
Università degli Studi di Padova Dipartimento di Fisica ed ...

1
Università degli studi di Padova
Dipartimento di Biologia
Dipartimento di Fisica e Astronomia
Corso di Laurea triennale in Ottica e Optometria
Tesi di Laurea
Miopia: fattori genetici e ambientali
Relatore: Prof.ssa Stefania Bortoluzzi Laureanda:
Correlatore: Prof.ssa Dominga Ortolan Dei Zotti Michela
Matr.:
1005342-OPT
A.A. 2013/2014

2

3
INDICE
Riassunto .............................................................................................. 5
1. Introduzione ..................................................................................... 7
1.1 Miopia: definizione, tipi e caratteri anatomici legati alla
miopia ............................................................................................... 7
1.2 Rischi legati alla miopia ............................................................11
1.3 Epidemiologia: prevalenza e incidenza ....................................12
1.3.1 Prevalenza .................................................................. 12
1.3.2 Incidenza e Progressione ........................................... 14
1.4 Impatto socio-economico della miopia .................................... 16
2. Fattori ambientali e miopia ............................................................ 18
2.1 Attività all’aperto e relazione con miopia ................................ 18
2.2 Lavoro prossimale e istruzione come fattori di rischio per lo sviluppo della miopia ...................................................................... 20
2.2.1 Lavoro prossimale e risposte accomodative e del
sistema di vergenza ............................................................ 31
3. Fattori genetici: la miopia come malattia multifattoriale ................. 35
3.1 Breve introduzione ai concetti chiave della genetica di malattie complesse ......................................................................... 35
3.1.1 Genotipo e fenotipo .................................................. 35
3.1.2 Valore fenotipico ....................................................... 38
3.1.3 Penetranza e espressività .......................................... 39
3.1.4 Malattie complesse e studi familiari ......................... 40
3.2 I componenti oculari refrattivi, in particolare LA, sono
caratteri geneticamente determinati ............................................. 42
3.3 Ricorrenza familiare degli errori refrattivi e loro ereditabilità.45
3.4 Geni candidati per la miopia..................................................... 49

4
4. Conclusione: fattori ambientali e genetici interagiscono in
modo complesso .................................................................................66
Bibliografia ..........................................................................................70

5
Riassunto
Tra i difetti refrattivi, la miopia è quella più studiata sia per l’influenza sulla vita
sociale sia per la molteplicità di fattori legati all’aumento progressivo e alle cause
che lo comportano. Date le evidenze sperimentali sull’ereditarietà della miopia e
le associazioni emerse tra prolungate attività da vicino, in questo elaborato sono
stati affrontati due temi importanti: il ruolo dell’ambiente e il ruolo della
genetica nello sviluppo e nella progressione della miopia.
Una delle possibili classificazioni della miopia considera l’età in cui l’errore
refrattivo inizia a svilupparsi. Generalmente si può suddividere in precoce e
tardiva [1]. La prima sembra essere più legata ad aspetti genetici e alla storia
famigliare dei difetti refrattivi, mentre nella seconda sembra essere
predominante la componente ambientale e in particolar modo il lavoro
prossimale [2].
I fattori ambientali, in base ai diversi studi visionati, hanno tendenzialmente un
ruolo protettivo o causativo dell’errore refrattivo preso in considerazione [3] [4]
[5]. Tra i fattori protettivi è stato analizzato quanto l’attività all’aperto sia
positivamente collegata alla miopia [6]. Invece, tra gli elementi causativi quello
che incide più negativamente è il lavoro prossimale [7] [8]. Come si può
riscontrare in molti studi, l’impegno eccessivo della vista in attività a distanza
ravvicinata può indurre un cambiamento refrattivo tendente alla miopia [9]. La
componente genetica sembra, però, prevalere su quella ambientale [10] [11]. Gli
studi approfondiscono l’ereditabilità delle componenti oculari, in particolar
modo della lunghezza assiale [12], e della ricorrenza familiare degli errori
refrattivi [13]. Emerge che la storia refrattiva dei genitori è determinante nel
possibile sviluppo e progressione miopica. Infatti, se entrambi i genitori sono
miopi il rischio per il bambino sarà più elevato sia di svilupparla sia di presentare
un valore maggiore della lunghezza assiale rispetto ai bambini emmetropi ed
ipermetropi [3] [14].

6
Sono stati analizzati anche i possibili geni implicati nell’errore refrattivo, noti in
base a studi di linkage o di associazione. Alcuni di questi geni sono stati
evidenziati da più studi, ma ancora oggi non è certo quali siano i più importanti e
soprattutto come le varianti geniche interagiscano nel determinare le
caratteristiche dell’occhio miope [15] [16].
La miopia, è considerata come un carattere complesso e multifattoriale. In
questo elaborato viene preso in considerazione non solo il ruolo di singoli fattori
genetici e ambientali nel determinare il suddetto errore refrattivo, ma anche che
diversi elementi interagiscono tra di loro, con meccanismi ed effetti ancora non
del tutto chiari [17] [18].

7
1. Introduzione.
1.1 Miopia: definizione, tipi e caratteri anatomici legati alla miopia.
Il potere refrattivo dell’occhio è determinato principalmente da quattro
componenti: la curvatura corneale (CC), la profondità della camera anteriore
(PCA), il potere della lente e la lunghezza assiale (LA, la distanza in millimetri
dalla cornea alla fovea). La relazione tra il potere refrattivo totale e la lunghezza
assiale determina lo stato refrattivo dell’occhio. Nell’occhio emmetrope i raggi di
luce entranti arrivano sulla retina a fuoco dando quindi una visione nitida
dell’oggetto posto all’infinito (figura 1) [19].
Nelle ametropie, invece, i raggi di luce non formano un’immagine a fuoco sulla
retina, bensì davanti alla retina (miopia) o dietro (ipermetropia). In entrambi i
casi, il risultato è una visione sfuocata dell’oggetto posto all’infinito.
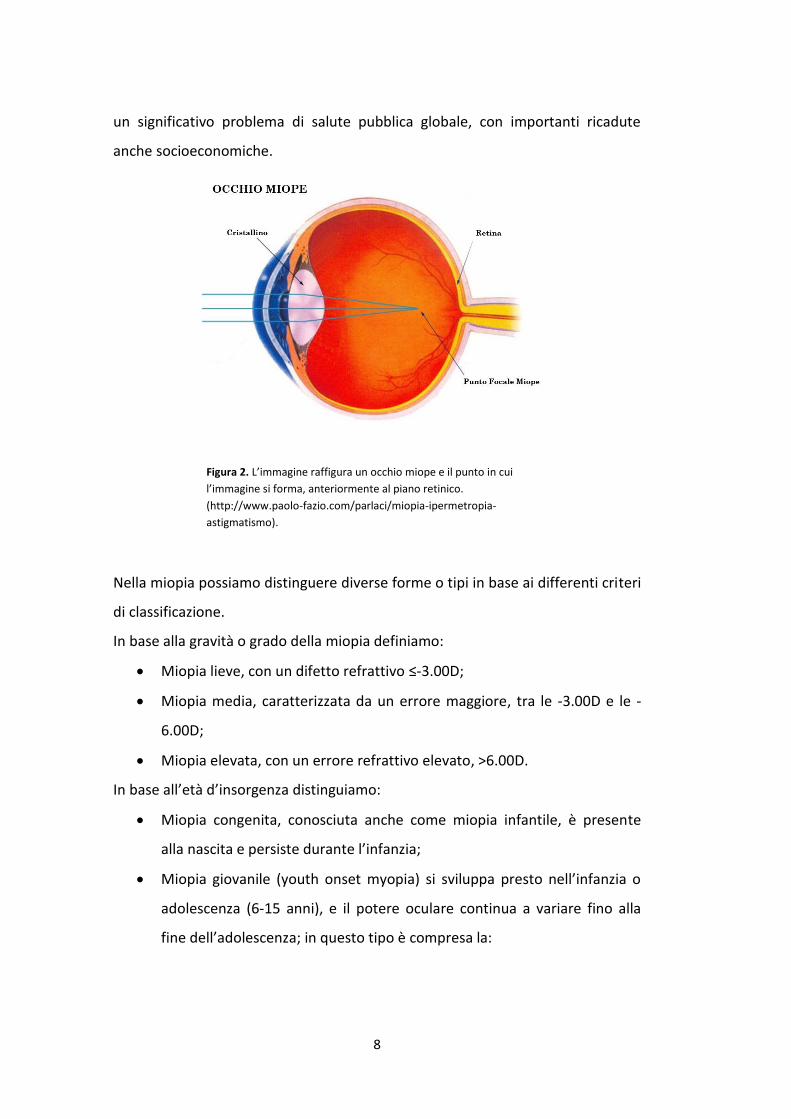
La miopia è un’ametropia sferica, in cui il piano focale dell’immagine, costruita
dal sistema ottico oculare, si trova prima del piano anatomico della retina e
rappresenta uno dei più comuni errori refrattivi (figura 2). La miopia è, di fatto,
Figura 1. L’immagine raffigura un occhio emmetrope e il punto in
cui si forma l’immagine. Essendo emmetrope essa si forma sul
piano retinico. (http://www.paolo-fazio.com/parlaci/miopia-
ipermetropia-astigmatismo).
8
un significativo problema di salute pubblica globale, con importanti ricadute
anche socioeconomiche.
Nella miopia possiamo distinguere diverse forme o tipi in base ai differenti criteri
di classificazione.
In base alla gravità o grado della miopia definiamo:
Miopia lieve, con un difetto refrattivo ≤-3.00D;
Miopia media, caratterizzata da un errore maggiore, tra le -3.00D e le -
6.00D;
Miopia elevata, con un errore refrattivo elevato, >6.00D.
In base all’età d’insorgenza distinguiamo:
Miopia congenita, conosciuta anche come miopia infantile, è presente
alla nascita e persiste durante l’infanzia;
Miopia giovanile (youth onset myopia) si sviluppa presto nell’infanzia o
adolescenza (6-15 anni), e il potere oculare continua a variare fino alla
fine dell’adolescenza; in questo tipo è compresa la:
Figura 2. L’immagine raffigura un occhio miope e il punto in cui
l’immagine si forma, anteriormente al piano retinico.
(http://www.paolo-fazio.com/parlaci/miopia-ipermetropia-
astigmatismo).

9
o Miopia scolastica (school myopia) appare durante l’infanzia,
particolarmente negli anni di scolarizzazione, è attribuita al lavoro
prossimale impegnativo durante gli anni scolastici;
Miopia adulta, distinguendo ulteriormente tra:
o Miopia adulta precoce inizia tra i 16 e i 40 anni;
o Miopia adulta tardiva il principio si ha dopo i 40 anni. [20].
Una terza classificazione si basa sui segni clinici:
Miopia semplice, la più comune, caratterizzata da un occhio troppo lungo
per il suo potere ottico o troppo potente per la sua lunghezza assiale.
Fattori genetici e ambientali sembrano contribuire entrambi allo sviluppo
della miopia semplice;
Miopia degenerativa, caratterizzata da un elevato grado di miopia
associato a cambiamenti degenerativi nel segmento posteriore
dell’occhio, con marcati segni visibili nel fondo oculare. Questa forma di
miopia peggiora progressivamente nel tempo. I cambiamenti
degenerativi possono risultare in funzioni visuali abnormali, come una
diminuzione dell’acuità visiva con migliore correzione o cambiamenti nel
campo visivo. Distacco di retina e glaucoma sono conseguenze molto
comuni in questa forma, considerata come una delle principali cause
d’invalidità visuale.
Miopia notturna, dovuta da un aumento di risposta accomodativa
associata a situazioni caratterizzate da bassi livelli di luminosità. A causa
di un contrasto insufficiente per uno stimolo accomodativo adeguato,
l’occhio assume la posizione di dark focus intermedio, invece di
focalizzare all’infinito.
Pseudomiopia, risultato di un aumento del potere refrattivo oculare
dovuto da sovrastimolazione del meccanismo accomodativo dell’occhio o
per spasmo ciliare. La condizione è così chiamata perché il soggetto
risulta essere miope solo a causa di risposte accomodative inadeguate.

10
Miopia indotta, secondaria all’esposizione a diversi agenti farmaceutici,
variazione degli zuccheri nel sangue, sclerosi nucleare della lente del
cristallino, o altre condizioni anomale. Questa miopia è spesso
temporanea e reversibile.
Queste tipologie di miopia sopra descritte, sono forme della cosiddetta miopia
non sindromica. La miopia sindromica, come ci suggerisce il nome, può risultare
come sintomo associato ad altri sintomi diversi in patologie complesse (sindromi)
che spesso coinvolgono più organi e apparati. Gli errori refrattivi sindromici sono
generalmente monogenici o poligenici e possono presentarsi all’interno di un più
ampio spettro di segni clinici. La miopia viene riportata come segno in numerose
sindromi oculari, può anche essere una caratteristica peculiare in malattie
ereditabili del tessuto connettivo. Ad alcune di queste sindromi sono stati
associati dei loci genetici, ma nessuno di questi loci trovati risulta essere
associato alla forma comune di miopia. In questo elaborato sarà presa in
considerazione specificamente la miopia non sindromica.
La compensazione della miopia deriva dall’utilizzo delle lenti negative che
allungando la focale, permettono la formazione di un’immagine retinica nitida
(figura3).
La prima e più usata forma di correzione è l’occhiale in quanto di facile uso e non
molto costoso; esso però comporta inevitabili limiti, legati al campo visivo della
lente, che aumentano quanto più è elevata la miopia che correggono. Le lenti
utilizzate per le miopie moderate ed elevate, inoltre fanno percepire al soggetto
Figura3. Occhio miope senza e con correzione di una lente negativa.(www.wikipedia.org).

11
un’immagine rimpicciolita, non reale e anche quest’effetto è proporzionale al
grado di severità dell’ametropia influenzando la qualità della vita dei pazienti che
ne fanno uso. Gli occhiali sono pesanti tanto da essere causa di antiestetici solchi
sul naso e spesso limitano il soggetto nella pratica di diverse attività sportive. La
qualità visiva migliora con l’applicazione delle lenti a contatto: non c’è
rimpicciolimento o distorsione periferica dell’immagine, vengono risolti i
problemi di natura estetica e funzionale. Scelta che, con le opportune valutazioni
da parte dell’ottico optometrista, riesce a soddisfare le più svariate esigenze
(applicazione morbida o rigida monofocale o multifocale, porto diurno, continuo
o notturno con le lenti ortocheratologiche), ma che comporta, comunque, un
costo per l’acquisto e la manutenzione.
1.2 Rischi e limiti legati alla miopia.
La miopia elevata congenita o acquisita può anche essere accompagnata da
diverse patologie oculari. Tale errore refrattivo è un fattore di rischio per lo
sviluppo di differenti condizioni oculari incluse: degenerazione periferica retinica,
cataratte associate all’età, glaucoma e neovascolarizzazione coroidea [21]. La
miopia progressiva è caratterizzata da un graduale allungamento del globo
oculare accompagnato da cambiamenti degenerativi, che potenzialmente
possono portare ad alterazioni fino alla cecità, nella retina e nella coroide [22].
Un occhio che soffre di miopia elevata con una maggiore lunghezza assiale
sembra essere più predisposto all’insorgenza di cataratte nucleari, che, inoltre, si
presentano con maggiore densità nucleare rispetto a quelle dei soggetti non
miopi [23].
Molte occupazioni richiedono requisiti visivi per poter accedere ad esse. Spesso,
quindi, soggetti con elevati, ma anche moderati, errori refrattivi devono fare i
conti con limitazioni nel lavoro oltre che nella vita personale. Ne sono un
esempio i concorsi alla polizia di stato, al ruolo degli agenti e assistenti è ritenuta
necessaria un’acuità visiva (AV) di 12/10 binocularmente, con non meno di 5/10
nell’occhio che vede meno ; un visus corretto di 10/10 per ciascun occhio per

12
una correzione massima complessiva di 1D quale somma dei singoli errori
refrattivi (www.docvadis.it). Un altro esempio sono i requisiti visivi richiesti
nell’accademia della marina militare che impongono un visus corretto di 10/10 in
ciascun occhio, se corretto con lenti ben tollerate di massimo -1.75 diottrie.
Invece nei concorsi di aeronautica militare, per il ruolo di naviganti normale, è
esclusa qualsiasi possibile correzione dell’errore refrattivo (anche con
laserterapia), avendo quindi un visus naturale di 10/10 (www.concorsimilitari.it).
Ai piloti civili viene richiesta un’AV, con o senza correzione, di almeno 7/10
monocularmente, e almeno 10/10 binocularmente, l’errore refrattivo in ogni
caso non deve superare le -3D (www.aviazionecivile.org). Una limitazione più
vicina alla vita di ogni giorno è quella imposta nelle patenti di guida dei veicoli.
Nella patente di tipo A e B (gruppo 1) il visus corretto o meno deve essere di
almeno 7/10, come somma dei due occhi, e l’occhio che vede peggio deve
riconoscere almeno i 2/10; per le patenti di tipo C, D e E (gruppo 2) occorre
possedere un’AV pari ad almeno 8/10 come somma dei due occhi e l’occhio che
vede peggio deve riconoscere almeno i 5/10, raggiungibile anche con correzione
purché la differenza tra le due lenti non sia superiore a tre (www.aci.it). I casi di
limitazione sono molteplici e, pur essendo spesso, comunque, parametri
facilmente superabili, rappresentano uno sbarramento per qualcuno creando
esigenze specifiche: necessità di cambiare o rinunciare al lavoro, agli
spostamenti, o ad un’attività. Non va dimenticato, inoltre, che la miopia limita
diverse abilità visive, non solo la capacità di distinguere dettagli. La sensibilità al
contrasto, la visione crepuscolare notturna, solo per fare alcuni esempi, ne
risultano coinvolte, peggiorando o modificando diversi aspetti e la qualità della
vita.
1.3 Epidemiologia: prevalenza e incidenza.
1.3.1 Prevalenza.
A metà del 2007, in tutto il mondo, The World Health Organisation ha stimato
circa 703milioni di casi di invalidità visuale a causa di errori refrattivi non corretti,

13
dei casi totali, 158milioni corrispondono a quelli di invalidità visiva in distanza
[24].
Uno studio evidenzia che entro l’anno 2020 i casi di miopia saranno 25 miliardi,
ovvero un terzo della popolazione del mondo sarà caratterizzata dalla miopia
[25]. Uno studio trasversale compiuto tra studenti australiani riporta una
prevalenza del 52.5% e del 59.1% nei dodicenni e diciasettenni rispettivamente,
provenienti dall’est asiatico; mentre la corrispondente stima di prevalenza nei
giovani di etnia europea-caucasica della stessa età è del 8.6% e del 17.7% [26].
Molti studi recenti hanno riportato una considerevole variazione nella prevalenza
della miopia tra bambini di diverse etnie, localizzazioni geografiche ed età.
Variazioni di prevalenza dovute da diverse localizzazioni geografiche sono state
largamente riportate da molti studi. I valori di prevalenza trovati nell’est e sud
est asiatico asiatico sono molto più elevati rispetto a quelli trovati nelle altre
regioni del mondo. Uno studio sulla prevalenza miopica in Cina mostra un valore
del 16.2% di alunni tra i 5-15 anni che presentano tale errore refrattivo e vivono
nelle zone rurali della Cina del nord [27]. Mentre nelle città metropolitane della
Cina del sud gli studenti della stessa età miopi risultano essere 38.1% in
Guangzhou [28] e del 36.7% in Hong Kong [29](i dati sono riassunti nella tabella
I).
Prevalenza
Area Geografica Età Prevalenza %
Studio
Est- Asia 12 anni, 17 anni
52.5/59.1 [26]
Europa/ Asia caucasica
12 anni, 17anni
8.6/17.7 [26]
Cina rurale 5-15 anni 16.2 [27]
Cina urbana 5- 15 anni 37.4 [28] [29]
Tabella I. Percentuale della prevalenza di miopia nelle diverse aree
geografiche.

14
Negli adulti i valori di prevalenza sembrano variare nelle diverse fasce d’età.
Nello studio “Beaver Dam Eye” la prevalenza di miopia passa dal 42.9%, negli
adulti di 43-54anni, a 25.1% negli adulti americani di 55-64anni; un’ulteriore
diminuzione risulta nei soggetti di 65-74 anni che mostrano una prevalenza di
miopia del 14.8% e poi diminuisce ancora leggermente negli individui di ≥75 anni,
stima del 14.4% [30]. Un altro studio, su larga scala, che prende come soggetti
individui di 40 anni o più delle aree urbane americane mostra un evidente
decremento nella prevalenza dei soggetti miopi aumentando l’età di donne di
diverse etnie e uomini bianchi. Emerge anche un modello bimodale tra gli
individui afro-americani, con picchi di prevalenza trovati nei soggetti di 40-49
anni e nei soggetti di 80 anni o più. Un modello simile descrive bene, anche la
prevalenza della miopia negli adulti di Singapore, che sembra essere, infatti, più
alta nei quarantenni e nei settantenni [31]. È ancora un argomento molto
dibattuto se queste variazioni di prevalenza miopica correlate all’età siano
dovute al tipo di studio longitudinale o al gruppo preso in considerazione [32].
Comunque la distribuzione bimodale della prevalenza miopica sembra
attribuibile a due fattori diversi che fanno sì che due età lontane siano i picchi di
prevalenza: la lunghezza assiale più lunga nei giovani e la sclerosi nucleare della
lente negli anziani che portano, di fatto, ad un aumento della prevalenza della
miopia nelle due classi d’età.
1.3.2 Incidenza e progressione.
Per quanto riguarda l’incidenza della miopia, un recente studio su studenti
australiani di 6 e 12 anni, riporta un’incidenza annuale di miopia del 2.2% nel
gruppo più giovane (6.7 ± 0.4 anni in media) mentre nell’altro gruppo (12.7 ± 0.4
anni in media) del 4.1%. I valori di incidenza annuale nell’est asiatico (6.9% nel
gruppo più giovane e 7.3% nel gruppo più vecchio) risultano essere molto più alti
rispetto ai bambini euro-caucasici (più giovani 1.3% e più vecchi 2.9%). In
entrambi i gruppi si è evidenziato un notevole aumento dell’incidenza nel tempo:
dal 1.4% al 14.4% nel gruppo più giovane (durante un periodo di 6.1±0.8 anni) e

