Scambi gassosi e trasporto gas - lucibello.me.it · La velocità di diffusione di un gas dipende...
59
Transcript of Scambi gassosi e trasporto gas - lucibello.me.it · La velocità di diffusione di un gas dipende...


Gli scambi gassosi avvengono per diffusione e sono dovuti esclusivamente a gradienti di pressione parziale dei singoli gas fra sangue ed alveoli

I gas quindi diffondono dal luogo in cui la pressione è maggiore verso il luogo a pressione minore e cioè dagli alveoli al sangue e viceversa.

L’ossigeno diffonde dall’alveolo al sangue venoso passando da una pressione di 100 mm/Hg alla pressione di 40 mm/Hg, mentre la CO2, al contrario, nel sangue venoso ha una pressione di 46 mm/Hg e passa negli alveoli dove la pressione è di 40 mm/Hg.

Il gradiente pressorio dell’O2 è quindi di 60 mm/Hg, mentre quello della CO2 di soli 6 mm/Hg; ciò è dovuto al fatto che la CO2 è 25 volte più solubile e 20 volte più diffusibile dell'O2, pertanto anche se il gradiente pressorio è decisamente inferiore, la velocità dello scambio gassoso sarà maggiore per la CO2 rispetto all’O2.
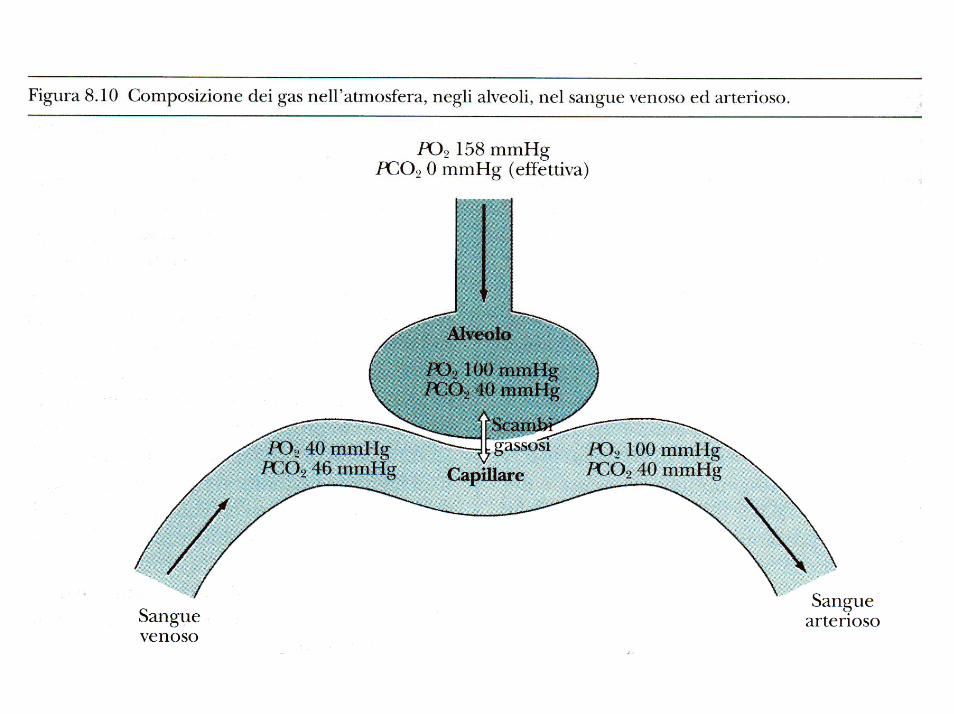

La pressione parziale della CO2 nell’alveolo è uguale a quella del sangue che abbandona i polmoni, cioè40 mm/Hg per cui si raggiunge un perfetto equilibrio;
per l’O2, invece, la pressione parziale negli alveoli èdi 100 mm/Hg, mentre quella che si ha nelle vene polmonari è 97 mm/Hg. Ciò è dovuto al fatto che il sangue che arriva ai bronchi dalle arterie bronchiali non va incontro ad ossigenazione nei capillari polmonari e ritorna al cuore non ossigenato (shunt fisiologico).

