
Espressione ed utilizzo della informazione genetica -...
48
1 Espressione ed utilizzo della informazione genetica Prof.ssa Flavia Frabetti Aa 2010-11 L’informazione genetica, contenuta nel DNA, ha lo scopo di: 1) mantenere lo stato vitale (strutture e funzioni) 2) realizzare l’adattamento (risposta ad un cambiamento) 3) determinare il differenziamento (specializzazione di strutture e funzioni) L’informazione genetica ereditata determina il manifestarsi di specifici caratteri inducendo la sintesi di certe proteine: le proteine sono l’anello di congiunzione tra informazione e la realizzazione/effettuazione di strutture ed attività.
Transcript of Espressione ed utilizzo della informazione genetica -...

1
Espressione ed utilizzodella
informazione genetica
Prof.ssa Flavia Frabetti
Aa 2010-11
L’informazione genetica, contenuta nel DNA, ha lo scopo di:1) mantenere lo stato vitale (strutture e funzioni)2) realizzare l’adattamento (risposta ad un cambiamento)3) determinare il differenziamento (specializzazione di strutture e funzioni)
L’informazione genetica ereditata determina il manifestarsidi specifici caratteri inducendo la sintesi di certe proteine: le proteine sono l’anello di congiunzione tra informazione e la realizzazione/effettuazione di strutture ed attività.
2
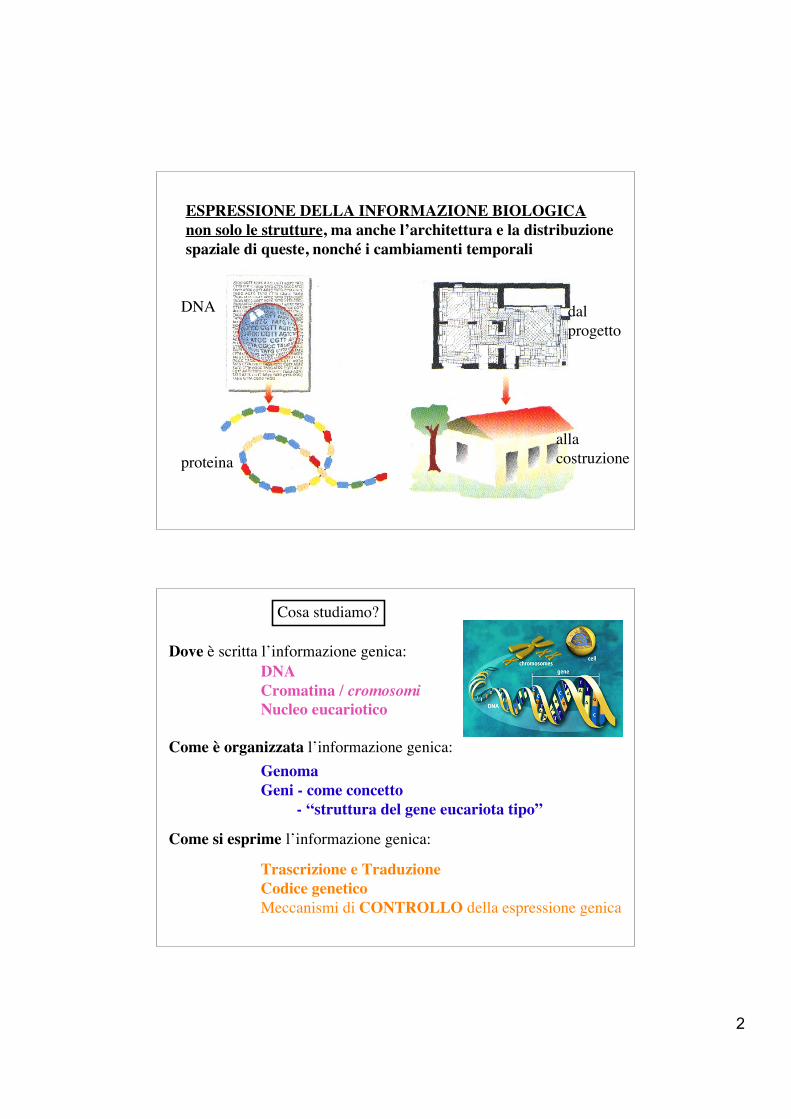
ESPRESSIONE DELLA INFORMAZIONE BIOLOGICAnon solo le strutture, ma anche l’architettura e la distribuzione spaziale di queste, nonché i cambiamenti temporali
dalprogetto
allacostruzioneproteina
DNA
Cosa studiamo?
Trascrizione e TraduzioneCodice geneticoMeccanismi di CONTROLLO della espressione genica
Dove è scritta l’informazione genica:DNACromatina / cromosomiNucleo eucariotico
Come è organizzata l’informazione genica:GenomaGeni - come concetto - “struttura del gene eucariota tipo”
Come si esprime l’informazione genica:
3
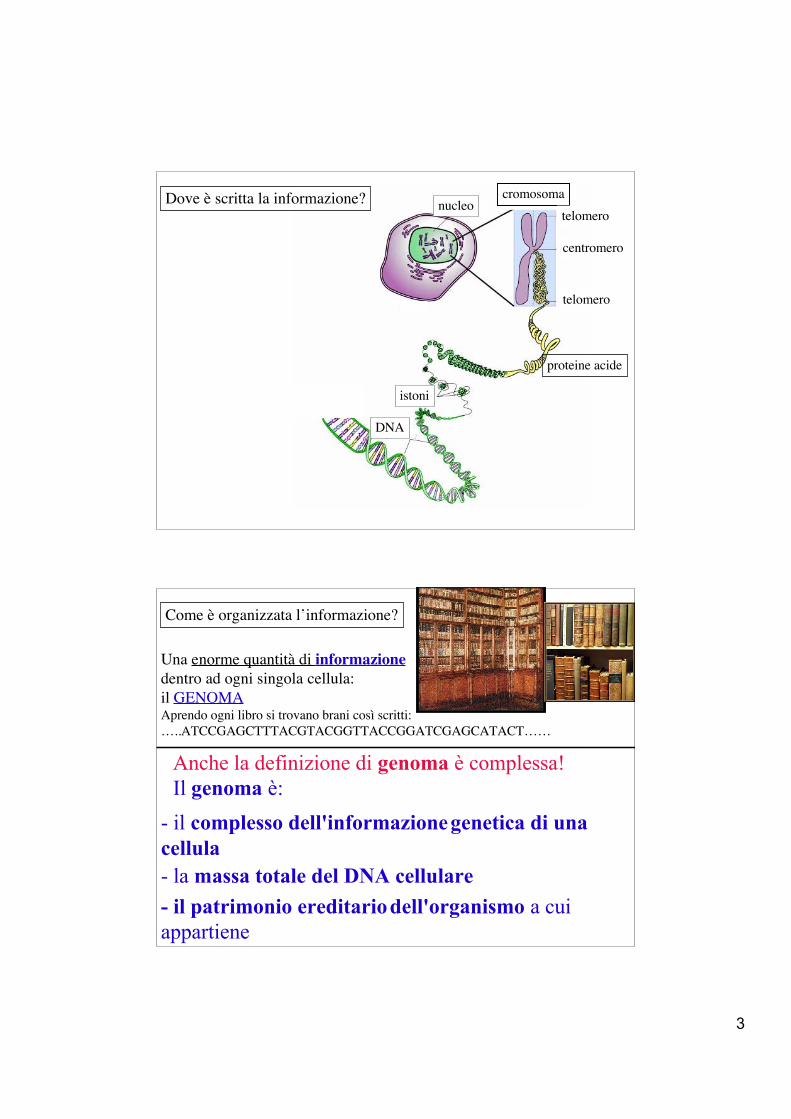
nucleo
DNA
istoni
cromosomatelomero
centromero
telomero
Dove è scritta la informazione?
proteine acide
Come è organizzata l’informazione?
Una enorme quantità di informazione dentro ad ogni singola cellula: il GENOMAAprendo ogni libro si trovano brani così scritti:…..ATCCGAGCTTTACGTACGGTTACCGGATCGAGCATACT……
- il complesso dell'informazione genetica di unacellula- la massa totale del DNA cellulare- il patrimonio ereditario dell'organismo a cuiappartiene
Anche la definizione di genoma è complessa!Il genoma è:
4
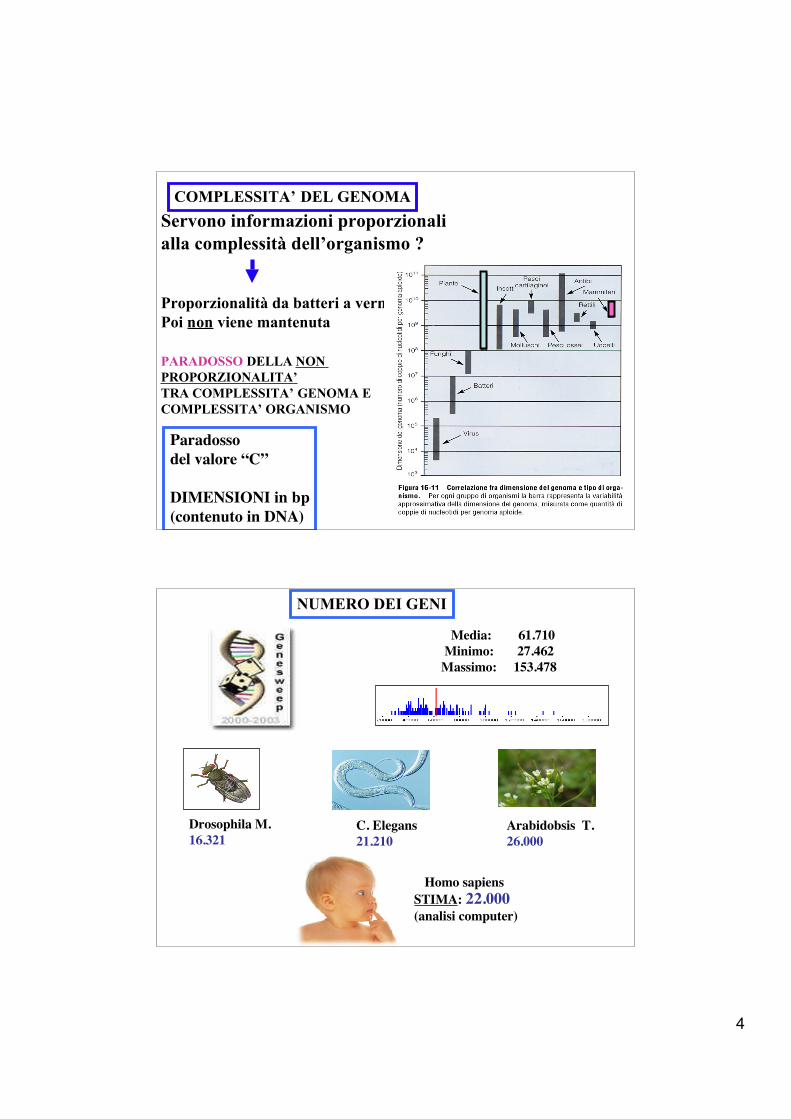
COMPLESSITA’ DEL GENOMAServono informazioni proporzionalialla complessità dell’organismo ?
Proporzionalità da batteri a vermiPoi non viene mantenuta
PARADOSSO DELLA NON PROPORZIONALITA’ TRA COMPLESSITA’ GENOMA ECOMPLESSITA’ ORGANISMO
Paradossodel valore “C”
DIMENSIONI in bp(contenuto in DNA)
NUMERO DEI GENI
Media: 61.710 Minimo: 27.462 Massimo: 153.478
Drosophila M.16.321
C. Elegans21.210
Arabidobsis T.26.000
STIMA: 22.000(analisi computer)
Homo sapiens
5
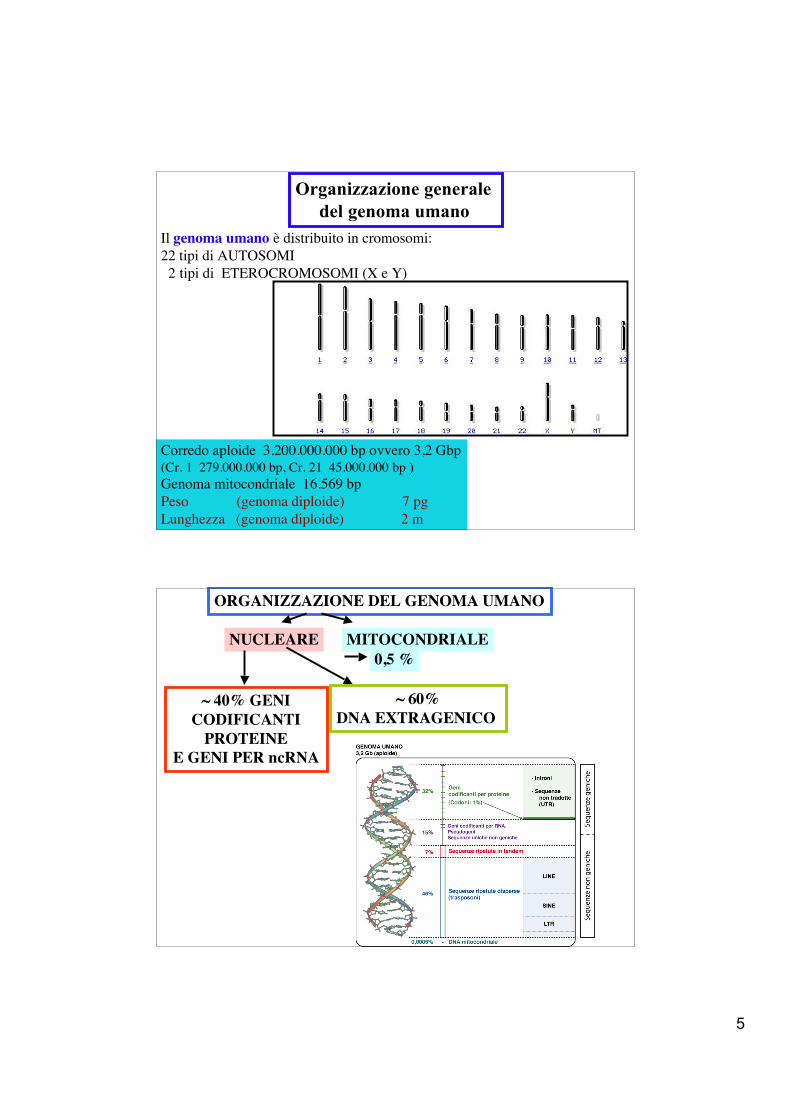
Organizzazione generale del genoma umano
Corredo aploide 3.200.000.000 bp ovvero 3,2 Gbp(Cr. 1 279.000.000 bp, Cr. 21 45.000.000 bp )Genoma mitocondriale 16.569 bpPeso (genoma diploide) 7 pgLunghezza (genoma diploide) 2 m
Il genoma umano è distribuito in cromosomi:22 tipi di AUTOSOMI 2 tipi di ETEROCROMOSOMI (X e Y)
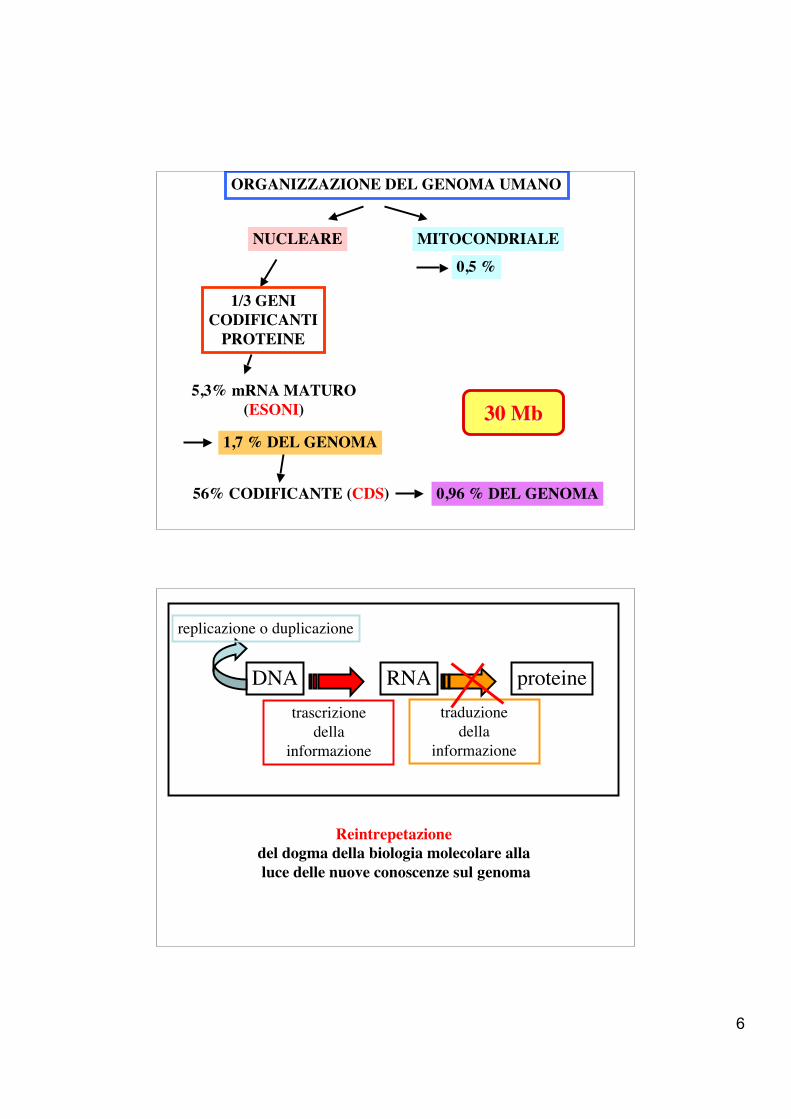
ORGANIZZAZIONE DEL GENOMA UMANO
NUCLEARE MITOCONDRIALE0,5 %
~ 40% GENICODIFICANTI
PROTEINEE GENI PER ncRNA
~ 60%DNA EXTRAGENICO
6
ORGANIZZAZIONE DEL GENOMA UMANO
NUCLEARE MITOCONDRIALE
0,5 %
1,7 % DEL GENOMA
5,3% mRNA MATURO(ESONI)
1/3 GENICODIFICANTI
PROTEINE
56% CODIFICANTE (CDS) 0,96 % DEL GENOMA
30 30 MbMb
DNA RNA proteine
replicazione o duplicazione
trascrizionedella
informazione
traduzionedella
informazione
Reintrepetazione del dogma della biologia molecolare alla luce delle nuove conoscenze sul genoma
7
Geni per RNA non tradotti o non-coding RNA (ncRNA, RNA noncodificanti proteine), nei mammiferirappresentano da 1/2 a 3/4 di tutti itrascritti
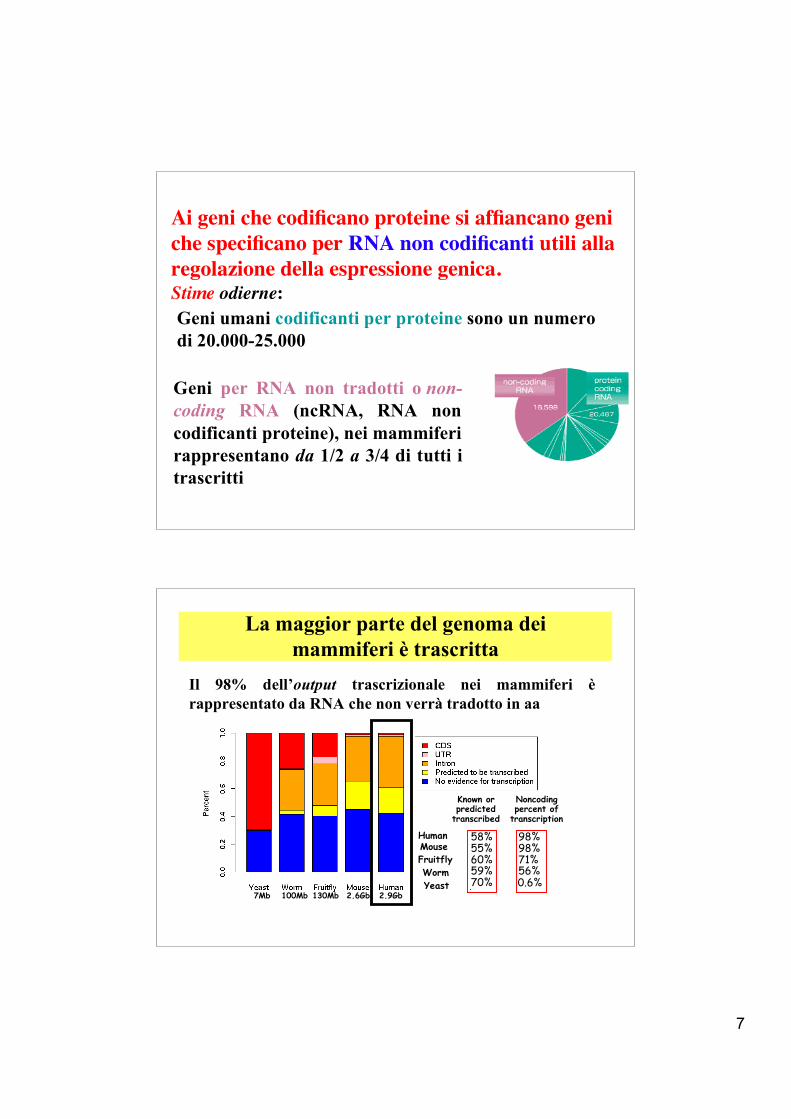
Ai geni che codificano proteine si affiancano geni che specificano per RNA non codificanti utili alla regolazione della espressione genica.Stime odierne:Geni umani codificanti per proteine sono un numerodi 20.000-25.000
La maggior parte del genoma deimammiferi è trascritta
Human 58% 98%Mouse 55% 98%Fruitfly 60% 71%Worm 59% 56%Yeast 70% 0.6%
Known orpredicted
transcribed
Noncodingpercent of
transcription
7Mb 100Mb 130Mb 2.6Gb 2.9Gb
Il 98% dell’output trascrizionale nei mammiferi èrappresentato da RNA che non verrà tradotto in aa

