
illu cap5 visione colore - fox.ino.it · Cap.5 La visione del colore 5.1 Il ruolo dei fotorecettori...
13
Cap.5 La visione del colore 5.1 Il ruolo dei fotorecettori Parlando di visione del colore la parte del nostro sistema visivo che svolge un compito importantissimo è la retina, cioè la membrana sensibile dell'occhio, in cui vengono generati i segnali nervosi in risposta allo stimolo visivo [Fiorentini]. Le cellule sensibili alla luce all'interno della retina sono i fotorecettori, di cui ne esistono due tipi: i coni e i bastoncelli. Complessivamente nell'occhio umano vi sono circa 6 milioni di coni e 125 milioni di bastoncelli, ma nella parte centrale della retina (la fovea) i coni sono in maggioranza. I bastoncelli sono i responsabili della visione notturna (visione scotopica), mentre i coni funzionano in condizioni di luce maggiore (visione fotopica). In realtà nel settore illuminotecnico ha creato grande interesse la scoperta del terzo recettore retinico [Berson]: è stato osservato che alcune cellule a livello di cellule gangliari (prima cioè che la luce giunga su coni e bastoncelli) assorbono la luce e ciò sembra avere interesse soprattutto per le sue possibili correlazioni pratiche legate al ciclo circadiano [Barinaga] [Van Bommel] [Rea]. Compito di questo recettore sarebbe quello di cambiare i livelli di melatonina, influenzando in questo modo anche la capacità di attenzione e di vigilanza delle persone. Si è anche cercato di determinare una curva di risposta di questo recettore, che mostrerebbe una maggiore sensibilità nella zona delle corte lunghezze d’onda (Fig.5.1). Fig.5.1 Sensibilità del “terzo”recettore retinico (Empirical circadian) paragonata con quella di coni (Photopic) e bastoncelli (Scotopic). Il terzo recettore ha un picco di sensibilità su lunghezze d’onda più corte rispetto alla sensibilità scotopica, dunque si è ipotizzato che illuminando con lampade che emettono maggiormente sul blu si possa favorire la produttività, poichè una persona è più ”sveglia”
Transcript of illu cap5 visione colore - fox.ino.it · Cap.5 La visione del colore 5.1 Il ruolo dei fotorecettori...

Cap.5 La visione del colore 5.1 Il ruolo dei fotorecettori Parlando di visione del colore la parte del nostro sistema visivo che svolge un compito importantissimo è la retina, cioè la membrana sensibile dell'occhio, in cui vengono generati i segnali nervosi in risposta allo stimolo visivo [Fiorentini]. Le cellule sensibili alla luce all'interno della retina sono i fotorecettori, di cui ne esistono due tipi: i coni e i bastoncelli. Complessivamente nell'occhio umano vi sono circa 6 milioni di coni e 125 milioni di bastoncelli, ma nella parte centrale della retina (la fovea) i coni sono in maggioranza. I bastoncelli sono i responsabili della visione notturna (visione scotopica), mentre i coni funzionano in condizioni di luce maggiore (visione fotopica). In realtà nel settore illuminotecnico ha creato grande interesse la scoperta del terzo recettore retinico [Berson]: è stato osservato che alcune cellule a livello di cellule gangliari (prima cioè che la luce giunga su coni e bastoncelli) assorbono la luce e ciò sembra avere interesse soprattutto per le sue possibili correlazioni pratiche legate al ciclo circadiano [Barinaga] [Van Bommel] [Rea]. Compito di questo recettore sarebbe quello di cambiare i livelli di melatonina, influenzando in questo modo anche la capacità di attenzione e di vigilanza delle persone. Si è anche cercato di determinare una curva di risposta di questo recettore, che mostrerebbe una maggiore sensibilità nella zona delle corte lunghezze d’onda (Fig.5.1).
