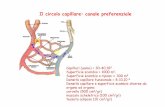
FONTI DI AMMINOACIDI: PROTEINE ASSUNTE CON GLI...
24
ENZIMI PROTEOLITICI: ZIMOGENI ATTIVATI NEL LUME INTESTINALE DURANTE LA DIGESTIONE DELLE PROTEINE ESOGENE Tripsina, chimotripsina, carbossipeptidasi A e B, amminopeptidasi, enteropeptidasi, elastasi FONTI DI AMMINOACIDI: PROTEINE ASSUNTE CON GLI ALIMENTI PROTEINE ENDOGENE (TURNOVER) BIOSINTESI Berg et al., BIOCHIMICA 6/E, Zanichelli editore S.p.A. Copyright © 2007 PROTEINE INGERITE CON GLI ALIMENTI
Transcript of FONTI DI AMMINOACIDI: PROTEINE ASSUNTE CON GLI...
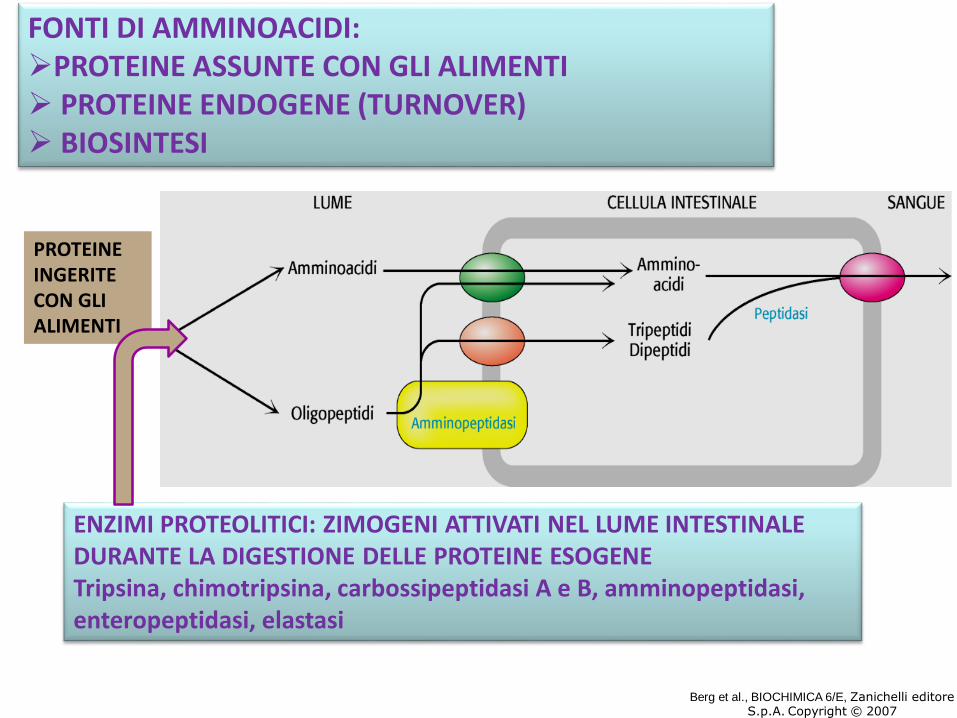
ENZIMI PROTEOLITICI: ZIMOGENI ATTIVATI NEL LUME INTESTINALE DURANTE LA DIGESTIONE DELLE PROTEINE ESOGENETripsina, chimotripsina, carbossipeptidasi A e B, amminopeptidasi, enteropeptidasi, elastasi
FONTI DI AMMINOACIDI:PROTEINE ASSUNTE CON GLI ALIMENTI PROTEINE ENDOGENE (TURNOVER) BIOSINTESI
Berg et al., BIOCHIMICA 6/E, Zanichelli editoreS.p.A. Copyright © 2007
PROTEINE INGERITE CON GLI ALIMENTI
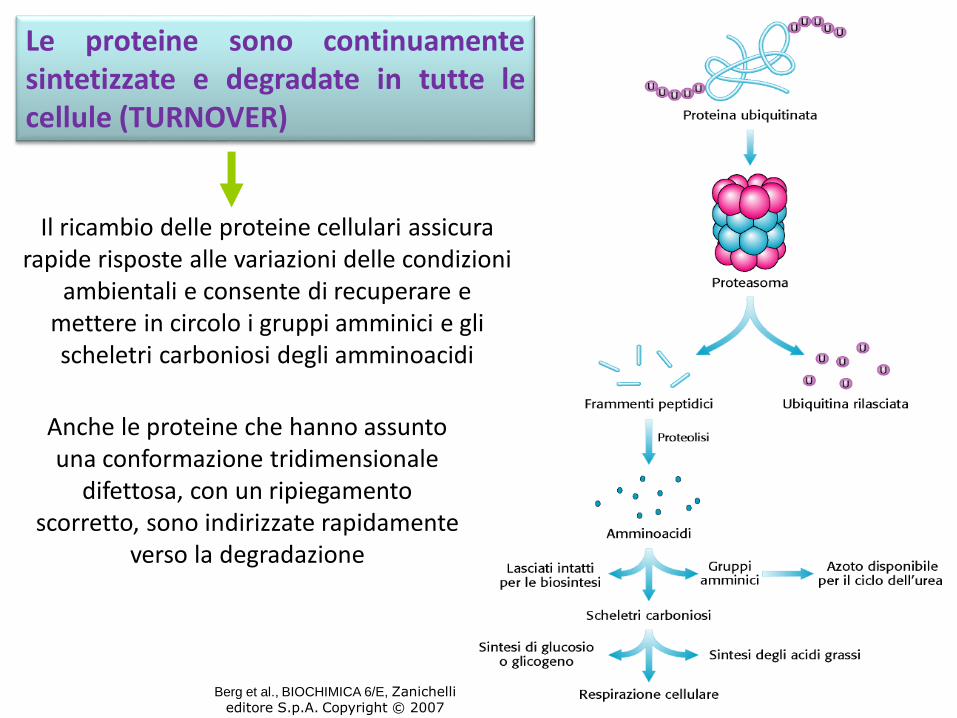
Le proteine sono continuamentesintetizzate e degradate in tutte lecellule (TURNOVER)
Il ricambio delle proteine cellulari assicura rapide risposte alle variazioni delle condizioni
ambientali e consente di recuperare e mettere in circolo i gruppi amminici e gli scheletri carboniosi degli amminoacidi
Anche le proteine che hanno assunto una conformazione tridimensionale
