
Nel Piu Profondo Dell'Anima - Lo Psichiatra Spirituale - Oscar Lepore

1
Gruppo Archeologico Albinetano
“Paolo Magnani”
STORIA EVOLUTIVA DELL’UMANITÀ Atti delle conferenze
dedicate al Prof. Paolo Magnani Relatore: dr Alberto Catalano
Giovedì 5 marzo 2015 Apparizione dei Primati. Ominidi Pre-Umani ancestrali.
Giovedì 12 marzo 2015
Ominidi Pre-Umani recenti.
Giovedì 19 marzo 2015 Popolazioni preistoriche scoperte ultimamente.

2
PRESENTAZIONE
In memoria del Prof. Paolo Magnani, scomparso nel 2013, il Gruppo Archeologico tutto ha voluto aggiungere
il suo nome a quello dell’associazione, a decorrere dal 2014. Inoltre intende dedicargli le serate di quest’anno
“ARGOMENTI DI ARCHEOLOGIA” ripercorrendo i temi che gli erano più cari.
Sarà relatore il dr Alberto Catalano – Medico Psichiatra - suo e nostro amico, socio del nostro Gruppo,
studioso in materia di evoluzione umana e strumenti litici preistorici.
PAOLO MAGNANI - (Reggio Emilia, 26 giugno 1926 – 7 agosto 2013)
Laureatosi in lettere classiche, entrò nel mondo della scuola e insegnò in diversi istituti superiori di Reggio
Emilia e Provincia.
L’ATTIVITA’ ARCHEOLOGICA
Suo principale interesse fu sempre l’archeologia, ma soprattutto i temi concernenti la storia evolutiva
dell’uomo. Ha collaborato agli scavi nei depositi paleolitici di Ghiardo di Bibbiano e alla terramara S. Rosa di
Poviglio, nonché alle ricerche della Società Reggiana di Archeologia. Per la sua competenza e il suo impegno
venne nominato Ispettore Onorario della Soprintendenza Archeologica dell’Emilia Romagna per il Comune di
Albinea.
Alla preistoria, ha dedicato i suoi studi come dimostrano le sue pubblicazioni:
- Preistoria di Reggio nell’Emilia (Ed. Nova et Vetera, 1993)
- Le terremare (con Renato Peroni, Ed. Nova et Vetera, 1996)
- Miti e riti della preistoria (con Lorenzo Facchini, Ed. Jaca Book, 2000)
- Gaetano Chierici - Tutti gli scritti di archeologia (Ed. Diabasis, 2007)
- Gaetano Chierici - Epistolario (Ed. Diabasis, 2010).
- Che cosa ci insegna la preistoria (La Nuova Tipolito, 2012)
Inoltre ha costantemente collaborato alla rivista “ Reggio Storia”.
IL GRUPPO ARCHEOLOGICO ALBINETANO
Nel Gruppo Archeologico Albinetano è entrato alla fine degli anni 90 e la sua presenza è stata determinante e fondamentale. E’ stato l’anima del Gruppo Archeologico, il leader carismatico, quello con le intuizioni e le idee più originali, sia proponendo azioni da intraprendere che suggerendo il modo migliore per realizzarle. Attento alle esigenze delle persone, capace di indirizzarne l’attività, in grado di rapportarsi con i problemi, interlocutore autorevole della Soprintendenza di cui è stato il tramite. Suo costante impegno è stato quello di sensibilizzare i giovani, ma non solo; per questo è stato relatore in numerose conferenze. Circa gli aspetti della sua personalità interpreto il pensiero di tutti i soci del Gruppo nel dire che aveva uno spirito vivace, ironico, attento alle novità, capace di restare al passo coi tempi, fondamentalmente giovane, mantenendo sempre però, nelle sue valutazioni, un atteggiamento obiettivo, critico, analitico, autonomo e ponderato. La mia personale conoscenza e frequentazione del Prof. Magnani risale ai primi anni 90, quando, sapendo che si occupava di archeologia l’ho contattato. Lui è stato per me un tutor d’eccezione. Da questo comune interesse è nato anche un rapporto sincero di amicizia e di stima che non si è mai interrotto. Nonostante la mia incompetenza in materia di preistoria, sono stata coinvolta ugualmente nella preparazione dei suoi primi libri e questo mi ha fatto molto piacere e resa orgogliosa e onorata per l’opportunità che mi offriva. Un altro, dei tanti ricordi che ho del Prof. Magnani, è quello della sua grande cultura letteraria e la conoscenza approfondita del latino a cui facevamo costantemente ricorso. Conosceva a memoria la Divina Commedia di Dante e più di una volta gli abbiamo chiesto di recitarne dei brani durante le camminate per raggiungere il Castello. I Canti dell’Inferno erano i preferiti e i più citati, questo ci forniva l’occasione per fare confronti e battute trovando dei collegamenti sui temi delle nostre indagini archeologiche o sugli eventi del momento. In conclusione, ricordandolo, a nome di tutti con affetto e rimpianto, leggo un brano da : “Cosa ci insegna l’archeologia”, il cui contenuto rispecchia il suo pensiero sul momento attuale che stiamo vivendo.

3
“La conoscenza del passato, anche del più lontano passato, può illuminare la lunga vicenda dell’ umanità da ogni punto di vista scientifico, può aiutare a conoscere sempre meglio noi stessi, i retaggi che ci condizionano, le cause dei nostri attuali problemi esistenziali. Ascoltare la voce dei millenni, sarebbe dunque saggio: purtroppo l'umanità attuale, protagonista e vittima di un progresso tecnologico vertiginoso, dagli esiti imprevedibili, sembra abbia rescisso i ponti con le proprie origini, vive intensamente e unicamente il presente”. Ringraziamo i componenti della sua famiglia qui presenti: la moglie Prof.ssa Licia Ayala e la figlia Prof.ssa Elena Magnani con il marito e i nipoti Pietro e Ilaria. Siamo loro molto grati e riconoscenti per aver contribuito a sostenere l’associazione in questa ricorrenza e fatto dono della Biblioteca personale del Prof. Paolo Magnani, contenente circa centocinquanta volumi, preziose testimonianze documentali degli studi preistorici, oltre a diverse copie delle sue pubblicazioni.
Gabriella Gandolfi
Presidente
Gruppo Archeologico Albinetano “Paolo Magnani”
RICORDO DELL’AMICO PAOLO MAGNANI
Credo di essere stato io la persona con la quale il prof. Magnani, negli ultimi tempi, ha avuto più occasioni di scambiare informazioni e valutazioni sulle nuove idee in ambito paleo-antropologico e archeologico. E’ stato nell’estate 2012, per quanto ricordo, che ho cominciato a parlargli dell’affermarsi, tra gli studiosi anglofoni, del MODELLO A CESPUGLIO DELLA STORIA EVOLUTIVA DELLA UMANITA’. Magnani, apparso subito perplesso di fronte all’abbandono del MODELLO LINEARE – che contempla una seriazione rigorosa di specie definite e che, a suo tempo, aveva adottato nel suo testo sulla “Preistoria di Reggio nell’Emilia” – ha presto dimostrato di condividere il mio interesse per questo nuovo modello; ne parlava volentieri, ma continuava a vagliarlo criticamente, finché un giorno si è presentato a casa mia, dichiarando di avere compreso che il termine “specie” è usato dai paleo-antropologi moderni in una accezione approssimativa, lontana dalla rigorosa definizione biologica, così da ridursi a una pura e semplice etichetta, peraltro assai comoda per connotare una popolazione in corso di evoluzione; l’adozione del MODELLO A CESPUGLIO – che spesso prescinde da una precisazione dei tipi biologici di ogni gradino del processo di Ominazione – gli appariva quindi coerente con un approccio pragmatico all’argomento, quindi perfettamente accettabile per omettere attribuzioni di specie, che sono solitamente destinate a essere smentite da qualche successivo ritrovamento. Il Modello Lineare dell'evoluzione umana – ritenuto valido fino a poco tempo fa ed espresso da una successione di ominidi, solitamente da H. HABILIS a H. SAPIENS, rappresentati in una postura gradualmente sempre più eretta che raggiunge la perfetta verticalizzazione solo con l'ultima specie (noi) – appare oggi completamente superato. Questa concezione è stata ormai abbandonata, in quanto non tiene conto dei più recenti contributi genetici alla biologia dell’Ominazione, attualmente ritenuti di importanza cruciale. Oggi, sappiamo, infatti, che la nostra preistoria si è sviluppata attraverso il concorso di differenti apporti da parte di molte specie e/o varianti; già il primo ominide del Genere Homo – forse rappresentato da LD 350-1 – conviveva nell’Africa Orientale assieme ad altri congeneri con i quali verisimilmente sono avvenuti molteplici scambi di materiale genetico; poiché qualche sorta di commistione genetica si è ripetuta a tutti i livelli dell’evoluzione umana, si ritiene più esatto ed esplicativo sostituire al concetto di Linea Evolutiva quello di FAMIGLIA UMANA e rappresentare l’Ominazione come un processo lungo e complesso, simbolicamente paragonabile alla crescita di un albero frondoso. Ritengo, pertanto, che la SCHEDA DELL’ALBERO DELLA FAMIGLIA UMANA, distribuita a tutti i presenti stassera, abbozzata insieme a Magnani nei suoi tratti essenziali, quindi aggiornata fino a fargli assumere la forma attuale, rappresenti un omaggio significativo alle doti critico-razionali dell’amico scomparso.
Alberto Catalano
Socio del Gruppo Archeologico Albinetano “Paolo Magnani”
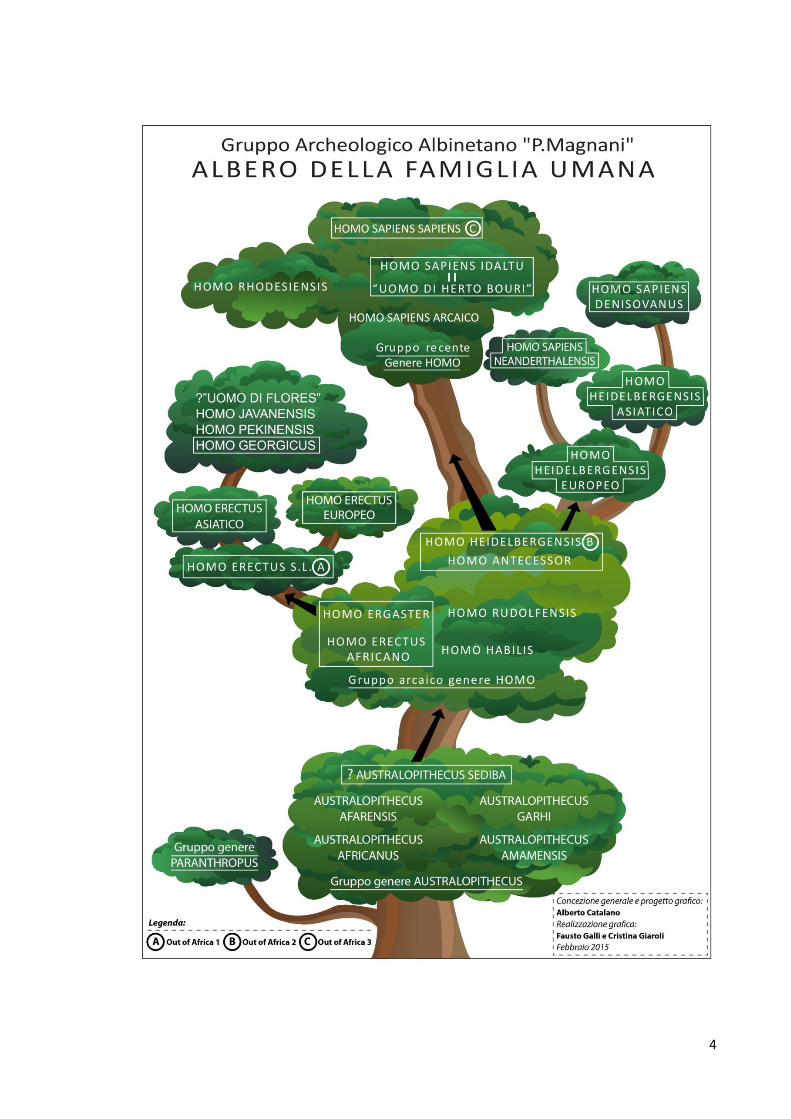
4