8
Geni:
definizione/concetto
struttura
regolazione
La complessità del genoma si riflette sulla espressione dell’informazione genica e sulla definizione stessa di gene
I geni sono leunità responsabili
delle caratteristicheereditarie
Il gene èuna regione di DNA
trascritta che contieneistruzioni per la sintesidi una proteina, di un
RNA o ignota

9
FUNZIONI DEI GENI
METABOLISMO 22%INFORMAZIONE GENETICA 25%STRUTTURA 21%SEGNALI 12%FUNZIONI TESSUTO-SPECIFICHE 20%
GENI PER RNA NON TRADOTTI(rRNA 85%, tRNA 10%, RNA non-codificanti)
Il gene è una delle tante istruzioni contenute in ogni cellula
2) Geni con limitazioni spazio-temporali nella espressione:
Espressione inducibile
Stadio del differenziamento
Stadio dello sviluppo
Stadio del ciclo cellulareTEMPO
Distribuzione intracellulare
Singole cellule (specificità dicellula es. Ab)
Specificità per tessuto,tipo cellulare
Espressione diversain diversi organi e tessuti
SPAZIO
Ci possono essere:1) Geni essenziali o geni housekeepingGeni che devono essere espressi praticamente in tutti i tipi cellularipoiché codificano prodotti utili al funzionamento generale delle cellule,per es. alla sintesi proteica o alla produzione di energia

10
Promotore
Gene strutturaleregione trascritta
Gene eucariota - la struttura nasconde complessità
I geni eucarioti hanno una struttura discontinua 1977
Ibrido molecolare DNA-mRNA

11
Il gene ha una natura discontinua:esoni tratti del gene indicati con Eintroni tratti del gene indicati con I
GENE - Struttura del gene eucariota tipo
E 1 E 2 E 3I 1 I 2
+1
Tratto di DNA che viene trascritto o TRASCRITTO PRIMARIO
DNA
+1
E 1 E 2 E 3
TRASCRITTOPRIMARIO o pre-mRNA
Esone 1 Esone 2 Esone 3
AUG UGA
Proteina NH2 COOH
I 1 I 2
ESONI (in giallo), intervallate da lunghi INTRONI (in grigio)
mRNA AUG UGA

