
Virus e malattie auto immuni -...
5
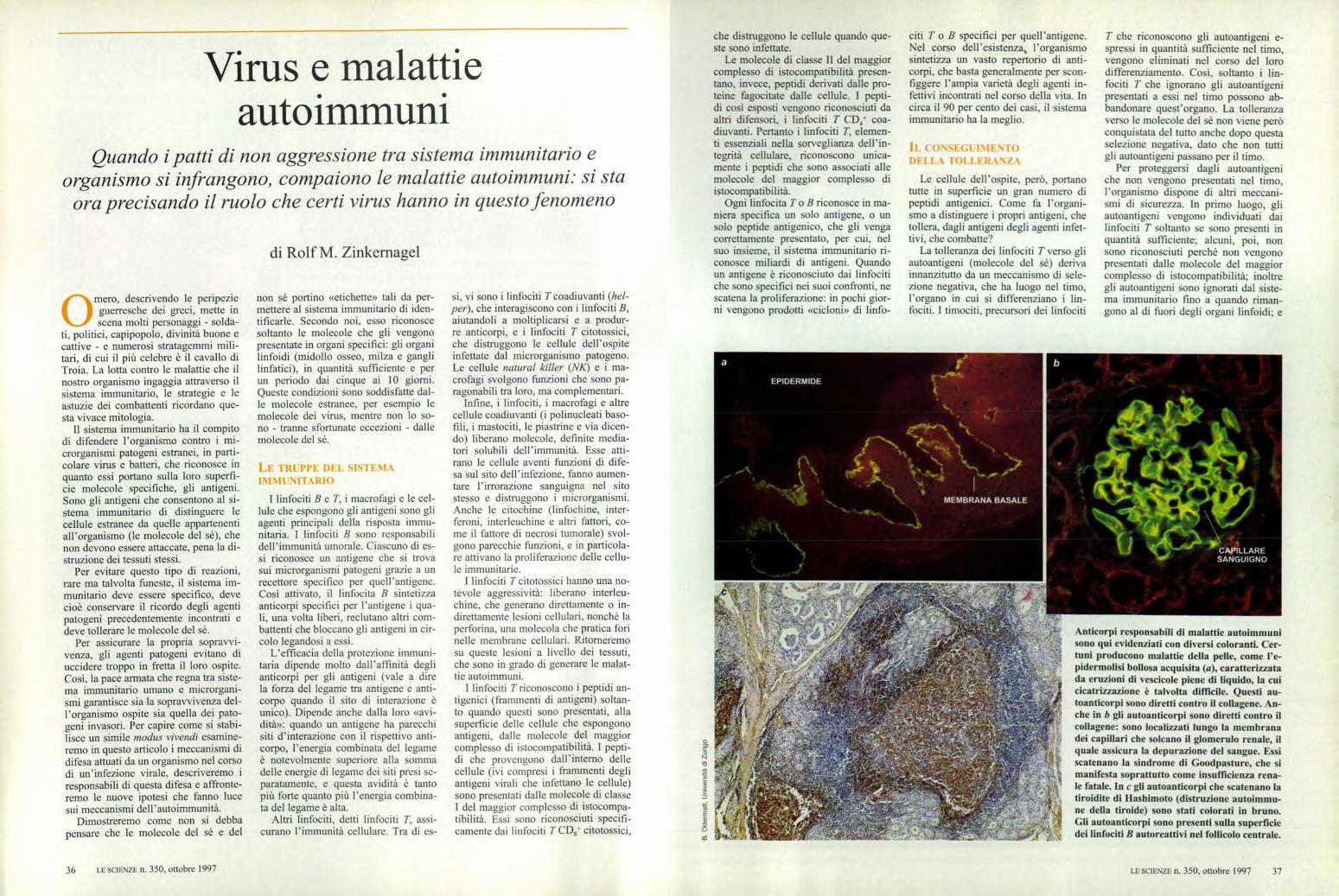
Virus e malattie auto immuni Quando i patti di non aggressione tra sistema immunitario e organismo si infrangono, compaiono le malattie autoimmuni: si sta ora precisando il ruolo che certi virus hanno in questo fenomeno di Rolf M. Zinkernagel Anticorpi responsabili di malattie autoimmuni sono qui evidenziati con diversi coloranti. Cer- tuni producono malattie della pelle, come l'e- pidermolisi bollosa acquisita (a), caratterizzata da eruzioni di vescicole piene di liquido, la cui cicatrizzazione è talvolta difficile. Questi au- toanticorpi sono diretti contro il collagene. An- che in b gli autoanticorpi sono diretti contro il coliagene: sono localizzati lungo la membrana ; dei capillari che solcano il glomerulo renale, il quale assicura la depurazione del sangue. Essi scatenano la sindrome di Goodpasture, che si manifesta soprattutto come insufficienza rena- le fatale. In e gli autoanticorpi che scatenano la tiroidite di Hashimoto (distruzione autoimmu- ne della tiroide) sono stati colorati in bruno. Gli autoanticorpi sono presenti sulla superficie dei linfociti B autoreattivi nel follicolo centrale. t""-.‘ ‘'‘ ,ervi \ i MEMBRANA BASALE a EPIDERMIDE () mero, descrivendo le peripezie guerresche dei greci, mette in scena molti personaggi - solda- ti, politici, capipopolo, divinità buone e cattive - e numerosi stratagemmi mili- tari, di cui il più celebre è il cavallo di Troia. La lotta contro le malattie che il nostro organismo ingaggia attraverso il sistema immunitario, le strategie e le astuzie dei combattenti ricordano que- sta vivace mitologia. Il sistema immunitario ha il compito di difendere l'organismo contro i mi- crorganismi patogeni estranei, in parti- colare virus e batteri, che riconosce in quanto essi portano sulla loro superfi- cie molecole specifiche, gli antigeni. Sono gli antigeni che consentono al si- stema immunitario di distinguere le cellule estranee da quelle appartenenti all'organismo (le molecole del sé), che non devono essere attaccate, pena la di- struzione dei tessuti stessi. Per evitare questo tipo di reazioni, rare ma talvolta funeste, il sistema im- munitario deve essere specifico, deve cioè conservare il ricordo degli agenti patogeni precedentemente incontrati e deve tollerare le molecole del sé. Per assicurare la propria sopravvi- venza, gli agenti patogeni evitano di uccidere troppo in fretta il loro ospite. Così, la pace armata che regna tra siste- ma immunitario umano e microrgani- smi garantisce sia la sopravvivenza del- l'organismo ospite sia quella dei pato- geni invasori. Per capire come si stabi- lisce un simile modus vivendi esamine- remo in questo articolo i meccanismi di difesa attuati da un organismo nel corso di un'infezione virale, descriveremo i responsabili di questa difesa e affronte- remo le nuove ipotesi che fanno luce sui meccanismi dell'autoimmunità. Dimostreremo come non si debba pensare che le molecole del sé e del non sé portino «etichette» tali da per- mettere al sistema immunitario di iden- tificarle. Secondo noi, esso riconosce soltanto le molecole che gli vengono presentate in organi specifici: gli organi linfoidi (midollo osseo, milza e gangli linfatici), in quantità sufficiente e per un periodo dai cinque ai 10 giorni. Queste condizioni sono soddisfatte dal- le molecole estranee, per esempio le molecole dei virus, mentre non lo so- no - tranne sfortunate eccezioni - dalle molecole del sé. LE TRUPPE DEL SISTEMA IMMUNITARIO I linfociti B e T, i macrofagi e le cel- lule che espongono gli antigeni sono gli agenti principali della risposta immu- nitaria. I linfociti B sono responsabili dell'immunità umorale. Ciascuno di es- si riconosce un antigene che si trova sui microrganismi patogeni grazie a un recettore specifico per quell'antigene. Così attivato, il linfocita B sintetizza anticorpi specifici per l'antigene i qua- li, una volta liberi, reclutano altri com- battenti che bloccano gli antigeni in cir- colo legandosi a essi. L'efficacia della protezione immuni- taria dipende molto dall'affinità degli anticorpi per gli antigeni (vale a dire la forza del legame tra antigene e anti- corpo quando il sito di interazione è unico). Dipende anche dalla loro «avi- dità»: quando un antigene ha parecchi siti d'interazione con il rispettivo anti- corpo, l'energia combinata del legame è notevolmente superiore alla somma delle energie di legame dei siti presi se- paratamente, e questa avidità è tanto più forte quanto più l'energia combina- ta del legame è alta. Altri linfociti, detti linfociti T, assi- curano l'immunità cellulare. Tra di es- si, vi sono i linfociti T coadiuvanti (hel- per), che interagiscono con i linfociti B, aiutandoli a moltiplicarsi e a produr- re anticorpi, e i linfociti T citotossici, che distruggono le cellule dell'ospite infettate dal microrganismo patogeno. Le cellule natural killer (NK) e i ma- crofagi svolgono funzioni che sono pa- ragonabili tra loro, ma complementari. Infine, i linfociti, i macrofagi e altre cellule coadiuvanti (i polinucleati baso- fili, i mastociti, le piastrine e via dicen- do) liberano molecole, definite media- tori solubili dell'immunità. Esse atti- rano le cellule aventi funzioni di dife- sa sul sito dell'infezione, fanno aumen- tare l'irrorazione sanguigna nel sito stesso e distruggono i microrganismi. Anche le citochine (linfochine, inter- feroni, interleuchine e altri fattori, co- me il fattore di necrosi tumorale) svol- gono parecchie funzioni, e in particola- re attivano la proliferazione delle cellu- le immunitarie. I linfociti T citotossici hanno una no- tevole aggressività: liberano interleu- chine, che generano direttamente o in- direttamente lesioni cellulari, nonché la perforina, una molecola che pratica fori nelle membrane cellulari. Ritorneremo su queste lesioni a livello dei tessuti, che sono in grado di generare le malat- tie autoimmuni. I linfociti T riconoscono i peptidi an- tigenici (frammenti di antigeni) soltan- to quando questi sono presentati, alla superficie delle cellule che espongono antigeni, dalle molecole del maggior complesso di istocompatibilità. I pepti- di che provengono dall'interno delle cellule (ivi compresi i frammenti degli antigeni virali che infettano le cellule) sono presentati dalle molecole di classe I del maggior complesso di istocompa- tibilità. Essi sono riconosciuti specifi- camente dai linfociti T CD8+ citotossici, che distruggono le cellule quando que- ste sono infettate. Le molecole di classe II del maggior complesso di istocompatibilità presen- tano, invece, peptidi derivati dalle pro- teine fagocitate dalle cellule. I pepti- di così esposti vengono riconosciuti da altri difensori, i linfociti T CR+ coa- diuvanti. Pertanto i linfociti T, elemen- ti essenziali