicoll, RobertS. Zucker e colleghi dell'Università del-la California a San Francisco. Il calciodà inizio all'LTP attivando almeno tretipi diversi di proteinchinasi.
L'induzione dell'LTP sembra dipen-dere dalla depolarizzazione postsinapti-ca, che porta all'ingresso del calcio e al-la successiva attivazione delle protein-chinasi che fungono da secondi messag-geri. Per il mantenimento dell'LTP, in-vece, vari gruppi di ricercatori (fra glialtri Bliss e colleghi, John Bekkers eCharles Stevens del Salk Institute e Ro-berto Malinow e Richard Tsien dellaStanford University) hanno scoperto cheè in gioco l'intensificazione del trasmet-titore della terminazione presinaptica.
Se l'induzione dell'LTP richiede unevento postsinaptico (ingresso di calcioattraverso i canali associati ai recettoriNMDA) e il suo mantenimento compor-ta un evento presinaptico (liberazione dimaggiori quantità di trasmettitore) allo-ra, come ha proposto per primo Bliss, ilneurone postsinaptico deve inviare qual-che tipo di messaggio al neurone presi-naptico. Ma allora gli esperti di neuro-scienze si trovano di fronte a un grossoproblema: sin da quando il grande ana-tomista spagnolo Santiago Ramón y
Cajal ha enunciato il principio della po-larizzazione dinamica, tutte le sinapsichimiche studiate si sono dimostrateunidirezionali. L'informazione fluiscesolo dal neurone presinaptico a quellopostsinaptico. Nel potenziamento a lun-go termine sembra che emerga un nuovoprincipio di comunicazione fra le cellulenervose. Le vie dei secondi messaggeriattivati dal calcio, o forse la stessa atti-vità diretta del calcio, sembrano provo-care la liberazione di un fattore di pla-sticità retrograda dal neurone postsi-naptico attivo. Questo fattore retrogradodiffonde quindi nelle terminazioni presi-naptiche e attiva uno o più secondi mes-saggeri che intensificano la liberazio-ne di trasmettitore e in tal modo man-tengono l'LTP (si veda l'illustrazione apagina 54).
A differenza delle terminazioni presi-naptiche, che immagazzinano trasmetti-tore in vescicole e lo liberano in siti spe-cializzati, le terminazioni postsinaptichenon hanno dispositivi speciali di libera-zione. Sembrava dunque attraente sup-porre che il messaggero retrogrado po-tesse essere una sostanza in grado di dif-fondere rapidamente dal neurone postsi-naptico, oltre la fessura sinaptica e finoalla terminazione presinaptica. Nel 1991quattro gruppi di ricercatori hanno otte-nuto dati sperimentali a sostegno dell'i-dea che questo messaggero retrogradosia l'ossido d'azoto: Thomas J. O'Dell eOttavio Arancio del nostro laboratorio,Erin M. Schuman e Daniel Madison del-la Stanford University, Paul F. Chapmane colleghi della School of Medicine del-l'Università del Minnesota e GeorgBóhme e colleghi in Francia. Se si ini-bisce la sintesi di ossido di azoto nelneurone postsinaptico o si assorbe ossi-do d'azoto nello spazio extracellularel'induzione di LTP viene bloccata, men-tre l'aggiunta di ossido d'azoto potenziala liberazione di trasmettitore dai neuro-ni presinaptici.
Mentre studiavamo gli effetti dell'e-sposizione di sezioni di ippocampo al-l'ossido d'azoto, insieme con Scott A.Small e Min Zhuo abbiamo fatto unascoperta sorprendente: l'ossido di azotoproduce LTP solo se è associato ad atti-vità dei neuroni presinaptici, come acca-de per la facilitazione presinaptica di-pendente dall'attività in Aplysia. Sembrache l'attività presinaptica, e forse l'in-gresso di calcio, siano determinanti per-ché l'ossido di azoto produca potenzia-mento. Questi esperimenti fanno pensa-re che il potenziamento a lungo termineutilizzi una combinazione di due mecca-nismi indipendenti di apprendimentosinaptico associativo: un meccanismohebbiano a recettore NMDA e uno nonhebbiano di facilitazione presinaptica di-pendente dall'attività. Secondo quest'i-potesi, l'attivazione di recettori NMDAnei neuroni postsinaptici produce un se-gnale retrogrado (ossido di azoto). Il se-gnale poi dà il via a un meccanismo pre-sinaptico dipendente dall'attività che fa-
cilita la liberazione di trasmettitore dalleterminazioni presinaptiche.