5
1^ Conversazione: 5 marzo 2015
Apparizione dei Primati. Ominidi Pre-Umani ancestrali
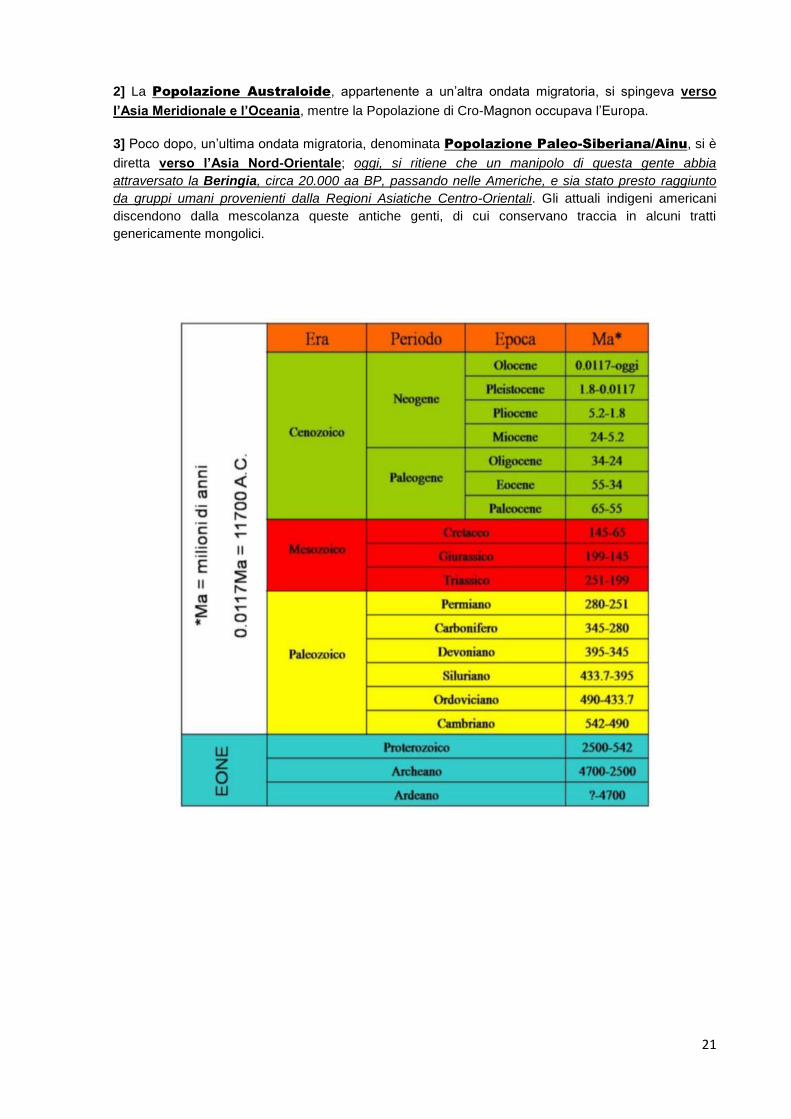
SCHEDA 01: TRANSIZIONE BIOTICA DI 65 MILIONI AA BP
La conversazione di stasera comincia in America.
Nel mare caraibico antistante quella che oggi è la parte estrema della Penisola Messicana dello
Yucatan, circa 65 milioni aa BP, è avvenuto uno dei più catastrofici impatti cosmici riguardanti il
nostro pianeta: un asteroide di circa 12 Km di diametro è entrato in collisione con la Terra a velocità
così elevata da produrre una cavitazione della crosta terrestre, denominata Cratere di Chicxulub,
profonda circa 20 Km, avente un diametro di oltre 300 Km, tuttora riconoscibile mediante prospezione
geologica.
1 2
1 – Localizzazione del Cratere di Chicxulub
2 – Mappa Gravimetrica della parte estrema dello Yucatan; la linea di costa corrisponde alla
variazione di colore, mentre le piccole chiazze bianche segnalano i cenotes (foibe, inghiottitoi
carsici) che contornano la parte centrale del cratere
L’impatto dell’asteroide è stato immediatamente seguito dalla formazione di una immensa nube di
polveri che ha avvolto l’intero pianeta per almeno una decina di anni e ha sconvolto l’ecosistema
terrestre in misura tale da determinare, da un lato, la scomparsa di un grande numero di specie viventi
e, dall’altro lato, la sopravvivenza di poche altre che si sono poi sviluppate a tal punto da diventare
dominanti.
Nella Storia Geologica della Terra (V. pag 21), il Passaggio MesozoicoCenozoico coincide con
questo evento cruciale, risalente appunto a 65 milioni aa BP.
Quella immane catastrofe è particolarmente significativa per il nostro argomento, in quanto ha segnato
la fine della dominazione dei Dinosauri e l’inizio della ascesa dei Mammiferi, Classe Zoologica alla
quale noi apparteniamo.
Tra gli animali scampati all’eccidio, perché hanno potuto proteggersi in tane sotterranee,alimentandosi
di insetti, larve, bulbi e radici, c’era una specie, denominata PURGATORIUS CERATOPS, dalla quale
discende l’Ordine dei Primati, che comprende l’Uomo e le categorie zoologiche affini.

6
Ricostruzione del probabile aspetto di Purgatorius Ceratops
Lentamente, Purgatorius è evoluto in forme appena più grandi, che hanno appreso ad arrampicarsi
sugli alberi per mangiarne i frutti e così hanno anche acquisito un indubbio vantaggio competitivo,
offerto dalla possibilità di fuga sui rami più alti per sottrarsi agli attacchi delle fiere terricole.
La seriazione successiva dei discendenti di Purgatorius comprende animali di taglia crescente, alcuni
anche privi di coda, che in questa sede non meritano menzione.
Mappa orientativa dei Primati Schema della Great Rift Valley
La rassegna dei nostri antenati inizia dai più antichi Primati Pre-umani, risalenti ad alcuni milioni di
anni fa, precisamente dagli Ominoidei; la differenziazione degli Ominidi nell’ambito degli Ominoidei è
avvenuta in Africa, circa 6 milioni aa BP, dopo un altro importante fenomeno geologico: l’apertura
della Great Rift Valley.
In seguito a tale evento tettonico, si è verificato un rilevante cambiamento climatico per cui la foresta
pluviale, che fino ad allora aveva coperto tutta la fascia equatoriale, ha lasciato scoperta la parte
orientale, divenuta ben presto una prateria semiarida, con radi alberi e/o arbusti, nota come savana.
Gli Ominoidei ben adattati alla vita arboricola si sono spostati, allora, a ovest, per rimanere nell’habitat
forestale, e si sono evoluti successivamente nelle Scimmie Antropomorfe Attuali (Gibboni, Gorilla,
Orango e Scimpanzè). I pochi Ominoidei che si sono adattati alle nuove condizioni ambientali della
savana sono, invece, rimasti nell’Africa Orientale, dove hanno cominciato a sviluppare i connotati
morfo-funzionali e comportamentali che contrassegnano la linea evolutiva dell’Uomo, così da ricevere
appunto la denominazione di OMINIDI.

7
.
L’Evoluzione Umana/OMINAZIONE, è avvenuta
all’interno della famiglia degli Ominidi, attraverso
un complesso processo di modificazioni
successive, durato circa 6 milioni di anni.
Oggi sappiamo che questo processo si è svolto
secondo uno schema discontinuo di
cambiamenti improvvisi e rapidi alternati a
pause più o meno prolungate, che traduce la
forma aggiornata della Teoria Darwiniana, detta
Modello degli Equilibri Punteggiati di Eldredge
e Gould.
SCHEDA 02: CRITERI RIVELATORI DI UMANITA’
L’evoluzione umana è avvenuta gradualmente, per successive acquisizioni.
1) Il primo tratto evolutivo umano, denominato Biped(al)ismo – consistente nella capacità di
assumere la stazione eretta e di camminare sui soli arti inferiori – è comparso in Africa Orientale
circa 4 milioni aa BP, ancora associato alla locomozione arboricola; il Bipedismo esclusivo si è
affermato solo 2 milioni di anni dopo, con Homo Erectus, e ha implicato l’importante
vantaggio di liberare gli arti superiori per utilissime attività, prima negate (afferrare,
trasportare, fabbricare, cacciare, pescare, etc.).
L’adozione del bipedismo è stato attribuito al passaggio dalla vita sugli alberi alla vita in
savana, che richiedeva la minore esposizione possibile al sole equatoriale (radiazione
pressoché verticale) e la necessità di proteggersi dalle fiere avvistandole in distanza per fuggire
velocemente sugli alberi. La maggiore mobilità offerta dal bipedismo ha implicato anche un
miglioramento dell’alimentazione, divenuta più ricca e variata, in funzione della quale si è
anche verificata una modificazione della dentatura, che ha consentito una più completa
assimilazione dei diversi nutrienti, che, a sua volta, ha incentivato lo sviluppo cerebrale.
2) Il passo successivo è rappresentato dall’assunzione della capacità di fabbricare e usare
strumenti, risalente a circa 2,5 milioni aa BP, presumibilmente dovuta a un primo ampliamento
dell’encefalo.
Si ritiene che un successivo, ulteriore ingrandimento cerebrale abbia dato inizio allo sviluppo
della comunicazione orale. Il linguaggio dei più antichi ominidi era certamente rappresentato da
suoni simili ai vocalizzi delle scimmie attuali, aventi funzione di identificazione all’interno del
gruppo e di avvertimento di specifici pericoli. Il vero e proprio linguaggio verbale si è sviluppato
tardivamente, verisimilmente solo nell’uomo anatomicamente moderno, in seguito a processi
biologici ancora incompletamente noti, circa 100.000 aa BP, presumibilmente in Sudafrica.
3) Un altro importante passo avanti verso una condotta pienamente umana è rappresentato dal
controllo del fuoco; presumibilmente, i più antichi Ominidi, per scaldarsi e forse cuocere i cibi,
raccoglievano materiale ardente da fonti naturali (alberi incendiati da fulmini e/o lava vulcanica
incandescente), e solo successivamente hanno appreso ad accendere il fuoco, con tecnica a
percussione litica prima, quindi col procedimento della frizione lignea. La data del primo utilizzo
del fuoco da parte dei nostri antenati continua a spostarsi indietro nel tempo, via via che si
allargano le nostre conoscenze archeologiche.

