
VOLUME 13 NO. 2 JULI 2016 - IPB University
35
VOLUME 13 NO. 2 JULI 2016
Transcript of VOLUME 13 NO. 2 JULI 2016 - IPB University

VOLUME 13 NO. 2 JULI 2016

JURNAL PRIMATOLOGI INDONESIA(The Indonesian Journal of Primatology)
ISSN 1410-5373
Volume 13, Nomor 2, Juli 2016
PENANGGUNG JAWABHuda S. Darusman
(Kepala Pusat Studi Satwa Primata LPPM-IPB)Chairul Saleh
(Perhimpunan Ahli dan Pemerhati Primata Indonesia)
KETUA EDITORSri Supraptini Mansjoer
ANGGOTA EDITORIrma H. Suparto, Dyah Perwitasari,
Entang Iskandar, Audrey Maria Ungerer
EDITOR TEKNIKWalberto Sinaga
Vallen Sakti Maulana
SEKRETARIAT DAN SIRKULASIRahayu Sulistina, R.M. Maulana,
Tita Ratnasari
ALAMAT REDAKSIPusat Studi Satwa Primata LPPM-IPBJalan Lodaya II No. 5, Bogor 16151
Telepon (0251) 8324358, 8313637, 8320417Faks. (0251) 8360712
Surat Elektronik: [email protected]: http://journal.ipb.ac.id/index.php/primata
PENERBITPusat Studi Satwa Primata LPPM-IPB
(PSSP LPPM-IPB)bekerja sama dengan
Perhimpunan Ahli dan Pemerhati Primata Indonesia
(PERHAPI)
Editorial
Hewan model merupakan salah satu bahan penelitian yang sangat dibutuhkan untuk pengujian respon biologis secara in vivo. Untuk penelitian-penelitian yang akan digunakan dalam pemecahan masalah kesehatan manusia, hewan model yang karakteristiknya mirip manusia adalah satwa primata. Namun, tidak semua dapat digunakan. Syarat hewan model yang dapat digunakan adalah (1) tidak dilindungi karena jumlahnya cukup banyak, (2) anatomi dan fisiologi mirip manusia, (3) mudah didapat, (4) mudah ditangani, (5) tidak berbahaya, dan (6) berespon terhadap perlakuan yang diberikan, (7) bukan yang berasal dari alam, namun berasal dari penangkaran. Salah satu satwa primata yang populer digunakan adalah monyet ekor panjang (MEP/Macaca fascicularis).
Jurnal Primatologi Indonesia (JPI) merupakan jurnal ilmiah primatologi yang diterbitkan oleh Pusat Studi Satwa Primata, Lembaga Penelitian dan Pengabdian kepada Masyarakat Institut Pertanian Bogor (PSSP LPPM-IPB) bekerjasama dengan Perhimpunan Ahli dan Pemerhati Primata Indonesia (PAPPI). Tujuan dan ruang lingkup penelaahan: 1) satwa primata sebagai model dalam pencegahan dan penyembuhan penyakit manusia, 2) patologi, imunologi, parasitologi, mikrobiologi dan kedokteran hewan primata, 3) morfologi, fisiologi, reproduksi, taksonomi, pertumbuhan dan perkembangan, evolusi dan sistematika serta genetika satwa primata, 4) penangkaran, penanganan, metodologi eksperimen serta manajemen koloni dan laboratorium satwa primata, 5) ekologi, demografi, pelestarian dan manajemen kawasan konservasi satwa primata, 6) neurologi, tingkah laku, sosiologi, komunikasi, psikologi dan kesejahteraan satwa primata, dan 7) kebijakan pemanfaatan, pelestarian dan pengembangan satwa primata.
Langganan per-tahun dapat dilakukan dengan penggantian biaya cetak Rp 60.000,-. Informasi lebih lanjut dapat diperoleh melalui Bagian Sekretariat dan Sirkulasi JPI.
Foto profil kedih diambil di Bukit Lawang, Sumatera Utara oleh Richard Gonzalez
dan diambil di Taman Margasatwa Ragunan, Jakarta oleh Walberto SinagaPusat Studi Satwa Primata LPPM-IPBJalan Lodaya II No. 5, Bogor 16151.E-mail: [email protected]

Indonesian Primate ProfilePresbytis thomasiCommon names: Kedih (Indonesia); Thomas’s Langur, North Sumatran Leaf Monkey, Thomas’s Langur, Sumatran Grizzled Langur, Thomas’s Leaf Monkey (British); Semnopithèque De Thomas (French); Langur De Thomas (Spanish)
Presbytis thomasi is one of the several langurs found in the oriental Islands; however, its geographic range is fairly limited. They are native to North Sumatera, Indonesia; more specifically, north of Wampu River and Simpang Kiri River (Colijn and Muchtar 1996, Nowak 1999).
When searching through tropical rainforests, rubber plantations, and both primary and secondary forests, one is able to encounter Thomas’s langurs. Since they are arboreal, we can find this unique species high in trees. However, they position in trees depends on the time of day. During the day, they select trees that have lots of twigs and leaves to protect them from the harsh sunlight. At night, they sleep in the top of tall trees in open areas (Colijn and Muchtar 1996, Flannery 2004, Gurmaya 1986).
This species have very distinctive appearance. Due to their unique facial coloration, it is easy to distinguish the North Sumatran leaf monkeys from other primates.
The white fur on the underside and arms (which contrasts the black fur surrounding the rest of the body) continues up around the neck. Two white stripes, starting from the top of the head, running down the sides, coming together in a V-shape and encircling the eyes. A purple-silver inner layer forms rings around their orange-brown eyes. Under their dark skin is a
pinkish skin which also covers their muzzle.The average weight of a Thomas's langur
is 6,69 kg for adult females, and 6,67 kg for adult males. The tail length is 500 to 850 mm, and body length is 420 to 620 mm (Colijn and Muchtar 1996, Flannery 2004, Nowak 1999).
The mating system of P. thomasi is still being disputed. In the Encyclopedia of Mammals, it is noted that the species is a monogamous primate. The female initiates the mating by performing various acts to persuade her male counterpart, such as releasing certain smells and displaying genitalia (Colobus and Leaf Monkeys 2001).
However, others dispute the monogamy of the species. Steenbeck et al. (1999) stated that within groups, there are often several females and one breeding male. A possible resolution between the two observations is that only one of the females in the group breeds with the male while the other females help raise the young (Steenbeek et al. 1999).
There is no specified breeding interval for Thomas’s langurs. However, even though the breeding season is not restricted, there tends to be an increase in mating when the following weaning period is expected to correspond with an abundance of food. The gestation period lasts 5 to 6 months. At the end of gestation, P. thomasi female produce one young at a time.
Jurnal Primatologi Indonesia, Vol. 13, No. 2, Juli 2016, hlm. 1-2ISSN 1410-5373
Picture 1 Left, Presbytis thomasi (kedih) at Bukit Lawang, North Sumatra (source: Richard Gonzalez); right, white kedih at Taman Margasatwa Ragunan, Jakarta (source: Walberto Sinaga)

Females rarely produce more than one offspring at a time, and never more than two. Weaning occurs at 12 to 15 months, after which an offspring is supposed to become independent. Although they are fully independent, juveniles do not reach sexual maturity until 4 or 5 years of age (Colobus and Leaf Monkeys 2001, Eimerl and DeVore 1965).
The average lifespan of P. thomasi is 20 years. In captivity, the average lifespan is 29 years. The nine-year difference may be due to numerous factors such as the destruction of habitat, hunting by humans, the presence of natural predators, and attacks between neighboring groups (Colobus and Leaf Monkeys 2001).
These langurs spend most of the day in groups either resting, feeding, or moving. These groups usually consist of five females and one male, but there can also be small groups of males or individual males living alone.
Within these groups, there is a dominance hierarchy that everyone, both male and female, are subject to a dominance in hierarchy. It has been suggested that an individual’s rank within the hierarchy may depend on its age or its ability to defend itself against others in the group (Colobus and Leaf Monkeys 2001, Eimerl and DeVore 1965, Gurmaya 1986, Steenbeek et al. 1999).
Even though this langur is known for having a calmer demeanor in gestures and responses to conspecifics than other primates, competition has been observed within groups. Competition increases with the size of the group, and the femlaes show a preference for smaller groups due to the decrease in risk of infanticide. Group sizes are often dependent upon female dispersal (Colobus and Leaf Monkeys 2001, Steenbeck and van Schaik 2001).
Many factors influence a female’s decision to leave her group and join another. These include competition for food, the risk of predation, and avoidance of inbreeding.
More important is the avoidance of infanticide. Often a female leaves her group to protect her young from infanticide (Sterck 1997).
ReferencesAndromeda Oxford Ltd. 2001. Colobus and
Leaf Monkeys. Pp. 380-393 in D Macdonald, ed. The Encyclopedia of Mammals, Vol. II: Primates and Large Herbivores, 2nd Edition. 132 West 31st Street, New York NY 10001: Facts on File, Inc.
Colijn E, Muchtar M. 1996. Primates of Indonesia-Presbytis Thomasi (Collet 1893) (On-line). The Indonesian Nature Conservation Database. Accessed February 08, 2004 at http://www.nature-conservation.or.id/primates/presbytis_thomasi.html.
Eimerl S, DeVore I. 1965. Life Nature Library: The Primates. New York: Time-Life Books.
Flannery S. 2004. Thomas Leaf-monkey (Presbytis thomasi) (On-line). The Primata. Accessed April 10, 2004 at http://members.tripod.com/uakari/presbytis_thomasi.html.
Gurmaya K. 1986. Ecology and Behavior of Presbytis thomasi in Northern Sumatra. Primates, 27(2): 151-172.
Nowak R. 1999. Primates; Family CERCOPITHECIDAE: Old World Monkeys. Pp. 599-600 in Walker›s Mammals of the World, Vol. I, 6th Edition. Baltimore and London: The Johns Hopkins University Press.
Steenbeck R, van Schaik C. 2001. Competition and group sive in Thomas’s langurs (Presbytis thomasi): the folivore paradox revisited. Behavioral Ecology and Sociobiology, 49(2-3): 100-110. Accessed March 07, 2004.
Steenbeek R, Piek R, van Buul M, van Hooff J. 1999. Vigilance in wild Thomas’s langurs (Presbytis thomasi): the importance of infanticide risk. Behavioral Ecology and Sociobiology, 45: 137-150. Accessed February 11, 2004.
Sterck E. 1997. Determinants of female dispersal in Thomas Langurs. Am J of Primatol, 42(3): 179-198.
Sterck E. 2002. Predator sensitive foraging in Thomas langurs. Pp. 74-91 in L Miller, ed. Eat or be eaten: Predator sensitive foraging among primates. Cambridge, UK: Cambridge University Press.
Swindler D. 1998. Introduction to the Primates. Seattle and London: University of Washington Press.
2 Jurnal Primatologi Indonesia, Volume 13, Nomor 2, Juli 2016, hlm. 1-2

Jurnal Primatologi Indonesia, Vol 13, Nomor 2, Juli 2016, hlm. 3-7ISSN 1410-5373
Aktivitas Harian Orangutan Sumatera (Pongo abelii, Lesson 1827) di Hutan Batang Toru Blok Barat Sumatera Utara
Daily Activities of the Sumatran Orangutans (Pongo abelii Lesson 1827) in Western Batang Toru Forest, North Sumatera
Khakim MFR1*, Mardiastuti A2, Iskandar E3
1Program Studi Primatologi, Program Multidisiplin, Sekolah Pascasarjana IPB2Departemen Konservasi Sumberdaya Hutan dan Ekowisata IPB
3Pusat Studi Satwa Primata IPB
*Korespondensi: [email protected]
Abstract. As the only surviving population in the southern Lake Toba, Sumatran orangutans in Batang Toru forest have a very high conservation value. The threat of poaching is still common in the surrounding forests of Batang Toru, including illegal mining by nearby community, habitat loss due to land conversion, as well as conflicts between humans and orangutans. This study aims to collect information of daily activity of orangutans by age and sex as part of the Sumatran orangutan conservation efforts. This study was conducted for 8 months, from October 2013 to June 2014. The daily activity data was collected using the focal animal sampling and ad libitum. The behaviour of orangutans was followed from the moment they woke up in the morning, till they return to their nest at night, whose data were recorded using the San Anselmo Standardization. The daily behaviour of the female orangutans consisted of feeding (65.2 to 67.3%), moving (15-16%), and resting (11-15%) while the males consisted of feeding (57-60%), moving (15-20%), and resting (13-18%). The feeding behavior has the highest proportion of all other behaviour due to the high activity of moving and being in the highland region. Based on statistical analysis, there was not significant differences in behaviour between adult females and males (χ² = 8.713; p-value = 0.190; α = 0.05) as well as the behaviour of adolescent females and males (χ² = 4.284; p-value = 0.369; α = 0.05).
Key words : activities, Batang Toru forest, behaviour, female, male, Sumatran Orangutan, to 7.34
Pendahuluan
Pada saat ini orangutan dibagi menjadi dua spesies, di Pulau Sumatera yaitu Pongo abelii dan di Pulau Kalimantan Pongo pygmaeus yang selanjutnya dibagi menjadi tiga subspesies yaitu P. p pygmaeus, P. p morio, dan P. p wurmbii (Goossen et al. 2009). Orangutan Sumatera tersebar di Kawasan Ekosistem Leuser dan sekitarnya, di sebelah utara Danau Toba, namun berdasarkan Wich et al. (2003), orangutan sumatera juga tersebar di bagian selatan Danau Toba, yaitu di Hutan Batang Toru.
Dalam publikasi terbaru disebutkan bahwa orangutan betina di Kawasan Hutan Batang Toru memiliki keunikan secara genetik, orangutan betina lebih dekat kekerabatannya dengan orangutan di Pulau Kalimantan dibanding dengan orangutan sumatera di kawasan lain (Nater et al. 2011). Populasi orangutan terbesar di Sumatera dijumpai di Leuser Barat (2,508 individu) dan Leuser Timur (1,052 individu), serta Rawa Singkil (1,500 individu). Menurut Departemen Kehutanan (2007) yang diperkuat
Wich et al. (2008) terdapat populasi lain yang diperkirakan potensial untuk bertahan dalam jangka panjang yaitu di Batang Toru Barat dengan populasi sekitar 400 individu dan Batang Toru Timur atau Sarulla Timur dengan estimasi populasi sekitar 150 individu.
Ancaman perburuan masih kerap terjadi di sekitar hutan Batang Toru, termasuk penambangan ilegal masyarakat di sekitar stasiun penelitian, serta kehilangan habitat akibat konversi lahan Etnis Nias yang datang ke hutan Batang Toru. Konflik antara manusia dengan orangutan juga kerap terjadi, khususnya saat musim panen durian. Hal tersebut menjadi bagian dari pemicu timbulnya ancaman terhadap keberadaan satwa di hutan Batang Toru. Sebagai satu-satunya populasi yang masih bertahan di kawasan selatan Danau Toba, orangutan Sumatera di hutan Batang Toru memiliki nilai konservasi yang sangat tinggi. Oleh karena itu, penelitian ini bertujuan untuk mengumpulkan informasi berupa aktivitas harian orangutan berdasarkan kelas umur dan jenis kelamin sebagai bagian upaya konservasi orangutan di hutan Batang Toru.

