DIPARTIMENTO DI FARMACIA - core.ac.uk · immunitario come milza, timo, tonsille e tessuto linfoide...
77
UNIVERSITA’ DI PISA DIPARTIMENTO DI FARMACIA Corso di Laurea Specialistica in FARMACIA Tesi di Laurea Studio del ruolo del sistema endocannabinoide in cellule staminali di melanoma cutaneo maligno. Relatore Candidato Prof. Stefano Fogli Sara Barbani Correlatore Dott.ssa Sara Carpi Anno accademico 2014/2015
Transcript of DIPARTIMENTO DI FARMACIA - core.ac.uk · immunitario come milza, timo, tonsille e tessuto linfoide...

UNIVERSITA’ DI PISA
DIPARTIMENTO DI FARMACIA
Corso di Laurea Specialistica in
FARMACIA
Tesi di Laurea
Studio del ruolo del sistema endocannabinoide in cellule
staminali di melanoma cutaneo maligno.
Relatore Candidato
Prof. Stefano Fogli Sara Barbani
Correlatore
Dott.ssa Sara Carpi
Anno accademico 2014/2015

1
INDICE
Capitolo 1. Introduzione 4
1.1 Il Sistema endocannabinoide 4
1.1.1 Endocannabinoidi 4
1.1.2 Recettori 5
1.1.3 Sintesi e metabolismo 7
1.1.4 Ruolo fisiologico 10
1.1.5 Sistema endocannabinoide e cancro 10
1.2 Melanoma cutaneo 12
1.2.1 Definizione di melanoma 12
1.2.2 Epidemiologia 13
1.2.3 Fattori di rischio 15
1.2.4 Patogenesi molecolare 18
1.3 Cellule staminali tumorali di melanoma 21
1.3.1 Cellule staminali tumorali e sistema
endocannabinoide 22
Capitolo 2. Scopo della ricerca 24
Capitolo 3. Materiali e metodi 25
3.1 Materiali per gli studi funzionali 25

2
3.1.1 Linee cellulari 25
3.1.2 Soluzioni e tamponi utilizzati 27
3.2 Materiali per gli studi di biologia molecolare 29
3.2.1 Kit e reagenti 29
3.2.2 Primer 30
3.3 Materiali per Western Blotting 32
3.4 Metodi utilizzati negli studi funzionali 33
3.4.1 Scongelamento della linea cellulare 33
3.4.2 Mantenimento in coltura 34
3.4.3 Congelamento cellulare 35
3.5 Metodi utilizzati negli studi di biologia molecolare 36
3.5.1 Estrazione dell’RNA totale 36
3.5.2 Retrotrascrizione 38
3.5.3 Real-Time PCR 39
3.5.4 TaqMan PCR 40
3.6 Metodi utilizzati per l’analisi tramite Western Blotting 42
3.6.1 Estrazione proteica 42
3.6.2 Quantizzazione proteica 43
3.6.3 Western Blotting 44
Capitolo 4. Risultati e discussione 51
4.1 Profilo di espressione genica dei componenti del sistema
endocannabinoide nel melanoma. 51

3
4.1.1 Valutazione dell’espressione del recettore CB1 in
cellule di melanoma. 51
4.1.2 Valutazione dell’espressione del recettore CB1
in cellule staminali di melanoma. 52
4.1.3 Valutazione dell’espressione del recettore CB2
in cellule di melanoma. 53
4.1.4 Valutazione dell’espressione degli enzimi
coinvolti nella biosintesi e nel catabolismo di AEA in
cellule di melanoma. 54
4.1.5 Valutazione dell’espressione degli enzimi
coinvolti nella biosintesi e nel catabolismo di 2-AG in
cellule di melanoma. 54
4.2 Profilo di espressione proteica del recettore CB1 nel
melanoma. 56
Capitolo 5. Conclusioni 57
Nomenclatura 59
Bibliografia 61
Indice tabelle 74
Indice figure 75
Ringraziamenti 76

Introduzione Capitolo 1
4
CAPITOLO 1
Introduzione
1.1 Il Sistema endocannabinoide
Il sistema endocannabinoide (ECS) è costituito da un insieme di elementi eterogenei
che nello specifico sono (Pertwee R.G., 2010):
- Cannabinoidi endogeni o endocannabinoidi in grado di attivare i rispettivi
recettori;
- Due tipi di recettori dei cannabinoidi, il recettore CB1 ed il recettore CB2;
- Proteine responsabili della sintesi, dell’uptake e del metabolismo degli
stessi endocannabinoidi.
1.1.1 Endocannabinoidi
Gli endocannabinoidi sono stati identificati come ligandi endogeni specifici in
grado di attivare i recettori dei cannabinoidi. Il primo endocannabinoide individuato
è stato l’anandamide (AEA) a cui ha fatto seguito l’osservazione di un metabolita
endogeno, il 2-arachidonoilglicerolo (2-AG).
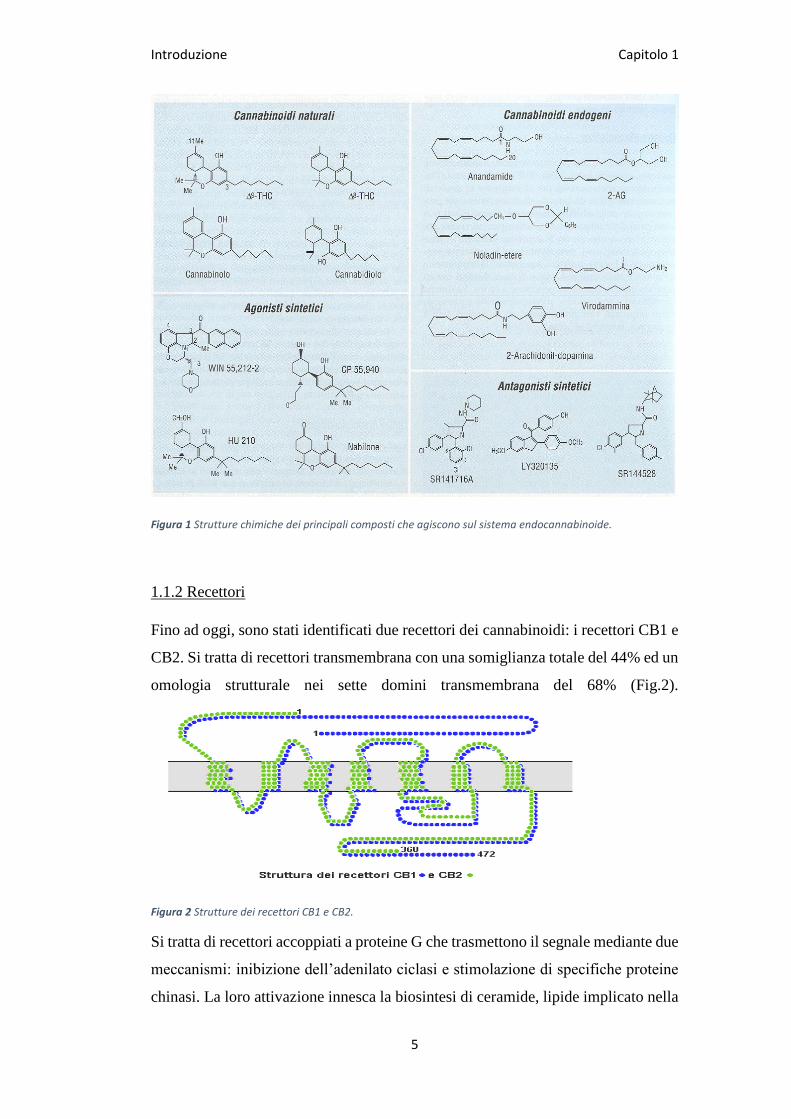
Introduzione Capitolo 1
5
Figura 1 Strutture chimiche dei principali composti che agiscono sul sistema endocannabinoide.
1.1.2 Recettori
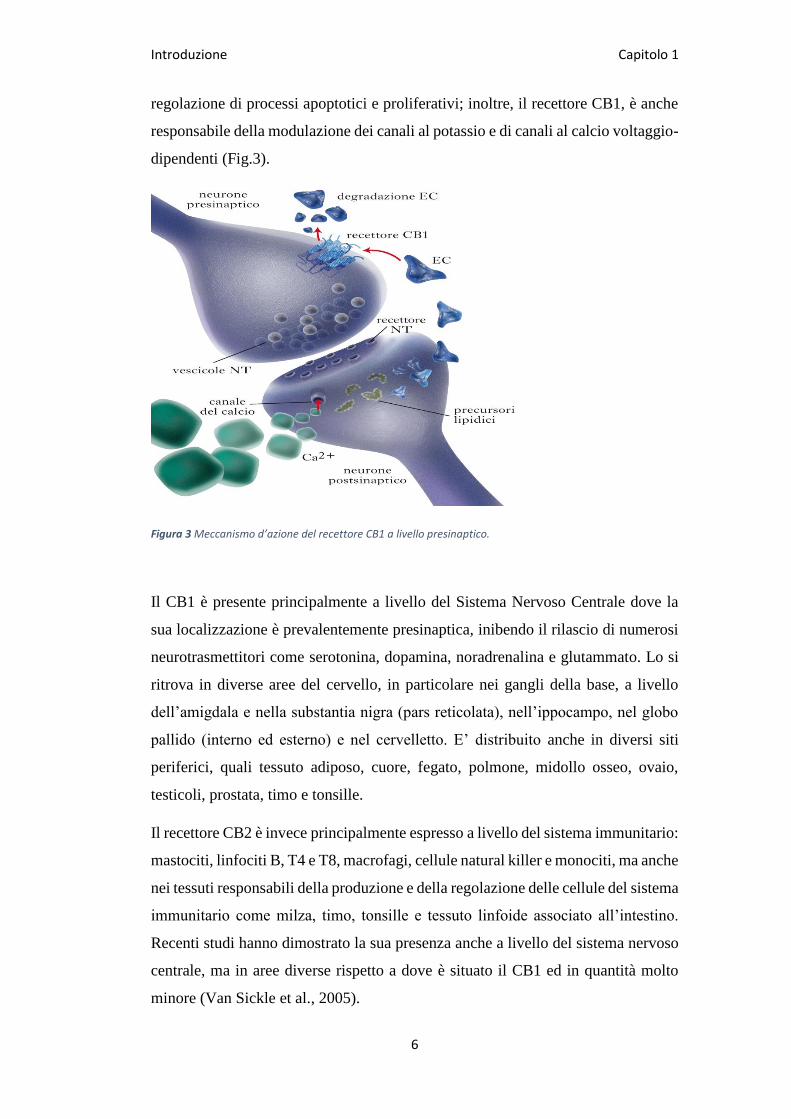
Fino ad oggi, sono stati identificati due recettori dei cannabinoidi: i recettori CB1 e
CB2. Si tratta di recettori transmembrana con una somiglianza totale del 44% ed un
omologia strutturale nei sette domini transmembrana del 68% (Fig.2).
Figura 2 Strutture dei recettori CB1 e CB2.
Si tratta di recettori accoppiati a proteine G che trasmettono il segnale mediante due
meccanismi: inibizione dell’adenilato ciclasi e stimolazione di specifiche proteine
chinasi. La loro attivazione innesca la biosintesi di ceramide, lipide implicato nella
Introduzione Capitolo 1
6
regolazione di processi apoptotici e proliferativi; inoltre, il recettore CB1, è anche
responsabile della modulazione dei canali al potassio e di canali al calcio voltaggio-
dipendenti (Fig.3).
Figura 3 Meccanismo d’azione del recettore CB1 a livello presinaptico.
Il CB1 è presente principalmente a livello del Sistema Nervoso Centrale dove la
sua localizzazione è prevalentemente presinaptica, inibendo il rilascio di numerosi
neurotrasmettitori come serotonina, dopamina, noradrenalina e glutammato. Lo si
ritrova in diverse aree del cervello, in particolare nei gangli della base, a livello
dell’amigdala e nella substantia nigra (pars reticolata), nell’ippocampo, nel globo
pallido (interno ed esterno) e nel cervelletto. E’ distribuito anche in diversi siti
periferici, quali tessuto adiposo, cuore, fegato, polmone, midollo osseo, ovaio,
testicoli, prostata, timo e tonsille.
Il recettore CB2 è invece principalmente espresso a livello del sistema immunitario:
mastociti, linfociti B, T4 e T8, macrofagi, cellule natural killer e monociti, ma anche
nei tessuti responsabili della produzione e della regolazione delle cellule del sistema
immunitario come milza, timo, tonsille e tessuto linfoide associato all’intestino.
Recenti studi hanno dimostrato la sua presenza anche a livello del sistema nervoso
centrale, ma in aree diverse rispetto a dove è situato il CB1 ed in quantità molto
minore (Van Sickle et al., 2005).