15
dal 13.0% al 29.6% nel gruppo più vecchio (in un periodo di follow-up di 4.5±0.3
anni) [26] (dati riassunti nella tabella II).
Incidenza
Area geografica Età Incidenza % Studio
Australia 12 anni, 17 anni
2.2 %, 4.1% [26]
Est- Asia 12 anni, 17 anni
6.9%, 7.3% [26]
Europa/ Asia caucasica
12 anni, 17 anni
1.3%, 2.9% [26]
Secondo una pubblicazione del 2004, in Taiwan tra il 1983 e il 2000 la prevalenza
di miopia nei giovani (7-18 anni) è aumentata significativamente: per gli
adolescenti tra i 16 e i 18 anni il valore è variato all’incirca del 14% mentre nei
bambini di 7 anni è variato del 262% [33]. Una simile tendenza si nota in un altro
resoconto sulle variazioni della prevalenza miopica in 30 anni negli Stati Uniti (tra
il 1971 e il 2004). In tutti i gruppi di differenti età la prevalenza della miopia ha
subito un significativo aumento in tre decadi. Ad esempio il valore di prevalenza
miopica nei giovani americani dai 12 ai 17anni aumenta del 12.0% (1971-1972) al
31.2% (tra il 2000 e il 2004) [34]. In Finlandia, la prevalenza nei bambini di 7-8
anni risulta invece essere rimasta costante, negli ultimi 20 anni, mentre nei
giovani di 14-15 anni la prevalenza risulta almeno raddoppiata (da 10.6% a
21.8%) [35].
In conclusione, è largamente riportato che la prevalenza della miopia nelle
recenti decadi è aumentata significativamente, se pur non in tutte le fasce d’età
e in tutte le aree geografiche considerate (tabella III).
Tabella II. Percentuale di incidenza di miopia nelle differenti aree
geografiche per studenti di 12 e 17 anni.
16
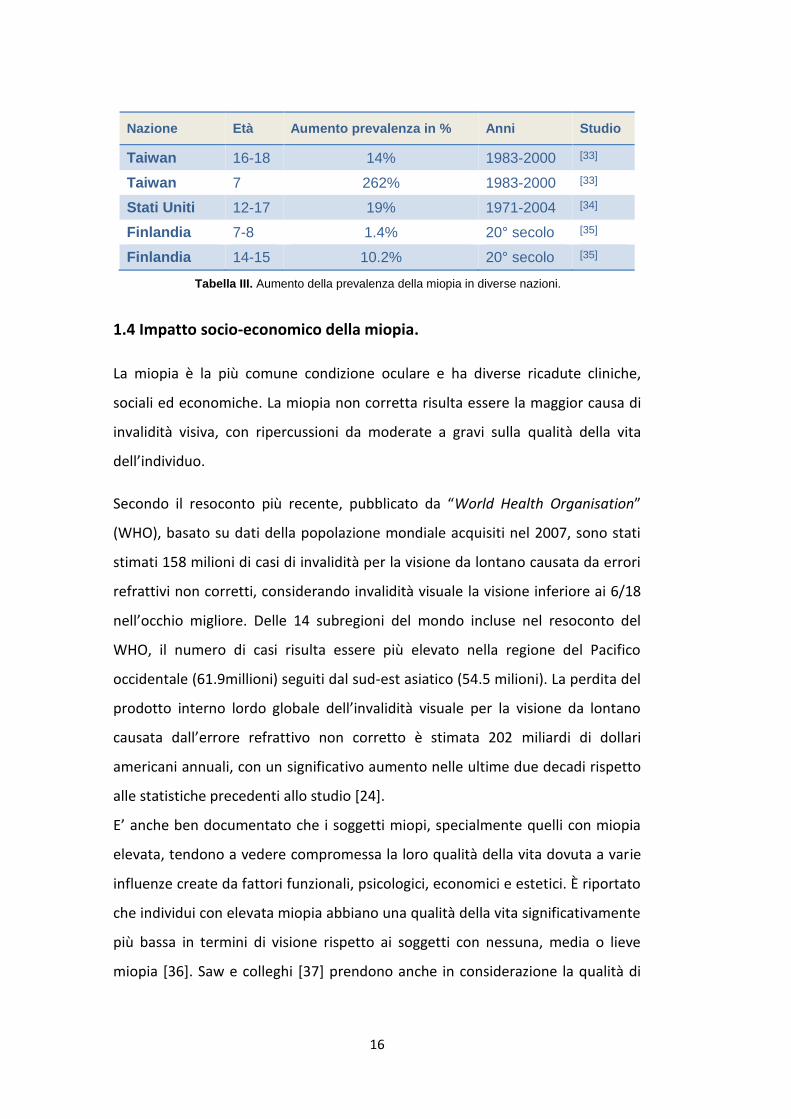
Nazione Età Aumento prevalenza in % Anni Studio
Taiwan 16-18 14% 1983-2000 [33]
Taiwan 7 262% 1983-2000 [33]
Stati Uniti 12-17 19% 1971-2004 [34]
Finlandia 7-8 1.4% 20° secolo [35]
Finlandia 14-15 10.2% 20° secolo [35]
1.4 Impatto socio-economico della miopia.
La miopia è la più comune condizione oculare e ha diverse ricadute cliniche,
sociali ed economiche. La miopia non corretta risulta essere la maggior causa di
invalidità visiva, con ripercussioni da moderate a gravi sulla qualità della vita
dell’individuo.
Secondo il resoconto più recente, pubblicato da “World Health Organisation”
(WHO), basato su dati della popolazione mondiale acquisiti nel 2007, sono stati
stimati 158 milioni di casi di invalidità per la visione da lontano causata da errori
refrattivi non corretti, considerando invalidità visuale la visione inferiore ai 6/18
nell’occhio migliore. Delle 14 subregioni del mondo incluse nel resoconto del
WHO, il numero di casi risulta essere più elevato nella regione del Pacifico
occidentale (61.9millioni) seguiti dal sud-est asiatico (54.5 milioni). La perdita del
prodotto interno lordo globale dell’invalidità visuale per la visione da lontano
causata dall’errore refrattivo non corretto è stimata 202 miliardi di dollari
americani annuali, con un significativo aumento nelle ultime due decadi rispetto
alle statistiche precedenti allo studio [24].
E’ anche ben documentato che i soggetti miopi, specialmente quelli con miopia
elevata, tendono a vedere compromessa la loro qualità della vita dovuta a varie
influenze create da fattori funzionali, psicologici, economici e estetici. È riportato
che individui con elevata miopia abbiano una qualità della vita significativamente
più bassa in termini di visione rispetto ai soggetti con nessuna, media o lieve
miopia [36]. Saw e colleghi [37] prendono anche in considerazione la qualità di
Tabella III. Aumento della prevalenza della miopia in diverse nazioni.

17
vita associata alla correzione ottica. Dimostrano che gli individui miopi che
indossano occhiali e/o lenti a contatto hanno livelli significativamente più alti di
preoccupazione di ferirsi, difficoltà nel far fronte alle esigenze della vita e meno
sicurezza nell’adempiere le attività di ogni giorno, se paragonati agli emmetropi.
Al contrario negli individui che hanno subito chirurgia refrattiva non risultano
particolari differenze di qualità di vita rispetto al gruppo di emmetropi. Questo
studio indica che la miopia corretta con occhiali o lenti a contatto ha un impatto
negativo negli aspetti relativi alla visione, della qualità della vita delle persone, se
paragonate a miopi che hanno subito chirurgia refrattiva. Non emerge
necessariamente corrispondenza tra miopia e carattere introverso, come
indagato tra i soggetti del GEM study (Genes in Myopia) [38].

18
2. Fattori ambientali e miopia.
2.1 Attività all’aperto e relazione con miopia.
Diversi fattori ambientali sembrano avere un ruolo protettivo o causativo
rispetto allo sviluppo o alla progressione della miopia.
Mentre lunghi e costanti periodi di lettura o attività prossimali aumentano il
rischio di sviluppare miopia, altri fattori ambientali hanno dimostrato un’azione
protettiva. Infatti, da studi recenti è emerso che il tempo trascorso in ambienti
ampi e all’esterno, immersi nella natura e la pratica di sport all’aperto durante
l’infanzia è associato a una diminuzione del rischio di sviluppare miopia [3] [39]
[40]. Inoltre il tempo impiegato nelle attività all’aperto, con il suo conseguente
effetto benefico sulla miopia, non sembra sostituire le ore dedicate nelle attività
prossimali. Interessante è l’evidenza che i fattori genetici possano interagire con
la variabile dell’attività all’esterno e il rischio di miopia. Jones et al. dimostrano
che la storia refrattiva miopica dei genitori è un importante predittore nei
modelli univariati e multivariati. Le ore alla settimana di sport e attività
all’esterno hanno differenti effetti quando vengono considerati insieme, come
variabile, al numero di genitori miopi: meno ore di sport e attività all’aperto
aumentano le probabilità di diventare miope nei bambini con due genitori miopi,
probabilità più elevata rispetto ai bambini con uno o nessun genitore miope. La
possibilità di divenire miope per i bambini con nessun genitore miope sembra
essere più bassa nei bambini con una più alta quantità in ore di sport e attività
all’aperto, paragonati a quelli con due genitori miopi. Sherwin et al. nel 2012 [6]
realizzano una meta-analisi, riassumendo in una revisione tutti i risultati
disponibili e rilevanti riguardanti la relazione tra tempo speso all’aperto e miopia
con relativa progressione. Esaminando tutti gli articoli sul tema arrivano alla
conclusione che l’aumento del tempo trascorso all’esterno può conferire una
modesta, ma significativa, riduzione del rischio dello sviluppo miopico e la sua
progressione. Trovano, inoltre, una significativa associazione protettiva tra
l’aumento del tempo all’esterno e la prevalenza della miopia in un gruppo di

19
circa 10.000 bambini e adolescenti con età inferiore ai 20 anni: ogni aumento di
un’ora alla settimana di tempo trascorso all’esterno è associato ad una riduzione
del 2% della probabilità di miopia. Anche se questa rappresenta una debole
associazione protettiva, la sua interpretazione dipende dall’unità della misura
presa in considerazione. Infatti aumentando di 7 ore il tempo trascorso
all’esterno, per settimana, l’associazione protettiva diviene più forte. Un’alta
percentuale degli studi disponibili sono stati effettuati nell’est asiatico, dove la
prevalenza di miopia è molto elevata e richiede, quindi, maggiori sforzi nella
ricerca dei vari meccanismi legati ad essa. Tale relazione sembra essere meno
marcata nelle popolazioni che non provengono dall’Asia dell’est. Questo fatto
può riflettere la scarsa quantità di tempo speso in attività all’aperto dai bambini
che abitano in quell’area, piuttosto che come conseguenza di differenze etniche
intrinseche. Rimane possibile che i bambini con miopia trascorrano meno tempo
in attività all’esterno rispetto ai bambini non miopi, riflessione supportata da
studi sul tipo di personalità tra persone aventi o meno miopia, in più
dall’evidenza che in aree urbane, dove si presentano spazi aperti meno ampi, si
concentrano maggiormente individui miopi che, conseguentemente, hanno
meno possibilità di partecipare a sport e ad attività all’aperto. Il tempo trascorso
all’esterno è influenzato da molti fattori, per esempio dall’etnia, da fattori
ambientali quali il clima, dai livelli di visibilità e dagli impegni scolastici. Questi
risultati, in generale, suggeriscono la necessità di studi controllati, randomizzati,
ben strutturati, in modo da investigare l’effetto dell’aumento delle ore di attività
all’aperto come un fattore protettivo, riducente il rischio di miopia e la sua
progressione per intervenire nella prevenzione in bambini e adolescenti. Esistono
miriadi di possibili meccanismi che giustifichino l’effetto protettivo del tempo
trascorso in attività all’aperto. Di questi fanno parte l’aumento del rilascio della
dopamina retinica in risposta alla luce del sole (la dopamina inibisce
l’allungamento assiale nella miopia sperimentale [41], e l’effetto protettivo può
essere bloccato dall’antagonista della dopamina, lo spiperone [42]); l’intensità
luminosa maggiore in ambienti all’aperto (miosi pupillare, aumento della

20
profondità di campo, diminuzione dello sfocamento e rallentamento della
crescita del bulbo) [43], e una richiesta accomodativa inferiore per la visione in
lontananza [44].
Una precisa comprensione dei fattori di rischio per la miopia e la sua
progressione è necessaria per potere rendere effettive delle strategie di
prevenzione all’errore refrattivo preso in considerazione, prevedendo magari
anche durante l’orario scolastico attività all’aperto.
2.2 Lavoro prossimale ed istruzione come fattori di rischio per sviluppo
della miopia.
Già negli anni ottanta era nota la correlazione tra miopia e istruzione infatti
Taylor, da un suo studio, riporta che la miopia è presente in maggior percentuale
tra i bambini scolarizzati ed istruiti e meno prevalente tra i bambini analfabeti
[45]. Si presume che lo sforzo accomodativo durante il lavoro prossimale possa
essere un fattore causativo nello sviluppo della miopia [46].
La visione prossimale può influenzare la miopia direttamente attraverso
l’accomodazione o indirettamente a causa dei fenomeni ad essa correlati. È stato
ipotizzato un processo in tre fasi dell’uso eccessivo dell’accomodazione:
Una condizione refrattiva simile alla miopia può essere raggiunta per un
eccesso di accomodazione, come se l’accomodazione rimanesse per un
tempo superiore al normale parzialmente attiva e, alternando la visione
da vicino a lontano, la focalizzazione in distanza risulti temporaneamente
difficile;
Successivamente, questa condizione si mantiene per lungo tempo e causa
un’alterazione della condizione di accomodazione in posizione di riposo o
dell’accomodazione minima tale da essere indistinguibile dalla condizione
refrattiva non accomodata;
Infine, questo stato di accomodazione prolungata causa un’alterazione
delle strutture anatomiche tale che l’occhio diviene anatomicamente
miope [1].

21
I dati epidemiologici attuali evidenziano l’aumento apprezzabile, nelle ultime 2/3
generazioni, della prevalenza miopica in diverse aree di tutto il mondo. Poiché i
caratteri genetici delle popolazioni non possono essere cambiati
significativamente nell’arco di così poco tempo, è improbabile che fattori
genetici abbiano giocato un ruolo in questa tendenza, ed è, quindi, più probabile
che fattori ambientali abbiano influenzato la maggiore incidenza di miopia che
caratterizza queste ultime generazioni. Infatti, molti studi riportano
l’associazione tra miopia e fattori ambientali come lo status socio-economico più
elevato e raggiungimento di livelli di educazione più elevati. All’interno del
contesto del processo di miopizzazione, l’istruzione, lo status socio-economico e
l’occupazione sono considerate come variabili correlate al maggiore utilizzo della
visione da vicino. Gli studi sulla prevalenza della miopia mostrano un aumento di
essa negli abitanti di Singapore con livelli di educazione più elevati, un reddito
personale mensile maggiore e occupazione associata al lavoro prossimale [7]. Alti
livelli di miopia sono stati trovati anche nei bambini coreani provenienti da
famiglie con reddito elevato [47]. I bambini miopi sembrano avere una più forte
storia refrattiva familiare nelle famiglie con genitori più istruiti, maggiore reddito,
e come occupazione impiegati o professionali.
Diversi studi sull’effetto della lettura hanno cercato di dimostrare la diretta
relazione tra miopia e attività prossimali, ma tale relazione è molto complessa e
ancora poco compresa. Le stime dell’esposizione al lavoro prossimale sono
soggette a considerevoli errori di misura. Gli effetti stimati possono variare
dipendendo dall’unità di misura scelta, dalla definizione dell’esito, o per l’età,
etnia e circostanze sociali dei soggetti studiati. Queste discrepanze nelle
caratteristiche e nelle metodologie degli studi possono portare a risultati
discordanti. Inoltre, la diffusione della tecnologia attuale come computer,
dispositivi di gioco, cellulari e smartphone ha aggiunto un livello di complessità
maggiore alla domanda di lavoro prossimale.
Lo studio di Saw e colleghi, esaminando le refrazioni di 1204 studenti cinesi tra i
10 e i 12 anni, stima che i fattori di rischio ambientali spiegano solo l’ 11.6% della

22
varianza miopica [37]. Similmente, Mutti trova che fattori come anno scolastico,
genitori miopi e lavoro prossimale spieghino solo il 12% della varianza miopica
[17].
Nel 2001, Hepsen et al. prendono in considerazione un campione di 177 individui
divisi in due gruppi in base all’errore refrattivo di partenza: il primo gruppo
(emmetropi) composto da soggetti caratterizzati da un difetto refrattivo tra
+0.50D e -0.50D (n=78) e il secondo gruppo formato da soggetti con errore
refrattivo tra +1.00D e -1.00D (n=114). Questi due gruppi, analizzati nei tre anni
successivi dall’inizio dello studio, vengono suddivisi in due sottogruppi ciascuno
separando gli studenti di una scuola privata dagli apprendisti lavoratori
specializzati. Nel primo gruppo, dei 41 studenti, 20 subiscono un cambiamento
miopico nella refrazione (48.8%) mentre i restanti non mostrano significativi
cambiamenti; sempre nel primo gruppo su 37 apprendisti 7 mostrano un
cambiamento refrattivo verso la miopia (18.9%), 27 non mostrano alcun
cambiamento (73.0%) e 3 subiscono una variazione verso l’ipermetropia (8.1%)
(tabella IV). Nel secondo gruppo dei 67 studenti, 40 subiscono un cambiamento
verso la miopia (59.7%), 27 non mostrano cambiamenti significativi; dei 47
apprendisti, 10 (21.3%) mostrano un cambiamento refrattivo verso la miopia, 34
(72.3%) non mostrano alcun cambiamento e 3 (6.4%) subiscono un cambiamento
verso l’ipermetropia (tabella V). Dai dati si evince che la progressione miopica
risulta essere più significativa nei soggetti frequentanti la scuola privata rispetto
agli apprendisti. La maggior parte dei cambiamenti refrattivi nel primo gruppo è
stata riscontrata nei primi 18 mesi di studio. Quest’analisi non mette in evidenza
l’esatta causa della progressione; la spiegazione più ragionevole è che i 18 mesi
considerati corrispondano al primo anno per questi studenti nella loro scuola
secondaria privata e quindi ad una maggiore quantità di studio. Anche gli
apprendisti devono comunque eseguire lavori da vicino, e nei risultati si
evidenzia una significativa differenza tra i due gruppi basata principalmente sulle
ore trascorse in attività prossimali. Il cambiamento miopico in questo studio è
stato associato ad un aumento statisticamente significativo nella media della

23
profondità della camera anteriore, della profondità della camera vitrea e
conseguentemente della lunghezza assiale. In conclusione quindi da questo
studio possiamo trarre che la lettura e le attività prossimali, ovvero fattori
ambientali, possano causare un cambiamento miopico nei giovani adolescenti,
che è attribuibile alla visione prossimale sostenuta [9].
Gruppo 1 (ER ± 0.50D)
Studenti (n=41) Apprendisti (n=37)
Cambio miopico
Nessun cambio
Cambio ipermetropico
Cambio miopico
Nessun cambio
Cambio ipermetropico
20 (48.8%)
21
(51.2%)
0 7
(18.9%)
27 (73.0%)
3
(8.1%)
Gruppo 2 (ER ± 1.00D)
Studenti (n=67) Apprendisti (n=47)
Cambio miopico
Nessun cambio
Cambio ipermetropico
Cambio miopico
Nessun cambio
Cambio ipermetropico
40 (59.7%)
27 (40.3%)
0 10 (21.3%)
34 (72.3%)
3
(6.4%)
Un altro studio prende come campione bambini dai 7 ai 9 anni. Dei 1005 soggetti
l’8.1% presenta miopia di grado elevato e il 24.3% miopia lieve. I valori di
prevalenza, in generale, sono più alti nei bambini cinesi (37.0%) che nei bambini
non cinesi (19.9%) presi in considerazione. È evidente anche un’associazione
positiva tra miopia elevata e status socioeconomico considerato nei termini dei
redditi familiari più elevati e istruzione avanzata dei genitori. Inoltre, nei bambini
cinesi emerge una significativa associazione tra miopia elevata e libri letti alla
settimana, utilizzo del computer (valore due volte più alto di miopia elevata) e
lezioni di musica; e se, in particolare hanno almeno un genitore miope,
presentano l’inizio dello sviluppo miopico in età precoce. Mentre nei bambini
Tabella V. Cambiamenti refrattivi degli studenti e degli apprendisti non emmetropi,
nell’arco di 3 anni. [9]
Tabella IV. Cambiamenti refrattivi degli studenti e degli apprendisti emmetropi, nell’arco di
3 anni. [9]

24
non cinesi nessuna di queste associazioni è significativa. Inoltre i bambini che
leggono più di due libri alla settimana e che hanno almeno un genitore miopie
presentano una lunghezza assiale maggiore, che non risulta essere una relazione
statisticamente significativa, se non associata alla miopia, mettendo in evidenza
che l’associazione tra miopia e lavoro prossimale è più forte nei bambini cinesi
che in quelli non cinesi. Questo suggerisce, ancora una volta, che il lavoro
prossimale e altri fattori ambientali possano influenzare diversamente il possibile
sviluppo di errori refrattivi nei bambini cinesi rispetto ai non cinesi. Nel modello
multivariato, il fattore di rischio più forte per miopia elevata sono i libri letti alla
settimana, seguiti dall’età, dalla miopia presente o meno nei genitori e dal tipo di
scuola frequentata. Tali fattori predicono, indipendentemente, la miopia di alto
grado. Altri studi mettono in evidenza che i bambini miopi hanno maggiore
probabilità di avere genitori miopi e di provenire da famiglie con un più elevato
status economico [31] [48]. È postulato che lo status socioeconomico possa
essere un surrogato di fattori ambientali di stili di vita come i traguardi
accademici, abilità intellettive o lavoro prossimale. Il tipo di scuola è stato
associato alla miopia e può essere un surrogato di altri fattori ambientali
correlati alla miopia: intelligenza, personalità, o altri non identificati fattori di
rischio ambientali. Questo studio in qualche modo prova ulteriormente la
correlazione tra miopia e lavoro prossimale, ma non risolve il dubbio se le attività
a distanza ravvicinata siano un fattore di rischio per lo sviluppo di miopia o se sia
un surrogato di altri fattori ambientali o genetici [49].
Sempre nel 2002, Mutti nota che i bambini miopi trascorrono più tempo in
attività prossimali, per compiti scolastici e lettura di piacere, e meno in attività
sportive, paragonati agli emmetropi [17]. Le ore impiegate nella visione della TV
o nel gioco con videogame è uguale in tutti e tre i gruppi (miopi, emmetropi,
ipermetropi). I bambini emmetropi e ipermetropi si eguagliano nell’utilizzo delle
ore per le diverse attività (tabellaVI).

25
Attività Tutti i soggetti (n=366)
Miopi
(n=67)
Emmetropi
(n=271)
Ipermetropi
(n=28)
Studio 9.4 ± 5.7 11.2 ± 7.2* 8.9 ± 5.2* 9.4 ± 4.9
Lettura di piacere
4.4 ± 4.5 5.8 ± 4.8* 4.1 ± 4.6* 3.6 ± 2.9
Tv 8.3 ± 5.9 9.2 ± 6.8 8.3 ± 5.7 6.6 ± 4.5
Video-game e computer
2.3 ± 3.3 2.7 ± 4.1 2.2 ± 3.2 1.4 ± 1.8
Sport 9.3 ± 6.4 7.4 ± 6.7* 9.7 ± 6.2* 9.8 ± 7.9
I punteggi dei test di profitto (utilizzando il test Iowa Tests of Basic Skills) sono
più alti nei bambini miopi che negli emmetropi, nell’area della lettura e della
lingua, sia considerando i percentili nazionali che quelli locali. Mentre paragonati
ai bambini ipermetropi, i bambini miopi realizzano un maggior punteggio
nell’area della lettura sia percentile nazionale che locale, nella lingua nazionale,
ma non in quella locale, questa inconsistenza può dipendere dalla fonte dei
punteggi, suggerendo che l’associazione tra miopia e i risultati di lettura misurati
da ITBS possa essere debole. Una delle difficoltà nel valutare questi fattori di
rischio è la loro interconnessione, e il loro potenziale, di confondere
l’associazione con la miopia. Infatti punteggi nell’area della matematica non
mostrano differenze tra i tre gruppi, mentre punteggi più alti dei bambini miopi
in lettura e lingua sembrano essere il risultato di un comfort visivo maggiore
durante il test. Pertanto risultati simili nella parte di matematica suggeriscono
che ogni gruppo refrattivo può vedere egualmente bene il test, ma che i gruppi
possano differire in abilità specifiche nel linguaggio (tabella VII).
Tabella VI. Paragone tra le ore trascorse da soggetti miopi, emmetropi e ipermetropi per le
attività prese in considerazione in questo studio. Tra emmetropi e ipermetropi nessun paragone
è significativo. Paragoni tra miopi e emmetropi sono significativi come sottolineato nella
tabella*. I dati sono espressi come ore in media ± DS. [17].