Fig.5.1 Sensibilità del “terzo”recettore retinico (Empirical circadian) paragonata con quella di coni (Photopic) e bastoncelli (Scotopic). Il terzo recettore ha un picco di sensibilità su lunghezze d’onda più corte rispetto alla sensibilità scotopica, dunque si è ipotizzato che illuminando con lampade che emettono maggiormente sul blu si possa favorire la produttività, poichè una persona è più ”sveglia”

e attenta. Sono stati fatti esperimenti sulla produttività dei lavoratori in funzione dell’illuminazione [Juslen]. Per tornare ai recettori noti da più tempo, i bastoncelli contengono tutti uno stesso pigmento (la rodopsina o porpora visiva), mentre nell'uomo i coni contengono tre tipi diversi di pigmenti, che assorbono in maniera diversa le varie radiazioni dello spettro. I tre diversi tipi di pigmenti sono in grado di assorbire lunghezze d’onda diverse. In particolare i coni detti lunghi o rossi (coni L) hanno un picco di assorbimento massimo per λmax.=565 nm, i coni medi o verdi (cono M) hanno λmax.=530 nm, per i coni corti o blu (coni S) λmax.=420. Il “colore” dei coni corrisponde alla zona dello spettro che sono in grado di assorbire. I bastoncelli hanno invece un massimo di assorbimento per λmax.=499 nm [Dartnall](Fig.5.2). La presenza di tre diversi pigmenti ci permette di distinguere i vari colori tra loro. Infatti un singolo tipo di pigmento non sarebbe in grado di discriminare i colori, poiché non saremmo in grado, ad esempio, di distinguere tra luce che giunge alla lunghezza d’onda ottimale di un cono (come i 565 nm) e luce che giunge in maggiore intensità, ma ad una lunghezza d’onda a cui l’occhio è meno sensibile. In pratica con un solo ricettore dei colori noi non potremmo ottenere una risposta univoca, non ottenendo una corretta visione dei colori, poichè il singolo cono non distingue la lunghezza d’onda che riceve, ma solo il numero di fotoni che assorbe. Questo fatto viene detto principio di univarianza di Rushton, ed è confermato dalle analisi sui soggetti umani anomali aventi un solo ricettore del colore così come dagli studi sugli animali con un solo tipo di cono. Per avere visione del colore la condizione minima è che la retina contenga due tipi di fotopigmenti con diverse proprietà di assorbimento spettrale.
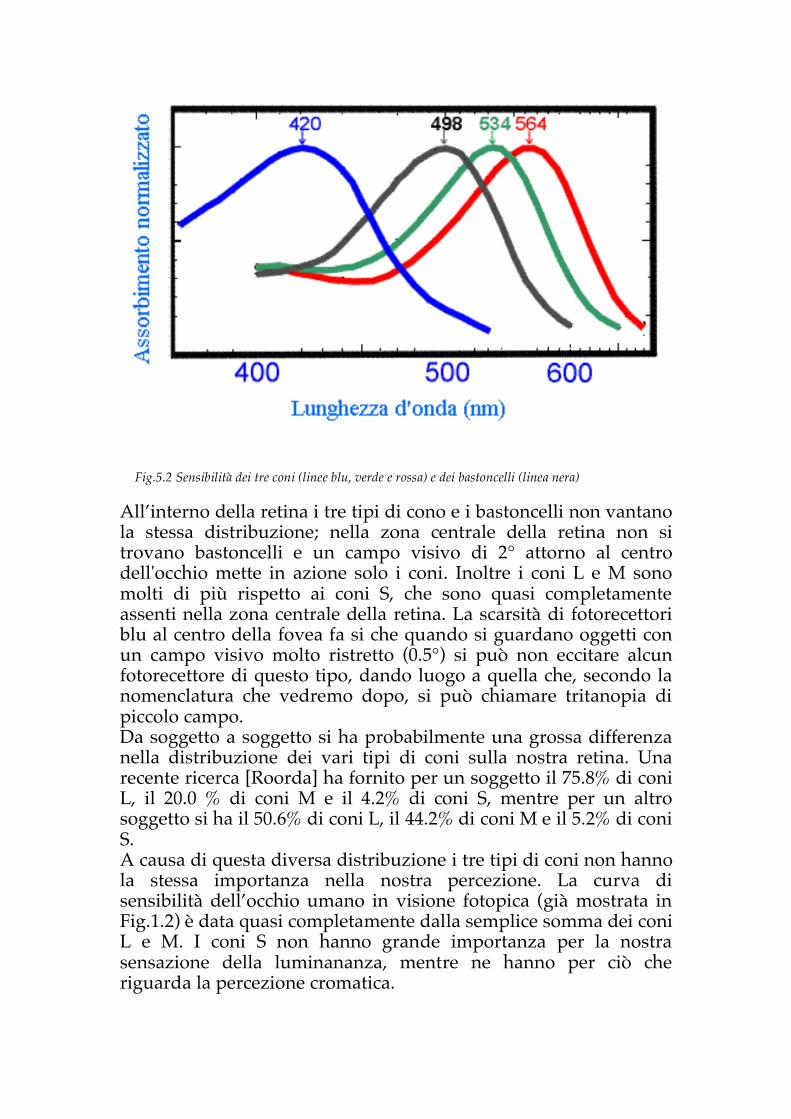
All’interno della retina i tre tipi di cono e i bastoncelli non vantano la stessa distribuzione; nella zona centrale della retina non si trovano bastoncelli e un campo visivo di 2° attorno al centro dell'occhio mette in azione solo i coni. Inoltre i coni L e M sono molti di più rispetto ai coni S, che sono quasi completamente assenti nella zona centrale della retina. La scarsità di fotorecettori blu al centro della fovea fa si che quando si guardano oggetti con un campo visivo molto ristretto (0.5°) si può non eccitare alcun fotorecettore di questo tipo, dando luogo a quella che, secondo la nomenclatura che vedremo dopo, si può chiamare tritanopia di piccolo campo. Da soggetto a soggetto si ha probabilmente una grossa differenza nella distribuzione dei vari tipi di coni sulla nostra retina. Una recente ricerca [Roorda] ha fornito per un soggetto il 75.8% di coni L, il 20.0 % di coni M e il 4.2% di coni S, mentre per un altro soggetto si ha il 50.6% di coni L, il 44.2% di coni M e il 5.2% di coni S. A causa di questa diversa distribuzione i tre tipi di coni non hanno la stessa importanza nella nostra percezione. La curva di sensibilità dell’occhio umano in visione fotopica (già mostrata in Fig.1.2) è data quasi completamente dalla semplice somma dei coni L e M. I coni S non hanno grande importanza per la nostra sensazione della luminananza, mentre ne hanno per ciò che riguarda la percezione cromatica.