nella sorveglianza dell'in- tegrità cellulare, riconoscono unica- mente i peptidi che sono associati alle molecole del maggior complesso di istocompatibilità. Ogni linfocita T o B riconosce in ma- niera specifica un solo antigene, o un solo peptide antigenico, che gli venga correttamente presentato, per cui, nel suo insieme, il sistema immunitario ri- conosce miliardi di antigeni. Quando un antigene è riconosciuto dai linfociti che sono specifici nei suoi confronti, ne scatena la proliferazione: in pochi gior- ni vengono prodotti «cicloni» di linfo- citi T o B specifici per quell'antigene. Nel corso dell'esistenza, l'organismo sintetizza un vasto repertorio di anti- corpi, che basta generalmente per scon- figgere l'ampia varietà degli agenti in- fettivi incontrati nel corso della vita. In circa il 90 per cento dei casi, il sistema immunitario ha la meglio. li CONSEGUIMENTO DELLA TOLLERANZA Le cellule dell'ospite, però, portano tutte in superficie un gran numero di peptidi antigenici. Come fa l'organi- smo a distinguere i propri antigeni, che tollera, dagli antigeni degli agenti infet- tivi, che combatte? La tolleranza dei linfociti T verso gli autoantigeni (molecole del sé) deriva innanzitutto da un meccanismo di sele- zione negativa, che ha luogo nel timo, l'organo in cui si differenziano i lin- fociti. I timociti, precursori dei linfociti T che riconoscono gli autoantigeni e- spressi in quantità sufficiente nel timo, vengono eliminati nel corso del loro differenziamento. Così, soltanto i lin- fociti T che ignorano gli autoantigeni presentati a essi nel timo possono ab- bandonare quest'organo. La tolleranza verso le molecole del sé non viene però conquistata del tutto anche dopo questa selezione negativa, dato che non tutti gli autoantigeni passano per il timo. Per proteggersi dagli autoantigeni che non vengono presentati nel timo, l'organismo dispone di altri meccani- smi di sicurezza. In primo luogo, gli autoantigeni vengono individuati dai linfociti T soltanto se sono presenti in quantità sufficiente; alcuni, poi, non sono riconosciuti perché non vengono presentati dalle molecole del maggior complesso di istocompatibilità; inoltre gli autoantigeni sono ignorati dal siste- ma immunitario fino a quando riman- gono al di fuori degli organi linfoidi; e 36 LE SCIENZE n. 350, ottobre 1997 LE SCIENZE n. 350, ottobre 1997 37
Transcript of Virus e malattie auto immuni -...
Virus e malattieauto immuni
Quando i patti di non aggressione tra sistema immunitario eorganismo si infrangono, compaiono le malattie autoimmuni: si sta
ora precisando il ruolo che certi virus hanno in questo fenomeno
di Rolf M. Zinkernagel
Anticorpi responsabili di malattie autoimmunisono qui evidenziati con diversi coloranti. Cer-tuni producono malattie della pelle, come l'e-pidermolisi bollosa acquisita (a), caratterizzatada eruzioni di vescicole piene di liquido, la cuicicatrizzazione è talvolta difficile. Questi au-toanticorpi sono diretti contro il collagene. An-che in b gli autoanticorpi sono diretti contro ilcoliagene: sono localizzati lungo la membrana
; dei capillari che solcano il glomerulo renale, ilquale assicura la depurazione del sangue. Essiscatenano la sindrome di Goodpasture, che simanifesta soprattutto come insufficienza rena-le fatale. In e gli autoanticorpi che scatenano latiroidite di Hashimoto (distruzione autoimmu-ne della tiroide) sono stati colorati in bruno.Gli autoanticorpi sono presenti sulla superficiedei linfociti B autoreattivi nel follicolo centrale.
t""-.‘ ‘'‘
,ervi\ i MEMBRANA BASALE
a
EPIDERMIDE
()
mero, descrivendo le peripezieguerresche dei greci, mette inscena molti personaggi - solda-
ti, politici, capipopolo, divinità buone ecattive - e numerosi stratagemmi mili-tari, di cui il più celebre è il cavallo diTroia. La lotta contro le malattie che ilnostro organismo ingaggia attraverso ilsistema immunitario, le strategie e leastuzie dei combattenti ricordano que-sta vivace mitologia.
Il sistema immunitario ha il compitodi difendere l'organismo contro i mi-crorganismi patogeni estranei, in parti-colare virus e batteri, che riconosce inquanto essi portano sulla loro superfi-cie molecole specifiche, gli antigeni.Sono gli antigeni che consentono al si-stema immunitario di distinguere lecellule estranee da quelle appartenentiall'organismo (le molecole del sé), chenon devono essere attaccate, pena la di-struzione dei tessuti stessi.
Per evitare questo tipo di reazioni,rare ma talvolta funeste, il sistema im-munitario deve essere specifico, devecioè conservare il ricordo degli agentipatogeni precedentemente incontrati edeve tollerare le molecole del sé.
Per assicurare la propria sopravvi-venza, gli agenti patogeni evitano diuccidere troppo in fretta il loro ospite.Così, la pace armata che regna tra siste-ma immunitario umano e microrgani-smi garantisce sia la sopravvivenza del-l'organismo ospite sia quella dei pato-geni invasori. Per capire come si stabi-lisce un simile modus vivendi esamine-remo in questo articolo i meccanismi didifesa attuati da un organismo nel corsodi un'infezione virale, descriveremo iresponsabili di questa difesa e affronte-remo le nuove ipotesi che fanno lucesui meccanismi dell'autoimmunità.
Dimostreremo come non si debbapensare che le molecole del sé e del
non sé portino «etichette» tali da per-mettere al sistema immunitario di iden-tificarle. Secondo noi, esso riconoscesoltanto le molecole che gli vengonopresentate in organi specifici: gli organilinfoidi (midollo osseo, milza e ganglilinfatici), in quantità sufficiente e perun periodo dai cinque ai 10 giorni.Queste condizioni sono soddisfatte dal-le molecole estranee, per esempio lemolecole dei virus, mentre non lo so-no - tranne sfortunate eccezioni - dallemolecole del sé.
LE TRUPPE DEL SISTEMAIMMUNITARIO
I linfociti B e T, i macrofagi e le cel-lule che espongono gli antigeni sono gliagenti principali della risposta immu-nitaria. I linfociti B sono responsabilidell'immunità umorale. Ciascuno di es-si riconosce un antigene che si trovasui microrganismi patogeni grazie a unrecettore specifico per quell'antigene.Così attivato, il linfocita B sintetizzaanticorpi specifici per l'antigene i qua-li, una volta liberi, reclutano altri com-battenti che bloccano gli antigeni in cir-colo legandosi a essi.
L'efficacia della protezione immuni-taria dipende molto dall'affinità deglianticorpi per gli antigeni (vale a direla forza del legame tra antigene e anti-corpo quando il sito di interazione èunico). Dipende anche dalla loro «avi-dità»: quando un antigene ha parecchisiti d'interazione con il rispettivo anti-corpo, l'energia combinata del legameè notevolmente superiore alla sommadelle energie di legame dei siti presi se-paratamente, e questa avidità è tantopiù forte quanto più l'energia combina-ta del legame è alta.