12
Dimensione media geni 57.000 bp (57 kb)istoni 100-400 bp (0,1-0,4 kb) distrofina 2.220.000 bp (2,22 Mb)ESONINumero medio 11(minimo)-(massimo) 1 - 363Dimensione media 280 bpINTRONIDimensione media ca. 6.000 bp
mRNA maturo (in media)Dimensione 3.000 basi Sequenza codificante 1.600 basi (56%)Proteina 540 AA
Nella realtà gli INTRONI sono molto più lunghidegli esoni
TTAGCACTACCGTATTTGCGCATTACCAGATTAGAGAATTAGCACTACCGTATTTGCGCATTACCAGATTAGAGAAATGCTAGTCGATCTATCGATCGGCTATTCGCAAAGCTGATGCTAGTCGATCTATCGATCGGCTATTCGCAAAGCTGCGCGACTGCGATGCGCTAGCATGCGATTCGCGATCGCCCGCGACTGCGATGCGCTAGCATGCGATTCGCGATCGCCGAGCGCTCGCGAGCGCGCTAGCGGAATACTATATAGCGAGCGCTCGCGAGCGCGCTAGCGGAATACTATATAGCGCGGATCAGTCTAGATCTATGAGATCGATAGCGATCTAGCGGATCAGTCTAGATCTATGAGATCGATAGCGATCTAGAGATAGGATCGAGATCGAGGCGAGATCATATGAGCGGAGATAGGATCGAGATCGAGGCGAGATCATATGAGCGCGGCTATTTAGGCTTAGAGGATTCGGAGATTCGGAGCTCGGCTATTTAGGCTTAGAGGATTCGGAGATTCGGAGCTTAGGATTACAGAGAGCTTCTTAGGCGCTCCCGGTATCGTAGGATTACAGAGAGCTTCTTAGGCGCTCCCGGTATCGCTCCCATCCCATATTAAAATCTATCGATCGAGCTCTCCACTCCCATCCCATATTAAAATCTATCGATCGAGCTCTCCAATGCGATCGATAGGACTAGTAGCTAGCTAGCTGAGCAATGCGATCGATAGGACTAGTAGCTAGCTAGCTGAGCATGATAGGCTCGATGAGCATGAGATGCATGTACGACTGTGATAGGCTCGATGAGCATGAGATGCATGTACGACTGCATAGGCATGACTGATCGACTGCATCATGACGCATGACCATAGGCATGACTGATCGACTGCATCATGACGCATGACTGCATGCATGACTGCATATGACGGACTCGCATTAGCACTGCATGCATGACTGCATATGACGGACTCGCATTAGCACTACCGTATTTGCGCATTACCAGATTAGAGAAATGCTAGTACCGTATTTGCGCATTACCAGATTAGAGAAATGCTAGTCGATCTATCGATCGGCTATTCGCAAAGCTGCGCGACTTCGATCTATCGATCGGCTATTCGCAAAGCTGCGCGACTGCGATGCGCTAGCATGCGATTCGCGATCGCCGAGCGCGCGATGCGCTAGCATGCGATTCGCGATCGCCGAGCGCTCGCGAGCGCGCTAGCGGAATACTATATAGCGCGGATTCGCGAGCGCGCTAGCGGAATACTATATAGCGCGGATCAGTCTAGATCTATGAGATCGATAGCGATCTAGAGATACAGTCTAGATCTATGAGATCGATAGCGATCTAGAGATAGGATCGAGATCGAGGCGAGATCATATGAGCGCGGCTAGGATCGAGATCGAGGCGAGATCATATGAGCGCGGCTATTTAGGCTTAGAGGATTCGGAGATTCGGAGCTTAGGATTTTAGGCTTAGAGGATTCGGAGATTCGGAGCTTAGGATTACAGAGAGCTTCTTAGGCGCTCCCGGTATCGCTCCCATACAGAGAGCTTCTTAGGCGCTCCCGGTATCGCTCCCATCCCATATTAAAATCTATCGATCGAGCTCTCCAATGCGTCCCATATTAAAATCTATCGATCGAGCTCTCCAATGCGATCGATAGGACTAGTAGCTAGCTAGCTGAGCATGATAATCGATAGGACTAGTAGCTAGCTAGCTGAGCATGATAGGCTCGATGAGCATGAGATGCATGTACGACTGCATAGGGCTCGATGAGCATGAGATGCATGTACGACTGCATAGGCATGACTGATCGACTGCATCATGACGCATGACTGCATGCATGACTGATCGACTGCATCATGACGCATGACTGCATGCATGACTGCATATGACGGACTCGCATTAGCACTACCGGCATGACTGCATATGACGGACTCGCATTAGCACTACCGTATTTGCGCATTACCAGATTAGAGAAATGCTAGTCGATTATTTGCGCATTACCAGATTAGAGAAATGCTAGTCGATCTATCGATCGGCTATTCGCAAAGCTGCGCGACTGCGATCTATCGATCGGCTATTCGCAAAGCTGCGCGACTGCGATGCGCTAGCATGCGATTCGCGATCGCCGAGCGCTCGCGGCGCTAGCATGCGATTCGCGATCGCCGAGCGCTCGCGAGCGCGCTAGCGGAATACTATATAGCGCGGATCAGTCAGCGCGCTAGCGGAATACTATATAGCGCGGATCAGTCTAGATCTATGAGATCGATAGCGATCTAGAGATAGGATCTAGATCTATGAGATCGATAGCGATCTAGAGATAGGATCGAGATCGAGGCGAGATCATATGAGCGCGGCTATTTAGGAGATCGAGGCGAGATCATATGAGCGCGGCTATTTAGGCTTAGAGGATTCGGAGATTCGGAGCTTAGGATTACAGCTTAGAGGATTCGGAGATTCGGAGCTTAGGATTACAGAGAGCTTCTTAGGCGCTCCCGGTATCGCTCCCATCCCGAGAGCTTCTTAGGCGCTCCCGGTATCGCTCCCATCCCATATTAAAATCTATCGATCGAGCTCTCCAATGCGATCGATATTAAAATCTATCGATCGAGCTCTCCAATGCGATCGATAGGACTAGTAGCTAGCTAGCTGAGCATGATAGGCTATAGGACTAGTAGCTAGCTAGCTGAGCATGATAGGCTCGATGAGCATGAGATGCATGTACGACTGCATAGGCATCGATGAGCATGAGATGCATGTACGACTGCATAGGCATGACTGATCGACTGCATCATGACGCATGACTGCATGCATGACTGATCGACTGCATCATGACGCATGACTGCATGCATGACTGCATATGACGGACTCGCAGACTGCATATGACGGACTCGCA
TTAGCACTACCGTATTTGCGCATTACCAGATTAGAGAATTAGCACTACCGTATTTGCGCATTACCAGATTAGAGAAATGCTAGTCGATCTATCGATCGGCTATTCGCAAAGCTGATGCTAGTCGATCTATCGATCGGCTATTCGCAAAGCTGCGCGACTGCGATGCGCTAGCATGCGATTCGCGATCGCCCGCGACTGCGATGCGCTAGCATGCGATTCGCGATCGCCGAGCGCTCGCGAGCGCGCTAGCGGAATACTATATAGCGAGCGCTCGCGAGCGCGCTAGCGGAATACTATATAGCGCGGATCAGTCTAGATCTATGAGATCGATAGCGATCTAGCGGATCAGTCTAGATCTATGAGATCGATAGCGATCTAGAGATAGGATCGAGATCGAGGCGAGATCATATGAGCGGAGATAGGATCGAGATCGAGGCGAGATCATATGAGCGCGGCTATTTAGGCTTAGAGGATTCGGAGATTCGGAGCTCGGCTATTTAGGCTTAGAGGATTCGGAGATTCGGAGCTTAGGATTACAGAGAGCTTCTTAGGCGCTCCCGGTATCGTAGGATTACAGAGAGCTTCTTAGGCGCTCCCGGTATCGCTCCCATCCCATATTAAAATCTATCGATCGAGCTCTCCACTCCCATCCCATATTAAAATCTATCGATCGAGCTCTCCAATGCGATCGATAGGACTAGTAGCTAGCTAGCTGAGCAATGCGATCGATAGGACTAGTAGCTAGCTAGCTGAGCATGATAGGCTCGATGAGCATGAGATGCATGTACGACTGTGATAGGCTCGATGAGCATGAGATGCATGTACGACTGCATAGGCATGACTGATCGACTGCATCATGACGCATGACCATAGGCATGACTGATCGACTGCATCATGACGCATGACTGCATGCATGACTGCATATGACGGACTCGCATTAGCACTGCATGCATGACTGCATATGACGGACTCGCATTAGCACTACCGTATTTGCGCATTACCAGATTAGAGAAATGCTAGTACCGTATTTGCGCATTACCAGATTAGAGAAATGCTAGTCGATCTATCGATCGGCTATTCGCAAAGCTGCGCGACTTCGATCTATCGATCGGCTATTCGCAAAGCTGCGCGACTGCGATGCGCTAGCATGCGATTCGCGATCGCCGAGCGCGCGATGCGCTAGCATGCGATTCGCGATCGCCGAGCGCTCGCGAGCGCGCTAGCGGAATACTATATAGCGCGGATTCGCGAGCGCGCTAGCGGAATACTATATAGCGCGGATCAGTCTAGATCTATGAGATCGATAGCGATCTAGAGATACAGTCTAGATCTATGAGATCGATAGCGATCTAGAGATAGGATCGAGATCGAGGCGAGATCATATGAGCGCGGCTAGGATCGAGATCGAGGCGAGATCATATGAGCGCGGCTATTTAGGCTTAGAGGATTCGGAGATTCGGAGCTTAGGATTTTAGGCTTAGAGGATTCGGAGATTCGGAGCTTAGGATTACAGAGAGCTTCTTAGGCGCTCCCGGTATCGCTCCCATACAGAGAGCTTCTTAGGCGCTCCCGGTATCGCTCCCATCCCATATTAAAATCTATCGATCGAGCTCTCCAATGCGTCCCATATTAAAATCTATCGATCGAGCTCTCCAATGCGATCGATAGGACTAGTAGCTAGCTAGCTGAGCATGATAATCGATAGGACTAGTAGCTAGCTAGCTGAGCATGATAGGCTCGATGAGCATGAGATGCATGTACGACTGCATAGGGCTCGATGAGCATGAGATGCATGTACGACTGCATAGGCATGACTGATCGACTGCATCATGACGCATGACTGCATGCATGACTGATCGACTGCATCATGACGCATGACTGCATGCATGACTGCATATGACGGACTCGCATTAGCACTACCGGCATGACTGCATATGACGGACTCGCATTAGCACTACCGTATTTGCGCATTACCAGATTAGAGAAATGCTAGTCGATTATTTGCGCATTACCAGATTAGAGAAATGCTAGTCGATCTATCGATCGGCTATTCGCAAAGCTGCGCGACTGCGATCTATCGATCGGCTATTCGCAAAGCTGCGCGACTGCGATGCGCTAGCATGCGATTCGCGATCGCCGAGCGCTCGCGGCGCTAGCATGCGATTCGCGATCGCCGAGCGCTCGCGAGCGCGCTAGCGGAATACTATATAGCGCGGATCAGTCAGCGCGCTAGCGGAATACTATATAGCGCGGATCAGTCTAGATCTATGAGATCGATAGCGATCTAGAGATAGGATTAGATCTATGAGATCGATAGCGATCTAGAGATAGGATCGAGATCGAGGCGAGATCATATGAGCGCGGCTATTTACGAGATCGAGGCGAGATCATATGAGCGCGGCTATTTAGGCTTAGAGGATTCGGAGATTCGGAGCTTAGGATTACGGCTTAGAGGATTCGGAGATTCGGAGCTTAGGATTACAGAGAGCTTCTTAGGCGCTCCCGGTATCGCTCCCATCCAGAGAGCTTCTTAGGCGCTCCCGGTATCGCTCCCATCCCATATTAAAATCTATCGATCGAGCTCTCCAATGCGATCCATATTAAAATCTATCGATCGAGCTCTCCAATGCGATCGATAGGACTAGTAGCTAGCTAGCTGAGCATGATAGGCGATAGGACTAGTAGCTAGCTAGCTGAGCATGATAGGCTCGATGAGCATGAGATGCATGTACGACTGCATAGGCATCGATGAGCATGAGATGCATGTACGACTGCATAGGCATGACTGATCGACTGCATCATGACGCATGACTGCATGCATGACTGATCGACTGCATCATGACGCATGACTGCATGCATGACTGCATATGACGGACTCGCATGACTGCATATGACGGACTCGCA
Nel “libro”
DNA,le pagine
sono scritte incodice/i
GENE