Introduzione Capitolo 1
7
Inizialmente la stimolazione del recettore CB1 era stata esclusivamente associata
agli effetti psicoattivi della cannabis, mentre il CB2 si pensava fosse coinvolto
esclusivamente in eventi relativi al sistema immunitario. Recentemente, questa idea
originaria si è evoluta nel concetto secondo cui entrambi i recettori sono in grado
di controllare funzioni centrali e periferiche, come sviluppo neuronale, processi
infiammatori e metabolici e rilascio di ormoni (Di Marzo V., 2009).
1.1.3 Sintesi e metabolismo
I principali agonisti endogeni del sistema endocannabinoide sono l’anandamide
(AEA) e il 2-arachidonoilglicerolo (2-AG).
La biosintesi di AEA comprende due fasi: la prima fase prevede la formazione di
N-arachidonoil-fosfatidiletanolammina (NArPE) dalla fosfatidiletanolammina
grazie all’N-aciltransferasi calcio-dipendente; la seconda fase è caratterizzata dalla
vera e propria conversione di NArPE in AEA grazie all’enzima l’N-
acylphosphatidylethanolamine-hydrolyzing phospholipase (NAPE-PLD)
La biosintesi del 2-AG, invece, inizia a partire dal sn-1-acyl-2-
arachidonoylglycerols (DAGs), il quale viene convertito in 2-AG mediante l’azione
di due enzimi DAG lipasi calcio-dipendenti, DAGL-a e DAGL-b.
I due endocannabinoidi sono inattivati attraverso due fasi: uptake cellulare
attraverso trasportatori di membrana, “endocannabinoid membrane transporter”
(EMT), ed idrolisi enzimatica intracellulare.
L’AEA viene principalmente inattivata dall’enzima Fatty Acid Amide Hydrolase
(FAAH) e portata ad acido arachidonico ed etanolammina; oltre a quest’ultimo,
anche l’enzima l’N-acylethanolamine acid amidase (NAAA) è coinvolto
nell’idrolisi di AEA con implicazioni fisiologiche non ancora ben conosciute.
Il 2-AG è inattivato e convertito in glicerolo ed acido arachidonico in parte,
dall’enzima FAAH, ma principalmente dal monoacylglycerol lipase (MAGL),
responsabile dell’idrolisi del 2-AG per l’85% nel cervello. Inoltre, il 2-AG, può
essere inattivato da due proteine integrali di membrana, l’ABHD6 e l’ABHD12.

Introduzione Capitolo 1
8
L’AEA e il 2-AG sono anche dei substrati per gli enzimi cyclooxygenase-2 (COX-
2), lipossigenasi (LOX), e citocromo P450; è possibile che questi ultimi giochino
un ruolo di controllo sui livelli degli endocannabinoidi attraverso processi di ossido-
riduzione producendo prostaglandine, esteri del glicerolo, idrossianandamidi e
idrossieicosanoidi, implicati in diverse attività biologiche e soprattutto coinvolti in
processi infiammatori (Hermanson DJ. et al., 2011).
Figura 4 Catabolismo degli endocannbinoide AEA e 2-AG.
Biosintesi, azione e degradazione di questi endocannbinoidi vengono innescate “on
demand” e sono normalmente limitate nel tempo a causa della natura lipofila di tali
composti, al percorso biosintetico fosfolipide-dipendente e alle variazioni delle
concentrazioni di calcio di alcuni enzimi implicati nella loro produzione (Alger BE.
et al., 2011).
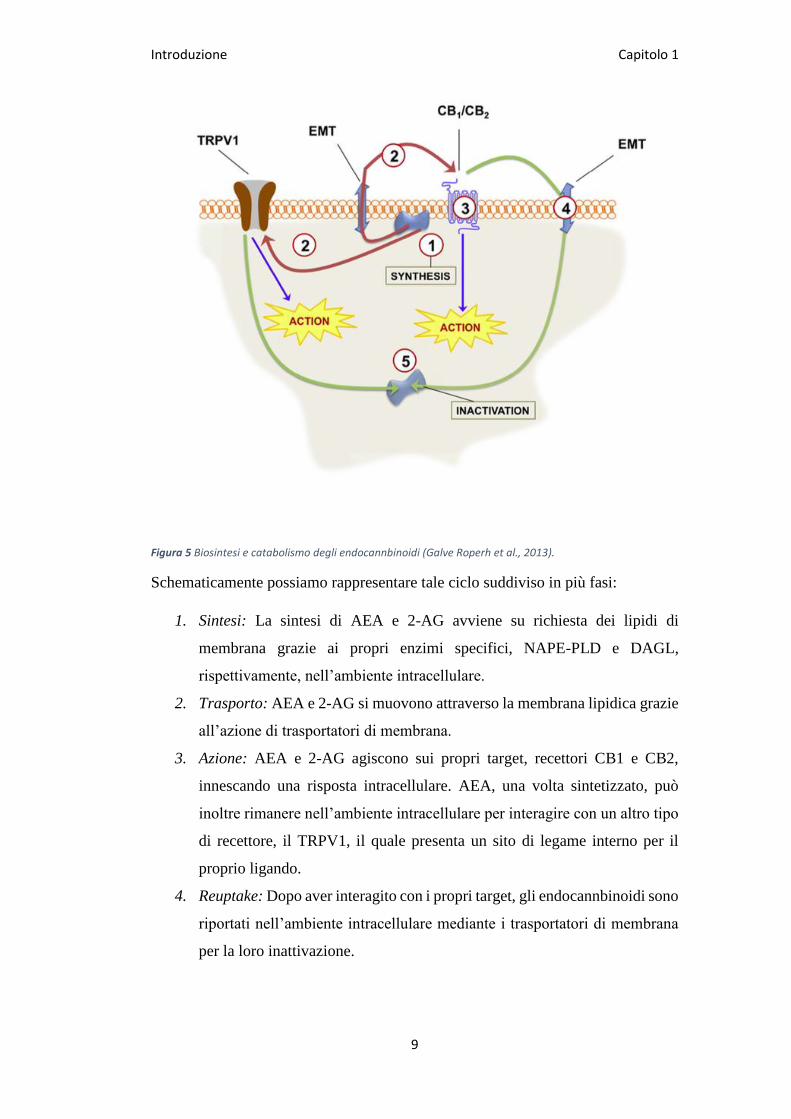
Introduzione Capitolo 1
9
Figura 5 Biosintesi e catabolismo degli endocannbinoidi (Galve Roperh et al., 2013).
Schematicamente possiamo rappresentare tale ciclo suddiviso in più fasi:
1. Sintesi: La sintesi di AEA e 2-AG avviene su richiesta dei lipidi di
membrana grazie ai propri enzimi specifici, NAPE-PLD e DAGL,
rispettivamente, nell’ambiente intracellulare.
2. Trasporto: AEA e 2-AG si muovono attraverso la membrana lipidica grazie
all’azione di trasportatori di membrana.
3. Azione: AEA e 2-AG agiscono sui propri target, recettori CB1 e CB2,
innescando una risposta intracellulare. AEA, una volta sintetizzato, può
inoltre rimanere nell’ambiente intracellulare per interagire con un altro tipo
di recettore, il TRPV1, il quale presenta un sito di legame interno per il
proprio ligando.
4. Reuptake: Dopo aver interagito con i propri target, gli endocannbinoidi sono
riportati nell’ambiente intracellulare mediante i trasportatori di membrana
per la loro inattivazione.

Introduzione Capitolo 1
10
5. Inattivazione: AEA viene idrolizzato dal FAAH in etanolammina ed acido
arachidonico, mentre il 2-AG è idrolizzato da MAGL, ed in minor parte
anche da FAAH, in glicerolo ed acido arachidonico.
1.1.4 Ruolo fisiologico
Il Sistema endocannabinoide risulta essere coinvolto in diversi processi fisiologici.
A livello del sistema nervoso centrale, gli endocannabinoidi intervengono nella
regolazione delle funzioni cognitive e delle emozioni nei circuiti neuronali della
corteccia, dell’ippocampo e dell’amigdala. Modulano il controllo dei movimenti e
della postura, la percezione delle sensazioni dolorifiche ed inoltre detiene funzioni
a livello gastrointestinale, cardiovascolare e respiratorio. Infine, attraverso
meccanismi di cross-talk con gli ormoni steroidei ed ipotalamici, gli
endocannabinoidi possono agire sull’assunzione di cibo e sulla riproduzione.
Parallelamente a tali funzioni fisiologiche, gli endocannabinoidi sembrano essere
implicati in diverse situazioni patologiche. E’ stata infatti riscontrata la presenza
dei recettori cannabinoidi sulle cellule del sistema immunitario, permettendo così
di ipotizzare un loro ruolo nel dolore cronico e nella neuroinfiammazione. Inoltre,
sono stati riscontrati alti livelli di endocannabinoidi in modelli sperimentali di
malattie neurodegenerative come il morbo di Parkinson, disordini gastrointestinali,
malattie metaboliche e disturbi del comportamento alimentare, come obesità ed
anoressia.
1.1.5 Sistema endocannabinoide e cancro
Il coinvolgimento del sistema endocannabinoide in processi tumorali è una
questione molto complicata che ad oggi rimane ancora aperta e necessita ulteriori
studi per quanto riguarda i meccanismi attraverso cui esso agisce; è stato comunque
dimostrato come questo sistema possa rappresentare un target significativo per
varie terapie in quanto, con i propri recettori, sembra essere coinvolto in processi
proliferativi e di migrazione delle cellule tumorali (Van Dross R. et al.,2013), in
particolare sembra che, vari tipi di neoplasie, presentino una regolazione alterata

Introduzione Capitolo 1
11
del sistema endocannabinoide. Molti studi stanno cercando di capire l’esistente
correlazione tra i livelli di espressione dei recettori cannabinoidi e la malignità
associata alle diverse tipologie di tumori; infatti è stato dimostrato come in alcuni
tumori, per esempio quello pancreatico e quello prostatico, elevati livelli dei
recettori CB1 e CB2, siano indice di un’elevata aggressività del tumore e quindi di
una precoce mortalità sui soggetti; nell’adenocarcinoma del colon, invece, non è
stata riscontrata alcuna correlazione, al contrario, sembra sia presente un
decremento dell’espressione di tali recettori(Daniel J. et al., 2011).
I cannabinoidi sembrano esercitare numerosi effetti dipendenti dalla linea cellulare
o dal tipo di tumore, ma la cosa interessante è il fatto che numerosi studi abbiano
riscontrato che, un’interazione con i propri recettori, determini un inibizione della
crescita del tumore attraverso diverse vie di segnale. Ciò è stato osservato in
differenti tipi di tumori quali glioma, glioblastoma, tumore al seno, cancro alla
prostata, tumore tiroideo, carcinoma del colon, leucemia e tumori ai linfoniodi. Una
scarsa regolazione del sistema endocannabinoide può promuovere la formazione di
un tumore sotto determinati tipi di condizioni fisiologiche che incrementano la
proliferazione e la migrazione delle cellule tumorali.
Differenti meccanismi sono implicati nell’azione anti-tumurogenica degli
endocannabinoidi e comprendono effetti citotossici e citostatici determinati dalla
modulazione della regolazione del normale ciclo cellulare, induzione dell’apoptosi
ed effetti anti-metastatici come inibizione della neo-angiogenesi e migrazione delle
cellule tumorali. Nello specifico, è stato visto, come i composti cannabinoidi siano
capaci di determinare un arresto del ciclo cellulare, in varie fasi, a seconda della
linea cellulare tumorale, per esempio in linee di tumori al seno è stato osservato un
arresto del ciclo cellulare in fase S, mentre in linee di glioblastoma, l’arresto,
avviene in fase di transizione G1/G0. Per quanto riguarda il fenomeno di induzione
apoptotica, invece, sembra dovuto all’inibizione di varie vie di segnale coinvolte
nella sopravvivenza delle cellule e che portano ad una down-regulation di alcune
proteine anti-apoptotiche, come la survivina. Ciò è stato evidenziato in alcuni studi
su glioma, leucemia e tumore pancreatico, utilizzando dei composti agonisti dei