Altri linfociti, detti linfociti T, assi-curano l'immunità cellulare. Tra di es-
si, vi sono i linfociti T coadiuvanti (hel-per), che interagiscono con i linfociti B,aiutandoli a moltiplicarsi e a produr-re anticorpi, e i linfociti T citotossici,che distruggono le cellule dell'ospiteinfettate dal microrganismo patogeno.Le cellule natural killer (NK) e i ma-crofagi svolgono funzioni che sono pa-ragonabili tra loro, ma complementari.
Infine, i linfociti, i macrofagi e altrecellule coadiuvanti (i polinucleati baso-fili, i mastociti, le piastrine e via dicen-do) liberano molecole, definite media-tori solubili dell'immunità. Esse atti-rano le cellule aventi funzioni di dife-sa sul sito dell'infezione, fanno aumen-tare l'irrorazione sanguigna nel sitostesso e distruggono i microrganismi.Anche le citochine (linfochine, inter-feroni, interleuchine e altri fattori, co-me il fattore di necrosi tumorale) svol-gono parecchie funzioni, e in particola-re attivano la proliferazione delle cellu-le immunitarie.
I linfociti T citotossici hanno una no-tevole aggressività: liberano interleu-chine, che generano direttamente o in-direttamente lesioni cellulari, nonché laperforina, una molecola che pratica forinelle membrane cellulari. Ritorneremosu queste lesioni a livello dei tessuti,che sono in grado di generare le malat-tie autoimmuni.
I linfociti T riconoscono i peptidi an-tigenici (frammenti di antigeni) soltan-to quando questi sono presentati, allasuperficie delle cellule che espongonoantigeni, dalle molecole del maggiorcomplesso di istocompatibilità. I pepti-di che provengono dall'interno dellecellule (ivi compresi i frammenti degliantigeni virali che infettano le cellule)sono presentati dalle molecole di classeI del maggior complesso di istocompa-tibilità. Essi sono riconosciuti specifi-camente dai linfociti T CD8+ citotossici,
che distruggono le cellule quando que-ste sono infettate.
Le molecole di classe II del maggiorcomplesso di istocompatibilità presen-tano, invece, peptidi derivati dalle pro-teine fagocitate dalle cellule. I pepti-di così esposti vengono riconosciuti daaltri difensori, i linfociti T CR+ coa-diuvanti. Pertanto i linfociti T, elemen-ti essenziali nella sorveglianza dell'in-tegrità cellulare, riconoscono unica-mente i peptidi che sono associati allemolecole del maggior complesso diistocompatibilità.
Ogni linfocita T o B riconosce in ma-niera specifica un solo antigene, o unsolo peptide antigenico, che gli vengacorrettamente presentato, per cui, nelsuo insieme, il sistema immunitario ri-conosce miliardi di antigeni. Quandoun antigene è riconosciuto dai linfocitiche sono specifici nei suoi confronti, nescatena la proliferazione: in pochi gior-ni vengono prodotti «cicloni» di linfo-
citi T o B specifici per quell'antigene.Nel corso dell'esistenza, l'organismosintetizza un vasto repertorio di anti-corpi, che basta generalmente per scon-figgere l'ampia varietà degli agenti in-fettivi incontrati nel corso della vita. Incirca il 90 per cento dei casi, il sistemaimmunitario ha la meglio.
li CONSEGUIMENTODELLA TOLLERANZA
Le cellule dell'ospite, però, portanotutte in superficie un gran numero dipeptidi antigenici. Come fa l'organi-smo a distinguere i propri antigeni, chetollera, dagli antigeni degli agenti infet-tivi, che combatte?
La tolleranza dei linfociti T verso gliautoantigeni (molecole del sé) derivainnanzitutto da un meccanismo di sele-zione negativa, che ha luogo nel timo,l'organo in cui si differenziano i lin-fociti. I timociti, precursori dei linfociti
T che riconoscono gli autoantigeni e-spressi in quantità sufficiente nel timo,vengono eliminati nel corso del lorodifferenziamento. Così, soltanto i lin-fociti T che ignorano gli autoantigenipresentati a essi nel timo possono ab-bandonare quest'organo. La tolleranzaverso le molecole del sé non viene peròconquistata del tutto anche dopo questaselezione negativa, dato che non tuttigli autoantigeni passano per il timo.
Per proteggersi dagli autoantigeniche non vengono presentati nel timo,l'organismo dispone di altri meccani-smi di sicurezza. In primo luogo, gliautoantigeni vengono individuati dailinfociti T soltanto se sono presenti inquantità sufficiente; alcuni, poi, nonsono riconosciuti perché non vengonopresentati dalle molecole del maggiorcomplesso di istocompatibilità; inoltregli autoantigeni sono ignorati dal siste-ma immunitario fino a quando riman-gono al di fuori degli organi linfoidi; e
36 LE SCIENZE n. 350, ottobre 1997 LE SCIENZE n. 350, ottobre 1997 37
ISOLA DI LANGERHANS
CELLULA BETAIN VIA DI DISTRUZIONE
infine, certi autoantigeni sono protettiin sedi immunologicamente privilegiatecome, per esempio, cervello, testicoli,ovaie e camera anteriore dell'occhio.
L'anergia è un altro meccanismo cheincrementa la tolleranza reciproca tragli autoantigeni isolati alla periferia e ilinfociti T specifici nei loro riguardi.Sono necessari due segnali perché ilinfociti T siano attivati: il segnale del-l'antigene e un altro segnale, per esem-pio la citochina o altre molecole defi-nite accessorie. Soltanto gli antigeni as-sociati a un secondo segnale, presenta-ti correttamente dalle cellule che li e-spongono, attivano gli specifici linfoci-ti T. In caso contrario, i linfociti T ri-mangono anergici, cioè silenti.
Nei cosiddetti «santuari» immunolo-gici non solo gli autoantigeni, ma an-che gli antigeni virali possono trovareprotezione. Diamo due esempi: l'infe-zione da parte del virus della rabbia,che colpisce gli assoni dei neuroni (lamalattia è rara, ma generalmente mor-tale) e l'infezione da papillomavirus,che colpisce alcune cellule della cute, icheratinociti (è un'infezione frequente,ma benigna). Quando queste infezionisi instaurano, i linfociti T non reagisco-no perché i virus sono isolati in sediprivilegiate, dove neppure gli antige-ni virali attivano il sistema immuni-tario. Per contro, quando i neuroni o icheratinociti infettati muoiono, liberanoantigeni virali che vengono fagocitatidai macrofagi. A questo punto gli an-tigeni, presi in carico dalle cellule cheli espongono, sono presentati al siste-ma immunitario, il quale attiva gli spe-cifici linfociti T che attaccano e di-struggono i neuroni o i cheratinociti.
Quindi, o il sistema immunitario eli-mina il virus dopo un episodio di infe-zione acuta, o il virus sfugge al sistema
immunitario: si instaura così un'infe-zione cronica, nel corso della quale so-pravvivono cellule infettate che libera-no saltuariamente particelle virali. Puòdarsi anche che i due scenari si alterni-no, con episodi di infezione acuta e diinfezione cronica.
INFEZIONE ACUTA,INFEZIONE CRONICA
I virus citopatogeni distruggono lecellule che infettano ma, quando le rea-zioni immunitarie sono efficaci, la di-struzione è limitata e le reazioni cessa-no non appena si interrompe la produ-zione di antigeni virali. L'infezione èstroncata e il tessuto colpito è solo lie-vemente danneggiato. Gli autoantigeni,ignorati fino a questo momento dailinfociti T, vengono liberati dalle cellu-le distrutte, ma la loro liberazione è dibreve durata e in quantità insufficientead attivare i linfociti B e T potenzial-mente autoreattivi.
Nel caso dei virus non citopatogeni, ilinfociti T CD8+ citotossici distruggo-no le cellule infettate prima che esse li-berino nuove particelle virali; quandoperò questa distruzione si prolunga, ri-schia di scatenare meccanismi immu-nopatologici o autoimmuni. Se i linfo-citi riescono a distruggere le cellule in-fettate più rapidamente di quanto il vi-rus si moltiplichi, questo finisce per es-sere eliminato. In caso contrario, il vi-rus persiste e l'infezione diventa croni-ca; la liberazione prolungata di antigenivirali attiva i linfociti B, che sintetizza-no anticorpi antivirali, e i linfociti Thelper, i quali producono sostanze chedanneggiano le cellule infettate e tal-volta anche quelle sane. Si instaura, al-lora, una malattia immunopatologica,nel corso della quale i tessuti vengono
lesi dalle sostanze liberate dallo stessosistema immunitario.