difettosa, con un ripiegamento scorretto, sono indirizzate rapidamente
verso la degradazione
Berg et al., BIOCHIMICA 6/E, Zanichellieditore S.p.A. Copyright © 2007
Ciclo dell’acido citrico
α-chetoglutaratoossalacetato
piruvato
Acetil-CoA
Glucosio 6-fosfato Ribosio 5-fosfato
Eritrosio 4-fosfato
Fosfoenolpiruvato
Istidina
3-fosfogliceratoSerina
Glicina Cisteina
FenilalaninaTriptofano
Tirosina
Valina Leucina
Alanina
Aspartato
Asparagina
Metionina Treonina
Isoleucina
Lisina
Glutammato
ArgininaProlinaGlutammina
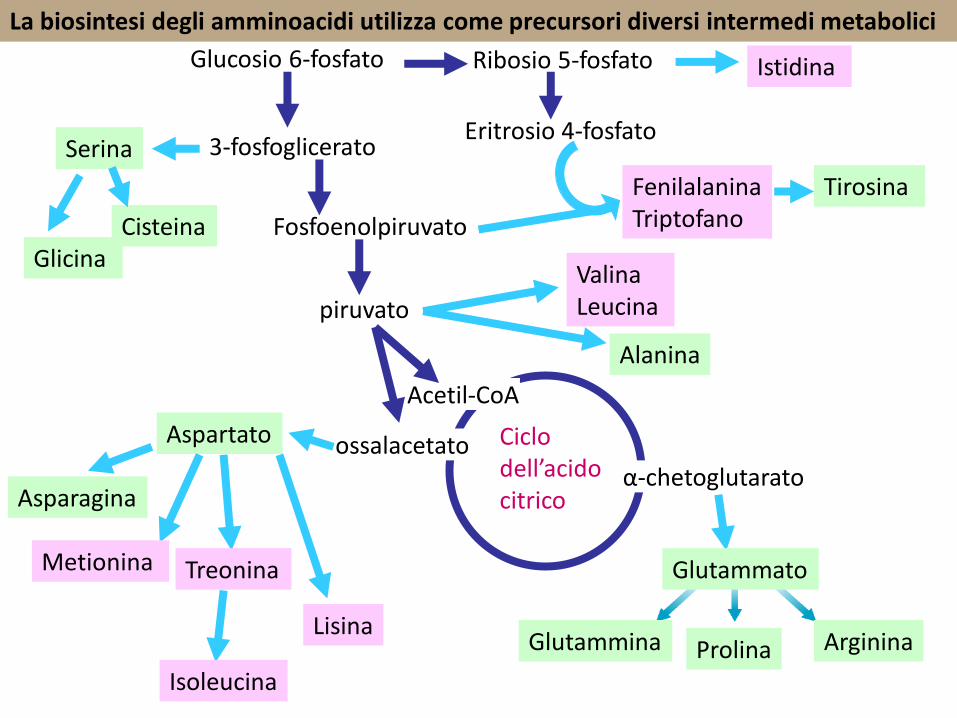
La biosintesi degli amminoacidi utilizza come precursori diversi intermedi metabolici

Esistono diverse vie biosintetiche degli aminoacidi, spesso differenti nei diversi tessuti e nei diversi organismi.
La gran parte dei processi di biosintesi utilizzano come precursori gli α-chetoacidi in reazioni di TRANSAMMINAZIONE
Consiste di 2 semireazioni:1) α-chetoacido convertito nel corrispondente α-amminoacido 2) α-amminoacido convertito nel corrispondente α-chetoacido
Scambio del gruppo –NH3+
Reazioni catalizzate dalle ammino-transferasi (o transamminasi)Coenzima che funge da scambiatore: PLP (piridossalfosfato)
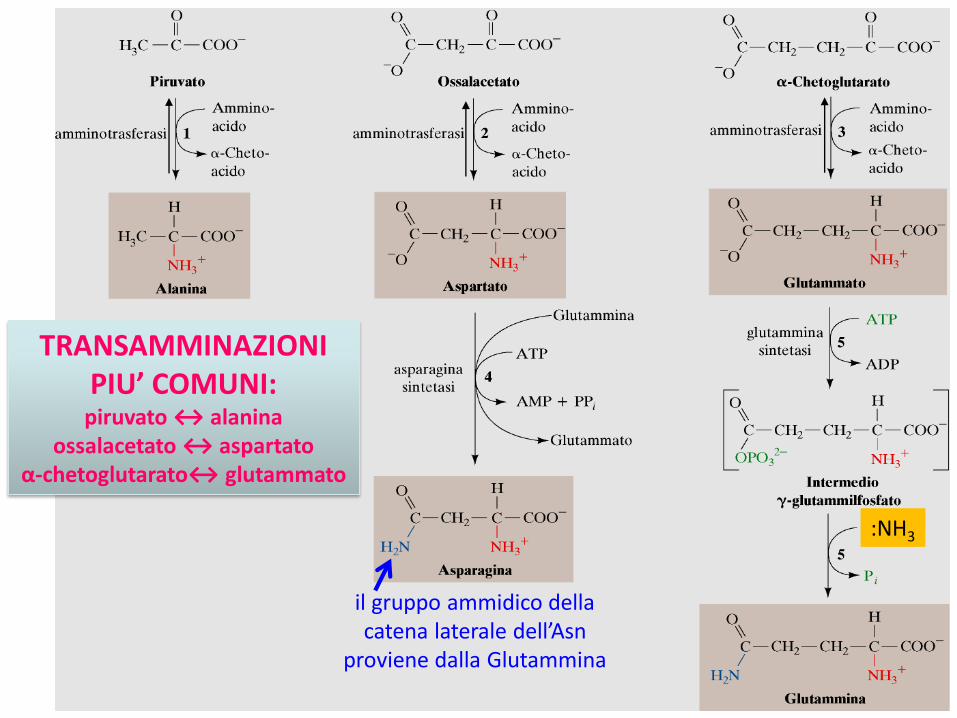
TRANSAMMINAZIONI PIU’ COMUNI:
piruvato ↔ alaninaossalacetato ↔ aspartato
α-chetoglutarato↔ glutammato
il gruppo ammidico della catena laterale dell’Asn
proviene dalla Glutammina
:NH3

α
Enzˡ
LyslNH2..
+
..