Quale può essere il vantaggio funzio-nale di combinare in questo modo duemeccanismi cellulari associativi, il recet-tore NMDA postsinaptico e la facilita-zione presinaptica dipendente dall'atti-vità? Se la facilitazione presinaptica èprodotta da una sostanza che può diffon-dersi, questa sostanza, in teoria, potreb-be raggiungere vie adiacenti. In effetti,studi effettuati da Tobias Bonhoeffere colleghi del Max-Planck-Institut fiirHimforschung di Francoforte indicanoche I'LTP iniziato in un neurone postsi-naptico si comunica a neuroni postsi-naptici adiacenti. Il fatto che la facilita-zione presinaptica sia dipendente dal-l'attività potrebbe essere un metodo pergarantire che vengano potenziate solospecifiche vie presinaptiche, cioè quelleche sono attive. Qualsiasi terminazionepresinaptica non attiva non ne sarebbeinfluenzata (si veda l'illustrazione apagina 56).
I cambiamenti nelle sinapsi che sipensa contribuiscano a questi casi di ap-prendimento implicito ed esplicito solle-vano una sorprendente possibilità ridu-zionista. Il fatto che i cambiamenti si-naptici associativi non richiedano reti dineuroni complesse fa pensare che possaesistere una corrispondenza diretta fraqueste forme associative di apprendi-mento e proprietà cellulari fondamentali.Nei casi che abbiamo passato in rasse-gna, le proprietà cellulari sembra derivi-no a loro volta dalle proprietà di proteinespecifiche - l'adenililciclasi e il recettoreNMDA - che sono in grado di risponde-re a due segnali indipendenti, comequelli che derivano dallo stimolo condi-zionato e dallo stimolo non condiziona-to. Ovviamente questi meccanismi asso-ciativi molecolari non agiscono isolata-mente: sono incorporati in cellule chepossiedono raffinati meccanismi mole-colari per elaborare il processo associa-tivo. Le cellule, a loro volta, sono inse-rite in reti complesse caratterizzate dauna notevole dose di ridondanza e di pa-rallelismo e da una enorme potenza dicalcolo, tutti elementi che aggiungonocomplessità sostanziale a questi mecca-nismi elementari.
Quando si è scoperto che il potenzia-mento a lungo termine si verifica nel-l'ippocampo, una regione che, come giàsi sapeva, è significativa per la memoria,si è cominciato a porre il problema sel'LTP fosse coinvolto nel processo di ar-chiviazione dei ricordi in quest'area delcervello. Dati sperimentali positivi inquesto senso sono stati forniti da Ri-chard Morris e colleghi della MedicalSchool dell'Università di Edimburgoche hanno utilizzato un compito di me-morizzazione spaziale. Quando i recet-tori NMDA dell'ippocampo sono bloc-cati, gli animali che eseguono l'esperi-mento non riescono ad apprendere ilcompito. Questi esperimenti indicanoche i meccanismi del recettore NMDA
nell'ippocampo, e forse FLTP, sono co-involti nell'apprendimento spaziale.
A vendo preso in considerazione i mec-canismi mediante i quali l'appren-
dimento può produrre cambiamenti nellecellule nervose, dobbiamo affrontare unultimo gruppo di interrogativi. Quali so-no i meccanismi grazie ai quali si man-tengono i cambiamenti sinaptici prodottidall'apprendimento esplicito e implici-to? Come si conserva la memoria nellungo termine?
Esperimenti condotti sia con Aplysiasia con mammiferi indicano che la me-morizzazione esplicita e quella implicitaprocedono per stadi. L'immagazzina-mento dell'informazione iniziale, un ti-po di memoria a breve termine, dura daqualche minuto a qualche ora e compor-ta cambiamenti nella forza di connessio-ni sinaptiche esistenti (attraverso modi-ficazioni mediate dai secondi messagge-ri, del tipo che abbiamo visto). I cam-biamenti di lungo termine (quelli chedurano settimane e mesi) sono imma-gazzinati nello stesso sito, ma richiedo-no qualcosa di completamente nuovo:l'attivazione di geni, l'espressione dinuove proteine e la crescita di nuoveconnessioni. In Aplysia, Craig H. Bailey,Mary C. Chen e Samuel M. Schacher ecolleghi della Columbia University eByrne e colleghi dello Health ScienceCenter dell'Università del Texas hannotrovato che gli stimoli che produconomemoria a lungo termine per sensibiliz-zazione e per condizionamento classicoconducono a un aumento del numerodelle terminazioni presinaptiche. Nel-l'ippocampo si verificano cambiamentianatomici simili dopo l'LTP.