Introduzione Capitolo 1
12
recettori cannabinoidi, i quali ne determinavano la loro attivazione e un
conseguente incremento della sintesi ex-novo di ceramide, un lipide pro-apoptotico.
Infine, altri tipi di studi, hanno osservato elevati livelli di AEA e 2-AG in differenti
tipi di tumori come glioblastoma, meningioma, carcinoma della prostata e del colon
e sarcoma endometriale. Tali livelli sono regolati dai corrispondenti enzimi che
sintetizzano e metabolizzano questi endocannabinoidi, perciò è stata valutata anche
la loro presenza, soprattutto in cancro alla prostata, ed è stato osservato un
incremento dell’espressione di FAAH in questo tessuto tumorale piuttosto che nello
stesso tessuto in condizioni sane. L’iperespressione di questi enzimi deputati al
catabolismo degli endocannabinoidi sta a significare una riduzione di questi ultimi
che non agiscono più sui propri recettori e quindi un aumento della proliferazione
cellulare e avanzamento del tumore stesso.
Per tutte queste ragioni, il sistema endocannabinoide, può rappresentare un
importante target per interventi farmacologici mirati al trattamento del cancro
(Daniel J., 2011).
1.2 Melanoma cutaneo
1.2.1 Definizione di melanoma
Il melanoma è una neoplasia maligna che origina da una crescita incontrollata dei
melanociti, cellule che sono deputate alla produzione del pigmento melanina (Fig.
6). Come mostrato in figura, tali cellule si trovano a livello della membrana basale
che separa l’epidermide dal derma sottostante e producono melanina che viene
distribuita alle cellule squamose e basali che compongono l’epidermide.
Il melanoma può originare a partire da un neo, dando così origine al melanoma
della pelle, ma anche da altri tipi di tessuti pigmentati nell’occhio o nell’intestino
(National Cancer Institute, Melanoma, 2014). Per la sua malignità è chiamato anche
melanoma maligno, melanocarcinoma, melanoepitelioma, melanosarcoma
(Merriam-Webster, 2014).
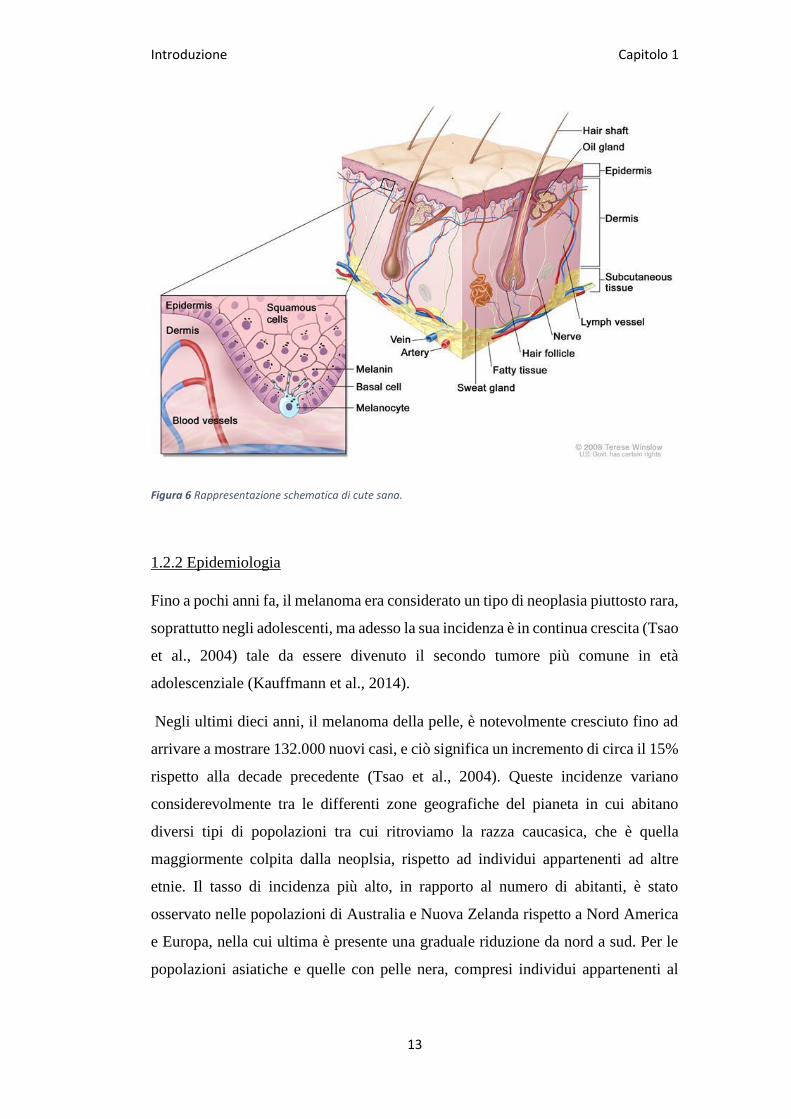
Introduzione Capitolo 1
13
Figura 6 Rappresentazione schematica di cute sana.
1.2.2 Epidemiologia
Fino a pochi anni fa, il melanoma era considerato un tipo di neoplasia piuttosto rara,
soprattutto negli adolescenti, ma adesso la sua incidenza è in continua crescita (Tsao
et al., 2004) tale da essere divenuto il secondo tumore più comune in età
adolescenziale (Kauffmann et al., 2014).
Negli ultimi dieci anni, il melanoma della pelle, è notevolmente cresciuto fino ad
arrivare a mostrare 132.000 nuovi casi, e ciò significa un incremento di circa il 15%
rispetto alla decade precedente (Tsao et al., 2004). Queste incidenze variano
considerevolmente tra le differenti zone geografiche del pianeta in cui abitano
diversi tipi di popolazioni tra cui ritroviamo la razza caucasica, che è quella
maggiormente colpita dalla neoplsia, rispetto ad individui appartenenti ad altre
etnie. Il tasso di incidenza più alto, in rapporto al numero di abitanti, è stato
osservato nelle popolazioni di Australia e Nuova Zelanda rispetto a Nord America
e Europa, nella cui ultima è presente una graduale riduzione da nord a sud. Per le
popolazioni asiatiche e quelle con pelle nera, compresi individui appartenenti al
Introduzione Capitolo 1
14
ceppo ispanico (nativi americani e popolazioni dell’Africa, eccetto Sud-Africa), il
tasso di incidenza diminuisce notevolmente (Globacan 2012, 2014).
Figura 7 Percentuali di casi stimati di melanoma cutaneo per tutte le età e per entrambi i sessi (Globacan 2012, 2014).
Nel 2014, negli Stati Uniti, sono stati diagnosticati circa 76.100 casi di melanoma,
dei quali 9.700 sono deceduti (Siegle et al.,2014); invece, in Italia, si stimano
11.000 nuovi casi con una piccola predominanza maschile (Aiom-airtum, 2014). E’
stato inoltre osservato come il melanoma stia rapidamente aumentando nell’uomo
a differenza di altri tumori maligni (Fig.8) (Jemal et al., 2011) e, alcuni studi
statistici, hanno evidenziato che i siti d’insorgenza di tale neoplasia, in ordine di
percentuale decrescente, sono: il tronco (43.5%), le estremità (33.9%), i siti sacrali,
quali siti sublinguali e palme (11.9%), testa e collo (10.7%).
Il melanoma può presentarsi in persone di qualsiasi età, ma principalmente negli
adulti che detengono un’età media al momento della diagnosi e con una successiva
e conseguente mortalità che si aggira tra i 61 e i 68 anni (Weinstock, 2012).
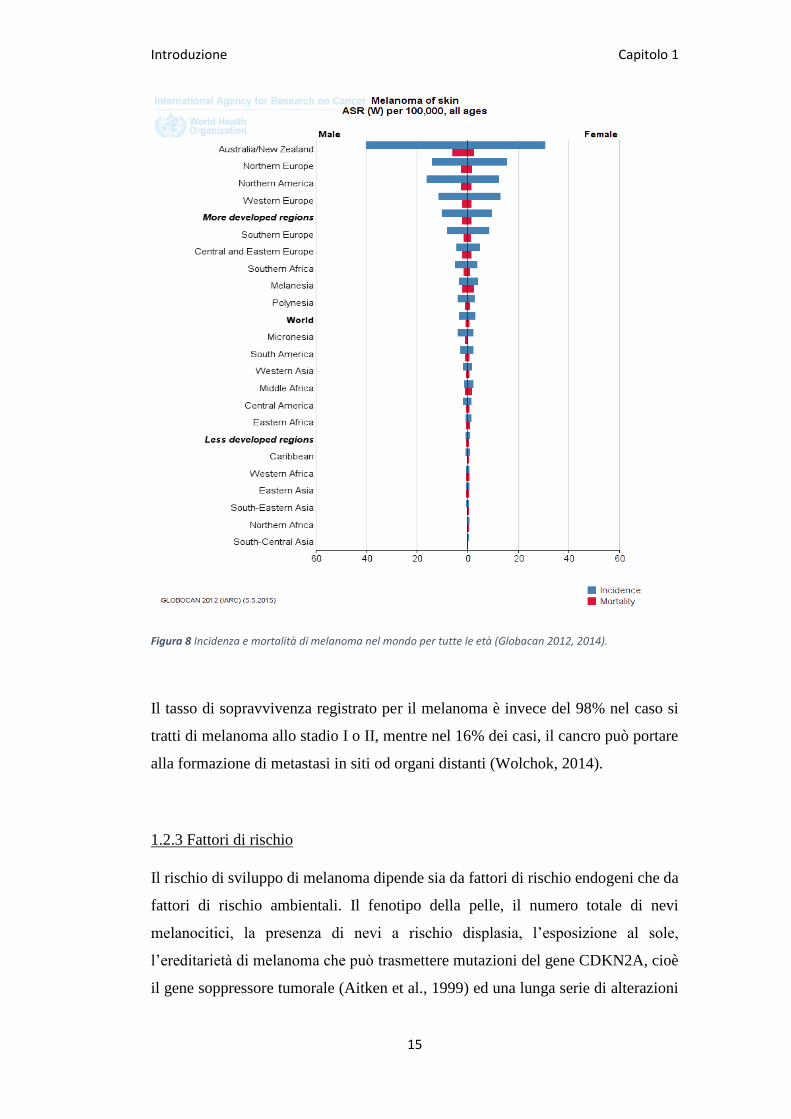
Introduzione Capitolo 1
15
Figura 8 Incidenza e mortalità di melanoma nel mondo per tutte le età (Globacan 2012, 2014).
Il tasso di sopravvivenza registrato per il melanoma è invece del 98% nel caso si
tratti di melanoma allo stadio I o II, mentre nel 16% dei casi, il cancro può portare
alla formazione di metastasi in siti od organi distanti (Wolchok, 2014).
1.2.3 Fattori di rischio
Il rischio di sviluppo di melanoma dipende sia da fattori di rischio endogeni che da
fattori di rischio ambientali. Il fenotipo della pelle, il numero totale di nevi
melanocitici, la presenza di nevi a rischio displasia, l’esposizione al sole,
l’ereditarietà di melanoma che può trasmettere mutazioni del gene CDKN2A, cioè
il gene soppressore tumorale (Aitken et al., 1999) ed una lunga serie di alterazioni

Introduzione Capitolo 1
16
genetiche (Gudbjartsson et al., 2008), possono aumentare lo stato di
immunodeficienza dell’individuo rappresentando fattori di rischio personali.
Questi fattori (Tab.1) sembra siano responsabili dell’aumento dell’incidenza del
melanoma cutaneo a livello globale (Gandini et al., 2005).
Tabella 1. Fattori di rischio per melanoma cutaneo (Helfand et al., 2001).
Per ridurre l’incidenza e la mortalità causata dal melanoma è importante anche
valutare e riconoscere l’esistenza di diversi gruppi di individui che detengono, per
diverse caratteristiche, differenti livelli di rischio; perciò è fondamentale il fattore
prevenzione (Tsao et al., 2004). Prendendo il singolo soggetto, gli elementi di
maggior interesse predisponenti alla malattia sono la presenza di numerose
lentiggini, associate spesso a carnagioni chiare che si abbronzano poco e con
tendenza all’ustione solare ed un elevato numero di nevi melanocitici e/o
displastici. Alcuni studi, hanno inoltre dimostrato come sia presente un’elevata
incidenza di melanoma in soggetti anziani e di sesso maschile (Fig.8) (Lasithiotakis
et al., 2008). Inoltre è importante anche il fattore ereditario e cioè la presenza
antecedente della stessa neoplasia in qualche familiare, questo perché possa
spingere il soggetto stesso a seguire idonee strategie di prevenzione; è stato infatti
calcolato che la probabilità di sviluppo di un secondo melanoma a distanza dai 5 ai
10 anni è del 2.8% e 3.6%, rispettivamente (Goggins et al., 2003).