Gambar 1 Peta Situasi Stasiun Riset Yayasan Ekosistem Lestari-Program Konservasi Orangutan Sumatera
Metode Penelitian
Lokasi dan waktu
Lokasi penelitian berada di Stasiun Riset Yayasan Ekosistem Lestari-Program Konservasi Orangutan Sumatera di Kawasan Hutan Batang Toru Blok Barat selama bulan Oktober 2013 sampai dengan bulan Februari 2014. Lokasi stasiun riset terletak di Kabupaten Tapanuli Utara dengan luas kawasan sekitar 12 km2 dengan total panjang transek lebih dari 30 km (Gambar 1). Stasiun riset berada pada ketinggian yang bervariasi antara 850-1100 mdpl (Khakim 2010).
Alat dan obyek penelitianAlat yang digunakan selama penelitian
satu unit binokuler 8x42, satu unit GPS Garmin 60 Map, kamera digital, jam tangan digital, piranti lunak SPSS 16 for windows dan ArcMap 10.1, alat tulis, kertas koran untuk spesimen tumbuhan pakan yang belum diidentifikasi, dan data. Obyek penelitian orangutan dan vegetasi yang menjadi komponen habitat orangutan.
Metode pengumpulan dataPengumpulan data perilaku harian
dilakukan dengan focal animal sampling dan ad libitum (Altmann 1974). Data perilaku dicatat sesuai dengan standarisasi San Anselmo (Morogh-Bernard dan McLardy 2002), dengan
mengikuti individu orangutan mulai dari sarang pagi hari hingga kembali ke sarang malam hari, dengan interval pencatatan tiap 2 menit (Martin dan Bateson 1993) dan bila memungkinkan terhadap individu lain yang dijumpai selama penelitian berlangsung (di Fiore 2004). Sampai dengan saat ini terdapat 17 individu yang telah diidentifikasi. Namun demikian hanya 7 individu yang telah terhabituasi, dengan susunan 1 individu betina dewasa dengan anak, 1 individu betina dewasa tanpa anak, 2 individu jantan dewasa, 1 individu remaja jantan, dan 1 individu remaja betina.
Analisis Data
Analisis berdasarkan pengamatan perilaku dilakukan secara deskriptif. Untuk membandingkan pola perilaku makan antara orangutan dewasa, remaja, dan anak menggunakan uji khi kuadrat (χ²) dengan bantuan program SPSS 16.0.
Hasil dan Pembahasan
Selama penelitian dicatat 113 perjump-aan dengan orangutan, dengan total mengikuti individu fokal selama 96.624 menit. Individu-fokal yang paling sering diikuti dijelaskan pada Tabel 1. Tujuh individu fokal terse-but merupakan orangutan yang telah terha-bituasi dengan pengamat, sehingga perilaku saat diikuti merupakan perilaku normal.
4 Khakim et al., Aktivitas harian Orangutan Sumatera

Perilaku HarianHasil pengamatan tiga kelas umur pada
individu betina menunjukkan proporsi makan terjadi antara 65,2 - 67,3%. Proporsi perilaku sosial dan bersuara menempati proporsi terkecil selama penelitian. Betina dewasa memiliki proporsi tertinggi dalam waktu makannya. Anakan memiliki proporsi berpindah yang lebih tinggi dibandingkan dua kelas umur lainnya. Oleh karena itu proporsi istirahat bernilai paling kecil. Perilaku sosial berupa berkutu-kutuan (grooming) masuk dalam perilaku sosial, namun proporsi selama penelitian sangat kecil (0,19%). Betina anak, masih sering dapat diamati menyusu pada Beta. Secara keseluruhan perilaku orangutan betina dewasa, orangutan betina remaja, dan orangutan anak dijelaskan pada Gambar 2.
Selama penelitian tidak dijumpai individu fokal anak jantan, sehingga penelitian fokus terhadap jantan dewasa dan jantan remaja. Jan-tan dewasa memiliki proporsi makan (60,11%) yang lebih tinggi dibanding dengan jantan remaja (56,56%). Hal ini berbeda dengan betina dewasa yang lebih banyak waktu makannya. Namun demikian jantan dewasa lebih tinggi proporsi berpindah dan istirahat. Vokalisasi
Tabel 1 Individu fokal yang paling sering dijumpai/diikuti
Gambar 2 Proporsi aktivitas betina pada tiga kelas umur (dewasa, remaja, dan anak)
lebih sering ketika mengeluarkan panggilan panjang (long call). Proporsi perilaku jantan dewasa dan jantan remaja dipaparkan pada Gambar 3.
Berdasarkan analisis statistik, tidak ada perbedaan perilaku yang nyata antara betina dewasa dengan jantan dewasa (χ² =8,713; p-value = 0,190; α = 0,05). Begitu juga dengan perilaku betina remaja dan jantan remaja yang menunjukkan tidak ada perbedaan perilaku nyata (χ²= 4,284; p-value = 0,369; α = 0,05).
PembahasanPopulasi orangutan Batang Toru pertama
kali disurvei pada tahun 1997 (Meijaard 1997 dalam Wich et al. 2014). Orangutan Batang Toru merupakan populasi yang terisolasi dari populasi orangutan sumatera lainnya, serta satu-satunya populasi orangutan yang hidup di habitat bagian selatan Danau Toba. Populasi orangutan di Batang Toru berkisar antara 400-600 individu yang tersebar di Blok Barat dan Blok Timur. Nilai populasi tersebut berkisar 15% dari total populasi orangutan Sumatera yang tersisa (Wich et al. 2008). Berdasarkan penelitian sebelumnya, nilai minimum populasi orangutan di Stasiun Penelitian
Jurnal Primatologi Indonesia, Volume 13, Nomor 2, Juli 2016, hlm. 3-7 5

SimpulanPerilaku harian orangutan betina terdiri
dari makan (65,2-67,3%), bergerak (15-16%), dan beristirahat (11-15%), dan yang jantan terdiri dari makan (57-60%), bergerak (15-20%), beristirahat (13-18%). Proporsi waktu makan lebih tinggi di banding lokasi yang lain karena tingginya aktivitas bergerak serta berada di wilayah dataran tinggi. Berdasarkan analisis statistik, tidak ada perbedaan perilaku yang nyata antara betina dewasa dengan jantan dewasa.
Ucapan Terimakasih
Ucapan terima kasih disampaikan kepada Yayasan Ekositem Lestari-Sumatran Orangutan Conservation Programme selaku sponsor atas terlaksananya penelitian ini. Terima kasih juga atas bantuan Gabriella Fredriksson, P.hD serta Matthew Nowak atas dukungannya. Tidak lupa ucapan terima kasih untuk asisten peneliti di Lapangan Alamsyah, Ulil Amri, Leonardy, Ettyson, dan Dosmaratua.
Daftar PustakaAltmann J. 1974. Observational study of
behavior: Sampling methods. Allee Laboratory of Animal Behavior, University of Chicago. U.S.A.
Departemen Kehutanan. 2007. Strategi dan Rencana Aksi Konservasi Orangutan Indonesia 2007-2017. Departemen Kehutanan. Jakarta.
Di Fiore A. 2004. Diet and feeding ecology of woolly monkeys in a Western Amazonian Rain Forest. International Journal of Primatology,Vol. 25(4) : 767-801.
Batang Toru 17 individu. Nilai rerata kepadatan populasi orangutan di Stasiun Penelitian Batang Toru 0,23 ind/km2 (Wich et al. 2014). Kepadatan populasi orangutan di Batang Toru memiliki nilai kepadatan terkecil untuk seluruh lokasi di Sumatera. Sebagai perbandingan, nilai kepadatan populasi orangutan di lokasi lain, yaitu Suaq Balimbing mencapai 7 ind/km2, sedangkan di Ketambe kepadatan populasi berkisar antara 3-5 ind/km2 (Atmoko dan van Schaik 2010).
Berdasarkan studi orangutan di Suaq Balimbing, orangutan jantan dewasa berbantalan pipi (flanged) menghabiskan 48,0% waktunya untuk makan, untuk beristirahat sebesar 34,9%, dan bergerak 14,9%, bagi jantan dewasa tidak berbantalan pipi (unflanged) menghabiskan 53,6% untuk makan, untuk beristirahat sebesar 25,7%, dan bergerak 17,9%. Betina dewasa yang aktif secara seksual di Suaq Balimbing menghabiskan 54,9% aktivitas hariannya untuk waktu makan 25,9% untuk beristirahat, dan untuk bergerak 16,9%. Studi lain di Ke-tambe orangutan jantan dewasa berbantalan pipi menghabiskan waktu 48,3% waktunya untuk makan, 41,9% untuk beristirahat, dan 9,2 untuk bergerak, untuk jantan dewasa tak berbantalan pipi menghabiskan 52,6%, untuk makan, beristirahat sebesar 33,5%, dan untuk bergerak sebesar 13,0%. Bagi betina dewasa yang aktif secara seksual 55,4% digunakan untuk waktu makannya, untuk beristirahat sebesar 32,0%, dan untuk bergerak sebesar 12,1% (Morrogh-Bernard et al. 2009). Berdasarkan penelitian Wich et al. (2014), orangutan di Batang Toru memiliki proporsi makan hanya bernilai 47,8%, istirahat 20,8%, berpindah 29,9%, dan 1,6% untuk aktivitas lain. Berdasarkan penelitian ini, diperoleh nilai proporsi waktu makan lebih tinggi di banding kedua lokasi lain di Sumatera. Hal ini kemungkinan karena daya jelajah harian yang tinggi, serta berada di wilayah dataran tinggi (850-1100 mdpl), sehingga membutuhkan asupan energi yang lebih tinggi pula.
Gambar 3 Proporsi aktivitas jantan dewasa (kiri) dan remaja (kanan)
6 Khakim et al., Aktivitas harian Orangutan Sumatera

Goosens B, Chikhi L, Jalil MF, James S, Acrenaz M, Lackman-Acrenaz I, Bruford MW. Taxonomy, geographic variation and population genetics of Bornean and Sumatran orangutan. Di dalam: Wich SA, Utami-Atmoko SS, Setia TM, van Shaik CP. (Editor). Orangutans, Geographic in Behavioral Ecology and Conservation. Oxford, hal 1 - 14.
Khakim MFR. 2010. Yearly Report Yayasan Ekosistem Lestari. Tidak Dipublikasikan.
Martin PR, Bateson P. 1993. Measuring Behaviour: An Introductory Guide. Cambridge (UK): Cambridge University Press.
Morogh-Bernard H, McLardy C. 2002. Orangutan data collection standardisation. San Anselmo (US).
Morogh-Bernard, Husson SJ, Knott CD, Wich SA, van Schaik CP, van Noordwijk MA, Lackman-Acrenaz I, Marshall AJ, Kanamori T, Kuze N, bin Sakong R. 2009. Di dalam: Wich SA, Utami-Atmoko SS, Setia TM, van Shaik CP. (Editor). .Orangutans, geographic in behavioral ecology and conservation. Oxford University Press, hal 119-134. New York.
Nater A, Nietlisbach P, Arora N, van Schaik CP, van Noordwijk MA, Willems, Erik P, Singleton I, Wich SA, Goossens B, Warren KS, Verschoor EJ, Farajallah DP, Pamungkas J, Krützen M. 2011. Sex-biased dispersal and volcanic activities shaped phylogeographic patterns of extant orangutans (genus: Pongo). Molecular Biology and Evolution. J Mol. Biol. Evol. 28(8):2275–2288.
Utami-Atmoko SS, van Schaik CP. 2010. The natural history of Sumatran orangutan. Dalam : Gursky-Doyen S dan Supriatna J. (Editor). Indonesian Primates. New York (US): Springer hal 41-55. New York.
Wich SA, Meijaard E, Marshall AJ, Husson S, Acrenaz M, Lacy RC, van Schaik CP, Sugardjito J, Simorangkir T, Traylor-Holzer K, Doughty M, Supriatna J, Dennis R, Gumal M, Knott CD, Singleton I. 2008. Distribution and conservation status of the orang-utan (Pongo spp.) on Borneo and Sumatra:how many remain?. Fauna & Flora International, Oryx, 42(3), 329–339.
Wich SA, Usher G, Peters HH, Mokhamad Faesal Rakhman Khakim, Nowak MG, Fredriksson GM. 2014. Preliminary data on the highland Sumatran Orangutans (Pongo abelii) of Batang Toru. Di dalam: Grow NB, Gursky-Doyen S, Krzton A (Editor). New York: Springer hal 265-283.
Jurnal Primatologi Indonesia, Volume 13, Nomor 2, Juli 2016, hlm. 3-7 7