8
Nella Grotta di Wonderwerk (Sudafrica) sono
state scoperte ceneri frammiste a resti
carbonizzati di ossame animale e di erbe,
evidenti indizi di un falò non accidentale, risalente
a circa un milione aa BP. Una sicura traccia di
uso intenzionale del fuoco tra 400.000 e
300.000 aa BP è stata trovata in Israele
(Focolare di Qesem Cave); si tratta di una sorta
di braciere di circa 2 metri di diametro, pieno di
ceneri e ossi combusti, usato ripetutamente. Veri
e propri focolari preistorici si repertano a far
tempo da circa 300.000 aa BP; uno dei più
antichi si trova in Europa, precisamente in
Francia (Capanna di Terra Amata)
4) L’ultimo, più significativo, tratto squisitamente umano è stato lo sviluppo della cognitività,
rivelata dalla comparsa di sempre più complesse capacità di progettazione, immaginazione e
pensiero simbolico, le cui espressioni più rilevanti sono le arti e le tcnologie; questo cruciale
passo evolutivo, compiuto solo dai membri della specie Homo Sapiens, sembrerebbe correlato
allo sviluppo del linguaggio, quindi risalirebbe a circa 100.000 aa BP, verisimilmente in
Sudafrica.
In questo schema, sono sintetizzate le principali funzioni cognitive, delle quali è anche
indicata la presunta distribuzione emisferica
SCHEDA 03: INIZIO DELLA STORIA DELL’UMANITA’: SCIMMIE AUSTRALOPITECINE
Per convenzione, la “Storia dell’Umanità” inizia ufficialmente dal momento in cui i nostri più
lontani antenati hanno costruito i primi manufatti.
Questi risalgono a circa 2,5 milioni aa BP e sarebbero stati prodotti da Ominidi Pre-umani
appartenenti al Genere Australopithecus, che oggi conosciamo abbastanza bene attraverso lo
studio dei numerosi fossili repertati, tutti provenienti dall’Africa Sud-Orientale.
Le Scimmie Australopitecine avevano statura poco superiore al metro e aspetto generale ancora
animalesco, ma – rispetto agli alti Primati contemporanei – la loro dentatura era già modificata in

9
senso umano (netta riduzione di incisivi e canini, associata ad aumento di premolari e molari) e il loro
encefalo, quantunque ancora piccolo (400-500 cc), presentava una certa complessità interna. Erano
capaci di locomozione bipede (sebbene fossero ancora in grado di vivere sugli alberi) come è ben
documentato dalla serie di orme rinvenute in Tanzania, a sud della Gola di Olduvai, risalenti a circa
3,6 milioni aa BP e note come Laetoli footprint trails.
Lucy Laetoli footprint trails
Le orme sono state impresse nelle ceneri tiepide del Vulcano Sadiman, poi cementate dalla pioggia,
da tre individui, due affiancati e uno che li seguiva mettendo i piedi nelle impronte del capofila di
destra; a fianco dei footprint trails, sulla destra, sono visibili anche orme di animali.
Sono le Scimmie Australopitecine di taglia più minuta che rivestono la massima importanza paleo-
antropologica, in quanto sono riuscite a sopravvivere in un ambiente piuttosto povero di risorse
alimentari, provvedendo a integrare la scarsa raccolta di nutrienti vegetali con brani di carne,
strappati dalle carogne abbandonate dalle fiere, mediante i primi rudimentali strumenti di
pietra. Si attribuisce a questa dieta parzialmente carnea lo sviluppo di più efficaci modalità di
funzionamento cerebrale e soprattutto di più strette modalità relazionali e collaborative tra i membri del
gruppo. Sono, appunto, questi importanti progressi somatici e sociali che hanno segnato l’inizio
della differenziazione del nuovo Genere Homo. Il reperimento recentissimo (2015) di una
mandibola frammentaria in Etiopia, contrassegnata con la sigla LD 350-1, rivoluziona le attuali
convinzioni sui primi membri del Genere Homo; questo fossile risale a più di 2,5 milioni aa BP e
presenta evidenti segni di transizione Genere Australopithecus Genere Homo.
Noti al grande pubblico sono:
Taung Child (Sudafrica), primo fossile australopitecino repertato, risalente a 2,5 milioni aa BP,
appartenente alla specie Australopithecus Africanus, presumibilmente ucciso da un rapace;
infatti, nelle orbite sono ben visibili lesioni ossee che i paleontologi ritengono di attribuire agli artigli di
un’aquila.
Taung Child Endocranio
Lucy (Etiopia), una giovane femmina, risalente a 3,2 milioni aa BP, appartenente alla specie
Australopithecus Afarensis.

10
Per le finalità della presente conversazione, devo però focalizzare la vostra attenzione su altre due
Australopitecine di peculiare interesse:
-Australopithecus Garhi, vissuto in Africa Orientale (Etiopia) circa 2,5 milioni aa BP, sebbene
maldocumentato dai reperti finora raccolti, rappresenta un membro cruciale nella storia evolutiva
dell’Umanità, perché rappresenta verisimilmente la prima specie a costruire utensili e a mangiare
carne; i suoi scarsi resti fossili, infatti, erano associati ad alcuni strumenti litici arcaici e a ossami di
animali che portano tracce di scarnificazione strumentale o frattura intenzionale per estrarne il midollo.
-Australopithecus Sediba si ritiene oggi il nostro più probabile antenato tra tutti i congeneri; ben
documentato da reperti fossili in ottime condizioni di conservazione, è vissuto in Sudafrica (Malapa
Valley) tra 1,97 e 1,98 milioni aa BP; secondo alcuni AA, nonostante il ridotto volume cranico (circa
450 cc.), sarebbe già un arcaico esponente del Genere Homo.
Olotipo di Australopithecus Sediba, conservato al CRADLE OF HUMANKIND (UNESCO,
1999), Maropeng, Sudafrica
SCHEDA 04: PRIMI ESPONENTI DEL GENERE HOMO
Homo Rudolfensis e Homo Habilis sono ritenuti finora i più antichi esponenti del Genere
Homo, al quale anche noi apparteniamo.
-Homo Rudolfensis, risalente a 1,9 milioni aa BP, viveva presso il Lago Turkana, già Lago
Rodolfo (Kenya), aveva volume cerebrale di circa 700 cc., ma aspetto ancora scimmiesco, così da
essere considerato da alcuni studiosi ancora una Australopitecina. Costruiva utensili litici tanto
rudimentali da non essere riuscito a sopravvivere a lungo e da essere soppiantato ben presto dal suo
rivale evolutivo H. Habilis, manualmente più efficiente.
-Homo Habilis – vissuto da circa 2,4 a 1,4 milioni aa BP, in Tanzania (Olduvai Gorge) – è stato il
primo sicuro produttore di utensili, forse preceduto da A.Garhi, più di 100.000 anni prima; i suoi
manufatti litici - choppers e chopping tools – devono essersi rivelati, sebbene ancora rudimentali, armi
sufficientemente efficaci per la difesa dalle fiere (non per la caccia vera e propria) e per sottrarre carne
a jene e sciacalli. Aveva capacità cranica di 700-750 cc, cosicché il suo encefalo è riuscito a
esprimere la abilità cognitiva e motoria necessaria a produrre una Industria litica definita
Olduvaiana/di Modo 1.

11
Chopping Tool Homo Habilis LD 350-1
Oggi, viene messa in dubbio, per ragioni cronologiche, la radicata tesi che Homo Habilis si collochi
sulla linea evolutiva umana, cosicché alcuni studiosi pensano addirittura che si sia estinto senza
lasciare discendenza.
E’ sicuramente un nostro progenitore Homo Ergaster, che discenderebbe – direttamente o
tramite una specie intermedia – da Australopithecus Sediba. Negli anni passati, si è lungamente
discusso se Homo Ergaster, a sua volta, sia il progenitore di Homo Erectus, disseppellito
inizialmente in Estremo Oriente (sebbene oriundo africano), oppure ne sia l’equivalente africano; oggi,
la questione è stata superata: Homo Ergaster e Homo Erectus sono ritenute due varianti,
rispettivamente africana ed euro-asiatica, della medesima specie, denominata Homo Erectus
sensu lato.
Homo Ergaster/Erectus era caratterizzato da una elevata variabilità regionale, oltre che
temporale, così da avere mostrato connotati tipo Ergaster fintanto che è vissuto in Africa e
connotati tipo Erectus dopo essere migrato dall’Africa, circa 1,8 milioni aa BP.
I suoi fossili africani sono un po’ più grandi (statura fino a 185 cm.) di quelli euro-asiatici
(statura media o appena inferiore); aveva corporatura e proporzioni pressoché simili alle nostre,
teca cranica di circa 900 cc di capacità; era dotato di facoltà cognitive nettamente superiori a Homo
Habilis, espresse da una comunicazione orale più efficace, da una maggiore disponibilità sociale, da
un ottimo senso di orientamento e soprattutto da una abilità manuale migliore, documentata da una
produzione litica innovativa e diversificata, denominata Industria Acheuleana/di Modo 2.
Homo Erectus S.L. (ricostruzione) Bifacciale Acheuleano/Amigdala
A Homo Ergaster/Erectus si attribuisce la scoperta dell’uso del fuoco, che forse sapeva
all’occorrenza anche accendere intenzionalmente, ma che più probabilmente preferiva mantenere
acceso in luoghi appartati, presumibilmente a cura delle femmine; Homo Ergaster/Erectus è stato
anche l’inventore della caccia, che praticava perlopiù in gruppo.
Oggi, i paleo-antropologi del Mondo Occidentale pensano che questo nostro antenato sia stato
artefice della prima, massiccia migrazione di Ominidi dall’Africa, avvenuta tra 1,8 e 1,3 milioni aa
BP.

12
Questo esodo (Out-of-Africa 1) ha avuto successo in virtù della elevata capacità di adattamento di
H.E/E. ai nuovi ambienti, climaticamente e geologicamente ben diversi da quelli africani; uscito dal
Continente Africano, infatti, si è diffuso in Asia Occidentale e Orientale e – seppure tardivamente –
anche nelle regioni meridionali dell’Europa.
Molti studiosi attribuiscono la sua uscita dall’Africa al carnivorismo, che impone di inseguire le
mandrie nei loro spostamenti nel territorio; l’alimentazione carnea ha conferito grossi vantaggi a
quei lontani progenitori, perché la carne rappresenta una fonte concentrata di calorie e di nutrienti, si
digerisce più rapidamente, rispetto ai vegetali, soprattutto dopo cottura, e consente così un risparmio
energetico per le attività cerebrali e muscolari.
Homo Ergaster/Erectus è sopravvissuto per oltre 1 milione 700.000 anni, da circa 1,89 milioni a
circa 140.000 aa BP, convivendo per alcune decine di migliaia di anni con i nostri antenati specifici
(Homo Sapiens Sapiens).
Voglio concludere questa conversazione segnalandovi una recente scoperta molto interessante
(2014): riesaminando il materiale archeologico raccolto, nel 1891, da M-E.Dubois, paleontologo
olandese, nel sito di ritrovamento dei primi fossili di Homo Erectus, a Java (Trinil), alcuni studiosi
hanno osservato che due conchiglie d’acqua dolce presentano tracce di una manipolazione
praticata circa 430.000 aa BP: una ha il margine affilato, per trasformarla in un raschiatoio o un
coltello; l’altra porta un disegno a zig-zag (forse inciso con un dente di squalo fossile), che sembra
rappresentare la più antica espressione d’arte conosciuta.
Territori colonizzati da Homo
Erectus