Fig.5.2 Sensibilità dei tre coni (linee blu, verde e rossa) e dei bastoncelli (linea nera)

Le strutture genetiche di tutti i pigmenti visivi che si incontrano in natura sono molto simili, il che fa pensare che tutti i pigmenti visivi derivino da uno stesso antico precursore [Applebury]. I fotopigmenti presenti in specie animali molto diverse hanno lo stesso λmax ed uno spettro di assorbimento identico. In generale si può considerare che la visione del colore è tanto più precisa quanto più tipi di pigmenti ci sono: molti mammiferi hanno due pigmenti, mentre alcuni uccelli, come i piccioni, hanno cinque pigmenti diversi. E' interessante chiedersi se vi sia una logica nell'evoluzione delle proprietà assorbenti dei coni. In tutti i mammiferi aventi un solo tipo di cono, il picco di sensibilità si trova vicino al picco di sensibilità dei bastoncelli, il che potrebbe far pensare che i coni siano derivati dai bastoncelli. Ma all'interno dei primati l'unico cono comune tra tutte le specie ha un λmax di circa 560 nm, il che fa pensare che quella sia la posizione primordiale dei coni del colore [Jacobs]. All'interno dei mammiferi vi sono molte diversità nella visione. Un tempo si pensava che molti mammiferi non avessero una visione del colore, in realtà la maggior parte tra essi vedono i colori. Una piccola parte (ad esempio i topi [Neitz]) non hanno visione del colore. Anche tutti i mammiferi che non hanno visione del colore presentano comunque dei coni sulla loro retina, si potrebbero cioè definire tecnicamente, usando le catergorie che vedremo dopo, come monocromati da coni e non da bastoncelli. La presenza dei coni in tutti i mammiferi può dare credito alle teorie che sostengono che i coni sono i recettori ancestrali. Anche se molti mammiferi hanno una visione del colore, solo alcuni primati hanno una visione di tipo tricromatico. Ad esempio le scimmie, a seconda della loro provenienza geografica, possono avere tre coni assai simili ai nostri, oppure due soli coni o, in certi casi, tre coni tutti nella zona del rosso verde. La presenza nell’uomo di tre tipi di fotorecettori si accorda benissimo con la necessità di avere tre colori linearmente indipendenti per poter ottenere tutti gli altri, secondo la prima legge di Grassmann [Ronchi]. La prima intuizione che la nostra visione dei colori non poteva basarsi su un'analisi puntuale di tutte le lunghezze d'onda si deve a Thomas Young. Egli scriveva “Poiché è quasi impossibile ammettere che ciascun punto sensibile della retina contenga un numero infinito di particelle ognuna capace di vibrare perfettamente all'unisono con ogni possibile lunghezza d'onda bisogna necessariamente concludere che il numero dei recettori sia limitato, per esempio, ai tre colori principali, rosso giallo e blu”.