Introduzione Capitolo 1
17
Da non trascurare è il fattore genetico: studi genetici molecolari hanno infatti
mostrato l’importante ruolo di due geni (CDKN2A e CDK4) (Fig.9), per i quali una
mutazione a carico può rappresentare un elevato fattore di rischio (Fargnoli et al.,
2006; Meyle et al., 2009).
Figura 9 Prevalenza di mutazioni a carico del gene CDKN2A come fattore ereditario di melanoma (Harland et al., 2014).
Tra i fattori di rischio ambientali ha un’importanza rilevante l’esposizione ai raggi
UV, la quale, generalmente, raddoppia il rischio di sviluppo di melanoma in
soggetti esposti e soprattutto nel fenotipo di persone con pelle chiara (Vainio et al.,
2000). Studi epidemiologici recenti confermano la presenza di una correlazione tra
intensa esposizione solare, che spesso porta ustioni e scottature, e lo sviluppo della
neoplasia (Gandini et al., 2005). Rimane tuttavia da identificare la componente
della radiazione solare capace di scatenare l’evento tumorale; gli UVR infatti
risultano essere suddivisi in tre bande di lunghezza d’onda diversa: UVA, UVB e
UVC. Grazie all’assorbimento da parte dell’ozonosfera, la superficie terrestre
riceve per il 99% UVA, una percentuale che va diminuendo a causa della riduzione
dello strato di ozono, portando così ad una maggiore permeazione dei raggi UVB.
Entrambe le lunghezze d’onda sono capaci di danneggiare i DNA: gli UVA, in
maniera indiretta, formando specie reattive dell’ossigeno, mentre gli UVB
direttamente inducendo la formazione di legami covalenti tra basi adiacenti. Questa

Introduzione Capitolo 1
18
loro azione e l’aumento della loro intensità, avvalora l’ipotesi di un ruolo decisivo
della radiazione solare nello sviluppo del melanoma cutaneo. Tuttavia,
considerando anche i numerosi fattori concomitanti, rimane da chiarire ancora oggi
quali siano le componenti maggiormente coinvolte ed il loro ruolo nello sviluppo
del tumore (Palmieri et al., 2009). Altri fattori di rischio ambientale possono essere
anche il contatto con agenti nocivi, come arsenico o radiazioni ionizzanti, con i
quali, spesso, il soggetto, viene a contatto per esigenze lavorative.
Insufficienze del sistema immunitario, dovute a chemioterapie, trapianti o AIDS,
e anomalie genetiche, il cui ruolo è ancora oggi materia d’indagine, rappresentano
altri fattori di rischio importanti per questo tipo di neoplasia (Markovic S.N et al.,
2007).
1.2.4 Patogenesi molecolare
Il melanoma è caratterizzato da diverse vie di segnale molecolari (signaling
pathways) coinvolte nel controllo della proliferazione e della morte cellulare.
L’iperattivazione o la riduzione dell’attività di questi componenti risultano essere
coinvolti nella trasformazione oncogenica dei melanociti. Le principali vie di
segnale alterate nel melanoma sono Rb/E2F, MAPK/ ERK, PI3K/ Akt/, NF-Kb,
Wnt/β-catenin, Notch, Jak/STAT, cyclin/CDK, JNK/c-Jun/ AP-1, MITF ed alcuni
fattori di crescita.
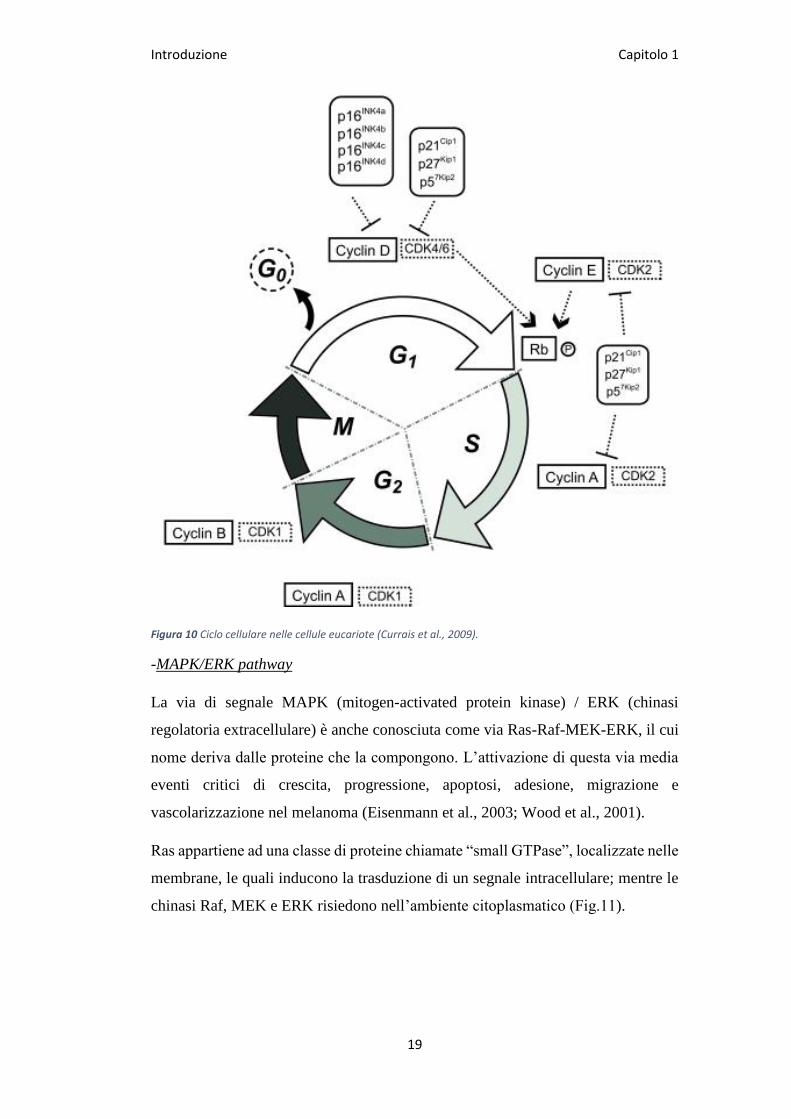
Introduzione Capitolo 1
19
Figura 10 Ciclo cellulare nelle cellule eucariote (Currais et al., 2009).
-MAPK/ERK pathway
La via di segnale MAPK (mitogen-activated protein kinase) / ERK (chinasi
regolatoria extracellulare) è anche conosciuta come via Ras-Raf-MEK-ERK, il cui
nome deriva dalle proteine che la compongono. L’attivazione di questa via media
eventi critici di crescita, progressione, apoptosi, adesione, migrazione e
vascolarizzazione nel melanoma (Eisenmann et al., 2003; Wood et al., 2001).
Ras appartiene ad una classe di proteine chiamate “small GTPase”, localizzate nelle
membrane, le quali inducono la trasduzione di un segnale intracellulare; mentre le
chinasi Raf, MEK e ERK risiedono nell’ambiente citoplasmatico (Fig.11).
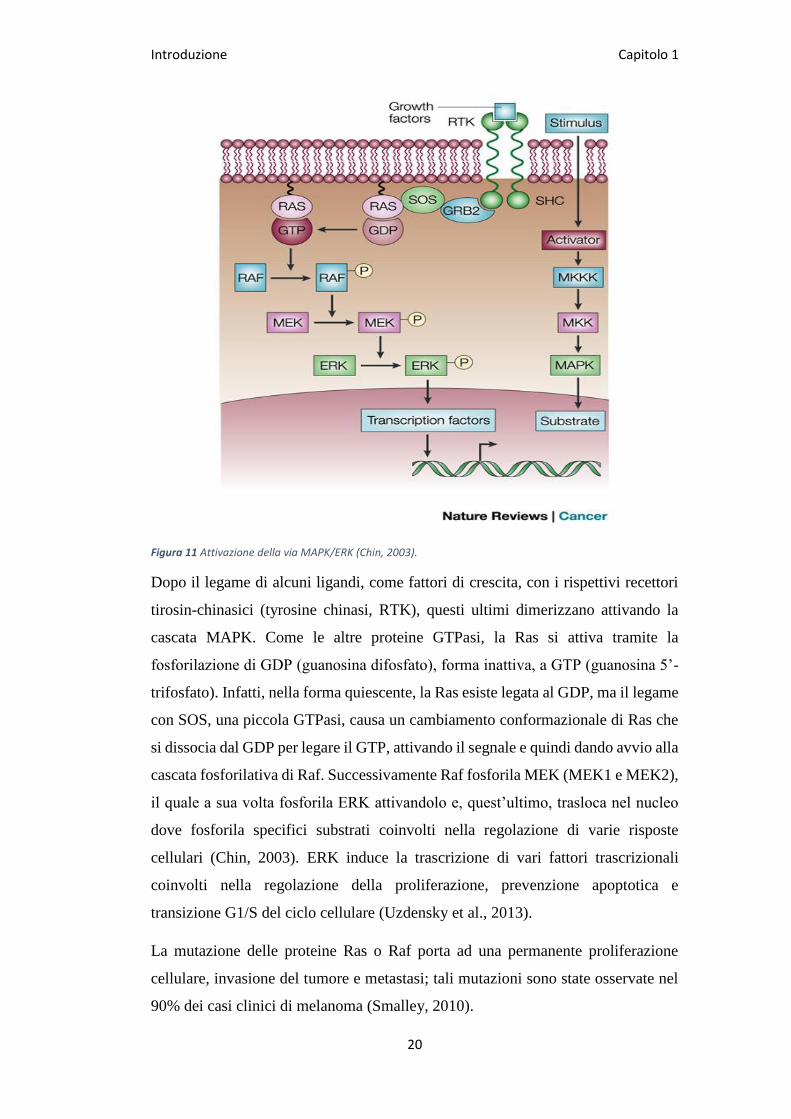
Introduzione Capitolo 1
20
Figura 11 Attivazione della via MAPK/ERK (Chin, 2003).
Dopo il legame di alcuni ligandi, come fattori di crescita, con i rispettivi recettori
tirosin-chinasici (tyrosine chinasi, RTK), questi ultimi dimerizzano attivando la
cascata MAPK. Come le altre proteine GTPasi, la Ras si attiva tramite la
fosforilazione di GDP (guanosina difosfato), forma inattiva, a GTP (guanosina 5’-
trifosfato). Infatti, nella forma quiescente, la Ras esiste legata al GDP, ma il legame
con SOS, una piccola GTPasi, causa un cambiamento conformazionale di Ras che
si dissocia dal GDP per legare il GTP, attivando il segnale e quindi dando avvio alla
cascata fosforilativa di Raf. Successivamente Raf fosforila MEK (MEK1 e MEK2),
il quale a sua volta fosforila ERK attivandolo e, quest’ultimo, trasloca nel nucleo
dove fosforila specifici substrati coinvolti nella regolazione di varie risposte
cellulari (Chin, 2003). ERK induce la trascrizione di vari fattori trascrizionali
coinvolti nella regolazione della proliferazione, prevenzione apoptotica e
transizione G1/S del ciclo cellulare (Uzdensky et al., 2013).
La mutazione delle proteine Ras o Raf porta ad una permanente proliferazione
cellulare, invasione del tumore e metastasi; tali mutazioni sono state osservate nel
90% dei casi clinici di melanoma (Smalley, 2010).

Introduzione Capitolo 1
21
La sottofamiglia di Ras comprende tre isoforme: H-Ras, K-Ras e N-Ras;
quest’ultima è maggiormente coinvolta nella patogenesi del melanoma, infatti,
mutazioni a suo carico, sono state trovate nel 15-30% dei casi (Sekulic et al., 2008).
La sottofamiglia di Raf, invece, consiste delle isoforme A-Raf, B-Raf e C-Raf
(chiamata anche Raf-1); tra queste le mutazioni a carico di Braf sono le più rilevanti
nel melanoma e ne sono state trovate nel 60-70% di casi di melanoma primario e
anche un’elevata percentuale nei nevi melanocitici cutanei. Questo è considerato un
fattore di rischio coinvolto nell’iniziazione e successivamente nella progressione
del melanoma; infatti sono state descritte più di 40 mutazioni per il gene BRAF e
tra queste la più rilevante è la BRAF T1799A, caratterizzata dalla sostituzione di
acido glutammico con valina (V600E) nel dominio chinasico della rispettiva
proteina. La proteina BRaf mutata attiva permanentemente ERK stimolando la
proliferazione cellulare Questa mutazione è stata trovata nell’80-90% delle
mutazioni a carico di BRAF nel melanoma (Platz et al., 2008).
1.3 Cellule staminali tumorali di melanoma
Le cellule staminali tumorali (CSC “Cancer stem cells”) sono una tipologia di
cellule responsabili non solo della formazione del tumore stesso, ma anche del suo
mantenimento e progressione, inoltre sono anche ritenute responsabili dei fenomeni
di resistenza a vari agenti tossici (Girouard et al., 2011). A causa di tali proprietà,
le CSC che sopravvivono in chemioterapia, sono in grado di ristabilire di nuovo il
tumore (Emma L. et al., 2015).
In origine, il tumore, viene avviato da una normale cellula staminale che trasforma
il proprio fenotipo in neoplastico ed assume le seguenti proprietà:
-Capacità di autorinnovamento costituendo così una “popolazione immortale”;
-Danno origine a cellule derivate (non-CSC) che possono dividersi un numero
limitato di volte dando luogo alla massa tumorale;
-sono resistenti alla chemioterapia standard (Roger S., 2015).