Jurnal Primatologi Indonesia, Vol 13, Nomor 2, Juli 2016, hlm. 8-13ISSN 1410-5373
Diseminasi dan Mutasi Gen ENV GP70 Simian retrovirus Serotipe-2 (SRV-2) pada Jaringan Macaca fascicularis
Dissemination and Mutation of Simian retrovirus serotype-2 (SRV-2) env gp70 Gene in Macaca fascicularis Tissues
Kristiana V1, Saepuloh U2*, Iskandriati D1,2, Pamungkas J2,3
1 Fakultas Teknobiologi, Universitas Katolik Indonesia Atma Jaya2 Laboratorium Mikrobiologi dan Imunologi, Pusat Studi Satwa Primata,
Lembaga Penelitian dan Pengabdian kepada Masyarakat, Institut Pertanian Bogor3Fakultas Kedokteran Hewan, Institut Pertanian Bogor
*Korespondensi: [email protected]
Abstract. Simian type D retroviruses (SRVs) cause simian acquired immune deficiency syndrome (SAIDS) in macaques with similar clinical signs to human AIDS. Thus, SRV can be used as a model of retrovirus infection in human. In this study, it was reported that tissues dissemination of the SRV serotype-2 naturally infect Macaca fascicularis from Indonesia. The presence of virus was detected from several tissues by polymerase chain reaction with specific primers that partially recognized env gp70 of SRV-2. Proviral SRV-2 was identified in peripheral blood mononuclear cell (PBMCs) and tissues, including lymphoid and non-lymphoid tissues. This showed broad distribution of SRV-2 in naturally infected macaques. The sequences of env gp70 were then aligned to determine variation among isolates that had been distributed in tissues. Nucleotide sequences alignment of env gp70 among isolates indicated high homology at the identities of 97.1 - 99.2% compared to PBMCs positive control (Mf ET1006). All sequences had close genetic relationship as indicated by the same cluster with SRV-2 reference sequences.
Key words : dissemination, env gp70 gene , Macaca fascicularis, SRV-2
Pendahuluan
Simian retrovirus Tipe D (SRV) adalah Betaretrovirus yang banyak ditemukan menginfeksi satwa primata genus Macaca. SRV merupakan patogen yang fatal bagi primata dan dilaporkan menyebabkan infeksi endemik di beberapa pusat penelitian primata di Amerika Serikat pada sekitar tahun 1980-an. Hingga kini, SRV telah ditemukan sebanyak tujuh serotipe.
SRV seringkali dikaitkan sebagai penyebab SAIDS (Simian Acquired Immuno Deficiency Syndrome), suatu sindrom penurunan sistem kekebalan tubuh atau imunodefisiensi pada simian (Marx et al. 1985, Gardner et al. 19888). SAIDS memiliki gejala klinis yang mirip dengan AIDS pada manusia (Daniel et al. 1984; Lerche et al.1995). Beberapa kesamaan gejala klinis ini menyebabkan SRV digunakan sebagai model untuk melihat aspek patogenitas retrovirus pada manusia.
Salah satu aspek patogenitas yang pen-ting adalah diseminasi virus dalam jaringan tu-buh. Beberapa jenis virus dapat menyebabkan infeksi yang terlokalisasi pada organ tertentu,
sementara beberapa lainnya dapat menyebab-kan infeksi yang tergeneralisasi (Guzman et al. 1999; Chopra & Mason, 1970). Untuk melihat diseminasi virus, pengamatan langsung pada berbagai organ manusia tidak memungkinkan untuk dilakukan. Penggunaan satwa primata, terutama genus Macaca, menjadi alternatif yang baik, karena kemiripan anatomi, fungsi fi-siologis, dan genetika dengan manusia.
Studi mengenai diseminasi SRV pada jaringan Macaca merupakan salah satu model untuk melihat penyebaran retrovirus pada manusia (Lackner et al. 1988). Pada studi yang dilakukan Wilkinson et al. (2003), sampel darah M. fascicularis yang berasal dari koloni endemik terinfeksi SRV-2 sengaja diinokulasikan melalui pembuluh darah (intravena) ke M. fascicularis yang bebas infeksi SRV untuk melihat penyebaran virus tersebut dalam jaringan. Hasilnya memperlihatkan adanya variasi penyebaran SRV ke beberapa jaringan tubuh, baik karena perbedaan respon antibodi tubuh maupun kemampuan transmisi virus. Pada penelitian ini, dilakukan analisis diseminasi SRV-2 pada M. fascicularis yang terinfeksi secara alami di habitat aslinya (Indonesia).

Dalam penyebarannya pada jaringan tubuh, SRV perlu mengenali reseptor pada sel inang untuk dapat masuk ke dalam sel dan melakukan proses replikasi. Bagian virus yang berfungsi untuk mengenali reseptor pada sel inang adalah selubung (envelope) virus. Selubung virus ini disandikan gen env pada genom SRV. Selubung virus menjadi faktor yang penting dalam tropis virus pada jaringan. Potensi patogenik virus salah satunya dipengaruhi variasi sekuen gen env (Philipp-Staheli et al. 2006). Data mengenai pengaruh diseminasi SRV terhadap terjadinya mutasi pada gen env belum tersedia. Oleh karena itu, analisis sekuen pun dilakukan pada gen env SRV yang telah terdistribusi pada jaringan.
Penelitian ini dilakukan untuk mengetahui diseminasi SRV-2 pada beberapa jaringan M. fascicularis asal Indonesia yang terinfeksi secara alami dengan mendeteksi keberadaan gen env gp70 SRV-2 menggunakan teknik PCR. Selain itu, ingin diketahui pula kemungkinan terjadinya mutasi pada gen env gp70 dari SRV-2 yang telah terdistribusi pada jaringan M. fascicularis dengan membandingkan sekuen gen env gp70 SRV-2 tiap jaringan yang terinfeksi dengan sekuen kontrol.
Materi dan Metode
Sampel jaringanSampel yang digunakan adalah sampel
jaringan (ginjal, hati, paru-paru, timus, yeyunum, kelenjar getah bening, limpa, otak, jantung) dan sel darah tepi berinti tunggal (peripheral blood mononuclear cells/PBMCs) M. fascicularis yang telah dibuktikan positif terinfeksi SRV-2 secara alami dengan uji PCR (data sekunder). Sampel merupakan koleksi Pusat Studi Satwa Primata, Lembaga Penelitian dan Pengabdian kepada Masyarakat-Institut Pertanian Bogor (PSSP, LPPM-IPB).Ekstraksi DNA
Ekstraksi DNA dilakukan dengan menggunakan QIAamp DNA Blood Mini Kit (Qiagen) dengan mengikuti cara kerja yang diberikan dalam manual.
Amplifikasi DNA dengan PCRDaerah target yang akan diamplifikasi
adalah sebagian daerah env gp70 dari SRV-2. Primer yang digunakan adalah SRV-1/3 5737U21 (5’-CCAGATGGCTACCAGAACGAY-3’) dan SRV-2 6243L22 (5’-CAAAACTGTGGGGACATTGCTA-3’) (Iskandriati et al. 2010). Master mix yang digunakan GoTaq Green Master Mix (Promega).
Kondisi PCR diatur dengan tahapan awal berupa pre-denaturasi pada suhu 94oC selama 10 menit. Tahap selanjutnya, adalah tahap amplifikasi dengan 40 siklus pengulangan, meliputi denaturasi pada suhu 94oC selama 30 detik, penempelan primer pada suhu 62oC selama 30 detik, dan ekstensi pada suhu 72oC selama 30 detik. Tahapan terakhir, pasca ekstensi pada suhu 72oC selama 7 menit dan dilanjutkan dengan penyimpanan pada suhu 4oC.
Elektroforesis dan visualisasi DNAHasil PCR lalu divisualisasi dengan
elektroforesis horisontal (100 Volt, 45 menit) pada gel agarosa 1,5% dengan penambahan 10 µL etidium bromida (10 mg/ml) dan diamati pada UV-transiluminator (GelDoc, Bio-Rad). Penanda DNA yang digunakan 100 bp DNA ladder (Invitrogen). Hasil PCR yang positif ditunjukkan adanya pita fragmen DNA yang berukuran 506 pasang basa.
Purifikasi DNA gel agarosaPita DNA positif yang dihasilkan
dipurifikasi gel agarosa menggunakan QIAquick Gel Extraction Kit (Qiagen) dengan mengikuti cara kerja yang diberikan dalam manual.
Desekuenan (Perunutan)DNA yang telah dipurifikasi dari gel
agarosa, kemudian dirunut dengan menggunakan primer SRV-1/3 5737U. Sampel dikirim untuk disekuen di P.T. Charoen Phokpand Indonesia.
Analisis bioinformatikaRunutan yang diperoleh diolah lebih
lanjut dengan menggunakan program BioEdit (Ibis Biosciences). Runutan tiap sampel lalu disejajarkan menggunakan program ClustalW (European Bioinformatics Institute/EBI) dengan kontrol runutan isolat SRV-2 asal M. fascicularis asli Indonesia (ET1006) (Iskandriati et al. 2010) dan runutan SRV-2 galur D2/MNE/WA yang dikulturkan pada sel A549 (A549/SRV2). Penyejajaran juga dilakukan dengan beberapa runutan pembanding dari basis data GenBank, yaitu SRV-2 galur D2/RHE/OR/V1 (AF126468.1), SRV-2 galur D2/RHE/OR (AF126467.1), SRV-2 complete genome (M16605.1), SRV-1 complete genome (M11841), SRV-3/Mason Pfizer Monkey Virus, MPMV (AF033815.1), SRV-4 (NC_014474.1), SRV-5 (AB611707.1) dan SRV-8 (NC_031326.1). Hasil penyejajaran digunakan untuk membuat pohon filogenetika dengan program MEGA 6.0. Pohon filogenetika dibangun menggunakan metode neighbor-joining dengan uji bootstrap pada 1000 kali pengulangan.
Jurnal Primatologi Indonesia, Volume 13, Nomor 2, Juli 2016, hlm. 8-13 9

Hasil dan PembahasanAmplifikasi dengan teknik PCR dilakukan
untuk mengidentifikasikan keberadaan DNA provirus SRV-2 pada sampel yang dianalisis. Hasil positif ditunjukkan adanya pita berukuran sekitar 506 pasang basa. Hasil amplifikasi menunjukkan terdapat tujuh sampel jaringan yang positif, yaitu paru-paru, yeyunum, kelenjar getah bening, limpa, otak, jantung, dan PBMCs (Gambar 1). Ini menunjukkan infeksi SRV-2 secara alami dapat berdiseminasi hingga ke jaringan-jaringan tersebut.
Purifikasi fragmen DNA dilakukan pada hasil PCR yang positif, kecuali hasil PCR sampel jaringan otak, karena hasilnya terlalu sedikit. Hasil purifikasi, seperti yang dapat dilihat pada Gambar 2, menunjukkan pita dengan variasi ketebalan pita yang mirip dengan hasil PCR. Ini menunjukkan keberhasilan proses purifikasi fragmen DNA yang dilakukan.
Diseminasi virus pada jaringan berhasil dilihat dengan mendeteksi keberadaan gen env gp70 SRV-2 pada sampel jaringan menggunakan teknik PCR. Ketebalan pita yang menunjukkan proviral SRV-2 paling besar teridentifikasi pada PBMCs dan jaringan limfoid lain seperti kelenjar getah bening dan limpa. Hal ini diperkirakan karena jalur penyebaran virus dalam tubuh melalui darah, disamping sel target dari retrovirus adalah sel-sel kekebalan tubuh. Akibatnya gejala utama infeksi retrovirus adalah penurunan sistem kekebalan tubuh hewan yang terinfeksi. SRV-2 juga menunjukkan diseminasi yang cukup luas ke beberapa organ non-limfoid, misalnya yeyunum dan paru-paru. Fenomena ini mendukung adanya infeksi oportunistik SAIDS yaitu diare yang persisten dan radang paru-paru (pneumonia). Diseminasi yang meluas ke beberapa beberapa jaringan ini dapat disebabkan rendahnya respon antibodi tubuh
yang dapat menetralisir virus pada M. fascicularis tersebut (Wilkinson et al. 2003).
Keenam fragmen DNA hasil purifikasi kemudian dirunut dengan menggunakan primer SRV-1/3 5737U. Runutan gen env SRV-2 dari enam jaringan kemudian disejajarkan secara bertingkat (multiple alignment) dengan runutan pembanding. Penyejajaran yang dilakukan adalah penyejajaran nukleotida gen env gp70 antar isolat asal jaringan dengan runutan kontrol ET1006 dan runutan A549/SRV-2.
Dari hasil penyejajaran yang dilakukan, dilihat persentase kemiripan antar sekuen isolat asal jaringan dengan isolat PBMCs asal Mf ET1006 sebgai kontrol positif, seperti yang dapat dilihat pada Tabel 1. Pada tabel tersebut dapat dilihat bahwa kemiripan (homology) antar sekuen isolat jaringan tersebut menunjukkan persen identitas nukleotida antara 97,4% sampai dengan 99,2%.
Penyejajaran nukleotida juga dilakukan antar isolat asal jaringan dengan isolat retrovirus referensi baik antar SRV-2 maupun SRV-1, SRV-3, SRV-4 dan SRV-5. Hasil penyejajaran kemudian digunakan untuk membangun pohon filogenetika. Pohon filogenetika yang dibentuk berdasarkan hasil penyejajaran nukleotida gen env gp70 antar isolat asal jaringan dengan sekuen kontrol ET1006 dan A549/SRV-2 maupun SRV 1-5 asal Genbank dapat dilihat pada Gambar 3.
Analisis penyejajaran nukleotida gen env gp70 antar sekuen isolat asal jaringan yang dibandingkan dengan isolat kontrol (PBMCs asal ET1006) menunjukkan kesamaan identitas nukleotida pada kisaran antar 96,0% sampai dengan 99,1%. Persentase kemiripan tertinggi adalah pada isolat PBMCs yang mencapai 99,1%, paru-paru 98,3% dan jejunum 97,6%; selanjutnya jaringan limpa 96,9% dan cairan getah bening sebesar 96,5%. Dari gambaran
Gambar 1 Hasil visualisasi PCR gen, env gp70 SRV-2, dari sampel beberapa jaringan asal M. fascicularis yang terinfeksi SRV-2. Marker 100 bp ladder (Invitrogen) (1), ginjal (3), paru-paru (4), hati (5), timus (6), yeyunum (7), kelenjar getah bening (8), limpa (9), otak (10), jantung (11), PBMCs (12), kontrol SRV-2 (13)
10 Kristiana et al., Diseminasi dan mutasi gen