6.3 Le cellule gangliari e la teoria di Hering I segnali generati dai coni vengono trasmessi alle cellule bipolari e da qui alle cellule gangliari, che poi passano i segnali direttamente al cervello tramite i loro assoni, che costituiscono il nervo ottico. Le cellule gangliari emettono verso il cervello gli impulsi nervosi; in particolare l'effetto di un segnale ricevuto dalla cellula gangliare è quello di modificare la frequenza degli impulsi. Dato che i recettori retinici sono circa 126 milioni, mentre gli assoni del nervo ottico sono un milione è evidente che ci deve essere un meccanismo "a collo di bottiglia" che permette di comprimere le informazioni. Ogni cellula gangliare risulta quindi connessa con un certo numero di fotorecettori, e stimolare uno di quei fotorecettori equivale a stimolare la cellula gangliare. L'area retinica connessa con una certa cellula gangliare è detta campo recettivo. Il campo recettivo è generalmente diviso in due zone: un cerchio centrale e un anello esterno. In questo modo in alcune cellule si ha un aumento di frequenza quando lo stimolo colpisce la parte centrale e una diminuzione quando lo stimolo colpisce l'anello (cellule centro on). In altre cellule si ha un processo esattamente opposto (cellule centro off). Questo sistema permette di migliorare la percezione dei contrasti locali. La visione dei colori è fortemente legata a questo tipo di struttura, dato che alcune cellule gangliari sono deputate a una discriminazione cromatica. In particolare sembra che esistano cellule con la parte centrale dedicata al rosso e l'anello interno dedicato al verde o viceversa (cellule rosso-verde) che sarebbero in grado di distinguere tra questi due colori, cellule invece in grado di confrontare il giallo (cioè la somma di rosso più verde) e il blu, e cellule che prendendo segnali da tutti e tre i tipi di coni forniscono un segnale di luminanza (Fig.5.?).

La presenza di queste cellule evidenzia che anche nella teoria di Hering, detta dell'opposizione dei colori, vi è qualcosa di vero. Secondo Hering il nostro sistema visivo conteneva due sistemi cromatici opponenti, il rosso-verde e il giallo-blu. Questo spiega anche perché nessun verde contenga mai del rosso e viceversa. In questo senso i colori fondamentali non sono più i tre della teoria tricromatica, ma diventano 4: rosso, verde, giallo, blu. La presenza di cellule centro on e centro off è legata alla capacità del nostro sistema visivo di interpretare i contrasti e di leggere i bordi delle immagini con precisione. 6.4 Anomalie della visione del colore Le deficienze nella visione del colore sono caratterizzate da un'anormale capacità di eguagliare certi colori e dall'impossibilità di distinguere colori che appaiono assai differenti per un soggetto normale. Tali problemi sono dovuti a un comportamento non regolare da parte dei fotopigmenti dell'occhio. Il soggetto può ad esempio avere nessun cono ricettore, o averne un solo tipo, oppure due rispetto ai tre delle persone dotate di visione normale. Oppure un soggetto può avere tre tipi di fotorecettori, ma uno di questi può comportarsi in maniera diversa dal comportamento atteso. In questo caso non si parla di veri dicromati, ma solo di tricromati anomali. I nomi delle anomalie variano a seconda del numero e del tipo di
Fig.5.? Schema della visione dei colori (da [Boynton])

fotorecettori assenti (vedi Tab.?). In Italia tali anomalie vengono detto Daltonismo, poiché il noto chimico John Dalton ne era affetto. Egli addirittura, credendo che il problema fosse a livello del cristallino, donò dopo la morte il suo occhio all'università perché fosse analizzato. L'occhio si trova tutt'ora, sotto spirito, in uno scaffale a Cambridge. Le deficienze più frequenti sono quelle dei dicromati e dei tricromati anomali. I dicromati hanno solo due tipi di fotorecettori, e eguagliano tutti i colori usando 2 sole variabili. In particolare i protanopi mancano del pigmento “rosso”, i deuteranopi del “verde” e i tritanopi del “blu”. I tricromati anomali necessitano invece di tre variabili per eguagliare tutti i colori. I colori che vengono percepiti come indistinguibili da parte dei dicromati possono essere identificati grazie alle linee isocromatiche tracciate sui diagrammi cromatici della CIE (vedi Fig.?) I colori che stanno lungo le linee risultano indistinguibili per il dicromate, a parte le differenze di luminanza. Paradossalmente vi sono situazioni in cui le persone affette da deficienza dei colori possono avere dei vantaggi rispetto agli osservatori