13
TTAGCACTACCGTATTTGCGCATTACCAGATTAGAGAATTAGCACTACCGTATTTGCGCATTACCAGATTAGAGAAATGCTAGTCGATCTATCGATCGGCTATTCGCAAAGCTGATGCTAGTCGATCTATCGATCGGCTATTCGCAAAGCTGCGCGACTGCGATGCGCTAGCATGCGATTCGCGATCGCCCGCGACTGCGATGCGCTAGCATGCGATTCGCGATCGCCGAGCGCTCGCGAGCGCGCTAGCGGAATACTATATAGCGAGCGCTCGCGAGCGCGCTAGCGGAATACTATATAGCGCGGATCAGTCTAGATCTATGAGATCGATAGCGATCTAGCGGATCAGTCTAGATCTATGAGATCGATAGCGATCTAGAGATAGGATCGAGATCGAGGCGAGATCATATGAGCGGAGATAGGATCGAGATCGAGGCGAGATCATATGAGCGCGGCTATTTAGGCTTAGAGGATTCGGAGATTCGGAGCTCGGCTATTTAGGCTTAGAGGATTCGGAGATTCGGAGCTTAGGATTACAGAGAGCTTCTTAGGCGCTCCCGGTATCGTAGGATTACAGAGAGCTTCTTAGGCGCTCCCGGTATCGCTCCCATCCCATATTAAAATCTATCGATCGAGCTCTCCACTCCCATCCCATATTAAAATCTATCGATCGAGCTCTCCAATGCGATCGATAGGACTAGTAGCTAGCTAGCTGAGCAATGCGATCGATAGGACTAGTAGCTAGCTAGCTGAGCATGATAGGCTCGATGAGCATGAGATGCATGTACGACTGTGATAGGCTCGATGAGCATGAGATGCATGTACGACTGCATAGGCATGACTGATCGACTGCATCATGACGCATGACCATAGGCATGACTGATCGACTGCATCATGACGCATGACTGCATGCATGACTGCATATGACGGACTCGCATTAGCACTGCATGCATGACTGCATATGACGGACTCGCATTAGCACTACCGTATTTGCGCATTACCAGATTAGAGAAATGCTAGTACCGTATTTGCGCATTACCAGATTAGAGAAATGCTAGTCGATCTATCGATCGGCTATTCGCAAAGCTGCGCGACTTCGATCTATCGATCGGCTATTCGCAAAGCTGCGCGACTGCGATGCGCTAGCATGCGATTCGCGATCGCCGAGCGCGCGATGCGCTAGCATGCGATTCGCGATCGCCGAGCGCTCGCGAGCGCGCTAGCGGAATACTATATAGCGCGGATTCGCGAGCGCGCTAGCGGAATACTATATAGCGCGGATCAGTCTAGATCTATGAGATCGATAGCGATCTAGAGATACAGTCTAGATCTATGAGATCGATAGCGATCTAGAGATAGGATCGAGATCGAGGCGAGATCATATGAGCGCGGCTAGGATCGAGATCGAGGCGAGATCATATGAGCGCGGCTATTTAGGCTTAGAGGATTCGGAGATTCGGAGCTTAGGATTTTAGGCTTAGAGGATTCGGAGATTCGGAGCTTAGGATTACAGAGAGCTTCTTAGGCGCTCCCGGTATCGCTCCCATACAGAGAGCTTCTTAGGCGCTCCCGGTATCGCTCCCATCCCATATTAAAATCTATCGATCGAGCTCTCCAATGCGTCCCATATTAAAATCTATCGATCGAGCTCTCCAATGCGATCGATAGGACTAGTAGCTAGCTAGCTGAGCATGATAATCGATAGGACTAGTAGCTAGCTAGCTGAGCATGATAGGCTCGATGAGCATGAGATGCATGTACGACTGCATAGGGCTCGATGAGCATGAGATGCATGTACGACTGCATAGGCATGACTGATCGACTGCATCATGACGCATGACTGCATGCATGACTGATCGACTGCATCATGACGCATGACTGCATGCATGACTGCATATGACGGACTCGCATTAGCACTACCGGCATGACTGCATATGACGGACTCGCATTAGCACTACCGTATTTGCGCATTACCAGATTAGAGAAATGCTAGTCGATTATTTGCGCATTACCAGATTAGAGAAATGCTAGTCGATCTATCGATCGGCTATTCGCAAAGCTGCGCGACTGCGATCTATCGATCGGCTATTCGCAAAGCTGCGCGACTGCGATGCGCTAGCATGCGATTCGCGATCGCCGAGCGCTCGCGGCGCTAGCATGCGATTCGCGATCGCCGAGCGCTCGCGAGCGCGCTAGCGGAATACTATATAGCGCGGATCAGTCAGCGCGCTAGCGGAATACTATATAGCGCGGATCAGTCTAGATCTATGAGATCGATAGCGATCTAGAGATAGGATCTAGATCTATGAGATCGATAGCGATCTAGAGATAGGATCGAGATCGAGGCGAGATCATATGAGCGCGGCTATTTAGGAGATCGAGGCGAGATCATATGAGCGCGGCTATTTAGGCTTAGAGGATTCGGAGATTCGGAGCTTAGGATTACAGCTTAGAGGATTCGGAGATTCGGAGCTTAGGATTACAGAGAGCTTCTTAGGCGCTCCCGGTATCGCTCCCATCCCGAGAGCTTCTTAGGCGCTCCCGGTATCGCTCCCATCCCATATTAAAATCTATCGATCGAGCTCTCCAATGCGATCGATATTAAAATCTATCGATCGAGCTCTCCAATGCGATCGATAGGACTAGTAGCTAGCTAGCTGAGCATGATAGGCTATAGGACTAGTAGCTAGCTAGCTGAGCATGATAGGCTCGATGAGCATGAGATGCATGTACGACTGCATAGGCATCGATGAGCATGAGATGCATGTACGACTGCATAGGCATGACTGATCGACTGCATCATGACGCATGACTGCATGCATGACTGATCGACTGCATCATGACGCATGACTGCATGCATGACTGCATATGACGGACTCGCAGACTGCATATGACGGACTCGCA
TTAGCACTACCGTATTTGCGCATTACCAGATTAGAGAATTAGCACTACCGTATTTGCGCATTACCAGATTAGAGAAATGCTAGTCGATCTATCGATCGGCTATTCGCAAAGCTGATGCTAGTCGATCTATCGATCGGCTATTCGCAAAGCTGCGCGACTGCGATGCGCTAGCATGCGATTCGCGATCGCCCGCGACTGCGATGCGCTAGCATGCGATTCGCGATCGCCGAGCGCTCGCGAGCGCGCTAGCGGAATACTATATAGCGAGCGCTCGCGAGCGCGCTAGCGGAATACTATATAGCGCGGATCAGTCTAGATCTATGAGATCGATAGCGATCTAGCGGATCAGTCTAGATCTATGAGATCGATAGCGATCTAGAGATAGGATCGAGATCGAGGCGAGATCATATGAGCGGAGATAGGATCGAGATCGAGGCGAGATCATATGAGCGCGGCTATTTAGGCTTAGAGGATTCGGAGATTCGGAGCTCGGCTATTTAGGCTTAGAGGATTCGGAGATTCGGAGCTTAGGATTACAGAGAGCTTCTTAGGCGCTCCCGGTATCGTAGGATTACAGAGAGCTTCTTAGGCGCTCCCGGTATCGCTCCCATCCCATATTAAAATCTATCGATCGAGCTCTCCACTCCCATCCCATATTAAAATCTATCGATCGAGCTCTCCAATGCGATCGATAGGACTAGTAGCTAGCTAGCTGAGCAATGCGATCGATAGGACTAGTAGCTAGCTAGCTGAGCATGATAGGCTCGATGAGCATGAGATGCATGTACGACTGTGATAGGCTCGATGAGCATGAGATGCATGTACGACTGCATAGGCATGACTGATCGACTGCATCATGACGCATGACCATAGGCATGACTGATCGACTGCATCATGACGCATGACTGCATGCATGACTGCATATGACGGACTCGCATTAGCACTGCATGCATGACTGCATATGACGGACTCGCATTAGCACTACCGTATTTGCGCATTACCAGATTAGAGAAATGCTAGTACCGTATTTGCGCATTACCAGATTAGAGAAATGCTAGTCGATCTATCGATCGGCTATTCGCAAAGCTGCGCGACTTCGATCTATCGATCGGCTATTCGCAAAGCTGCGCGACTGCGATGCGCTAGCATGCGATTCGCGATCGCCGAGCGCGCGATGCGCTAGCATGCGATTCGCGATCGCCGAGCGCTCGCGAGCGCGCTAGCGGAATACTATATAGCGCGGATTCGCGAGCGCGCTAGCGGAATACTATATAGCGCGGATCAGTCTAGATCTATGAGATCGATAGCGATCTAGAGATACAGTCTAGATCTATGAGATCGATAGCGATCTAGAGATAGGATCGAGATCGAGGCGAGATCATATGAGCGCGGCTAGGATCGAGATCGAGGCGAGATCATATGAGCGCGGCTATTTAGGCTTAGAGGATTCGGAGATTCGGAGCTTAGGATTTTAGGCTTAGAGGATTCGGAGATTCGGAGCTTAGGATTACAGAGAGCTTCTTAGGCGCTCCCGGTATCGCTCCCATACAGAGAGCTTCTTAGGCGCTCCCGGTATCGCTCCCATCCCATATTAAAATCTATCGATCGAGCTCTCCAATGCGTCCCATATTAAAATCTATCGATCGAGCTCTCCAATGCGATCGATAGGACTAGTAGCTAGCTAGCTGAGCATGATAATCGATAGGACTAGTAGCTAGCTAGCTGAGCATGATAGGCTCGATGAGCATGAGATGCATGTACGACTGCATAGGGCTCGATGAGCATGAGATGCATGTACGACTGCATAGGCATGACTGATCGACTGCATCATGACGCATGACTGCATGCATGACTGATCGACTGCATCATGACGCATGACTGCATGCATGACTGCATATGACGGACTCGCATTAGCACTACCGGCATGACTGCATATGACGGACTCGCATTAGCACTACCGTATTTGCGCATTACCAGATTAGAGAAATGCTAGTCGATTATTTGCGCATTACCAGATTAGAGAAATGCTAGTCGATCTATCGATCGGCTATTCGCAAAGCTGCGCGACTGCGATCTATCGATCGGCTATTCGCAAAGCTGCGCGACTGCGATGCGCTAGCATGCGATTCGCGATCGCCGAGCGCTCGCGGCGCTAGCATGCGATTCGCGATCGCCGAGCGCTCGCGAGCGCGCTAGCGGAATACTATATAGCGCGGATCAGTCAGCGCGCTAGCGGAATACTATATAGCGCGGATCAGTCTAGATCTATGAGATCGATAGCGATCTAGAGATAGGATTAGATCTATGAGATCGATAGCGATCTAGAGATAGGATCGAGATCGAGGCGAGATCATATGAGCGCGGCTATTTACGAGATCGAGGCGAGATCATATGAGCGCGGCTATTTAGGCTTAGAGGATTCGGAGATTCGGAGCTTAGGATTACGGCTTAGAGGATTCGGAGATTCGGAGCTTAGGATTACAGAGAGCTTCTTAGGCGCTCCCGGTATCGCTCCCATCCAGAGAGCTTCTTAGGCGCTCCCGGTATCGCTCCCATCCCATATTAAAATCTATCGATCGAGCTCTCCAATGCGATCCATATTAAAATCTATCGATCGAGCTCTCCAATGCGATCGATAGGACTAGTAGCTAGCTAGCTGAGCATGATAGGCGATAGGACTAGTAGCTAGCTAGCTGAGCATGATAGGCTCGATGAGCATGAGATGCATGTACGACTGCATAGGCATCGATGAGCATGAGATGCATGTACGACTGCATAGGCATGACTGATCGACTGCATCATGACGCATGACTGCATGCATGACTGATCGACTGCATCATGACGCATGACTGCATGCATGACTGCATATGACGGACTCGCATGACTGCATATGACGGACTCGCA
GENE
1 ccctgtggag ccacacccta gggttggcca atctactccc aggagcaggg agggcaggag 61 ccagggctgg gcataaaagt cagggcagag ccatctattg cttacatttg cttctgacac
121 aactgtgttc actagcaacc tcaaacagac accATGgtgc acctgactcc tgaggagaag
181 tctgccgtta ctgccctgtg gggcaaggtg aacgtggatg aagttggtgg tgaggccctg
241 ggcaggttgg tatcaaggtt acaagacagg tttaaggaga ccaatagaaa ctgggcatgt
301 ggagacagag aagactcttg ggtttctgat aggcactgac tctctctgcc tattggtcta
361 ttttcccacc cttaggctgc tggtggtcta cccttggacc cagaggttct ttgagtcctt
421 tggggatctg tccactcctg atgctgttat gggcaaccct aaggtgaagg ctcatggcaa
481 gaaagtgctc ggtgccttta gtgatggcct ggctcacctg gacaacctca agggcacctt
541 tgccacactg agtgagctgc actgtgacaa gctgcacgtg gatcctgaga acttcagggt
601 gagtctatgg gacccttgat gttttctttc cccttctttt ctatggttaa gttcatgtca
661 taggaagggg agaagtaaca gggtacagtt tagaatggga aacagacgaa tgattgcatc
721 agtgtggaag tctcaggatc gttttagttt cttttatttg ctgttcataa caattgtttt
781 cttttgttta attcttgctt tctttttttt tcttctccgc aatttttact attatactta
841 atgccttaac attgtgtata acaaaaggaa atatctctga gatacattaa gtaacttaaa
901 aaaaaacttt acacagtctg cctagtacat tactatttgg aatatatgtg tgcttatttg
961 catattcata atctccctac tttattttct tttattttta attgatacat aatcattata
1021 catatttatg ggttaaagtg taatgtttta atatgtgtac acatattgac caaatcaggg
1081 taattttgca tttgtaattt taaaaaatgc tttcttcttt taatatactt ttttgtttat
1141 cttatttcta atactttccc taatctcttt ctttcagggc aataatgata caatgtatca
1201 tgcctctttg caccattcta aagaataaca gtgataattt ctgggttaag gcaatagcaa
1261 tatttctgca tataaatatt tctgcatata aattgtaact gatgtaagag gtttcatatt
1321 gctaatagca gctacaatcc agctaccatt ctgcttttat tttatggttg ggataaggct
1381 ggattattct gagtccaagc taggcccttt tgctaatcat gttcatacct cttatcttcc
1441 tcccacagct cctgggcaac gtgctggtct gtgtgctggc ccatcacttt ggcaaagaat
1501 tcaccccacc agtgcaggct gcctatcaga aagtggtggc tggtgtggct aatgccctgg
1561 cccacaagta tcacTAAgct cgctttcttg ctgtccaatt tctattaaag gttcctttgt
1621 tccctaagtc caactactaa actgggggat attatgaagg gccttgagca tctggattct
1681 gcctaataaa aaacatttat tttcattgca atgatgtatt taaattattt ctgaatattt
1741 tactaaaaag ggaatgtggg aggtcagtgc atttaaaaca taaagaaatg atgagctgtt
1801 caaaccttgg gaaaatacac tatatcttaa actccatgaa agaaggtgag gctgcaacca
1861 gctaatgcac attggcaaca gcccctgatg cctatgcctt attcatccct cagaaaagga
1921 ttcttgtaga ggcttgattt gcaggttaaa gttttgctat gctgtatttt acattactta
1981 ttgttttagc tgtcctcatg aatgtctttt cactacccat ttgcttatcc tgcatctctc
2041 tcagccttga ct
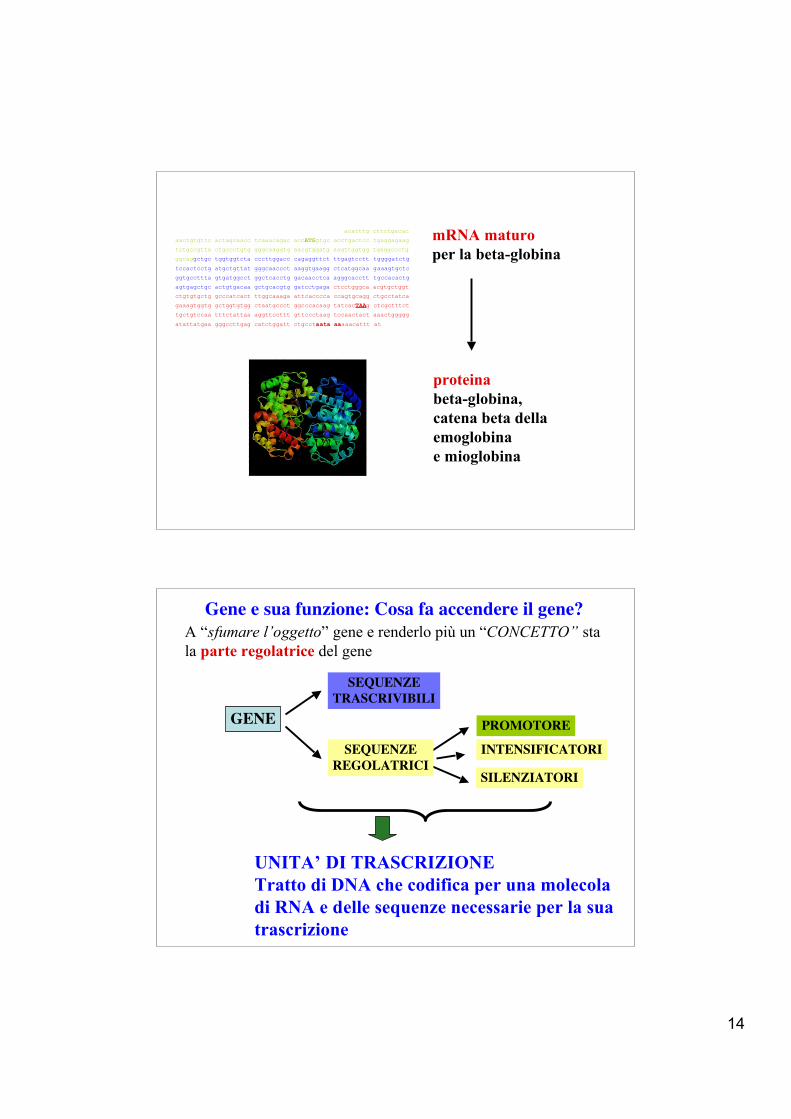
Gene della beta-globina (Homo sapiens) da 1 a 2052 nucleotidi
Sequenza GenBank n. NM_000518
DNA a filamento doppio (l’altro si ricava per complementarità)
Cromosoma 12
14
acatttg cttctgacac
aactgtgttc actagcaacc tcaaacagac accATGgtgc acctgactcc tgaggagaag
tctgccgtta ctgccctgtg gggcaaggtg aacgtggatg aagttggtgg tgaggccctg
ggcaggctgc tggtggtcta cccttggacc cagaggttct ttgagtcctt tggggatctg
tccactcctg atgctgttat gggcaaccct aaggtgaagg ctcatggcaa gaaagtgctc
ggtgccttta gtgatggcct ggctcacctg gacaacctca agggcacctt tgccacactg
agtgagctgc actgtgacaa gctgcacgtg gatcctgaga ctcctgggca acgtgctggt
ctgtgtgctg gcccatcact ttggcaaaga attcacccca ccagtgcagg ctgcctatca
gaaagtggtg gctggtgtgg ctaatgccct ggcccacaag tatcacTAAg ctcgctttct
tgctgtccaa tttctattaa aggttccttt gttccctaag tccaactact aaactggggg
atattatgaa gggccttgag catctggatt ctgcctaata aaaaacattt at
mRNA maturoper la beta-globina
proteinabeta-globina,catena beta della emoglobina e mioglobina
GENE
SEQUENZETRASCRIVIBILI
SEQUENZEREGOLATRICI
PROMOTORE
INTENSIFICATORI
SILENZIATORI
Gene e sua funzione: Cosa fa accendere il gene?
UNITA’ DI TRASCRIZIONETratto di DNA che codifica per una molecola di RNA e delle sequenze necessarie per la sua trascrizione
A “sfumare l’oggetto” gene e renderlo più un “CONCETTO” stala parte regolatrice del gene