Gambar 2 Hasil purifikasi fragmen gen, env gp70 SRV-2. Marker 100 bp ladder (1), ginjal (3), paru-paru (3), yeyunum (4), kelenjar getah bening (5), limpa (6), jantung (7), PBMCs (8)
Tabel 1 Persentase kemiripan hasil penyejajaran runutan nukleotida di antara keenam isolat asal jaringan dengan kontrol isolat SRV asal Indonesia (ET1006) dan dibandingkan dengan SRV-2 referensi Genbank
Gambar 3 Pohon filo-genetik hasil penyejajaran nukleotida gen env gp70 SRV-2 isolat asal jaringan dengan isolat SRV-2 asal referensi dan SRV-1, 3, 4, dan 5
Jurnal Primatologi Indonesia, Volume 13, Nomor 2, Juli 2016, hlm. 8-13 11
data tersebut dapat dilihat bahwa persentase identitas homologi pada jaringan non-limfoid (PBMCs, yeyunum dan paru-paru) lebih besar dibandingkan dengan jaringan jaringan limfoid (limpa dan cairan getah bening). Ini berarti mutasi nukleotida virus SRV-2 pada jaringan non-limfoid lebih kecil dibandingkan dengan
mutasi pada jaringan asal limfoid, walaupun mutasi yang terjadi sangat kecil, sehingga masih berada dalam kluster yang sama dalam pohon filogenetik. Hal ini kemungkinan berhubungan dengan kemampuan virus untuk menghindar dari sistem kekebalan tubuh pada organ-organ limfoid melalui mekanisme mutasi.
1 2 3 4 5 6 7 8 9 10 1. Jantung - 95,7 96,4 95,0 95,5 97,8 96,0 94,8 95,5 92,0 2. Jejunum - 97,6 95,8 96,2 98,6 97,6 95,1 95,8 93,0 3. Paru-paru - 96,5 96,9 99,2 98,3 95,8 96,5 93,7 4. Kelenjar getah
bening - 95,1 97,4 96,5 94,2 94,8 92,1
5. Limpa - 97,8 96,9 94,4 95,1 92,3 6. PBMCs - 99,1 96,7 97,4 94,6 7. ET1006 - 97,4 98,1 94,2 8. SRV2_AF126467 - 97,7 93,1 9. SRV2_AF126468 - 92,9
10. SRV2_M16605 -

Pada pohon filogenetika hasil penyejajaran nukleotida env gp70 isolat asal jaringan (Gambar 3), isolat dapat dilihat membentuk satu kluster bersama-sama dengan kontrol positif ET1006 dan SRV-2 referensi yaitu AF126467, AF126468, dan M16605. Variasi nukleotida env gp70 antar isolat jaringan sangat kecil dan memiliki kemiripan yang tinggi dengan isolat lapang ET1006. Hal ini diperkirakan bahwa proviral SRV-2 pada jaringan bersifat stabil dan hanya sedikit mengalami mutasi padahal daerah gen yang dideteksi adalah daerah envelop yang memiliki variabilitas cukup tinggi dibanding daerah gen lainnya (Philipp-Staheli et al. 2006). Adapun sekuen kontrol yang digunakan yaitu SRV-2 ET1006 yang merupakan isolat lapang asal Indonesia menunjukkan adanya kedekatan antara isolat SRV-2 asal Indonesia dengan isolat SRV-2 strain D2/CEL/OR, vD2/RHE/OR dan D2/RHE/ORVI. Hal ini memperkuat asumsi bahwa isolat ET1006 merupakan isolat lapang SRV-2 yang diisolasi di pusat penelitian primata Amerika Serikat (Iskandriati et al. 2010).
SRV-1 dan SRV-3 membentuk kelompok sendiri yang terpisah dengan kelompok SRV-2. Ini menunjukkan kekerabatan yang lebih dekat antara SRV-1 dengan SRV-3, dibandingkan dengan SRV-2. Demikian juga dengan SRV-4 yang memiliki kedekatan dengan SRV-8 dan membentuk kluster tersendiri terpisah dari kelompok SRV-2 (Zao et al. 2010; Zao et al. 2016). SRV-5 juga membentuk kluster yang terpisah dengan SRV-2 demikian juga terhadap SRV-1/SRV-3 dan SRV-4/SRV-8 (Takano et al. 2013).
Secara umum, gen env gp70 dari SRV-2 menunjukkan adanya sedikit variasi. Tingkat variasi yang rendah mengindikasikan genom SRV-2 yang cukup stabil. Hasil ini sesuai dengan penelitian sebelumnya yang menunjukkan adanya variasi pada gen env gp70 dengan tingkat variasi yang tidak terlalu signifikan (Rosenblum et al. 2000).
Simpulan
Diseminasi SRV-2 yang menginfeksi M. fascicularis secara alami berhasil diamati dengan mengamplifikasi daerah gen env gp70 SRV-2 asal beberapa jaringan. SRV-2 menunjukkan diseminasi yang cukup luas ke beberapa jaringan, yaitu jaringan paru-paru, yeynum, kelenjar getah bening, limpa, otak, jantung, dan PBMCs. Analisis bioinformatika menunjukkan adanya homologi yang tinggi pada gen env gp70 SRV-2 yang terdistribusi pada jaringan tersebut.
Untuk menggambarkan dengan jelas diseminasi virus dan pengaruhnya pada patogenitas virus, pengamatan diseminasi perlu dilakukan secara menyeluruh pada seluruh jaringan tubuh.
Ucapan Terimakasih
Penelitian ini didanai Pusat Studi Satwa Primata, Lembaga Penelitian dan Pengabdian kepada Masyarakat–Institut Pertanian Bogor (PSSP, LPPM-IPB).
Daftar Pustaka
Chopra HC, Mason MM. 1970. A new virus in a spontaneous mammary tumor of a rhesus monkey. J Cancer Res 30:2081-2086.
Daniel MD, King NW, Letvin NL, Hunt RD, Seghal PK, Desrosiers RC. 1984. A new type D retrovirus isolated from macaques with an immunodeficiency syndrome. J Science 223:602-605
Gardner MB, Luciw P, Lerche N, Marx P. 1988. Nonhuman primate retrovirus isolates and AIDS. J Adv Vet Sci and Comp Med 32:171-190
Guzman RE, Kerlin RL, Zimmerman TE. 1999. Histologic lesions in cynomolgus monkeys (Macaca fasccularis) naturally infected with simian retrovirus type D: Comparison of seropositive, virus-positive, and uninfected animals. J Toxicol Pathol 27(6):672-677.
Iskandriati D, Saepuloh U, Mariya S, Grant RF, Solihin DD, Sajuthi D, Pamungkas J. 2010. Isolation and characterization of simian retrovirus type D from Macaca fascicularis and M. nemestrina in Indonesia. J Microbiol Indones 4(3): 132-136
Lackner AA, Rodriguez MH, Bush CE, Munn RJ, Kwang HS, Moore PF, Osborn KG, Marx PA, Gardner MB, Lowenstine LJ. 1988. Distribution of a macaque immunosuppressive type D retrovirus in neural, lymphoid, and salivary tissues. J Virol 62(6): 2134-2142
Lerche NW, Heneine W, Kaplan JE, Spira T, Yee JL, Khabbaz RF. 1995. An expanded search for human infection with simian type D retrovirus. AIDS Res Hum Retroviruses 11(4):527-529
12 Kristiana et al., Diseminasi dan mutasi gen

Jurnal Primatologi Indonesia, Volume 13, Nomor 2, Juli 2016, hlm. 8-13 13
Marx PA, Bryant ML, Osborn KG, Maul DH, Lerche NW, Lowenstine LJ, Kluge JD, Zaiss CP, Henrickson RV, Shiigi SM. 1985. Isolation of a new serotype of simian acquired immune deficiency syndrome type D retrovirus from Celebes black macaques (Macaca nigra) with immune deficiency and retroperitoneal fibromastosis. J Virol 56(2):571-578.
Philipp-Staheli J, Marquardt T, Thouless ME, Bruce AG, Grant RF, Tsai CC, Rose TM. 2006. Genetic variability of the envelope gene of type D simian retrovirus-2 (SRV-2) subtypes associated with SAIDS-related retroperitonial fibromatosis in different macaque species. J Virol 3:11.
Rosenblum LS, Weiss RA, McClure MO. 2000. Virus load and sequence variation in simian retrovirus type 2 infection. J Virol 74(8):3449-3454.
Takano JI, Leon A, Kato M, Abe Y, Fujimoto K. 2013. Isolation and DNA characterization of a simian retrovirus 5 from a Japanese monkey (Macaca fuscata). J Gen Virol 94: 955–959.
Wilkinson RC, Murrell CK, Guy R, Davis G, Hall JM, North DC, Rose NJ, Almond N. 2003. Persistence and dissemination of simian retrovirus type 2 DNA in relation to viremia, seroresponse, and experimental transmissibility in Macaca fascicularis. J Virol 77 (20):10751-10759.
Zao CL, Armstrong K , Tomanek L, Cooke A, Berger R, Estep JS, Marx PA, Trask JS, Smith DG, Yee J, Lerche NW. 2010. The Complete Genome and Genetic Characteristics of SRV-4 Isolated from Cynomolgus Monkeys (Macaca fascicularis. J Virol 405(2): 390–396.
Zao CL, Tomanek L,Cooke A, Berger R, Yang L, Xie C, Chen S, Shi C, Rong R. 2016. A novel simian retrovirus subtype discovered in cynomolgus monkeys (Macaca fascicularis). J Gen Virol (manuscript).

Pendahuluan
Kukang adalah satwa nokturnal, primata prosimian arboreal yang tersebar di sepanjang Asia Tenggara. Kukang Sumatera dapat ditemukan di Pulau Sumatera hingga semenanjung Malaysia. Walaupun status konservasi kukang Sumatera saat ini tergolong rentan (vulnerable), jumlah kukang di pasar hewan jauh melampui kestabilan populasi mereka di alam. Hal ini menyebabkan peningkatan perhatian dunia untuk memasukkan semua genus Nycticebus ke dalam status Appendix I di CITES pada tahun 2007. Pusat Rehabilitasi Satwa Primata Yayasan Inisiasi Alam Rehabilitasi Indonesia (PRS YIARI) yang berdiri sejak tahun 2008, telah menjalankan program konservasi kukang Indonesia secara holistik, meliputi kegiatan penyelamatan, rehabilitasi, pelepasliaran, monitoring, edukasi, dan kampanye kepada masyarakat. Hasil pemeriksaan umum kukang eks peliharaan yang telah diterima PRS YIARI sejak tahun 2011-2015, menyatakan bahwa 34 ekor dari total 68 ekor kukang menderita metabolic bone disease (MBD) [3]. Studi kasus ini membahas tentang kejadian komplikasi MBD dan urolithiasis pada seekor kukang sumatera di YIARI.
Materi dan MetodeSinyalemen
Seekor kukang sumatera jantan dan dewasa bernama Omen dengan bobot 500 gram (Gambar 1), diserahkan oleh pemiliknya ke Pusat Rehabilitasi Satwa Primata YIARI pada tanggal 14 Mei 2016. Sebelumnya kukang tersebut telah dipelihara selama setahun.
Komplikasi MBD dan Urolithiasis pada Kukang Sumatera(Nycticebus coucang) di YIARI Bogor
Complications of MBD and Urolithiasis of Sumatran Kukang(Nycticebus coucang) in YIARI Bogor
Priambada NP¹*, Prameswari W¹, Sanchez KL¹
¹Yayasan Inisiasi Alam Rehabilitasi Indonesia Bogor
*Korespondensi: [email protected]
Abstract. The sumatran loris (Nycticebus coucang) is a nocturnal prosimian arboreal primate that can be found in Sumatra to Peninsular Malaysia. The number of hunting and trading of Sumatran lorises as a very high-priced pet causes its conservation to be protected under UU No. 5 year 1990. The loris conditions that are maintained by the community do not fulfill the aspect of animal welfare, causing poor condition of the slow loris, which lead to death. Primate Rehabilitation Center of Indonesia, the Rehabilitation Initiation Foundation (PRS YIARI), which was established in 2008, has run the conservation program of lemurs Indonesia, including rescue, rehabilitation, release and education. 50% of the total 68 pet lorises received by PRS YIARI in 2011-2015 were found to have bone disease (MBD). This case study discussed the incidence of MBD complications and urolithiasis in a Sumatra loris in YIARI. The diagnosis refered to an initial prosthesis examination that showed bending in the long bones, and a thin bone cortex. And there was a clearly bounded radiopaque mass in the abdominal area that was pressed to one side only, which indicated a full vesica urinaria. The results of blood biochemical examination showed a low calcium value with a ratio of imbalanced calcium and phosphorus. In addition, a urine analysis also showed an alkaline pH, and many struvit crystals were found through a urine cytology. Administration of calcium, urolith dissolution and antibiotics gave a positive result in this case.
Key words : hypocalcemia, loris, MBD, struvit, urolithiasis
Gambar 1 Kukang sumatera jantan yang sakit
Amnesis
Saat dipelihara biasa diberi makan serangga dan buah-buahan. Beberapa minggu
Jurnal Primatologi Indonesia, Vol 13, Nomor 2, Juli 2016, hlm. 14-18ISSN 1410-5373