26
ITBS subtest
Tutti i soggetti
(n=366)
Miopi
(n=67)
Emmetropi
(n=271)
Ipermetropi
(n=28)
Nazionale
Lettura 79.6 ± 23.2 82.9 ± 23.7!^ 79.2 ± 23.1! 75.3 ± 22.9^
Lingua 82.8 ± 19.0 86.6 ± 17.7!^ 82.2 ± 19.2! 790 ± 20.2^
Matematica 83.8 ± 19.8 84.1 ± 21.4 83.5 ± 20.0 86.3 ± 13.6
Locale
Lettura 53.7 ± 29.6 62.5 ± 31.0! 52.6 ± 28.9! 41.5 ± 26.6
Lingua 55.1 ± 28.8 64.2 ± 29.7!^ 53.2 ± 28.1! 52.2 ± 29.2^
Matematica 54.5 ± 28.5 57.4 ± 29.6 53.6 ± 28.4 59.7 ± 26.7
Ip studia un campione di studenti australiani di 12anni, dimostrando che le
principali attività prossimali sono lo studio e l’uso del computer, anche se ci sono
differenze nei sottogruppi per etnie, sesso e scolarizzazione [50]. Le ragazze
dedicano più tempo dei loro coetanei di sesso maschile nelle attività prossimali;
per quanto riguarda l’etnia, in totale gli studenti euro-caucasici trascorrono
meno ore in attività da vicino rispetto agli studenti dell’est asiatico. In generale
esiste solamente una debole correlazione tra equivalente sferico e lavoro
prossimale, quest’ultimo si associa debolmente anche con la lunghezza assiale e
la curvatura corneale. I bambini che riportano una lettura continuativa per lunghi
periodi di tempo tendono a sviluppare miopia, quest’attitudine però non
raggiunge una significatività statistica e non risulta neppure avere una relazione
con la lunghezza assiale tanto meno con la curvatura corneale. Trascorrere molto
tempo nella pratica di attività all’esterno, invece, tra gli studenti adolescenti di
Singapore, viene associato ad una refrazione ipermetropica più lieve. Eseguendo
il modello multivariato, le associazioni significative con la miopia risultano essere
l’etnia, il tipo di scuola, la presenza di miopia nei genitori, la lettura continuativa
per 30min o più e la distanza di lettura ravvicinata (meno di 30 cm). In
Tabella VII Paragone tra i punteggi dei test locali e nazionali, di diversi ambiti, tra i soggetti
miopi, emmetropi ipermetropi. Tra emmetropi e ipermetropi i paragoni non sono significativi. Tra
miopi e emmetropi sono significativi come sottolineato nella tabella !. Tra miopi e ipermetropi i
dati significativi sono sottolineati nella tabella^. I dati sono espressi in punteggi medi ± DS. [17].

27
conclusione, in questo studio, vengono associate alla miopia le ore trascorse
nella lettura continuata (>30 min) (debole correlazione) e la distanza ravvicinata
(correlazione significativa). [51].
A un anno dallo studio di IP, Lu prendendo in considerazione un campione di,
ragazzi con età media 14.7 anni, provenienti da un area rurale cinese, non trova
nessuna evidenza nella relazione tra maggior tempo trascorso in attività
prossimali e miopia. Trova, invece, associazione tra una minore distanza per le
attività prossimali e la miopia. L’associazione può essere interpretata in due
modi: come un’ulteriore evidenza dello sviluppo della miopia causata dal lavoro
prossimale, oppure che la distanza di lettura ridotta sia una conseguenza della
più corta lunghezza focale del sistema ottico, propria dei miopi non corretti da
lenti [52].
Anche studi su studenti universitari supportano la teoria che il lavoro prossimale
sostenuto influenzi la progressione e lo sviluppo miopico nel soggetto. Per
esempio Klinge et al. paragonano lo stato refrattivo tra gli studenti universitari in
Norvegia e la popolazione generale nei paesi della Scandinavia. La popolazione
generale mostra una prevalenza miopica significativamente più bassa (33%) nei
confronti degli studenti (47%) [53]. Oltre all’alta prevalenza di miopia tra gli
studenti universitari, i risultati di questi studi dimostrano che, mentre sono
impegnati in attività che richiedono periodi significativi di lavoro prossimale
concentrato (per esempio: 7ore alla settimana di lettura di articoli scentifici [54],
la maggior parte degli studenti (da 40% a 80%) mostra un peggioramento del
proprio errore refrattivo e diventa più miope [55] [56] [57] [58]. Basandosi su
queste investigazioni, Jiang et al. studiano un campione di studenti al primo anno
di optometria dividendoli in tre gruppi: soggetti miopi, ipermetropi ed
emmetropi. Dopo i primi nove mesi di studio l’errore refrattivo tra gli studenti
universitari miopi subisce un cambiamento significativo, per gli altri due gruppi le
variazioni refrattive non sono significative. La progressione miopica è continua
durante ogni semestre, ma cessa durante le vacanze invernali. Gli autori
confrontano anche sette diverse attività visive e il loro impiego nel primo

28
semestre e nelle vacanze invernali (lavoro in classe, lettura e scrittura, guardare
la televisione, lettura di piacere, attività all’aperto, attività all’interno e lavoro al
computer); tutte, eccetto il lavoro al computer, in numero di ore per giorno,
sono significativamente differenti tra il semestre e il periodo di vacanza. Durante
il periodo di studio gli studenti spendono 10.23 ± 2.26 ore al giorno per il lavoro
in classe, per la lettura e per la scrittura collegata allo studio accademico e per il
lavoro al computer, mentre in vacanza trascorrono solo 3.14 ± 3.37 ore al giorno
per queste attività correlate al lavoro prossimale. Al contrario per le attività quali
guardare la televisione, lettura di piacere, attività all’esterno e all’interno
durante il semestre le ore che utilizzano sono 4.15 ± 2.03 al giorno, mentre in
vacanza sono 8.56 ± 3.81 ore al giorno (tabella VIII).
Periodo dell’anno Attività prossimali legate allo studio
Attività all’aperto, ore di TV e lettura di piacere
Semestre di studio 10.23 ± 2.26 4.15 ± 2.03
Vacanze invernali/estive
3.14 ± 3.37 8.56 ± 3.81
Questi dati indicano chiaramente che la richiesta di lavoro prossimale cala
significativamente durante il periodo di vacanza. Tutto ciò conferma che il lavoro
prossimale è un importante fattore nello sviluppo miopico per gli studenti
universitari e suggerisce che la progressione miopica in questi studenti possa
essere interrotta da un semplice evento, come per esempio le vacanze [8].
Un altro aspetto da valutare come fattore dell’attività prossimale è la miopia
transiente indotta dal lavoro prossimale (nearwork-induced transient myopia,
NITM). Questo tipo di miopia si riferisce ai piccoli e momentanei cambiamenti
miopici in un punto lontano riscontrato dopo un periodo di lavoro prossimale
sostenuto. La NITM riflette un’inabilità della lente cristallina di ridurre il suo
potere appropriatamente e rapidamente, risultando in un fenomeno
Tabella VIII. Ore dedicate alle diverse attività dagli studenti nel
periodo di studio universitario e durante le vacanze estive e
invernali. I dati sono espressi in ore in media ± DS. [8].

29
effetto/isteri accomodativa. La miopia transiente può essere influenzata da vari
fattori: la durata del compito a distanza prossimale, errore refrattivo, età,
richiesta diottrica e fattori cognitivi. L’ampiezza della NITM iniziale e la durata del
decadimento sono due parametri chiave utilizzati per descrivere la risposta
accomodativa per un punto lontano dopo una mansione di lavoro prossimale
sostenuta. L’ampiezza iniziale della NITM è definita come la differenza diottrica
tra lo stato refrattivo immediato pre- e l’immediato post-compito prossimale. La
durata di decadimento rappresenta la quantità di tempo per questa risposta
accomodativa momentanea utile a ritornare ai livelli base prima del compito
prossimale.
Lin et al. nel 2012, prendono come campione bambini miopi, ipermetropi e
emmetropi di 7 anni per investigare alcuni aspetti della miopia transiente indotta
dal lavoro prossimale. Trovano che l’ampiezza della NITM iniziale è
significativamente e consistentemente più alta nel gruppo di bambini miopi
rispetto agli altri bambini ipermetropi e emmetropi (tabella IX) [59]. Risultati
simili sono stati trovati anche riguardo al tempo di decadimento considerando
tutte e tre i gruppi. Questi risultati concordano con uno studio precedente di
Wolffsohn et al. [60] che ricerca le caratteristiche della miopia transiente in 45
preadolescenti miopi ed emmetropi con età media di 7.5 anni. Viene riportato
che l’ampiezza della NITM iniziale è significativamente più elevata nei miopi
rispetto agli emmetropi; inoltre non risulta esserci una significativa riduzione
nella NITM durante l’intervallo di tre minuti dopo il compito prossimale. Questi
risultati suggeriscono che i preadolescenti miopi di Hong Kong sono più
suscettibili, rispetto ai loro compagni emmetropi, allo sfocamento indotto.
Questo è coerente con una ricerca precedente. Quest’ultima evidenzia che il 30%
dei bambini bianchi americani, tra i 4 e i 10 anni, mantengono un’elevata, e non
decaduta NITM, durante l’intervallo dei tre minuti, dopo la mansione di 5 minuti
di lavoro prossimale sostenuto eseguito binocularmente [61]. La differenza tra i
risultati riguardanti la miopia transiente sono collegati alla condizione visiva (test
eseguito monocularmente o binocularmente). Nel secondo studio viene utilizzata

30
la visione binoculare, nella quale la vergenza accomodativa indotta potrebbe
aver ridotto il generale sfocamento causato e, quindi, ridotto anche la NITM.
Anche se la NITM non è stata indotta in tutti gli studenti in ognuno dei tre gruppi
refrattivi, la più bassa porzione di NITM non indotta si trova nel gruppo dei miopi
paragonati agli emmetropi e ipermetropi. La percentuale di studenti che
esprimono l’incompleto decadimento della NITM è significativamente più alto
nei miopi rispetto ai coetanei ipermetropi e emmetropi (tabella IX). Perciò gli
occhi miopi dimostrano essere suscettibili all’errore refrattivo sia rispetto
all’ampiezza della NITM iniziale sia al tempo di decadimento di essa. Simili
risultati sono stati riportati in altri due studi, nei quali la miopia transiente è stata
testata in giovani adulti. Vera-Diaz et al. riportano che dopo 10 minuti della
mansione del lavoro prossimale in un gruppo di 13 miopi progressivi, con età tra i
18 e i 27 anni, la miopia transiente media non decade completamente ai valori
base durante l'intervallo di 2 minuti dopo il compito [62]. Più recentemente,
Vasudevan e Ciuffreda [63] testano la possibile conseguenza dell'additività della
miopia transiente e le caratteristiche del decadimento, caratteristica nei giovani
adulti miopi e emmetropi dopo due ore di lettura continuativa.
Approssimativamente il 35% dei miopi e il 33% degli emmetropi mostra un
aumento della miopia transiente con incompleto decadimento dopo la prima ora
di lettura. Queste percentuali aumentano bruscamente fino al 80% e al 60% dei
miopi e degli emmetropi, rispettivamente, dopo la seconda ora di lettura
continuativa. Questa tendenza comune tra miopi ed emmetropi trova valore
valutando cosa si intende per emmetropi, ossia soggetti con un errore refrattivo
-0.50D < ER≤ +0.50D.Le percentuali di studenti nel presente studio con
decadimento incompleto della NITM in entrambi i gruppi refrattivi, mostrano
simili tendenze agli studenti dello studio sopra citato, suggerendo che la NITM sia
un fenomeno accomodativo considerevole. Un’altra interessante scoperta dello
studio di Lin et al. è che circa il 30% degli ipermetropi mostra un decadimento
incompleto. La ragione di ciò rimane sconosciuta.

31
NITM Miopi Emmetropi Ipermetropi
Ampiezza NITM iniziale (D) 0.18 ± 0.16 0.9 ± 0.13 0.10 ± 0.19
Decadimento incompleto (%) 37.9 19.6 29.0
Tuttavia tutti gli ipermetropi presi in considerazione in questo studio sono
studenti della scuola primaria (8.4±1.1 anni di età media) con potenziale
predisposizione alla miopia, all’aumentare del livello scolastico e dell’aumento
del lavoro prossimale corrispondente. Una verifica della NITM e dell’errore
refrattivo nei seguenti due anni dovrebbe provare il cambiamento miopico
previsto precedentemente. Concludendo, l’incompleto decadimento della miopia
transiente sembra essere una caratteristica relativamente comune e un possibile
guadagno nella conoscenza della natura potenzialmente miopica nel futuro. Una
successiva valutazione di questi bambini nei prossimi 3 anni porterà alla
conoscenza del possibile ruolo della NITM nello sviluppo della miopia
permanente durante il periodo scolastico [59].
2.2.1 Lavoro prossimale e risposte accomodative e del sistema di vergenza.
Nel lavoro prossimale un aspetto fondamentale è il processo
dell’accomodazione. Le attività da vicino, infatti, richiedono l’attivazione
dell’accomodazione e del sistema di vergenza per raggiungere una chiara e
singola visione binoculare. Numerosi studi hanno dimostrato che una fissazione
prossimale sostenuta induce adattamento dell’accomodazione e dei sistemi di
vergenza. Nel 2002 M. Rosenfield studia le risposte accomodative di target a
distanza prossimale in 23 giovani (età media 23.0 anni) emmetropi, miopi e un
ipermetrope, nessuno dei soggetti presenta astigmatismo superiore alle 0.75D. I
risultati non portano ad una significativa correlazione tra il gradiente richiesta-
risposta accomodativa né della progressione miopica o, complessivamente,
cambi nell’errore refrattivo. L’unica significativa evidenza è un basso gradiente
stimolo-responso accomodativo nei miopi stabili. Anche se il periodo di
Tabella IX. Valori della miopia transiente indotta dal lavoro prossimale nei bambini
miopi, emmetropi e ipermetropi. I valori dell’ampiezza iniziale di NITM sono espressi in
diottrie in media ± DS [59].

32
sperimentazione è relativamente breve, questi risultati suggeriscono che prima
dello sviluppo della miopia non si verificano cambiamenti nel gradiente. Quando
vengono presentati i marcatori prossimali, convergenti, d’induzione dello
sfocamento per l’accomodazione, si evidenzia che ogni variazione che si verifica
nel gradiente stimolo-responso possa essere in funzione dell’essere miope,
piuttosto che rappresentare uno stimolo per un possibile sviluppo di un errore
refrattivo. Emerge anche che lo sviluppo della miopia non è associato a un
aumento del lag accomodativo. Infatti, i soggetti che mostrano una elevata
progressione miopica hanno più piccoli lag di accomodazione. Di nuovo, questo
sta ad indicare che il cambiamento della risposta accomodativa è influenzato
dalla miopia. In conclusione dai dati di questo studio possiamo dedurre che
quando tutti i marcatori per l’accomodazione vengono presentati, l’ampiezza
dell’errore accomodativo è indipendente dallo sviluppo dell’errore refrattivo. I
dati non supportano, quindi, la proposta che lo sviluppo della miopia in giovani
adulti sia accompagnata da una risposta accomodativa ridotta durante il lavoro
prossimale [64].
Un altro studio indaga, in un periodo 12 mesi, se le anomalie accomodative sono
collegate all’errore refrattivo, all’età di inizio della miopia o alla progressione
della miopia, misurando un’ampia gamma di funzioni accomodative, in soggetti
miopi e non miopi, per determinare se esistano differenze tra i due gruppi.
Esamina anche la correlazione tra le varie funzioni accomodative per vedere se
queste sono co-dipendenti, e, mediante un modello a regressione multipla per
vedere se le funzioni accomodative sono influenzate dalla progressione della
miopia. Emerge una correlazione significativamente positiva tra l’errore
refrattivo e l’ampiezza accomodativa, accomodazione attraverso foro stenopeico
e il lag accomodativo binoculare a 33 cm. I non miopi mostrano in media delle
ampiezze accomodative significativamente più elevate dei miopi. Le due migliori
funzioni accomodative indipendenti che fungono da predittori della progressione
miopica sono il lag binoculare di accomodazione in distanza prossimale (33cm) e
la facilità accomodativa monoculare prossimale (tabella X).

33
Soggetti Facilità accomodativa (risposta positiva)
(cicli/minuto)
Ampiezza accomodativa
(D)
Monoculare Binoculare
Miopi 2.53 ± 3.21 2.17 ± 1.08 8.12 ± 0.96
Emmetropi 1.86 ± 0.96 1.77 ± 0.81 8.88 ± 1.01
Tali risultati sono in accordo con studi passati che dimostrano un aumento del lag
accomodativo negli individui con miopia progressiva. Il tempo di risposta positiva
nella facilità accomodativa è più lungo nei partecipanti che diventano più miopi. I
ritardi nell’ottenere l’immagine a fuoco, quando cambia la fissazione da lontano
a vicino, possono portare a brevi periodi di defocus ipermetropico. Se questo
defocus retinico si accumula in aggiunta al defocus che si presenta durante
l’esecuzione del test, allora possono verificarsi periodi considerevoli di defocus. Il
defocus accumulato dovrebbe essere massimo nelle persone che trascorrono più
tempo durante il giorno cambiando continuamente punti di fissazione, per
esempio tra la lettura e un punto visuale più distante, come succede all’interno
della classe agli studenti. Si conferma quindi che il lag accomodativo è un
significante predittore di progressione miopica. Riassumendo, le due funzioni
accomodative chiave, che differenziano gli individui con un errore refrattivo
stabile e individui che mostrano un aumento di esso verso la miopia, sono la
facilità accomodativa e il lag accomodativo [65].
Nel 2006 Harb et al., prendendo in considerazione un campione di adulti miopi
ed emmetropi non trovano nessuna differenza statisticamente rilevante tra i lag
accomodativi dei due gruppi, anche dopo un periodo di 2-3 min di lettura
prossimale sostenuta. I miopi adulti mostrano una maggiore variabilità della
risposta accomodativa, durante periodi estesi di lettura in termini sia della
deviazione standard della risposta accomodativa sia del potere delle
microfluttuazioni accomodative. In conclusione in questo ricerca emerge che nei
soggetti adulti con miopia stabile, la media dell’ampiezza accomodativa della
Tabella X. Valori delle funzioni accomodative significativamente diverse tra i soggetti miopi
e emmetropi. I dati della facilità accomodativa sono espressi in media dei cicli al minuto ±
DS. I dati dell’ampiezza accomodativa sono espressi in media delle diottrie ± DS. [65].

34
risposta, durante un periodo esteso di lettura, è virtualmente identico a quello
degli adulti emmetropi. Tuttavia, trovano più variabilità accomodativa nei miopi,
concordemente con l’ipotesi che i miopi siano meno sensibili al defocus. Inoltre
le piccole fluttuazioni nella risposta accomodativa può essere individualmente
troppo piccole per stimolare la crescita del bulbo oculare e l’integrazione nel
tempo delle fluttuazioni durante periodi di lettura sostenuta possa essere
sufficiente per produrre un segnale sfocato che induce miopia [46].
Similmente, nel 2008 Langaas et al. non trovano nessuna differenza nel lag
accomodativo tra i giovani miopi e quelli non miopi, mentre, tra i due gruppi
emergono differenze significative nella variabilità della risposta accomodativa.
Quest’ultima aumenta all’aumentare del grado di miopia. Il gruppo di bambini
emmetropi mostra un aumento lineare nella variabilità della risposta
accomodativa con l’aumento della domanda dei target; invece i bambini miopi
dimostrano di avere un’ampia variabilità nella risposta accomodativa al target di
0.25D, nessun aumento nella variabilità al target da 2D e un più grande aumento,
e un più grande aumento al target da 4D, rispetto al gruppo emmetrope [66]. Lo
stesso pattern della variabilità della risposta accomodativa viene seguito da
soggetti adulti emmetropi e miopi presi in considerazione nello studio di Day et
al., in particolare il modello di variabilità degli adulti con miopia tardiva è simile a
quello dei bambini con miopia precoce, questo porta a supportare un ruolo di un
aumento della variabilità della risposta accomodativa nell’eziologia di entrambe
le miopie: precoce e tardiva [67],poiché l’aumento di essa risulta in un’immagine
retinica sfuocata quando viene integrata nel tempo. Questo potrebbe causare il
fallimento nell’emmetropizzazione producendo un significativo allungamento del
bulbo oculare [68]. Tuttavia, è possibile anche che l’aumento della variabilità sia
prodotto dalla miopia come dimostrato per il lag accomodativo sia nella miopia
sia tardiva sia in quella precoce [69] [70].

35
3. Fattori genetici: la miopia come malattia multifattoriale.
3.1 Breve introduzione ai concetti chiave della genetica di malattie
complesse.
3.1.1 Genotipo e fenotipo.
Il genotipo di un individuo può essere definito come l’insieme delle
caratteristiche genetiche di un individuo. Un gene è definibile come una regione
del genoma (locus ovvero una sequenza di DNA) che codifica per un prodotto
proteico o per un RNA funzionale. Sequenze diverse del gene, le varianti
alleliche, loci allelici o alleli, possono essere osservate in diversi individui di una
specia. Le varianti alleliche possono determinare differenze nel fenotipo (dal
greco phainein, che significa "apparire", e týpos, che significa "impronta"). Un
carattere ereditario, ovvero una caratteristica osservabile del fenotipo di un
individuo, può essere ereditato nelle generazioni in modo Mendeliano o in modo
più complesso. Un carattere determinato da un singolo gene si dice Mendeliano
(monogenico), mentre se è dipendente da più geni si dice poligenico. Un tratto
poligenico riflette l’attività di più di un gene. I caratteri Mendeliani possono
essere riconosciuti dalle tipiche modalità di trasmissione negli alberi genealogici.
Un esempio delle modalità di trasmissione dei caratteri mendeliani negli alberi
genealogici può essere la malattia Huntington che è una patologia a ereditarietà
autosomica dominante.
Figura 4. Albero
genealogico
della malattia di
Huntington.
(http://upload.wi
kimedia.org/wiki
pedia/commons/
thumb/e/e0/Auto
somal_Dominant
_Pedigree_Char
t2.svg/513px-
Autosomal_Dom
inant_Pedigree_
Chart2.svg.png).