15
Gene networks - un gene può esprimere le proprieinformazioni solo nel contesto dell’intero genoma
della cellula
Promot 3 Promot 4Promot 2gene1 gene2 gene3 gene4Promot 1
FATTORI DITRASCRIZIONE
- +DNA
RNA
proteine
Come si esprime l’informazione?
Per i geni classici vedremo:I meccanismi di Trascrizione e TraduzioneCosa è il Codice geneticoI principali meccanismi di CONTROLLO della espressione genica
16
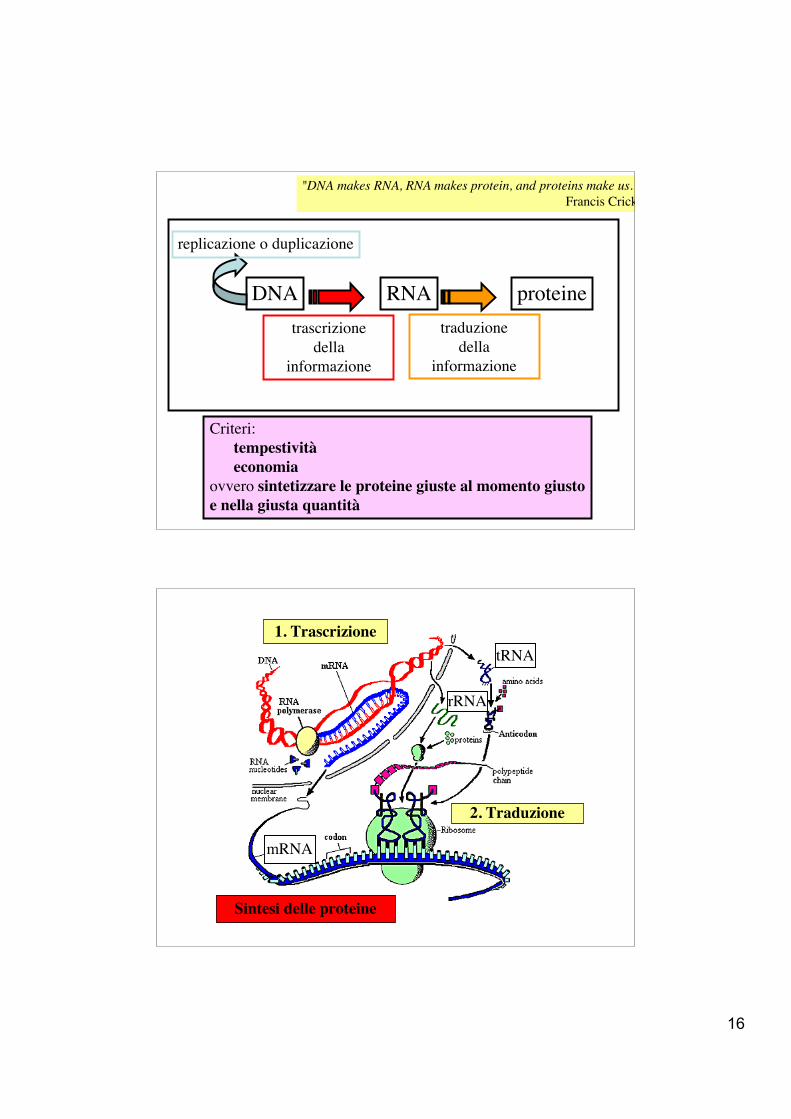
Criteri: tempestività economiaovvero sintetizzare le proteine giuste al momento giustoe nella giusta quantità
DNA RNA proteine
replicazione o duplicazione
trascrizionedella
informazione
traduzionedella
informazione
"DNA makes RNA, RNA makes protein, and proteins make us." Francis Crick
1. Trascrizione
Sintesi delle proteine
2. Traduzione
tRNA
rRNA
mRNA

17
TRASCRIZIONE (eucarioti)“assemblaggio catalizzato”
Nella trascrizione l’informazione genetica contenutanel DNA viene trascritta in una sequenza di RNA in base al principio della complementarietà delle basi
1- riconoscimento del gene da esprimere2- individuazione di inizio e termine del gene specifico
3- trascrizione nella giusta quantità del gene4- alto grado di fedeltà, cioè accuratezza
è un processo controllato, regolatopoiché le necessità sono:

18
La trascrizione genera diversi tipi di RNA:
rRNA o RNA ribosomiale
tRNA o RNA transfer o RNA ditrasporto
mRNA o RNA messaggero
HnRNA o RNA eterogeneo nucleare opre-RNA
snRNA o piccoli RNA non codificanti
gene
proteina in crescita
aa legatoal tRNA
ribosomi
Il processo è realizzato da enzimi chiamati
RNA polimerasi DNA-dipendenti
Negli eucarioti:RNA polimerasi I
RNA polimerasi II
RNA polimerasi III
rRNA : 28 S, 18 S, 5.8 S
precursori degli mRNA, ncRNA, diversi snRNA
tRNA e rRNA 5 S e alcuni snRNA
l’enzima si lega ad una sequenza sul DNA detta promotore
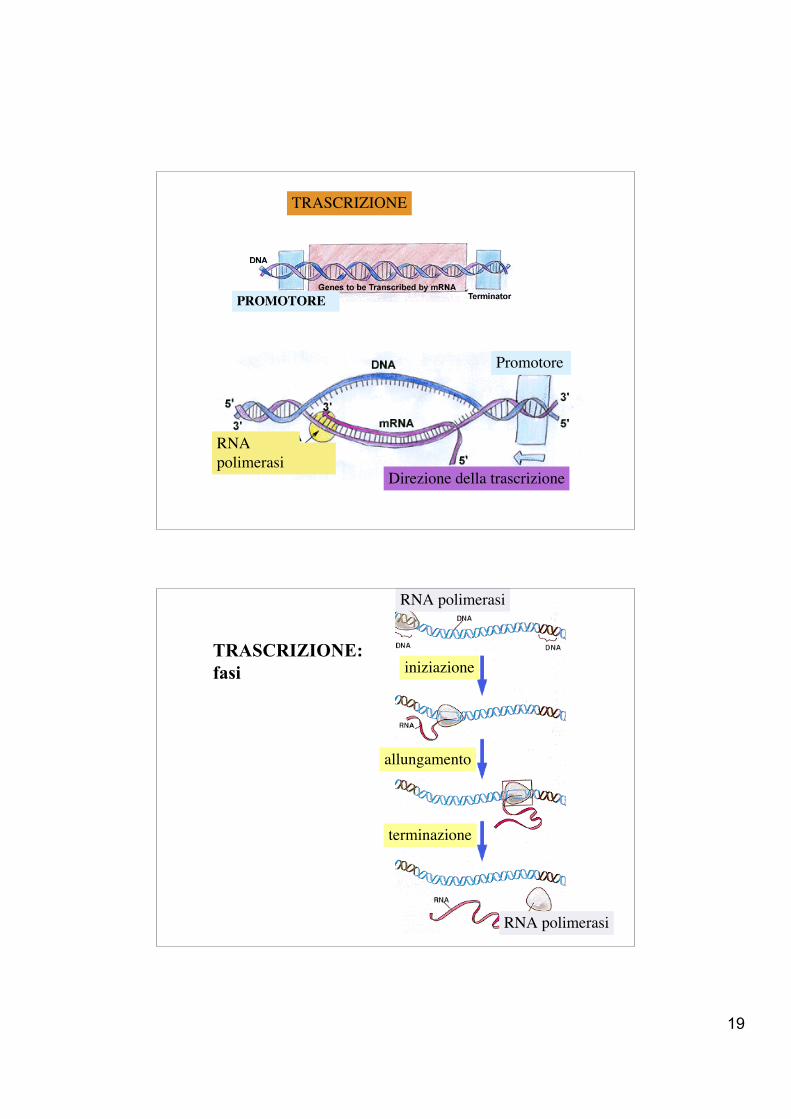
19
Promotore
RNApolimerasi
Direzione della trascrizione
TRASCRIZIONE
PROMOTORE
TRASCRIZIONE:fasi
RNA polimerasi
iniziazione
allungamento
terminazione
RNA polimerasi

20
ORIENTAMENTO DELLE UNITA’
5'
5'
3'
3'5' 3'
filamento CODING o SENSO
filamento STAMPO o TEMPLATEo ANTI SENSO
RNA POLIMERASI
VELOCITA’ca. 40 nucleotidi al secondoEs. gene distrofina lungo ca. 2.200.000 bp???Quante ore per trascriverlo?
17 ore!
filamento non-senso serve da stampo
filamento senso
basi
Entrambi i filamenti del DNA possono costituire stampoper la sintesi dell’RNA�, in funzione del gene, saranno sempre letti in direzione 3'- 5', poiché la crescita del filamento di RNA di neosintesi è in direzione 5'-3'
5' 3'
5'
Complementarietà delle basi

21
Meccanismo della trascrizione:
sintesi di RNA sulla base di uno stampo di DNA
Enzima: RNA polimerasiTrascrizione:lo stampo di DNA è letto da 3' a 5',l’RNA è sintetizzato da 5' a 3'
1° nucleotide
2° nucleotide
5'
3'
3'
5'
1° nucleotide
Uscita del pirofosfato Formazione legame fosfodiesterico
5'
Codice = sistema di segnali, o segni, o simboli, che, per convenzione, è destinato a rappresentareuna informazione tra la fonte dei segnali e ilpunto di destinazione.Es. comunicare in codice/ c. linguistico,formato da suoni/ c.grafico, c.fiscale, c. a barre, c.morse, ecc.
Definizioni
Codice geneticosequenza di codoni contenenti le informazioni genetiche del gene e determinanti la sequenza degli aa che origina la proteina in base alla lettura dei codoni stessi
CODICE GENETICO oCodice di traduzione (1964)

22
codice genetico anni ‘60
Codice genetico è un codice a triplette di nucleotidi, detti codoni
I codoni specificano per gli aa. In totale 43=64 codoni.
Caratteristiche del codice genetico
• il codice è ridondante• il codice non è ambiguo• il codice non ha punteggiatura ovvero interruzioni• il codice è letto senza sovrapposizioni • per interpretarlo è fondamentale la cornice di lettura o quadro di lettura (reading frame)• il codice è universale (ad eccezione di quello dei mitocondri)
Trp Phe Gly
ACCAAACCG
UGGUUUGGC
DNA
mRNA
proteina
Cornice di lettura