Introduzione Capitolo 1
22
Le cellule staminali tumorali di melanoma, chiamate anche cellule iniziatrici di
melanoma (Fig.12) e per la loro morfologia sfere, sono cellule capaci di
autorinnovarsi cioè di formare altre cellule multipotenti e dare origine, sotto
opportuni stimoli, alla formazione ed alla progressione del melanoma (Dong Fang
et al., 2005).
Figura 12 Cellule iniziatrici di melanoma (MMICs) (Lee et al., 2014).
La chemioterapia e/o la radioterapia utilizzate per debellare il tumore, risultano
essere funzionali solo sulle cellule coinvolte nella proliferazione del tumore stesso,
ma non sulle CSC che sono più resistenti e possono far progredire e dare un nuovo
avvio al tumore stesso (Schatton et al., 2008). Le biopsie di melanoma hanno
rivelato come questo contenga dall’1 al 20% di cellule staminali tumorali (Na et al.,
2009; Schatton et al., 2008). Inoltre nel 2008 Quintana con i suoi collaboratori
hanno dimostrato che 1 cellula di melanoma su 4 può iniziare un nuovo tumore in
animali con immuno-deficienza e più del 25% delle cellule di melanoma sono
potenzialmente tumorogeniche. Pertanto lo sviluppo futuro di nuove terapie contro
il melanoma mira alla ricerca di specifici marker espressi selettivamente in cellule
iniziatrici di melanoma.
1.3.1 Cellule staminali tumorali e sistema endocannabinoide
Il sistema endocannabinoide sembra essere coinvolto in meccanismi di regolazione
appartenenti alle cellule iniziatrici/staminali nel sistema nervoso centrale (Galve et

Introduzione Capitolo 1
23
al., 2013) e nella differenziazione delle cellule staminali di glioma (Compagnucci
et al., 2013).
Altri studi hanno dimostrato la correlazione tra ECS e cellule staminali dimostrando
come il trattamento con fitocannabinoidi, possa arrestare lo sviluppo dei primi
embrioni in blastociti, probabilmente attraverso un meccanismo CB1-dipendente
(Paria et al.,1998; Nones J. et al., 2010). Tuttavia, il blocco farmacologico dei
recettori CB1 e CB2 risulta indurre morte delle cellule staminali, in quanto il
sistema endocannabinoide è coinvolto nei meccanismi di sopravvivenza di queste
ultime (Jiang S. et al., 2007). Su queste basi è stato ipotizzato che la presenza dei
recettori CB1 e CB2, indice della presenza del sistema endocannabinoide, a livelli
basali, sia collegata alla sopravvivenza delle cellule staminali mentre la presenza
del sistema endocannabinoide ad elevate concentrazioni innesca una cascata
autofagica a livello embrionale (Oh HA et al., 2013).

Scopo della Ricerca Capitolo 2
24
Capitolo 2
Scopo della ricerca
Lo scopo di questo progetto è stato quello di valutare la presenza ed il significato
del sistema endocannabinoide (con i suoi recettori CB1 e CB2 e gli enzimi deputati
alla sintesi ed al metabolismo degli endocannabinoidi endogeni AEA e 2-AG) in
melanociti, linee cellulari di melanoma, cellule isolate da singolo paziente e in
cellule staminali tumorali. In particolare abbiamo valutato il coinvolgimento del
sistema endocannabinoide nello sviluppo del fenotipo neoplastico ed il possibile
cross-talk con la via B-RAF, coinvolta nella trasmissione del segnale di
proliferazione, in cellule portatrici o meno della mutazione attivante V600E.

Materiali e metodi Capitolo 3
25
CAPITOLO 3
Materiali e metodi
3.1 Materiali per gli studi funzionali
3.1.1 Linee cellulari
A375
Le A375 (American Type Culture Collection, ATCC, Rockville; MA, USA) sono
cellule di melanoma cutaneo umano che crescono adese alla superficie della fiasca,
aderendo al pavimento di poli-lisina ed hanno un aspetto allungato. La linea
cellulare è caratterizzata da un tempo di duplicazione di circa 16-20 ore (Goodall,
Carreira et al. 2008 Cancer research). Le cellule sono state mantenute in coltura in
incubatore con il 5% di CO2 a 37°C in un terreno costituito dal mezzo base RPMI
1640 (Rosewell Park Memorial Institute 1640 Medium) ricostituito con 10% di siero
fetale bovino (FBS, Life Technologies, Monza) e 1% della miscela 1:1 degli
antibiotici penicillina (50 UI/ml) e streptomicina (50 µg/ml).
Figura 13. Cellule A375 a bassa densità (sinistra) ed alta densità (destra) di crescita [Sito ATCC: http://www.lgcstandards-atcc.org/~/media/Attachments/4/9/9/0/1823.ashx].

Materiali e metodi Capitolo 3
26
MeWo
Le MeWo sono cellule derivanti da cellule di metastasi linfonodali di melanoma
cutaneo maligno umano e crescono aderendo al pavimento della fiasca con un
tempo di replicazione di circa 30 ore. Tali cellule sono state mantenute in coltura in
incubatori a 37°C con il 5% di CO2 in un terreno costituito dal mezzo base di RPMI
1640 ricostituito con il 10% di FBS e 1% degli antibiotici Penicillina e
Streptomicina.
Figura 14. MeWo a bassa densità (sinistra) e ad alta densità di crescita (destra). [http://cellbank.nibio.go.jp/legacy/celldata/jcrb0066.htm#reference].
501 Mel
Le 501Mel sono una linea maligna di melanoma cutaneo umano e crescono adese
al pavimento della fiasca con un tempo di replicazione di circa 40 ore. Vengono
mantenute in coltura in incubatori a 37°C con il 5% di CO2 in un terreno di coltura
costituito dal mezzo base Dulbecco’s Modified Medium (DMEM, Sigma Aldrich,
Milano) ricostituito con il 10% di FBS e 1% della miscela di antibiotici penicillina
e streptomicina.

Materiali e metodi Capitolo 3
27
Melanociti NHEM
I melanociti (NHEM primary normal human epidermal melanocytes PromoCell
GmbH, Germany) sono cellule umane sane isolate dell’epidermide di donatori sani,
giovani o adulti, derivanti da differenti locazioni tra le quali viso, seno, addome e
dita. Si trovano nello strato basale, ma si diramano tra i cheratinociti in strati
sovrastanti. Circa il 5-10% delle cellule dell’epidermide sono melanociti ed hanno
la funzione principale di produrre la melanina, la proteina responsabile per la
pigmentazione della pelle, occhi e capelli. Essa protegge le cellule della pelle e, in
profondi strati, esercita tale funzione contro gli effetti pericolosi delle radiazioni
UV (http://www.promocell.com/products/human-primary-cells/melanocytes).
Queste cellule crescono in incubatore a 37°C con 5% di CO2 nel proprio mezzo di
coltura specifico, Melanocyte Growth Medium M2 (PromoCell GmbH, Germany),
ed hanno un tempo di replicazione di 7-14 giorni.
Figura 15 Melanociti NHEM (http://www.promocell.com/products/human-primary-cells/melanocytes).
3.1.2 Soluzioni e tamponi utilizzati
RPMI
Il Rosewell Park Memorial Institute 1640 Medium (RPMI, Life Technologies,
Monza) è il terreno di coltura usato per consentire la crescita e la proliferazione
delle cellule A375 e MeWo. Il mezzo utilizza un sistema tampone con bicarbonato
e come indicatore rosso fenolo che per valori di pH intorno a 7.4, in cui le cellule
crescono bene, presenta un colore rosso-arancio che vira al giallo quando, in seguito
a proliferazione cellulare, si ha acidificazione del mezzo per produzione della CO2
proveniente dal metabolismo cellulare. Assume, invece, una colorazione violacea

Materiali e metodi Capitolo 3
28
in presenza di pH alcalino indicando che le cellule non sono metabolicamente attive
o che la regolazione della CO2 è alterata.
DMEM
Il Dulbecco's Modified Medium (DMEM, Sigma-Aldrich, Milano) è il terreno di
coltura usato per consentire la crescita e la proliferazione delle cellule 501Mel.
Contiene amminoacidi, vitamine, sali, glucosio, glutammina e, come indicatore di
pH, il rosso fenolo. Per valori di pH intorno a 7.4, in cui le cellule crescono bene,
presenta un colore rosso-arancio che vira al giallo quando, in seguito a
proliferazione cellulare, si ha acidificazione del mezzo per produzione della CO2
proveniente dal metabolismo cellulare. Assume, invece, una colorazione violacea
in presenza di pH alcalino indicando che le cellule non sono metabolicamente attive
o che la regolazione della CO2 è alterata.
Melanocyte Growth Medium M2
Il mezzo M2 (Promocell GmbH, Germany) è il mezzo di coltura utilizzato per il
mantenimento e la crescita dei melanociti umani, privo di siero e privo di PMA
(Phorbol Myristate Acetate). Tale mezzo contiene tutti i fattori di crescita e
supplementi necessari per la crescita di tali melanociti ad eccezione di antibiotici
ed antimicotici. E’ stato formulato per essere utilizzato in incubatore a 37°C e 5%
di CO2.
FBS
Il fetal bovine serum (FBS, Life Technologies, Monza) è costituito da fattori di
crescita e proteine che hanno il compito di facilitare la sopravvivenza, la crescita e
la divisione cellulare e per questo viene addizionato al mezzo.
Penicillina/Streptomicina
La miscela 1:1 degli antibiotici penicillina (50 UI/ml) e streptomicina (50µg/ml)
(Penicillina/Streptomicina, Sigma Aldrich, Milano) viene usata per evitare la
proliferazione batterica all’interno della coltura cellulare.
Tripsina
La miscela tripsina-EDTA (Trypsin-Versene (EDTA) Mix 10X, Life Technologies,
Monza) è una soluzione contenente una proteasi, la tripsina, capace di produrre tagli

Materiali e metodi Capitolo 3
29
proteolitici a livello del legame tra l’arginina (membrana cellulare) e la lisina
(supporto trattato) permettendo il distacco delle cellule. Tale funzione è sfruttata
per degradare le proteine della matrice su cui le cellule crescono in adesione. Nella
soluzione è presente anche EDTA, un agente chelante, aggiunto per migliorare
l’attività proteolitica della tripsina poiché, legando cationi come calcio e magnesio
presenti nell’ambiente extra-cellulare, gli impedisce di nascondere i legami
peptidici su cui agisce la tripsina.
PBS
Il phosfate saline buffer (PBS, Sigma Aldrich, Milano) è un tampone costituito da
una soluzione salina acquosa contenente cloruro di sodio, cloruro di potassio e
fosfato di sodio. La sua funzione è quella di mantenere i valori di pH costanti e
isotonici. Non essendo tossico per le cellule, viene usato per i lavaggi delle colture
cellulari con lo scopo di eliminare i detriti.
3.2 Materiali per gli studi di biologia molecolare
3.2.1 Kit e reagenti
L’estrazione dell’RNA totale dalle cellule è stata eseguita con RNeasy MiniKit
(Quiagen, Milano) che comprende: RNeasy Mini Spin Columns, Collection Tubes
da 1,5 e 2 ml, acqua RNase-Free e buffer (buffer RLT, buffer RW1 e buffer RPE).
La retrotrascrizione è stata eseguita utilizzando il kit QuantiTect Reverse
Transcription Kit (Quiagen, Milano), comprendente gDNA Wipeout Buffer 7x,
QuantiscriptR, Reverse Transcriptase, Quantiscript RT Buffer 5x, RT Primer Mix e
acqua RNase-Free.
Le reazioni di PCR sono state effettuate usando HotStarTaq® PCR (Quiagen,
Milano). Il Kit è costituito da: PCR Buffer 10x, HotStarTaq MasterMix, acqua
RNase-Free.
Il gel per la corsa elettroforetica è stato ottenuto utilizzando agarosio (EuroClone,
Milano) e tampone TBE 0,5x contenente acido borico 0,9M, EDTA 0,01M, Tris

Materiali e metodi Capitolo 3
30
1M (Sigma-Aldrich, Milano). Ladder 100pb e blu di Bromofenolo-xilene sono stati
forniti da Sigma Aldrich (Milano).
La Real-Time PCR è stata effettuata usando due tipi di sonde: la sonda SsoFast™
Eva GreenR Supermix (Bio-Rad, Hercules, USA), e le sonde TaqMan®Gene
Expression Assays (Applied Biosystems®, Life Technologies) dirette verso CB1R,
CB2R e 18S.
Oltre ad ottenere i cDNA mediante retrotrascrizione degli mRNA ottenuti dalle
rispettive cellule in coltura, ci sono stati forniti i cDNA di linee cellulari isolate da
singolo paziente nell’ambito di una collaborazione con il laboratorio di Biologia
della Cellula Tumorale, Core Research Laboratory, Istituto Toscano Tumori
(Firenze) diretto dalla dott.ssa Barbara Stecca (Tab.2).
Tabella 2. Cellule isolate da singolo paziente (Stecca et al., 2013). .
3.2.2 Primer
I primer sono oligonucleotidi sintetici che hanno il compito di innescare la sintesi
del nuovo filamento di cDNA durante la fase di amplificazione delle real-time PCR.
Per ogni sequenza bersaglio serve una coppia di primer, il forward (complementare
al filamento in direzione 3’-5’) ed il reverse (complementare al filamento in
direzione 5’-3’). I primer del nostro oggetto di studio sono stati scelti facendo
riferimento a sequenze presenti in letteratura. Le sequenze scelte sono state
analizzate utilizzando il programma “Primer Blast”
(http://www.ncbi.nlm.nih.gov/tools/primer-blast/index.cgi), un programma che
permette di valutare la specificità di ogni coppia per il target ed evitare la
formazione di prodotti di amplificazione genica indesiderati. Le sequenze dei
primer utilizzati sono riportate in Tabella 3. GAPDH e β-actina sono geni