36
L’albero genealogico mostrato in figura 4 illustra il modello di ereditarietà della
malattia: ogni discendente di un individuo affetto avrà un rischio del 50% di
ereditare l'allele mutante e quindi di essere colpito dalla malattia. Questa
probabilità è indipendente dal sesso e il fenotipo “non salta” generazioni.
La maggior parte dei caratteri determinati geneticamente nell’uomo non si
comporta in modo mendeliano poiché controllata da più loci genici. I caratteri
poligenici comprendono caratteri determinati da due o pochi geni con un effetto
importante (oligenici) e caratteri sotto il controllo di numerosi geni diversi,
ciascuno dei quali contribuisce al fenotipo in modo più o meno decisivo. Sia i
caratteri monogenici sia quelli poligenici possono anche essere multifattoriali,
ovverosia influenzati anche dall’ambiente. I caratteri poligenici non influenzati
dall’ambiente si dicono poligenici puri e sono molto rari: ad esempio, il colore
degli occhi è vicino a essere puramente poligenico. La maggior parte dei caratteri
comuni come l’altezza, il colore della pelle, il peso corporeo sono caratteri
poligenici multifattoriali. La predisposizione alla maggior parte delle malattie più
diffuse, quali cancro, diabete e malattie mentali, così come alcune tendenze
comportamentali hanno ereditarietà multifattoriale. Nel caso dei caratteri
multifattoriali, la relazione tra genotipo e ambiente può essere molto complessa,
e il contributo al fenotipo di un singolo allele di un gene è più difficile da
identificare e prevedere, a causa delle azioni combinate degli altri geni e
dell’ambiente. Una condizione poligenica multifattoriale può riflettere i
contributi additivi di diversi geni. Come già accennato, ogni gene conferisce un
certo grado di suscettibilità, ma l’influenza di diversi geni non è necessariamente
di ugual peso. Inoltre gli effetti di più geni possono non essere semplicemente
additivi, ma prevedere delle sinergie più complesse.
I caratteri monogenici sono qualitativi o discreti, e spesso forniscono un fenotipo
“tutto o niente”, così come “normale” rispetto ad “affetto”. Per un tratto
poligenico, l’azione combinata di molti geni produce spesso una “variazione
continua” del fenotipo, che viene indicato anche come carattere quantitativo. In
alcuni casi, la presenza del fenotipo qualitativo (ad es. labiopalatoschisi, “labbro

37
leporino” o palato normale) è probabilmente dovuta ad un effetto soglia, oltre il
quale si manifesta il fenotipo. Le sequenze di DNA che contribuiscono ai caratteri
poligenici sono chiamate loci di caratteri quantitativi o continui, o QTL (da
Quantitative Trait Locus). Se un carattere mostra una variazione continua
generalmente è anche poligenico. In altre parole, è la componente multigenica
del carattere che contribuisce alla variazione continua del fenotipo. Presi
singolarmente, i geni che conferiscono un carattere poligenico a un individuo
seguono le leggi di Mendel ma, nel loro insieme, non producono rapporti
fenotipici tipici di un singolo gene. Essi sono tutti fattori che contribuiscono al
fenotipo, senza che relazioni di dominanza e recessività tra i diversi alleli di
ciascun gene siano facilmente riconoscibili. Alcuni geni contribuiscono più di altri
ad un carattere poligenico. Considerando un singolo gene, diversi alleli possono
inoltre avere effetti diversi sul fenotipo a seconda di come modificano
esattamente una proteina codificata, le caratteristiche funzionali o l’espressione
di questa.
Il tipo di variabilità riscontrata nella popolazione differenzia in modo sostanziale i
caratteri quantitativi da quelli qualitativi: mentre i fenotipi di caratteri
mendeliani possono essere facilmente raggruppati in classi distinte sulla base di
un solo criterio di discriminazione (variabilità discontinua), nel caso di caratteri
quantitativi ciò non è possibile perché fenotipi si distribuiscono entro un certo
ambito di valori-limite senza soluzione di continuità (variabilità continua), tanto
che l’osservatore è costretto ad utilizzare criteri arbitrari di discriminazione per
procedere alla classificazione. La maggior parte delle misure di caratteri
quantitativi presenta distribuzioni rapportabili a Gaussiane, per cui possono
essere descritte usando i parametri tipici delle distribuzioni normali.
I caratteri quantitativi presentano, dunque, una variabilità continua, tuttavia alla
base di questa variabilità sta la variabilità discontinua generata dalla
segregazione di ogni singolo locus. Aumentando il numero di loci aumenta anche
il valore di classi genotipiche. Le differenze di valore fenotipico tra le diverse
classi diminuiscono con l’aumentare del numero di loci implicati nella

38
determinazione del carattere. Inoltre, la variabilità dovuta a cause non genetiche
ma ambientali contribuisce ad annullare eventuali discontinuità tra i fenotipi
associati ai diversi genotipi (combinazione di alleli in diversi geni), rendendo
spesso molto difficile l’analisi dei determinanti genetici dei caratteri quantitativi
[71].
3.1.2 Valore fenotipico.
La misura del carattere quantitativo viene definita “valore fenotipico”. Esso è
determinato dall’azione di due componenti: il genotipo (G), che rappresenta un
particolare assortimento di geni posseduto da un individuo; l’ambiente (E) che è
l’insieme delle circostanze non-genetiche in grado di influenzare l’espressione
del carattere. Le due componenti, allora, possono agire additivamente nel senso
che l’espressione fenotipica è la somma del genotipo e delle deviazioni casuali
che l’ambiente esercita sul carattere in esame. L’Equazione fondamentale della
genetica quantitativa è pertanto:
in cui P: il fenotipo; G: valore genotipico; E: deviazione che l’ambiente esercita
sull’espressione del carattere.
Poiché le deviazioni ambientali calcolate su tutta la popolazione animale sono
distribuite normalmente e, in quanto casuali, la loro somma è zero, la media del
valore fenotipico in una popolazione è uguale alla media del valore genotipico; in
altri termini per una popolazione animale abbastanza numerosa, il fenotipo
medio è uguale al genotipo medio e quindi (P = G).
µ : media delle condizioni ambientali comuni al gruppo di animali cui l’individuo i
appartiene;
Gi: il valore genotipico dell’individuo i;
Ei: rappresenta l’insieme degli effetti ambientali (E = environment) che hanno
influenzato il fenotipo dell’animale [72]. Assumendo per semplicità che non vi

39
siano interazioni genotipo/ambiente la quota di origine genetica della varianza
fenotipica osservata in una popolazione, viene definita ereditabilità (h2):
Il valore h2 può variare da 0 a 1; il valore h2 sarà ovviamente tanto maggiore
quanto più rilevante sarà la varianza di origine genetica rispetto alla varianza
fenotipica globale. Quando h2=1, il carattere è determinato geneticamente in
modo stretto e non presenta variabilità dovuta a effetti ambientali. Dall’altra
parte, se le differenze tra gli ambienti cui gli individui sono esposti tendono a
zero (ad es. in un allevamento di animali in condizioni standard e controllate)
l’ereditabilità dei caratteri sarà massima. L’ereditabilità stima la percentuale di
variazione fenotipica per un carattere particolare che è dovuta a differenze
genetiche di una certa popolazione in un determinato momento. Pur essendo le
stime di ereditabilità estremamente utili esse non possono avere valore assoluto,
ma al massimo valore relativo alla popolazione e all’ambiente in cui lo studio è
stato condotto.
3.1.3 Penetranza e espressività.
Per penetranza s’intende la frequenza con cui individui di un dato genotipo
manifestano il carattere a livello fenotipico. Teoricamente un carattere con
eredità dominante dovrebbe manifestarsi in tutti gli individui nel cui genotipo è
presente l’allele dominante in questione. Al contrario in alcune malattie
ereditarie si nota una penetranza incompleta o ridotta. La causa di tale
fenomeno può essere ricercata in un effetto “ambientale”, ma più spesso deve
essere attribuita allo sfondo genetico, costituito dai molti geni ancora ignoti che,
con le loro varianti, possono influire positivamente o negativamente sulla
manifestazione del genotipo nel fenotipo.
L’espressività si riferisce al grado con cui un dato genotipo si manifesta a livello
fenotipico. Si distinguono caratteri con espressività costante, qualora la
manifestazione sia molto simile in tutti gli individui che mostrano il carattere, e

40
caratteri con espressività variabile se invece esiste una forte variabilità nella
manifestazione.
3.1.4 Malattie complesse e studi familiari.
Per poter analizzare i fattori genetici coinvolti nelle malattie complesse comuni è
necessario stabilire i criteri tramite i quali le persone possono essere classificate
come affette o non affette. Nel caso di sindromi mendeliane è abbastanza ovvio
stabilire quali caratteristiche di un paziente facciano parte della sindrome e quali
invece siano solo delle coincidenze. Nel caso di malattie non mendeliane, invece,
le cose sono definite molto meno chiaramente. Una volta stabiliti dei criteri
chiari diagnostici per una malattia complessa, è necessario verificare se la
genetica abbia un qualche ruolo nell’eziologia della malattia. Sotto quest’ipotesi
maggiore è la quota di DNA che due persone hanno in comune, più alta
dovrebbe essere la probabilità che queste due persone condividano anche il
fenotipo in questione. La strada più ovvia per affrontare questo problema è
quella di dimostrare che il carattere si trasmetta all’interno di famiglie e questo
comporta studi di familiarità, oppure sui gemelli e su individui adottati.
Il grado con cui una malattia si riscontra e ricorre nelle famiglie può essere
espresso dal rapporto di rischio (R) ovvero il rischio che un familiare (R) di un
probando malato ha di presentare la malattia, in confronto al rischio nella
popolazione generale. Un rischio di 1 indica che non vi è alcun rischio aggiuntivo
oltre a quello che caratterizza la popolazione; valori di R superiori a 1 mostrano
la ricorrenza familiare della malattia. Il rischio dipende dal grado di parentela con
il probando e generalmente i valori tendono a 1 per i parenti più lontani del
malato, come nipoti e cugini.
Non bisogna dimenticare che le persone forniscono ai loro figli anche l’ambiente,
oltre che ai geni. Molti caratteri si manifestano all’interno delle famiglie in
quanto i membri della famiglia condividono lo stesso ambiente. Ci si dovrebbe
sempre chiedere se la condivisione dello stesso ambiente può essere la ragione
di familiarità di un carattere. Questa considerazione diventa ancora più

41
importante nel caso di caratteristiche comportamentali, come il cosiddetto
quoziente d’intelligenza (QI) o per le malattie mentali.
L’analisi della segregazione è uno strumento statistico per analizzare
l’ereditarietà di qualsiasi carattere, può fornire prove a favore o contro il
coinvolgimento di un locus principale di suscettibilità e permette, almeno in
parte, di definirne le proprietà. I risultati di questo tipo di analisi possono essere
utili per indirizzare studi futuri di linkage o di associazione. Gli studi di linkage
analizzano i soggetti appartenenti alla stessa famiglia, gemelli omozigoti ed
eterozigoti basandosi sul concetto “identity by descent”. Altri studi si avvalgono
di soggetti adottati con genitori biologici ed adottivi inclusi nello studio.
Un approccio diverso e molto in voga è utilizzato dagli studi di associazione.
Questi si basano sul concetto di “identity by status” ovvero sul fatto che individui
non imparentati di una popolazione possano condividere alcune caratteristiche
genotipiche in quanto condividono gli stessi alleli nel gene o nei geni che
determinano il fenotipo (figura 5).
Figura 5. Le analisi di linkage testano la co-segregazione degli alleli all’interno
dei membri della stessa famiglia mentre le analisi per associazione ricercano le
differenze nelle frequenze alleliche tra individui non imparentati affetti e non
affetti [180].

42
3.2 I componenti oculari refrattivi, in particolare LA, sono caratteri
geneticamente determinati?
La refrazione è dovuta alla coordinazione di componenti oculari biometrici come
la lunghezza assiale, la profondità della camera anteriore, la curvatura corneale e
lo spessore della lente (figura 6). La lunghezza assiale (LA) è considerata come la
maggiore determinante dell’errore refrattivo. L’ereditabilità della LA è stimata in
un intervallo tra il 40 e il 94% [12]. La miopia e gli altri errori refrattivi sono le
conseguenze di una disarmonia errata dei componenti e strutture oculari.
In uno studio longitudinale Sorsby [73] raccolse i dati refrattivi di 68 bambini di
età tra i 3 e gli 8 anni alle loro prime visite e li rivalutò la seconda volta dopo 6.5
anni in media. I bambini vennero suddivisi in due gruppi: nel primo emerse uno
sviluppo normale o vicino all’emmetropia; mentre il secondo mostrò segni di
miopia. Nel primo gruppo si evidenziò uno stabile decremento di ipermetropia
(media di 0.09D per anno) nei 6.5 anni e un aumento della lunghezza assiale
(media= 0.14mm per anno). Nel secondo gruppo invece, si verifica un maggiore
decremento di ipermetropia (media=0.38D per anno) e un maggiore incremento
della lunghezza assiale (media=0.24D per anno) che risulta essere quasi il doppio
rispetto al primo gruppo. Molti altri studi riportano la correlazione tra LA e
miopia, ovvero tanto maggiore la lunghezza assiale tanto maggiori sono la
Figura 6. Illustrazione della
lunghezza assiale LA. PCA=
profondità della camera
anteriore; SL= spessore della
lente; PCV= profondità della
camera vitrea.

43
severità e il grado della miopia. Un aumento di 1mm di LA senza altre
compensazioni è equivalente ad un cambiamento miopico di -2 o -2.5D.
La lunghezza assiale viene analizzata come fattore poligenico e come un tratto
quantitativo. La previsione di tale parametro per poter anticipare un possibile
sviluppo di miopia, si ha solamente nei 2-4 anni che precedono l’inizio dell’errore
refrattivo in questione. Si nota infatti che LA raggiunge il suo valore di maggior
variazione nell’anno precedente al principio della miopia, mentre le variazioni
successive all’esordio della miopia sono più limitate [74].
Diverse evidenze supportano la forte componente genetica nella determinazione
di LA. Bambini con genitori miopi presentano un LA più lungo rispetto ai bambini
che non hanno genitori miopi [75]. Molti studi, che hanno analizzato la
componente genetica della miopia e della LA, hanno fornito stime
dell’ereditabilità dell’errore refrattivo notevolmente variabili. Le variazioni
derivano dai diversi gradi di parentela, considerate tra i soggetti, ovvero i valori
risultano differenti negli studi tra gemelli e quelli tra genitore e figlio. La varianza
di LA causata da fattori ambientali risulta essere solo del 6% circa contrastando
quella della miopia tra il 14% e il 33% (tabella XI) [10] [76].
Tratto Ereditabilità Studio
Gemelli Fratelli Nuclei familiari
LA 0.88 0.73 0.75 [11]
Miopia 0.82 0.50 0.21 [10]
Uno studio, con campione dei bambini cinesi di età tra i 5 e i 16 anni, rivela che la
lunghezza assiale è maggiore nei bambini con due genitori miopi (media
23.65mm) seguita da quella dei bambini con un solo genitore miope (media
23.51mm) e, infine, la lunghezza assiale dei bambini con nessuno dei due genitori
miope (media 23.47mm). I bambini con due genitori miopi tendono ad avere
anche la camera vitrea più profonda, mentre i valori della camera anteriore e
dello spessore della lente sono simili tra i bambini con le diverse storie familiari.
Tabella XI. Ereditabilità paragonata tra LA (lunghezza assiale) e
miopia.

44
Nel sottogruppo di bambini pre-miopi, quelli che non hanno nessuno dei due
genitori miope hanno LA più lungo (media=23.15 mm), ma una minore crescita
(media=0.20 mm per anno) rispetto sia ai bambini con due genitori miopi
(media=23.11 mm e crescita media di 0.37 mm per anno) che a quelli con un solo
genitore miope (media=23.07 mm e crescita media di 0.26 mm per anno) (tabella
XII).
Genitore/i miope/i (num.)
LA in mm Crescita annuale in mm
Nessuno 23.15 0.20
Uno 23.07 0.37
Due 23.11 0.25
In conclusione, in questo studio la storia refrattiva familiare mostra avere una
correlazione positiva con la lunghezza assiale e il suo cambiamento nei bambini
non miopi, in quanto bambini pre-miopi con due genitori miopi mostrano avere
una maggiore crescita annua della lunghezza assiale rispetto agli altri bambini
[14].
Diversi studi su gemelli supportano l’importanza della componente genetica
nell’errore refrattivo e nel determinare la LA. Ne è esempio lo studio di Dirani e
colleghi del 2008, in cui gemelli di entrambi i sessi vengono esaminati per
determinare in quale proporzione l’associazione tra LA e miopia sia dovuta a
fattori genetici o fattori ambientali. Si evidenzia che, come già visto in
precedenza, il 50% della varianza dell’equivalente sferico è spiegato dalla forte
correlazione negativa tra errore refrattivo e LA [77] [78] [79] [80]. Questo
suggerisce che LA sia uno dei maggiori determinanti dell’errore refrattivo.
L’ereditabilità stimata dell’equivalente sferico risulta essere del 75%-88%, e
concorda con studi precedenti [81]. Viene anche evidenziata una forte base
genetica della LA, confermando studi precedenti di gemelli [82] [83] [84]. In
conclusione da questo studio emerge che una grande proporzione della relazione
Tabella XII. Confronto tra il numero di genitori miopi e la lunghezza
assiale, con relativa crescita annuale. LA e crescita annuale sono
dati espressi in media. [92]

45
tra equivalente sferico e lunghezza assiale è spiegata da effetti genetici per
entrambi i sessi, mentre in piccola parte sembra essere dovuta da fattori
ambientali unici [76].
3.3 Ricorrenza familiare degli errori refrattivi e loro ereditabilità.
Il contributo dei determinanti genetici nello sviluppo della miopia e degli errori
refrattivi è stato ipotizzato da oltre un secolo grazie ad osservazioni riguardanti
numerosi studi su gemelli. L’evidenza del ruolo della genetica nello sviluppo delle
miopia è emerso anche negli studi su soggetti appartenenti alla stessa famiglia.
Sebbene il fattore ereditarietà sia ormai noto essere importante nella refrazione
oculare, l’ambiente e il comportamento giocano un ruolo di rilievo nello sviluppo
refrattivo anche se le dinamiche non sono ancora chiare. Nel tentativo di
determinare il ruolo di fattori genetici nello sviluppo della miopia Pacella et al.
nel 1999 esaminano la genealogia di 43 soggetti con miopia giovanile,
utilizzando i dati raccolti da un campione di soggetti giovani, partecipanti ad uno
studio longitudinale durato 24 anni. Ne emerge che avere due genitori miopi è il
fattore predittivo per lo sviluppo della miopia nel periodo scolastico, fornendo
l’evidenza di una componente genetica nell’eziologia della miopia. Inoltre, si può
evidenziare che livelli moderati di miopia giovanile possono essere ereditati
come un fattore complesso, che coinvolge fattori sia genetici sia ambientali.
Queste scoperte sono in accordo con le evidenze emerse in “The Orinda
longitudinal study of myopia” [85]: i fattori genetici giocano un ruolo
nell’eziologia della miopia, anche se i fattori ambientali sono comunque
importanti. E’ possibile che, essendo un tratto complesso, la suscettibilità alla
miopia possa essere dovuta a più di un gene. I geni possono predisporre un
individuo alla miopia e fattori ambientali, quali lavoro prossimale, possono
attivare l’espressione di tale condizione. Il risultato del 30% di bambini con
nessuno dei due genitori miopi che effettivamente diventa miope per l’età di 15
anni è in contrasto con l’ipotesi che la miopia segua le leggi classiche mendeliane
di ereditabilità, ovvero una modalità di trasmissione autosomica dominante.

46
Infatti con questa modalità di trasmissione ci si aspetterebbe che i soggetti di
ogni generazione fossero affetti. I risultati di questo studio non mostrano tale
schema e anche i dati dell’analisi genealogica sono inconsistenti con il modello di
ereditarietà autosomica dominante con penetranza completa. Tutti questi
risultati sulla natura genetica della miopia sono largamente empirici [13].
Studi sull’ereditabilità, basati sullo studio di gemelli ed individui della stessa
famiglia, hanno riportato una stima in percentuale di ereditabilità elevata,
variabile tra il 50% e il 90%. Infatti attraverso l’utilizzo di essi può essere
dimostrato che gemelli identici o monozigoti (MZ), i quali condividono il 100% dei
loro geni, presentano stati refrattivi più simili paragonati ai gemelli non identici o
dizigoti (DZ), che condividono fino al 50% dei loro geni. In questo modo si può
stimare la relativa importanza dei fattori genetici e ambientali paragonando
gemelli MZ con gemelli DZ per la concordanza della malattia per tratti qualitativi
o per la correlazione per tratti quantitativi. I gemelli sono descritti come
concordanti se entrambi mostrano il tratto identico, e discordanti se solo uno dei
due mostra il carattere. Una concordanza per un carattere che sia
significativamente maggiore nei gemelli MZ che nei gemelli DZ è una prova della
base genetica per quel tratto. Bisogna notare che gli studi basati sui classici
modelli sui gemelli hanno un potenziale errore verso il “trovare effetti genetici”
in quanto il modello assume che la correlazione degli effetti ambientali condivisa
all’interno della coppia di gemelli non dipenda dal tipo di gemelli, e
conseguentemente ogni eccessiva correlazione in gemelli MZ paragonata a
quella dei gemelli DZ è attribuita solo a fattori genetici e non a fattori ambientali
condivisi dai gemelli MZ come quelli relativi alla vita in utero. Questo problema
può risultare in una sovrastima del contributo genetico nella variazione
dell’equivalente sferico come in altri tratti oculari biometrici (LA, curvatura
corneale e la profondità della camera anteriore). Un altro studio sui gemelli
prende come campione 226 coppie di gemelle monozigoti e 280 coppie di
gemelle con un età media di 62.4 e 62.1 anni per MZ e DZ rispettivamente; è
emerso che l’effetto additivo genetico è responsabile del 86% della variazione

47
dell’equivalente sferico. Il fattore genetico sembra dare il contributo maggiore
nella variabilità degli errori refrattivi nella popolazione [86].
Molti studi confermano che genitori miopi tendono ad avere figli miopi. La
prevalenza di miopia nei bambini con due genitori miopi è del 30- 40%,
diminuendo nei bambini con un genitore miope ( 20-25%) e meno del 10% nei
bambini con nessuno dei due genitori miope [87] [88] [89]. Nello studio di Wu,
del ’99, vengono analizzati soggetti della stessa famiglia appartenenti a tre
generazioni diverse (bambino= 3^generazione, genitore= 2^ generazione e
nonno= 1^ generazione); tutte e tre le generazioni vengono suddivise in due
sottogruppi: miopi (≥-1.00D) e non miopi. La percentuale di miopia è 5.8%, 20.8%
e 26.2% nella prima, seconda e terza generazione rispettivamente (tabella XIII).
Generazione Miopia in %
Bambino (3^) 26.2
Genitore (2^) 20.8
Nonno (1^) 5.8
Si nota che la probabilità di essere miope nella terza generazione e sei volte più
alta rispetto alle altre due generazioni. In generale nella terza generazione la
probabilità di diventare miope aumenta con l’aumentare del numero di genitori
miopi. Nella seconda generazione si nota lo stesso meccanismo anche se i valori
risultano più bassi rispetto a quelli della terza generazione. Queste grandi
differenze nei valori devono essere dovute a fattori ambientali e non a fattori
genetici, in quanto non sarebbe possibile un cambiamento così veloce in così
poco tempo nella componente genetica dell’errore refrattivo. Questo studio
sottolinea la maggiore possibilità di sviluppare miopia se entrambi i genitori sono
miopi rispetto a bambini con un solo genitore miope o con nessun genitore
miope. Ne risulta anche che la miopia debba essere considerata come un tratto
poligenico multifattoriale nel quale l’input genetico è rimasto più o meno lo
stesso nelle tre generazioni, mentre è aumentata l’influenza ambientale dallo
Tabella XIII. Confronto tra le percentuali di
soggetti miopi tra tre generazioni. [90].