Jurnal Primatologi Indonesia, Volume 13, Nomor 2, Juli 2016, hlm. 14-18 15
Gambaran hematologi pada tanggal 17 dan 25 Mei 2016 pada Tabel 1 menunjukkan kondisi leukosit sedikit di bawah normal dan perlahan meningkat ke arah normal. Selain itu satwa juga ditemukan dalam kondisi anemik di awal perawatan.
sebelum diserahkan, kondisinya melemah, cara jalan berbeda dan rambut rontok. Hingga tiba di YIARI, kukang sering dapat dilihat tidak aktif dan sulit untuk bergerak, nafsu makan masih baik, tetapi jarang ditemukan feses di kandang setiap harinya.
Gejala klinis
Kelemahan, sulit berjalan, alopesia dan posisi tidur abnormal. Seminggu kemudian ditemukan stranguria hingga disuria dan juga konstipasi.
Hasil pemeriksaan fisik
Secara umum satwa terlihat kurus (BCS 2), dengan bobot sekitar 500 gram. Lemah, dehidrasi dan stres dengan kondisi rambut kusam dan rontok, terutama di area perut dan inguinal. Kondisi gigi sudah terpotong, tulang panjang seperti radius-ulna terasa membengkok, satwa terlihat kesulitan untuk bergerak secara normal. Palpasi area abdomen secara umum terasa besar dan keras, kondisi usus penuh feses dapat teraba, namun juga terasa bentukan seperti kantong penuh air.
Uji pendukung
Pemeriksaan rongen posisi ventrodorsal dan lateral kiri, pemeriksaan hematologi dan biokimia darah, uji kandungan urin (dip stick), sitologi urin di lab PSSP.
Hasil dan PembahasanHasil
Pemeriksaan rontgen awal dapat dilihat pada Gambar 2 menunjukkan adanya pembengkokan di tulang panjang dan korteks tulang yang tipis. Serial rontgen di periode minggu-minggu selanjutnya menunjukkan terdesak hanya kesatu sisi oleh massa radiopaque berbatas jelas, yang kemudian terindikasi sebagai vesika urinaria yang penuh.
Gambar 2 Pemeriksaan rontgen awal
Pada Tabel 2, biokimia darah menunjukkan nilai kalsium yang rendah dengan rasio kalsium dan fosfor yang tidak berimbang. Pada Tabel 3 dapat dilihat hasil analisis uji urin pada tiga hari yang berbeda.
Diagnosis
Komplikasi MBD dengan urolithiasis
Diagnosis Diferensial
Infeksi saluran perkemihan, cystitis
Prognosis
Dubious
Terapi
Pemberian Calcium Borogluconate 100 mg/kg bobot badan, dilanjutkan dengan suplemen kalsium oral, antibiotik (enrofloxacin 5 mg/BB), analgesik (tramadol 5 mg), vitamin D 0,1 ml , disolusi medik (Batugin elixir 1 ml), laksatif (0,3ml) dan enema, terdapat pada Tabel 4.
Kataterisasi telah dilakukan hingga 2 kali namun selalu tertahan pada kedalaman 7 cm, tanpa ada cairan yang bisa tertarik ataupun komponen lainnya (Gambar 3).
Tabel 1Gambaran Hematologi

16 Priambada et al., Komplikasi MBD dan Urolithiasis pada Kukang Sumatera
Cystocentesis dilakukan 3 kali pada tanggal 24 Mei 2016, 2 Juni 2016 dan 12 Juni 2016 dengan jumlah cairan urin 15-45ml dengan endapan kristal.
Pembahasan
Metabolic bone disease (MBD) adalah penyakit yang umumnya dikaitkan dengan hilangnya densitas tulang (seperti yang ditemui pada osteopenia) ataupun pelunakan tulang baik lokal maupun sistemik seperti yang biasa ditemui pada rickets dan osteomalacia. MBD dapat disebabkan berbagai macam faktor, tetapi pada umumnya diakibatkan oleh abnormalitas mineral seperti kalsium dan fosfor, ataupun kekurangan vitamin D. Selain itu MBD juga biasa dikaitkan dengan pemberian pakan yang tidak tepat, kurangnya ekspos sinar matahari ataupun sinar ultraviolet buatan, serta penyakit perkemihan yang sangat parah.
Tabel 2 Biokimia darah
Tabel 3 Uji urin analisis
Uji urin analisis 24 Mei 2016 menunjukkan hasil SG 1.005, pH 6, haemoglobinuria, eritrosituria, nitrituria.Uji urin analisis 2 Juni 2016 menunjukkan hasil SG 1.005, pH 8, leukosituria, bilirubinuria, urobilinogenuria.Uji urin analisis 12 Juni 2016 menunjukkan hasil SG 1.000, pH 8, bilirubinuria, urobilinogenuria.
Gambar 3 Proses pemberian katerisasi

Jurnal Primatologi Indonesia, Volume 13, Nomor 2, Juli 2016, hlm. 14-18 17
Urolithiasis merupakan penyakit yang ditemukan di berbagai macam spesies dan ditandai dengan adanya bentukan polikristal di saluran perkemihan. Urolithiasis adalah peyakit metabolik yang jarang ditemukan pada primata non manusia, tetapi mempunyai kecendrungan untuk lebih sering terjadi pada hewan jantan. Pada primata non manusia, penyakit ini umumnya terjadi di vesika urinaria dan uretra, tetapi memungkinkan juga untuk ditemukan pada ginjal dan ureter. Faktor predisposisi urolithiasis sangat beragam mulai dari supersaturasi faktor predisposisi di urin, metabolisme senyawa-senyawa yang tidak umum dan peningkatan konsumsi diet tertentu. Penderita urolithiasis sering ditemukan dengan gejala stranguria, hematuria, pollakiuria dan rasa sakit di perut, ataupun gejala kolik. Diagnosa dapat dilakukan melalui kombinasi pemeriksaan fisik dan pencitraan melalui radiografi dan ultrasonografi. Pengobatan dapat dilakukan dengan disolusi medis dan juga pengangkatan batu secara fisik.
Urolithiasis jarang ditemui pada primata non-manusia, sehingga dokumentasinya sangat terbatas. Beberapa kasus telah dilaporkan terjadi secara sporadis di berbagai spesies seperti lemur, monyet ekor panjang dan monyet cebus. Menurut hasil studi urolith yang dilakukan Universitas Minnesota, tipe urolith yang paling sering ditemukan pada primata antara lain, calcium carbonat (62%), calcium oxalate (16,6%), calcium fosfat (9,5%), struvit (4,7%) dan cysteine (4,7 %). Batu struvit (MgNH4PO4 6H2O) merupakan bentuk urolith yang umum terjadi pada anjing, kucing dan manusia. Urolit struvit dalam urin steril umumnya hanya ditemukan pada kucing, sedangkan pada anjing pada tubulus distal ginjal. Urolith struvit dapat
ditangani dengan disolusi, pemberian pakan yang dapat menghasilkan pH urine <6,8 sangat dianjurkan sebagai pencegahan.
Pada kasus ini MBD terjadi ditandai dengan menurunnya kadar kalsium dan imbalans rasio antara jumlah kalsium dan fosfor beserta gejala klinis lainnya. Seiring dengan perjalanan MBD, diikuti pula dengan munculnya gangguan saluran perkemihan berupa obstruksi saluran dan manusia kecenderungannya terkait dengan infeksi bakteri penghasil urease. Pembentukan urolith struvit merupakan hasil kombinasi faktor predisposisi seperti spesies, jenis kelamin dan pakan. Struvit lebih mudah larut dalam kondisi urine yang sedikit asam (pH<6,8), sehingga ini juga terkait erat dengan pembentukan urine basa, seperti riwayat sejarah keluarga dengan kasus urolith struvit, diet rendah protein, ataupun kondisi asidosis perkemihan akibat urolith struvit. Kandungan fosfor pada urolith struvit dapat dikorelasikan dengan kondisi satwa yang mengalami abnormalitas metabolisme mineral dalam tubuh sebagai faktor predisposisi penyebab urolithiasis.
Satu bulan pasca terapi, kukang Omen berangsur-angsur membaik (Gambar 4). Konstipasi perlahan-lahan menghilang, bekas urine mulai sering ditemukan di kandang dan palpasi abdomen sudah tidak terasa kondisi pembesaran abdomen. Empat bulan pasca terapi, kondisi kadar kalsium dan fosfor dalam darah juga sudah mulai meningkat dan berimbang. Lima bulan pasca terapi, kukang Omen sudah mulai bergerak dan berjalan normal dengan berat bobot sudah mencapai 800 serta BCS 4 (berat), tanpa ditemukan masalah lainnnya.
Tabel 4 Terapi metabolic bone disease (MBD) dan urolithiasis

18 Priambada et al., Komplikasi MBD dan Urolithiasis Pada Kukang Sumatera
Gambar 4 Kukang sumatera satu bulan pasca terapi
Simpulan
Merujuk pada kejadian kasus dan pembahasan, dapat disimpulkan bahwa telah terjadi komplikasi MBD dan urolithiasis pada seekor kukang sumatra di Yayasan IAR Indonesia. Penanganan berupa pemberian kalsium, disolusi urolith dan pemberian antibiotik, memberikan hasil yang positif pada kasus ini.
Daftar Pustaka
Barker MJM, Herbert RT. 1972. Diseases of the Skeleton. In R. N. T-W-Fienes (Ed.), Pathology of simian primates (pp. 433–519). Basel: S. Karger
Bernard JB. 1997. Vitamin D and ultraviolet radiation: Meeting lighting needs for captive animals. In Nutrition Advisory Group handbook fact sheet 002
Chesney RW. 1984. Metabolic bone diseases. Pediatrics in Review, 5(8), 227–237.Non-Human Primates in Biomedical Research: Diseases. Christian R. Abee, Keith Mansfield, Suzette D. Tardif, Timothy Morris. Academic Press, 2012
Deng F, Ouyang JM. 2006. Comparative investigations of ultrafine crystals in urine of healthy human and lithogenic patients Mater. Sci. Eng.C, vol. 26: 688–69
Ettinger SJ, Feldman C. 2010. Textbook of Veterinary Internal Medicine, 7th edition. Pp 1979-2000.Guillermo CC, Nelson RW. 2013. Small Animal Internal Medicine, 4th edition, pp. 609-611.
Faltas NH. 2000. Urolithiasis in cynomolgus monkey (Macaca fascicularis): a case report. Contemp Top Lab Anim Sci 39, 18-9
Farrel M, Rando C, Garrord B. 2015. Lessons from the Past: Metabolic Bone Disease in Historical Captive Primates. Int J of Primatol. Springer
Hesse A, Tiseluis HG, Jahnen A. 1997b Urinary stones, Diagnosis, Treatment, and Prevention of Recurrence. Karger, Basel, pp. 131-147
Hosey G, Melf V, Pankhurst S. 2009. Zoo animals: Behaviour, management and welfare. Oxford: Oxford University Press
Johnson MM. 2014. Obstructive Urolithiasis in Non-Human Primates. Clinicopathologic conference. November 21st 2014
Nekaris KAI, Nijman V. 2007. CITES proposal highlights rarity of Asian nocturnal primates (Lorisidae : Nycticebus). J Folia Primatol 78:211-214
Pintilie (Popescu) GS, Varga I, Ionescu I, Preda. 2010. Urinary calculi as consequence of renal infection and relationship between their incidence and different dietary factors. Proceedings of the Nutrition Society 69: (OCE3), E269
Prameswari W, Priambada NP, Sachez KL, Winarti I. 2015. Studi kasus : Gambaran kondisi kesehatan kukang (Nycticebus sp) eks peliharaan pada tahun 2011-2015 di YIARI Bogor. Prosiding Konferensi Ilmiah Veteriner Nasional (KIVNAS) ke-17.
Roos C, Boonratana R, Supriatna J, Fellowes JR, Groves CP, Nash SD, Rylands AB, Mittermeier RA. 2014. An updated taxonomy and conservation status review of Asian primates. Asian Primates J 4
Theiler A. 1934. Osteodystrophic disease of domesticated animals. Vet J, 90, 159–175
Zachary JF, McGavin MD. 2011. Pathologic Basis of Veterinary Disease, 5th edition., pp. 643-645.

Jurnal Primatologi Indonesia, Vol 13, Nomor 2, Juli 2016, hlm. 19-23 ISSN 1410-5373
The Value of Hematological and Serum Chemistry of Orangutan (Pongo sp.) Among Ages Groups and Sex Differences – A Case Study in
Taman Safari Indonesia
The Value of Hematological and Serum Chemistry of Orangutan (Pongo sp.) among Age Groups and Sex Differences – A Case Study at
Taman Safari IndonesiaHastuti YT1, Mulia BH1, Widianti A1, Manansang J1, Sajuthi D2,3, Mansjoer SS2, Darusman HS2,3*
1Taman Safari Indonesia, Cisarua, Bogor, Indonesia2Primate Research Center, Bogor Agricultural University (IPB), Bogor, Indonesia
3Faculty of Veterinary Medicine, Bogor, Indonesia
*Korespondensi: [email protected]
Abstract. Orangutan is a world heritage, and any information related to its health, such as physiological profile is valuable to promote the conservation status. This is a case report on Taman Safari Indonesia (TSI) which studied the hematological and blood chemistry value among age groups (adult n=7 and juvenile n=2) and sex (male n=9 and female n=8). Compared to normal reference, overall, differences across all parameters were discovered less in sex differences than in the age conspecific.
Key words : blood chemistry, hematological, Orangutan.
Introduction
As a part of mega-biodiversity country in the world, Indonesia inherited enormous resources, several among which are regarded as world-iconic or world-heritage, and orangutan (Pongo sp.) is known among them (Delegado and van Schaik 2000, Rijksen and Meijaard 1999). It then becomes the responsibility of all nations to keep a decent welfare and conservation of orangutan (van Schaik 2004; Wich et al. 2009).
Taman Safari Indonesia (TSI) is a well-established facility of an ex-situ conservation, typically for several endangered species in Indonesia, including orangutans. TSI also performs the conservation medicine plan on orangutan by providing database of the physiological value, which is regarded highly relevant for the health management plan (Russon 2009) a complete blood test, including hematology and blood chemistry – emphasizing liver and kidney functions.
This study contains the physiological value of hematology and serum chemistry of the orangutans at TSI. From this documentation, it is hoped that the values can be used as a reference for orangutans in ex-situ conservation facility, and finally may enrich and support the conservation status of orangutans, and add to the evidence of their blood profile. There are a few studies that publish the data (McPherson 2013).
Materials and Methods
The study used 9 orangutan which were further grouped by age – 7 adults and 2 juveniles; and 17 orangutan which differed by sex – 9 males and 8 females. This study was conducted at TSI, and blood was taken under sedation. The procedures follow the institutional animal care and use committee at TSI, which assured animal welfare during maintenance and sampling procedures.
Results and DiscussionHematology
The values of hematology that are based on sex and age group differences are presented in Table 1, Table 2 and Table 3, respectively.
Overall, the hematology value between male and female were considered normal, with the female being higher than the male, and a high variety between both sexs – showing that there is a potential difference between the male and female orangutans. On the other hand, the MCV values varied less between male and female, indicating that these values were relatively similar although the value itself – compared with the reference – was high. Above trend it was also found in the MPV data that the value was higher than reference, and the differences between sexs were less.