Materiali e metodi Capitolo 3
31
housekeeping, cioè geni costitutivamente trascritti e tradotti poiché codificano
proteine ed enzimi fondamentali per la vita della cellula.
Tabella 3.Sequenza dei primer utilizzati in RT-PCR e real-time PCR; primer forward (F) e primer reverse (R).
Per quanto riguarda le sequenze delle sonde TaqMan CB1 e CB2, queste ci sono
state fornite dalla parte ThermoFisher Scientific di Life Technologies, i quali ne
sono i diretti proprietari, quindi l’intera sequenza è segreta e non può essere fornita
al cliente. Quello che possiamo fare è controllare dove sia disegnato il saggio,
utilizzando i dati provenienti dall’Assay Location” (nella tabella “View Details”
per ognuno dei saggi), che dà la posizione, la base centrale su cui si appaia la sonda,
la lunghezza del prodotto e con questi dati ricostruire la sequenza di legame.
-CNR1(esempio)
Prodotto di amplificazione: 113 pb
Assay location: 5576 (che significa la posizione 5576 della sequenza di riferimento
di GeneBank).
Quindi, considerando la metà della lunghezza del prodotto di amplificazione, conto
56 pb a monte e a valle della sequenza, e quindi troviamo la regione di riferimento
riconosciuta dal saggio: 5576-56=5520; 5576+56=5632 (dalla 5520esima alla
5632esima base della sequenza di riferimento).
Materiali e metodi Capitolo 3
32
Tabella 4. Sonde TaqMan utilizzate in Real-Time PCR.
3.3 Materiali per Western blotting
SDS
Il sodio dodecil sulfate (SDS) è un detergente anionico che, rompendo i legami ad
idrogeno e le interazioni idrofobiche, provoca la distruzione delle strutture
secondarie e terziarie delle proteine. L’utilizzo di questo detergente rende costante
il rapporto massa/carica permettendo la separazione proteica in base al peso
molecolare. E’ usato in soluzione al 10% in di acqua milliQ.
Soluzione di acrilammide
L’acrylamide-solution al 30% (Euroclone spa, Italia) è costituita da monomeri di
acrilammide (l’ammide dell’acido acrilico) e da piccole quantità di N,N’-
metilenbisacrilammide, più comunemente nota con il nome di bis-acrilammide.
Nello specifico, quest’ultima è un composto essenzialmente formato da due
molecole di acrilammide legate da un gruppo metilenico.
Catalizzatori: TEMED e APS
La tetrametilendiammina (TEMED, Bio-Rad, CA, USA) ed il persolfato di
ammonio (APS, Bio-Rad, CA, USA) sono due catalizzatori utilizzati per
velocizzare il meccanismo di formazione dei gel di poliacrilamide.
Specificatamente, il TEMED catalizza la decomposizione dello ione persolfato
portando alla formazione di un radicale libero che reagisce con un monomero di
acrilammide, generando così un nuovo radicale libero, che reagirà con un’ulteriore
molecola di acrilammide.

Materiali e metodi Capitolo 3
33
Anticorpi
Gli anticorpi primari utilizzati sono: un anticorpo monoclonale di coniglio diretto
verso l’intera lunghezza del recettore CB1 umano (Cod.172970, Abcam,
Cambridge, Regno Unito), un anticorpo monoclonale di topo diretto verso la β-
actina di origine umana (Cod. MAB1501, Merck Millipore, Darmstadt, Germany)
e sempre un anticorpo monoclonale di topo diretto verso la proteina BRAF (Cod.
sc-5284, Sanya Cruz Biotechnology).
I rispettivi anticorpi secondari sono anticorpi coniugati con enzima HRP
(Horseradish peroxidase) anti-rabbit (Cod.28603588, Millipore S.A.S, Molsheim)
e anti-mouse (Cod.sc-5284, Santa Cruz Biotechnology). Questo enzima è capace di
amplificare un segnale debole aumentando così la rilevabilità di una molecola
bersaglio. La sua presenza è resa visibile con l’utilizzo di un substrato, il quale viene
ossidato dall’HRP determinando dei cambiamenti rilevabili con metodi
spettrofotometrici. In questo studio il substrato che viene convertito è il Luminolo
in condizioni di Chemiluminescenza (ECL, Enhanced Chemiluminescence, Santa
Cruz Biotechnology) producendo, al termine della reazione una luce rilevabile.
3.4 Metodi utilizzati negli studi funzionali
3.4.1 Scongelamento della linea cellulare
Lo scongelamento è la procedura necessaria per riportare le cellule in uno stato
metabolico attivo dopo che sono state conservate congelate in azoto liquido o in
congelatore a -80°C per un determinato periodo di tempo.
Il criotubo viene agitato meccanicamente a mano con moto rotativo per facilitare lo
scongelamento, visibile per formazione di una soluzione liquida all’interno. A
questo punto il contenuto viene aspirato ed introdotto in una provetta sterile da 15
ml aggiungendo 5-6 ml di mezzo di coltura completo, preventivamente riscaldato a
37° C a bagno maria, agitando completamente la provetta in modo da equilibrare la
sospensione cellulare con il mezzo. La soluzione contenente le cellule viene quindi
risospesa e poi lasciata riposare a temperatura ambiente per qualche minuto prima
di centrifugare la provetta a 1100 rpm per 5 minuti. A centrifugazione ultimata, il
sovranatante viene eliminato mediante aspirazione, mentre il pellet cellulare viene

Materiali e metodi Capitolo 3
34
risospeso in 3-4 ml di mezzo completo. La sospensione cellulare viene poi trasferita
in una fiasca per coltura contenente circa 5 ml di mezzo completo arrivando ad un
totale di 9-10 ml di contenuto e posta nell’incubatore (37°C, 5% CO2).
3.4.2 Mantenimento in coltura
La linea cellulare è mantenuta in una fase di crescita esponenziale in fiasche per
colture cellulari T75 (Starstedt, Verona, Italia) all’interno dell’incubatore. Le
cellule vengono controllate quotidianamente: il mezzo di coltura consumato,
arricchito di sostanze di scarto del metabolismo cellulare che causano il viraggio
dell’indicatore acido-base dal rosso al giallo, viene regolarmente sostituito con
mezzo fresco.
Quando le cellule raggiungono circa un 80% di confluenza, un’ulteriore crescita
potrebbe comportare la formazione di multistrati cellulari che si sovrappongono
l’uno sull’altro.
Per evitare tale condizione, tutto il mezzo di coltura è aspirato dalla fiasca e viene
eseguito un lavaggio con 5-6 ml di tampone fosfato (PBS, Sigma Aldrich, Milano)
per pulire la superficie di crescita da eventuali detriti rimasti. Rimosso il tampone,
si aggiunge 1 ml di Tripsina/EDTA (Sigma Aldrich, Milano) diluita in 1 ml di
tampone PBS per staccare le cellule dalla superficie di crescita. La fiasca viene
portata nell’incubatore all’incirca per 1 minuto, in modo da attivare l’azione della
tripsina.
Dopo aver verificato al microscopio che le cellule si siano staccate (se così non
fosse facilitiamo il meccanismo con agitazione meccanica a mano), aggiungiamo
5-6 ml di mezzo di coltura per inattivare l’azione proteolitica dell’enzima, quindi la
sospensione cellulare viene trasferita in una provetta da 15 ml e centrifugata a 1100
rpm per 5 minuti a temperatura ambiente. Al termine della centrifugazione, il
sovranatante viene eliminato e il pellet cellulare risospeso in mezzo di coltura
completo. Le cellule possono essere seminate nuovamente in fiasca in modo tale da
raggiungere il numero idoneo per poi utilizzarle per gli esperimenti, oppure essere
congelate per conservarne una certa aliquota.

Materiali e metodi Capitolo 3
35
Tutte le operazioni riguardanti la coltura cellulare sono condotte sotto cappa a
flusso laminare, con un sistema di sterilizzazione a raggi U.V. quando il flusso è
interrotto o in assenza dell’operatore.
3.4.3 Congelamento cellulare
Il congelamento cellulare è la procedura necessaria qualora si voglia conservare le
cellule per un lungo periodo di tempo in uno stato di quiescenza.
Il mezzo di congelamento utilizzato per le cellule è costituito da FBS e 10% di
DMSO. Le componenti del mezzo di congelamento contribuiscono a stabilizzare le
cellule per il loro mantenimento a bassissime temperature (in azoto liquido o
congelatore a -80°C): l’FBS contiene proteine che possono esercitare una funzione
stabilizzante e protettiva sulla membrana in seguito ai danni che il congelamento
può provocare ed il DMSO agisce da criopreservante avendo la funzione di ridurre
le dimensioni dei cristalli di ghiaccio che si formano all’interno delle cellule.
Di norma, viene preparato, già aliquotato in eppendorf, 1,53 ml di FBS e mantenuto
a temperatura di -20°C; al momento della preparazione della sospensione cellulare
da congelare si aggiungono 170µl di DMSO raggiungendo e la sospensione
cellulare, raggiungendo così il volume massimo che può essere occupato all’interno
del criotubo.
La procedura di congelamento consiste di una prima parte in cui le cellule vengono
staccate e centrifugate come nella normale procedura del mantenimento in coltura.
Dopo centrifugazione (1100 rpm/5 minuti), il sovranatante viene rimosso ed il
pellet risospeso in 1,53 ml di FBS contenuto all’interno delle eppendorf aliquotate
e mantenute a -20°C indipendentemente dalla dimensione del pellet (se il
precipitato è eccessivamente abbondante si possono preparare più criotubi). Infine
viene aggiunta il restante volume di DMSO ed il criotubo viene alloggiato nel
contenitore per la conservazione in congelatore e tale procedura deve essere
abbastanza veloce in quanto il DMSO risulta essere non ottimale per le cellule a
temperatura ambiente.

Materiali e metodi Capitolo 3
36
3.5 Metodi utilizzati negli studi di biologia molecolare
3.5.1 Estrazione dell’RNA totale
L’estrazione di RNA totale da cellule è un processo che viene eseguito con lo scopo
di ottenere una quantità sufficiente di RNA da poter utilizzare per la
retrotrascrizione di c-DNA.
L’RNA totale viene estratto in condizioni e con materiali RF (RNase-Free) per
limitare la degradazione dell’RNA da parte dell’enzima RNase (Fig.16).
Il protocollo di estrazione si articola nelle seguenti fasi:
1. Staccare le cellule in coltura mediante il processo di tripsinizzazione, quindi
centrifugare a 1100 rpm per 5 minuti.
2. Eliminare il sovranatante e sottoporre il pellet a tre lavaggi consecutivi con
PBS, quindi risospenderlo alla fine in mezzo di coltura completo RPMI, in
volume variabile a seconda delle dimensioni del pellet.
3. Procedere alla conta cellulare con la camera di Burker, dopodichè
centrifugare a 1100 rpm per 5 minuti.
4. Eliminare il sovranatante ed aggiungere 1ml di PBS, quindi centrifugare a
1100 rpm per 3 minuti; tale procedura viene eseguita due volte.
5. Al termine dei due lavaggi, aspirare il sovranatante ed aggiungere il buffer
RLT ricostituito (10µl di beta-mercaptoetanolo per 1 ml di buffer RLT non
ricostituito), per indurre la lisi cellulare. Il volume di RLT da utilizzare per
la lisi dipende dal numero di cellule presenti nel campione, secondo la
proporzione fornita dalle istruzioni del fornitore del kit (Tab.5)
Numero di cellule (n) Volume RLT
n < 5x106 350 µl
5x106< n < 1x107 600 µl
Tabella 5. Volume di tampone RLT da utilizzare in base al numero di cellule o al supporto utilizzato.
6. Agitare la provetta con il buffer, in modo da lisare il pellet, ma dato che alla
lisi cellulare corrisponde un aumento della viscosità della soluzione, per

Materiali e metodi Capitolo 3
37
evitare la formazione di grumi o filamenti, è necessario utilizzare una
bacchetta in modo da omogenizzare quanto più possibile il contenuto del
lisato.
7. Trasferire il lisato in una eppendorf, quindi si aggiunge un volume di etanolo
RF 70%, pari a quello di buffer RLT utilizzato, agitando continuamente per
evitare la formazione di due fasi.
8. Caricare una quantità massima di 700 µl di lisato cellulare per ogni spin
column, quindi inserire la colonna in un collection tube da 2 ml e
centrifugare per 1 minuto a velocità maggiore di 10000 rpm.
9. Eliminare l’eluato e procedere con la digestione del DNA genomico
attraverso il kit RNase Free DNase Set (Qiagen, Milano):
- Aggiungere 350 µl di Buffer RW1 e centrifugare per 15 secondi a
velocità maggiore di 10000 rpm e si lascia agire per 15 minuti.
- Dopo questo breve lasso di tempo si aggiungono 350 µl di Buffer RW1
e si centrifuga nuovamente.
- Eliminato l’eluato si aggiungono 500 µl di Buffer RPE e si procede con
una nuova centrifugazione per 15 secondi ad una velocità maggiore di
10000 rpm e si elimina nuovamente l’eluato.
- Si caricano quindi altri 500 µl di Buffer RPE e si procede con una nuova
centrifugazione per 2 minuti a velocità maggiore di 10000 rpm.
- A questo punto, senza aggiungere Buffer, si centrifuga per 1 minuto
sempre a velocità maggiore di 10000 rpm, si elimina il tubo collettore,
ed utilizzando una eppendorf si aggiungono 40 µl di acqua RNase Free
sulla membrana, che viene lasciata idratare per qualche minuto.
- Si centrifuga un’ultima volta per 1 minuto a velocità maggiore di 10000
rpm e l’eluato ottenuto contiene l’RNA, quindi si elimina la colonna e
si alloggia la eppendorf in ghiaccio.
Per determinare la concentrazione e la purezza dei campioni ottenuti sono state
effettuate delle misurazioni spettrofotometriche con NanoQuant Infinite 2000
(TECAN, Salzsburg, Austria) a due lunghezze d’onda: 260 nm e 280 nm. La stima
della purezza del campione è data dal rapporto fra l’assorbanza a 260 nm e quella a
280 nm ed il valore ottenuto dovrà essere compreso tra 1,8 e 2: quanto più il valore
si avvicina a 2, tanto maggiore sarà la purezza dell’RNA estratto.