48
zero nella prima generazione ad un livello molto significativo nella terza
generazione, con la seconda generazione come via di mezzo [90].
Uno studio più recente indaga la relazione tra la storia dei genitori e la miopia
giovanile precoce, in bambini che sono diventati miopi e bambini che rimangono
non miopi durante la durata dello studio (dal I al VIII anno di scuola). Nei bambini
rimasti non miopi il 48% ha la madre miope, mentre nei bambini che hanno
sviluppato la miopia la percentuale di madri miopi è 69%; risultati simili si
ottengono considerando i padri miopi e non. Il numero dei genitori miopi, quindi,
è fortemente associato al possibile sviluppo futuro della miopia del figlio. Infatti
tra i bambini rimasti non miopi solo il 21% ha due genitori miopi, mentre nel
gruppo dei bambini che hanno sviluppato la miopia il 45% ha due genitori miopi.
Paragonato con il gruppo di riferimento (bambini con nessuno dei due genitori
miopi) i valori di probabilità per la futura miopia dei bambini con uno e due
genitori miopi sono significativi. Un altro aspetto su cui vanno ad indagare è se la
storia refrattiva dei genitori e le ore spese a praticare sport all’esterno sono in
qualche modo relazionabili tra di loro. Il risultato è che meno ore di sport o
attività all’esterno aumentano la probabilità di sviluppare miopia nei bambini con
due genitori miopi, rispetto ai bambini con uno o nessuno dei genitori miope. La
probabilità di diventare miopi per i bambini con nessuno dei due genitori miopi,
paragonata con quella dei bambini con uno o due genitori miopi, appare più
bassa nei bambini con la più alta quantità di ore trascorse praticando sport e
attività all’aperto. Da questo studio si può concludere che la storia refrattiva dei
genitori è un importante predittore nel possibile sviluppo di miopia del figlio, con
un effetto accentuato considerando come altra variabile le ore di sport o di
attività all’aperto per settimana [3].
Un altro esempio dell’importanza della storia refrattiva dei genitori rispetto
all’errore refrattivo del figlio, si ha in uno studio tra australiani di 12 anni da cui
emerge che i giovani con due genitori miopi hanno il più alto grado negativo di
errore refrattivo e la più lunga lunghezza assiale(aggiustando per fattori
ambientali e demografici) [91]. Un altro studio svolto su giovani cinesi di 5-16

49
anni residenti a Hong Kong concorda con i risultati descritti nel precedente
studio, ma, diversamente dai bambini australiani, evidenzia che storia refrattiva
dei genitori non è significativamente associata con una maggiore velocità di
crescita oculare e cambiamento miopico nel tempo [92]. Anche se una maggiore
severità della miopia del genitore aumenta il rischio di sviluppare miopia nel
figlio, l’impatto della miopia del genitore sulla miopia elevata nei bambini rimane
ancora indeterminato. Da tutti questi studi si evince che la probabilità in un
bambino di sviluppare miopia aumenta all’aumentare del numero dei genitori
miopi. Ciò sottolinea, dunque, la forte componente genetica nell’errore
refrattivo.
3.4 Geni candidati per la miopia.
La scoperta dei possibili geni implicati in malattie complesse come la miopia che
risultano da una componente sia genetica che ambientale si è dimostrata
difficile. Un approccio di studio è quello della Systems Genetics, utilizzato per
comprendere il flusso delle informazioni biologiche che determinano i tratti
complessi. Gli studi che hanno utilizzato questo tipo di metodo hanno fornito la
prima visione globale dell’architettura molecolare sottostante a caratteri
complessi e sono utili per l’identificazione dei geni, delle pathway e i network che
stanno alla base di malattie umane comuni. L’identificazione dei geni e delle
varianti genetiche responsabili di molte malattie monogeniche Mendeliane come
la fibrosi cistica, la malattia di Huntington e l’emocromatosi è stata ottenuta con
successo già da alcuni anni. Gli studi volti a identificare i geni coinvolti negli
errori refrattivi hanno utilizzato prevalentemente approcci di associazione
genetica. Un esempio sono gli studi di associazione genica a vasto raggio
(Genome-wide association studies, GWASs) che hanno identificato migliaia loci
genici che contribuiscono alle malattie umane comuni. L’ampia maggioranza dei
loci identificati per le malattie comuni mostrano degli effetti modesti che
potrebbero essere difficili da riprodurre. Questo spiega la limitata
sovrapposizione tra i risultati di studi diversi. Anche se questa tipologia di studio

50
può essere una potente metodologia per scoprire le possibili cause della miopia,
gli studi di associazione hanno diversi limiti. In questo tipo di studio possono
risultare dei falsi positivi che non sono replicati in studi successivi oppure risultati
veritieri possono non essere replicati in studi successivi a causa di assetti
sperimentali meno potenti o a causa di eterogeneità della popolazione presa
come campione negli studi seguenti. I risultati degli studi di associazione devono
essere replicati in ulteriori studi su distinte etnie prima che un gene possa essere
definitivamente considerato associato, positivamente o negativamente, con la
miopia. I geni candidati sono stati scelti, in questi studi, per la loro posizione
fisica in regioni cromosomiche conosciute per la miopia, grazie alla loro funzione
biologica conosciuta o in base al pattern di espressione del gene.
Negli ultimi anni, molti studi hanno riportato numerosi possibili geni implicati
nello sviluppo della miopia. Una relazione positiva sembra essere stata
riscontrata nei geni coinvolti nella crescita della matrice extracellulare e vie di
segnale coinvolte nel rimodellamento della matrice. Tra questi sono inclusi geni
che codificano per diversi costituenti extracellulari comprendendo:
collagene (COL2A1, COL1A1);
fattori di crescita transformanti (TGFB1, TGFB2, TGIF1);
hepatocyte growth factor (HGF) e i suoi recettori (cMET);
insuline-like growth factor (IGF1);
metalloproteinasi della matrice (MMP1, MMP2, MMP3, MMP9) e
proteoglicano lumicano (LUM).
Il 90% della sclera nei mammiferi è costituita dalla relazione tra i prodotti di
questi geni e il collagene. Questi risultati sottolineano fortemente che la base
genetica dell’errore refrattivo umano è parzialmente spiegata dalle variazioni
genetiche che colpiscono direttamente la composizione della matrice
extracellulare nel tessuto sclerale, portando a differenti valori di ingrandimento
oculare e di suscettibilità alla miopia. La maggior parte dei possibili geni implicati
nella miopia fino a ora identificati riguarda la miopia elevata. Almeno due di

51
questi geni (HGF, hepatocyte growth factor e COL2A1,collagen, type II, alpha 1)
hanno mostrato una correlazione anche con i gradi più lievi di miopia.
Verhoeven, nel 2013, pubblica una meta-analisi internazionale utilizzando i dati
degli equivalenti sferici di più di 45000 partecipanti, dei quali 37382 individui da
27 popolazioni di discendenza europea, e 8376 individui da cinque popolazioni di
discendenza asiatica. In tutti i partecipanti allo studio sono stati identificati 26
loci significativi, includendo 15 regioni cromosomiche già identificate in studi
precedenti. I geni identificati sono coinvolti:
nella neurotrasmissione ( GRIA4),
nel trasporto degli ioni (KCNQ5, CD55, CHNRG),
nel metabolismo dell’acido retinoico ( RDH5, RORB, CYP26A1),
nel rimodellamento della matrice extracellulare (LAMA2, BMP2) e
nello sviluppo dell’occhio (SIX4, PRSS56, CHD7).
Nonostante queste scoperte, così come per la maggior parte delle malattie
complesse, le associazioni significative spiegano solo il 3-4% della varianza [16].
Nello stesso anno, Kiefer compie un più ampio studio di associazione genomica
usufruendo dei dati raccolti dal progetto commerciale per la ricerca di parenti
basata su DNA 23andMe. Con quasi 26000 casi di miopia e 20000 casi controllo,
identifica 22 loci associati significativi [15]. La somiglianza dei risultati, di due
differenti studi che hanno utilizzato metodologie completamente diverse, è
degna di nota: dei 22 loci scoperti da Verhoeven, 14 sono stati replicati da Kiefer
e quelle regioni non confermate hanno una suggestiva associazione in altri studi.
Come già discusso, negli ultimi anni sono stati condotti numerosi studi sui
determinanti ereditari della miopia, ciascuno dei quali ha evidenziato in maniera
più o meno chiara il possibile coinvolgimento di uno o più geni. In questo
elaborato si è cercato di svolgere una ricerca per quanto possibile esaustiva dei
risultati pubblicati e, sulla base di questi, di raccogliere e riportare un gruppo di
geni verosimilmente associati allo sviluppo e alla progressione della miopia e la
sua progressione, abbiamo raccolto una lista di nomi e simboli di possibili geni
implicati nello sviluppo di tutti i gradi di miopia. Gli 82 geni riportati in almeno
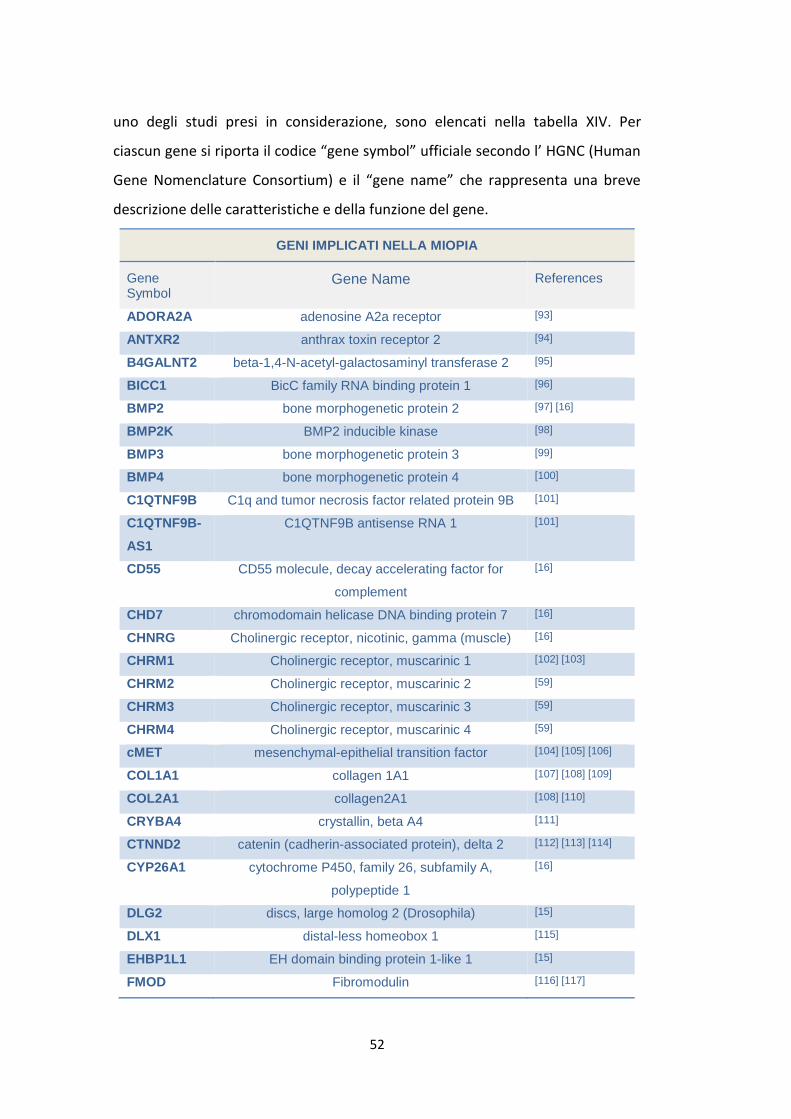
52
uno degli studi presi in considerazione, sono elencati nella tabella XIV. Per
ciascun gene si riporta il codice “gene symbol” ufficiale secondo l’ HGNC (Human
Gene Nomenclature Consortium) e il “gene name” che rappresenta una breve
descrizione delle caratteristiche e della funzione del gene.
GENI IMPLICATI NELLA MIOPIA
Gene Symbol
Gene Name References
ADORA2A adenosine A2a receptor [93]
ANTXR2 anthrax toxin receptor 2 [94]
B4GALNT2 beta-1,4-N-acetyl-galactosaminyl transferase 2 [95]
BICC1 BicC family RNA binding protein 1 [96]
BMP2 bone morphogenetic protein 2 [97] [16]
BMP2K BMP2 inducible kinase [98]
BMP3 bone morphogenetic protein 3 [99]
BMP4 bone morphogenetic protein 4 [100]
C1QTNF9B C1q and tumor necrosis factor related protein 9B [101]
C1QTNF9B-
AS1
C1QTNF9B antisense RNA 1 [101]
CD55 CD55 molecule, decay accelerating factor for
complement
[16]
CHD7 chromodomain helicase DNA binding protein 7 [16]
CHNRG Cholinergic receptor, nicotinic, gamma (muscle) [16]
CHRM1 Cholinergic receptor, muscarinic 1 [102] [103]
CHRM2 Cholinergic receptor, muscarinic 2 [59]
CHRM3 Cholinergic receptor, muscarinic 3 [59]
CHRM4 Cholinergic receptor, muscarinic 4 [59]
cMET mesenchymal-epithelial transition factor [104] [105] [106]
COL1A1 collagen 1A1 [107] [108] [109]
COL2A1 collagen2A1 [108] [110]
CRYBA4 crystallin, beta A4 [111]
CTNND2 catenin (cadherin-associated protein), delta 2 [112] [113] [114]
CYP26A1 cytochrome P450, family 26, subfamily A,
polypeptide 1
[16]
DLG2 discs, large homolog 2 (Drosophila) [15]
DLX1 distal-less homeobox 1 [115]
EHBP1L1 EH domain binding protein 1-like 1 [15]
FMOD Fibromodulin [116] [117]

53
GJD2 gap junction protein, delta 2, 36kDa [118]
GPR25 G protein-coupled receptor 25 [15]
GRIA4 glutamate receptor, ionotropic, AMPA 4 [16]
GRM6 glutamate receptor, metabotropic 6 [119]
HGF hepatocyte growth factor (hepapoietin A; scatter
factor)
[120] [121] [104]
[122] [123]
HLA-DQB1 major histocompatibility complex, class II, DQ beta
1
[124] [125]
IGF1 insulin-like growth factor 1 (somatomedin C) [126]
KCNMA1 potassium large conductance calcium-activated
channel, subfamily M, alpha member 1
[15]
KCNQ5 potassium voltage-gated channel, KQT-like
subfamily, member 5
[16]
LAMA1 Laminin, alpha 1 [127]
LAMA2 laminin, alpha 2 [16]
LEPREL1 leprecan-like 1 [128]
LRFN5 leucine rich repeat and fibronectin type III domain
containing 5
[15]
LRRC4C leucine rich repeat containing 4C [15]
LUM LUM [129] [121] [103]
[130] [102] [117]
LYPLAL1 lysophospholipase-like 1 [131]
MFRP membrane frizzled-related protein [132]
MIPEP mitochondrial intermediate peptidase [101]
MMP1 matrix metalloproteinase [133]
MMP2 matrix metalloproteinase [133]
MMP3 matrix metalloproteinase [109] [134]
MMP9 matrix metalloproteinase [134]
MYO1D myosin ID [134]
MYOC myocilin, trabecular meshwork inducible
glucocorticoid response
[135] [136] [137]
[138]
NPLOC4 nuclear protein localization 4 homolog (S.
cerevisiae)
[139]
NYX Nyctalopin [140]
OPTC opticin [117]
PABPCP2 poly(A) binding protein, cytoplasmic 1 pseudogene
2
[15]
PAX6 Paired box 6 [110] [141] [142]
[143] [144] [145]

54
[146] [147] [148]
PDE11A phosphodiesterase 11A [15]
PRELP proline/arginine-rich end leucine-rich repeat
protein
[117]
PRSS56 protease, serine, 56 [16]
PZP pregnancy-zone protein [149]
QKI QKI, KH domain containing, RNA binding
RASGRF1 Ras protein-specific guanine nucleotide-releasing
factor 1
[150]
RBFOX1 RNA binding protein, fox-1 homolog (C. elegans) 1 [151]
RDH5 retinol dehydrogenase 5 (11-cis/9-cis) [16]
RGR retinal G protein coupled receptor [152]
RORB RAR-related orphan receptor B [16]
SERPINI2 serpin peptidase inhibitor, clade I (pancpin),
member 2
[153]
SETMAR SET domain and mariner transposase fusion gene [15]
SFRP1 secreted frizzled-related protein 1 [154]
SHISA6 shisa family member 6 [15]
SIX4 sine oculis-related homeobox 4 [16]
SLC30A10 solute carrier family 30, member 10 [131]
TCF7L2 transcription factor 7-like 2 (T-cell specific, HMG-
box)
[155]
TGFB1 Trasforming growth factor, beta 1 [156] [157] [121]
[158] [157] [158]
[159]
TGFB2 Trasforming growth factor, beta 2 [116]
TJP2 tight junction protein 2 [15]
UMODL1 uromodulin-like 1 [160]
VDR vitamin D (1,25- dihydroxyvitamin D3) receptor [161]
ZBTB38 zinc finger and BTB domain containing 38 [15]
ZC3H11B zinc finger CCCH-type containing 11B
pseudogene
[131]
ZIC2 zinc finger protein of the cerebellum 2 [162]
ZNF644 zinc finger protein 644 [163] [164]
A partire da questo gruppo di geni raccolto in letteratura, per questo elaborato
sono state condotte anche alcune analisi originali per ottenere una migliore
Tabella XIV. Lista dei possibili geni candidati nella miopia.

55
interpretazione del significato funzionale dei geni rappresentati nel loro insieme
e per superare il livello di granularità relativo al singolo gene, con il fine ultimo di
caratterizzare i possibili elementi funzionali di circuiti regolativi o di interazione
proteina-proteina che vedono rappresentati più geni implicati nella miopia.
I geni non agiscono infatti in isolamento: il prodotto o i prodotti genici, le
proteine (o gli RNA funzionali), esplicano le loro funzioni all’interno di complessi
molecolari più grandi, comprendenti diverse proteine, oppure fanno parte di vie
biosintetiche o regolative che vedono la partecipazione di molti elementi
codificati da geni diversi. L’espressione dei geni è inoltre regolata da RNA e
proteine prodotti da altri geni.
Le funzioni geniche sono codificate in diversi database.
La Gene Ontology [165]riporta, per molti geni, le Funzioni Molecolari (MF), i
Processi Biologici in cui sono coinvolti (BP) e le Componenti Cellulari di cui fanno
parte (CC).
Il database KEGG [166] è il più completo database di mappe che ricapitolano
vie biosintetiche e vie regolative che modellizzano le interazioni tra geni e
prodotti genici in diversi contesti biologici. Entrambi i database utilizzano
linguaggi controllati e organizzati in gerarchie ordinate e “machine readable”.
Una branca della biologia moderna detta Systems Biology, utilizza metodi
mutuati dalle scienze esatte, come la matematica e la fisica, per la ricostruzione
e l’analisi di reti (grafi in cui i nodi rappresentano i geni e gli archi delle relazioni)
in grado di ricapitolare diversi tipi di relazioni tra geni e prodotti genici e
modellizzare, in ultima analisi, il funzionamento dei sistemi biologici. Dopo aver
raccolto la lista di geni implicati nella miopia abbiamo eseguito un’analisi di
arricchimento funzionale con un webtool chiamato David. Quest'analisi
considera le funzioni note dei geni nel loro insieme e va a ricercare ontologie
arricchite nel gruppo specifico di geni associati alla miopia, rispetto a quanto
atteso, considerando la numerosità dei geni per funzione, in tutto il genoma
umano (background gene set). Il webtool David utilizza il Gene Ontology, un
progetto bioinformatico atto ad unificare la descrizione delle caratteristiche dei

56
geni in tutte le specie. Infatti la mancanza di una terminologia universale
standard nel campo della biologia e i domini correlati, rende la comunicazione e
la condivisione dei dati più difficile. Il progetto Gene Ontology fornisce una
ontologia di termini definiti, che descrivono le funzioni dei geni in maniera
organizzata mappando la conoscenza su una struttura ad albero basata su un
dizionario controllato, che rappresenta le proprietà dei prodotti dei geni.
L'ontologia è suddivisa in tre ambiti:
Cellular component (CC): le parti di una cellula o dal suo ambiente
extracellulare;
Molecular function (MF): le attività elementari di un prodotto genico a
livello molecolare, come legante o catalisi;
Biological process (BP): le operazioni o complessi di eventi molecolari con
un inizio e una fine definiti, pertinenti al funzionamento di unità viventi
integrate: cellule, tessuti, organi e organismi.
Ogni termine GO all'interno dell’ontologia ha una nomenclatura precisa, che può
essere una parola o una stringa di parole, un identificativo alfanumerico univoco,
una definizione con le fonti citate, e un namespace che indica il dominio cui
appartiene.
Dei nostri 82 geni, 5 non sono ritrovati nel database di David, che ne annota
quindi 77. Per ciascuna delle tre ontologie, viene eseguito un test di
arricchimento, che identifica i termini funzionali (GO terms) significativamente
arricchiti nel gruppo di geni considerati rispetto all’ipotesi nulla, basata su un
gruppo di geni scelti a caso nel genoma umano. Per ogni test viene calcolato un
p-value, quest’ultimo viene poi corretto per i test multipli, ottenendo un valore
FDR (False Discovery Rate) [167]. Nella nostra piccola ricerca abbiamo
considerato significativamente arricchiti i termini con FDR<0.05. Come esempio
dalla tabella prendiamo il processo biologico della percezione sensoriale dello
stimolo luminoso (GO:0050953). Con questo termine sono associati 12 geni della
nostra lista (GJD2, LUM, GRM6, PAX6, RORB, COL2A1, COL1A1, RGR, MFRP,
CRYBA4, RDH5, NYX). Il termine risulta significativamente arricchito nella lista