Se la memoria a lungo termine provo-ca cambiamenti anatomici, questo signi-fica forse che il nostro cervello cambiacostantemente dal punto di vista anato-mico quando apprendiamo e quandoscordiamo qualcosa? Ci saranno deicambiamenti nell'anatomia del nostrocervello se ci ricorderemo qualcosa diquel che è scritto in queste pagine, do-po aver letto questo fascicolo di «LeScienze»?
Molti scienziati si sono posti questadomanda, ma più di tutti se ne è occu-pato Michael Merzenich dell'Universitàdella California a San Francisco. Merze-nich ha esaminato la rappresentazionedella mano nell'area sensoriale dellacorteccia cerebrale. Fino a poco tempoaddietro, gli studiosi di neuroscienzepensavano che questa rappresentazionerimanesse costante per tutto il corso del-la vita, ma Merzenich e colleghi hannodimostrato che le mappe corticali sonocostantemente soggette a modifiche inbase all'uso delle vie sensoriali. Poichétutti noi cresciamo in ambienti diversi,siamo esposti a combinazioni diverse distimoli, e probabilmente esercitiamo lenostre capacità sensoriali e motorie inmodi diversi, l'architettura del cervellodi ciascuno di noi cambierà in modi leg-
germente diversi. Questa modificazionecaratteristica dell'architettura del cervel-lo, unita alla costituzione genetica, con-tribuisce alle basi biologiche dell'e-spressione dell'individualità.
Questa concezione è dimostrata nelmigliore dei modi in uno studio nel cor-so del quale Merzenich induceva unascimmia a toccare un disco in rotazionecon le sole tre dita centrali della mano.Dopo varie migliaia di rotazioni del di-sco, l'area della corteccia dedicata alletre dita centrali si era espansa, a spesedi quella dedicata alle altre dita. L'eser-cizio pratico, quindi, può portare a cam-biamenti nella rappresentazione conica-le delle dita più attive. Quali sono i mec-canismi sottostanti al fenomeno? Datirecenti indicano che le connessioni cor-ticali nel sistema somatosensoriale sonocostantemente modificate e aggiornatesulla base dell'attività correlata, attra-verso un meccanismo che appare similea quello che genera il potenziamento alungo termine.
In effetti, come spiega l'articolo diCarla J. Shatz (si veda Lo sviluppo delcervello a pagina 26), i primi risultati de-gli studi di biologia cellulare sullo svi-luppo fanno pensare che i meccanismidell'apprendimento possano portare unulteriore vantaggio. Vi è motivo per pen-sare che la regolazione fine delle con-nessioni nelle fasi avanzate dello svilup-po possa aver bisogno di un meccanismosinaptico associativo, dipendente dall'at-tività, forse simile all'LTP. Se questo èvero anche a livello molecolare, se cioèl'apprendimento ha meccanismi mole-colari in comune con aspetti dello svi-luppo e della crescita, lo studio dell'ap-prendimento può aiutarci a collegare lapsicologia cognitiva alla biologia mole-colare dell'organismo in generale. Que-sta grande unificazione biologica dissol-verebbe l'alone di mistero che circondai processi mentali e ne inserirebbe deci-samente lo studio nel quadro evoluzio-nistico della biologia.
BIBLIOGRAFIA
HAWKINS R. D., ABRAMS T. W., CA-REW T. J. e KANDEL E. R., A CellularMechanism of Classica! Conditioningin Aplysia: Activity-Dependent Amplifi-cation of Presynaptic Facilitation in«Science», 219, pp. 400-405, 28 gennaio1983.
NICOLL R. A., KAUER J. A. e MALENKA R.C., The Current Excitement in Long--Term Potentiation in «Neuron», 1, n. 2,aprile 1988.
SQUIRE LARRY R., Memory and theH ippocampus: A Synthesis from Find-ings with Rats, Monkeys, and Humans in«Psychological Review», 99, n. 2, aprile1992.
58 LE SCIENZE n. 291, novembre 1992
LE SCIENZE n. 291, novembre 1992 59


