The hematology values in juveniles and adult orangutans were basically normal; however, the adults have higher MCV than the juveniles, while the MPV were higher than the adult orangutans. The MPV value differences were found mostly between male and female.
*Mean Corpuscular Volume**Mean Corpuscular Hemoglobin*** Mean Corpuscular Hemoglobin Concentration
Table 2 Mean values of hematology classified by age groups
*Mean Corpuscular Volume**Mean Corpuscular Hemoglobin*** Mean Corpuscular Hemoglobin Concentration
Among the values, there was one baby-female with high level of WBC, MCV, MCH, Plt, lymphocyte and MPV, compared with the references, while the others were limited to MCV and MPV only. The RBC values were reasonably more different among age groups in female.
Table 1 Mean values of hematology classified by sex groups
20 Yohanna et al., The value of hematological and serum chemistry of Orangutan

Jurnal Primatologi Indonesia, Volume 13, Nomor 2, Juli 2016, hlm. 20-23 21
Blood Chemistry
The results of blood chemistry of the liver and kidney functions as well as other metabolic indicators are presented in Table 4 and Table 5.
Table 3 Mean values of hematology classified by age groups
*Mean Corpuscular Volume**Mean Corpuscular Hemoglobin*** Mean Corpuscular Hemoglobin Concentration
The values were found to be similar among age groups, and both share the high level of GPT and creatinine, compared with the references. Higher value of serum chemistry in both sexs was also found of cholesterol level.
Table 4 Mean values of Blood Chemistry among sex groups
*Glutamate-Oksaloasetat Transaminase**Glutamate-Piruvat Transaminase

22 Yohanna et al., The value of hematological and serum chemistry of Orangutan
Table 5 Mean Values of Blood Chemistry among Age Groups of Male Orangutan
*Glutamate-Oksaloasetat Transaminase**Glutamate-Piruvat Transaminase
Compared with the adults, the values of GOT, GPT and cholesterol in juvenile were found to be higher. This can be caused by the differences in the amount of fat tissue among
Tabel 6 Mean Values of Blood Chemistry among Age Groups of Male Orangutan
*Glutamate-Oksaloasetat Transaminase**Glutamate-Piruvat Transaminase
the age group. In female orangutans the values of blood chemistry were uniformly high in GPT and cholesterol in all age groups (Table 6).
Conclusion
The results of hematology and blood chemistry in orangutans at TSI showed that the differences were greater among age groups compared with sex groups. The age and sex interaction was also found as a factor to the values which were indicated by several hematological and blood chemistry levels in male and female age group – which was marked by higher liver and lipid metabolism in younger age groups, in both male and female.
References List
Delagado R, van Schaik CP. 2000. The behavioral ecology and conservation of the orangutan (Pongo pygmaeus): A tale of two islands. J Evol. Anthrop. 9, 201–218.
McPherson JF. 2013. Normal Blood Parameters, Common Disease and Parasites Affecting Captive Non-human Primates. J Primatol. 2:112

Jurnal Primatologi Indonesia, Volume 13, Nomor 2, Juli 2016, hlm. 20-23 23
Rijksen HD, Meijaard E. 1999. Our Vanishing Relative: The Status of Wild Orangutans at the Close of the Twentieth Century (Dordrecht: Kluwer Academic Publishers).
Russon A. 2009. Quick guide – Orangutans. Curr. J Biol. 19(20), R925-927
Van Schaik CP. 2004. Among Orangutans (Cambridge, MA: Harvard University Pr)
Wich SA, Utami SS, Mitra ST, van Schaik CP. 2009. Orangutans: Geographic Variation in Behavioral Ecology and Conservation (Oxford: Oxford University Pr).

Jurnal Primatologi Indonesia, Vol 13, Nomor 2 Juli 2016, hlm. 24-31 ISSN 1410-5373
Pendahuluan
Beruk (Macaca nemestrina) merupakan satwa primata yang termasuk ke dalam famili Cercopithecidae dan termasuk kelompok monyet dunia lama (Napier dan Napier 1967; Grooves 2001). Satwa ini memiliki persebaran di wilayah Asia Tenggara, terutama di dua pulau besar Indonesia, Pulau Sumatera, dan Kalimantan. Habitat beruk berupa dataran rendah dan perbukitan hutan hujan primer (Crockett dan Wilson 1980). Di Pulau Kalimantan, monyet ini biasa dijadikan sebagai alat yang membantu kerja manusia di kebun atau di lahan pertanian, seperti misalnya membantu mengambil buah kelapa dari pohonnya (Nadine Rupert, pers kom). Selain dijadikan sebagai hewan yang diperbantukan, perburuan kepada satwa ini juga terus meningkat, dilihat dari status konservasi yang meningkat menjadi vulnerable (Richardson et al. 2008).
Perilaku harian dibedakan menjadi 5 kelas, yaitu makan (feeding), mencari makan (foraging), berpindah (moving), istirahat (resting), dan sosial (social) (O’Brien dan Kinnaird 1997). Perilaku makan adalah perilaku monyet mengambil makanan, memasukkan makanan kedalam mulut, menyimpannya dalam kantung pipi, dan mengunyah, serta menelan makanan. Mencari makan adalah perilaku monyet memanjat pohon untuk mencari makanan, meraih makanan, menangkap serangga, membolak-
balik daun atau serasah. Berpindah merupakan perilaku berjalan, berlari atau melompat untuk meninggalkan satu lokasi menuju lokasi lainnya. Istirahat merupakan perilaku duduk di tanah atau pohon, autogrooming, tidur, atau tiduran tanpa melakukan perilaku lainnya. Tingkah laku sosial merupakan perilaku yang berkaitan dengan kehidupan kelompok, seperti allogrooming, berkelahi, seksual, dan bermain (O’Brien dan Kinnaird 1997). Perilaku harian anakan menunjukkan interaksi bermain dengan induk betina dan mulai meniru tindakan dan suara jantan dewasa (Syaukani 2012).
Tokuda et al. (1968) melaporkan hasil pengamatan perilaku M. nemestrina di dalam kandang penangkaran di Seattle, Amerika Serikat, tetapi hanya perilaku seksualnya. Bernstein (1972) melaporkan perilaku harian kelompok beruk di kandang buatan menunjukkan pola perilaku diurnal yang jelas dan dipengaruhi keadaan cuaca. Perilaku harian kelompok beruk lain juga diamati dan menunjukkan beberapa tipe perilaku termasuk pola perilaku diurnal (Martenson et al. 1977).
Penelitian untuk mengamati perilaku harian beruk di Indonesia, khususnya di tempat penangkaran belum banyak dilaporkan. Status konservasi beruk yang termasuk rentan menuju kepunahan (vulnerable) mendukung pentingnya penelitian mengenai perilaku harian. Informasi yang didapatkan akan berkontribusi ke dalam upaya konservasi M. nemestrina. Penelitian ini bertujuan mengkaji perilaku harian M.
Perilaku Harian Beruk (Macaca nemestrina) di Fasilitas Penangkaran Pusat Studi Satwa Primata, Institut Pertanian Bogor
The Behavior of the Pig-Tail Macaque (Macaca nemestrina) at The Ex-Situ Captive Breeding Facility, Primate Research Center Bogor Agricultural
UniversityPasetha A1*, Sandriliana D1, Mulyana JS1, Ummah RI1, Anaktototy Y1, Widayati KA1
1Departemen Biologi, Fakultas Matematika dan Ilmu Pengethauan MIPA-IPB
*Korespondensi : [email protected]
Abstract. Macaca nemestrina is classified as a species susceptible to extinction (vulnerable) by IUCN 2008; however, research on the daily behavior of M. nemestrina in Indonesian captivity has not been widely reported. Therefore, this study is important as a contribution to the breeding program and conservation efforts of these animals. Observations of the M. nemestrina group were made in captivity at the Primate Animal Study Center, Institute for Research and Community Service, IPB, consisting of 1 adult male, 11 adult females and 3 juveniles. The daily behavior used the Focal Animal Sampling method, and recording the daily behavior used the Instant Security Sampling method. The behavior data includes autogrooming (a), aggressive (ag), feeding (fe), feeding (fo), maintenance (g), motion (l), from the seen (oos), position / rest (p) social (s), and drinking (d). Observational data were then analyzed using Pearson Correlation test.
Key words : daily behavior, M. nemestrina, PSSP IPB

nemestrina pada kondisi penangkaran dan mendapatkan gambaran pola dan proporsi perilaku harian M. nemestrina selama berada di dalam penangkaran. Selain itu, tujuan penelitian ini juga untuk menganalisis kemungkinan perbedaan proporsi perilaku harian, perilaku makan dan mencari makan pada M. nemestrina jantan, betina, dan juvenil.
Materi dan Metode
Penelitian dilaksanakan pada bulan Maret – Mei 2017 di penangkaran Pusat Studi Satwa Primata, Lembaga Penelitian dan Pengabdian kepada Masyarakat, IPB (PSSP LPPM-IPB). Analisis data dilakukan di Divisi Fungsi Hayati dan Perilaku Hewan Departemen Biologi, FMIPA IPB. Materi yang diamati yaitu M. nemestrina jantan, betina, dan anakan dari yang berada di kandang koral B (kB).
Alat untuk penelitian meliputi: timer, alat tulis, pakaian scrub, sepatu boot, head cap, gloves, masker, dan face shield.
Habituasi dan Identifikasi Individu
Habituasi dilakukan selama dua minggu untuk mengenalkan dan membiasakan satwa dengan observer dan observer dengan satwa. Masing-masing individu kemudian diidentifikasi dan diklasifikasikan menurut tingkatan umur (Andrade et al. 2004). Identifikasi individu dimulai pada bulan Maret. Identifikasi dilakukan dengan mengamati ciri-ciri fisik tertentu pada masing-masing individu seperti adanya bentuk wajah, bekas luka, bentuk dan warna puting susu, ukuran tubuh, bentuk dan ukuran hidung, bentuk dan ukuran pantat, serta raut wajah. Identifikasi individu dilakukan pada semua individu.
Pengamatan dan Pencatatan Data Perilaku
Pengamatan perilaku harian menggunak-an Focal Animal Sampling, yaitu mengikuti dan mengamati satu individu yang diobservasi dalam durasi waktu tertentu. Pencatatan perilaku har-ian menggunakan Instantaneous sampling yaitu mencatat perilaku dengan interval setiap satu
Tabel 1 Komposisi kelompok kB berdasarkan perbedaan kelompok umur dan jenis kelamin di PSSP IPB
menit (Martin dan Bateson 1993). Data perilaku yang dicatat meliputi autogrooming (a), agresif (ag), makan (fe), mencari makan (fo), grooming (g), lokomosi (l), out of seen (oos), posisi atau istirahat (p), sosial (s), dan minum (d). Data ditabulasi menggunakan pivot table. Proporsi perilaku harian dikelompokkan dan dihitung berdasarkan perbedaan waktu diurnal: pagi hari (08.00 – 09.30), siang hari (11.00 – 12.30), dan sore hari (14.00 – 15.30).
Analisis data
Data perilaku harian akan disajikan dalam bentuk persentase, kemudian dianalisis secara deskriptif. Frekuensi tingkah laku adalah banyaknya tingkah laku yang dilakukan setiap individu (Xi) dibagi seluruh tingkah laku yang diamati pada individu tersebut (Yi) dikalikan 100% (Riendriasari et al. 2009). Data selanjutnya dianalisis dengan uji Korelasi Pearson digunakan untuk melihat korelasi antar tingkah perilaku makan dengan kelas umur yang berbeda selama berada di penangkaran.
Hasil dan Pembahasan
Perilaku Harian M. nemestrina Berdasarkan Perbedaan Waktu Diurnal
Dari data yang telah dikoleksi dari bulan Maret sampai dengan Mei 2017 diperoleh data perilaku harian sebanyak 7020 poin atau setara dengan 117 jam. Pada pagi hari M. nemestrina memiliki proporsi perilaku harian yaitu autogrooming (8,60%), agresif (6,10%), minum (0,64%), makan (8,43%), mencari makan (6,42%), grooming (1,81%), lokomosi (18,11%), posisi (47,92%), sosial (1,85%) dan kawin (0,07%). Tengah hari M. nemestrina memiliki proporsi perilaku harian yaitu autogrooming (6,07%), agresif (5,82%), minum (2,14%), makan (23,27%), mencari makan (12,55%), grooming (4,92%), lokomosi (11,01%), posisi (32,74%), sosial (1,39%) dan kawin (0,04%). Pada petang hari M. nemestrina memiliki proporsi perilaku harian yaitu autogrooming (8,67%), agresif (3,88%), minum (0,51%), makan (25,72%), mencari
Jurnal Primatologi Indonesia, Volume 13, Nomor 2, Juli 2016, hlm. 24-31 25