La velocità di diffusione di un gas dipende dai seguenti fattori:
Gradiente pressorio solubilità gas areaVelocità di diffusione = K -----------------------------------------------------------------
____________√Densità del gas spessore parete alveolare
K = cost. = 0,139

Oltre alla differenza di pressione parziale dei gas fra sangue e tessuto, nello scambio sono importanti:
•la solubilità del gas, •la densità, •lo spessore della parete •l’area

Bisogna inoltre considerare:
•la superficie di scambio,•la lunghezza del percorso diffusionale,•la resistività diffusionale,•la velocità del flusso sanguigno attraverso i capillari.
Le prime due dipendono dal numero dei capillari perfusi (entità della capillarizzazione).

La membrana respiratoria è costituita:
1) da uno strato di liquido che riveste l’alveolo e che contiene il fattore surfactante il cui compito è quello di abbassare la tensione superficiale (per il principio di La Place) nell’alveolo stesso,
2) dall’epitelio alveolare, 3) dalla membrana basale,4) da un piccolo spazio interstiziale tra l’alveolo e la parete
capillare, 5) dalla membrana basale del capillare e 6) dall’endotelio del capillare.


Lo spessore della membrana respiratoria, nonostante sia formata da molti strati, in alcuni punti è di circa 0,2 µm, mentre l’area totale della superficie di scambio, in un adulto, è di circa 70 -100 m2; e poiché la quantità di sangue, presente nei capillari polmonari, (60 e i 140 ml ad ogni istante) deve essere distribuita all’intera superficie ci si spiega del perché gli scambi avvengano tanto rapidamente.


Il trasporto dei gas metabolicamente attivi comprende processi sia fisici che chimici.

L’ossigeno è trasportato dal sangue in due modi:
1) fisicamente disciolto nel plasma2) legato all’emoglobina degli eritrociti.
Il primo è trascurabile (0,3 ml% nel sangue arterioso, pO2 = 100 mm Hg, 0,13 ml% nel sangue venoso, pO2 = 40 mmHg) e dipende dalla solubilità del gas, dalla temperatura e dalla sua pressione parziale nel sangue in accordo con la legge di Henry.
Per cui quasi tutto il potere ossiforico del sangue è dovuto all’emoglobina alla quale l’O2 si lega labilmente.

L’emoglobina è un cromoprotide capace di legare, al gruppo eme, labilmente e reversibilmente l’ossigeno, quando la pO2 è elevata l’ossigeno si combina con l’emoglobina, quando la pO2 si abbassa, come avviene nei tessuti periferici, l’ossigeno viene ceduto.
1 gr di emoglobina fissa 1,34 ml di ossigeno e poichénel sangue si trovano 14, 15 gr di Hb%, moltiplicando si ottengono 19, 20 volumi di ossigeno, cioè quello trasportato dagli eritrociti in 100 cc di sangue.


E’ possibile costruire un diagramma cartesiano in cui si pone la pressione parziale dell’ossigeno sulle ascisse e la percentuale di saturazione dell’emoglobina con l’ossigeno sulle ordinate.
Tale diagramma prende il nome di curva di dissociazione dell’emoglobina, ed assume una forma caratteristica ad S italica.


Le relazioni tra pressione parziale dell'ossigeno ed entità dell'ossigenazione dell'emoglobina possono essere indagate costruendo un diagramma cartesiano con la pressione parziale dell'02 sull'asse dell’ascissa e la percentuale di saturazione dell’Hb con ossigeno sull'ordinata.
Si afferma che l’Hb è completamente satura di ossigeno quando è tutta convertita in ossiemoglobina. Se invece coesistono emoglobina ridotta ed ossiemoglobina, allora si parla di saturazione parziale.