13
2^ Conversazione: 12 marzo 2015
Ominidi Pre-Umani recenti
SCHEDA 05: NUOVA TEORIA EVOLUTIVA DOPO IL RITROVAMENTO DI HOMO ERECTUS
IN SITI EXTRA-AFRICANI
In questi ultimi anni, si è fatto molto acceso il dibattito paleo-antropologico non tanto sui primi Homo
quanto sulla loro evoluzione.
Mentre il Modello Out-of-Africa – che sostiene l’origine africana dell’Umanità ed è confortato dai
risultati delle più moderne ricerche genetiche – è oggi accettato pressoché universalmente, una
nuova tesi del 2013, denominata Teoria dell’Origine Unica dei membri del Genere Homo, propone
che i congeneri Homo Rudolfensis, Homo Habilis, Homo Ergaster, Homo Erectus e Homo
Antecessor siano tutti altrettante varianti dell’unica specie Homo Erectus S. L., dal quale
deriverebbe, per successive approssimazioni, l’Uomo Moderno. Finora pochi archeologi hanno
accettato questa teoria.
Modello Out-of-Africa
SCHEDA 06: ESPONENTI PIU’ RECENTI DEL GENERE HOMO: (1) HOMO ANTECESSOR E
HOMO HEIDELBERGENSIS
Tra i congeneri convissuti con Homo Ergaster/Erectus nel continente africano, riveste particolare
interesse un ominide, denominato Homo Heidelbergensis, detto anche Homo Sapiens
Arcaico per alcuni suoi tratti anatomici già moderni. Il suo più antico fossile, la Mandibola di Mauer
(Heidelberg/Germania), è stato magistralmente studiato, nel 1882, da O. Schoetensack, che sulla
sola base di quell’unico resto fossile ha ipotizzato la sua appartenenza a questa nuova specie, che poi
è stata confermata dal ritrovamento di molteplici fossili.
Mandibola di Mauer

14
H. Heidelbergensis è vissuto in Africa, Europa e Asia Occidentale, da 800.000 a 100.000 aa BP,
adattandosi a sopravvivere anche in ambienti freddi, dove gli è stato certamente di grande utilità
sapere accendere e controllare il fuoco. Aveva capacità cranica di 1100-1250 cc; è stato un abile
cacciatore, capace di abbattere grandi erbivori (cervidi, equidi, pachidermi, etc.), servendosi – già
600.000 aa BP – di lance di legno, che da 300.000 aa BP ha nettamente migliorato armandone le
estremità utili con punte litiche.
Homo Heidelbergensis (ricostr.) “Excalibur” Lance di Schoeningen
L’industria litica di Homo Heidelbergensis era, tuttavia, ancora alquanto primitiva, sebbene più
avanzata rispetto a quella dei predecessori, così da essere denominata Acheuleano Evoluto; un
esempio della sua abilità litotecnica è offerto dallo splendido bifacciale noto come “Excalibur”.
Questa popolazione abitava non solo in ripari naturali, ma anche in ricoveri appositamente
costruiti, di cui l’esempio più dimostrativo è la Capanna di Terra Amata (Nizza, Francia), dotata
anche di un focolare.
Scapola equina perforata di Eartham Pit Ricostruzione della Capanna di Terra Amata
Poiché sono assai poco noti i fossili di Homo Heidelbergensis, in questa sede ritengo opportuno
presentarvi almeno i più significativi:
Lance di Schoeningen (Germania), zagaglie da caccia in legno, perfettamente conservate,
risalenti a circa 400.000 aa BP, consistenti in fusti sottili e diritti di abete rosso e di pino, lunghi 1,8-2,5
metri, adatti a essere scagliati.
Scapola equina perforata di Eartham Pit/Boxgrove (Gran Bretagna), consistente nella placca
di una scapola di cavallo, risalente a circa 500.000 aa BP, nella quale è visibile un foro pressoché
circolare prodotto da una arma con le caratteristiche delle lance di cui sopra.
15
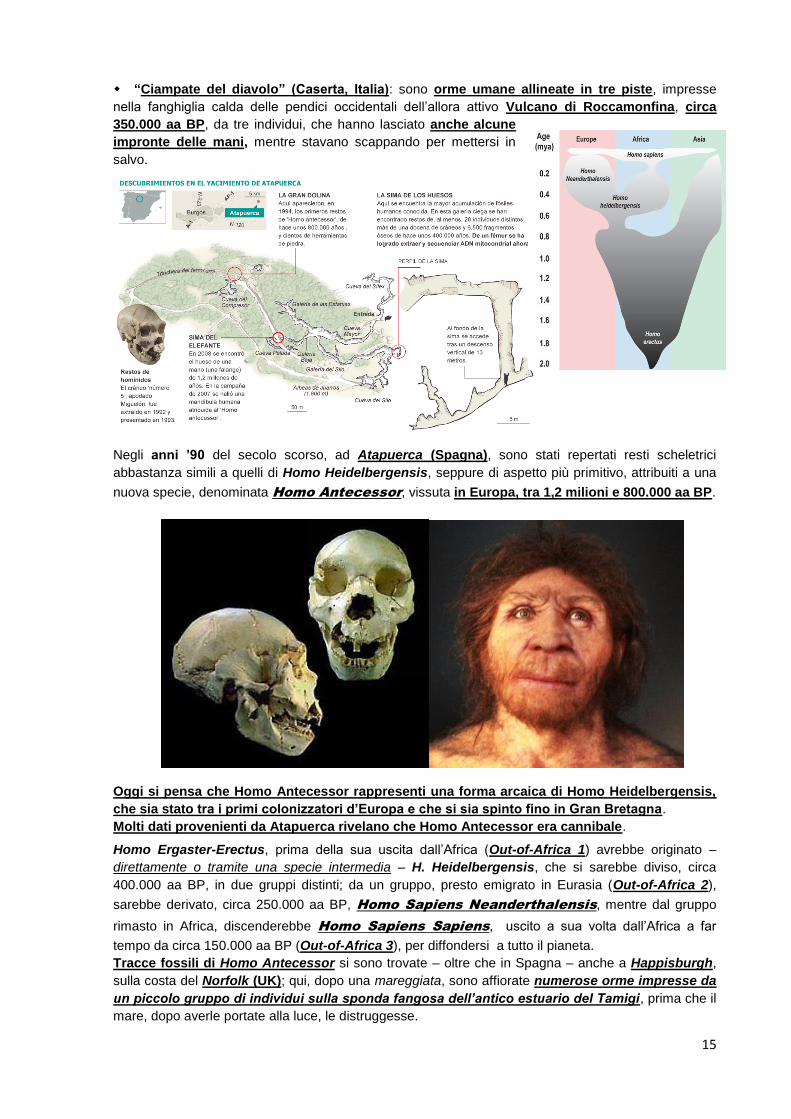
“Ciampate del diavolo” (Caserta, Italia): sono orme umane allineate in tre piste, impresse
nella fanghiglia calda delle pendici occidentali dell’allora attivo Vulcano di Roccamonfina, circa
350.000 aa BP, da tre individui, che hanno lasciato anche alcune
impronte delle mani, mentre stavano scappando per mettersi in
salvo.
Negli anni ’90 del secolo scorso, ad Atapuerca (Spagna), sono stati repertati resti scheletrici
abbastanza simili a quelli di Homo Heidelbergensis, seppure di aspetto più primitivo, attribuiti a una
nuova specie, denominata Homo Antecessor, vissuta in Europa, tra 1,2 milioni e 800.000 aa BP.
Oggi si pensa che Homo Antecessor rappresenti una forma arcaica di Homo Heidelbergensis,
che sia stato tra i primi colonizzatori d’Europa e che si sia spinto fino in Gran Bretagna.
Molti dati provenienti da Atapuerca rivelano che Homo Antecessor era cannibale.
Homo Ergaster-Erectus, prima della sua uscita dall’Africa (Out-of-Africa 1) avrebbe originato –
direttamente o tramite una specie intermedia – H. Heidelbergensis, che si sarebbe diviso, circa
400.000 aa BP, in due gruppi distinti; da un gruppo, presto emigrato in Eurasia (Out-of-Africa 2),
sarebbe derivato, circa 250.000 aa BP, Homo Sapiens Neanderthalensis, mentre dal gruppo
rimasto in Africa, discenderebbe Homo Sapiens Sapiens, uscito a sua volta dall’Africa a far
tempo da circa 150.000 aa BP (Out-of-Africa 3), per diffondersi a tutto il pianeta.
Tracce fossili di Homo Antecessor si sono trovate – oltre che in Spagna – anche a Happisburgh,
sulla costa del Norfolk (UK); qui, dopo una mareggiata, sono affiorate numerose orme impresse da
un piccolo gruppo di individui sulla sponda fangosa dell’antico estuario del Tamigi, prima che il
mare, dopo averle portate alla luce, le distruggesse.

16
Happisburgh footprints Bifacciale di Happisburgh
SCHEDA 07: ESPONENTI PIU’ RECENTI DEL GENERE HOMO: (2) HOMO SAPIENS NEAN-
DERTHALENSIS
Con Homo Heidelbergensis si conclude il periodo definito Paleolitico Inferiore, al quale
succede il periodo Paleolitico Medio, dominato da un importante membro della specie Homo
Sapiens: “Uomo di Neanderthal”, ovvero Homo Sapiens Neanderthalensis.
Questo è vissuto in Europa (Centro-Meridionale) e in Asia (Sud-Occidentale e Centrale) da
circa 250.000 a 39.000 aa BP; quest’ultima data, precisata da studi recentissimi, rivela che è
convissuto con Homo Sapiens Sapiens – almeno nel corridoio del Levante e nell’Europa
Mediterranea – per circa 5.000 anni.
Homo Sapiens Neanderthalensis è una variante di Homo Sapiens Sapiens; aveva statura media,
corporatura tozza, teschio allungato con capacità cranica di circa 1500 cc. (quindi superiore alla
nostra attuale) e arcate sopracciliari marcate; probabilmente destrimane e di carnagione chiara, con
capelli rossicci e occhi verde-azzurri, era dotato di un repertorio comportamentale piuttosto ricco:
costruzione di ripari estivi all’aperto; inumazione delle salme; allestimento di grandi focolari;
buona competenza litotecnica, rivelata da un discreto repertorio di strumenti, perlopiù prodotti
mediante la scheggiatura “Levallois”, tipica dell’industria detta Mousteriana/di Modo 3.

17
Manufatti Mousteriani
I Neanderthaliani vivevano in piccoli gruppi famigliari isolati (clan), erano essenzialmente carnivori
e si alimentavano prevalentemente di carne di ungulati. Esistono sporadiche prove archeologiche di
cannibalismo; il sito più dimostrativo di tale comportamento è la Cueva de El Sidron (Spagna): la
carneficina ha riguardato un intero clan – forse una sorta di vendetta dopo uno sgarro – ma non si può
escludere la antropofagia per fame, in quanto tutti gli scheletri portano segni evidenti di
malnutrizione.
I siti neanderthaliani scavati – numerosi in Europa – sono relativamente scarsi in Italia; segnalo,
tuttavia, il Campo-base di Ghiardo (Reggio Emilia). Recentemente è stata recuperata una calotta
cranica neanderthaliana dalle sabbie del Po (San Daniele Po, Cremona).
SCHEDA 08: ESTINZIONE DI HOMO SAPIENS NEANDERTHALENSIS
L’estinzione dell’Uomo di Neanderthal rimane a tutt’oggi per molti aspetti enigmatica; dopo
l’arrivo in Eurasia di Homo Sapiens Sapiens, i Neanderthaliani sono stati progressivamente
emarginati dai migliori territori di caccia e indotti a ritirarsi alla periferia delle regioni allora abitabili,
spesso in zone inospitali.
Molti dati archeologici, paleontologici e geologici oggi disponibili suggeriscono che i
Neanderthaliani in Europa – già prima dell’arrivo dei Sapiens Sapiens – fossero stati decimati
da eventi catastrofici, principalmente rappresentati da:
1] Eventi di Heinrich – Sono picchi di temperature bassissime, che si sono susseguiti da 55.000
aa BP al termine dell’ultima Glaciazione; i Neanderthaliani sarebbero stati, se non incapaci,
perlomeno lenti ad adattare le proprie tattiche di caccia ai cambiamenti ecologici indotti da tali bruschi
sbalzi climatici; inoltre, poiché il loro metabolismo richiedeva un apporto energetico di circa 350
calorie/giorno in più rispetto agli uomini moderni, l’apporto alimentare corrispondente non poteva
essere sempre assicurato in tempi di carestia; infine, i Neanderthaliani erano forti e robusti, ma
mancavano di agilità, così da risultare penalizzati quando la grossa selvaggina si era molto ridotta,
sostituita da piccoli e veloci roditori come i Leporidi.
2] Violenta eruzione del Supervulcano dei Campi Flegrei – Questa imponente eruzione, risalente a
39.000 aa BP, ha innescato un lungo e freddissimo inverno vulcanico; le ceneri eruttate hanno
oscurato il sole in misura così intensa da determinare, per qualche anno, un gravissimo
sconvolgimento dell’intero eco-sistema delle zone abitate da Homo Sapiens Neanderthalensis.