normali: sono quelle circostanze in cui il colore è più un effetto di mascheramento che un'informazione in più. Ad esempio nella seconda guerra mondiale i soggetti dicromati gravi venivano spinti a prestare sevizio sui bombardieri, proprio per la loro capacità di vedere attraverso i camuffamenti colorati e di non farsi distrarre da quelle che, agli occhi degli osservatori normali, sarebbero apparse configurazioni cromatiche ingannevoli. Il nervo ottico contiene gli assoni delle celle dei ganglioni, e comprende circa un milione di fibre. Il piccolo numero di fibre rispetto al numero di ricettori provoca un fenomeno detto “di collo di bottiglia” per cui più segnali elettrici convergono a una singola cellula dei ganglioni. Questo tipo di schema permette di capire non solo le anomalie nella visione del colore, come vedremo in seguito, ma anche i fenomeni di colore soggettivi, non consistenti cioè con una distribuzione spettrale, ma [7] prodotti all'interno del sistema visivo senza un legame con la lunghezza d'onda che arriva effettivamente sull'occhio. Un esempio sono le righe verdi che si notano chiudendo gli occhi dopo aver osservato per un certo tempo delle righe rosse e bianche, o i colori che sembrano comparire all'interno di una serie fitta di righe bianche e nere. Per confermare tale teoria della visione del colore sono state realizzate alcune misure psicofisiche, quale quella illustrata nella Fig.? che mostra la capacità dell'occhio di discriminare le varie lunghezze d'onda. L'andamento dei massimi e dei minimi è coerente con la teoria sopra esposta.

L'analisi della retina e delle zone immediatamente successive non permette però di capire per intero il funzionamento nella visione dei colori. L'origine vera e propria della visione dei colori sembra ritrovarsi a un livello più elevato nell'organizzazione cerebrale. Un esempio di fenomeno la cui validità va ricercata a livello più alto è la costanza di colore. Per fortuna vi è un’altra capacità estremamente interessante del nostro cervello, detta costanza di colore, che ci permette di continuare a vedere un oggetto dello stesso colore anche sotto sorgenti spettralmente molto diverse, come può essere il caso di una lampada ad incandescenza, del cielo blu, del sole al tramonto o di una lampada fluorescente. Uno degli esperimenti più interessanti in questo settore fu realizzato da Land nel 1974, utilizzando mosaici di pezzi di carta colorata simili a un quadro di Mondrian. Proviamo a darne una breve descrizione. Per illuminare il dipinto si utilizzano, in una stanza buia, tre proiettori con tre filtri cromatici abbastanza stretti su corte (filtro blu), medie (filtro verde) e lunghe (filtro rosso) lunghezze d’onda. I tre proiettori sono guidati da tre potenziometri in modo da poterne variare l’intensità di emissione con continuità per un ampio intervallo di valori. Per prima cosa con tutti e tre i proiettori accesi in modo da poter simulare una sorgente luminosa “bianca” si valuta il colore di una delle macchie: immaginiamo che tale macchia appaia blu. Possiamo misurare, utilizzando uno spettrofotometro, la risposta di intensità luminosa della macchia tenendo accessi alternativamente ognuno dei tre proiettori ed ottenendo così una terna di valori corrispondente alla nostra percezione blu. A questo punto si misura con il nostro spettrofotometro un’altra macchia di colore diverso, ad esempio verde. Accendendo solo il proiettore rosso si modifica il valore del potenziometro in modo da riprodurre esattamente la stessa lettura che abbiamo avuto per la macchia blu. La stessa procedura si ripete per gli altri due proiettori. E’ facile immaginare che, a questo punto, se accendiamo tutti e tre i proiettori contemporaneamente la nostra macchia apparirà blu, dato che le letture coincidono: in realtà la macchia continua ad apparire verde, senza preoccuparsi dell’equivalenza delle letture. Per Land questo provava che il colore di una superficie all’interno di una scenario complesso non dipendeva solo dallo stimolo fisico. In un certo qual modo il nostro cervello deve essere in grado di valutare le caratteristiche spettrali di un illuminante e deve poter ricostruire le caratteristiche della superficie illuminata, deve ciò saper “sottrarre l’illuminante”. Questo tipo di capacità è di fondamentale importanza per poter assegnare un valore all’informazione legata al colore, e per questa ragione è una capacità che non è esclusiva degli essere umani, come ha provato un interessante esperimento di Ingle del 1985 condotto sui pesci. Egli addestrò gli animali a raggiungere sempre, all’interno di un dipinto