Nel corso di un'infezione cronicavengono liberate notevoli quantità diautoantigeni, i quali - se raggiungono itessuti linfoidi, vi restano per un temposufficientemente lungo e vengono cor-rettamente presentati - rischiano di atti-vare linfociti specifici che in preceden-za li ignoravano. Vengono così stimo-lati linfociti T e B autoreattivi, con laconseguente massiccia produzione diautoanticorpi e la moltiplicazione dilinfociti T autoreattivi, che mantengonoin vita il meccanismo autoimmune.
Ogni volta che si ignora il virus re-sponsabile dei meccanismi immunopa-tologici, la malattia è considerata au-toimmune, ma io ritengo che alcunemalattie qualificate come autoimmunisiano, in realtà, direttamente causate dareazioni immunitarie contro un agenteinfettivo sconosciuto.
Negli anni settanta, alcuni immuno-logi avevano ipotizzato che certi anti-geni virali siano talmente simili ad al-cuni autoantigeni da far sì che il siste-ma immunitario scateni contro di essila produzione di autoanticorpi: graziea questo meccanismo di mimetismomolecolare, i linfociti attivati contro il
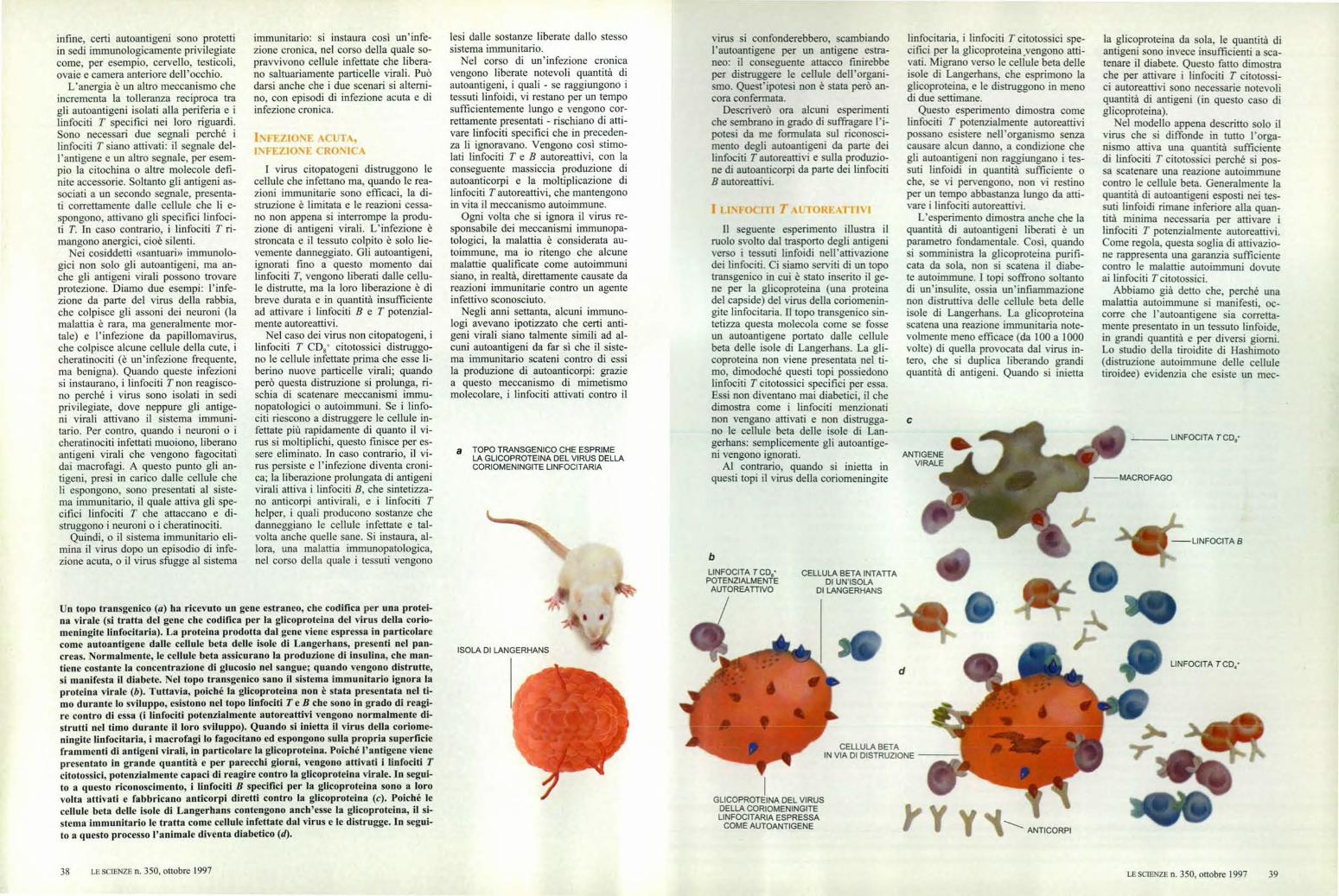
a TOPO TRANSGENICO CHE ESPRIMELA GLICOPROTEINA DEL VIRUS DELLACORIOMENINGITE LINFOCITARIA
virus si confonderebbero, scambiandol'autoantigene per un antigene estra-neo: il conseguente attacco finirebbeper distruggere le cellule dell'organi-smo. Quest'ipotesi non è stata però an-cora confermata.
Descriverò ora alcuni esperimentiche sembrano in grado di suffragare l'i-potesi da me formulata sul riconosci-mento degli autoantigeni da parte deilinfociti T autoreattivi e sulla produzio-ne di autoanticorpi da parte dei linfocitiB autoreattivi.
I LINFOCITI T AUTOREATTIVI
Il seguente esperimento illustra ilruolo svolto dal trasporto degli antigeniverso i tessuti linfoidi nell'attivazionedei linfociti. Ci siamo serviti di un topotransgenico in cui è stato inserito il ge-ne per la glicoproteina (una proteinadel capside) del virus della coriomenin-gite linfocitaria. Il topo transgenico sin-tetizza questa molecola come se fosseun autoantigene portato dalle cellulebeta delle isole di Langerhans. La gli-coproteina non viene presentata nel ti-mo, dimodoché questi topi possiedonolinfociti T citotossici specifici per essa.Essi non diventano mai diabetici, il chedimostra come i linfociti menzionatinon vengano attivati e non distrugga-no le cellule beta delle isole di Lan-gerhans: semplicemente gli autoantige-ni vengono ignorati.
Al contrario, quando si inietta inquesti topi il virus della coriomeningite
LINFOCITA T CID,:POTENZIALMENTE
AUTOREATTIVO
se
GLICOPROTEINA DEL VIRUSDELLA CORIOMENINGITELINFOCITARIA ESPRESSA
COME AUTOANTIGENE
linfocitaria, i linfociti T citotossici spe-cifici per la glicoproteina yengono atti-vati. Migrano verso le cellule beta delleisole di Langerhans, che esprimono laglicoproteina, e le distruggono in menodi due settimane.
Questo esperimento dimostra comelinfociti T potenzialmente autoreattivipossano esistere nell'organismo senzacausare alcun danno, a condizione chegli autoantigeni non raggiungano i tes-suti linfoidi in quantità sufficiente oche, se vi pervengono, non vi restinoper un tempo abbastanza lungo da atti-vare i linfociti autoreattivi.
L'esperimento dimostra anche che laquantità di autoantigeni liberati è unparametro fondamentale. Così, quandosi somministra la glicoproteina purifi-cala da sola, non si scatena il diabe-te autoimmune. I topi soffrono soltantodi un'insulite, ossia un'infiammazionenon distruttiva delle cellule beta delleisole di Langerhans. La glicoproteinascatena una reazione immunitaria note-volmente meno efficace (da 100 a 1000volte) di quella provocata dal virus in-tero, che si duplica liberando grandiquantità di antigeni. Quando si inietta
la glicoproteina da sola, le quantità diantigeni sono invece insufficienti a sca-tenare il diabete. Questo fatto dimostrache per attivare i linfociti T citotossi-ci autoreattivi sono necessarie notevoliquantità di antigeni (in questo caso diglicoproteina).
Nel modello appena descritto solo ilvirus che si diffonde in tutto l'orga-nismo attiva una quantità sufficientedi linfociti T citotossici perché si pos-sa scatenare una reazione autoimmunecontro le cellule beta. Generalmente laquantità di autoantigeni esposti nei tes-suti linfoidi rimane inferiore alla quan-tità minima necessaria per attivare ilinfociti T potenzialmente autoreattivi.Come regola, questa soglia di attivazio-ne rappresenta una garanzia sufficientecontro le malattie autoimmuni dovuteai linfociti T citotossici.