23
TRADUZIONE
TRASCRIZIONE
TRADUZIONE“assemblaggio codificato”
Nella traduzione l’informazione genetica contenuta nellasequenza di codoni lungo l’mRNA viene decodificata otradotta in una sequenza di aa costituenti la proteina, unitiin una sequenza precisa determinata dalla sequenza deicodoni
Protagonisti principali di questo processo:
mRNAtRNA
ribosomi
porta nel citoplasmail messaggio geneticosotto forma di unaspecifica sequenza dicodoni
sono gli interpreti del linguaggio
facilitano l’appaiamentospecifico tra gli anticodonidel tRNA e i codoni dell’mRNA
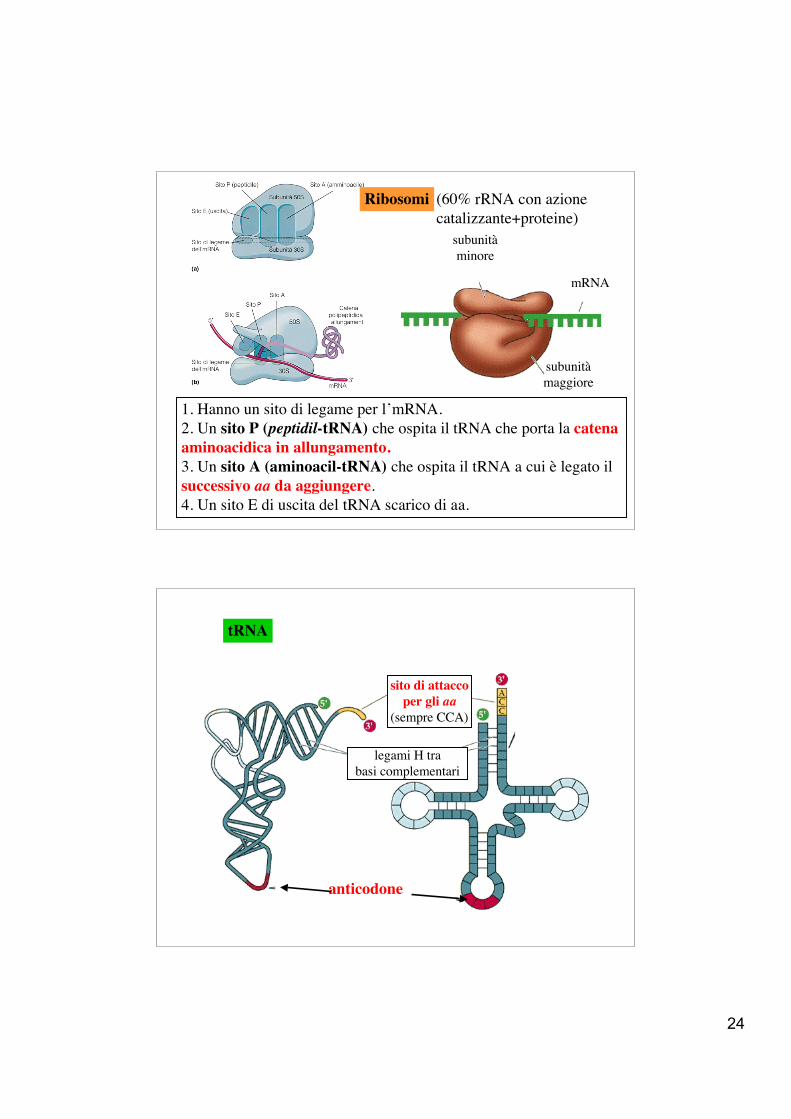
24
Ribosomi (60% rRNA con azione catalizzante+proteine)
1. Hanno un sito di legame per l’mRNA.2. Un sito P (peptidil-tRNA) che ospita il tRNA che porta la catena aminoacidica in allungamento.3. Un sito A (aminoacil-tRNA) che ospita il tRNA a cui è legato il successivo aa da aggiungere.4. Un sito E di uscita del tRNA scarico di aa.
mRNA
subunitàmaggiore
subunitàminore
anticodone
sito di attaccoper gli aa
(sempre CCA)
legami H trabasi complementari
tRNA
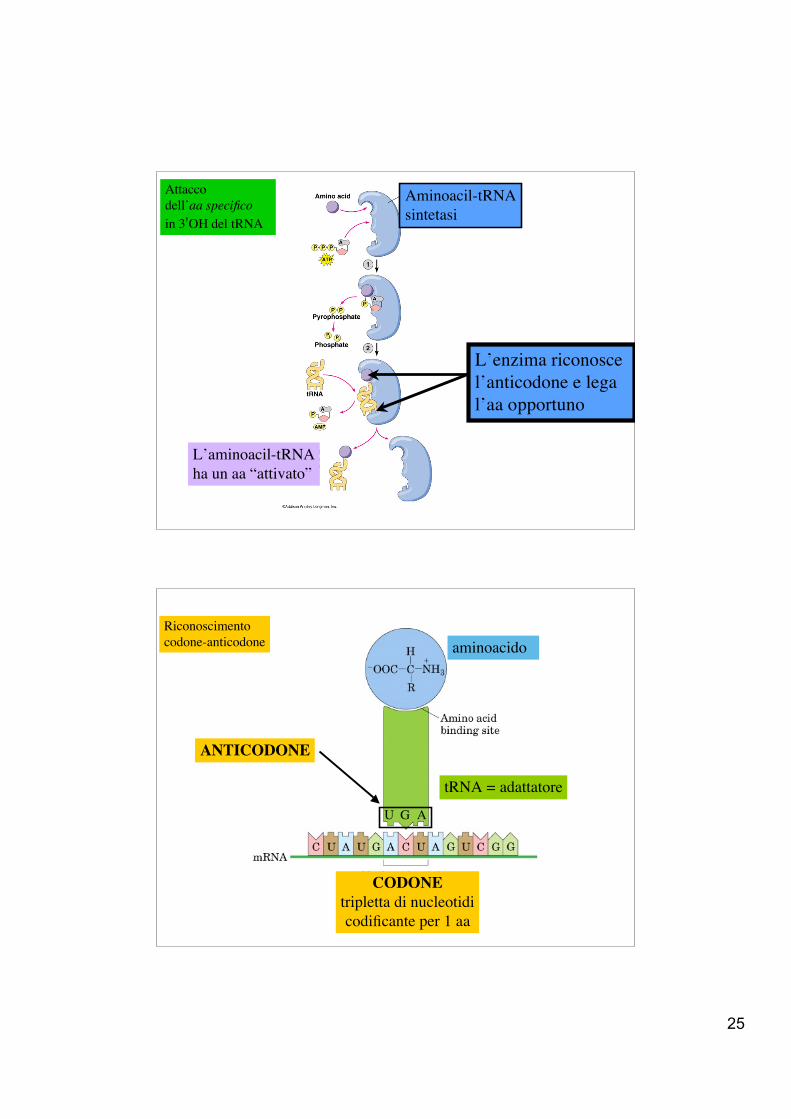
25
Attaccodell’aa specificoin 3'OH del tRNA
L’enzima riconosce l’anticodone e legal’aa opportuno
L’aminoacil-tRNAha un aa “attivato”
Aminoacil-tRNAsintetasi
tRNA = adattatore
aminoacido
CODONEtripletta di nucleotidicodificante per 1 aa
Riconoscimento codone-anticodone
ANTICODONE
U G A

26
Inizio sintesi proteica
Allungamento sintesi proteica
Terminazione sintesi proteica
Met
5’ 3’mRNA
AUG
Met
3’
Ser
5’mRNA
Met-Ser Pro Met-Ser-Pro-Thr
5’mRNA
Met-Ser-Pro-Thr-...-...-...-
UGA
STOP!
3’
NH3 COO-
Per individuare il sito di INIZIO della TRADUZIONE: Sequenza consenso di M.Kozak: R--AUGG, R sta per una purina A o G
Destino proteico:l’mRNA può essere tradottoda ribosomi liberi o ribosomi legati al RER
nucleo
mitocondri cloroplasti
citoplasma
vescicolesecretorie lisosomim.p.
RIBOSOMI LIBERI
RIBOSOMI SUL RETICOLO ENDOPLASMATICO

27
Le proteine ed i loro segnali di smistamento
Proteina non ripiegata Proteina ripiegata
zona segnale
sequenza segnale
Ad ogni segnale deve corrispondere un recettore proteicocomplementare
REGOLAZIONEESPRESSIONE GENICA
eucarioti
28
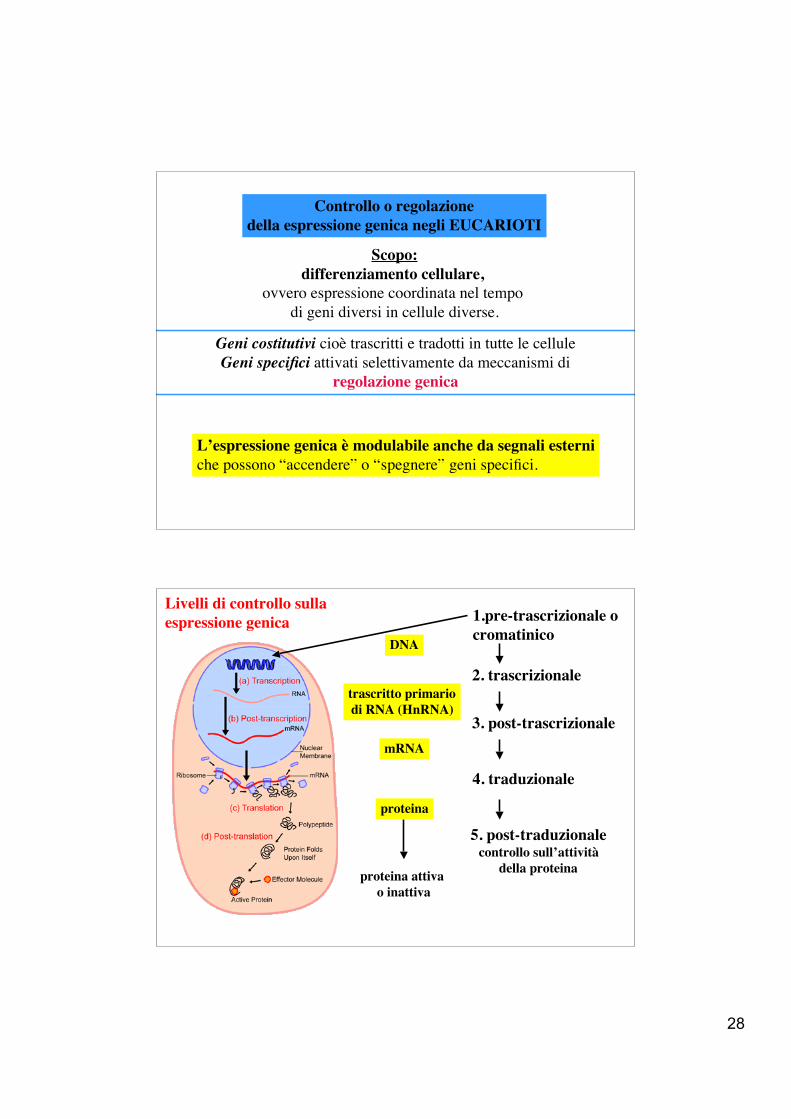
Controllo o regolazionedella espressione genica negli EUCARIOTI
Scopo:differenziamento cellulare,
ovvero espressione coordinata nel tempo di geni diversi in cellule diverse.
Geni costitutivi cioè trascritti e tradotti in tutte le celluleGeni specifici attivati selettivamente da meccanismi di
regolazione genica
L’espressione genica è modulabile anche da segnali esterniche possono “accendere” o “spegnere” geni specifici.
Livelli di controllo sulla espressione genica 1.pre-trascrizionale o
cromatinico
2. trascrizionale
3. post-trascrizionale
4. traduzionale
5. post-traduzionalecontrollo sull’attività
della proteina
trascritto primariodi RNA (HnRNA)
mRNA
proteina
DNA
proteina attiva o inattiva
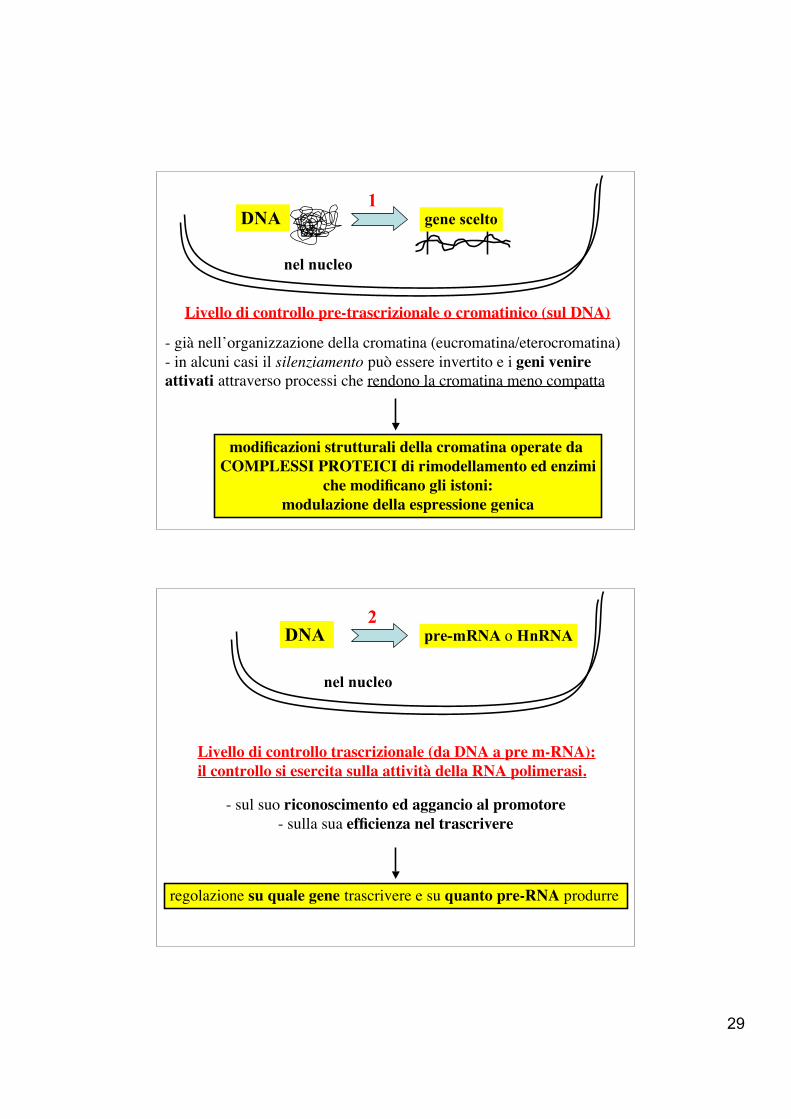
29
- già nell’organizzazione della cromatina (eucromatina/eterocromatina)- in alcuni casi il silenziamento può essere invertito e i geni venireattivati attraverso processi che rendono la cromatina meno compatta
modificazioni strutturali della cromatina operate da COMPLESSI PROTEICI di rimodellamento ed enzimi
che modificano gli istoni:modulazione della espressione genica
DNA
nel nucleo
Livello di controllo pre-trascrizionale o cromatinico (sul DNA)
gene scelto1
- sul suo riconoscimento ed aggancio al promotore- sulla sua efficienza nel trascrivere
regolazione su quale gene trascrivere e su quanto pre-RNA produrre
DNA pre-mRNA o HnRNA
nel nucleo
Livello di controllo trascrizionale (da DNA a pre m-RNA):il controllo si esercita sulla attività della RNA polimerasi.
2

