Studio biochimico dellattivazione dei recettori Le molecole segnale idrofiliche non possono superare...
30
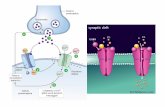
udio biochimico dell’attivazione dei recetto Le molecole segnale idrofiliche non possono superare il doppio strato lipidico della membrana cellulare. Il sito recettoriale delle molecole idrosolubili è situato sulla parte extracellulare di recettori transmembrana. Molecole segnale lipofiliche attraversano facilmente la membrana cellulare: i loro recettori sono quindi localizzati all'interno della cellula.
-
Upload
gioacchino-ferretti -
Category
Documents
-
view
219 -
download
0
Transcript of Studio biochimico dellattivazione dei recettori Le molecole segnale idrofiliche non possono superare...

Studio biochimico dell’attivazione dei recettori
Le molecole segnale idrofiliche non possono superare il doppio
strato lipidico della membrana cellulare. Il sito recettoriale
delle molecole idrosolubili è situato sulla parte
extracellulare di recettori transmembrana.
Molecole segnale lipofiliche attraversano facilmente la
membrana cellulare: i loro recettori sono quindi localizzati
all'interno della cellula.

Differenti tipi di recettori
- Legati a canali ionici
- Legati alla proteina G
- Recettori steroidei
- Recettori legati ad enzimi

Recettori legati a canali ionici con effetto di tipo:
eccitatorio acetilcolina, glutammato, serotonina(apertura dei canali Na+ con
conseguente depolarizzazione)
inibitorio Gaba, glicina(apertura dei canali Cl- con
conseguente iperpoplarizzazione)

Recettori steroidei
Gli steroidi sono molecole
con elevata lipofilia, in
grado di attraversare le
membrane plasmatiche e
a livello nucleare
legandosi a recettori
specifici (DNA-binding
protein) regolano la
trascrizione di un gene
adiacente al legame

Quando attivati funzionano direttamente come enzimi (RTK) o sono associati ad enzimi (PTK)
Recettori legati ad enzimi

Il ligando endogeno (1) lega specificamente il recettore (2). Viene attivato il sistema G-protein (3). Viene attivato l’adenilato ciclasi (4) che produce il secondo messaggero cAMP (5) a partire dall’ATP (6). Avvengono una serie di reazioni enzimatiche (7) e via fosforilazione (8) il glicogeno (9) viene trasformato in glucosio (10). La fosforilazione può interessare proteine di membrana come canali ionici (11).
Recettori legati alla G-protein

Attivazione della Proteine G
La G-protein è composta da tre subunità legata quando non attivata al GDP
L’interazione ligando recettore attiva la G-protein liberando GDP...
..legando GTP. Il trimero si suddivideIl GTP legato ad viene idrolizzato a GDP+ P. Le subunità si ricombinano.

Gq
R
GTP
GDP
+ -
GTP GDP
Gi RAC
ATP cAMP
R Gs
GTP GDP
PK-A+
Ca++ RS
+Ca++
Ca++
PL-C
PIP2 IP3 +
DAG PK-C+

Sfruttando il meccanismo di attivazione illustrato si può
determinare l’attività agonista, agonista inverso, agonista
parziale, antagonista.
Utilizzando GTPS35, analogo non idrolizzabile del GTP, si può
misurare l’attività di un ligando in base all’accumulo di
GTPS35.
Nel caso in cui il composto sia antagonista è necessario
realizzare la curva di attività in presenza di un agonista pieno e
verificare lo spostamento parallelo a destra a dosi crescenti di
antagonista.
CH2O P O P O P
O O 35S
OH
OHOHOHO
N
N N
N
O
H
NH2
OHOH

10 -11 10 -10 10 -9 10 -8 10 -7 10 -6 10 -5
0
50
100Agonista pieno
Agonista parziale
Agonista inverso
log dose
Bo
un
d
(GT
P
S35
)

cAMP
La reazione è basata sulla competizione per l’anti-anticorpo tra
cAMP legato alla fosfatasi alcalina e quello libero proveniente
dalla stimolazione recettoriale
Dosaggio del secondo messaggero
APAP
AP
AP+ S
S
S
S

cAMP coniugato alla fosfatasi alcalina
Anticorpo primario legato al pozzetto
cAMP indotto dall’attivazione
recettoriale
del ligando
Anticorpo secondario
La quantità di cAMP cellulare è inversamente proporzionale
alla quantità di colorazione gialla svolta.
AP

Attivazione di PK-A
Glucagone
Recettore
G-Protein(, , , subunità)
PK-A inattivaPK-A
Adenilato ciclasi(inattivo)
G-Protein (, subunità)
Fosforilazione di substrati

Substrato PK-A-R110 (bisammide Rhodamina 110 peptide)
PK-A R110non fluorescente
PK-A
PK-A R110fluorescente
PK-A R110non fluorescente fosforilato
proteasi proteasi
non fluorescente
Eccitazione 485 nm Emissione 530 nM