Abbiamo già detto che, perché unamalattia autoimmune si manifesti, oc-corre che l'autoantigene sia corretta-mente presentato in un tessuto linfoide,in grandi quantità e per diversi giorni.Lo studio della tiroidite di Hashimoto(distruzione autoimmune delle celluletiroidee) evidenzia che esiste un mec-
LINFOCITA T CID,'
MACROFAGO
-LINFOCITA
Un topo transgenico (a) ha ricevuto un gene estraneo, che codifica per una protei-na v irale (si tratta del gene che codifica per la glicoproteina del virus della conio-meningite linfocitaria). La proteina prodotta dal gene viene espressa in particolarecome autoantigene dalle cellule beta delle isole di Langerhans, presenti nel pan-creas. Normalmente, le cellule beta assicurano la produzione di insulina, che man-tiene costante la concentrazione di glucosio nel sangue; quando vengono distrutte,si manifesta il diabete. Nel topo transgenico sano il sistema immunitario ignora laproteina virale (b). Tuttavia, poiché la glicoproteina non è stata presentata nel ti-mo durante lo sviluppo, esistono nel topo linfociti T e B che sono in grado di reagi-
re contro di essa (i linfociti potenzialmente autoreattivi vengono normalmente di-strutti nel timo durante il loro sviluppo). Quando si inietta il virus della coriome-ningite linfocitaria, i macrofagi lo fagocitano ed espongono sulla propria superficieframmenti di antigeni virali, in particolare la glicoproteina. Poiché l'antigene vienepresentato in grande quantità e per parecchi giorni, vengono attivati i linfociti Tcitotossici, potenzialmente capaci di reagire contro la glicoproteina virale. In segui-
to a questo riconoscimento, i linfociti B specifici per la glicoproteina sono a lorovolta attivati e fabbricano anticorpi diretti contro la glicoproteina (e). Poiché lecellule beta delle isole di Langerhans contengono anch'esse la glicoproteina, il si-stema immunitario le tratta come cellule infettate dal virus e le distrugge. In segui-to a questo processo l'animale diventa diabetico (d).
CELLULA BETA INTATTADI UN'ISOLA
DI LANGERHANS
ANTIGENEVIRALE
r y rA ANTICORPIMi0
• LINFOCITA T CD4'
7",e4
38 LE SCIENZE n. 350, ottobre 1997 LE SCIENZE n. 350, ottobre 1997 39
• #4 LINFOCITA T
AUTOANTIGENE
VASILINFATICI
ATTIVATOLINFOCITA B
TIMO CELLULA CHE ESPONE-L'AUTOANTIGENE -
LINFOCITA T AUTOREATTIVO DISTRUTTO
GANGLIO LINFOCITA T AUTOREATTIVOLINFATICO
MIDOLLOOSSEO
GANGLIOMESENTERICO
4\4101).„,i,MILZA
PLACCADI PEYER
NUMEROSIAUTOANTIGENI41.4141 •4. 4.
LINFOCITA T AUTOREATTIVO
ESPOSIZIONEPROLUNGATA
SISTEMA LINFOIDESECONDARIO
MOLTIPLICAZIONEDEI LINFOCITI TAUTOREATTIVI
ORGANO BERSAGLIO
Cà
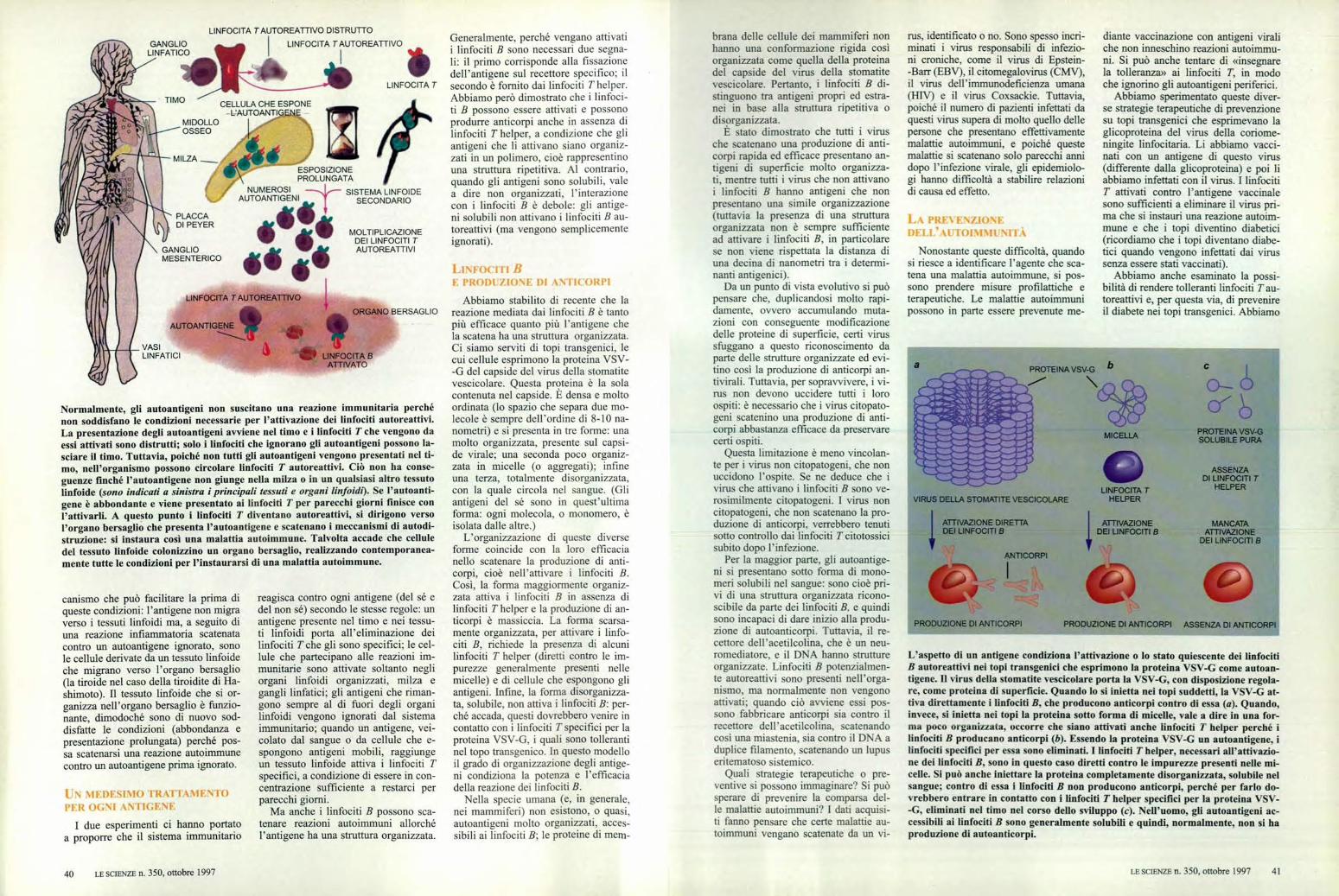
Normalmente, gli autoantigeni non suscitano una reazione immunitaria perchénon soddisfano le condizioni necessarie per l'attivazione dei linfociti autoreattivi.La presentazione degli autoantigeni avviene nel timo e i linfociti T che vengono daessi attivati sono distrutti; solo i linfociti che ignorano gli autoantigeni possono la-sciare il timo. Tuttavia, poiché non tutti gli autoantigeni vengono presentati nel ti-mo, nell'organismo possono circolare linfociti T autoreattivi. Ciò non ha conse-guenze finché l'autoantigene non giunge nella milza o in un qualsiasi altro tessutolinfoide (sono indicati a sinistra i principali tessuti e organi linfoidi). Se l'autoanti-gene è abbondante e viene presentato ai linfociti T per parecchi giorni finisce conl'attivarli. A questo punto i linfociti T diventano autoreattivi, si dirigono versol'organo bersaglio che presenta l'autoantigene e scatenano i meccanismi di autodi-struzione: si instaura così una malattia autoimmune. Talvolta accade che celluledel tessuto linfoide colonizzino un organo bersaglio, realizzando contemporanea-mente tutte le condizioni per l'instaurarsi di una malattia autoimmune.