stile Mondrian, le macchie di un determinato colore. Quello che notò fu che i pesci tendevano sempre a dirigersi verso le macchie di quel colore anche quando il cambiamento di illuminazione provocava una variazione nelle lunghezze d’onda riflesse dalla superfici: anche i pesci sono quindi dotati della costanza di colore. Questo fenomeno permette al progettista illuminotecnico di scegliere, entro certi limiti, lampade con una diversa resa del colore senza modificare eccessivamente la percezione cromatica delle persone (ed eventualmente anche degli animali) presenti nella stanza. Anche la costanza del colore ha dei limiti. Il più interessante è quello per cui essa non può funzionare quando il nostro campo visivo è occupato da una singola superficie colorata illuminata da una sorgente nascosta: in questo caso il sistema visivo non ha alcuna informazione utile per scomporre il contributo della superficie da quello della sorgente. Per avere costanza di colore è quindi assolutamente necessario avere a disposizione almeno due superfici di colore diverso all’interno della scena osservata, il che suggerisce che il colore di una superficie sia determinato non solo dalla composizione spettrale della luce riflesse dalla superficie, ma anche dalla composizione spettrale di tutto ciò che la circonda. E’ come se il nostro sistema visivo in questo modo fosse in grado di valutare i rapporti di riflettanza tra i vari colori. Molti tentativi sono stati fatti per evidenziare quali lampade lasciassero maggiormente intatta la costanza del colore. Quando due colori sono uguali? Si potrebbe pensare di rispondere a questa domanda attraverso gli spettri di riflettanza, cioè la percentuale di luce che viene riflessa indietro per ogni lunghezza d’onda. Ma tale definizione presupporrebbe una visione del colore basata su molti sensori che prendono in considerazione una singola lunghezza d’onda, mentre la risposta del nostro sistema visivo è mediata dalla presenza di soli tre coni (alcuni uccelli, come ad esempio i piccioni, possono vantare 5 tipi diversi di coni e possono quindi riconoscere una grande quantità di sfumature di colore) e non è possibile analizzare lo spettro in dettaglio. In realtà, come visto precedentemente, quando osserviamo un oggetto il nostro sistema visivo lo “vede” attraverso la risposta dei tre coni, osservando in pratica una media pesata dai coni dello spettro di riflettanza; per questa ragione può capitare che due colori risultino percettivamente uguali anche se fisicamente sono diversi. Si parlerà in questo caso di un’eguaglianza metamerica. A causa di questo funzionamento del nostro sistema visivo la frase “il bianco è la somma di tutti i colori” deve essere analizzata criticamente. Se è sicuramente vero che riunendo insieme i colori dello spettro precedentemente divisi da un prisma si ottiene luce bianca, è però vero che per ottenere la sensazione di bianco possono bastare due o tre luci colorate scelte intelligentemente. In fig.3.1 è illustrato ad esempio come

sia possibile ottenere il bianco nei LED utilizzando tre sorgenti (caso a sinistra) o in pratica due sorgenti (caso a destra)
Fig.3.1 Produzione del bianco nei LED. E’ quindi evidente che Il funzionamento del sistema visivo umano per ciò che riguarda la visione del colore può far si che due colori, pur avendo spettri di riflettanza completamente diversi appaiano dello stesso colore: si tratta del fenomeno del metamerismo e i due stimoli che appaiono identici si dicono metamerici. E’ un fenomeno che gli americani chiamano “problema della giacca e dei pantaloni”. Infatti capita che due capi di abbigliamento (si pensi in particolare a abiti scuri) possano apparire dello stesso colore sotto una particolare sorgente artificiale di illuminazione e diversi sotto un’altra. E’ abbastanza tipico in situazioni come queste esaminare gli oggetti sotto un’altra sorgente, tipicamente il Sole, per poter cogliere eventuali differenze. In Fig.3.2 sono rappresentati gli spettri di due oggetti di colore diverso. Questi due oggetti possono apparire dello stesso colore quando sono visti sotto una lampada ad alta temperatura di colore (cfr.capitolo 6), contenente cioè molta radiazione alle basse lunghezze d’onda, dove i due spettri sono simili. Ma se illuminiamo i due oggetti con una lampada ad incandescenza che contiene molta radiazione nel rosso, dove i due spettri differiscono, la differenza risulterà evidente anche percettivamente.