Materiali e metodi Capitolo 3
38
Figura 16. Schema della procedura per l'estrazione dell'RNA (RNeasy MiniKit, Quiagen, Milano).
3.5.2 Retrotrascrizione
La retrotrascrizione consente di ottenere dall’RNA estratto dalle cellule il cDNA,
che sarà, a sua volta, usato come stampo per la successiva reazione di
amplificazione. La retrotrascrizione viene effettuata seguendo la procedura
descritta dal QuantiTect® Reverse Transcription Handbook (Quiagen, Milano),
riportata di seguito:
1. Scongelare sia l’RNA sia i componenti del kit (gDNA Wipeout Buffer,
QuantiScript Reverse Transcriptase, QuantiScript RT Buffer, RT Primer
Mix, RNase Free Water) e porli in ghiaccio, centrifugandoli prima
brevemente così da far scendere eventuali gocce e si posizionano poi
nuovamente in ghiaccio. E’ necessario inoltre preparare anche la reazione
del DNA genomico (2 µl di gDNA Wipeout Buffer 7x, 1µg di RNA totale,

Materiali e metodi Capitolo 3
39
RNase Free Water q.b a 14 µl) e, come fatto precedentemente, si centrifuga
per far scendere eventuali gocce ed infine si posiziona il tutto in ghiaccio.
2. Incubare i campioni con l’RNA per 2 minuti nel termociclizzatore a 42°C e
posizionarli poi nuovamente in ghiaccio; tale procedura consente di
eliminare il DNA genomico dai campioni.
3. Preparare la Reverse Transcriptase Master Mix: 1 µl di QuantiScript
Reverse Transcriptase, 4 µl di QuantiScript RT Buffer 5x, 1 µl di RT Primer
Mix, i quali vengono aggiunti all’RNA totale privo di DNA genomico (14
µl), ottenuto al punto precedente, per un volume totale di 20 µl.
4. Centrifugare per far scendere eventuali gocce e posizionare in ghiaccio.
5. Incubare nel termociclizzatore seguendo il protocollo specifico per la
retrotrascrizione, in modo tale da attivare l’enzima QuantiScript Reverse
Transcriptase: 30 minuti a 42°C, 3 minuti a 95°C e infinito per 4°C.
I campioni di cDNA ottenuti saranno poi utilizzati per le reazioni di PCR o
conservati a -20°C.
3.5.3 Real-Time PCR
La real-time PCR è stata eseguita per quantificare l’espressione genica del NAPE-
PLD, DAGL-a, DAGL-β, MGLL e FAAH, e degli housekeeping GAPDH
(gliceraldeide-3-fosfato deidrogenasi) e -actina.
I primer forward e reverse, l’acqua, cDNA e la mix sono stati scongelati
preventivamente in ghiaccio.
In ogni well si addizionano:
-5 µl di sonda Syber Green (che va aggiunta per ultima perché è fotosensibile);
-400 nM di primer forward;
-400 nM di primer reverse;
-20 ng di cDNA ottenuto dalla retrotrascrizione dell’RNA estratto e diluito con un
rapporto 1:16;
-acqua quanto basta per ottenere un volume finale di 10 µl.
Materiali e metodi Capitolo 3
40
I protocolli di real-time PCR eseguiti si compongono delle seguenti fasi (Tab.6)
Tabella 6.Protocollo per real-time PCR con sonda SYBER Green.
La temperatura di annealing utilizzata per la GAPDH è di 59°C, DAGL-A (59°C),
DAGL-β (62°C), MGLL (62,8°C), FAAH (53,8°C) e NAPE-PLD (47°C).
3.5.4 Real-time PCR con sonde Taq-man
Le TaqMan sono dei tipi di sonde che sono state progettate per aumentare la
specificità della PCR quantitativa, sono infatti sonde ad ibridazione specifica per il
frammento d’interesse marcate con molecole fluorescenti (Fig.16).

Materiali e metodi Capitolo 3
41
Figura 17. Meccanismo d'azione della sonda TaqMan utilizzata in PCR quantitativa.
Negli esperimenti è stata utilizzata la Taqman® Gene Expression Assays
contenente:
-Master Mix II, no UNG, 2X
-TaqMan®Gene Expression Assay Mix 20X contenente i due primer d’interesse e
6-FAM™;
-RNase-free water;
Si procede seguendo le seguenti fasi:
1.Preparazione della piastra per Real-Time PCR
Ogni pozzetto della piastra da riempire deve contenere un volume finale di 20µl e
si introducono le componenti nel seguente ordine con le rispettive concentrazioni:
-acqua e cDNA (50 ng) (linee tumorali di A375, 501Mel, MeWo, SSM2C, M26C,
Me5 e Melanociti; e sfere di A375, SSM2C, M26C, Me5) per un volume di 9µl;
-Mix 10µl;

Materiali e metodi Capitolo 3
42
-Sonda TaqMan 1µl (CB1, CB2 e il 18s).
2. Protocollo e parametri della Thermal Cycling
-Attivazione della Polimerasi 95°C per un tempo di 10 minuti (eseguto per un solo
ciclo);
-Denaturazione 95°C per un tempo di 15 secondi;
-Annealing 60°C per un tempo di 1 minuto; questi ultimi due step ripetuti per 40
cicli.
Tabella 7. Protocollo di Real-Time PCR con sonde TaqMan.
3.6 Metodi utilizzati per l’analisi tramite Western Blotting
3.6.1 Estrazione proteica
L’estrazione proteica è stata eseguita per ottenere i lisati proteici da utilizzare per
lo studio dell’espressione proteica con Western Blotting.
Tale processo avviene secondo le seguenti fasi:
1. I componenti del kit vengono sciolti e si prepara la soluzione di lisi
addizionando 500µl di Ripa Lysis Buffer, 5µl di PMFS, 5µl del cocktail di
inibitori delle proteasi, 5µl di sodium othovanadate e 5 µl di NaF (inibitore
delle fosfatasi).
2. Dai dischi dove sono state precedentemente seminate e trattate le cellule si
aspira il mezzo ed, i dischi stessi, vengono collocati in ghiaccio, lavati con
PBS freddo ed infine aggiunti del tampone appena preparato.
3. Staccare le cellule con lo scraper e la sospensione cellulare ottenuta viene
trasferita in una eppendorf e mantenuta in ghiaccio.

Materiali e metodi Capitolo 3
43
4. Sonicare le eppendorf con la sospensione per 5 secondi quattro volte,
alternando la sonicazione in ghiaccio; infine incubare per 1 ora a 4°C.
5. Centrifugare a 4°C a 1100 rpm per 10 minuti e trasferire il surnatante,
costituito dal lisato proteico, in una nuova eppendorf che sarà conservata a
-20°C.
3.6.2 Quantizzazione proteica
La quantizzazione delle proteine è un metodo che viene eseguito per la
determinazione del contenuto proteico che si ritrova all’interno del lisato ottenuto
dalle nostre cellule. Nei nostri esperimenti abbiamo utilizzato il “Metodo di
Bradford”, un metodo che rileva la presenza delle proteine mediante il legame di
queste ultime con il colorante Coomassie Blue. Il procedimento è costituito da due
fasi:
1. La prima fase prevede di costruire una retta di taratura utilizzando soluzioni
standard di albumina bovina (BSA). Le soluzioni standard sono preparate
nell’intervallo di quantità tra 1 e 10 µg di proteina per una buona applicabilità del
metodo.
- si preparano tante eppendorf quanti sono i punti della curva di taratura, compreso
il bianco.
- aggiungere ad ogni eppendorf le quantità di acqua indicate in tabella (Tab.8) e
successivamente le quantità di soluzione madre di BSA (1mg/10 ml); aggiungere
infine per ognuna 200 µl di Bradford e vortexare fino a quando si verifica una
variazione di colore da marrone al blu, indice della presenza delle proteine.
L’intensità di tale colorazione è direttamente proporzionale al contenuto proteico.
- attendere 10 minuti e trasferire il contenuto delle eppendorf in cuvette per la lettura
spettrofotometrica a 620 nm.

Materiali e metodi Capitolo 3
44
Tabella 8.Valori per taratura con BSA secondo metodo Bradford.
2. La seconda fase ha lo scopo di dare una lettura di assorbanza allo
spettrofotometro in base al contenuto proteico del nostro lisato. Si procede nel
seguente modo:
- prelevare 10 µl di lisato proteico e metterli in una eppendorf, denominandola con
il nome della linea cellulare da cui deriva.
- aggiungere per ognuna 790 µl di acqua e 200 µl di Bradford, e vortexare.
- attendere 10 minuti e trasferire la soluzione, che ha virato colore da marrone al
blu, in cuvette per poi eseguire la lettura allo spettrofotometro a 620 nm. Ricordarsi
di eseguire la lettura anche del bianco ne quale si aggiunge PBS al posto del lisato
proteico.
Al termine di queste due fasi, viene costruita una retta di taratura nella quale si
interpolano i dati di assorbanza ottenuti dai nostri campioni per determinarne la
concentrazione proteica incognita.
3.6.3 Western Blotting
Il Western Blotting è una tecnica che consente di valutare ed analizzare le proteine
separate con elettroforesi grazie al riconoscimento da parte di anticorpi specifici.
1. La prima parte del protocollo prevede di separare le proteine mediante la
procedura di SDS- PAGE (SDS polyacrylamide gel electrophoresis), un tipo di
elettroforesi che si basa sulla capacità denaturante dell’SDS. L’elettroforesi

Materiali e metodi Capitolo 3
45
pertanto avviene su gel di poliacrilammide, in presenza di sodio dodecil solfato
(SDS), un composto denaturante in grado di legarsi alle proteine e distruggere i loro
legami ad idrogeno e ponti di solfuro, promuovendo così la perdita della loro
struttura secondaria o terziaria. Questo step è indispensabile per permettere ai
complessi proteina-SDS che si formano, di separarsi tra loro solo in funzione della
massa, annullando le cariche degli amminoacidi che influenzerebbero la
migrazione.
2. A questo punto inizia la corsa dove i complessi carichi negativamente migrano
verso il polo positivo del campo elettrico applicato e la loro mobilità risulta essere
inversamente proporzionale al Log10 del loro peso molecolare.
2.1 Per eseguire tale corsa vengono utilizzati due diversi tipi di gel: lo Stacking gel,
che permette di posizionare e concentrare i campioni negli appositi pozzetti,
affinchè questi possano iniziare la loro corsa elettroforetica dallo stesso punto; il
Resolving gel sottostante, ha invece la funzione di separare le proteine dei vari
campioni sulla base del loro peso molecolare (Fig.18).
Inizialmente si prepara quindi il gel di corsa secondo quanto è riportato dal
protocollo (Tab.9).
Tabella 9. Sostanze e relative quantità usate nella preparazione dei gel.
-Il Resolving Buffer è costituito da Tris base 1,5 M diluito in acqua milliQ e portato
a pH di 8.8.
-Lo Stacking Buffer è formato da Tris base 0,5 M diluito con acqua milliQ e portato
a pH di 6.8.