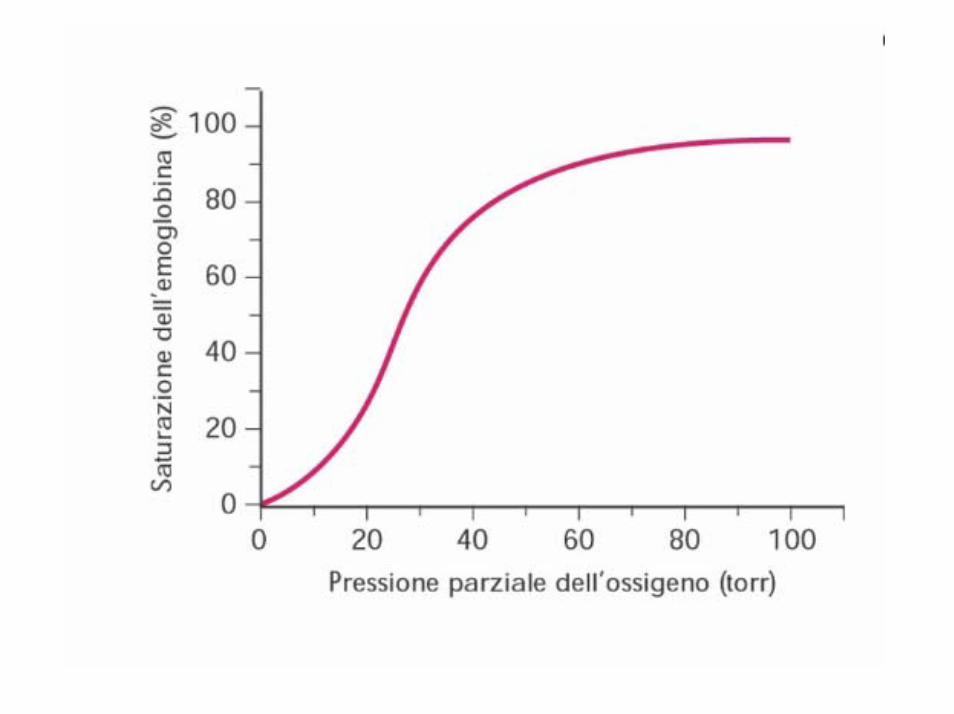

Quando la pO2 è bassa la pendenza è ripida, piatta o livellata quando la pO2 è alta. L’affinità dell’emoglobina per l’O2 aumenta con l’ossigenazione.
La parte superiore della curva, relativamente piatta, può essereconsiderata la parte associativa, cioè la parte che riguarda l’assunzione di O2.
La parte più bassa della curva, ripida, può essere considerata la parte dissociativa, cioè la parte che riguarda la liberazione di O2 dall’Hb.

Ad una pO2 di 100 mm/Hg l’Hb è saturata al 97,4%.
Quando la pO2 si abbassa fino a circa 60 mm/Hg l’emoglobina del sangue arterioso è ancora saturata per l’89%.
Ciò spiega come sia possibile la vita alle grandi altezze, dove la pO2 tende a diminuire senza una riduzione importante di O2 legato all’Hb. Ad una pO2 compresa tra 10 e 40 la curva è, invece, molto ripida e minime variazioni di pO2 provocano il distacco di relativamente grandi quantitàdi O2.

Ossigenoterapia iperbarica
Somministrazione terapeutica di ossigeno a pressione superiore aquella atmosferica. Scopo di questa forma di terapia è di aumentare
la pressione parziale dell’ossigeno (aumentando la pressione assoluta del gas inalato) per facilitarne il passaggio nel sangue e
nei globuli rossi.
A un regime pressorio di 3 atmosfere, la quantità di ossigeno presente in forma disciolta nel plasma (non legato quindi
all’emoglobina) corrisponde a circa 20 volte quella che si ha in condizioni di pressione normale.

Attualmente le principali indicazioni per l’ossigenoterapia iperbarica sono costituite dall’intossicazione da monossido di carbonio, dalle necrosi ossee e dall’osteomielite, dalla gangrena gassosa (da Clostridium perfringens), dalle lesioni nervose e midollari (traumi, polinevriti ecc.).


La CO2 rappresenta il prodotto finale del metabolismo aerobico, e viene quindi eliminata, come prodotto di rifiuto, attraverso ipolmoni. Essa viene trasportata dal sangue (diffondendo da questo agli alveoli grazie a gradienti di pressione) sotto diverse forme:
1. disciolta fisicamente nel plasma (5-8%), 2. sotto forma di acido carbonico, 3. come bicarbonati di sodio nel plasma, di potassio nelle
emazie, 4. sotto forma di carbamino- composti.