30
PROMOTOREnatura modulare
Sequenza nel DNA di almeno 40 nucleotidi, con affinità più o meno elevata per la RNA polimerasi, di solito posta a monte del gene.
Hanno:siti di riconoscimento (es. TATA box)sito di legame stabilesito di inizio della trascrizionesiti di regolazione (simili a intensificatori e/o silenziatori)
GENE
SEQUENZEREGOLATRICI
SEQUENZETRASCRIVIBILI
PROMOTORE
INTENSIFICATORI
SILENZIATORI
Gene: Cosa fa accendere il gene?
Il PROMOTORESequenza data dalla combinazioni di corti elementi solitamente collocati nella regione immediatamente a monte del gene, spesso entro 200 bp dal sito di inizio della trascrizione. I PROMOTORI HANNO NATURA MODULARE!
Risposta: NECESSITA’ DI SEQUENZE SEGNALE
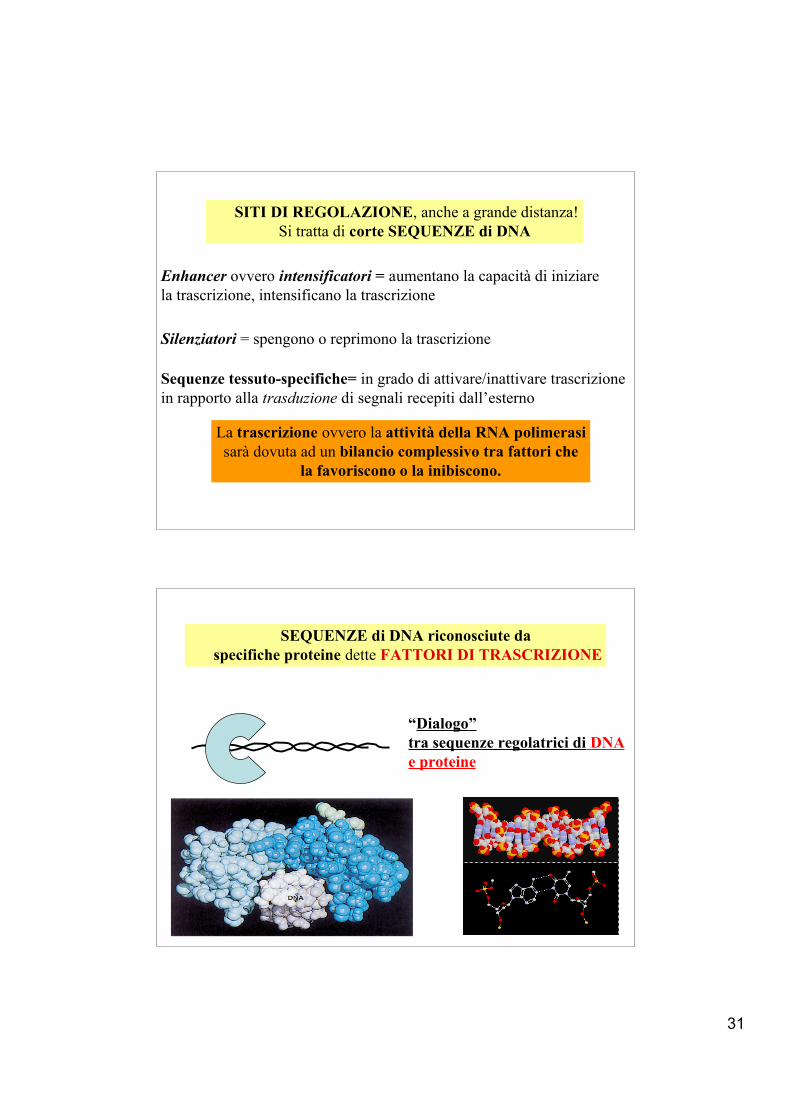
31
Enhancer ovvero intensificatori = aumentano la capacità di iniziarela trascrizione, intensificano la trascrizione
Sequenze tessuto-specifiche= in grado di attivare/inattivare trascrizionein rapporto alla trasduzione di segnali recepiti dall’esterno
Silenziatori = spengono o reprimono la trascrizione
La trascrizione ovvero la attività della RNA polimerasi sarà dovuta ad un bilancio complessivo tra fattori che
la favoriscono o la inibiscono.
SITI DI REGOLAZIONE, anche a grande distanza!Si tratta di corte SEQUENZE di DNA
SEQUENZE di DNA riconosciute da specifiche proteine dette FATTORI DI TRASCRIZIONE
“Dialogo”tra sequenze regolatrici di DNAe proteine

32
FATTORI GENERALIo BASALI
di trascrizione
PROTEINE REGOLATRICISPECIFICHE
Nella regolazione della trascrizione, ad attivare o reprimere la RNA polimerasi, intervengono
proteine di 2 tipi:
RNA polimerasi II
Inizio della trascrizioneFATTORI GENERALIdi trascrizione
promotore
Questi fattori sono richiesti per l’inizio della trascrizionee sono simili per tutti i geni trascritti dalla RNApolimerasi II
TATA box
TFIID TFIIB
DNAgene X

33
promotore
TATA box
TFIID TFIIB
DNA
gene XRNA polimerasi II
Sequenze regolatrici
PROTEINE REGOLATRICISPECIFICHE
trascritto di RNA
Le proteine specifiche si possono legare a sequenze specifiche, anche lontane sia a valle che a monte del gene.Il ripiegamento del DNA le porta poi vicine al promotore per influenzare la polimerasi.
Integrazione al PROMOTORE di:- RNA pol- fattori generali di trascrizione - serie multiple di proteine regolatrici
EFFETTO COMBINATORIOUn singolo promotore può essere regolato da molte sequenze regolatrici sparse lungo il DNA e riconosciute da più proteine regolatrici
proteine regolatrici
+ + - +
sinergia trascrizionale
Come agiscono i fattori di trascrizione o proteine regolatrici?

34
EFFETTO DI COORDINAZIONEUna singola proteina regolatrice/fattore di trascrizionepuò regolare e quindi coordinare l’espressione genica di parecchi geni diversi
proteina regolatrice
Gene networks - un gene può esprimere le proprieinformazioni solo nel contesto dell’intero genoma
della cellula
Promot 3 Promot 4Promot 2gene1 gene2 gene3 gene4Promot 1
FATTORI DITRASCRIZIONE
- +DNA
RNA
proteine

35
pre-mRNA o HnRNA mRNA (maturo)
nel nucleo
Livello di controllo post-trascrizionale (da pre m-RNA a mRNA):
-controllo sulla elaborazione dell’mRNA,la maturazione comporta 3 tipi fondamentali di modificazionichimiche del trascritto primario a ottenere mRNA maturo
- controllo del trasporto dell’mRNA al citoplasma e sulla suastabilità nel citoplasma (importante il 3’UTR)
3
4
Eventi post-trascrizionali:dal preRNA all’mRNA
1- aggiunta del CAP (m7G)aggiunta in 5’ di una 7metil-guanosina:preserva il trascritto dalla degradazione ed è segnale diaggancio per il ribosoma
3- splicingprocesso di taglia e cuci per eliminare gli introni
2- poli-adenilazione in 3’OHaggiunta di una coda di poliA (200-250):aiuta il passaggo al citoplasma, influenza la stabilità dell’mRNA

36
TATADNAPROMOTORE
+1-25
E 1 E 2 E 3
TrascrittoPrimario o pre-mRNA
Esone 1 Esone 2 Esone 3
AUG UGA
I 1 I 2
Da pre-mRNA a mRNA maturo!!!
mRNA AUG UGACAP5’
aaaaaaaaa3’
SPLICING= TAGLIA E CUCI degli introni/esoni

37
Spliceosoma = complesso ribonucleoproteico150 proteine, 5 RNA snRNA (50-200 nucleotidi): U1, U2, U4, U5, U6Dimensioni simili ad un ribosoma
snRNAsmall nuclear RNA
Proteine
snRNPs
SPLICING ALTERNATIVO: lo splicing mostra unanotevole flessibilità
12
12
12
12Aggiunta di un esone
Aggiunta di un tratto intronico
Esclusione a vicendadi esoni
Sito di splicing interno
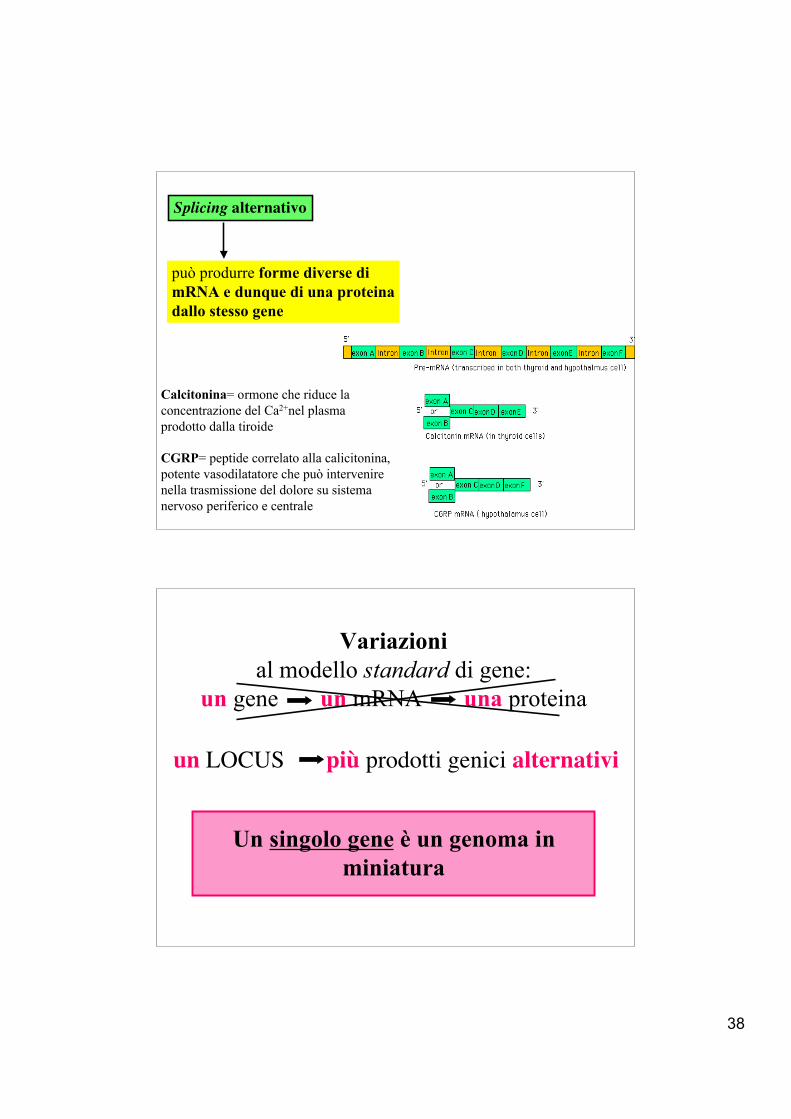
38
Splicing alternativo
può produrre forme diverse dimRNA e dunque di una proteinadallo stesso gene
Calcitonina= ormone che riduce la concentrazione del Ca2+nel plasmaprodotto dalla tiroide
CGRP= peptide correlato alla calicitonina, potente vasodilatatore che può intervenire nella trasmissione del dolore su sistema nervoso periferico e centrale
Variazionial modello standard di gene:
un gene un mRNA una proteina
Un singolo gene è un genoma inminiatura
un LOCUS più prodotti genici alternativi

39
L’mRNA al termine della maturazione presenta: - Regione non tradotta in 5´, 5´ UTR o leader - Regione codificante, CDS (coding sequence) - Regione non tradotta in 3´, 3´ UTR o trailer
N C
PROTEINA
TRADUZIONE
CAP AAAAA
5’UTR 3’UTR
5’ 3’SEQUENZA CODIFICANTE
AUG
mRNA MATURO
UAACDS UGAUAG
STOP
DOPO MATURAZIONE
Funzione delle regioni dell'mRNA
5´UTR Efficienza della traduzione
CDS Traduzione (1%)
3´UTR Stabilità (velocità di degradazione;concetto di emivita da pochi minutia 20-30 ore)

40
mRNA (maturo)5
proteina
nel citoplasma
5. Livello di controllo traduzionale (da m-RNA a proteina):Operato da fattori proteici che possono influire sull’inizio della traduzione e quindi modularla, legandosi alla regione 5´UTR emagari “mascherandola” al ribosoma
6. Livello di controllo post-traduzionale (sulla proteina):- controllo sulla modificazione delle proteine sintetizzate-controllo del ripiegamento (aiutato da proteine chaperon) e trasporto nella cellula al sito di funzione
6proteina attiva
Livello di controllo post-traduzionale (sulla proteina):modificazione delle proteine sintetizzate che risultano essenzialiperché la proteina possa funzionare
Fosforilazionein TIROSINATREONINASERINA

41
Il nuovo “dogma”
ncRNA
microRNA
A modulare la espressione genica delle proteine cellulariintervengono anche i geni non codificanti proteine oncRNA (non coding RNA)
Ipotesi
ncRNA non sono semplice “rumore trascrizionale”, mapiù probabilmente svolgono un ruolo regolatorio e sonocoinvolti in numerosi processi cellulari e biologici, inparticolare durante lo sviluppo e il differenziamento