ANTICORPI
PRODUZIONE DI ANTICORPI PRODUZIONE DI ANTICORPI ASSENZA DI ANTICORPI
c
O-- O
PROTEINA VSV-GSOLUBILE PURA
ASSENZADI LINFOCITI T
HELPER
MANCATAATTIVAZIONE
DEI LINFOCITI B
VIRUS DELLA STOMATITE VESCICOLARE
ATTIVAZIONE DIRETTADEI LINFOCITI
LINFOCITA THELPER
Ir ATTIVAZIONEDEI LINFOCITI B
a PROTEINA VSV-G
MICELLA
L'aspetto di un antigene condiziona l'attivazione o lo stato quiescente dei linfocitiB autoreattivi nei topi transgenici che esprimono la proteina VSV-G come autoan-tigene. Il virus della stomatite vescicolare porta la VSV-G, con disposizione regola-re, come proteina di superficie. Quando lo si inietta nei topi suddetti, la VSV-G at-tiva direttamente i linfociti B, che producono anticorpi contro di essa (a). Quando,invece, si inietta nei topi la proteina sotto forma di micelle, vale a dire in una for-ma poco organizzata, occorre che siano attivati anche linfociti T helper perché ilinfociti B producano anticorpi (b). Essendo la proteina VSV-G un autoantigene, ilinfociti specifici per essa sono eliminati. I linfociti T helper, necessari all'attivazio-ne dei linfociti B. sono in questo caso diretti contro le impurezze presenti nelle mi-celle. Si può anche iniettare la proteina completamente disorganizzata, solubile nelsangue; contro di essa i linfociti B non producono anticorpi, perché per farlo do-vrebbero entrare in contatto con i linfociti T helper specifici per la proteina VSV--G, eliminati nel timo nel corso dello sviluppo (c). Nell'uomo, gli autoantigeni ac-cessibili ai linfociti B sono generalmente solubili e quindi, normalmente, non si haproduzione di autoanticorpi.
canismo che può facilitare la prima diqueste condizioni: l'antigene non migraverso i tessuti linfoidi ma, a seguito diuna reazione infiammatoria scatenatacontro un autoantigene ignorato, sonole cellule derivate da un tessuto linfoideche migrano verso l'organo bersaglio(la tiroide nel caso della tiroidite di Ha-shimoto). Il tessuto linfoide che si or-ganizza nell'organo bersaglio è funzio-nante, dimodoché sono di nuovo sod-disfatte le condizioni (abbondanza epresentazione prolungata) perché pos-sa scatenarsi una reazione autoimmunecontro un autoantigene prima ignorato.
UN MEDESIMO TRATTAMENTOPER OGNI ANTIGENE
I due esperimenti ci hanno portatoa proporre che il sistema immunitario
reagisca contro ogni antigene (del sé edel non sé) secondo le stesse regole: unantigene presente nel timo e nei tessu-ti linfoidi porta all'eliminazione deilinfociti T che gli sono specifici; le cel-lule che partecipano alle reazioni im-munitarie sono attivate soltanto negliorgani linfoidi organizzati, milza egangli linfatici; gli antigeni che riman-gono sempre al di fuori degli organilinfoidi vengono ignorati dal sistemaimmunitario; quando un antigene, vei-colato dal sangue o da cellule che e-spongono antigeni mobili, raggiungeun tessuto linfoide attiva i linfociti Tspecifici, a condizione di essere in con-centrazione sufficiente a restarci perparecchi giorni.
Ma anche i linfociti B possono sca-tenare reazioni autoimmuni allorchél'antigene ha una struttura organizzata.
Generalmente, perché vengano attivatii linfociti B sono necessari due segna-li: il primo corrisponde alla fissazionedell'antigene sul recettore specifico; ilsecondo è fornito dai linfociti T helper.Abbiamo però dimostrato che i linfoci-ti B possono essere attivati e possonoprodurre anticorpi anche in assenza dilinfociti T helper, a condizione che gliantigeni che li attivano siano organiz-zati in un polimero, cioè rappresentinouna struttura ripetitiva. Al contrario,quando gli antigeni sono solubili, valea dire non organizzati, l'interazionecon i linfociti B è debole: gli antige-ni solubili non attivano i linfociti B au-toreattivi (ma vengono semplicementeignorati).
LINFOCITI BE PRODUZIONE DI ANTICORPI
Abbiamo stabilito di recente che lareazione mediata dai linfociti B è tantopiù efficace quanto più l'antigene chela scatena ha una struttura organizzata.Ci siamo serviti di topi transgenici, lecui cellule esprimono la proteina VSV--G del capside del virus della stomatitevescicolare. Questa proteina è la solacontenuta nel capside. È densa e moltoordinata (lo spazio che separa due mo-lecole è sempre dell'ordine di 8-10 na-nometri) e si presenta in tre forme: unamolto organizzata, presente sul capsi-de virale; una seconda poco organiz-zata in micelle (o aggregati); infineuna terza, totalmente disorganizzata,con la quale circola nel sangue. (Gliantigeni del sé sono in quest'ultimaforma: ogni molecola, o monomero, èisolata dalle altre.)
L'organizzazione di queste diverseforme coincide con la loro efficacianello scatenare la produzione di anti-corpi, cioè nell'attivare i linfociti B.Così, la forma maggiormente organiz-zata attiva i linfociti B in assenza dilinfociti T helper e la produzione di an-ticorpi è massiccia. La forma scarsa-mente organizzata, per attivare i linfo-citi B, richiede la presenza di alcunilinfociti T helper (diretti contro le im-purezze generalmente presenti nellemicelle) e di cellule che espongono gliantigeni. Infine, la forma disorganizza-ta, solubile, non attiva i linfociti B: per-ché accada, questi dovrebbero venire incontatto con i linfociti T specifici per laproteina VSV-G, i quali sono tollerantinel topo transgenico. In questo modelloil grado di organizzazione degli antige-ni condiziona la potenza e l'efficaciadella reazione dei linfociti B.
Nella specie umana (e, in generale,nei mammiferi) non esistono, o quasi,autoantigeni molto organizzati, acces-sibili ai linfociti B; le proteine di mem-
brana delle cellule dei mammiferi nonhanno una conformazione rigida cosìorganizzata come quella della proteinadel capside del virus della stomatitevescicolare. Pertanto, i linfociti B di-stinguono tra antigeni propri ed estra-nei in base alla struttura ripetitiva odisorganizzata.
È stato dimostrato che tutti i virusche scatenano una produzione di ami-corpi rapida ed efficace presentano an-tigeni di superficie molto organizza-ti, mentre tutti i virus che non attivanoi linfociti B hanno antigeni che nonpresentano una simile organizzazione(tuttavia la presenza di una strutturaorganizzata non è sempre sufficientead attivare i linfociti B, in particolarese non viene rispettata la distanza diuna decina di nanometri tra i determi-nanti antigenici).
Da un punto di vista evolutivo si puòpensare che, duplicandosi molto rapi-damente, ovvero accumulando muta-zioni con conseguente modificazionedelle proteine di superficie, certi virussfuggano a questo riconoscimento daparte delle strutture organizzate ed evi-tino così la produzione di anticorpi an-tivirali. Tuttavia, per sopravvivere, i vi-rus non devono uccidere tutti i loroospiti: è necessario che i virus citopato-geni scatenino una produzione di anti-corpi abbastanza efficace da preservarecerti ospiti.
Questa limitazione è meno vincolan-te per i virus non citopatogeni, che nonuccidono l'ospite. Se ne deduce che ivirus che attivano i linfociti B sono ve-rosimilmente citopatogeni. I virus noncitopatogeni, che non scatenano la pro-duzione di anticorpi, verrebbero tenutisotto controllo dai linfociti T citotossicisubito dopo l'infezione.
Per la maggior parte, gli autoantige-ni si presentano sotto forma di mono-meri solubili nel sangue: sono cioè pri-vi di una struttura organizzata ricono-scibile da parte dei linfociti B, e quindisono incapaci di dare inizio alla produ-zione di autoanticorpi. Tuttavia, il re-cettore dell'acetilcolina, che è un neu-romediatore, e il DNA hanno struttureorganizzate. Linfociti B potenzialmen-te autoreattivi sono presenti nell'orga-nismo, ma normalmente non vengonoattivati; quando ciò avviene essi pos-sono fabbricare anticorpi sia contro ilrecettore dell'acetilcolina, scatenandocosì una miastenia, sia contro il DNA aduplice filamento, scatenando un lupuseritematoso sistemico.
Quali strategie terapeutiche o pre-ventive si possono immaginare? Si puòsperare di prevenire la comparsa del-le malattie autoinununi? I dati acquisi-ti fanno pensare che certe malattie au-toimmuni vengano scatenate da un vi-
rus, identificato o no. Sono spesso incri-minati i virus responsabili di infezio-ni croniche, come il virus di Epstein--Barr (EBV), il citomegalovirus (CMV),il virus dell'immunodeficienza umana(FIN) e il virus Coxsackie. Tuttavia,poiché il numero di pazienti infettati daquesti virus supera di molto quello dellepersone che presentano effettivamentemalattie autoimmuni, e poiché questemalattie si scatenano solo parecchi annidopo l'infezione virale, gli epidemiolo-gi hanno difficoltà a stabilire relazionidi causa ed effetto.
LA PREVENZIONEDELL'A UTOIMMUNITÀ
Nonostante queste difficoltà, quandosi riesce a identificare l'agente che sca-tena una malattia autoimmune, si pos-sono prendere misure profilattiche eterapeutiche. Le malattie autoimmunipossono in parte essere prevenute me-
diante vaccinazione con antigeni viraliche non inneschino reazioni autoimmu-ni. Si può anche tentare di «insegnarela tolleranza» ai linfociti T, in modoche ignorino gli autoantigeni periferici.