18
Crateri vulcanici flegrei Area di deposizione delle ceneri vulcaniche flegree
La prova più dimostrativa della devastazione prodotta dall’eruzione flegrea si trova nella Grotta
Mezmaiskaya (Russia) nella quale gli archeologi russi hanno trovato una evidente coltre di ceneri
flegree: mentre le tracce neanderthaliane sono numerose sotto di essa (prima dell’eruzione), invece
nei livelli sovrastanti (dopo l’eruzione) sono completamente assenti.
Durante lo svolgimento della catastrofe, i primi gruppi di uomini moderni in uscita dall’Africa
stavano lentamente percorrendo il Corridoio del Levante, così da non risentirne le
conseguenze e semmai da ricavarne il vantaggio di trovare le regioni euro-asiatiche spopolate.
I fattori implicati nell’estinzione dell’Uomo di Neanderthal considerati finora rappresentano
certamente concause importanti, ma quella che le ultime ricerche indicano come causa più
probabile e determinante della sua scomparsa è stato l’Interbreeding con Homo Sapiens
Sapiens; è ormai provato, infatti, che – durante il periodo di convivenza con quest’ultimo – i
Neanderthaliani, ormai nettamente ridotti di numero, siano andati incontro a una progressiva
ibridazione con i nuovi arrivati, fino a essere completamente riassorbiti dalla specie moderna;
questo spiega la presenza, nel nostro genoma, di una quota del 2-4% di materiale genetico
neanderthaliano. Studi condotti sul DNA mitocondriale di Homo Sapiens Sapiens – che si
trasmette solo per via materna – suggeriscono che gli incontri intimi più frequenti avvenivano tra
maschi moderni e femmine neanderthaliane.
Una nuova ipotesi (2015) – peraltro compatibile con la precedente – sostiene che il fattore
determinante dell’estinzione dei Neanderthaliani sia stato la incapacità di competere per la
sussistenza con gli ultimi arrivati uomini moderni; questi, infatti, godevano di una netta
superiorità venatoria, che li ha enormemente avvantaggiati: mentre quelli continuavano a usare armi
e tecniche di caccia tradizionali, questi non solo disponevano di più efficaci e maneggevoli armi da
lancio, ma soprattutto erano accompagnati da mute di canidi, addestrati a inseguire le prede e a
immobilizzarle così da facilitarne l’abbattimento; questo nuova modalità di caccia ha consentito ai
Sapiens Sapiens – durante le fasi più fredde dell’ultima Glaciazione – di nutrirsi della carne degli
enormi e pericolosi mammut, completamente preclusa ai Neanderthaliani, che hanno dovuto
accontentarsi di sporadiche catture di renne fino alla morte per inedia.
19
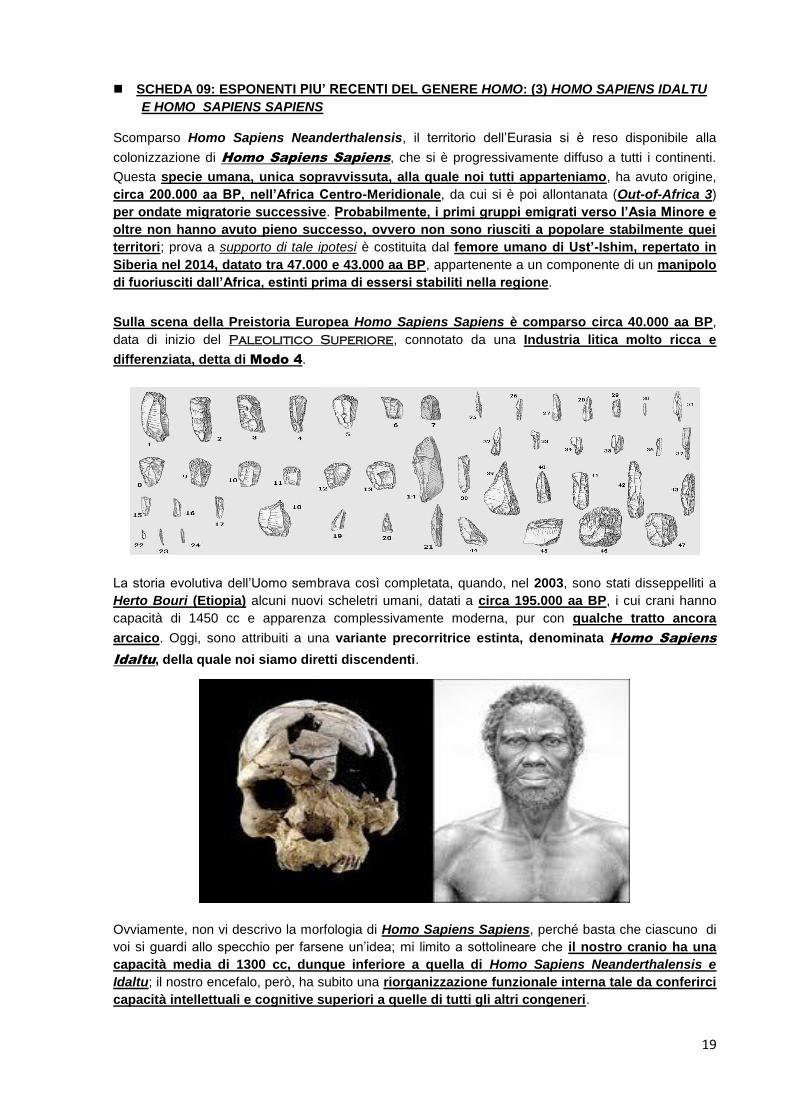
SCHEDA 09: ESPONENTI PIU’ RECENTI DEL GENERE HOMO: (3) HOMO SAPIENS IDALTU
E HOMO SAPIENS SAPIENS
Scomparso Homo Sapiens Neanderthalensis, il territorio dell’Eurasia si è reso disponibile alla
colonizzazione di Homo Sapiens Sapiens, che si è progressivamente diffuso a tutti i continenti.
Questa specie umana, unica sopravvissuta, alla quale noi tutti apparteniamo, ha avuto origine,
circa 200.000 aa BP, nell’Africa Centro-Meridionale, da cui si è poi allontanata (Out-of-Africa 3)
per ondate migratorie successive. Probabilmente, i primi gruppi emigrati verso l’Asia Minore e
oltre non hanno avuto pieno successo, ovvero non sono riusciti a popolare stabilmente quei
territori; prova a supporto di tale ipotesi è costituita dal femore umano di Ust’-Ishim, repertato in
Siberia nel 2014, datato tra 47.000 e 43.000 aa BP, appartenente a un componente di un manipolo
di fuoriusciti dall’Africa, estinti prima di essersi stabiliti nella regione.
Sulla scena della Preistoria Europea Homo Sapiens Sapiens è comparso circa 40.000 aa BP,
data di inizio del Paleolitico Superiore, connotato da una Industria litica molto ricca e
differenziata, detta di Modo 4.
La storia evolutiva dell’Uomo sembrava così completata, quando, nel 2003, sono stati disseppelliti a
Herto Bouri (Etiopia) alcuni nuovi scheletri umani, datati a circa 195.000 aa BP, i cui crani hanno
capacità di 1450 cc e apparenza complessivamente moderna, pur con qualche tratto ancora
arcaico. Oggi, sono attribuiti a una variante precorritrice estinta, denominata Homo Sapiens
Idaltu, della quale noi siamo diretti discendenti.
Ovviamente, non vi descrivo la morfologia di Homo Sapiens Sapiens, perché basta che ciascuno di
voi si guardi allo specchio per farsene un’idea; mi limito a sottolineare che il nostro cranio ha una
capacità media di 1300 cc, dunque inferiore a quella di Homo Sapiens Neanderthalensis e
Idaltu; il nostro encefalo, però, ha subito una riorganizzazione funzionale interna tale da conferirci
capacità intellettuali e cognitive superiori a quelle di tutti gli altri congeneri.
20
A riprova della superiorità intellettiva della nostra specie, accenno solo a due conquiste culturali
molto significative nella prospettiva archeologica:
Domesticazione di animali e vegetali: questa complessa operazione comprende una serie di
interventi intenzionali successivi, focalizzati a un determinato risultato; così, il cane è stato
domesticato per la caccia, la guardia e il controllo delle greggi forse già 45.000 aa BP, mentre gli
ovini sono stati domesticati circa 10.000 aa BP per la carne, la lana, il latte; i cereali, domesticati
mediante selezione in epoche più recenti, hanno richiesto un impegno intellettuale superiore e una
prolungata sperimentazione.
Arte preistorica parietale e mobiliare: veri capolavori artistici, eseguiti dagli antenati del
Paleolitico Superiore, sono stati incisi/dipinti sulle pareti di alcune caverne, mentre in diversi siti sono
state recuperati piccoli, ma preziosi manufatti, in materiali diversi, quali ornamenti corporei, statuette
di veneri e animali, etc..
SCHEDA 10: PIU’ ANTICA POPOLAZIONE MODERNA EUROPEA
I nostri primi antenati, comparsi nell’Africa Meridionale, erano individui complessivamente
piuttosto gracili, glabri, dotati di un linguaggio evoluto.
Questa Primigenia Gente Moderna, autrice dell’Out-of-Africa 3, comprendeva almeno tre diverse
popolazioni:
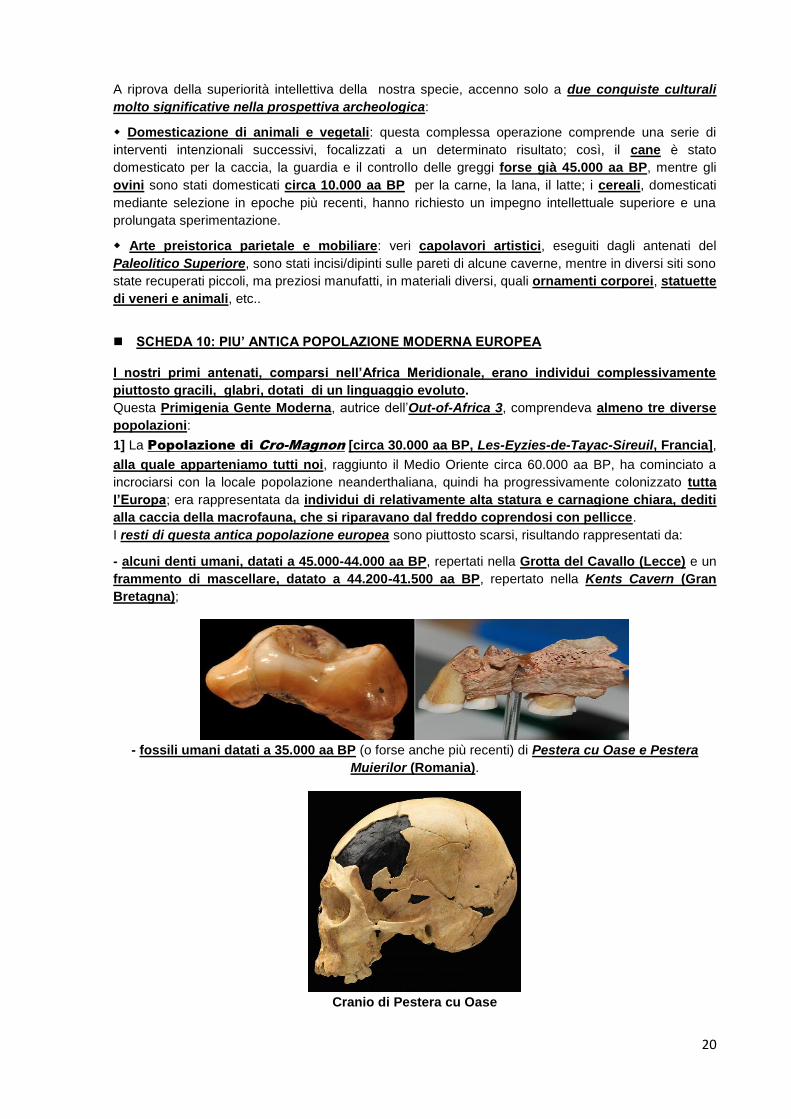
1] La Popolazione di Cro-Magnon [circa 30.000 aa BP, Les-Eyzies-de-Tayac-Sireuil, Francia],
alla quale apparteniamo tutti noi, raggiunto il Medio Oriente circa 60.000 aa BP, ha cominciato a
incrociarsi con la locale popolazione neanderthaliana, quindi ha progressivamente colonizzato tutta
l’Europa; era rappresentata da individui di relativamente alta statura e carnagione chiara, dediti
alla caccia della macrofauna, che si riparavano dal freddo coprendosi con pellicce.
I resti di questa antica popolazione europea sono piuttosto scarsi, risultando rappresentati da:
- alcuni denti umani, datati a 45.000-44.000 aa BP, repertati nella Grotta del Cavallo (Lecce) e un
frammento di mascellare, datato a 44.200-41.500 aa BP, repertato nella Kents Cavern (Gran
Bretagna);
- fossili umani datati a 35.000 aa BP (o forse anche più recenti) di Pestera cu Oase e Pestera
Muierilor (Romania).
Cranio di Pestera cu Oase
21
2] La Popolazione Australoide, appartenente a un’altra ondata migratoria, si spingeva verso
l’Asia Meridionale e l’Oceania, mentre la Popolazione di Cro-Magnon occupava l’Europa.
3] Poco dopo, un’ultima ondata migratoria, denominata Popolazione Paleo-Siberiana/Ainu, si è
diretta verso l’Asia Nord-Orientale; oggi, si ritiene che un manipolo di questa gente abbia
attraversato la Beringia, circa 20.000 aa BP, passando nelle Americhe, e sia stato presto raggiunto
da gruppi umani provenienti dalla Regioni Asiatiche Centro-Orientali. Gli attuali indigeni americani
discendono dalla mescolanza queste antiche genti, di cui conservano traccia in alcuni tratti
genericamente mongolici.