+HH-N—Lys-Enz
H
α-chetoacido
La piridossamina-fosfatotrasferirà il gruppo
amminico su un secondo α-chetoacido che diventerà un α-amminoacido
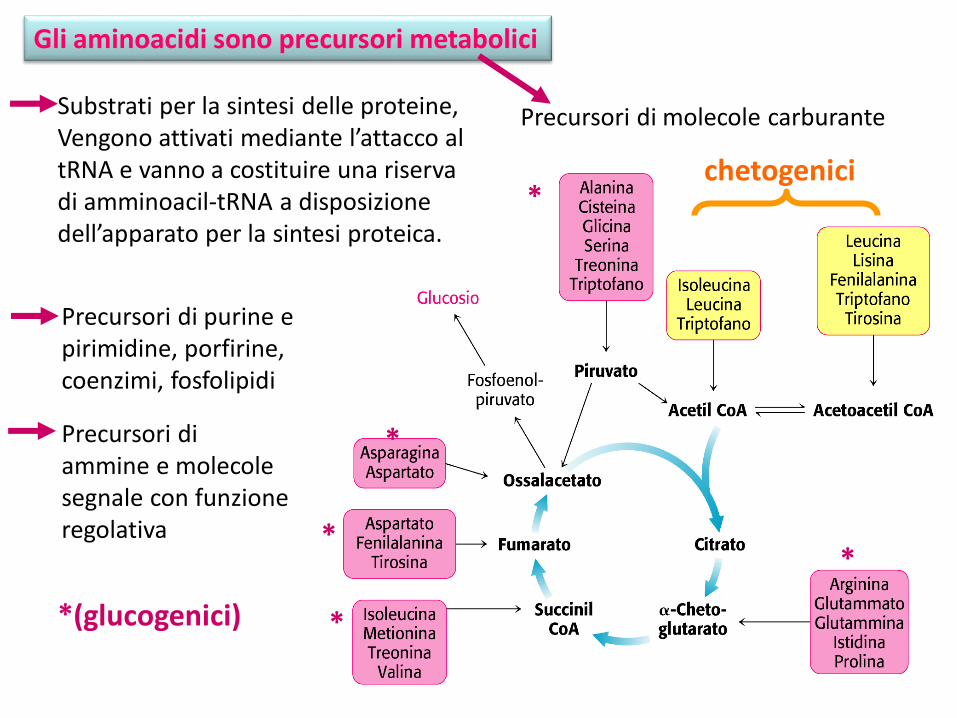
Gli aminoacidi sono precursori metabolici
Precursori di ammine e molecole segnale con funzione regolativa
Precursori di purine e pirimidine, porfirine, coenzimi, fosfolipidi
Precursori di molecole carburanteSubstrati per la sintesi delle proteine, Vengono attivati mediante l’attacco al tRNA e vanno a costituire una riserva di amminoacil-tRNA a disposizione dell’apparato per la sintesi proteica.
chetogenici*
*
*
*
*
*(glucogenici)

L’ARGININA è precursore dell’OSSIDO NITRICO >> •N=0
Radicale libero instabile e gassoso dell’azotoMolto reattivo, in soluzione acquosa esiste per pochi secondi: forma nitriti e nitrati con l’H2O. È Potenzialmente tossico ma anche importante dal punto di vista fisiologico
Nei mammiferi la OSSIDO NITRICO SINTASI (NOS) produce N=Oa partire da Arginina e O2
arginina citrullina
Ossido nitricoN=O
Altri cofattori della NOS: FAD, FMN, EME, tetraidrobiopterina BH4
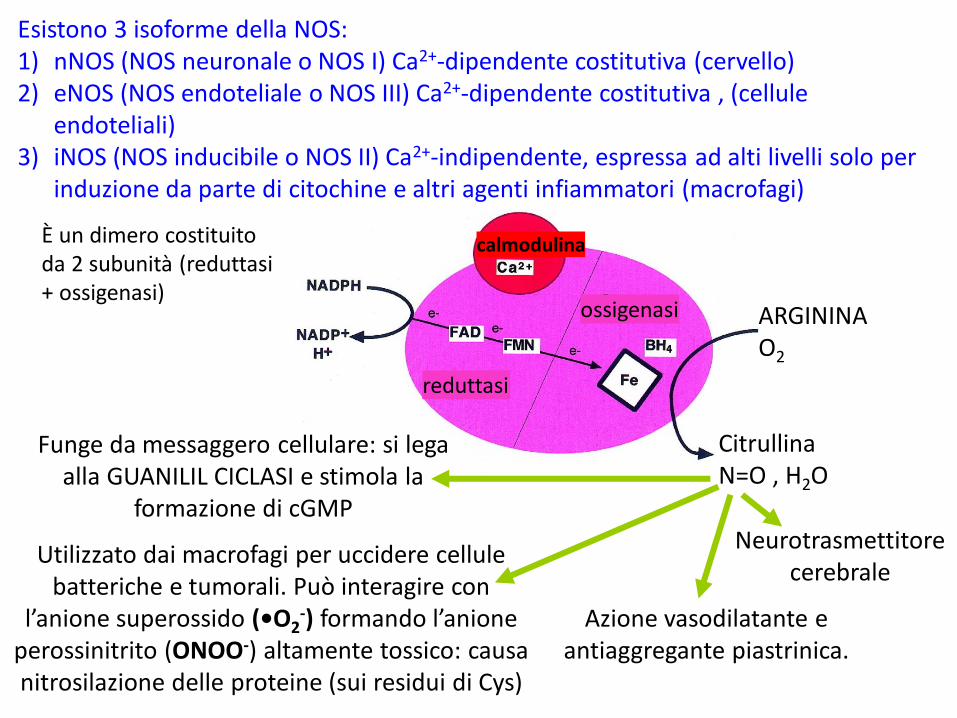
Esistono 3 isoforme della NOS:1) nNOS (NOS neuronale o NOS I) Ca2+-dipendente costitutiva (cervello)2) eNOS (NOS endoteliale o NOS III) Ca2+-dipendente costitutiva , (cellule
endoteliali)3) iNOS (NOS inducibile o NOS II) Ca2+-indipendente, espressa ad alti livelli solo per
induzione da parte di citochine e altri agenti infiammatori (macrofagi)
reduttasi
ossigenasi
calmodulina
ARGININAO2
CitrullinaN=O , H2O
Funge da messaggero cellulare: si lega alla GUANILIL CICLASI e stimola la
formazione di cGMP
Utilizzato dai macrofagi per uccidere cellule batteriche e tumorali. Può interagire con
l’anione superossido (•O2-) formando l’anione
perossinitrito (ONOO-) altamente tossico: causa nitrosilazione delle proteine (sui residui di Cys)
Azione vasodilatante e antiaggregante piastrinica.