Abbiamo sperimentato queste diver-se strategie terapeutiche di prevenzionesu topi transgenici che esprimevano laglicoproteina del virus della coriome-ningite linfocitaria. Li abbiamo vacci-nati con un antigene di questo virus(differente dalla glicoproteina) e poi liabbiamo infettati con il virus. I linfocitiT attivati contro l'antigene vaccinalesono sufficienti a eliminare il virus pri-ma che si instauri una reazione autoim-mune e che i topi diventino diabetici(ricordiamo che i topi diventano diabe-tici quando vengono infettati dai virussenza essere stati vaccinati).
Abbiamo anche esaminato la possi-bilità di rendere tolleranti linfociti T au-toreattivi e, per questa via, di prevenireil diabete nei topi transgenici. Abbiamo
40 LE SCIENZE n. 350, ottobre 1997
LE SCIENZE n. 350, ottobre 1997 41
I libri di
LE SCIENZE
QUARK 2000
()
la fisica fondamentale dell'anoe le sfide del nuovo millennio
12 SCII?. 1..1E1,170M
QUARK 2000la fisica fondamentale italianae le sfide del nuovo millennio
in collaborazione conl'Istituto Nazionale di Fisica Nucleare
IL QUADRO CONCETTUALE
La relativitàTullio Regge
La meccanica quantisticaGiancarlo GhirardiQuark e leptoniNicola Cabibbo
MOMENTI STORICI DELLA FISICA ITALIANA
La vicenda del gruppo FermiFabio Sebastiani e Carlo Tarsitani
Rossi a FirenzeAlberto Bonetti
Amaldi e la ricostruzioneGiovanni Battimelli e Michelangelo De Maria
AdA e FrascatiCarlo Bemardini
L'ATTUALITÀ
La violazione della simmetria CPPaolo Franzini
Il neutrino solareGian Paolo Bellini
Simmetrie dinamicheFrancesco lachello
APE: i supercalcolatori paralleli dell'INFNNicola Cabibbo e Raffaele Tripiccione
La scoperta del quark top e il contributo del-l'INFN
Giorgio BellettiniLe onde gravitazionali
Alberto GiazottoIl progetto EUROBALL
Carlo Rossi Alvarez
LA MOSTRA
QUARK 2000La fisica fondamentale italianae le sfide del nuovo millennio
Alessandro Pascolim
160 pagine, L. 25 000
per le ordinazioni utilizzare la cartolinainserita in questo fascicolo
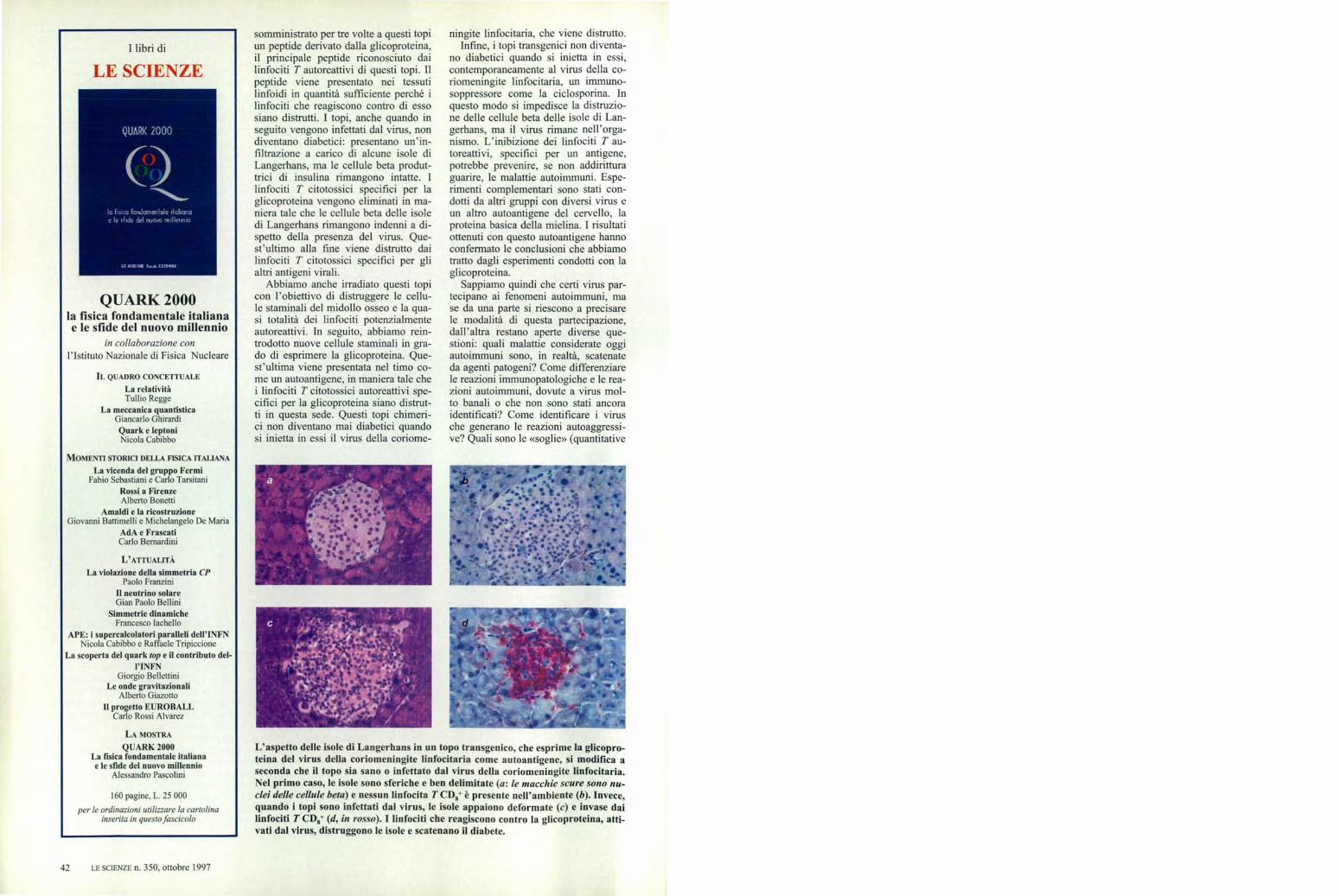
L'aspetto delle isole di Langerhans in un topo transgenico, che esprime la glicopro-teina del virus della coriomeningite linfocitaria come autoantigene, si modifica aseconda che il topo sia sano o infettato dal virus della coriomeningite linfocitaria.Nel primo caso, le isole sono sferiche e ben delimitate (a: le macchie scure sono nu-clei delle cellule bela) e nessun linfocita T CD8+ è presente nell'ambiente (b). Invece,quando i topi sono infettati dal virus, le isole appaiono deformate (c) e invase dailinfociti T OZI,* (d, in rosso). I linfociti che reagiscono contro la glicoproteina, atti-vati dal virus, distruggono le isole e scatenano il diabete.
somministrato per tre volte a questi topiun peptide derivato dalla glicoproteina,il principale peptide riconosciuto dailinfociti T autoreattivi di questi topi. Ilpeptide viene presentato nei tessutilinfoidi in quantità sufficiente perché ilinfociti che reagiscono contro di essosiano distrutti. I topi, anche quando inseguito vengono infettati dal virus, nondiventano diabetici: presentano un'in-filtrazione a carico di alcune isole diLangerhans, ma le cellule beta produt-trici di insulina rimangono intatte. Ilinfociti T citotossici specifici per laglicoproteina vengono eliminati in ma-niera tale che le cellule beta delle isoledi Langerhans rimangono indenni a di-spetto della presenza del virus. Que-st'ultimo alla fine viene distrutto dailinfociti T citotossici specifici per glialtri antigeni virali.
Abbiamo anche irradiato questi topicon l'obiettivo di distruggere le cellu-le staminali del midollo osseo e la qua-si totalità dei linfociti potenzialmenteautoreattivi. In seguito, abbiamo rein-trodotto nuove cellule staminali in gra-do di esprimere la glicoproteina. Que-st'ultima viene presentata nel timo co-me un autoantigene, in maniera tale chei linfociti T citotossici autoreattivi spe-cifici per la glicoproteina siano distrut-ti in questa sede. Questi topi chimeri-ci non diventano mai diabetici quandosi inietta in essi il virus della coriome-
ningite linfocitaria, che viene distrutto.Infine, i topi transgenici non diventa-
no diabetici quando si inietta in essi,contemporaneamente al virus della co-riomeningite linfocitaria, un immuno-soppressore come la ciclosporina. Inquesto modo si impedisce la distruzio-ne delle cellule beta delle isole di Lan-gerhans, ma il virus rimane nell'orga-nismo. L'inibizione dei linfociti T au-toreattivi, specifici per un antigene,potrebbe prevenire, se non addiritturaguarire, le malattie autoimmuni. Espe-rimenti complementari sono stati con-dotti da altri gruppi con diversi virus eun altro autoantigene del cervello, laproteina basica della mielina. I risultatiottenuti con questo autoantigene hannoconfermato le conclusioni che abbiamotratto dagli esperimenti condotti con laglicoproteina.