22
3^ Conversazione: 19 marzo 2015
Popolazioni preistoriche scoperte ultimamente
SCHEDA 11: UOMO DI FLORES: HOMO FLORESIENSIS? VARIANTE PIGMEA
(FLORESIENSE) DI HOMO SAPIENS SAPIENS?
Nel 2003, sull’Isola di Flores, appartenente alle Isole Minori della Sonda, archeologi australiani e
indonesiani hanno repertato, nella ampia Grotta Liang Bua, una decina di scheletri incompleti e mal
fossilizzati di un ominide così piccolo da essere subito soprannominato “Hobbit”, assieme a
strumenti litici e resti di animali.
Negli anni successivi, sono stati dissotterrati nell’isola altri resti fossili umani ugualmente
minuscoli.
I fossili umani di Flores risalgono a un periodo compreso tra 38.000 e 13.000 aa BP, ma alcuni
archeologi retrodatano i più antichi resti dello Hobbit a oltre 95.000 aa BP. Molto probabilmente
l’Uomo di Flores è convissuto, per un discreto lasso di tempo, con Homo Sapiens Sapiens, ma
finora ne manca la prova; qualche archeologo ipotizza che Homo Sapiens Sapiens abbia forse
sfruttato il piccolo ominide in un ruolo ancillare.
Nel 2010, da uno scavo condotto nella Grotta Callao, sull’Isola Filippina di Luzon/Kalusunan, è
emerso un piccolo osso umano metatarsale, risalente a circa 67.000 aa BP, del tutto simile al
corrispondente ossicino di Flores. Questo reperto documenta che – più di 60.000 aa BP – anche
un’altra isola dell’Oceano Pacifico Centro-Occidentale era abitata da ominidi di aspetto
pigmoide.
Il primo paleo-antropologo che ha studiato il materiale scheletrico di Liang Bua, Teuku Jacob
(Indonesia) – per ragioni politico-nazionalistiche – l’ha danneggiato intenzionalmente con lo scopo di
“modernizzarlo” e di potere sostenere che esso appartiene a una popolazione Sapiens Sapiens
Pigmea [simile a quella Malese e Andamana] e che l’unico cranio è patologico (microcefalo), quindi
antropologicamente inutilizzabile.

23
Studiato successivamente da Yousuke Kaifu (Tokyo), il cranio (LB1) è stato, invece, considerato
indenne da patologie, ma fisiologicamente minuscolo a causa del nanismo insulare.
Gli animali che vivono lungamente relegati nelle isole, infatti, vanno incontro a riduzione dimensionale;
anche l’encefalo impicciolisce, ma subisce una riorganizzazione neurologica, che gli assicura il
mantenimento della funzionalità acquisita e spesso gli conferisce anche qualche miglioria/vantaggio;
la competenza dell’Uomo di Flores nell’uso del fuoco, nella lavorazione della pietra e nella caccia ad
animali di taglia medio-grande ne sarebbero la prova. Secondo l’Autore giapponese, la popolazione
nana discenderebbe da Homo Erectus Javanensis, che era già di taglia piuttosto ridotta rispetto al
corrispettivo africano, presumibilmente migrato nell’isola in tempi remoti. Confermerebbe questa
ipotesi il ritrovamento di manufatti litici, datati a circa 1 milione aa BP, sia al fondo di Liang Bua
che in altri vicini siti archeologici (Mata Menge e Wolo Sege). Homo Erectus Javanensis aveva
capacità cerebrale di circa 850 cc, suscettibile di riduzione teorica volumetrica di oltre la metà per
effetto del nanismo da segregazione insulare. A sostegno di queste vedute, Kaifu enfatizza il
nanismo di altri animali convissuti con l’Uomo di Flores, per esempio gli Stegodonti pigmei.
Oggi sappiamo che il cosiddetto “Effetto Isola” è un fenomeno reale, non solo a Flores; si tratta
di un processo biologico ancora incompletamente noto, che si manifesta con variazioni dimensionali
estreme, mai però finora documentato nel Genere Homo.
Secondo studi recenti (2008), le specie più grandi tendono a ridurre la propria stazza per non
sfruttare eccessivamente le limitate risorse insulari, mentre le specie più piccole tendono ad
accrescersi per l’assenza di predatori o competitori esistenziali; entrambe le modalità – nanismo e
gigantismo insulare – si sono verificate nell’Isola di Flores, dove oltre agli elefanti nani, si sono
trovati i fossili di un enorme marabù (Leptoptilos Robustus) e gli ultimi esemplari di un ratto
gigantesco (Papagomys Amandvillei); i varani (Varanus Komodoensis), lucertoloni aggressivi,
oggi relegati nelle Isole di Komòdo e Rinca, hanno abitato pure Flores in passato.
Oggi, dopo più di dieci anni di studi e discussioni, il problema dello Hobbit di Flores è tutt’altro che
risolto:
- Secondo alcuni Autori (Robert B. Eckhardt & Coll.), l’Uomo di Flores appartiene alla specie
Homo Sapiens Sapiens, sebbene alcuni fossili presentino i segni di una patologia
malformativa [Malattia di L. Down (Trisomia 21)].
- Altri Autori (Katerina Harvati, Karen L. Baab, Dean Falk) ipotizzano che l’ominide
appartenga a una nuova specie del Genere Homo, denominata da P. Brown, Homo
Floresiensis, differente da Homo Sapiens Sapiens, seppure discendente da un antenato
africano comune.