La quantità di CO2 disciolta è di circa 2,7 ml per 100 ml di sangue e la pCO2 è di 46 mm/Hg. Ad una pCO2di 40 mm/Hg la quantità di gas disciolto è circa 2,4 ml.

La CO2 disciolta nel sangue reagisce con l’acqua per formare acido carbonico, grazie all’enzima anidrasi carbonica presente nel globulo rosso. L’acido carbonico si dissocia in idrogenioni ed in bicarbonationi.
Molti ioni idrogeno si combinano con l’emoglobina deossigenata nei globuli rossi mentre molti ioni bicarbonato diffondono dai globuli rossi nel plasma, sostituiti da cloro ioni i quali compiono il percorso inverso (passaggio dei cloruri o fenomeno di Hamburger).
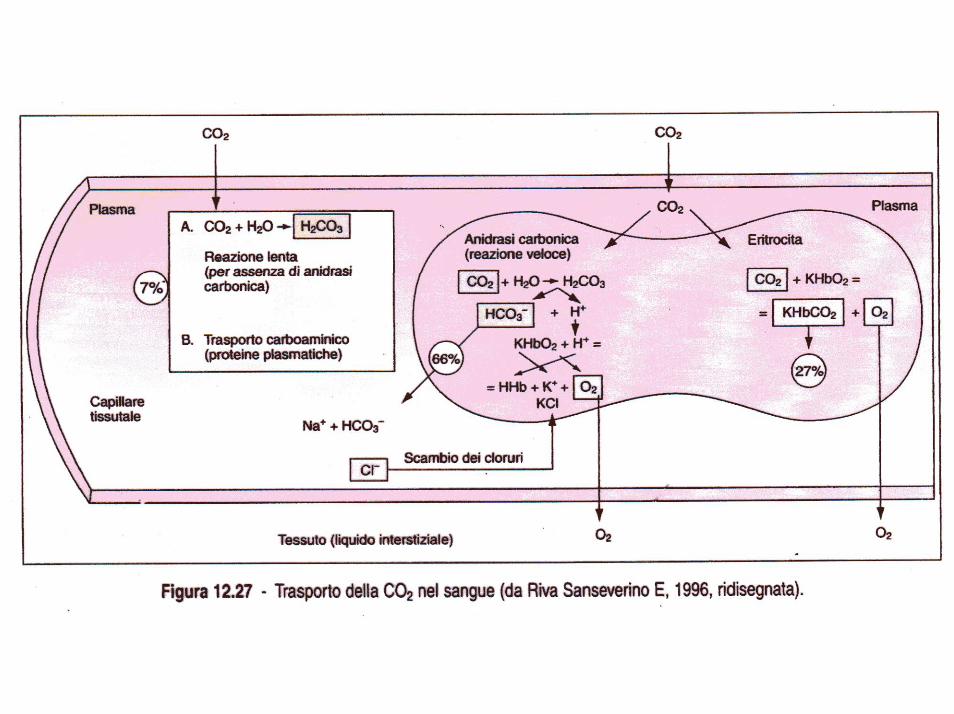

Altra possibilità dell’anidride carbonica è quella di combinarsi all’interno del globulo rosso con la parte protidica dell’emoglobina e cioè con la globina formando carbaminoemoglobina, e in minor quantità, con le proteine plasmatiche.

Anche per la CO2 esiste una curva di dissociazione. Essa è importante perché così come il distacco dell’ossigeno dall’emoglobina facilita il passaggio di ossigeno ai tessuti, il legame dell’ossigeno con l’emoglobina tende ad eliminare la CO2 dal sangue.

Questo effetto è detto effetto Haldane ed è dovuto al fatto che il legame dell’ossigeno con l’Hb rende l’emoglobina più acida e ciò facilita l’eliminazione della CO2 con due meccanismi:
1) l’Hb più acida ha una minore affinità per la CO2 la quale viene quindi eliminata;
2) l’aumentata acidità dell’emoglobina provoca il rilascio di un maggior numero di idrogenioni i quali si combinano con i bicarbonationi del plasma formando acido carbonico; in seguito questo dissocia in H2O e CO2 liberando quest’ultima negli alveoli.