Neurotrasmettitore cerebrale
È un dimero costituito da 2 subunità (reduttasi + ossigenasi)

Dalla decarbossilazione di alcuni amminoacidi si ottengono importanti ammine biologiche
Decarbossilazione dell’ISTIDINA ISTIDINA DECARBOSSILASI(PLP-dipendente)
CO2
Aminoacidi come precursori di ammine con funzione regolativa
CO2
Decarbossilazione del TRIPTOFANO
Idrossilazione in C-5Monoossigenasi
PLP
SEROTONINA(neurotrasmettitore)
Mediatore dell’infiammazione e delle reazioni allergiche
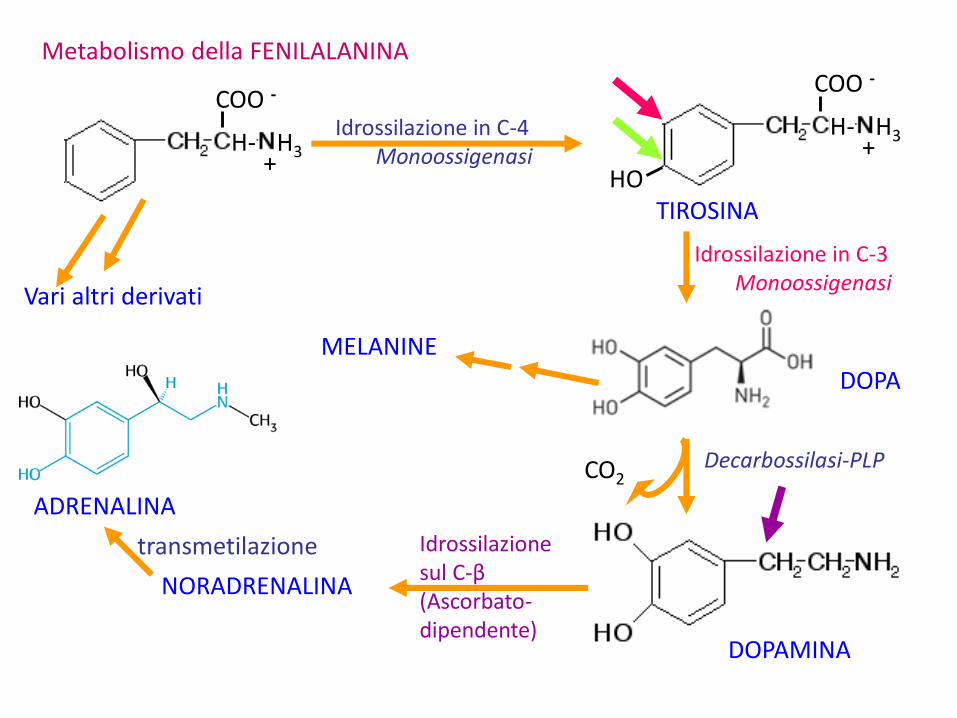
Metabolismo della FENILALANINA
CO2
transmetilazione
MELANINE
Idrossilazione in C-4Monoossigenasi
Decarbossilasi-PLP
COO -
H- H3+
COO -
H- H3+
HOTIROSINA
Idrossilazione in C-3Monoossigenasi
DOPA
DOPAMINA
Idrossilazione sul C-β(Ascorbato-dipendente)
NORADRENALINA
ADRENALINA
Vari altri derivati
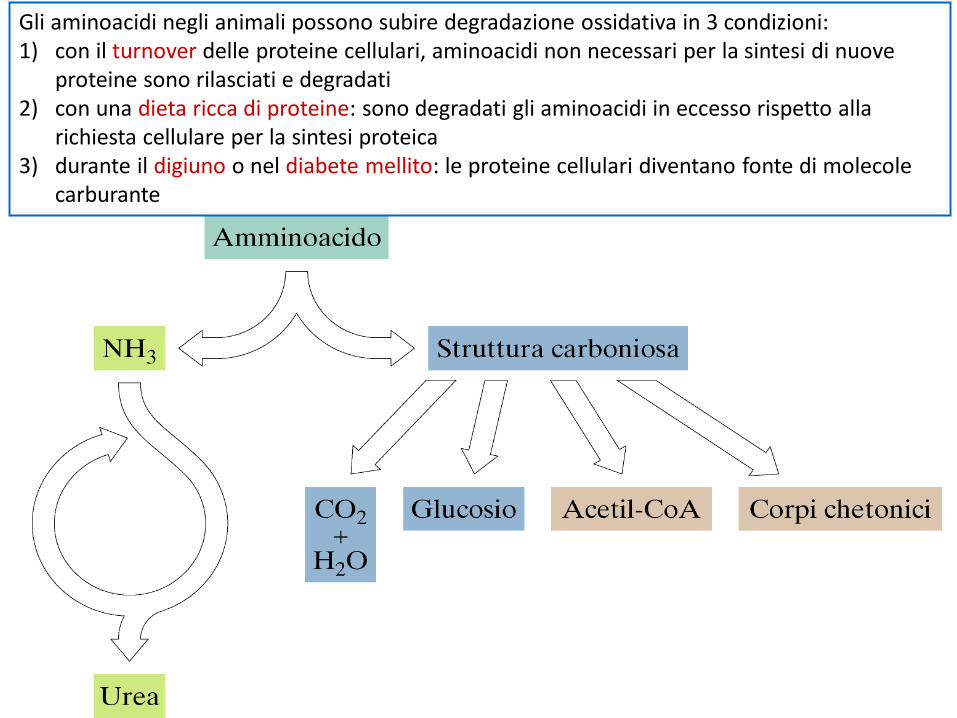
Gli aminoacidi negli animali possono subire degradazione ossidativa in 3 condizioni:1) con il turnover delle proteine cellulari, aminoacidi non necessari per la sintesi di nuove
proteine sono rilasciati e degradati2) con una dieta ricca di proteine: sono degradati gli aminoacidi in eccesso rispetto alla
richiesta cellulare per la sintesi proteica3) durante il digiuno o nel diabete mellito: le proteine cellulari diventano fonte di molecole
carburante
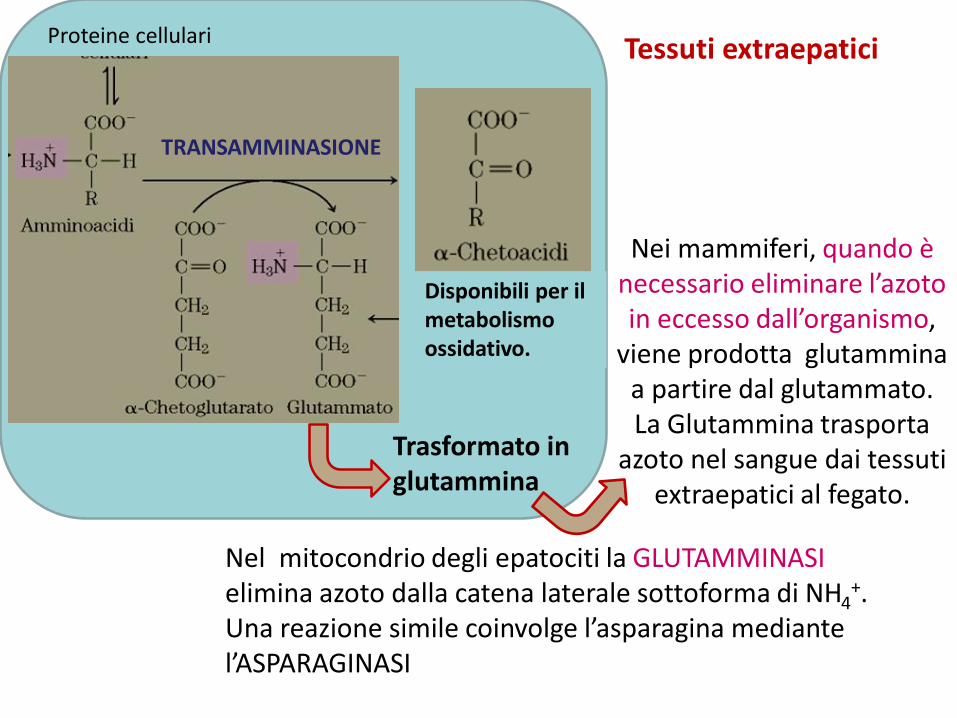
Proteine cellulari
Disponibili per il metabolismo ossidativo.