57
(FDR = 4,55 E-06), in quanto osserviamo 12 geni su 77 associati al termine
funzionale, 12,1 volte di più di quanto atteso per caso, cioè scegliendo 77 geni
dal genoma umano in modo casuale. Gli stessi geni trovati nella percezione
sensoriale dello stimolo luminoso li troviamo anche nella percezione visuale. Nei
processi biologici quali il processo del sistema neurologico e la cognizione oltre
ad altri geni sono presenti anche quelli sopra citati, eccetto per il COL1A1 (tabella
XV). Anche gli arricchimenti in termini riguardanti GO CC e GO MF sono
interessanti, rispettivamente in quanto risultano arricchiti termini riguardanti la
matrice extracellulare, struttura importante per l'istologia dell'occhio e termini
relativi alla funzione dei recettori muscarinici dell'acetilcolina accoppiati e
proteine G. Da notare che, tra questi, il gene CHRM3 codifica per un recettore
che controlla la contrazione del muscolo liscio e risulta coinvolto
nell'accomodamento [168].
Un’analisi simile ci ha permesso di cercare le mappe KEGG significativamente
arricchite nei geni considerati. L’unica KEGG pathway arricchita dove ritroviamo
12 geni presenti nella lista iniziale, con FDR < 0.05 è quella che descrive tutta una
serie di processi regolativi fondamentali alterati in diversi tipi di tumori e
caratteristici delle cellule tumorali (tabella XVI).
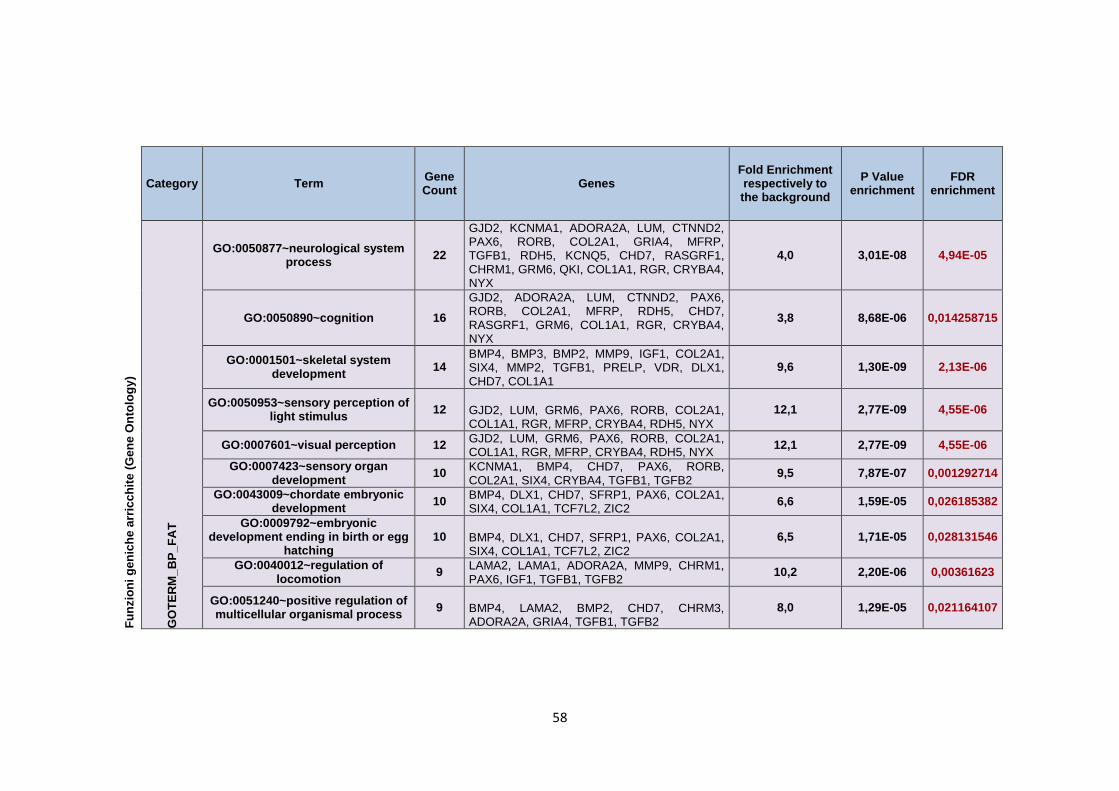
58
Category Term Gene Count
Genes Fold Enrichment respectively to the background
P Value enrichment
FDR enrichment
Fu
nzio
ni
gen
ich
e a
rric
ch
ite
(G
en
e O
nto
log
y)
GO
TE
RM
_B
P_
FA
T
GO:0050877~neurological system process
22
GJD2, KCNMA1, ADORA2A, LUM, CTNND2, PAX6, RORB, COL2A1, GRIA4, MFRP, TGFB1, RDH5, KCNQ5, CHD7, RASGRF1, CHRM1, GRM6, QKI, COL1A1, RGR, CRYBA4, NYX
4,0 3,01E-08 4,94E-05
GO:0050890~cognition 16
GJD2, ADORA2A, LUM, CTNND2, PAX6, RORB, COL2A1, MFRP, RDH5, CHD7, RASGRF1, GRM6, COL1A1, RGR, CRYBA4, NYX
3,8 8,68E-06 0,014258715
GO:0001501~skeletal system development
14 BMP4, BMP3, BMP2, MMP9, IGF1, COL2A1, SIX4, MMP2, TGFB1, PRELP, VDR, DLX1, CHD7, COL1A1
9,6 1,30E-09 2,13E-06
GO:0050953~sensory perception of light stimulus
12 GJD2, LUM, GRM6, PAX6, RORB, COL2A1, COL1A1, RGR, MFRP, CRYBA4, RDH5, NYX
12,1 2,77E-09 4,55E-06
GO:0007601~visual perception 12 GJD2, LUM, GRM6, PAX6, RORB, COL2A1, COL1A1, RGR, MFRP, CRYBA4, RDH5, NYX
12,1 2,77E-09 4,55E-06
GO:0007423~sensory organ development
10 KCNMA1, BMP4, CHD7, PAX6, RORB, COL2A1, SIX4, CRYBA4, TGFB1, TGFB2
9,5 7,87E-07 0,001292714
GO:0043009~chordate embryonic development
10 BMP4, DLX1, CHD7, SFRP1, PAX6, COL2A1, SIX4, COL1A1, TCF7L2, ZIC2
6,6 1,59E-05 0,026185382
GO:0009792~embryonic development ending in birth or egg
hatching 10 BMP4, DLX1, CHD7, SFRP1, PAX6, COL2A1,
SIX4, COL1A1, TCF7L2, ZIC2
6,5 1,71E-05 0,028131546
GO:0040012~regulation of locomotion
9 LAMA2, LAMA1, ADORA2A, MMP9, CHRM1, PAX6, IGF1, TGFB1, TGFB2
10,2 2,20E-06 0,00361623
GO:0051240~positive regulation of multicellular organismal process
9 BMP4, LAMA2, BMP2, CHD7, CHRM3, ADORA2A, GRIA4, TGFB1, TGFB2
8,0 1,29E-05 0,021164107
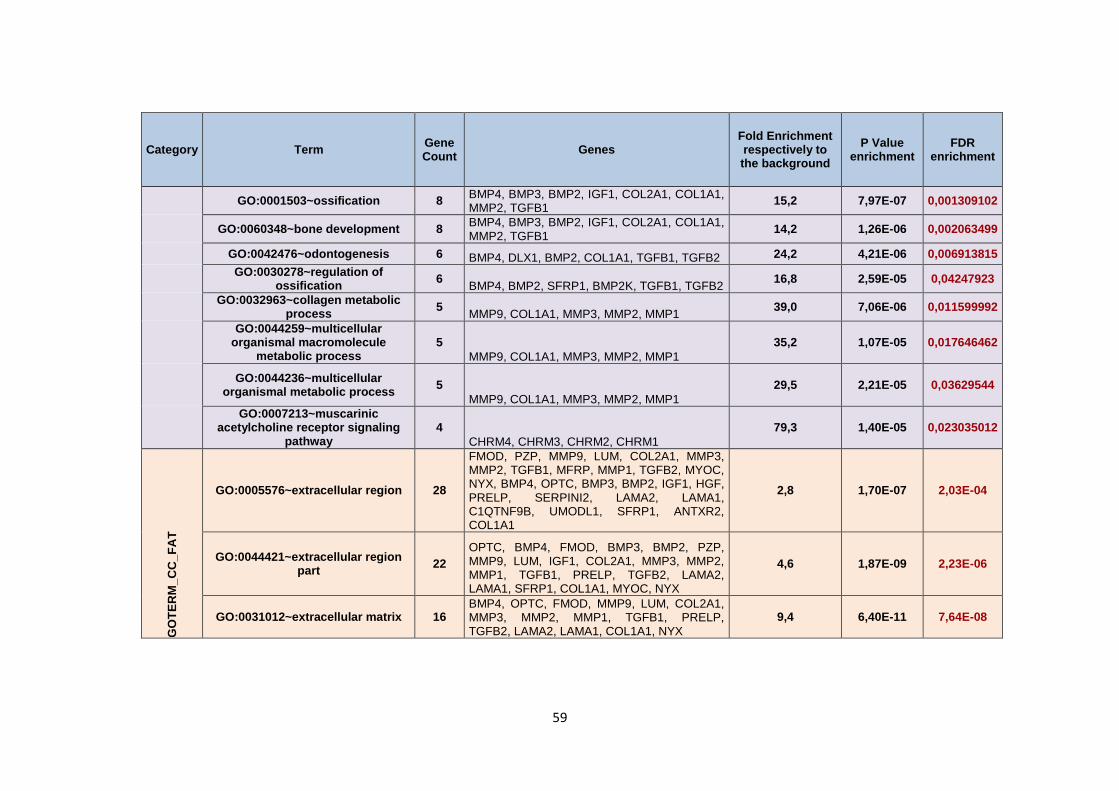
59
Category Term Gene Count
Genes Fold Enrichment respectively to the background
P Value enrichment
FDR enrichment
GO:0001503~ossification 8 BMP4, BMP3, BMP2, IGF1, COL2A1, COL1A1, MMP2, TGFB1
15,2 7,97E-07 0,001309102
GO:0060348~bone development 8 BMP4, BMP3, BMP2, IGF1, COL2A1, COL1A1, MMP2, TGFB1
14,2 1,26E-06 0,002063499
GO:0042476~odontogenesis 6 BMP4, DLX1, BMP2, COL1A1, TGFB1, TGFB2 24,2 4,21E-06 0,006913815
GO:0030278~regulation of ossification
6 BMP4, BMP2, SFRP1, BMP2K, TGFB1, TGFB2
16,8 2,59E-05 0,04247923
GO:0032963~collagen metabolic process
5 MMP9, COL1A1, MMP3, MMP2, MMP1
39,0 7,06E-06 0,011599992
GO:0044259~multicellular organismal macromolecule
metabolic process 5
MMP9, COL1A1, MMP3, MMP2, MMP1
35,2 1,07E-05 0,017646462
GO:0044236~multicellular organismal metabolic process
5
MMP9, COL1A1, MMP3, MMP2, MMP1
29,5 2,21E-05 0,03629544
GO:0007213~muscarinic acetylcholine receptor signaling
pathway 4
CHRM4, CHRM3, CHRM2, CHRM1
79,3 1,40E-05 0,023035012
GO
TE
RM
_C
C_
FA
T
GO:0005576~extracellular region 28
FMOD, PZP, MMP9, LUM, COL2A1, MMP3, MMP2, TGFB1, MFRP, MMP1, TGFB2, MYOC, NYX, BMP4, OPTC, BMP3, BMP2, IGF1, HGF, PRELP, SERPINI2, LAMA2, LAMA1, C1QTNF9B, UMODL1, SFRP1, ANTXR2, COL1A1
2,8 1,70E-07 2,03E-04
GO:0044421~extracellular region part
22
OPTC, BMP4, FMOD, BMP3, BMP2, PZP, MMP9, LUM, IGF1, COL2A1, MMP3, MMP2, MMP1, TGFB1, PRELP, TGFB2, LAMA2, LAMA1, SFRP1, COL1A1, MYOC, NYX
4,6 1,87E-09 2,23E-06
GO:0031012~extracellular matrix 16 BMP4, OPTC, FMOD, MMP9, LUM, COL2A1, MMP3, MMP2, MMP1, TGFB1, PRELP, TGFB2, LAMA2, LAMA1, COL1A1, NYX
9,4 6,40E-11 7,64E-08

60
Category Term Gene Count
Genes Fold Enrichment respectively to the background
P Value enrichment
FDR enrichment
GO:0005578~proteinaceous extracellular matrix
15 BMP4, OPTC, FMOD, LUM, MMP9, COL2A1, MMP3, MMP2, MMP1, TGFB1, PRELP, LAMA2, LAMA1, COL1A1, NYX
9,5 2,82E-10 3,36E-07
GO:0043005~neuron projection 10 KCNMA1, CHRM4, CHRM3, ADORA2A, CHRM2, RASGRF1, CHRM1, GRIA4, TGFB1, TGFB2
5,9 3,68E-05 0,043889317
GO:0030424~axon 9 KCNMA1, CHRM4, CHRM3, ADORA2A, CHRM2, CHRM1, GRIA4, TGFB1, TGFB2
11,5 9,19E-07 0,001097657
GO:0045211~postsynaptic membrane
8 KCNMA1, CHRM4, CHRM3, ADORA2A, CHRM2, CHRM1, GRIA4, DLG2
12,0 3,80E-06 0,004540728
GO:0033267~axon part 7 KCNMA1, CHRM4, CHRM3, ADORA2A, CHRM2, CHRM1, GRIA4
26,3 2,19E-07 2,62E-04
GO:0043679~nerve terminal 6 KCNMA1, CHRM4, CHRM3, CHRM2, CHRM1, GRIA4
32,9 8,81E-07 0,001052485
GO:0032279~asymmetric synapse 5 CHRM4, CHRM3, ADORA2A, CHRM2, CHRM1
112,7 6,21E-08 7,41E-05
GO
TE
RM
_M
F_
FA
T
GO:0016907~G-protein coupled acetylcholine receptor activity
4
CHRM4, CHRM3, CHRM2, CHRM1
164,9 1,03E-06 0,001283589
GO:0004981~muscarinic acetylcholine receptor activity
4
CHRM4, CHRM3, CHRM2, CHRM1
164,9 1,03E-06 0,001283589
Tabella XV. Nella tabella sono riportati i termini divisi nei tre ambiti delle tre ontologie. Per ogni termine sono espressi: il numero dei geni della lista trovati
in quel particolare processo biologico, componente cellulare e funzione molecolare; i nomi dei geni; l’arricchimento rispetto al background; il p-value e il
valore FDR. Quest’ultimo indica la significatività statistica dell’arricchimento osservato.

61
KEGG PATHWAY Gene Count Genes Fold
Enrichment P Value FDR
hsa05200 Pathways in
cancer 12
BMP4, LAMA2, LAMA1, BMP2, MMP9, IGF1, HGF, TCF7L2, MMP2, MMP1, TGFB1, TGFB2
5,3 5.08E-06 0,0051
Tabella XVI. Per il termine pathways in cancer sono riportati il numero dei geni, della lista iniziale, trovati; il nome dei geni; il p-value e il valore FDR per
l’arricchimento osservato.

62
Figura 7. In questa
pathway KEGG
intitolata “Pathways
in cancer” sono
mostrati i geni noti
per essere coinvolti
nello sviluppo dei
tumori e le relazioni
tra geni e prodotti
genici. I 12 geni,
mostrati con una
stellina a fianco al
nome, sono presenti
anche nella lista di
geni implicati per la
miopia. La pathway
risulta
significativamente
arricchita in geni
collegati alla miopia,
molti dei quali hanno
importanti ruoli
regolativi in processi
fisiologici che sono
alterati nei tumori.

63
A partire dalla lista dei geni abbiamo costruito una rete con il programma String
(http://string-db.org/), che mette in evidenza le relazioni tra i prodotti dei geni in
base a diversi tipi di evidenze sperimentali e di dati di conoscenza e letteratura.
Noi abbiamo cercato le coppie di geni che potessero essere collegati in base a
evidenze di:
co-espressione genica;
interazioni proteina-proteina;
co-rappresentazioni dei nomi dei geni in database di conoscenza e in
letteratura.
In questo modo abbiamo ottenuto una rete che collega 41 su 77 (53.24%) dei
geni considerati e ne descrive le relazioni. Nella figura 8 sono rappresentati tutti i
geni con le possibili interazioni tra di essi, in questa immagine sono raffigurati
anche i geni che non mostrano avere relazioni di nessun tipo con un altro gene
presente nella lista. Poi abbiamo eliminato i nodi della rete non connessi e
abbiamo calcolato gli arricchimenti funzionali usando solo i 41 geni della
componente connessa della rete, quindi abbiamo ricalcolato la rete String
mostrando gli archi che collegano i nodi con spessore proporzionale al peso delle
evidenze sperimentali che supportano le interazioni (Figura 9).

64
Legenda
Interazione proteina-proteina sperimentalmente dimostrata
Geni co-espressi quindi potenzialmente e funzionalmente correlati
Gruppo di interazioni proteina-proteina da database
Gruppo interazioni proteina-proteina estratti dagli abstract scientifici
Omologia delle proteine
Figura 8. Evidenzia la presenza/ assenza di legami dei geni candidati per essere
implicati nella miopia e i prodotti genici corrispondenti. Ogni nodo rappresenta un gene e
in particolare la principale proteina codificata. Gli archi che connettono coppie di nodi
mostrano relazioni funzionali note. Con i diversi colori si identificano diversi tipi di
relazioni tra i nodi, come spiegato dalla legenda.

65
Figura 9. Rete di interazioni proteina-proteina supportate sperimentalmente, del
sottoinsieme dei geni implicati nello sviluppo e nella progressione della miopia che
risultano connessi ad almeno un altro gene in base alle analisi con il programma String.
Lo spessore degli archi è direttamente proporzionale al peso delle informazioni
disponibili riguardo a ciascuna interazione.

66
4. Conclusione: fattori ambientali e genetici interagiscono in modo
complesso.
Nell’elaborato abbiamo dimostrato, grazie a vari studi presi in considerazione,
che la refrazione è un fenotipo complesso influenzato sia da fattori ambientali sia
da fattori genetici.
Numerose evidenze, da modelli di miopia sperimentale e studi epidemiologici,
hanno dimostrato che l’esposizione ambientale gioca un ruolo cruciale nella
crescita oculare e lo sviluppo refrattivo. Il preciso meccanismo biologico
attraverso il quale i fattori ambientali influenzano la refrazione umana è,
tuttavia, ancora tema di discussione. È probabile che variabili esogene
interagiscano con fattori ereditabili al fine di modulare la crescita del bulbo
oculare durante lo sviluppo dell’occhio. L’evidenza a favore di un ruolo della
predisposizione genetica nello sviluppo refrattivo è anch’essa convincente.
Diversi studi di linkage hanno mappato circa 20 loci per la miopia elevata e per la
miopia moderata considerando la refrazione come un tratto quantitativo. Gli
studi di associazione genetica hanno identificato delle varianti in almeno 25 geni
presumibilmente coinvolti nella refrazione oculare. In ogni caso, solo poche
associazioni positive sono state replicate in maniera convincente da campioni
indipendenti. Gli studi di associazione Genome-wide che sono in corso al
momento, aiuteranno a scoprire nuove variabili coinvolte negli errori refrattivi,
facendo chiarezza sulla relazione tra le variabili conosciute di suscettibilità per la
miopia, e di offrire una maggiore conoscenza nel meccanismo complesso che sta
alla base dello sviluppo refrattivo.
Per sviluppare l’immagine completa dell’epidemiologia dell’errore refrattivo
bisognerà considerare sia i fattori ambientali che quelli genetici.
Nello studio di Mutti e colleghi, del 2002 [17], la più importante associazione,
potenzialmente contradditoria, è tra il lavoro prossimale e l’errore refrattivo dei
genitori. Forse i genitori miopi hanno figli miopi solo perché hanno trasmesso
loro un “ambiente miope” caratterizzato da richieste di lavoro prossimale
intense. Le variabili sono state valutate in un modello di regressione multivariato.

67
L’associazione tra miopia e numero di genitori miopi, lavoro prossimale in
diottrie-ore per settimana, il numero di ore trascorse in attività sportive per
settimana, e i risultati locali del ITBS (Iowa tests of basic skilles) per la lettura
sono stati aggiustati per gli effetti di ogni altro fattore in questo modello.
Nonostante la loro correlazione, i fattori di rischio hanno un effetto lievemente
contradditorio nell’associazione con la miopia. Questo suggerisce che
l’associazione tra la miopia dei bambini e quella dei genitori possa essere dovuta
all’ereditarietà piuttosto che a maggiori richieste di lavoro prossimale in bambini
miopi con genitori miopi. Il lavoro prossimale sembra avere un’associazione
indipendente con la miopia che non viene spiegata da attitudini accademiche
maggiori nei miopi o miopia nei genitori. Similmente, i miopi riescono ad
ottenere punteggi più alti nei test di lettura indipendentemente dalla quantità di
tempo che dedicano alle attività prossimali.
L’ipotesi di ereditabilità della suscettibilità al lavoro prossimale può essere
valutata statisticamente testando se sia presente un’interazione significativa tra
lavoro prossimale e la storia refrattiva dei genitori. Se l’ipotesi fosse vera, gli
odds ratio associati al lavoro prossimale dovrebbero essere molto più elevati nei
bambini con due genitori miopi e i valori più bassi nei bambini con nessuno dei
due genitori miopi. In questo studio livelli ottenuti non corrispondono alle
aspettative, quindi, non trovano evidenze che supportino la teoria che
l’ereditabilità sia importante solo perché i genitori miopi hanno figli che
trascorrono più tempo in attività prossimali. Bambini con genitori non miopi
dedicano lo stesso tempo, dei bambini con genitori miopi, allo svolgimento di
attività prossimali. In conclusione sia il lavoro prossimale che i genitori miopi
sono associati alla miopia ritrovata nei bambini, l’ereditabilità dell’errore
refrattivo però rimane un fattore più importante nello sviluppo e progressione
miopica. Non emergono evidenze a supporto né della teoria che i bambini con
miopia somiglino ai genitori poiché svolgono più attività prossimali né che essi
possano ereditare una suscettibilità all’ambiente.

68
Fino ad oggi, la maggior parte degli studi hanno indagato separatamente sui
fattori ambientali e genetici della la miopia. Rimane ancora da delucidare se
esistano interazioni tra i segni genetici e l’esposizione ambientale sul rischio della
miopia. Nello studio di Fan [18], si esamina se il livello di istruzione modifichi
l’associazione tra i polimorfismi a singolo nucleotide (SNPs), recentemente
scoperti, e l’errore refrattivo, utilizzando i dati da cinque studi di coorte basati
sulla popolazione di Singapore. I partecipanti allo studio sono cinesi, malesiani e
indiani con un età tra i 30 e i 55 anni. In questo studio, le interazioni più
promettenti tra SNPs e l’educazione sono state individuate su tre loci genetici
(SHISA6-DNAH9, GJD2, ZMAT4-SFRP1). Modelli di interazioni simili sono stati
osservati anche per la lunghezza assiale e la miopia. Di questi tre loci genetici
GJD2 è stato identificato come il primo gene consistente per la suscettibilità della
miopia. La sua importanza è stata scoperta per la prima volta in uno studio di
associazione genome-wide su soggetti europei al marker genetico rs634990
[118], seguito da diverse replicazioni in plurimi studi [168] [169] [170]. I geni
all’interno o vicini ai loci SHISA6-DNAH9 e ZMAT4-SFRP1 sono connessi al
processo della trasduzione del segnale nella retina e quindi probabilmente
coinvolti nello sviluppo degli errori refrattivi. I geni DNAH9, SFRP1 e GJD2
giocano un ruolo cruciale nella retina mantenendo le funzioni dei fotorecettori
[171], determinando la degenerazione delle cellule dei fotorecettori [172] o
abilitando la trasduzione molecolare tra queste cellule neuronali [118]. Gli alleli,
che aumentano il rischio della miopia (varianti di SNPs: rs524952 in GJD2,
rs2969180 in SHISA6-DNAH9 e rs2137227 in ZMAT4-SFRP1) mostrano una
significativa associazione con tale errore refrattivo negli individui con un’elevata
istruzione (25% del campione preso in considerazione). Questo suggerisce che
l’influenza dei geni sulla suscettibilità della miopia può essere altamente
dipendente da fattori ambientali presenti. Inoltre gli alleli di rischio in questo
studio hanno una minore o nulla influenza sul cambiamento miopico negli
individui con un’istruzione minore. Possiamo supporre quindi che la
predisposizione genetica alla miopia possa essere latente o soppressa con una

69
minore esposizione ad un ambiente miope, che viene associato in questo caso al
livello di istruzione. Al contrario, è possibile che un’esposizione maggiore ad un
ambiente protettivo, come l’attività prossimale, possa colpire differentemente gli
individui che maggiormente sono predisposti geneticamente allo sviluppo della
miopia paragonati a quelli con un minor rischio. La chiave determinante del
potere statistico negli studi che prendono in considerazione l’interazione
ambiente-genetica sono:
le dimensioni del campione,
la rilevanza dell’interazione,
la frequenza allelica,
la forza dell’associazione per i principali effetti e
la soglia significativa.
Riassumendo, da questo studio si evince che il livello di educazione influenza
l’associazione tra i tre recenti loci individuati e l’errore refrattivo. L’effetto
genetico di questi tre loci in una refrazione miopica è significativamente più
elevato all’interno del gruppo di soggetti con un’istruzione elevata rispetto a
quelli con un’educazione minore. Queste scoperte sottolineano l’importanza del
ruolo dell’interazione ambiente-genetica nello sviluppo della miopia.

70
Bibliografia
[1] A. Rossetti e P. Gheller, Manuale di optometria e contattologia, Zanichelli, 2003.
[2] S. Saw, G. Gazzard, K. Au Eong e D. Tan, «Myopia: attempts to arrest progression,»
Br J Ophthalmol, vol. 86, pp. 1306-1311, 2002.
[3] L. Jones, L. Sinnot, D. Mutti, G. Mitchell, M. Moeschberger e K. Zadnik, «Parental
history of myopia, sports and outdoor activities, and future myopia,» Invest
Ophthalmol Vis Sci, vol. 48, pp. 3524-3532, 2007.
[4] K. Rose, I. Morgan e J. Ip, «Outdoor activity reduces the prevalence of miopya in
children,» Ophthalmol, pp. 1279-1285, 2008.
[5] M. Dirani, L. Tong e G. Gazzard, «Outdoor activity and myopia in singapore
teenage children,» Br J Ophthalmol, pp. 997-1000, 2009.
[6] J. Sherwin, M. Reacher, R. Keogh, A. Khawaja, D. Mackey e P. Foster, «The
association between time spent outdoors and myopia in children and
adolescents,» Ophthalmology, vol. 119, pp. 2141-2151, 2012.
[7] T. Wong, P. Foster, G. Johnson e S. Seah, «Refractive errors, axial ocular
dimensions, and age-related cataracts: the Tanjong Pagar survey,» Invest
Ophthalmol Vis Sci, vol. 41, pp. 1479-1485, 2003.
[8] B. Jiang, S. Schatz e K. Seger, «Myopic progression and dark focus variation in
optometric students during the firts academic year,» Clin Exp Optom, vol. 3, pp.
153-159, 2005.
[9] I. Hepsen, C. Evereklioglu e H. Bayramlar, «The effect of reading and near-work on
the development of myopia in emmetropic boys: a prospective, controlled, three-
year follow-up study,» Vision Research (Elsevier Science), vol. 41, pp. 2511-2520,
2001.
[10] C. Chen, K. Scurrah, J. Stankovich, P. Garoufalis, M. Dirani, K. Pertile, A. Richardson,
P. Mitchell e P. Baird, «Heritability and sharde environmental estimates for myopia
and associated ocular biometric traits: the genes in myopia (GEM) family study,»
Hum Genet, vol. 121, pp. 511-520, 2007.
[11] M. Dirani, S. Shekar e P. Baird, «Evidence of shared genes in refraction and axial
length: the genes in myopia (GEM) twin study,» Invest Ophthalmol Vis Sci, vol. 49,
pp. 4336-4339, 2008.