makan (9,39%), grooming (9,07%), lokomosi (10,53%), posisi atau istirahat (28,72%), sosial (3,47%) dan kawin (0,00%).
Proporsi perilaku harian M. nemestrina berdasarkan perbedaan waktu diurnal menunjukkan perbedaan proporsi untuk setiap waktu (Gambar 1). Berdasarkan uji statistika juga tidak ada perbedaan antara proporsi perilaku posisi (P < 0,05).
Menurut O’Brien dan Kinnaird (1997) satwa primata terestrial dan semi arboreal biasanya menunjukkan suatu siklus diurnal pada perilaku hariannya. M. nemestrina merupakan hewan yang aktif pada siang hari. Waktu aktif tersebut digunakan dalam proporsi yang berbeda-beda (O’Brien dan Kinnaird 1997). Dari tiga kategori waktu diurnal, proporsi perilaku harian tertinggi yaitu posisi. Hal tersebut menunjukkan bahwa dalam kondisi penangkaran, M. nemestrina tidak menggunakan strategi hidupnya untuk mencari makan (karena mereka telah terbiasa dengan jadwal makan yang ada) dan juga karena keterbatasan luasan habitat.
Pola perilaku makan memiliki kecenderungan meningkat dari pagi sampai dengan petang hari. Proporsi perilaku makan pada tengah dan petang hari memiliki proporsi yang lebih besar dibandingkan saat pagi hari. Pola perilaku mencari makan menunjukkan sesuatu yang konstan pada tiga kategori waktu diurnal. Suatu organisme ketika mempunyai banyak sumberdaya makanan maka organisme tersebut cenderung sedikit untuk bergerak mencari makanan (Muhlenberg et al. 2004).
Perilaku Harian M. nemestrina Jantan di PSSP IPB
Dari data yang telah dikoleksi dari bulan Maret sampai dengan Mei 2017 diperoleh data perilaku harian jantan sebanyak 2275 poin. Proporsi perilaku harian M. nemestrina jantan selama berada di penangkaran terlihat memiliki perbedaan berdasarkan tiga waktu diurnal (Gambar 2).
Hasil penelitian menunjukkan bahwa perilaku posisi memiliki proporsi paling besar dibandingkan perilaku lainnya pada tiga waktu diurnal. Pada setiap waktu diurnal, perilaku posisi memiliki proporsi yang berbeda. Namun, hasil uji statistika menunjukkan tidak ada perbedaan antara proporsi perilaku posisi pada tiga waktu diurnal (P>0,05 ). Proporsi perilaku makan pada tiga waktu diurnal (10,82%; 22,83%; 24,12%). Berdasarkan uji statistika terdapat perbedaan antara proporsi perilaku makan saat pagi hari dengan siang-sore hari (P<0,05), tetapi tidak terdapat perbedaan proporsi perilaku makan siang dengan sore hari (P>0,05).
Hasil penelitian menunjukkan bahwa perilaku posisi memiliki proporsi paling besar dibandingkan perilaku lainnya pada tiga waktu diurnal. Pada setiap waktu diurnal, perilaku posisi memiliki proporsi yang berbeda. Namun, hasil uji statistika menunjukkan tidak ada perbedaan antara proporsi perilaku posisi pada tiga waktu diurnal (P>0,05). Proporsi perilaku posisi menunjukkan pola seragam pada waktu pengamatan diurnal (Bernstein 1972). Perilaku
26 Pasetha et al., Perilaku harian beruk di fasilitas penangkaran PSSP IPB
Gambar 1 Proporsi perilaku harian M. nemestrina berdasarkan perbedaan waktu diurnal (pagi hari, tengah hari, dan petang hari)
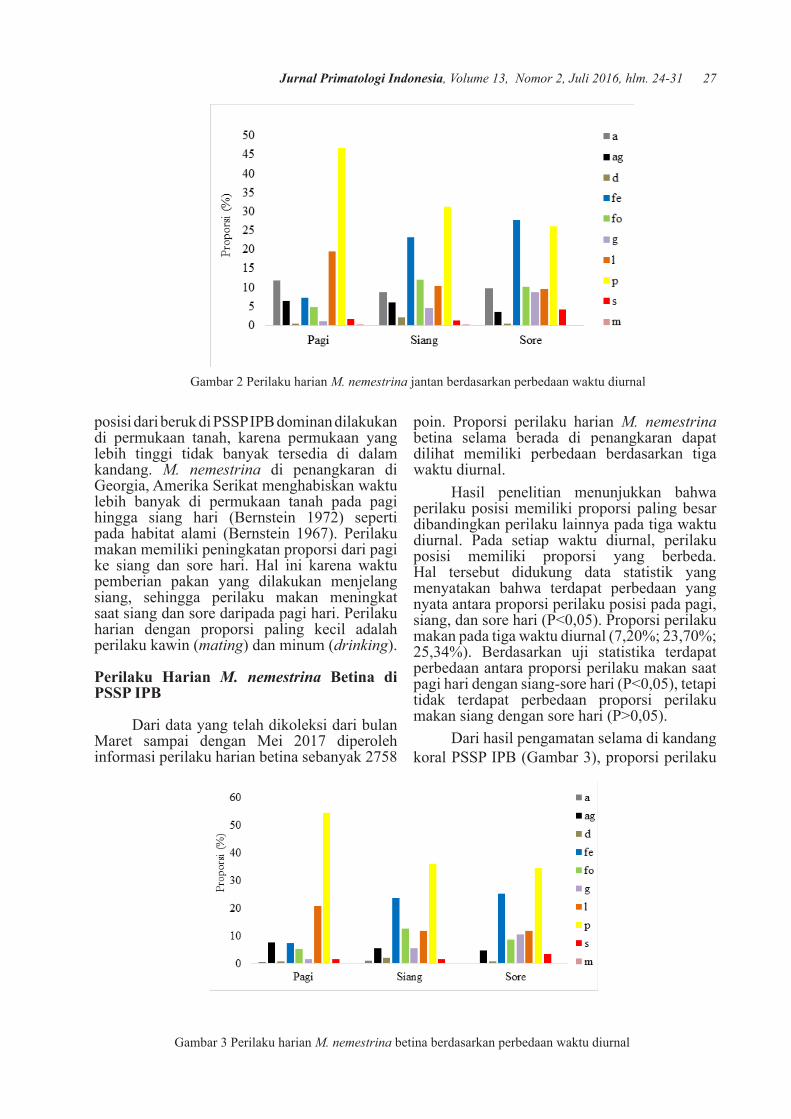
posisi dari beruk di PSSP IPB dominan dilakukan di permukaan tanah, karena permukaan yang lebih tinggi tidak banyak tersedia di dalam kandang. M. nemestrina di penangkaran di Georgia, Amerika Serikat menghabiskan waktu lebih banyak di permukaan tanah pada pagi hingga siang hari (Bernstein 1972) seperti pada habitat alami (Bernstein 1967). Perilaku makan memiliki peningkatan proporsi dari pagi ke siang dan sore hari. Hal ini karena waktu pemberian pakan yang dilakukan menjelang siang, sehingga perilaku makan meningkat saat siang dan sore daripada pagi hari. Perilaku harian dengan proporsi paling kecil adalah perilaku kawin (mating) dan minum (drinking).
Perilaku Harian M. nemestrina Betina di PSSP IPB
Dari data yang telah dikoleksi dari bulan Maret sampai dengan Mei 2017 diperoleh informasi perilaku harian betina sebanyak 2758
poin. Proporsi perilaku harian M. nemestrina betina selama berada di penangkaran dapat dilihat memiliki perbedaan berdasarkan tiga waktu diurnal.
Hasil penelitian menunjukkan bahwa perilaku posisi memiliki proporsi paling besar dibandingkan perilaku lainnya pada tiga waktu diurnal. Pada setiap waktu diurnal, perilaku posisi memiliki proporsi yang berbeda. Hal tersebut didukung data statistik yang menyatakan bahwa terdapat perbedaan yang nyata antara proporsi perilaku posisi pada pagi, siang, dan sore hari (P<0,05). Proporsi perilaku makan pada tiga waktu diurnal (7,20%; 23,70%; 25,34%). Berdasarkan uji statistika terdapat perbedaan antara proporsi perilaku makan saat pagi hari dengan siang-sore hari (P<0,05), tetapi tidak terdapat perbedaan proporsi perilaku makan siang dengan sore hari (P>0,05).
Dari hasil pengamatan selama di kandang koral PSSP IPB (Gambar 3), proporsi perilaku
Jurnal Primatologi Indonesia, Volume 13, Nomor 2, Juli 2016, hlm. 24-31 27
Gambar 2 Perilaku harian M. nemestrina jantan berdasarkan perbedaan waktu diurnal
Gambar 3 Perilaku harian M. nemestrina betina berdasarkan perbedaan waktu diurnal

28 Pasetha et al., Perilaku harian beruk di fasilitas penangkaran PSSP IPB
makan (fe) pada M. nemestrina betina meningkat pada siang dan sore hari. Hal ini karena pemberian makan dilakukan setiap pukul 10.00 WIB yang mempengaruhi pola perilaku makan M. nemestrina, sedangkan proporsi perilaku mencari makan (fo) lebih rendah dibandingkan perilaku makan. Hal ini terjadi karena pihak PSSP telah menyediakan makanan untuk M. nemestrina. Kedua perilaku ini mempengaruhi proporsi perilaku posisi (p) yang menurun pada siang dan sore hari yang pada waktu tersebut, mereka lebih banyak melakukan aktivitas makan dan grooming (g).
Penurunan perilaku autogrooming (a) pada siang dan sore hari dipengaruhi meningkatnya proporsi perilaku grooming pada waktu tersebut. Hal ini terjadi karena M. nemestrina betina lebih banyak melakukan grooming dengan sesamanya. Pada pagi hari, proporsi perilaku lokomosi (l) lebih banyak dilakukan daripada siang dan sore hari. Proporsi perilaku ini seiring dengan proporsi perilaku agresif (ag) yang lebih tinggi pada pagi hari. Kedua perilaku ini menunjukkan bahwa M. nemestrina lebih aktif berinteraksi pada pagi hari daripada sore hari. Pada sore hari, proporsi perilaku sosial (s) dari M. nemestrina betina lebih banyak dilakukan terutama pada betina yang memiliki anakan. Perilaku sosial ini lebih banyak terjadi antara indukan dan anakan yang mulai mengambil posisi untuk beristirahat karena M. nemestrina juga merupakan hewan diurnal.
Perilaku Harian M. nemestrina Anakan di PSSP IPB
Dari data yang telah dikoleksi dari bulan Maret sampai dengan Mei 2017 diperoleh
data perilaku harian juvenil sebanyak 2100 poin. Proporsi perilaku harian M. nemestrina juvenil selama berada di penangkaran terlihat memiliki perbedaan berdasarkan tiga waktu diurnal (Gambar 5).
Hasil penelitian menunjukkan bahwa perilaku posisi memiliki proporsi paling besar dibandingkan perilaku lainnya pada tiga waktu diurnal. Pada setiap waktu diurnal, perilaku posisi memiliki proporsi yang berbeda. Berdasarkan uji statistika menunjukkan ada perbedaan antara proporsi perilaku posisi pada tiga waktu diurnal (P<0,05 ). Pada grafik dapat dilihat adanya kecenderungan perilaku posisi yang berbanding terbalik dengan perilaku makan. Berdasarkan uji statistik, terdapat perbedaan yang nyata antara perilaku makan dan perilaku posisi (P<0,05).
Berdasarkan data yang diperoleh selama pengamatan, tiga perilaku M. nemestrina anakan dengan proporsi tertinggi selama di penangkaran yakni posisi, makan dan autogroming. Ketiga perilaku tersebut termasuk kedalam kategori individual activity (Bernstein 1972). Hasil penelitian yang sama dilaporkan oleh Bernstein pada kelompok Pigtail Macaque di Amerika yang diamati di alam, dimana yang proporsinya individual activity sebesar 21% terjadi di pagi hari (Bernstein 1972).
Proporsi perilaku posisi menurun saat siang dan sore hari, namun terjadi peningkatan perilaku makan, karena pada rentang pagi menjelang siang (sekitar 10.00 WIB) petugas memberikan makan pada M. nemestrina yang ada di penangkaran terebut. Peningkatan proporsi perilaku makan ini terjadi karena ketersediaan makanan yang ada di kandang.
Gambar 5 Perilaku harian M. nemestrina juvenil berdasarkan perbedaan waktu diurnal
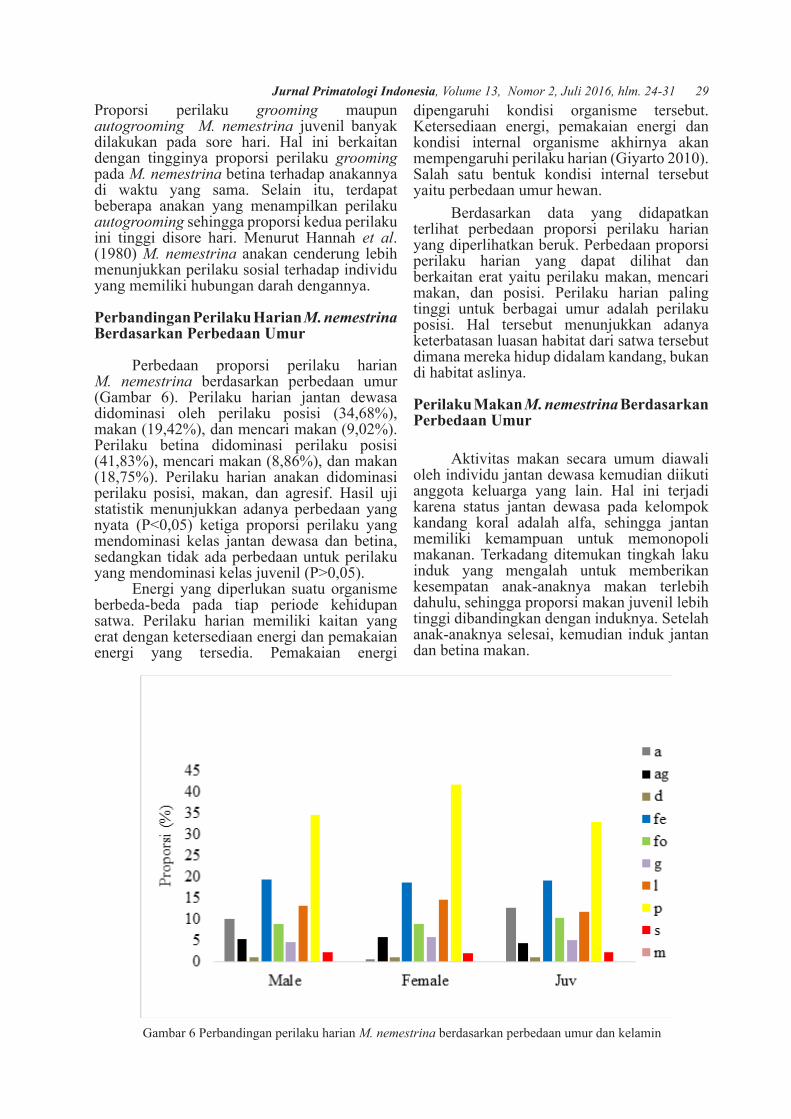
Proporsi perilaku grooming maupun autogrooming M. nemestrina juvenil banyak dilakukan pada sore hari. Hal ini berkaitan dengan tingginya proporsi perilaku grooming pada M. nemestrina betina terhadap anakannya di waktu yang sama. Selain itu, terdapat beberapa anakan yang menampilkan perilaku autogrooming sehingga proporsi kedua perilaku ini tinggi disore hari. Menurut Hannah et al. (1980) M. nemestrina anakan cenderung lebih menunjukkan perilaku sosial terhadap individu yang memiliki hubungan darah dengannya.
Perbandingan Perilaku Harian M. nemestrina Berdasarkan Perbedaan Umur
Perbedaan proporsi perilaku harian M. nemestrina berdasarkan perbedaan umur (Gambar 6). Perilaku harian jantan dewasa didominasi oleh perilaku posisi (34,68%), makan (19,42%), dan mencari makan (9,02%). Perilaku betina didominasi perilaku posisi (41,83%), mencari makan (8,86%), dan makan (18,75%). Perilaku harian anakan didominasi perilaku posisi, makan, dan agresif. Hasil uji statistik menunjukkan adanya perbedaan yang nyata (P<0,05) ketiga proporsi perilaku yang mendominasi kelas jantan dewasa dan betina, sedangkan tidak ada perbedaan untuk perilaku yang mendominasi kelas juvenil (P>0,05).
Energi yang diperlukan suatu organisme berbeda-beda pada tiap periode kehidupan satwa. Perilaku harian memiliki kaitan yang erat dengan ketersediaan energi dan pemakaian energi yang tersedia. Pemakaian energi
dipengaruhi kondisi organisme tersebut. Ketersediaan energi, pemakaian energi dan kondisi internal organisme akhirnya akan mempengaruhi perilaku harian (Giyarto 2010). Salah satu bentuk kondisi internal tersebut yaitu perbedaan umur hewan.
Berdasarkan data yang didapatkan terlihat perbedaan proporsi perilaku harian yang diperlihatkan beruk. Perbedaan proporsi perilaku harian yang dapat dilihat dan berkaitan erat yaitu perilaku makan, mencari makan, dan posisi. Perilaku harian paling tinggi untuk berbagai umur adalah perilaku posisi. Hal tersebut menunjukkan adanya keterbatasan luasan habitat dari satwa tersebut dimana mereka hidup didalam kandang, bukan di habitat aslinya.
Perilaku Makan M. nemestrina Berdasarkan Perbedaan Umur
Aktivitas makan secara umum diawali oleh individu jantan dewasa kemudian diikuti anggota keluarga yang lain. Hal ini terjadi karena status jantan dewasa pada kelompok kandang koral adalah alfa, sehingga jantan memiliki kemampuan untuk memonopoli makanan. Terkadang ditemukan tingkah laku induk yang mengalah untuk memberikan kesempatan anak-anaknya makan terlebih dahulu, sehingga proporsi makan juvenil lebih tinggi dibandingkan dengan induknya. Setelah anak-anaknya selesai, kemudian induk jantan dan betina makan.
Gambar 6 Perbandingan perilaku harian M. nemestrina berdasarkan perbedaan umur dan kelamin
Jurnal Primatologi Indonesia, Volume 13, Nomor 2, Juli 2016, hlm. 24-31 29