% peptide fosforilato
0 10 20 30 40 50 60 70 80 90 100
FLU
100,000
50,000
0
L’intensità di fluorescenza misurata nel saggio è inversamente correlata all’attività della PK-A

Dosaggio del Ca++ intracellulare
IP2
DAGIP3
Calmodulina-proteina Kinasi (inattivoa)
Calmodulina-proteina Kinasi
Proteina kinasi C
Proteina kinasi C inattiva
Gq-
REL

Saggio dell’Aequorina
L’Aequorina è una fotoproteina riscontrata nella medusa
Aequorea Victoria.
L’apoaequorina è una proteina di 21 KDalton e necessita di un
gruppo prostetico idrofobico coeletranzina per essere convertita
nella forma enzimatica attiva: aequorina.
Dosaggio del Ca++

In presenza di Ca++ aequorina ossida la coeletranzina in
coelentramide con produzione di CO2, ed emissione di luce.
Questi saggi vengono effettuati in linee cellulari stabili in grado
di esprimere il GPCR ed apo-aequorina

Sistemi di modulazione del segnale
Oltre a sistemi di attivazione recettoriale e di amplificazione del segnale sono presenti meccanismi di interruzione del segnale
•Autoregolazione del rilascio del neurotrasmettitore (feedback negativo)
Questo meccanismo si realizza mediante recettori
presinaptici che possono essere controllati dallo stesso
neurotrasmettitore (autorecettori) o da un
neurotrasmettitotre differente (eterorecettori)

•Sistemi enzimatici preposti alla degradazione
•Sistemi di recupero del neurotrasmettitore
•Desensitizzazione recettoriale (in recettori accoppiati alla G-protein)
La desensitizzazione può essere:
• omologa (specifica per un recettore) oppure eterologa (non specifica);• rapida (~ 20 minuti) oppure lenta (giorni);• perdita funzione recettoriale (uncoupling) o con diminuzione dei recettori (down-regulation)

• Uncoupling del recettore (desensitizzazione rapida)
La fosforilazione non altera la capacità del recettore di attivare la G-protein ma aumenta l’affinità del recettore fosforilato per la proteina -arrestina che inibisce l’interazione del recettore con la G-protein determinando il disaccoppiamento).

• Uncoupling del recettore (desensitizzazione lenta)
Molti sistemi recettoriali sono fosforilati dai loro stessi enzimi effettori PKA e PKC
Questo tipo di fosforilazione è un feedback diretto negativo che al tempo stesso sopprime l’attivazione dell’enzima effettore PKA.
Questo meccanismo di desensitizzazione è eterologo in quanto l’attivazione di PKA PKC è sufficiente a determinare la fosoforilazione del recettore.
La desensitizzazione da PKA è più lenta ma è molto più sensibile alla concentrazione di agonista.

Saggi funzionali su organo isolato
In questi esperimenti in seguito ad uno stimolo (farmacologico o
elettrico) deve essere apprezzabile e misurabile una risposta in
un preparato biologico
variazioni del volume del preparato biologico che viene
valutato mediante semplici registratori a galleggiante
variazione di lunghezza o di tensione del preparato biologico
L’effetto che si dovrà valutare può essere rappresentato da:
variazioni di pressione


La stimolazione del preparato biologico può essere effettuata
Elettricamente
Farmacologicamente
In alcuni esperimenti può essere necessario stimolare
elettricamente il preparato. La forma e la durata degli impulsi
sono parametri importanti soprattutto in preparati
neuromuscolari.
Le concentrazioni di farmaco necessarie in questo tipo di
esperimento sono molto basse anche perché quando impiegati
a dosaggi elevati sono dotati di vari effetti ed è quindi
importante scegliere quelle concentrazioni alle quali agiscono
in modo specifico.

Esperimenti farmacologici
Qualitativi
Quantitativi
Studio del meccanismo di azioneQuantizzazione
dell’attività caratterizzata
Agonistiproducono effetti attivi o diretti sui preparati biologici
Antagonisti
producono effetti negativi o indiretti e inibizione o alterazione
degli effetti degli agonisti

Curva dose-risposta di un agonista pieno
Eff
ett
o m
ass
imo
pe
rce
ntu
ale
Log dose
Effetto massimo
In rosso la curva dose-risposta dell’agonista.
In nero le curve dose risposta dell’antagonista in presenza
di dosi crescenti di antagonista.

dose dose
carbacolo 1 M carbacolo 1 M in presenza di 0.1 M di atropina
Risposta dell’ileo di cavia ad una dose di carbacolo

Impulso elettrico Contrazione
Attività pre- post- giunzionale
Agonista pre-
Un agonista pre- è in grado di ridurre il rilascio di mediatore chimico successivo alla stimolazione elettrica.Es: 8-OH-DPAT sui recettori serotoninergici 5-HT1A nella vescica
o ileo di cavia

Agonista post-
Un agonista post è in grado di attivare autonomamente l’effetto biologico
Es: carbacolo su recettori muscarinici nell’ ileo di cavia