71
[12] T. Young e D. Hornbeak, «Myopia Genetics: A Review of Current Research and
Emerging Trends,» Curr Opin Ophthalmol., vol. 20, pp. 356-362, September 2009.
[13] R. Pacella, J. McLellan, K. Grice, E. Del Bono, J. Wiggs e J. Gwiazda, «Role of genetic
factors in the etiology of juvenile-onset myopia based on a longitudinal study of
refractive error,» Optom Vis Sci, vol. 76, pp. 381-386, 1999.
[14] D. Lam, D. Fan, R. Lam, S. Rao, K. Chong, J. Lau, R. Lai e E. Cheung, «The effect of
parental history of myopia on children's eye size and growth: results of a
lungitudinal study,» Invest Ophthalmol Vis Sci, vol. 49, pp. 873-876, 2008.
[15] A. Kiefer, J. Tung, C. Do, D. Hinds, J. Mountain e U. Francke, «Genome-wide
analysis points to roles for extracellular matrix remodeling, the visual cycle, and
neuronal development,» Plos Genet, 2013.
[16] V. Verhoeven, P. Hysi, R. Wojciechowski, Q. Fan e J. Guggenheim, «Genome-wide
meta-analysis of multiancestry cohorts identify multiple new susceptibility loci for
refractive error and myopia,» Nat Genet, vol. 45, pp. 314-318, 2013.
[17] D. Mutti, G. Mitchell, M. Moeschberger, L. Jones e K. Zadnik, «Parental myopia,
near work, school achievement and children's refractive error,» Invest Ophthalmol
Vis Sci, vol. 43, pp. 3633-3640, 2002.
[18] Q. Fan, R. Wojciechowski, M. Ikram Kamran, C. Cheng, P. Chen, X. Zhou, C. Pan, C.
Khor, E.-S. Tai, T. Aung, T. Wong, Y. Teo e S. Saw, «Education influences the
association between genetic variants and refractive error: a meta-analysis of five
Singapore studies,» Hum Mol Gen, vol. 23, pp. 546-554, 2014.
[19] P. Baird, M. Schaeche e M. Dirani, «The GEnes in Myopia (GEM) study in
understanding the aetiology of refractive errors,» Elsevier( progress in retinal and
eye research), vol. 29, pp. 540-542, 2010.
[20] A. O. Association, Optometric Clinical Practice Guideline: Care of the Patient with
Myopia, 1997.
[21] S. Saw, G. Gazzard e E. Shih-Yen, «Myopia and associated pathological
complications,» Ophthalmic Physiol Opt, vol. 25, pp. 381-391, 2005.
[22] B. Curtin, The myopias: basic science and clinical management, Philadelphia:
Harper & Row, 1985.
[23] M. Praveen, A. Vasavada, U. Jani, R. Trivedi e P. Choudhary, «Prevalence of
cataract type in relation to axial length in subjects with high myopia and
emmetropia in an Indian population.,» Am J Ophthalmol, vol. 145, pp. 176-181,

72
2008.
[24] T. Fricke, B. Holden, D. Wilson, G. Schlenther, K. Naidoo e S. Resnikoff, «Global
cost of correcting vision impairment from uncorrected refractive error,» Bull world
Health Organ, vol. 90, pp. 728-738, 2012.
[25] J. Kempen, P. Mitchell e K. Lee, «The prevalence of refractive errors among adult
in the United States, Western Europe and Australia,» Arch Ophthalmol, pp. 495-
505, 2004.
[26] A. French, I. Morgan, G. Burlutsky, P. Mitchell e K. Rose, «Prevalence and 5- to 6-
year incidence and progression of myopia and hyperopia in australian
schoolchildre,» Ophthalmology, vol. 120, pp. 1482-1491, 2013.
[27] J. Zhao, X. Pan, R. Sui, S. Munoz, R. Sperduto e L. Ellwein, «Refractive error study in
children: results from Shunyi district, China,» Am J Ophthalmol, vol. 129, pp. 427-
435, 2000.
[28] M. He, J. Zeng, Y. Liu, J. Xu, G. Pokharel e L. Ellwein, «Refractive error and visual
impairment in urban children in southern china,» Invest Ophthalmol Vis Sci, vol.
45, pp. 793-799, 2004.
[29] D. Fan, D. Lam, R. Lam, J. Lau, K. Chong e E. Cheung, «Prevalence, incidence, and
progression of myopia of schoolchildren in Hong Kong,» Invest Ophthalmol Vis Sci,
vol. 45, pp. 1071-1075, 2004.
[30] Q. Wang, B. Klein, R. Klein e S. Moss, «Refractive status in the Beaver Dam Eye
study,» Invest Ophthalmol Vis Sci, vol. 35, pp. 4344-4347, 1994.
[31] T. Wong, P. Foster, J. Hee, T. Ng, J. Tielsch e S. Chew, «Prevalence and risk factors
for refractive errors in adult Chinese in Singapore,» Invest Ophthalmol Vis Sci, vol.
41, pp. 2486-2494, 2000.
[32] D. Mutti e K. Zadnik, «Age-related decreases in the prevalence of myopia:
longitudinal change or cohort effect?,» Invest Ophthalmol Vis Sci, vol. 41, pp.
2103-2107, 2000.
[33] L. Lin, Y. Shih, C. Hsiao e C. Chen, «Prevalence of myopia in Taiwanese
schoolchildren: 1983 to 2000.,» Ann Acad Med Singapore, vol. 33, pp. 27-33, 2004.
[34] S. Vitale, R. Sperduto e 3. F. Ferris, «Increased prevalence of myopia in the United
States between 1971-1972 and 1999-2004. Arch Ophthalmol 2009; 127(12): 1632–
1639.,» Arch Ophthalmol, vol. 127, pp. 1632-1639, 2009.

73
[35] O. Parssinen, «The increased prevalence of myopia in Finland,» Acta Ophthalmol,
vol. 90, pp. 497-502, 2012.
[36] K. Rose, R. Harper, C. Tromans, C. Walterman, D. Goldberg e C. Haggerty, «Quality
of life in myopia,» Br J Ophthalmol, pp. 1031-1034, 2000.
[37] S. Saw, S. Tan, D. Fung, S. Chia, D. Koh, D. Tan e R. Stone, «IQ and the association
with myopia in children,» Invest Ophthalmol Vis Sci, vol. 45, pp. 2943-2948, 2004.
[38] V. D. R. Berg, M. Dirani, C. Chen, N. Haslam e P. Baird, «Myopia and personality:
the genes in myopia (GEM) personality study,» Invest Ophthalmol Vis Sci, vol. 49,
pp. 882-886, 2008.
[39] K. Rose, I. Morgan, J. Ip, A. Kifley, S. Huynh, W. Smith e P. Mitchell, «Outdoor
activity reduces the prevalence of myopia in children,» Ophthalmol, vol. 115, pp.
1279-1285, 2008.
[40] M. Dirani, L. Tong, G. Gazzard, X. Zhang, A. Chia, T. Young, K. Rose, P. Mitchell e S.
Saw, «Outdoor activity and myopia in singapore teenage children,» Br J
Ophthalmol, vol. 93, pp. 997-1000, 2009.
[41] C. McCarthy, P. Megaw, M. Devadas e I. Morgan, «Dopaminergic agents affect the
ability of brief periods of normal vision to prevent form-deprivation myopia,» Exp
Eye Res, vol. 84, pp. 100-107, 2007.
[42] R. Ahby e F. Schaeffel, «The effect of bright light on lens compensation in chicks,»
Invest Ophthalmol Vis Sci, vol. 51, pp. 5247-5253, 2010.
[43] K. Rose, I. Morgan, J. Ip, A. Kifley, S. Huynh, W. Smith e P. Mitchell, «Outdoor
activity reduces the prevalence of myopia in children,» Ophthalmology, vol. 115,
pp. 1279-1285, 2008.
[44] L. Deng, J. Gwiazda e F. Thorn, «Children's refractions and visual activities in the
school year and summer,» Optom Vis Sci, vol. 87, pp. 406-413, 2010.
[45] H. Taylor, «Racial variations in vision,» American Journal Of Epidemiology, vol. 113,
pp. 62-80, 1981.
[46] E. Harb, F. Thorn e D. Troilo, «Characteristics of accommodative behavior during
sustained reading in emmetropes and myopes,» Vision Research (Elsevier), vol. 46,
pp. 2581-2592, 2006.
[47] H. Lim, J. Yoon, S. Hwang e S. Lee, «Prevalence and associated sociodemographic
factors of myopia in Korean children: the 2005 third Korea National Health and

74
Nutrition Examination Survey,» Jpn J Ophthalmol, vol. 56, pp. 76-81, 2012.
[48] K. Au Eong, T. Tay e M. Lim, «Education and myopia in 110236 young singaporean
males,» Singapore Med J, vol. 6, pp. 489-492, 1993.
[49] S. Saw, W. Chua, C. Hong, H. Wu, W. Chan, K. Chia, R. Stone e D. Tan, «Nearwork in
early-onset myopia,» Invest Ophthalmol Vis Sci, vol. 43, pp. 332-339, 2002.
[50] J. Ip, S. Saw, K. Rose, I. Morgan, A. Kifley, J. Wang e P. Mitchell, «Role of near work
in myopia: findings in a sample of australian school children,» Invest Ophthalmol
Vis Sci, vol. 49, pp. 2903-2910, 2008.
[51] T. Queek, C. Chua e C. Chong, «Prevalence of refractive errors in teenage high
school students in Singapore,» Ophthalmic Physiol Opt., vol. 24, pp. 47-55, 2004.
[52] B. Lu, N. Congdon, X. Liu, K. Choi, D. Lam, M. Zhang, M. Zheng, Z. Zhou, L. Li, X. Liu,
A. Sharma e Y. Song, «Associations between near work, outdoor activity, and
myopia among adolescent students in rural china,» Arc Ophthalmol, vol. 127, pp.
769-775, 2009.
[53] B. Kinge, A. Midelfart e G. Jacobsen, «Refractive errors among young adults and
university students in Norway,» Acta Ophthalmol Scand, vol. 76, pp. 692-695,
1998.
[54] B. Kinge, A. Midelfart, G. Jacobsen e J. Rystad, «The influence of near-work on
development of myopia among university students. A three year longitudinal
study among engeeniring students in Norway,» Acta Ophthalmol Scand, pp. 26-29,
2000.
[55] E. Dunphy e M. Stoll, «Myopia among american male graduate students,» Am J
Ophthalmol, vol. 65, pp. 518-521, 1968.
[56] K. Zadnik e D. Mutti, «Refractive error changes in law students,» Am J Optom
Physiol Opt, vol. 64, pp. 558-561, 1987.
[57] B. Kinge e A. Midelfart, «Refractive changes among Norwegian university students:
a three-year longitudinal study,» Acta Ophthalmol Scandal, vol. 77, pp. 302-305,
1999.
[58] J. Loman, G. Quinn, L. Kamoun, G. Ying, M. Maguire, D. Hudesman e R. Stone,
«Darkness and near work, myopia and its progression in third-year law students,»
Ophthalmology, vol. 109, pp. 1032-1038, 2002.
[59] Z. Lin, B. Vasudevan, Y. Liang, Y. Zhang, L. Qiao, S. Rong, S. Li, N. Wang e K.
Ciuffreda, «Baseline characteristics of nearwork-induced transient myopia,»

75
Optom Vis Sci, vol. 89, pp. 1725-1733, 2012.
[60] J. Wolffsohn, B. Gilmartin, R. Wing-hong Li, M. Edwards, S. Wing-shan Chat, J.
Kwok-fai Lew e B. Sing-ying Yu, «Nearwork-induced transient myopia in
preadolescent Hong Kong chinese,» Invest Ophthalmol Vis Sci, vol. 44, pp. 2284-
2289, 2003.
[61] K. Ciuffreda e V. Thunyalukul, «Myopic nearwork aftereffects in children,» Invest
Ophthalmol Vis Sci, vol. 40, 1999.
[62] F. Vera Diaz, N. Strang e B. Winn, «Nearwork induced transient myopia during
myopia progression,» Curr Eye Res, vol. 24, pp. 372-379, 2002.
[63] B. Vasudevan e K. Ciuffreda, «Additivity of nearwork-induced transient myopia and
its decay characteristics in different refractive group,» Invest Ophthalmol Vis Sci,
vol. 49, pp. 836-841, 2008.
[64] M. Rosenfield, R. Desai e J. Portello, «Do progressing myopes show reduced
accomodative response?,» Optom Vis Sci, vol. 79, pp. 268-273, 2002.
[65] P. Allen e D. O'Leary, «Accomodation functions: co-dependency and relationship
to refractive error,» Vision Research (Elsevier), vol. 46, pp. 491-505, 2006.
[66] T. Langaas, P. Riddell, E. Svarverud, A. Ystenaes, I. Langeggen e J. Bruenech,
«Variability of the accomodation response in early onset myopia,» Optom Vis Sci,
vol. 85, pp. 37-48, 2008.
[67] M. Day, N. Strang, D. Seidel, L. Gray e E. Mallen, «Refractive group differences in
accomodation and microfluctuations with changing accomodation stimulus,»
Ophthalmic Physiol Opt, vol. 26, pp. 88-96, 2006.
[68] T. Norton e P. Gamlin, «The near response, emmetropia and myopia,» Ophtalmic
Physiol Opt, vol. 19, pp. 79-80, 1999.
[69] M. Abbot, K. Schmid e N. Strang, «Differences in the accommodation stimulus
response curves of adult myopes and emmetropes,» Ophthalmic Physiol Opt, vol.
18, pp. 13-20, 1998.
[70] D. Mutti, G. Mitchell, J. Hayes, L. Jones, M. Moeschberger, S. Cotter, R. Kleinstein,
R. Manny, J. Twelker e K. Zadnik, «Accomodative lag before and after the onset of
myopia,» Invest Ophthalmol Vis Sci, vol. 47, pp. 837-846, 2006.
[71] T. Fantoni, Genetica vol II, Piccin, 2009, pp. 719-727.

76
[72] http://www.federica.unina.it/medicina-veterinaria/zootecnica-generale-e-
miglioramento-genetico/genetica-caratteri-quantitativi/,
«http://www.federica.unina.it/medicina-veterinaria/zootecnica-generale-e-
miglioramento-genetico/genetica-caratteri-quantitativi/,» [Online].
[73] L. G. Sorsby A, A longitudinal study of refraction and its components during
growth, London: HMSO, 1970.
[74] D. Mutti, J. Hayes, G. Mitchell, L. Jones, M. Moeschberger, S. Cotter, R. Kleinstein,
R. Manny, J. Twelker, K. Zadnik e C. S. Group, «Refractive error, axial length, and
relative peripheral refractive error before and after the onset of myopia,» Invest
Ophthalmol Vis Sci, vol. 48, pp. 2510-2519, 2007.
[75] D. Kurtz, L. Hyman, J. Gwiazada, R. Manny, L. Dong, Y. Wang, M. Scheiman e C.
Group, «Role of parental myopia in the progression of myopia and its interaction
with treatement in COMET children,» Invest Ophthalmol Vis Sci, vol. 48, pp. 562-
570, 2007.
[76] M. Dirani, S. Shekar e P. Baird, «Evidence of shared genes in refraction and axial
length: the genes in myopia (GEM) twin study,» Invest Ophthalmol Vis Sci, n. 49,
pp. 4336-4339, 2008.
[77] E. Ojaimi, K. Rose, I. Morgan, W. Smith e F. Martin, «Distribution of ocular
biometric parameters and refraction in a population-based study of Australian
children,» Invest Ophthalmol Vis Sci, vol. 46, pp. 2748-2754, 2005.
[78] D. Atchison, N. Pritchard, K. Schimd, D. Scott, C. Jones e J. Pope, «Shape of the
retinal surface in emmetropia and myopia,» Invest Ophthalmol Vis Sci, vol. 46, pp.
2698-2707, 2005.
[79] T. Eysteinsson, F. Jonasson, A. Arnasson, H. Sasaki e K. Sasaki, «Relationships
between ocular dimensions and adult stature among partecipants in the Reykjavik
Eye Study,» Acta Ophthalmol Scand, vol. 83, pp. 734-738, 2005.
[80] E. Mallen, Y. Gammoh, M. Al-Bdour e F. Sayegh, «Refractive error and ocular
biometry in Jordan adults,» Ophthalmic Physiol Opt, vol. 25, pp. 302-309, 2005.
[81] C. Hammond, H. Snieder, C. Gilbert e T. Spector, «Genes and environment in
refractive error: the twin eye study,» Invest Ophthalmol Vis Sci, pp. 1232-1239,
2001.
[82] A. Sorsby, B. Benjamin, M. Sheridan, G. Leary e J. Stone, «Refractions and its
components during growth of the eye after the age three,» Med Res Council
Special Rep Ser., vol. 301, pp. 1-18, 1962.

77
[83] N. Lyhne, A. Sjolie, O. Kyvik e A. Green, «The importance of genes and
environment for ocular refraction and its determiners: a population based study
among 20-45 year old twins,» Br J Ophthalmol, vol. 85, pp. 1470-1476, 2001.
[84] S. Valluri, B. Minkovitz, K. Budak, L. Essary, R. walker e E. Chansue, «Comparative
corneal topography and refractive variables in monozygotic and dizygotic twins,»
Am J Ophthalmol, vol. 127, pp. 158-163, 1999.
[85] K. Zadnik, «Myopia development in childhood,» Optom Vis Sci, vol. 74, pp. 603-
608, 1997.
[86] C. Hammond, H. Snieder, C. Gilbert e T. Spector, «Genes and environment in
refractive error: the twin eye study,» Invest Ophthalmol Vis Sci, vol. 42, pp. 1232-
1236, 2001.
[87] G. Ashton, «Segregation analysis of ocular refractionand myopia,» Hum Hered, vol.
35, pp. 232-239, 1985.
[88] J. Gwiazda, F. Thorn, J. Bauer e R. Held, «Emmetropization and the progression of
manifest refraction i children followed from infancy to puberty,» Clin Vis Sci, vol. 8,
pp. 337-344, 1993.
[89] D. Mum e K. Zadnik, «The utility of three predictors of childhood myopia: a
Bayesian analysis,» Vision Res, vol. 35, pp. 1345-1352, 1995.
[90] M. Wu e M. Edwards, «The effect of having myopic parents: an analysis of myopia
in three generations,» Optom Vis Sci, vol. 76, pp. 387-392, 1999.
[91] J. Ip, S. Huynh, D. Robaei, K. Rose, I. Morgan e W. Smith, «Ethnic differences in the
impact of parental myopia:findings from a population-based study of 12-year-
old,» Invest Ophthalmol Vis Sci, vol. 48, pp. 2520-2528, 2007.
[92] D. Lam, D. Fan, R. Lam, S. Rao, K. Chong e J. e. a. Lau, «The effect of parental
history of myopia on children’s eye size and growth: results of a longitudinal
study,» Invest Ophthalmol Vis Sci, n. 49, pp. 873-876, 2008.
[93] X. Chen, A. Xue, W. Chen, Y. Ding, D. Yan, J. Peng, C. Zeng, J. Qu e X. Zhou,
«Assessment of exonic single nucleotide polymorphisms in the adenosine A(2A)
receptor gene to high myopia susceptibility in Chinese subjects,» Mol Vis, vol. 17,
pp. 486-491, 2011a.
[94] S. Bell, A. Mavila, R. Salazar, K. Bayless e S. Kanagala, «Differential gene expression
during capillary morphogenesis in 3D collagen matrices: regulated expression of
genes involved in basement membrane matrix assembly, cell cycle progression,
cellular differentiation and G-protein signaling,» J Cell Sci, vol. 114, pp. 2755-2773,

78
2001.
[95] P. Paluru, S. Ronan, E. Heon, M. Devoto e S. Wildenberg, «New locus for
autosomal dominant high myopia maps to the long arm of chromosome 17,»
Invest Ophthalmol, vol. 44, pp. 1830-1836, 2003.
[96] S. Nallasamy, P. Paluru, P. Devoto, N. Wasserman e J. Zhou, «Genetic linkage study
of high-grade myopia in a Hutterite population from South Dakota,» Mol Vis, vol.
13, p. 229–236, 2007.
[97] H. Liu, Y. Lin, W. Lin, L. Wan, J. Sheu, H. Lin, Y. Tsai, C. Tsai e F. Tsai, «A novel
genetic variant of BMP2K contributes to high myopia,» J Clin Lab Anal, vol. 23, pp.
362-367, 2009.
[98] H. Liu, Y. Lin, W. Lin, L. Wan, J. Sheu, H. Lin, Y. Tsai, C. Tsai e F. Tsai, «A novel
genetic variant of BMP2K contributes to high myopia,» J. Clin. Lab Anal., pp. 362-
367, 2009.
[99] S. Scott, A. Jun e S. Chakravarti, «Sphere formation from corneal keratocytes and
phenotype specific markers,» Exp Eye Res, vol. 93, pp. 898-905, 2011.
[100] Z. Yang, X. Xiao, S. Li e Q. Zhang, «Clinical and linkage study on a consanguineous
Chinese family with autosomal recessive high myopia,» Mol Vis, vol. 15, pp. 312-
318, 2009.
[101] Y. Shi, J. Qu, D. Zhang, P. Zhao, Q. Zhang, P. Tam, L. Sun, X. Zuo, X. Zhou, X. Xiao, J.
Hu, Y. Li, L. Cai, X. Liu, F. Lu, S. Liao, B. Chen, F. He, B. Gong, H. Lin, S. Ma, J. Cheng,
J. Zhang, Y. Chen, F. Zhao e X. e. a. Yang, «Genetic variants at 13q12.12 are
associated with high myopia in the Han Chinese population,» am J Hum Genet, vol.
88, pp. 888-892, 2011b.
[102] H. Lin, L. Wan, Y. Tsai, W. Chen, S. Tsai e F. Tsai, «Muscarinic acetylcholine
receptor 1 gene polymorphisms associated with high myopia,» Mol Vis, vol. 15, pp.
1774-1780, 2009b.
[103] H. Lin, Y. Kung, Y. Lin, J. Sheu, B. Chen, Y. Lan, C. Lai, Y. Shu, L. Wan e F. Tsai,
«Association of the Lumican gene functional 3’ UTR polymorphism with high
myopia,» Invest Ophthalmol Vis Sci, p. 51, 2009a.
[104] T. Yanovitch, Y. Li, R. Metlapally, D. Abbott, K. Viet e T. Young, «Hepatocyte growth
factor and myopia: genetic association analyses in a Caucasian population,» Mol
Vis, vol. 15, pp. 1028-1035, 2009.
[105] C. Khor, R. Grignani, D. Ng, K. Toh, K. Chia, D. Tan, D. Goh e S. Saw, «cMet and
refractive error progression in children,» Ophtalmology, vol. 116, pp. 1469-1474,