Trasformato in glutammina
Tessuti extraepatici
Nei mammiferi, quando è necessario eliminare l’azoto in eccesso dall’organismo,
viene prodotta glutammina a partire dal glutammato. La Glutammina trasporta
azoto nel sangue dai tessuti extraepatici al fegato.
Nel mitocondrio degli epatociti la GLUTAMMINASIelimina azoto dalla catena laterale sottoforma di NH4
+. Una reazione simile coinvolge l’asparagina mediante l’ASPARAGINASI
TRANSAMMINASIONE
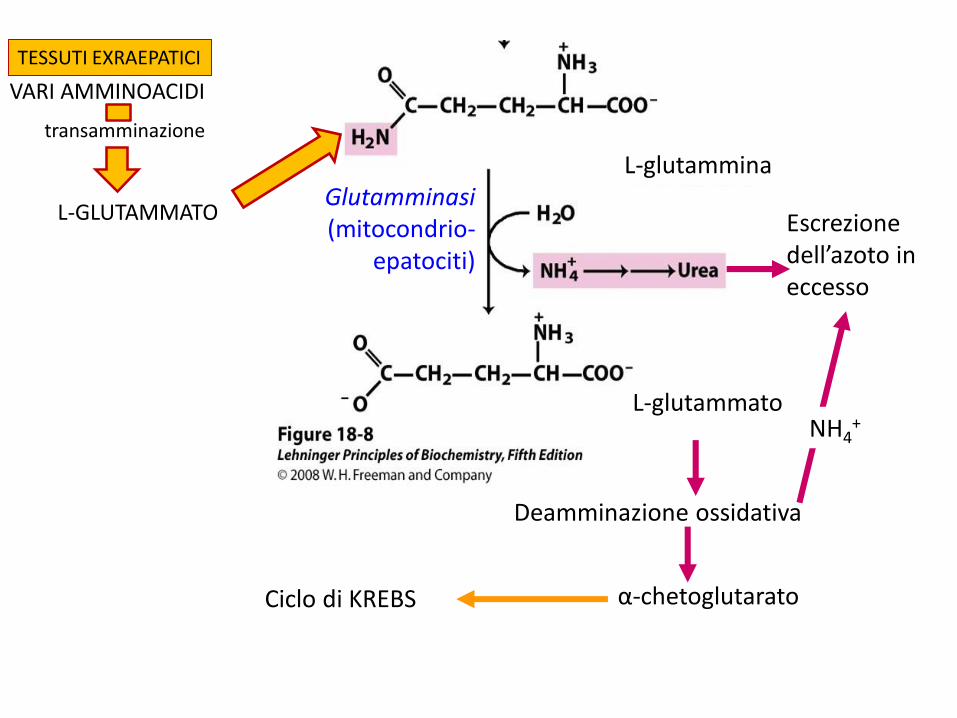
L-glutamminaGlutamminasi(mitocondrio-
epatociti)
L-glutammato
Escrezione dell’azoto in eccesso
Deamminazione ossidativa
α-chetoglutarato
NH4+
Ciclo di KREBS
L-GLUTAMMATO
VARI AMMINOACIDI
transamminazione
TESSUTI EXRAEPATICI
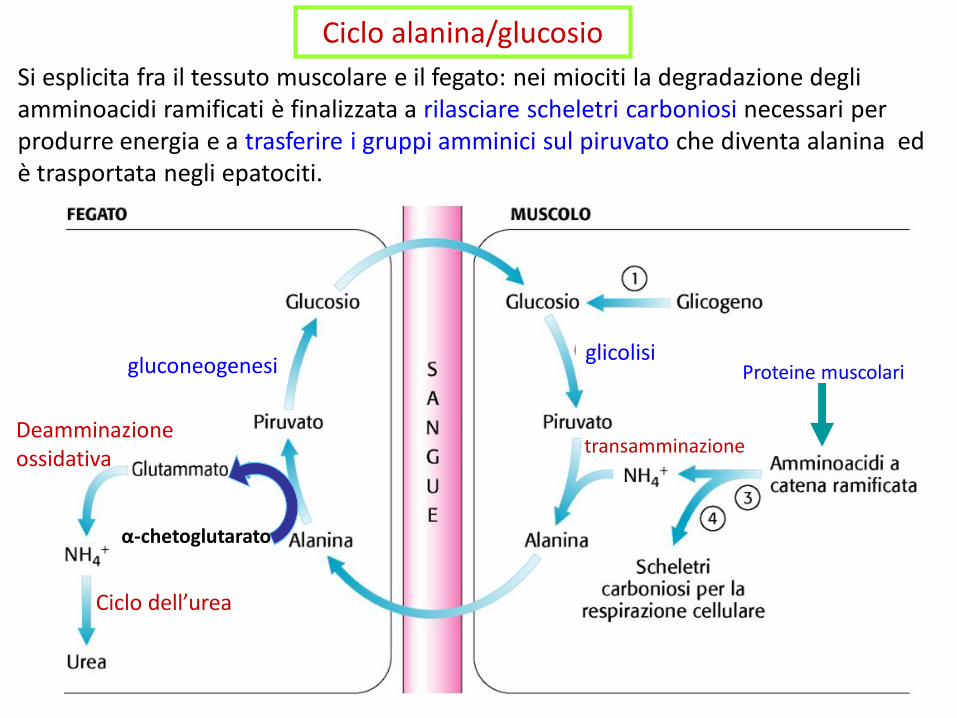
Ciclo alanina/glucosio
Si esplicita fra il tessuto muscolare e il fegato: nei miociti la degradazione degli amminoacidi ramificati è finalizzata a rilasciare scheletri carboniosi necessari per produrre energia e a trasferire i gruppi amminici sul piruvato che diventa alanina ed è trasportata negli epatociti.