Simpulan
Perilaku harian yang erat kaitannya dengan kondisi penangkaran yaitu posisi, makan dan mencari makan. Pola dan proporsi perilaku harian M. nemestrina selama berada di penangkaran berbeda untuk setiap kategori umur dan jenis kelamin. Proporsi perilaku makan anakan selalu lebih kecil daripada jantan dan betina dewasa, sedangkan perilaku posisi pada ketiganya sama. Proporsi perilaku mencari makan lebih kecil daripada perilaku makan.
Daftar Pustaka
Altman J. 1974. Observational Study of Behavior. Sampling Methods. Illinois: Alle Laboratory of Animal Behavior
Bernstein IS. 1967. A field study of the pigtail monkey (Macaca nemestrina). Primates. 8:217-228.
Bernstein IS. 1972. Daily activity cycles and weather influences on a pigtail monkey group. Folia Primat. 18:390-415.
Crockett CM, Wilson WL. 1980. The ecological separation of Macaca nemestrina and Macaca fascicularis in Sumatra. In: The Macaques: Studies in Ecology, Behaviour and Evolution, D.G.L Lindburg (ed). Van Nostrand Reinhold Company. New York. P. 148.
Giyarto. 2010. Penggunaan home range dan aktivitas harian monyet hitam sulawesi (Macaca nigra Desmarest, 1822) di Cagar Alam Tangkoko-Batuangus, Sulawesi Utara [skripsi]. Yogyakarta (ID): Universitas Gadjah Mada.
Groves C. 2001.Primate Taxonomy. Washington (US): Smithsonian Institution Pr.
Hannah MH, Warren W, Holmes G, Medina SR, Sackett GP. 1980. Kin Preference in infant Macaca nemestrina. Nature, 285: 225-227.
[IUCN] International Union for Conservation of Nature. 2008. IUCN Red List of Threatened Species, Pig-Tailed Macaques (Macaca nemestrina) [Internet]. [diunduh 2017 March 20]. Tersedia pada: http://www.iucnredlist.org
Martenson J, Oswald M, Sackett D, Erwin J. 1977. Diurnal variation of common behaviors of pigtail mmonkeys (Macaca nemestrina). Primates. 18:875-882.
Martin P, Bateson P. 1993. Measuring Behaviour: An Introductory guide Second Edition. United Kingdom: Cambridge University Press.
Napier JR, Napier PH. 1967. A Handbook of Living Primates. London (UK): Academic Press.
O’Brien TG, Kinnaird MF. 1997. Behav-ior, diet, and movements of the Su-lawesi crested black macaque (Macaca nigra). Int J Primatol. 18:321-351.
Richardson M, Mittermeier RA, Rylands AB, Konstant B. 2008. Macaca nemes-trina. The IUCN Red List of Threatened Species 2008: e.T12555A3356892.
Riendriasari SD, Entang I, Jansen M, Joko P. 2009. Tingkah Laku Owa Jawa (Hylobates moloch) di Fasilitas Penangkaran Pusat Studi Satwa Primata, Institut Pertanian Bogor. J Primatol Indones. 6:9-13.
Gambar 7 Perilaku harian M. nemestrina juvenil berdasarkan perbedaan waktu diurnal
30 Pasetha et al., Perilaku harian beruk di fasilitas penangkaran PSSP IPB

Syaukani. 2012. Study of population and homerange of thomas langur (Presbytis thomasi) at Soraya Research Station, Leuser Ecosystem. J Natural. 12:37-41
Tokuda K, Simons RC, Jensen GD. 1968. Sexual behavior in a captive group of pigtailed monkeys (Macaca nemestrina). J Primates. 9:283-294.
Jurnal Primatologi Indonesia, Volume 13, Nomor 2, Juli 2016, hlm. 24-31 31
MITRA BESTARI JPI
(Volume 13, Nomor 2, Juli 2016)
Prof Dr Ir Sri Supraptini Mansjoer (PSSP LPPM IPB)Prof Dr Ir Muhammad Bismark, MS (Balitbanghut Dept. Kehutanan)
Prof Dr Ir Ani Mardiastuti, MSc (Fahutan IPB)Prof Dr drh Agik Suprayogi, MSc (FKH IPB)Prof Dr Ir Dewi Apri Astuti, MS (Fapet IPB)
Dr dr Irma H. Suparto, MS (PSSP LPPM IPB)Dr Ir RR Dyah Perwitasari, MSc (FMIPA IPB)
Dr drh Diah Iskandriati (PSSP LPPM IPB)Dr drh Ernie Sulistyowati, MSi (PSSP LPPM IPB)
Dr drh Setyo Widodo (FKH IPB)Dr Ir Entang Iskandar, MSi (PSSP LPPM IPB)
drh Audrey Maria Ungerer (IPB)

1. Jurnal Primatologi Indonesia (JPI) menerima naskah dalam bentuk: a) hasil penelitian, b) catatan penelitian, c) ulasan atau tinjauan pusta-ka, d) laporan kasus, e) paparan program/keg-iatan, dan f) resensi buku. Naskah dapat ditulis dalam bahasa Indonesia maupun Inggris.
2. Naskah tidak sedang dikirim ke atau dievaluasi oleh berkala ilmiah untuk penerbitan. Naskah yang dikirim ke JPI dan dinyatakan diterima oleh Redaksi JPI untuk dimuat dalam JPI men-jadi milik JPI.
Naskah dikirim sebanyak tiga eksemplar ke-pada Dewan Editor JPI, dengan alamat:
Dewan EditorJurnal Primatologi Indonesia
Pusat Studi Satwa Primata Lembaga Penelitian dan Pengabdian kepada
Masyarakat Institut Pertanian BogorJalan Lodaya II No. 5 Bogor 16151
Telepon/Faks: (0251) 8313-637 / (0251) 8360-712Surat Elektronik: [email protected]
Website: http://journal.ipb.ac.id/index.php/primata 3. Naskah ditulis dengan Times New Roman uku-
ran 12 pt, jarak dua spasi, dengan jarak pinggir dua centimeter dan dicetak pada kertas HVS ukuran A4. Gambar, grafik dan tabel disertakan bersama naskah di bagian akhir naskah pada lembar terpisah. Redaksi JPI akan meminta naskah pengetikan (dalam format word for windows: windows 2003/ME/XP) bagi naskah yang dinyatakan diterima untuk dimuat. Pen-giriman naskah (file) dapat melalui e-mail atau compact disc (CD) melalui jasa kantor pos.
4. Naskah hasil penelitian disusun dengan urutan sebagai berikut ini.
a. Judul ditulis dalam bahasa Indonesia dan In-ggris.
b. Nama lengkap penulis. Bila penulis lebih dari satu orang perlu dibubuhkan angka se-cara berurutan untuk keterangan afiliasi lem-baga/institusi tempat bekerja penulis.
c. Nama lembaga/institusi tempat penulis bekerja, disertai dengan alamat, kode pos, telepon, dan faksimili.
d. Nama penulis untuk korespodensi. Kore-spodensi hanya kepada salah satu penulis, maka perlu diberikan tanda khusus bagi yang bersangkutan.
e. Abstrak ditulis tidak lebih dari 300 kata dalam bahasa Indonesia dan Inggris. Pada
abstrak tersebut dicantumkan kata kunci atau key word yang tidak melebihi lima buah kata mengikuti huruf abjad.
f. Pendahuluan. g. Materi dan Metode. h. Hasil dan Pembahasan. i. Simpulan. j. Ucapan Terima Kasih (jika ada), dan k. Daftar Pustaka.
5. Artikel hasil penelitian ditulis maksimum 25 halaman, termasuk lampiran gambar, grafik dan tabel. Artikel lain ditulis maksimum 20 hala-man dan diperbolehkan tidak mencantumkan subjudul sebagaimana tersebut di atas.
6. Penyertaan gambar dalam bentuk foto (hitam putih atau bewarna) harus jelas dan tajam, dengan kontras yang tinggi, berukuran standar kartu pos. Gambar yang dikehendaki dicetak bewarna akan dikenakan biaya tambahan yang dibebankan kepada pengirim naskah.
7. Daftar Pustaka disusun dengan mencantum-kan semua nama penulis yang disusun secara berurutan abjad dan diikuti tahun penerbitan, judul, nama berkala ilmiah atau penerbit buku dan nomor halaman. Penyingkatan nama berka-la ilmiah harus mengikuti singkatan yang ber-laku.
Contoh Penulisan Daftar Pustaka:
Berkala ilmiahBenveniste RE, Morton WR, Clark EA, Tsai CC,
Ochs HD, Ward JM, Kuller L, Knott WB, Hill RW, Gale MJ, Thouless ME. 1988. In-oculation of baboons and macaques with simian immunodeficiency virus/Mne, a primate lenti-virus closely related to Human Immunodefi-ciency Virus Type-2. J Virol. 62:2091-2101.
Suatu bab bukuBarker IK, Van-Dreumel AA, Palmer N. 1996.
The Alimentary System. Di dalam: Jubb KVF, Kennedy PC, Palmer N, editor. Pathology of Domestic Animals. Ed ke-4. New York (US): hlm 91.
BukuHunt TK. 1980. Wound Healing and Wound Infec-
tion: Theory and Surgical Practice. New York: Appleton-Century-Crofts.
PEDOMAN PENULISAN NASKAH

DAFTAR ISI
(Table of Contents)
Volume 13, Nomor 2, Juli 2016
ARTIKEL ASLI (Original Articles)
Khakim MFR, Mardiastuti A, Iskandar E Aktivitas Harian Orangutan Sumatera (Pongo abelii, Lesson 1827) di Hutan Batang Toru Blok Barat Sumatera Utara
Daily Activity of Sumatran Orangutan (Pongo abelii Lesson 1827) in West Batang Toru Forest Block
Kristiana V, Saepuloh U, Iskandariati D, Pamungkas J Diseminasi dan Mutasi Gen ENV GP70 Simian retrovirius Serotipe-2 (SRV-2) pada Jaringan Macaca fascicularis
Dissemination and Mutation of Simian retrovirus serotype-2 ENV GP70 Gene in Macaca fascicularis Tissues
Priambada NP, Prameswari W, Sanchez KL Komplikasi MBD dan Urolithiasis pada Kukang Sumatera (Nycticebus coucang) di YIARI Bogor
Complications MBD and Urolithiasis of Sumatran Kukang (Nycticebus coucang) in YIARI Bogor
Hastuti YT, Mulia BH, Widianti A, Manansang J, Sajuthi D, Mansjoer SS, Darusman HS
Pusat Studi Satwa Primata,
Lembaga Penelitian dan Pengabdian kepada Masyarakat - Institut Pertanian Bogor
JI. Lodaya II, No.5
Bogor, Jawa Barat - 16151
Halaman (Pages)
3
8
The Value of Hematological and Serum Chemistry of Orangutan (Pongo sp.)
Among Ages Groups and Sex Differences – A Case Study in Taman Safari
Indonesia
Pasetha A, Sandriliana D, Mulyana JS, Ummah RI, Anaktototy Y, Widayati KA
Perilaku Harian Beruk (Macaca nemestrina) di Fasilitas Penangkaran Pusat Studi
Satwa Primata, Institut Pertanian Bogor
Behavior of The Pig Tail Macaque (Macaca nemestrina) at The Ex-Situ Captive
Breeding Facility, Primate Research Center Bogor Agricultural University
19
24
14


