Sappiamo quindi che certi virus par-tecipano ai fenomeni autoimmuni, mase da una parte si riescono a precisarele modalità di questa partecipazione,dall'altra restano aperte diverse que-stioni: quali malattie considerate oggiautoimmuni sono, in realtà, scatenateda agenti patogeni? Come differenziarele reazioni immunopatologiche e le rea-zioni autoimmuni, dovute a virus mol-to banali o che non sono stati ancoraidentificati? Come identificare i virusche generano le reazioni autoaggressi-ve? Quali sono le «soglie» (quantitative
42 LE SCIENZE n. 350, ottobre 1997
aINIEZIONE
DI ANTIGENI VIRALI INIEZIONEDI GLICOPROTEINA\
GLICOPROTEINAA, /
DEL VIRUSDELLA CORIOMENINGITE
44:é
TRAPIANTO DI NUOVE
è CELLULE STAMINALIà à ANTIGENI VIRALI
LINFOCITA TAUTOREATTIVO DISTRUTTO
PRODUZIONEDI ANTICORPI
E DI LINFOCITI TE B
1111‘ DISTRUZIONE DEL VIRUS
Alle • 301)
•à à
INIEZIONEeIk‘'• DEL VIRUS
\CELLULA BETA
DI LANGERHANS INTATTA
DISTRUZIONE DELLECELLULE STAMINALIDEL MIDOLLO OSSEO
INIEZIONE‘I›. DI CICLOSPORINA
'è 41
àà è
ANTIGENI VIRALII.
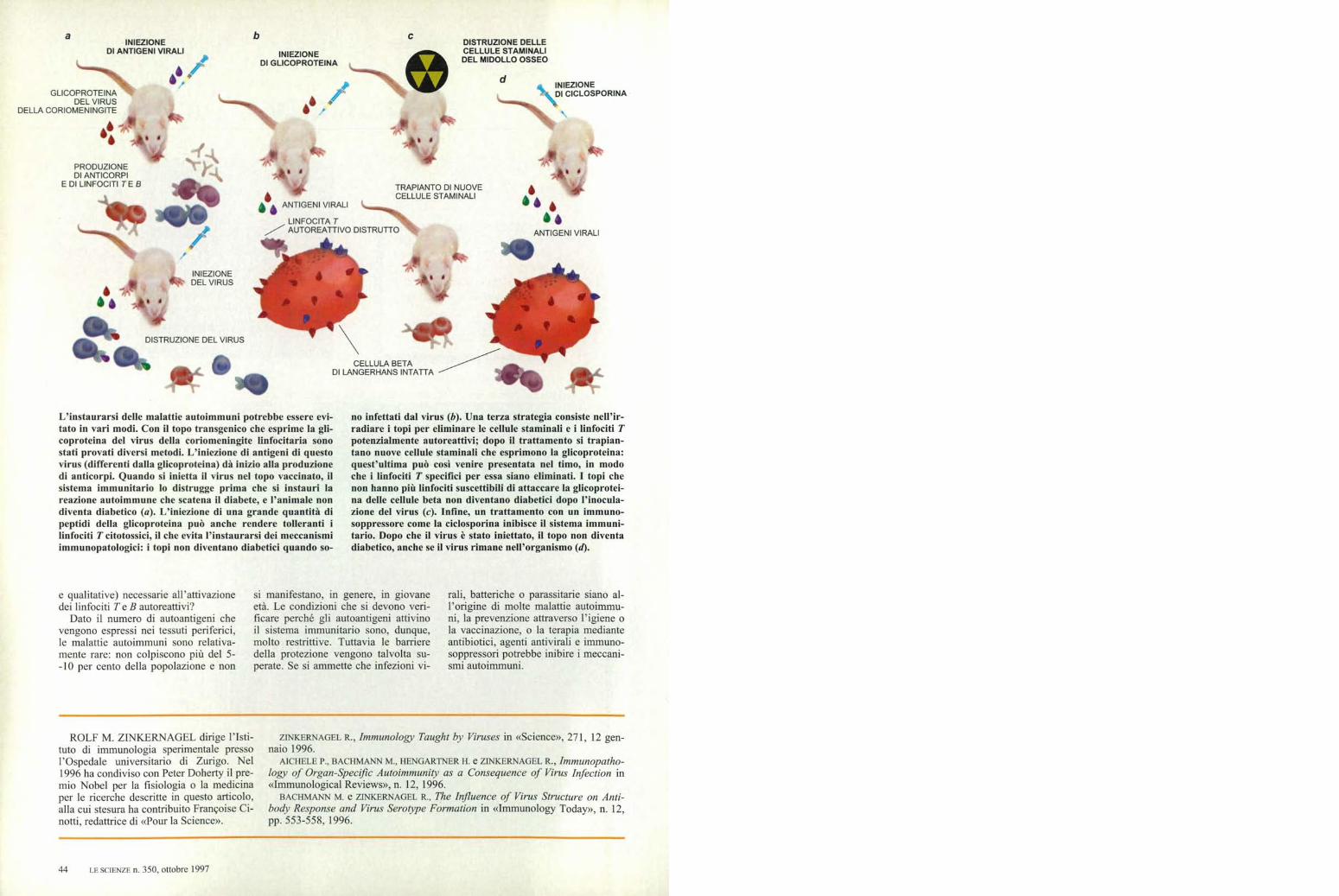
L'instaurarsi delle malattie autoimmuni potrebbe essere evi-tato in vari modi. Con il topo transgenico che esprime la gli-coproteina del virus della coriomeningite linfocitaria sonostati provati diversi metodi. L'iniezione di antigeni di questovirus (differenti dalla glicoproteina) dà inizio alla produzionedi anticorpi. Quando si inietta il virus nel topo vaccinato, ilsistema immunitario lo distrugge prima che si instauri lareazione autoimmune che scatena il diabete, e l'animale nondiventa diabetico (a). L'iniezione di una grande quantità dipeptidi della glicoproteina può anche rendere tolleranti ilinfociti T citotossici, il che evita l'instaurarsi dei meccanismiimmunopatologici: i topi non diventano diabetici quando so-
no infettati dal virus (b). Una terza strategia consiste nell'ir-radiare i topi per eliminare le cellule staminali e i linfociti Tpotenzialmente autoreattivi; dopo il trattamento si trapian-tano nuove cellule staminali che esprimono la glicoproteina:quest'ultima può così venire presentata nel timo, in modoche i linfociti T specifici per essa siano eliminati. I topi chenon hanno più linfociti suscettibili di attaccare la glicoprotei-na delle cellule beta non diventano diabetici dopo l'inocula-zione del virus (e). Infine, un trattamento con un immuno-soppressore come la ciclosporina inibisce il sistema immuni-tario. Dopo che il virus è stato iniettato, il topo non diventadiabetico, anche se il virus rimane nell'organismo (d).
e qualitative) necessarie all'attivazionedei linfociti T e B autoreattivi?
Dato il numero di autoantigeni chevengono espressi nei tessuti periferici,le malattie autoimmuni sono relativa-mente rare: non colpiscono più del 5--10 per cento della popolazione e non
si manifestano, in genere, in giovaneetà. Le condizioni che si devono veri-ficare perché gli autoantigeni attivinoil sistema immunitario sono, dunque,molto restrittive. Tuttavia le barrieredella protezione vengono talvolta su-perate. Se si ammette che infezioni vi-
rali, batteriche o parassitarie siano al-l'origine di molte malattie autoimmu-ni, la prevenzione attraverso l'igiene ola vaccinazione, o la terapia medianteantibiotici, agenti antivirali e immuno-soppressori potrebbe inibire i meccani-smi autoimmuni.
ROLF M. ZINKERNAGEL dirige l'Isti-tuto di immunologia sperimentale pressol'Ospedale universitario di Zurigo. Nel1996 ha condiviso con Peter Doherty il pre-mio Nobel per la fisiologia o la medicinaper le ricerche descritte in questo articolo,alla cui stesura ha contribuito Fran9oise Ci-notti, redattrice di «Pour la Science».
Z1NKERNAGEL R., Immunology Taught by Viruses in «Science», 271, 12 gen-naio 1996.
AICHELE P., BACHMANN M., HENGARTNER H. e ZINKERNAGEL R., Immunopatho-logy of Organ-Specific Autoimmunity as a Consequence of Virus Infection in«Immunological Reviews», n. 12, 1996.
BACHMANN M. e ZINKERNAGEL R., The Influence of Virus Structure on Anti-body Response and Virus Serotype Formation in «Immunology Today», n. 12,pp. 553-558, 1996.
44 LE SCIENZE n. 350, ottobre 1997


