24
Purtroppo, il clima caldo-umido di Flores ha danneggiato il DNA dei fossili umani in misura tale
da impedire qualsiasi analisi genetica.
Museo Scienze Naturali, Tokyo
La popolazione preistorica sarebbe stata sterminata – assieme agli altri animali peculiari di Flores – da
un’eruzione vulcanica risalente a circa 13.000 aa BP, geologicamente documentata. Molti
ricercatori e cripto-zoologi, però, sono convinti che qualche Hobbit sia scampato e viva
nascosto nelle foreste pluviali inesplorate dell’isola, dalle quali spunterebbe ogni tanto. Questa
ipotesi è alimentata dalla credenza popolare nell’esistenza di una sorta di nanetto scimmiesco di
sorprendente agilità, noto come Ebu Gogo, del tutto simile all’Orang Pendek della vicina Isola di
Sumatra, di cui è stata trovata solo qualche traccia concreta. La possibile esistenza di questo
misterioso essere vivente ha anche interessato qualche Società Scientifica Anglo-Americana, che ha
organizzato spedizioni speciali con lo scopo di verificare l’ipotesi; la scrittrice inglese, Deborah Martyr
è riuscita ad avvistare il mostriciattolo tre volte e a eseguire il calco di alcune sue orme.
A conclusione del paragrafo, desidero farvi notare che la diatriba sull’esistenza di Homo Floresiensis
ha relegato in secondo piano il problema dell’arrivo della popolazione nana nell’Isola di Flores;
infatti, il mare circostante è tanto profondo da non essersi mai prosciugato neppure durante
l’Ultimo Massimo Glaciale, così da costituire una barriera di alcune decine di chilometri
insormontabile a piedi. La presenza della popolazione nana preistorica a Flores precedentemente
all’arrivo dell’uomo moderno, si spiega solo ipotizzando che sia stato un suo antenato, come Homo
Erectus, a raggiungere l’isola moltissimi anni prima, quando ciò era reso possibile da qualche
collegamento geologico. A sostegno di questa tesi depongono due dati paleontologici: (I) alcuni
utensili litici arcaici recuperati nel sito archeologico di Mata Menge sono costituiti di selce
sicuramente non-locale; (II) nello stesso giacimento, sono stati repertati resti fossili di animali
tipici della terraferma dell’Asia Meridionale (coccodrilli, stegodonti giganti, molluschi d’acqua
dolce, etc.).
Supporta questa tesi anche il recentissimo ritrovamento (2014) di una mandibola frammentaria, ma
ben conservata (Penghu 1), ripescata casualmente presso l’Isola di Taiwan (Repubblica della
Cina), da un bassofondo che era sicuramente emerso durante il Pleistocene Medio; il reperto
documenta la presenza, nelle isole dell’Oceano Pacifico Centro-Occidentale, di un ominide arcaico
del Genere Homo, tra 450.000 e 200.000 aa BP, assegnato a una variante finora sconosciuta di
Homo Erectus.
25
SCHEDA 12: HOMO SAPIENS DENISOVANUS
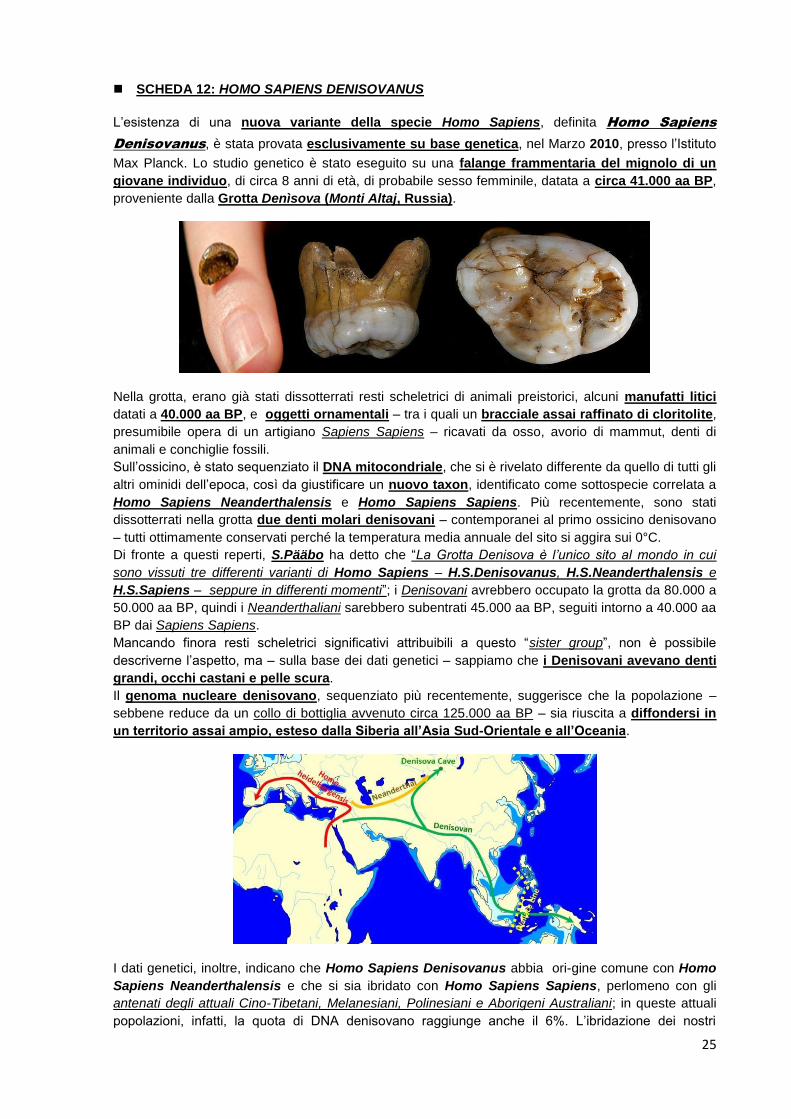
L’esistenza di una nuova variante della specie Homo Sapiens, definita Homo Sapiens
Denisovanus, è stata provata esclusivamente su base genetica, nel Marzo 2010, presso l’Istituto
Max Planck. Lo studio genetico è stato eseguito su una falange frammentaria del mignolo di un
giovane individuo, di circa 8 anni di età, di probabile sesso femminile, datata a circa 41.000 aa BP,
proveniente dalla Grotta Denìsova (Monti Altaj, Russia).
Nella grotta, erano già stati dissotterrati resti scheletrici di animali preistorici, alcuni manufatti litici
datati a 40.000 aa BP, e oggetti ornamentali – tra i quali un bracciale assai raffinato di cloritolite,
presumibile opera di un artigiano Sapiens Sapiens – ricavati da osso, avorio di mammut, denti di
animali e conchiglie fossili.
Sull’ossicino, è stato sequenziato il DNA mitocondriale, che si è rivelato differente da quello di tutti gli
altri ominidi dell’epoca, così da giustificare un nuovo taxon, identificato come sottospecie correlata a
Homo Sapiens Neanderthalensis e Homo Sapiens Sapiens. Più recentemente, sono stati
dissotterrati nella grotta due denti molari denisovani – contemporanei al primo ossicino denisovano
– tutti ottimamente conservati perché la temperatura media annuale del sito si aggira sui 0°C.
Di fronte a questi reperti, S.Pääbo ha detto che “La Grotta Denisova è l’unico sito al mondo in cui
sono vissuti tre differenti varianti di Homo Sapiens – H.S.Denisovanus, H.S.Neanderthalensis e
H.S.Sapiens – seppure in differenti momenti”; i Denisovani avrebbero occupato la grotta da 80.000 a
50.000 aa BP, quindi i Neanderthaliani sarebbero subentrati 45.000 aa BP, seguiti intorno a 40.000 aa
BP dai Sapiens Sapiens.
Mancando finora resti scheletrici significativi attribuibili a questo “sister group”, non è possibile
descriverne l’aspetto, ma – sulla base dei dati genetici – sappiamo che i Denisovani avevano denti
grandi, occhi castani e pelle scura.
Il genoma nucleare denisovano, sequenziato più recentemente, suggerisce che la popolazione –
sebbene reduce da un collo di bottiglia avvenuto circa 125.000 aa BP – sia riuscita a diffondersi in
un territorio assai ampio, esteso dalla Siberia all’Asia Sud-Orientale e all’Oceania.
I dati genetici, inoltre, indicano che Homo Sapiens Denisovanus abbia ori-gine comune con Homo
Sapiens Neanderthalensis e che si sia ibridato con Homo Sapiens Sapiens, perlomeno con gli
antenati degli attuali Cino-Tibetani, Melanesiani, Polinesiani e Aborigeni Australiani; in queste attuali
popolazioni, infatti, la quota di DNA denisovano raggiunge anche il 6%. L’ibridazione dei nostri
26
progenitori Sapiens Sapiens con Neanderthaliani e
Denisovani ha determinato effetti sicuramente
benefici per la nostra specie; segnatamente: (1) A
seguito del flusso genetico H.S.Denisovanus
H.S.Sapiens, si riscontra la presenza del gene
EPAS1 (Cromosoma 2) nel genoma sia dei
Denisovani che del 20% dei Cinesi attuali e
soprattutto dei Tibetani; questo gene si attiva in
condizioni di ipossia e promuove l’adattamento a
sopravvivere a grandi altitudini. (2) Gli scambi
genetici con Denisovani e Neanderthaliani hanno
conferito alla nostra specie una importante arma di
difesa da infezioni batteriche e virali e
degenerazioni neoplastiche, rappresentata dalla
acquisizione dei geni HLA (Cromosoma 6), che
stimolano il nostro apparato immunitario a
riconoscere gli antigeni patogeni, a mobilitare il
sistema linfocitario e a bloccarli/inattivarli.
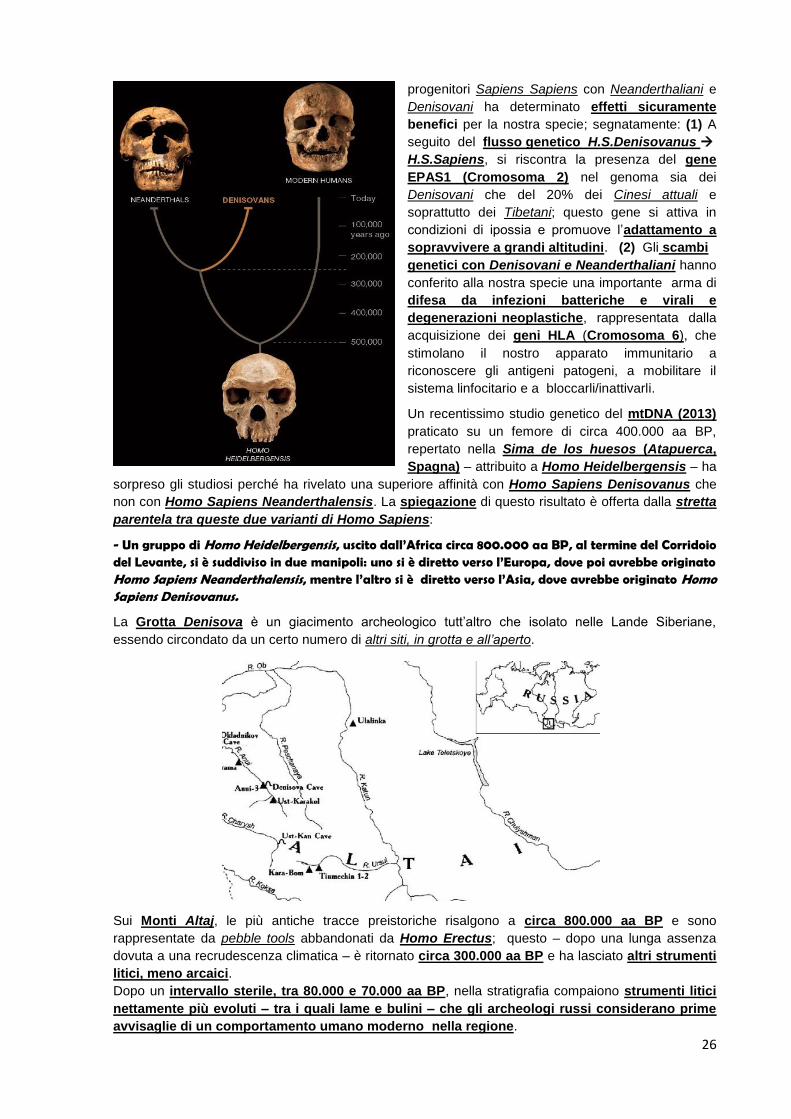
Un recentissimo studio genetico del mtDNA (2013)
praticato su un femore di circa 400.000 aa BP,
repertato nella Sima de los huesos (Atapuerca,
Spagna) – attribuito a Homo Heidelbergensis – ha
sorpreso gli studiosi perché ha rivelato una superiore affinità con Homo Sapiens Denisovanus che
non con Homo Sapiens Neanderthalensis. La spiegazione di questo risultato è offerta dalla stretta
parentela tra queste due varianti di Homo Sapiens:
- Un gruppo di Homo Heidelbergensis, uscito dall’Africa circa 800.000 aa BP, al termine del Corridoio del Levante, si è suddiviso in due manipoli: uno si è diretto verso l’Europa, dove poi avrebbe originato Homo Sapiens Neanderthalensis, mentre l’altro si è diretto verso l’Asia, dove avrebbe originato Homo Sapiens Denisovanus.
La Grotta Denisova è un giacimento archeologico tutt’altro che isolato nelle Lande Siberiane,
essendo circondato da un certo numero di altri siti, in grotta e all’aperto.
Sui Monti Altaj, le più antiche tracce preistoriche risalgono a circa 800.000 aa BP e sono
rappresentate da pebble tools abbandonati da Homo Erectus; questo – dopo una lunga assenza
dovuta a una recrudescenza climatica – è ritornato circa 300.000 aa BP e ha lasciato altri strumenti
litici, meno arcaici.
Dopo un intervallo sterile, tra 80.000 e 70.000 aa BP, nella stratigrafia compaiono strumenti litici
nettamente più evoluti – tra i quali lame e bulini – che gli archeologi russi considerano prime
avvisaglie di un comportamento umano moderno nella regione.