transamminazione
glicolisigluconeogenesi
Deamminazione ossidativa
Ciclo dell’urea
Proteine muscolari
α-chetoglutarato

RUOLO CENTRALE DEL FEGATO NELMETABOLISMO DEGLI
AMMINOACIDI
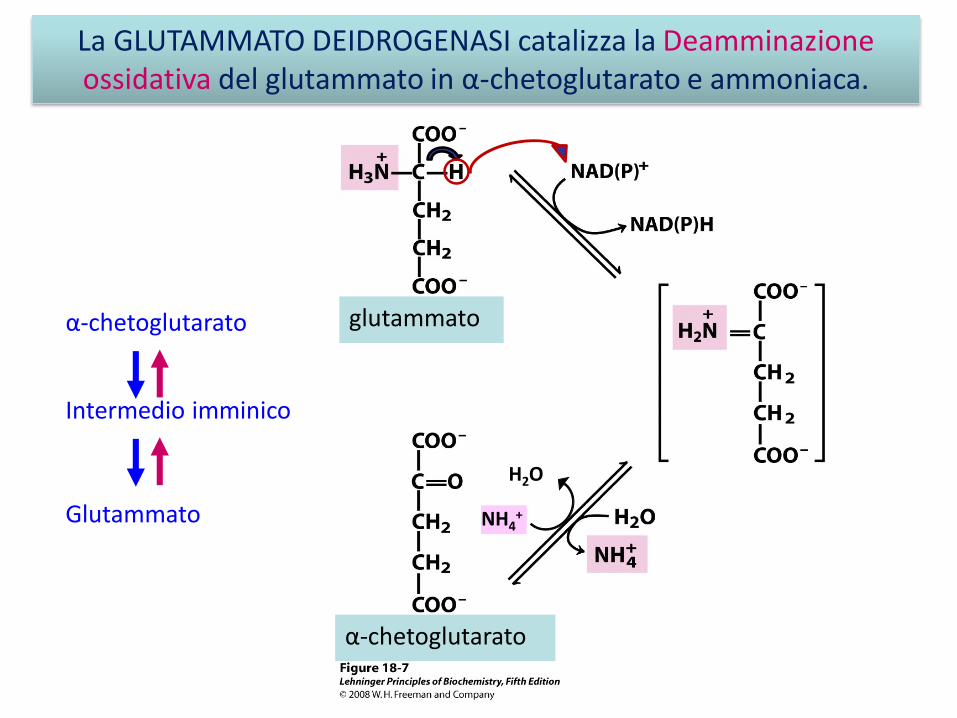
La GLUTAMMATO DEIDROGENASI catalizza la Deamminazioneossidativa del glutammato in α-chetoglutarato e ammoniaca.
α-chetoglutarato
glutammato
NH4+
H2O
α-chetoglutarato
Intermedio imminico
Glutammato
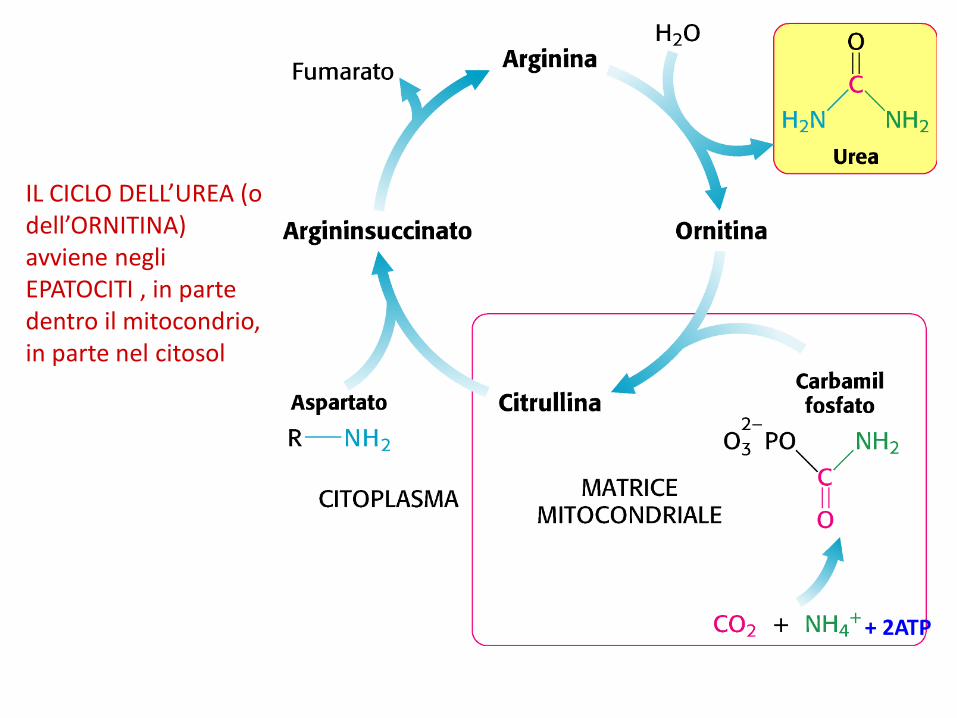
IL CICLO DELL’UREA (o dell’ORNITINA) avviene negli EPATOCITI , in parte dentro il mitocondrio, in parte nel citosol
+ 2ATP
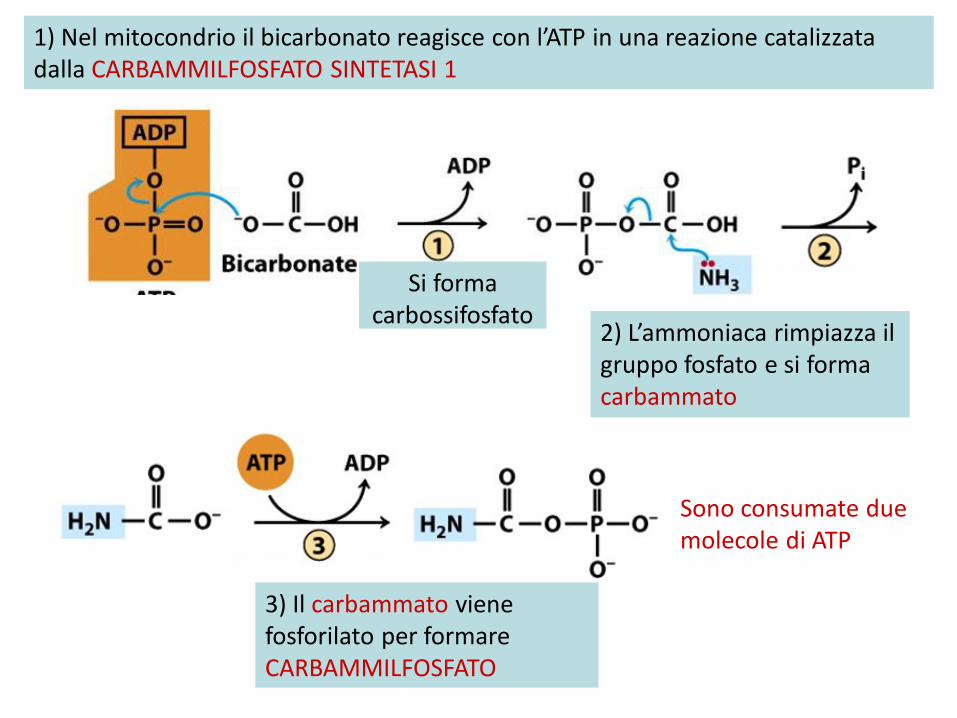
1) Nel mitocondrio il bicarbonato reagisce con l’ATP in una reazione catalizzata dalla CARBAMMILFOSFATO SINTETASI 1
Si forma carbossifosfato
2) L’ammoniaca rimpiazza il gruppo fosfato e si forma carbammato