Una corretta attività respiratoria garantisce l’omeostasi gassosa, cioè le concentrazioni ideali di O2 e CO2 nel sangue arterioso.

Ogni qualvolta si hanno modificazioni della composizione dell’aria inspirata, con conseguenti variazioni delle rispettive pressioni parziali dei gas nell’aria alveolare e a livello ematico, viene informato, tramite i chemocettori o direttamente, tramite alcuni nervi cranici (vago e glossofaringeo), il centro bulbare il quale per via riflessa invia segnali lungo il midollo spinale ai centri cervicali e toracici dei nervi che raggiungono i muscoli respiratori affinchè venga modificato il respiro.
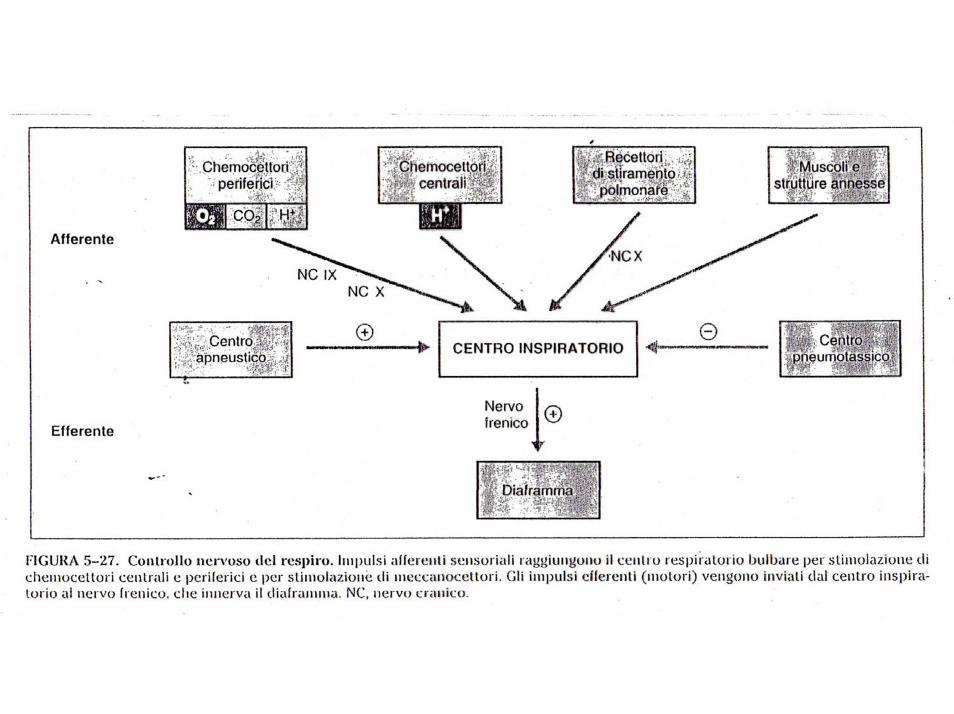