42
Gruppo eteogeneo per dimensioni e funzione: difficile classificazione
Caratterizzati spesso da una alta densità di codoni di STOP e quindi dalla assenza di ORF (cornici di lettura aperte) estese
Grande importanza per il ruolo da protagonisti nella REGOLAZIONE GENICA
Possono formarsi dalle zone introniche di geni coding o da vere e proprie unità geniche
Sempre di più nei database internazionali
ncRNA - generalità
La loro trascrizione ed elaborazione è sottoposta a controllo
ncRNA - meccanismi
Silenziamento genico post-trascrizionale: stabilità mRNA Attraverso meccanismi di “Interferenza da RNA”
Cambiamenti nei livelli di espressione degli ncRNA sono associaticon forme diverse di cancro e patologie neurodegenerative
Modulazione della trascrizione (interazioni a triplice elica)
Dinamica della struttura della cromatina (regolatori epigenetici)
Regolazione espressione genica

43
ncRNAs
2- small RNAsnoRNA
alcuninon noti
~20-300 nt
~300->10.000 nt
~18-25 nt
3-Medium e largeRNAs
1- microRNAsiRNA
Studi funzionali
Nuovi ncRNAsfunzionali
F.F. Costa“Non-coding RNAs: lost in translation?”Gene 386 (2007) 1-10
ncRNAs
alcuninon noti
3-Medium e largeRNAs
~18-25 nt- silenziamentogenico post-trascrizionale(interferenza daRNA)
~300 - >10000 nt
- inattivazione X- regolazionetrascrizionale
~20- 300 nt- modificazioni in RNA- regolazione trascrizionale- ruolo strutturale
2- small RNAsnoRNA,
1- microRNAsiRNA
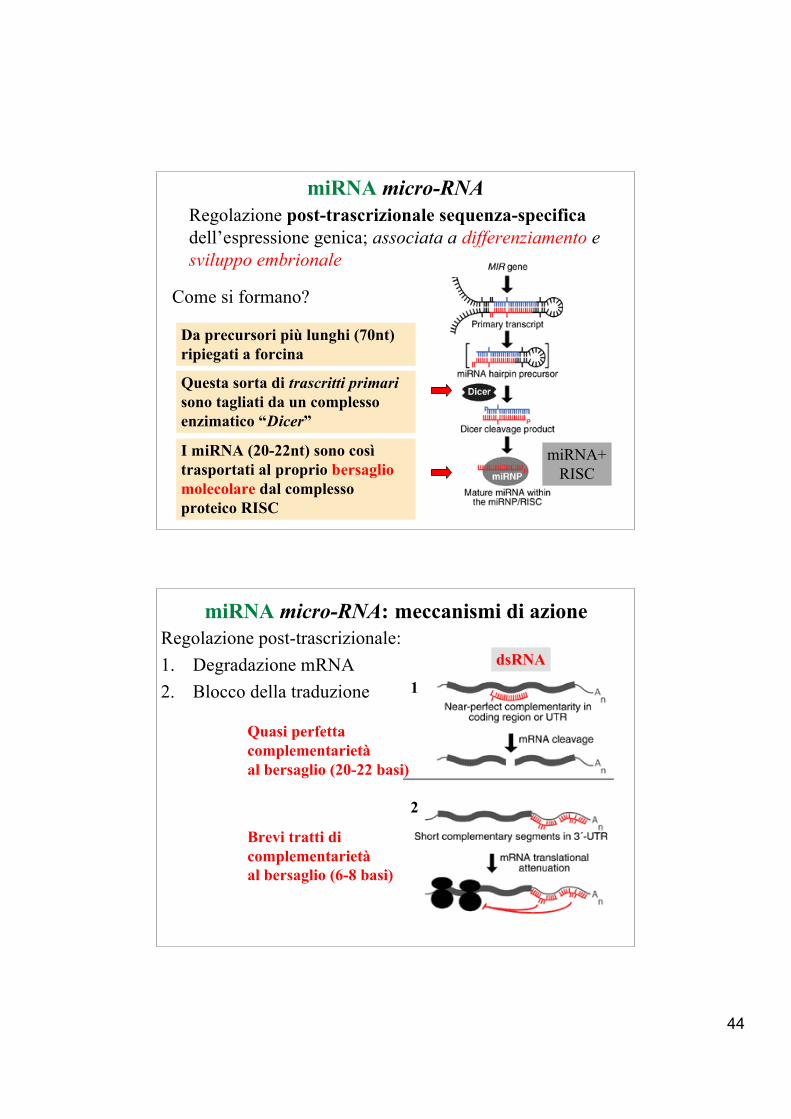
44
miRNA micro-RNARegolazione post-trascrizionale sequenza-specificadell’espressione genica; associata a differenziamento esviluppo embrionale
Da precursori più lunghi (70nt)ripiegati a forcina
Come si formano?
Questa sorta di trascritti primarisono tagliati da un complessoenzimatico “Dicer”
I miRNA (20-22nt) sono cosìtrasportati al proprio bersagliomolecolare dal complessoproteico RISC
miRNA+RISC
miRNA micro-RNA: meccanismi di azioneRegolazione post-trascrizionale:1. Degradazione mRNA2. Blocco della traduzione 1
2
Quasi perfetta complementarietà al bersaglio (20-22 basi)
Brevi tratti di complementarietà al bersaglio (6-8 basi)
dsRNA

45
miRNA
Curiosità e spunti:• la maggioranza di quelli specificamente espressi incellule staminali embrionali umane sono down-regolatidurante il proseguo dello sviluppo embrionale• molti specificamente implicati nello sviluppo delcervello come dimostrato in diversi lavori sul modellomurino• altri legati a sviluppo di cancro: agirebbero comeoncogeni od onco-soppressori (es. miR-21, let-7, miR-122a)
Andrew Fire, 47 anniStanford California
Craig Mello, 46 anniMassachusetts Medical School
Premio Nobel per la medicina 2006
Nature, feb. 1998“Potent and specific genetic interference by double-
stranded RNA (dsRNA)in Caenorhabditis elegans”

46
Cosa si intende per RNA interference(RNAi)
Sistema di silenziamento genico basato su RNA
RNAi è un processo mediante il quale RNA a doppio filamento (doublestrand RNA o dsRNA) silenzia, in modo sequenza-specifico,l’espressione di geni omologhi per sequenza attraverso l’appaiamentocon l’mRNA bersaglio, seguito dalla sua degradazione; quindi:silenziamento post-trascrizionale.
E’ un meccanismo conservato a livello evolutivodi degradazione specifica di mRNA bersaglio cheavviene in 2 stadi:
1°stadio-iniziale. I dsRNAs sonoprocessati da un enzima dimerico,DICER (RNasiIII). Si generano isiRNAs.
2°stadio-effettore. siRNAs o smallinterfering RNA (< 22nt), vengonoincorporati in un secondo complessoenzimatico (endonucleasico), RISC(complesso di silenziamento indotto daRNA) che degrada uno dei duefilamenti e usa l’altro per riconoscere,in base all’appaiamento delle basi,il substrato da degradare attraversol’azione in base alla complementarietàdelle basi.
dsRNA
siRNA
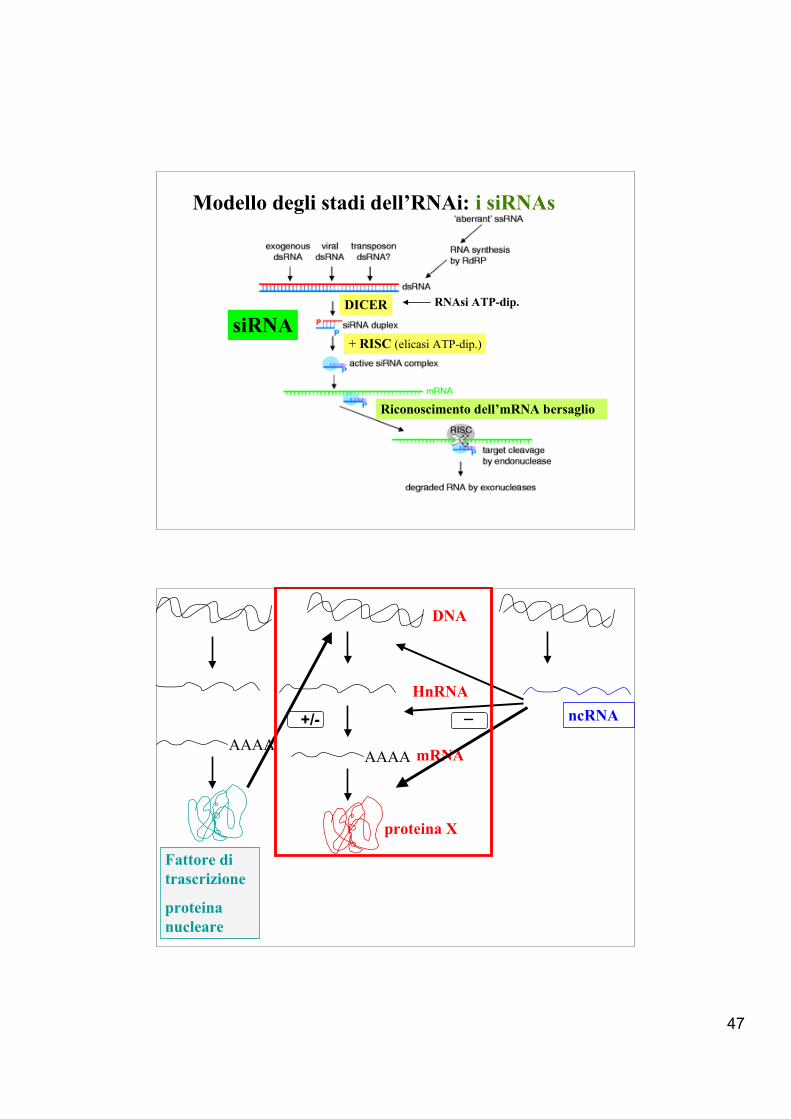
47
Modello degli stadi dell’RNAi: i siRNAs
RNAsi ATP-dip.
+ RISC (elicasi ATP-dip.)
DICER
Riconoscimento dell’mRNA bersaglio
siRNA
AAAA
HnRNA
proteina X
DNA
ncRNA
AAAA
Fattore ditrascrizione
proteinanucleare
mRNA
_+/-
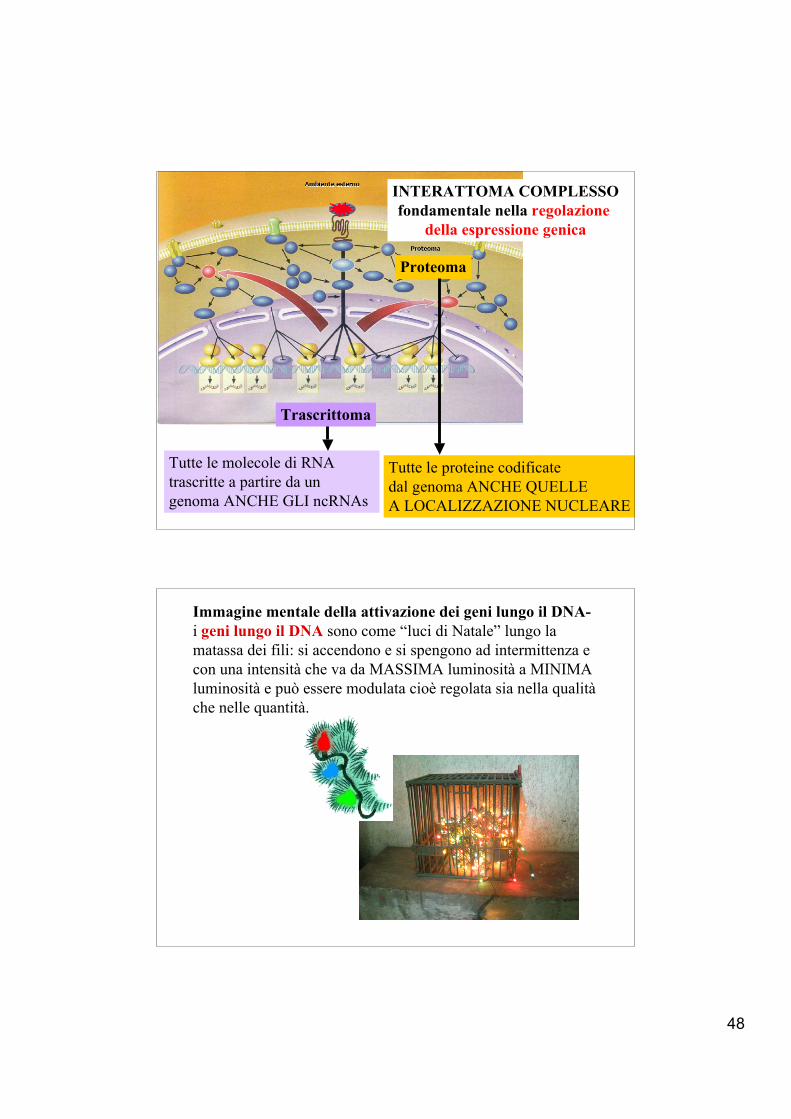
48
Tutte le molecole di RNAtrascritte a partire da ungenoma ANCHE GLI ncRNAs
Tutte le proteine codificate dal genoma ANCHE QUELLEA LOCALIZZAZIONE NUCLEARE
Trascrittoma
Proteoma
INTERATTOMA COMPLESSOfondamentale nella regolazione
della espressione genica
Immagine mentale della attivazione dei geni lungo il DNA-i geni lungo il DNA sono come “luci di Natale” lungo lamatassa dei fili: si accendono e si spengono ad intermittenza e con una intensità che va da MASSIMA luminosità a MINIMAluminosità e può essere modulata cioè regolata sia nella qualità che nelle quantità.


