3) Il carbammato viene fosforilato per formare CARBAMMILFOSFATO
Sono consumate due molecole di ATP
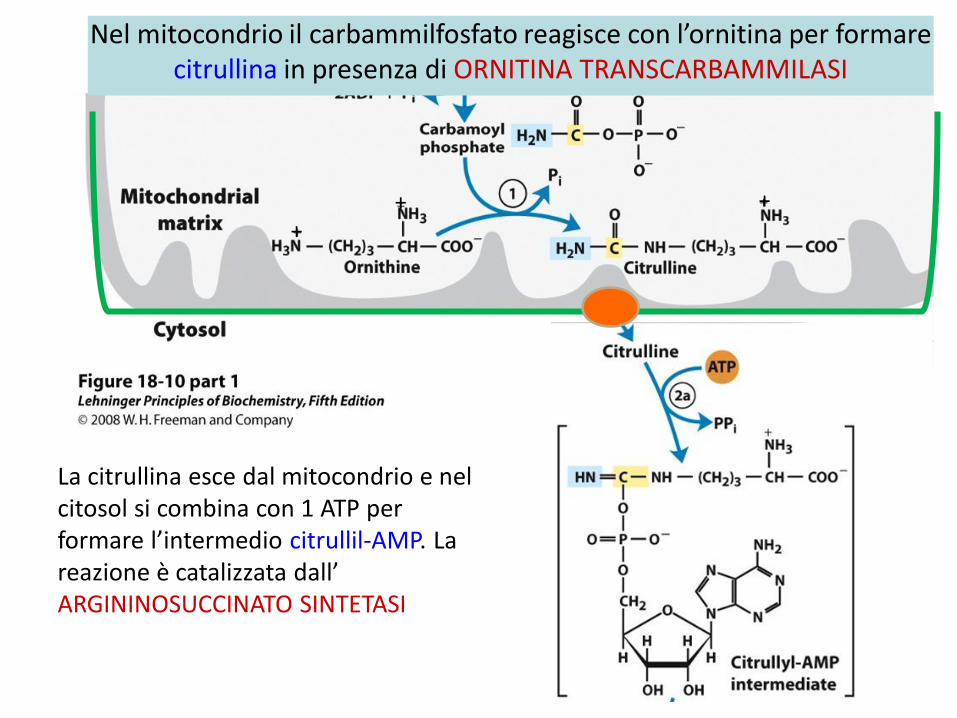
Nel mitocondrio il carbammilfosfato reagisce con l’ornitina per formare citrullina in presenza di ORNITINA TRANSCARBAMMILASI
++
+
La citrullina esce dal mitocondrio e nel citosol si combina con 1 ATP per formare l’intermedio citrullil-AMP. La reazione è catalizzata dall’ ARGININOSUCCINATO SINTETASI
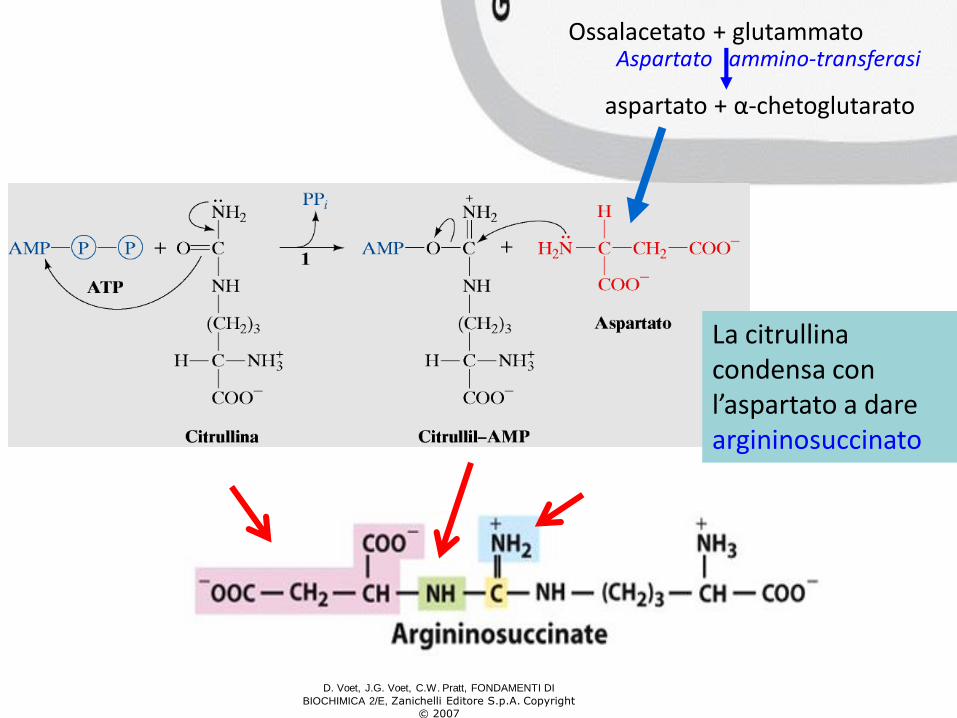
Ossalacetato + glutammatoAspartato ammino-transferasi
aspartato + α-chetoglutarato
D. Voet, J.G. Voet, C.W. Pratt, FONDAMENTI DI
BIOCHIMICA 2/E, Zanichelli Editore S.p.A. Copyright © 2007
La citrullinacondensa con l’aspartato a dare argininosuccinato
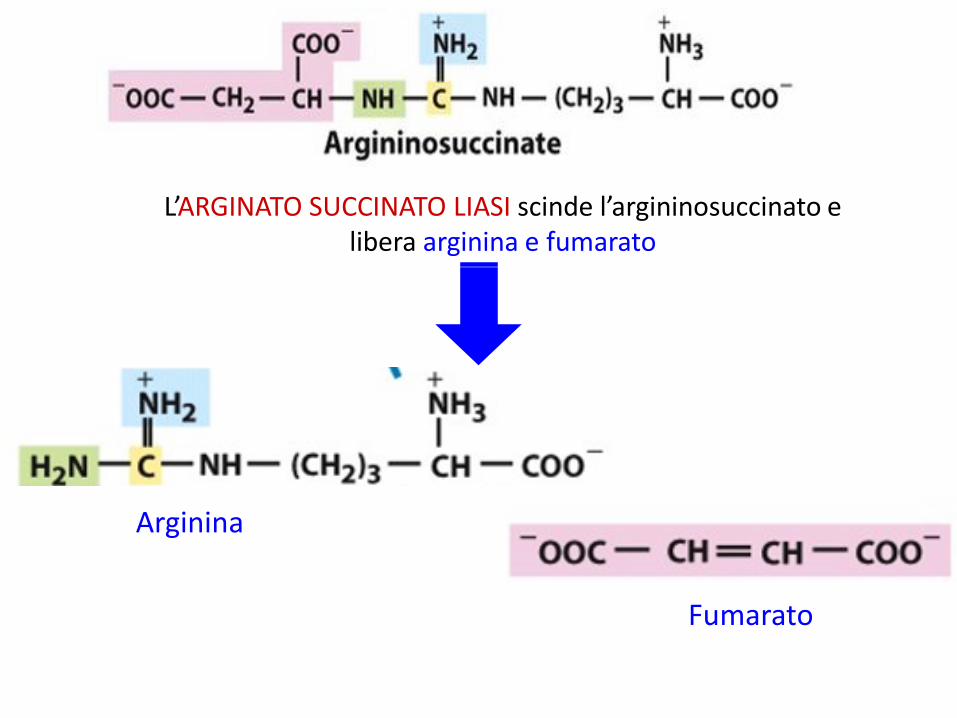
L’ARGINATO SUCCINATO LIASI scinde l’argininosuccinato e libera arginina e fumarato
Arginina