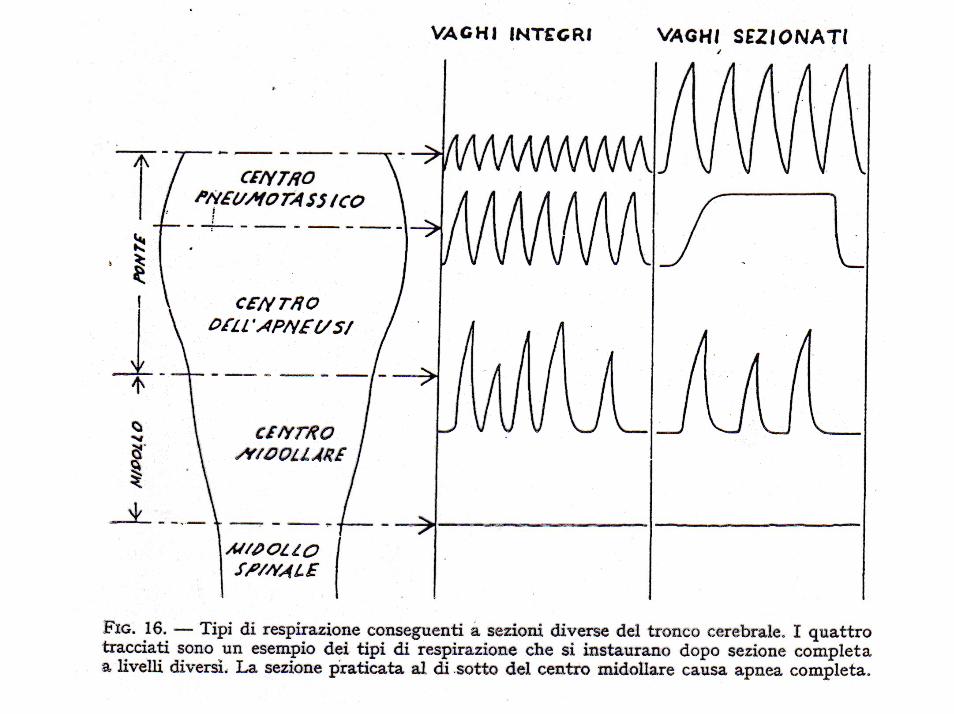
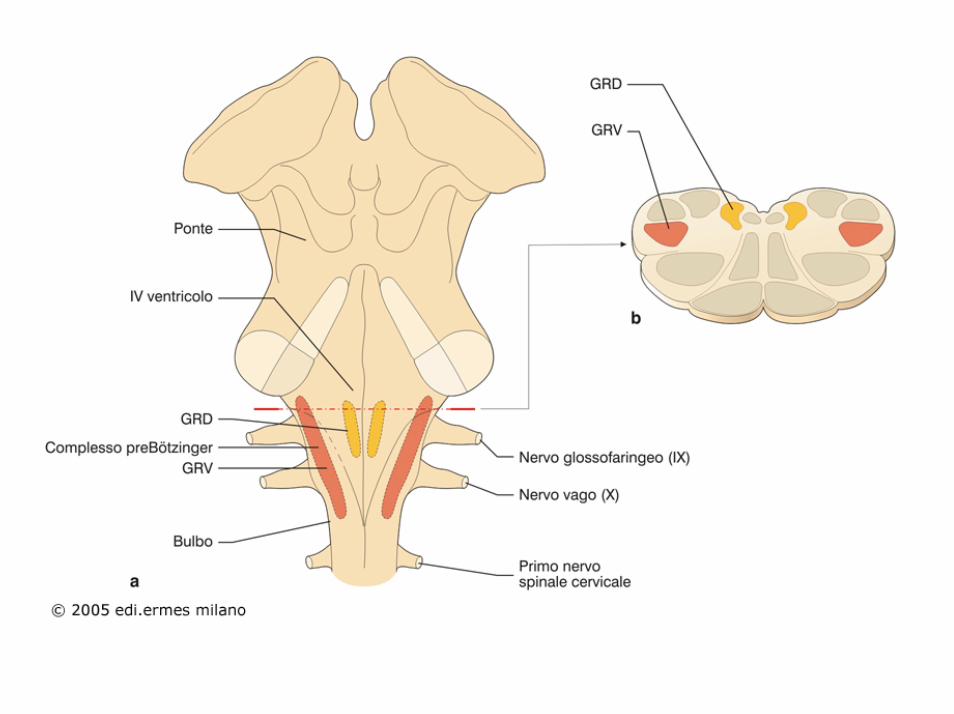
Il centro respiratorio è costituito da diversi gruppi di neuroni situati nella formazione reticolare del bulbo e del ponte.

Si distinguono in particolare quattro gruppi di neuroni:
1. un gruppo respiratorio dorsale, situato nella parte dorsale del bulbo a funzione inspiratoria,
2. un gruppo respiratorio ventrale, situato nella parte ventrolaterale del bulbo, a funzione sia inspiratoria che espiratoria,
3. il centro apneustico, situato nella parte caudale del ponte, capace di prolungare l’atto inspiratorio
4. il centro pneumotassico, situato nella parte superiore del ponte, che ha il compito di controllare la frequenza del respiro e di limitare la durata dell’inspirazione.