Fig.3.2 Due spettri metamerici Questo fenomeno sembra suggerire una robustissima influenza nella nostra percezione del colore da parte del tipo di sorgente luminosa che stiamo prendendo in considerazione, dipendenza che appare evidente quando si vogliono realizzare fotografie sotto la luce artificiale: un oggetto bianco, fotografato con una pellicola tradizionale, ma ritratto sotto una lampadina ad incandescenza, apparirà rosa-arancione. Uno spettroradiometro permette di evidenziare immediatamente se siamo di fronte a stimoli metamerici, dato che è in grado di fornirci lo spettro di riflettanza della superficie. Esistono però anche strumenti che, pensati per un funzionamento che ricordi la nostra visione, non rivelano il metamerismo e che vengono generalmente detti colorimetri. In tali strumenti non esiste l’elemento disperdente: la luce filtrata viene inviata su tre diverse fotocellule che permettono di elaborare tre numeri corrispondenti alla risposta sulle lunghezze d’onda corte, medie e lunghe. Non ha molto senso chiedersi quale misura sia più “vera”: quello che è certo è che il mestiere dei progettisti illuminotecnici risulterebbe estremamente impegnativo se il funzionamento del nostro sistema visivo si fermasse alla stretta dipendenza dallo spettro di riflettanza, dato che tutto la fatica andrebbe a ricadere nel tentativo, spesso utopico, di riprodurre con le nostre sorgenti artificiali l’illuminazione solare. A conferma di questo va ricordato che, come esistono acromatopsie congenite che descriveremo inseguito e che sono legate a problemi a livello retinico, sono state osservate acromatopsie di origine cerebrale. Si tratta di casi di persone che hanno perso completamente la visione a colori a causa di un danno cerebrale più o meno esteso. La neurologia ha a lungo escluso la possibile esistenza di tali patologie, che avrebbero provato la presenza di un centro di analisi del colore nel nostro cervello. Una prova inconfutabile fu fornita solo da Semir Zeki nel 1973 [8]. Egli riusci ad identificare, nella corteccia prestriata di ciascun emisfero cerebrale della scimmia, una piccola area (indicata con la sigla V4) che sembrava specializzata nella risposta al colore. Si tratta di un'area molto piccola anche nell'uomo, non più grande di un fagiolo, ma dalla sua integrità dipende probabilmente tutta la nostra percezione del colore [9]. Le persone affette da questo tipo di acromatopsia non perdono completamente la capacità di distinguere certe lunghezze d'onda, risultano anzi estremamente sensibili a fenomeni quali la variazione della sorgente luminosa che illumina la scena, come se fosse perduta la costanza di colore. Ma sono impossibilitati a ricostruire il colore di una scena. Questo poiché la zona V4 è il centro generatore del colore, centro

però che per fornire la corretta informazione deve dialogare con un centinaio di altri sistemi mesencefalici. L'integrazione vera e propria avviene poi a livelli superiori: la V4, scrive Sacks, "non è un punto di arrivo: essa non è che una stazione di passaggio, che proietta a sua volta a livelli sempre più alti, per arrivare infine all'ippocampo (essenziale per la memoria), ai centri emozionali del sistema limbico e dell'amigdala, e a molte altre aree della corteccia". La visione del colore sembra non richiedere alcuna conoscenza a priori, non è determinata dall'apprendimento o dall'esperienza, anche se l'aspettativa e la disposizione mentale alla visione del colore sono fattori importantissimi. Generalmente nelle persone affette da tale anomalia sia ha uno sviluppo maggiore di altre capacità visive. Recentemente è stato anche ipotizzato che una acromatopsia provocata dalla perdita della zona V4 possa essere guarita "addestrando" un'altra zona del cervello a svolgere questo compito. Il problema della visione