Fumarato
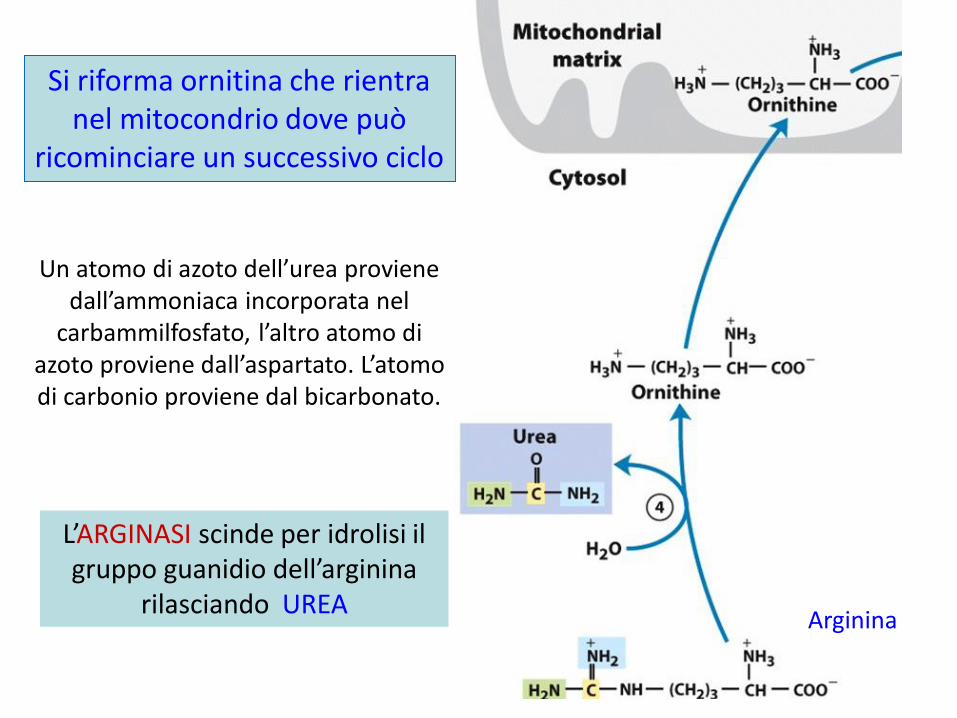
Arginina
L’ARGINASI scinde per idrolisi il gruppo guanidio dell’arginina
rilasciando UREA
Si riforma ornitina che rientra nel mitocondrio dove può
ricominciare un successivo ciclo
Un atomo di azoto dell’urea proviene dall’ammoniaca incorporata nel
carbammilfosfato, l’altro atomo di azoto proviene dall’aspartato. L’atomo di carbonio proviene dal bicarbonato.
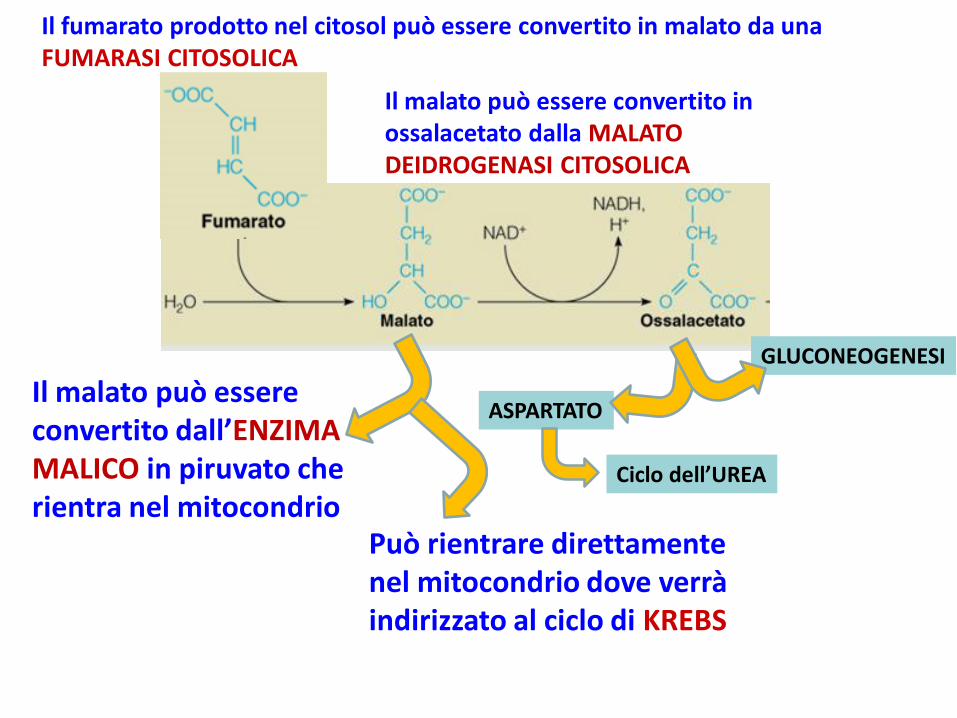
Il fumarato prodotto nel citosol può essere convertito in malato da una FUMARASI CITOSOLICA
Il malato può essere convertito dall’ENZIMA MALICO in piruvato che rientra nel mitocondrio
Il malato può essere convertito in ossalacetato dalla MALATO DEIDROGENASI CITOSOLICA
GLUCONEOGENESI
ASPARTATO
Può rientrare direttamente nel mitocondrio dove verrà indirizzato al ciclo di KREBS
Ciclo dell’UREA