79
2009.
[106] M. Schache, C. Chen, M. Dirani e P. Baird, «The hepatocyte growth factor receptor
(MET) gene is not associated with refractive error and ocular biometrics in a
Caucasian population,» Mol Vis, vol. 15, pp. 2599-2605, 2009a.
[107] Y. Inamori, M. Ota, H. Inoko, E. Okada, R. Nishzaki, T. Shiota, J. Mok, A. Oka, S.
Ohno e N. Mizuki, «The COL1A1 gene and high myopia susceptibility in Japanese,»
Hum Genet, vol. 122, pp. 151-157, 2007.
[108] R. Metlapally, Y. Li, K. Tran-Viet, D. Abbott, G. Czaja, F. Malecaze, P. Calvas, D.
Mackey, T. Rosenberg, M. Paget, J. Zayats, T. Owen, Guggenheim e Young,
«COL1A1 and COL2A1 genes and myopia susceptibility: evidence of association
and suggestive linkage to the COL2A1 locus,» Invest Phthalmol Vis Sci, vol. 50, pp.
4080-4086, 2009.
[109] C. Liang, K. Hung, Y. Tsai, W. Chang, H. Wang e S. Juo, «Systematic assessment if
the tagged polymorphism of the COL1A1 gene for high myopia,» J Hum Genet, vol.
52, pp. 374-377, 2007.
[110] D. Mutti, M. Cooper, S. O'Brien, L. Jones, M. Mariazita, J. Murray e K. Zadnik,
«Candidate gene and locus analysis of myopia,» Mol Vis, vol. 13, pp. 1012-1019,
2007.
[111] D. Ho, M. Yap, P. Ng, W. Fung e S. Yip, «Association of high myopia with crystallin
beta A4 (CRYBA4) gene polymorphisms in the linkage-identified MYP6 locus,»
PLoS, 2012.
[112] B. Lu, D. Jiang, P. Wang, Y. Gao, W. Sun, X. Xiao, S. Li, X. Jia, X. Guo e Q. Zhang,
«Replication study supports CTNND2 as a susceptibility gene for high myopia,»
Invest Ophthalmol Vis Sci, vol. 52, pp. 8258-8261, 2011.
[113] Y. Li, L. Goh, C. Khor, Q. Fan, M. Yu, S. Han, X. Sim, R. Ong, T. Wong, E. Vithana, E.
Yap, H. Nakanishi, F. Matsuda, K. Ohno-Matsui, N. Yoshimura, M. Seielstad, E. Tai,
T. Young e S. Saw, «Genome-wide association studies reveal genetic variants in
CTNND2 for high myopia in Singapore Chinese,» Ophthalmology, vol. 118, pp. 368-
375, 2011a.
[114] Z. Yu, J. Zhou, X. Chen, X. Zhou, X. Sun e R. Chu, «Polymorphisms in the CTNND2
gene and 11q24.1 genomic region are associated with pathological myopia in a
chinese population,» Journal international d’ophthalmologie, vol. 228, pp. 123-
129, 2012.
[115] S. Lapan e P. Reddien, « DLX and sp6-9 Control optic cup regeneration in a

80
prototypic eye,» PLoS Genet, 2011.
[116] H. Lin, L. Wan, Y. Tsai, S. Liu, W. Chen, S. Tsai e F. Tsai, «Sclera-related gene
polymorphisms in high myopia,» Mol. V, vol. 15, pp. 1655-1663, 2009c.
[117] M. Majava, P. Bishop, P. Hagg, P. Scott, A. Rice, C. Inglehearn, C. Hammond, T.
Spector, L. Ala-Kokko e M. Mannikko, «Novel mutations in the small leucine-rich
repeat protein/proteoglycan (SLRP) genes in high myopia,» Hum Mutat, vol. 28,
pp. 336-344, 2007.
[118] A. Solouki, V. Verhoeven, V. C. Duijn, A. Verker, M. Ikram, P. Hysi, D. Despriet, V. L.
Koolwijk, L. Ho e D. Ramdan, «A genome-wide association study identifies a
susceptibility locus for refractive errors and myopia at 15q14,» Nat Genet, vol. 42,
pp. 897-901, 2010.
[119] X. Xu, S. Li, X. Xiao, P. Wang, X. Guo e Q. Zhang, «Sequence variations of GRM6 in
patients with high myopia,» Mol Vis, vol. 15, pp. 2094-2100, 2009.
[120] W. Han, M. Yap, J. Wang e S. Yip, «Family-based association analysis of hepatocyte
groth factor (HGF) gene polymorphisms in high myopia,» Invest Ophthalmol Vis
Sci, vol. 47, pp. 2291-2299, 2006.
[121] P. Wang, S. Li, X. Xiao, X. Jia, X. Jiao, X. Guo e Q. Zhang, «High myopia is not
associated with the SNPs in the TGIF, lumican, TGFB1, and HGF genes,» Invest
Ophthalmol Vis Sci, vol. 50, pp. 1546-1551, 2009.
[122] S. Veerappan, K. Pertile, A. Islam, M. Schache, C. Chen, P. Mitchell, M. Dirani e P.
Baird, «Role of the hepatocyte growth factor gene in refractive error,»
Ophthalmology, vol. 117, 2009a.
[123] S. Veerappan, K. Pertile, A. Islam, M. Schache, C. Chen, P. Mitchell, M. Dirani e P.
Baird, «Role of the hepatocyte growth factor gene in refractive error,»
Ophthalmology, vol. 117, pp. 239-245, 2010.
[124] X. Li, B. Ji, D. Zhu, Z. Geng e R. Chu, «Studies of the association of pathological
myopia in Chinese patients with HLA alleles,» Acta genetica Sinica, vol. 27, pp.
189-194, 2000.
[125] S. Li, B. Ji, R. Chu, T. Chen, Y. Luo, Y. Lu, D. Zhu e Z. Geng, «Study on the association
of pathologic myopia with HLA-DQB1 gene,» Chin J Ophthalmol, vol. 37, pp. 263-
266, 2001.
[126] J. Mak, M. Yap, W. Fung, P. Ng e S. Yip, «Association of IGF1 gene haplotypes with
high myopia in Chinese adults.,» Arch Ophthalmol, vol. 130, pp. 209-216, 2012.

81
[127] Y. Zhao, F. Zhang, S. Zhu, H. Duan, Y. Li, Z. Zhou, W. Ma e N. Li Wang, «The
association of a single nucleotide polymorphism in the promoter region of the
LAMA1 gene with susceptibility to Chinese high myopia,» Mol Vis, vol. 17, pp.
1003-1010, 2011.
[128] S. Mordechai, L. Gradstein, A. Pasanen, R. Ofir, K. El Amour, J. Levy, N. Belfair, T.
Lifshitz, S. Joshua, G. Narkis, K. Elbedour, J. Myllyharju e O. Birk, «High myopia
caused by a mutation in LEPREL1, encoding prolyl 3-hydroxylase 2.,» Am J Hum
Genet, vol. 89, pp. 438-445, 2011.
[129] I. Wang, T. Chiang, Y. Shih, C. Hsiao, S. Lu, Y. Hou e L. Lin, «The association of single
nucleotide polymorphisms in the 5’-regulatory region of the lumican gene with
susceptibility to high myopia in Taiwan,» Mol Vis, vol. 12, pp. 852-857, 2006.
[130] Z. Chen, I. Wang, Y. Shih e L. Lin, «The association of haplotype at the lumican
gene with high myopia susceptibility in Taiwanese patients.,» Ophthalmology, vol.
116, pp. 1920-1927, 2009.
[131] Q. Fan, V. Barathi, C. Cheng, X. Zhou, A. Meguro, I. Nakata, C. Khor, L. Goh, Y. Li, W.
Lim, C. Ho, F. Hawthorne, Y. Zheng, D. Chua, H. Inoko, K. Yamashiro, K. Ohno-
Matsui, K. Matsuo, F. Matsuda e E. Vithana, «Genetic variants on chromosome
1q41 influence ocular axial length and high myopia,» PLoS, 2012.
[132] O. Sundin, G. Leppert, E. Silva, J. Yang, S. Dharmaraj, I. Maumenee, L. Santos, C.
Parsa, E. Traboulsi, K. Broman, C. Dibernardo, J. Sunness, J. Toy e E. Weinberg,
«Extreme hyperopia is the result of null mutations in MFRP, which encodes a
Frizzled-related protein.,» Proc Natl Acad Sci USA, vol. 102, pp. 9553-9558, 2005.
[133] R. Wojciechowski, J. Bailey-Wilson e D. Stambolian, «Association of matrix
metalloproteinase gene polymorphisms with refractive error in Amish and
Ashkenazi families,» Invest Ophthalmol Vis Sci, vol. 51, pp. 4989-495, 2010.
[134] N. Hal, C. Gale, S. Ye e C. Martyn, «Myopia and polymorphisms in genes for matrix
metalloproteinases,» Invest Ophthalmol Vis sci, vol. 50, pp. 1232-1236, 2009.
[135] W. Tang, S. Yip, K. Lo, P. Ng, P. Choi, S. Lee e M. Yap, «Linkage and association of
myocilin (MYOC) polymorphisms with high myopia in a chinese population,» Mol
Vis, vol. 13, pp. 534-544, 2007.
[136] Y. Leung, P. Tam, L. Baum, D. Shun-Chiu e C.-P. Pang, «TIGR/MYOC proximal
promoter GT-repeat polymorphism is not associated with myopia,» Hum Mut
Mutation and Polymorphism Report, vol. 16, p. 533, 2000.
[137] T. Zayats, T. Yanovitch, R. Creer, G. McMahon, Y. Li, T. Young e J. Guggenheim,

82
«Myocilin polymorphisms and high myopia in subjects of european origin,» Mol
Vis, vol. 15, pp. 213-222, 2009.
[138] Z. Vatavuk, J. Skunca Herman, G. Bencic, B. Andrijevic Derk, V. Lacmanovic Loncar,
I. Petric Vickovic, K. Bucan, K. Mandic, A. Mandic, I. Skegro, J. Pavicic Astalos, I.
Merc, M. Martinovic, P. Kralj, T. Knezevic, K. Barac-Juretic e L. Zgaga, «Common
variant in myocilin gene is associated with high myopia in isolated population of
Korcula Island, Croatia,» Croat Med, vol. 50, pp. 17-22, 2009.
[139] W. Meng, J. Butterworth, P. Calvas e F. Malecaze, «Myopia and iris colour: A
possible connection?,» Med Hyp, vol. 78, pp. 778-780, 2012.
[140] W. Tang, S. Yip, Lo, N. K.K., P.W., P. Choi, S. Lee e M. Yap, «Linkage and association
of myocilin (MYOC) polymorphisms with high myopia in a Chinese population,»
Mol Vis, vol. 13, pp. 534-544, 2007.
[141] C. Hammond, T. Andrew, Y. Mak e T. Spector, «A susceptibility locus for myopia in
the normal population is linked to the PAX6 gene region on chromosome 11: a
genome-wide scan of dyzigotic twins,» Am J Hum Genet, vol. 75, pp. 294-304,
2004.
[142] Y. Tsai, C. Chiang, H. Lin, J. Lin, L. Wan e F. Tsai, «A PAX6 gene polymorphism is
associated with genetic predisposition to extreme myopia,» Eye, vol. 22, pp. 576-
581, 2008.
[143] C. Simpson, P. Hysi, S. Bhattacharya, C. Hammond, A. Webster, C. Peckham, P.
Sham e J. Rahi, «The roles of PAX6 and SOX2 in myopia: lessons from,» Invest
Ophthalmol Vis Sci, vol. 48, pp. 4421-4425, 2007.
[144] A. Hewitt, L. Kearns, R. Jamieson, K. Williamson, v. V. Heyningen e D. Mackey,
«PAX6 mutations may be associated with high myopia,» Ophtalmic Genet, vol. 28,
pp. 179-182, 2007.
[145] W. Han, K. Leung, W. Fung, J. Mak, Y. Li, M. Yap e S. Yip, «Association of PAX6
polymorphisms with high myopia in Han Chinese nuclear families,» Invest
Ophthalmol Vis Sci, vol. 50, pp. 47-56, 2009.
[146] T. Ng, C. Lam, D. Lam, S. Chiang, P. Tam, D. Wang, B. Fan, G. Yam, D. Fan e C. Pang,
«AC and AG dinucleotide repeats in the PAX6 P1 promoter are associated with
high myopia,» Mol Vis, vol. 15, pp. 2239-2248, 2009.
[147] B. Jiang, M. Yap, K. Leung, P. Ng, W. Fung, W. Lam, Y. Gu e S. Yip, «PAX6
haplotypes are associated with high myopia in Han chinese,» PLoS One, 2011.

83
[148] C. Liang, E. Hsi, K. Chen, Y. Pan e Y. J. S. Wang, «A functional polymorphism at
3’UTR of the PAX6 gene may confer risk for extreme myopia in the chinese,» Invest
Ophthalmol Vis Sci, vol. 52, pp. 3500-3505, 2011.
[149] X. Duan, Q. Lu, P. Xue, H. Zhang, Z. Dong, F. Yang e N. Wang, «Proteomic analysis
of aqueous humor from patients with myopia,» Molecular Vision, vol. 14, pp. 370-
377, 2008.
[150] P. Hysi, T. Young, D. Mackey, T. Andrew, A. Fernandez-Medarde, A. Solouki, A.
Hewitt, S. Macgregor, J. Vingerling, Y. Li, M. Ikram, L. Fai, P. Sham, L. Manyes, A.
Porteros, M. Lopes, F. Carbonaro, S. Fahy e e. al., «A genome-wide association
study for myopia and refractive error identifies a susceptibility locus at 15q25,»
Nat Get, vol. 42, pp. 902-905, 2010.
[151] D. Stambolian, R. Wojciechowski, K. Oexle, M. Pirastu, X. Li, L. Raffel, M. Cotch, E.
Chew e B. Klein, «Meta-analysis of genome-wide association studies in five cohorts
reveals common variants in RBFOX1, a regulator of tissue-specific splicing,
associated with refractive error,» Hum Mol Genet, vol. 22, pp. 2754-2764, 2013.
[152] O. Strauss, «The retinal pigment epithelium in visual function,» Physiol Rev, vol.
85, pp. 845-881, 2005.
[153] P. Hysi, C. Simpson, Y. Fok, D. Gerrelli, A. Webster, S. Bhattacharya, C. Hammond,
P. Sham e J. Rahi, «Common polymorphisms in the SERPINI2 gene are associated
with refractive error in the 1958 British Birth Cohort,» Invest Ophthalmol Vis Sci,
vol. 53, pp. 440-447, 2012.
[154] P. Esteve, F. Trousse, J. Rodriguez e P. Bovolenta, «SFRP1 modulates retina cell
differentiation through a beta-catenin-independent mechanism,» J Cell Sci, vol.
116, pp. 2471-2481, 2003.
[155] M. Buraczynska, A. Swatowski, D. Markovska-Gosik, A. Kuczmaszewska e A.
Ksiazek, «Transcriptor factor 7-like 2 (TCF7L2) gene polymorphism and
complication/comorbidity in type 2 diabetes patients,» J Diab Res, vol. 93, pp. 390-
395, 2011.
[156] T. Hayashi, H. Inoko, R. Nishizaki, S. Ohno e N. Mizuki, «Exclusion of transforming
growth factor-b1 as a candidate gene for myopia in the japanese,» Jpn J
Ophthalmol, vol. 51, pp. 96-99, 2007.
[157] H. Lin, L. Wan, Y. Tsai, Y. Tsai, S. Fan, C. Tsai e F. Tsai, «the TGFbeta1 gene codon
10 polymorphism contributes to the genetic predisposition to high myopia,» Invest
Ophthalmol Vis Sci, vol. 12, pp. 698-703, 2006.

84
[158] Y. Zha, K. Leung, K. Lo, W. Fung, P. Ng, M. Shi, M. Yap e S. Yip, «TGFB1 as a
susceptibility gene for high myopia: a replication study with new findings,» Arch
Ophthalmol, vol. 127, pp. 541-548, 2009.
[159] C. Khor, Q. Fan, L. Goh, D. Tan, T. Young, Y. Li, M. Seielstad, D. Goh e S. Saw,
«Support for TGFB1 as a susceptibility gene for high myopia in individuals of
Chinese descent,» arch Ophthalmol, vol. 128, pp. 1081-1084, 2010.
[160] R. Nishizaki, M. Ota, H. Inoko, A. Meguro, T. Shiota, E. Okada, J. Mok, A. Oka, S.
Ohno e N. Mizuki, «New susceptibility locus for high myopia is linked to the
uromodulin-like 1 (UMODL1) gene region on chromosome 21q22.3.,» Eye, vol. 23,
pp. 222-229, 2009.
[161] D. Mutti, M. Cooper, E. Dragan, L. Jones-Jordan, M. Bailey, M. Marazita e J.
Murray, «Vitamin D receptor (VDR) and group-specific component (GC, vitamin D-
binding protein) polymorphisms in myopia,» Invest Ophthalmol, vol. 52, pp. 3818-
3124, 2011.
[162] C. Garcia-Frigola, M. Carreres, C. Vegar, C. Mason e E. Herrera, «Zic2 promotes
axonal divergence at the optic chiasm midline by EphB1-dependent and -
independent mechanisms,» Development, vol. 135, pp. 1833-1841, 2008.
[163] Y. Shi, Y. Li, D. Zhang, H. Zhang, F. Lu, X. Liu, F. He, B. Gong, L. Cai, R. Li, S. Liao, S.
Ma, H. Lin, J. Cheng, H. Zheng, Y. Shan, B. Chen, J. Hu, X. Jin, P. Zhao, Y. Chen e e.
al., «Exome sequencing identifies ZNF644 mutations in highmyopia,» PLoS Genet,
2011a.
[164] K. Tran-Viet, E. St Germain, V. Soler, C. Powell, S. Lim, T. Klemm, S. Saw e T. Young,
«Study of a US cohort supports the role of ZNF644 and high-grade myopia
susceptibility,» Mol Vis, vol. 12, pp. 937-944, 2012.
[165] www.geneontology.org, «Gene Ontology Consortium,» [Online].
[166] www.genome.jp/kegg/pathway, «http://www.genome.jp/kegg/pathway.html,»
[Online].
[167] J. Storey e R. Tibshirani, «Statistical significance for genome-wide studies,»
Proceeding of the national academy of sciences, vol. 100, pp. 9440-9445, 2003.
[168] H. Hayashi, K. Yamashiro, H. Nakanishi, I. Nakata, Y. Kurashige, A. Tsujikawa, M.
Moriyama, K. Ohno-Matsui, M. Mochizuki e M. Ozaki, «association of 15q14 and
15q25 with high myopia in Japanese,» Invest Ophthalmol Vis Sci, vol. 52, pp. 4853-
4858, 2011.

85
[169] X. Jiao, P. Wang, S. Li, A. Li, X. Guo, Q. Zhang e J. Hejmancik, «Association of
markers at chromosome 15q14 in Chinese patients with moderate to high
myopia,» Mol Vis, vol. 18, pp. 2633-2646, 2012.
[170] V. Verhoeven, P. Hysi, S. Saw, V. Vitart, A. Mirshahi, J. Guggenheim, M. Cotch, K.
Yamashiro, D. Baird e P. Mackey, «Large scale international replication and meta-
analysis study confirms association of the 15q14 locus with myopia. The CREAM
consortium,» Hum Genet, vol. 131, pp. 1467-1480, 2012.
[171] D. Supp, D. Wittte, S. Potter e M. Brueckner, «Mutation of an axonemal dynein
affects left-right asymmetry in inversus viscerum mice,» Nature, vol. 389, pp. 963-
966, 1997.
[172] S. Jones, C. Jomary, J. Grist, H. Stewart e M. Neal, «Modulated expression of
secreted frizzled-related proteins in human retinal degeneration,» Neuroreport,
vol. 11, pp. 3963-3967, 2000.
[174] F. P. H. J. e. a. Wong TY, «Prevalence and risk factors for refractive errors in adult
chinese in Singapore,» Invest Ophthalmol Vis Sci, pp. 2486-2494, 2000.
[175] H. B. W. D. S. G. N. K. R. S. e. a. Fricke TR, «Global cost of correcting vision
impairment from uncorrected refractive error,» Bull world Health Organ, pp. 728-
738, 2012.
[176] H. Nakanishi, R. Yamada, N. Gotoh, H. Hayashi, K. Yamashiro, N. Shimada, K. Ohno-
Matsui, M. Mochizuki, M. Saito, T. Iida, K. Matsuo, K. Tajima, N. Yoshimura e F.
Matsuda, «A genome-wide association analysis identified a novel susceptible locus
for pathological myopia at 11q24.1,» Plos Genet, 2009.
[177] E. Goldschimdt e N. Jacobsen, «Genetic and environmental effects on myopia
development and progression,» Eye, vol. 28, pp. 126-133, 2014.
[178] A. Kiefer, J. Tung, B. Do, D. Hinds, J. Mountain, U. Francke e N. Eriksson, «Genetics
of Myopia in a Participant driven, web-based cohort,» 23andMe, 2012.
[179] S. Cheng, C. Lam e M. Yap, «Prevalence of myopia-related retinal changes among
12-18 year old Hong Kong Chinese high myopes,» Ophthalmic PHysiolo Opt, vol.
33, pp. 652-660, 2013.
[180] P. Concannon, S. Rich e G. Nepom, «Genetics of type 1A diabetes,» The New
England Journal of Medicine, vol. 360, pp. 16-46-1654, 2009.
[181] P. Greene, «Mechanical considerations i myopia: relative effects of accomodation,
convergence, intraocular pressure, and extraocular muscles,» American Journal Of

86
Optom and Physiol Optics, vol. 57, pp. 902-914, 1980.
[182] B. Holde, T. Fricke e S. Ho, «Global vision impairment due to uncorrected
presbyopia,» Arch Ophthalmol, vol. 126, pp. 1731-1739, 2008.
[183] G. Hung e K. Ciuffreda, «Model of human refractive error development,» Curr Eye
Res, vol. 19, pp. 41-52, 1999.
[184] D. Lam, W. Lee, Y. Leung, P. Tam, D. Fan, B. Fan e C. Pang, «TGFbeta-induced
factor: a candidate gene for high myopia.,» Invest Ophthalmol Vis Sci, vol. 44, pp.
1012-1015, 2003.
[185] H. Nakanishi, R. Yamada, N. Gotoh, H. Hayashi, A. Otani, A. Tsujikawa, K.
Yamashiro, N. Shimada, K. Ohno-Matsui, M. Mochizuki, M. Saito, K. Saito, T. Iida, F.
Matsuda e N. Yoshimura, «Absence of association between COL1A1
polymorphisms and high myopia in the Japanese population,» Invest Ophthalmol
Vis Sci, vol. 50, pp. 544-550, 2008.
[186] R. Rose, C. Harper, C. Tromans e C. Waterman, «Quality of life in myopia,» Br J
Ophthalmol, vol. 84, pp. 1031-1034, 2000.
[187] T. Smith, K. Frick, B. Holden, T. fricke e K. Naidoo, «Potential lost productivity
resulting from the global burden of uncorrected refractive error,» Bull world
health organ, vol. 87, pp. 431-437, 2009.
[188] S. Veerappan, M. Schache, K. Pertile, F. Islam, C. Chen, P. Mitchell, M. Dirani e P.
Baird, «The retinoic acid receptor alpha (RARA) gene is not associated with
myopia, hypermetropia, and ocular biometric measures,» Mol Vis, vol. 15, pp.
1390-1397, 2009b.
[189] R. Wojciechowski, «Nature and nurture: the complex genetics of myopia and
refractive error,» Clin Genet, vol. 79, pp. 301-320, April 2011.
[190] G. Zhu, A. Hewitt, J. Ruddle, L. Kearns, S. Brown e J. MacKinnon, «Genetic
dissection of myopia: evidence for linkage of ocular axial lenght to chromosome
5q,» Ophthalmology, vol. 115, pp. 1053-1057, Jun 2008.
[191] D. Hornbeak e T. Young, «Myopia genetics: a review of current research and
emerging,» Curr Opin Ophthalmol, vol. 20, pp. 356-362, september 2009.
[192] P. Foster e Y. Jiang, «Epidemiology of myopia,» Eye, vol. 28, pp. 202-208, 2014.

87


