Il centro respiratorio è assai sensibile alle variazioni delle concentrazioni di ossigeno, anidride carbonica ed idrogenioni. In particolare un eccesso di CO2 o di idrogenioni agisce direttamente sul centro respiratorio, causando un aumento dei segnali sia stimolatori, sia inibitori.
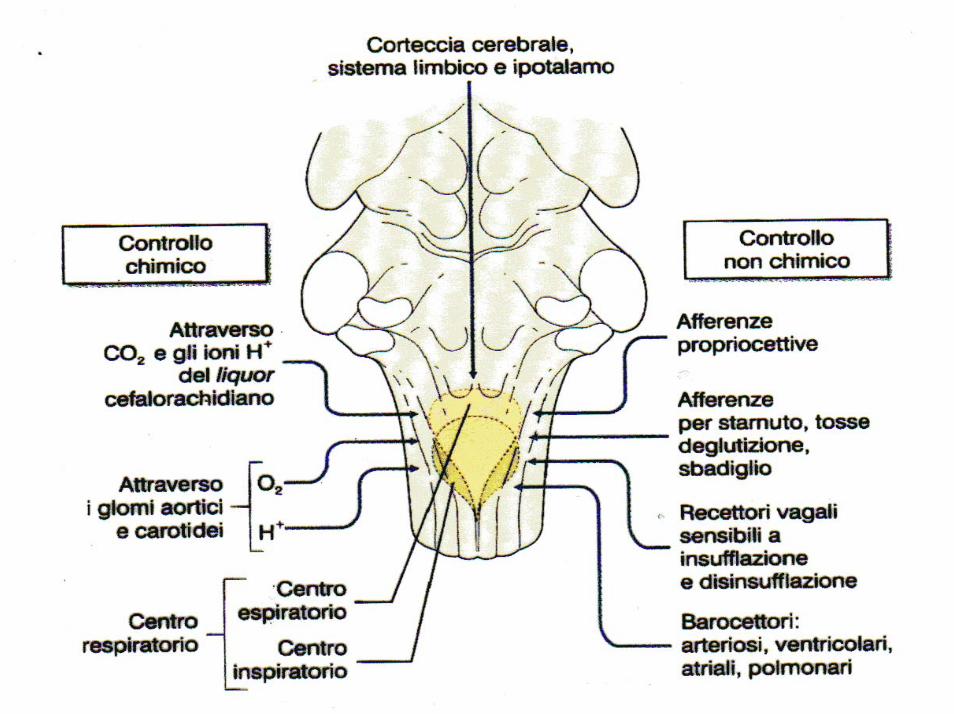

L’O2, pur non avendo un effetto diretto sul centro, agisce sui chemocettori dei glomi aortici e carotidei, i quali inviano segnali al centro respiratorio per la regolazione del respiro.
In realtà nessuno dei gruppi neuronali (gruppo dorsale, gruppo ventrale, centro apneustico e centro pneumotassico) menzionati è in grado di captare le variazioni di CO2 e di idrogenioni, per cui esiste un’area neuronale chemocettrice localizzata bilateralmente nel bulbo molto sensibile alle variazioni di CO2 e di idrogenioni.

I neuroni dell’area chemocettrice sono eccitati principalmente dagli idrogenioni i quali, però, non oltrepassano le barriere emato-encefalica ed emato-liquorale anche se nel liquor, in parte per la scarsità del potere tamponante, in parte per l’assenza di barriera liquor-interstizio essi agiscono rapidamente.

La CO2 non agisce direttamente, ma poiché reagisce con l’acqua tissutale formando acido carbonico che si dissocia in bicarbonationi ed in idrogenioni, tramite questi ultimi esercita un potente effetto stimolatorio.

Gli idrogenioni a livello ematico, invece, vengono facilmente tamponati dai sistemi tampone del sangue, ed è questo il motivo per cui è più efficace l’aumento ematico della concentrazione di CO2 piuttosto che quello della concentrazione ematica di idrogenioni.