dei colori resta estremamente impegnativo. Dice ancora Sacks "dopo aver descritto “il celebrato fenomeno dei colori” Newton evitò ogni ipotesi teorica sulla sensazione e non azzardò alcuna ipotesi sul “modo o l'azione con i quali la luce produce nella nostra mente i fantasmi dei colori”. A distanza di tre secoli, ancora non abbiamo alcuna ipotesi e può darsi che queste domande sino destinate a non trovare risposta". Allo stesso modo scrive Nordby “Sebbene abbia acquisito una conoscenza teorica approfondita della fisica dei colori e della fisiologia dei processi di recezione dei colori, nulla di tutto ciò può aiutarmi a comprendere quale sia la loro vera natura” [10]. 4 Test sulla visione del colore Dalle discussioni precedenti si può arguire il funzionamento e l'utilità dei vari test per la visione del colore. Esistono vari tipi di test che possono essere più o meno veloci, e conseguentemente più o meno precisi, e quindi adatti alle varie esigenze. I test più noti sono quelli basati sulle tavole pseudoisocromatiche, realizzate con l'uso di spot di vari colori, che risultano diversi per le persone dotate di visione normale, ma che appaiono uguali ai soggetti dicromati. Le tavole più conosciute sono quelle di Ishiara, che sono realizzate in modo tale da dar luogo a figure diverse (in particolare numeri) a seconda se chi guarda è un soggetto normale o anomalo. Tali tavole sono assai utili per la determinazione delle anomalie Deutan o Protan.

Molto usato, anche se leggermente più laborioso sia per l'effettuazione che per l'interpretazione è il cosiddetto test dei 100 toni, che si basa sulla capacità di discriminare e ordinare le varie tinte. In tutti questi tipi di test riveste ovviamente un'importanza fondamentale il tipo di illuminazione che viene utilizzato. I test sono solitamente realizzati per poter essere usati in maniera ottimale con un'illuminazione che secondo la catalogazione della CIE viene indicata di tipo C. Altri tipi di lampade possono alterare il risultato del test; ad esempio una illuminazione realizzata con lampade al tungsteno altera l'apparenza delle tavole di Ishiara, rendendo difficile la discriminazione non tanto dei dicromatici, il cui difetto rimane evidente, ma dei tricromati anomali. I manuali dei test non sempre riportano il livello di illuminamento, ma quasi tutti i laboratori lavorano con valori che oscillano tra 250 e 650 lux. Qualora vi sia la necessita di valutazioni più precise si deve ricorrere all'anomaloscopio. Esistono poi i test vocazionali, che riproducono il compito visuale di una certa occupazione, e permettono di determinare se una persona possa completarlo in maniera soddisfacente. Molti dei test vocazionali si basano sul ''color naming'', cioè sul saper indicare il nome di certi colori. Tale tecnica, inizialmente utilizzata appunto solo nel campo dei test vocazionali, è recentemente assurta a una certa rilevanza scientifica. Si è notato [11] che molti veri dicromati sono in grado di categorizzare i colori chiamandoli per nome. Ovviamente i loro risultati sono inferiori rispetto a quelli dei normali tricromati, ma molte volte migliori rispetto a quello che ci si sarebbe potuto aspettare da dei dicromati. In un primo tempo si attribuì tali risultati alle capacita di discriminazione dei colori da parte dei bastoncelli. Un successivo lavoro [12] ha mostrato l'importanza, per un più corretto riconoscimento da parte dei dicromati, di stimoli di colore estesi spazialmente e temporalmente, attribuendo importanza ai ricettori contenenti pigmenti anomali.















![Il colore [1] Il colore La percezione del colore La psicologia del colore La cromoterapia.](https://static.fdocumenti.com/doc/165x107/5542eb65497959361e8d06ae/il-colore-1-il-colore-la-percezione-del-colore-la-psicologia-del-colore-la-cromoterapia.jpg)