27
La Grotta Denisova è stata presumibilmente prescelta da tanti ominidi sia perché (1) aveva una
apertura superiore per l’illuminazione interna e lo sfogo del fumo del focolare, sia perché (2) era
collocata in una posizione strategica per intercettare uomini/animali nella sottostante valle del fiume
Anuj.
Nei vicini siti di Kara Bom (Transizione P. Medio-P. Sup./Paleolitico Superiore Precoce) e di Ust’-
Karakol (Paleolitico Superiore Iniziale = Aurignaziano), entrambi all’aperto, il materiale
archeologico, raccolto dagli strati cronologicamente contemporanei a quelli dei fossili umani
denisovani, appartiene a una sofisticata cultura, finora attribuita a Homo Sapiens Sapiens; ulti-
mamente, però, alcuni archeologi russi si stanno domandando se questa industria – almeno in
parte – possa essere opera di Homo Sapiens Denisovanus; la conferma di questa ipotesi si
attende da nuovi ritrovamenti archeologici.
SCHEDA 13: POPOLAZIONE/GENTE DELLA CAVERNA DEL CERVO ROSSO [PCCR]
Oggi viene chiamata “Gente della Caverna del Cervo Rosso” un manipolo di umani vissuti al
termine del Pleistocene, noti anche come Uomini di Maludong-Longlin, strettamente somiglianti a
noi uomini moderni, estinti circa 11.000 aa BP.
I primi fossili della Popolazione della Caverna del Cervo Rosso (PCCR) – sebbene siano stati
presentati alla comunità scientifica solo nel Marzo 2012 – sono stati recuperati nel Sud-Ovest della
Cina, nelle Grotte di Longlin (Regione Autonoma di Guangxi), nel 1979; altri resti scheletrici fossili
della medesima popolazione sono stati recuperati – assieme a ossame di Cervo Rosso Gigante
Preistorico, a tracce di ceramica e a un focolare con cenere e carboni – dieci anni dopo, nel 1989,
nella piccola Caverna Maludong/Grotta del Cervo Rosso (Provincia di Yunnan), distante circa 300
Km da Longlin. Numerosissimi resti scheletrici e odontologici, in parte combusti, di cervo rosso
preistorico, raccolti nei due siti, suggeriscono che l’alimentazione carnea di questi ominidi fosse
essenzialmente assicurata dalla caccia al grosso ungulato.

28
I resti scheletrici solo recentemente sono stati riesaminati, datati con precisione tra 14.500 e 11.500
aa BP, e infine sottoposti a studio anatomico attento da parte di Ji Xueping (paleo-antropologo
cinese) e Darren Curnoe (paleo-antropologo australiano). I crani presentano qualche connotato
primitivo, mentre appare già più moderno il calco dell’encefalo, perlomeno per quanto attiene
ai lobi frontali.
Queste differenze risaltano chiaramente dal confronto del cranio fossile di Longlin con un
cranio contemporaneo. Perlopiù, i resti scheletrici di Longlin e Maludong appaiono manomessi
o comunque alterati: alcuni sembrano spezzati, altri sono stati colorati con ocra e/o combusti;
escludendo la improbabile ipotesi del cannibalismo, è logico pensare a un rituale funerario o
crematorio.
E’ ancora inspiegate la differenza dimensionale tra le mandibole repertate nei due siti di Longlin (più
grande) e di Maludong (più piccola) e la lavorazione della calotta cranica di Maludong, così da
ricavarne una sorta di tazza (con due piccoli fori ai lati); a proposito di quest’ultima, poiché tutti gli
altri esemplari conosciuti di coppe ricavate dalla manipolazione di teschi sono opere di uomini
moderni, anche questo deve considerarsi prodotto da Homo Sapiens Sapiens.
I paleo-antropologi sono stati finora riluttanti ad assegnare questa gente a una nuova specie
umana. Oggi, quattro sono le ipotesi prevalenti:
Questa popolazione rappresenta il risultato della ibridazione tra Homo Sapiens
Denisovanus e Homo Sapiens Sapiens (C.Stringer).
La PCCR discende da una ennesima, antica e tuttora ignota migrazione di ominidi
dall’Africa, che avrebbero mantenuto connotati anatomici arcaici (J.Xueping e D.Curnoe).
Questa gente è una “variante robusta” di Homo Sapiens Sapiens (E.Trinkaus (USA),
P.Gunz (D), etc.).
La PCCR rappresenta il punto d’arrivo di una linea evolutiva originale sviluppatasi
autonomamente in Oriente da Homo Erectus, così da costituire una nuova specie; questa tesi
è propugnata dai paleo-antropologi dell’Asia Orientale, segnatamente Cinesi.
La maggioranza degli archeologi europei ritiene che questa gente appartenga a una variante
della specie Homo Sapiens Sapiens. La soluzione del problema non può venire che dalla
verifica del genoma, che però non può essere sequenziato sul materiale fossile oggi disponibile,
perché tutte le le ossa sono combuste e/o malconservate.
Alcuni dati archeologici suggeriscono che la PCCR sia vissuta in pieno isolamento sociale dalle
altre popolazioni vicine, rappresentate presumibilmente da gruppi di uomini moderni ormai di
Cultura Mesolitica.
Il ritrovamento di un mucchietto di chicchi di riso selvatico nella Grotta Maludong, ha fatto
supporre che quegli ominidi stessero iniziando i primi tentativi di sviluppare una cultura agricola,
29
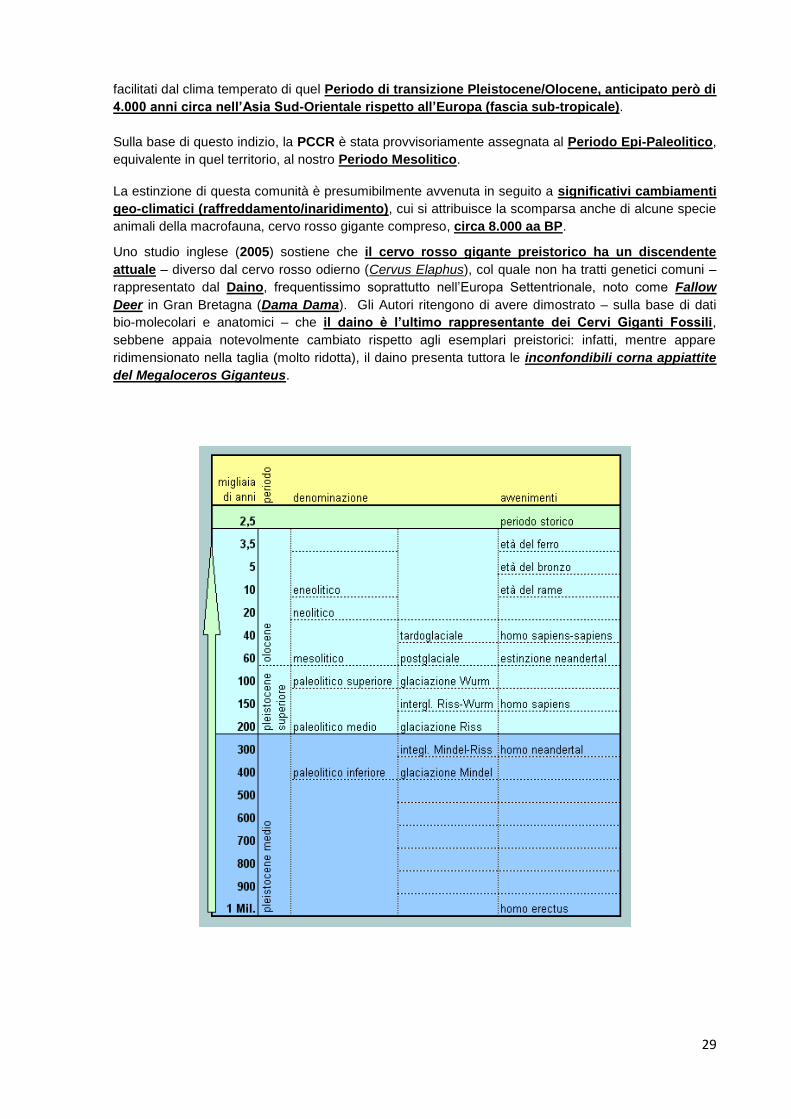
facilitati dal clima temperato di quel Periodo di transizione Pleistocene/Olocene, anticipato però di
4.000 anni circa nell’Asia Sud-Orientale rispetto all’Europa (fascia sub-tropicale).
Sulla base di questo indizio, la PCCR è stata provvisoriamente assegnata al Periodo Epi-Paleolitico,
equivalente in quel territorio, al nostro Periodo Mesolitico.
La estinzione di questa comunità è presumibilmente avvenuta in seguito a significativi cambiamenti
geo-climatici (raffreddamento/inaridimento), cui si attribuisce la scomparsa anche di alcune specie
animali della macrofauna, cervo rosso gigante compreso, circa 8.000 aa BP.
Uno studio inglese (2005) sostiene che il cervo rosso gigante preistorico ha un discendente
attuale – diverso dal cervo rosso odierno (Cervus Elaphus), col quale non ha tratti genetici comuni –
rappresentato dal Daino, frequentissimo soprattutto nell’Europa Settentrionale, noto come Fallow
Deer in Gran Bretagna (Dama Dama). Gli Autori ritengono di avere dimostrato – sulla base di dati
bio-molecolari e anatomici – che il daino è l’ultimo rappresentante dei Cervi Giganti Fossili,
sebbene appaia notevolmente cambiato rispetto agli esemplari preistorici: infatti, mentre appare
ridimensionato nella taglia (molto ridotta), il daino presenta tuttora le inconfondibili corna appiattite
del Megaloceros Giganteus.

30
INDICE
PRESENTAZIONE
1^ Conversazione :5 marzo 2015 pag. 5
Apparizione dei Primati. Ominidi Pre-Umani ancestrali
SCHEDA 01: TRANSIZIONE BIOTICA DI 65 MILIONI AA BP
SCHEDA 02: CRITERI RIVELATORI DI UMANITA’
SCHEDA 03: INIZIO DELLA STORIA DELL’UMANITA’
SCHEDA 04: PRIMI ESPONENTI DEL GENERE HOMO
2^ Conversazione :12 marzo 2015 pag. 13
Ominidi Pre-Umani recenti
SCHEDA 05: TEORIE EVOLUTIVE DOPO IL RITROVAMENTO DI HOMO
ERECTUS IN SITI EXTRA-AFRICANI
SCHEDA 06: ESPONENTI PIU’ RECENTI DEL GENERE HOMO: (1) HOMO
ANTECESSOR E HOMO HEIDELBERGENSIS
SCHEDA 07: ESPONENTI PIU’ RECENTI DEL GENERE HOMO: (2) HOMO
SAPIENS NEANDERTHALENSIS
SCHEDA 08: ESTINZIONE DI HOMO SAPIENS NEANDERTHALENSIS
SCHEDA 09: ULTIMI ESPONENTI DEL GENERE HOMO: (3) HOMO
SAPIENS IDALTU E HOMO SAPIENS SAPIENS
SCHEDA 10: PIU’ ANTICA POPOLAZIONE MODERNA EUROPEA
3^ Conversazione :19 marzo 2015 pag. 22
Popolazioni preistoriche scoperte ultimamente
SCHEDA 11: UOMO DI FLORES: HOMO FLORESIENSIS? VARIANTE
NANA/PIGMEA (FLORESIENSE) DI HOMO SAPIENS SAPIENS?
SCHEDA 12: HOMO SAPIENS DENISOVANUS
SCHEDA 13: POPOLAZIONE/GENTE DELLA CAVERNA DEL CERVO ROSSO
I testi sono stati elaborati e sintetizzati dal dr Alberto Catalano con la collaborazione grafica di Isa Montanari


