L’eccitazione del centro respiratorio da parte della CO2 èmolto efficace all’inizio, ma poi si attenua nei successivi due giorni poiché i reni aumentano la concentrazione di bicarbonati nel sangue, i quali si combinano con gli idrgenioni riducendone la concentrazione.

Le variazioni della concentrazione di O2 non hanno un effetto diretto sul centro respiratorio, ma quando la pressione parziale dell’ossigeno nel sangue arterioso scende al di sotto dei 60-70 mm Hg scattano i meccanismi periferici di controllo del respiro

Si tratta di speciali recettori, detti chemocettori, situati nei glomi aortici e carotidei, in particolare a livello dell’arco aortico e del seno carotideo, che rispondono alle variazioni di O2, CO2 e di idrogenioni e trasmettono segnali al centro respiratorio.
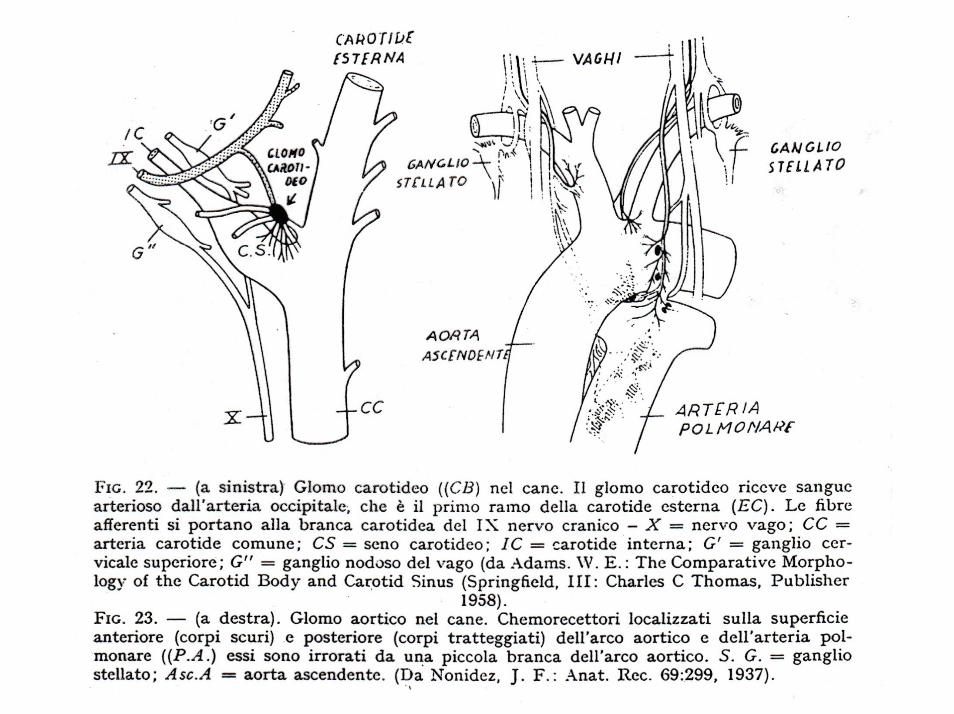

Questi chemocettori sono stimolati anche da concentrazioni elevate di CO2 (maggiore di 50 mm/Hg) e di idrogenioni e, anche se meno potenti, rispondono in maniera molto più rapida (5 volte più rapida) della stimolazione diretta centrale, per cui non possono essere considerati di secondaria importanza.

Per quanto riguarda le variazioni di O2, la risposta da parte del chemocettore è piuttosto debole perchècompensata dall’enorme flusso ematico e dall’effetto frenante dei meccanismi di regolazione del respiro attivati dalla pCO2 e dal pH (bassi pH esaltano il respiro, alti pH lo deprimono)

Se il flusso ematico è normale, i chemocettori non sono stimolati poiché la loro irrorazione è molto grande; se la pO2 scende al di sotto di 60 mm/Hg (a livello arterioso) vengono stimolati; se la pO2 è normale e la perfusione ridotta, come avviene nelle emorragie o nell’ipotensione, vengono stimolati.