Materiali e metodi Capitolo 3
46
Le due miscele vengono agitate e, solo al momento dell’uso, vengono aggiunti i
catalizzatori APS (ammonioperisolfato 10%) e TEMED (Tetrameti-Etilen-
Diammina), il quale, una volta attivato da APS, funziona da catalizzatore rendendo
reattiva l’acrilamide ed inducendo così una polimerizzazione radicalica e la
formazione del gel.
Si procede quindi prima al caricamento del Resolving gel ed, una volta che il gel si
è solidificato, viene caricato lo Stacking gel, all’interno del quale si inserisce il
pettine che permette la formazione dei pozzetti al cui interno saranno caricati i
campioni.
2.2 Una volta preparato il gel, questo viene introdotto nella camera di elettroforesi,
immerso nel tampone di Running (TRIS 25 mM, 0,1% SDS e glicina 192 mM, il
tutto portato a pH di 8.3).
Prima di procedere al caricamento dei campioni è opportuno verificare che il livello
del liquido all’interno della camera sia più elevato di quello esterno. Si procede
quindi al caricamento dei campioni nei pozzetti, dopo averli scaldati per 3 minuti a
37°C così da favorire ulteriormente la completa denaturazione proteica per poi
effettuare la corsa elettroforetica a 110 V per circa 90 minuti.
Figura 18. Schema dell'SDS page.

Materiali e metodi Capitolo 3
47
3. Terminata la corsa elettroforetica, il gel di poliacrilamide contenente le proteine
separate, viene trasferito su membrana di nitrocellulosa, riequilibrata
precedentemente in tampone di Blotting (TRIS 25 mM, 0,1% di SDS, 20% di
Metanolo, 192 mM di glicina, il tutto portato a pH di 8.3).
3.1 Si procede dunque alla preparazione del Sandwich costituito nell’ordine dal
catodo verso l’anodo di spugna/ carta da filtro/ membrana di nitrocellulosa/ gel/
carta da filtro/ spugna. Durante tale procedura bisogna fare molta attenzione ad
evitare la formazione di bolle (Fig.19).
Figura 19. Componenti del sandwich e camera di trasferimento.
3.2 Il sandwich viene inserito all’interno di una camera di trasferimento immerso
in soluzione di Blotting fredda e si fa passare corrente a 330 mA per 90 minuti
perpendicolarmente al gel, provocando così il trasferimento delle proteine dal gel
alla nitrocellulosa.
3.3 Terminato il blotting, il sandwich viene aperto e la membrana viene colorata
per verificare il corretto trasferimento delle proteine, usando come colorante una
soluzione composta da: 0,1% di naphtol blue black, 10% di metanolo e 2% di acido
acetico. Successivamente si procede con la fase di decolorazione utilizzando una
soluzione formata al 50% da metanolo e 7% di acido acetico.
3.4 Si prosegue con la fase di Blocking che serve a bloccare tutti i siti di legame
aspecifici sulla nitrocellulosa. La membrana è quindi incubata in soluzione di

Materiali e metodi Capitolo 3
48
blocking (TRIS 20mM 2,42 g/l, NaCl 500mM 29,4 g/l pH 8.0, Tween-20 0,05% e
no-fat milk 5%) per 45 minuti su una bascula, così da essere mantenuta
costantemente in agitazione.
4. Trattamento con gli Anticorpi
4.1 Durante il tempo precedente nel quale la nitrocellulosa si trova sulla bascula in
soluzione di blocking, si procede con la preparazione della soluzione diluita con
anticorpo primario per β-actina (1:1000), CB1 (1:1000) e BRAF (1:1000) in
soluzione di blocking all’ 1%.
Terminato il periodo di incubazione, la soluzione viene rimossa e si aggiunge
successivamente quella contenente l’anticorpo primario e si lascia incubare tutta la
notte su una bascula a 4°C.
4.2 Il mattino successivo, una volta recuperata la soluzione con l’anticorpo in cui è
stata incubata la nitrocellulosa, vengono effettuati 3 lavaggi, ciascuno di 5 minuti,
con la soluzione di TBS- Tween 0,1% con il fine di eliminare l’anticorpo primario
non legato, lavorando con i campioni collocati su una bascula a temperatura
ambiente.
4.3 Terminata tale procedura si aggiunge la soluzione contenente l’anticorpo
secondario specifico precedentemente preparato alla opportuna diluizione, β-actina
Ab II anti-mouse (1:4000), CB1 Ab II anti-rabbit (1:4000), BRAF Ab II anti-mouse
(1:4000), lasciando incubare per 2 ore (Fig.20).
4.4 Finita questa incubazione, si effettuano tre lavaggi ciascuno con la soluzione di
TBS- Tween 0,1% a temperatura ambiente e della durata di 5 minuti.

Materiali e metodi Capitolo 3
49
Figura 20. Rappresentazione del trattamento delle nitrocellulose con anticorpo primario e secondario.
5. L’ultima fase del Western blotting consiste nella rilevazione delle bande
corrispondenti alle proteine di interesse con lo strumento ImageQuant LAS 4000
(Fig.21).
Figura 21. Strumento ImageQuant LAS 4000.
Tale strumento ha una risoluzione elevata, fino a 6,3 MPel (3072 ×2048 pixel) ed
una profondità di fuoco pari a 480mm. Il rumore di fondo è ridotto grazie al sistema
di raffreddamento a -25°C; la quantificazione è uniforme ed è possibile eseguire

Materiali e metodi Capitolo 3
50
rilevazioni multiplex nelle quali, la sensibilità, è tale da permettere il rilevamento
anche di 40 ng di proteine.
Le membrane vengono poste su pellicola e su di esse viene versata goccia a goccia
la soluzione con reagente luminescente ECL (preparata in rapporto 1:1) per 1
minuto. Al termine del tempo stabilito, le membrane vengono ricoperte da pellicola
ed inserite nello strumento che eseguirà la rilevazione delle bande corrispondenti
alla proteina di interesse nei diversi campioni.

Risultati e discussione Capitolo 4
51
CAPITOLO 4
Risultati e Discussione
4.1 Profilo di espressione genica dei componenti del sistema endocannabinoide
nel melanoma
4.1.1 Valutazione dell’espressione del recettore CB1 in cellule di melanoma
Lo studio di real-time PCR, usando sonde TaqMan, ha dimostrato che il recettore
dei cannabinoidi di tipo 1 (CB1R) è espresso nelle linee cellulari tumorali A375 e
501Mel e nelle cellule derivanti dal paziente Me5. I melanociti, la linea cellulare
MeWo e le cellule derivanti dal paziente SSM2c e M26c non esprimono, o
esprimono il recettore CB1 a livelli molto inferiori (Fig.22).
Figura 22. Espressione del recettore CB1 in diverse linee di melanoma cutaneo maligno.
I risultati suggeriscono una possibile associazione tra l’espressione del recettore
CB1 e la presenza della mutazione BRAF V600E. Infatti, il recettore risulta essere
espresso a più alti livelli nelle linee tumorali mutanti rispetto a quelle con genotipo
BRAF wild type. Questi risultati sono in parte in linea con quelli riportati nella
Risultati e discussione Capitolo 4
52
letteratura (Haskò et al., 2014; Kenessey et al., 2011), anche se la possibile
associazione tra CB1 e BRAF non è stata ancora chiaramente dimostrata.
4.1.2 Valutazione dell’espressione del recettore CB1 in cellule staminali di
melanoma.
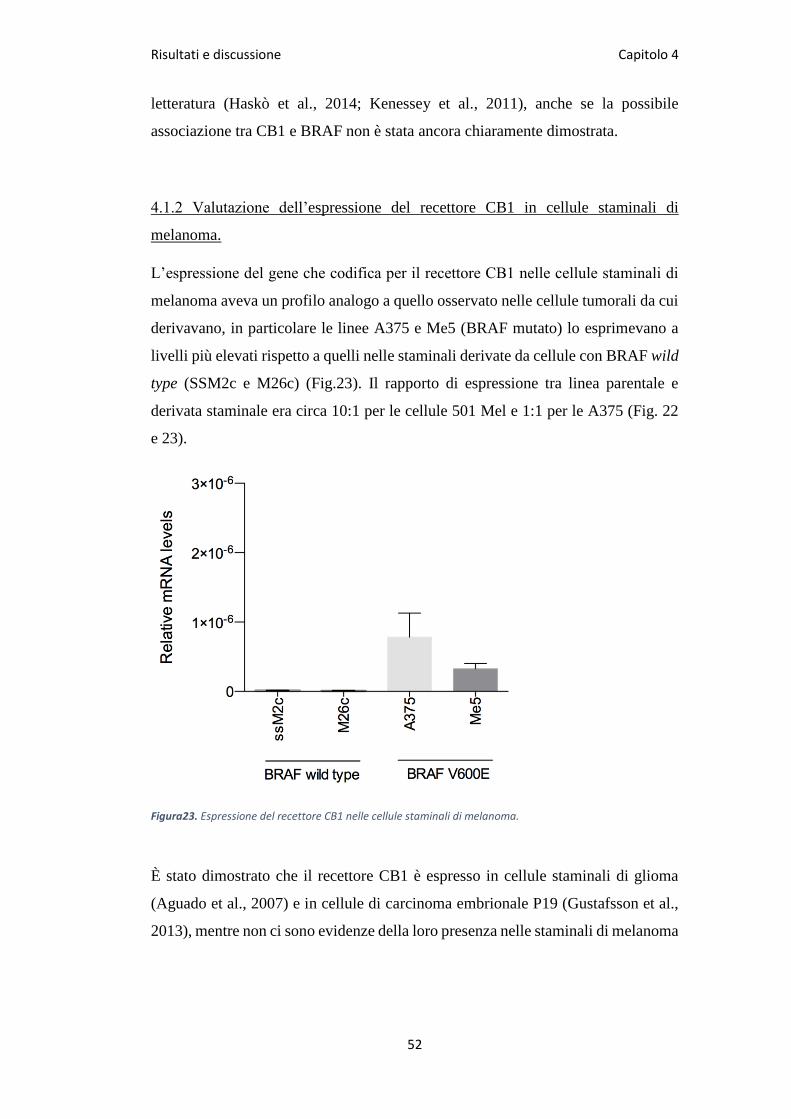
L’espressione del gene che codifica per il recettore CB1 nelle cellule staminali di
melanoma aveva un profilo analogo a quello osservato nelle cellule tumorali da cui
derivavano, in particolare le linee A375 e Me5 (BRAF mutato) lo esprimevano a
livelli più elevati rispetto a quelli nelle staminali derivate da cellule con BRAF wild
type (SSM2c e M26c) (Fig.23). Il rapporto di espressione tra linea parentale e
derivata staminale era circa 10:1 per le cellule 501 Mel e 1:1 per le A375 (Fig. 22
e 23).
Figura23. Espressione del recettore CB1 nelle cellule staminali di melanoma.
È stato dimostrato che il recettore CB1 è espresso in cellule staminali di glioma
(Aguado et al., 2007) e in cellule di carcinoma embrionale P19 (Gustafsson et al.,
2013), mentre non ci sono evidenze della loro presenza nelle staminali di melanoma

Risultati e discussione Capitolo 4
53
cutaneo maligno. Questi risultati quindi incoraggiano allo studio del ruolo del
sistema endocannabinoide nelle cellule iniziatrici di melanoma.
4.1.3 Valutazione dell’espressione del recettore CB2
Dagli studi in real-time PCR non emergeva la presenza del recettore CB2 sia nelle
cellule staminali, sia nelle rispettive tumorali primarie e nelle linee A375, 501 Mel
e MeWo (Tab.10). Come controllo positivo è stato utilizzato cDNA estratto da
cellule Jurkat (linfociti T immortalizzati) nelle quali è stata precedentemente
dimostrata la presenza del recettore CB2 (Börner et al., 2007).
Il ruolo indiretto dei recettori CB2 nel controllo della progressione e disseminazione
del melanoma nel SNC è stato comunque dimostrato in vitro (Haskò et al., 2014).
Tabella 10. Espressione del recettore CB2.

Risultati e discussione Capitolo 4
54
4.1.4 Valutazione dell’espressione degli enzimi coinvolti nella biosintesi e nel
catabolismo di AEA
L’espressione dei geni codificanti per gli enzimi NAPE-PDL e FAAH è stata
dimostrata nella linea di melanoma A375 ma non nelle cellule staminali
corrispondenti, nei melanociti, nelle altre linee cellulari tumorali e nelle cellule
derivanti da pazienti utilizzate nel nostro studio (Fig.24).
Figura 24. Espressione degli enzimi NAPE-PLD e FAAH deputati rispettivamente alla sintesi e al catabolismo di AEA.
Il rapporto tra i livelli di espressione di NAPE-PDL e FAAH nelle cellule A375 era
circa 300:1 indicando una propensione da parte di questa linea a sintetizzare AEA
in eccesso rispetto alla capacità di catabolismo cellulare.
4.1.5 Valutazione dell’espressione degli enzimi coinvolti nella biosintesi e nel
catabolismo di 2-AG
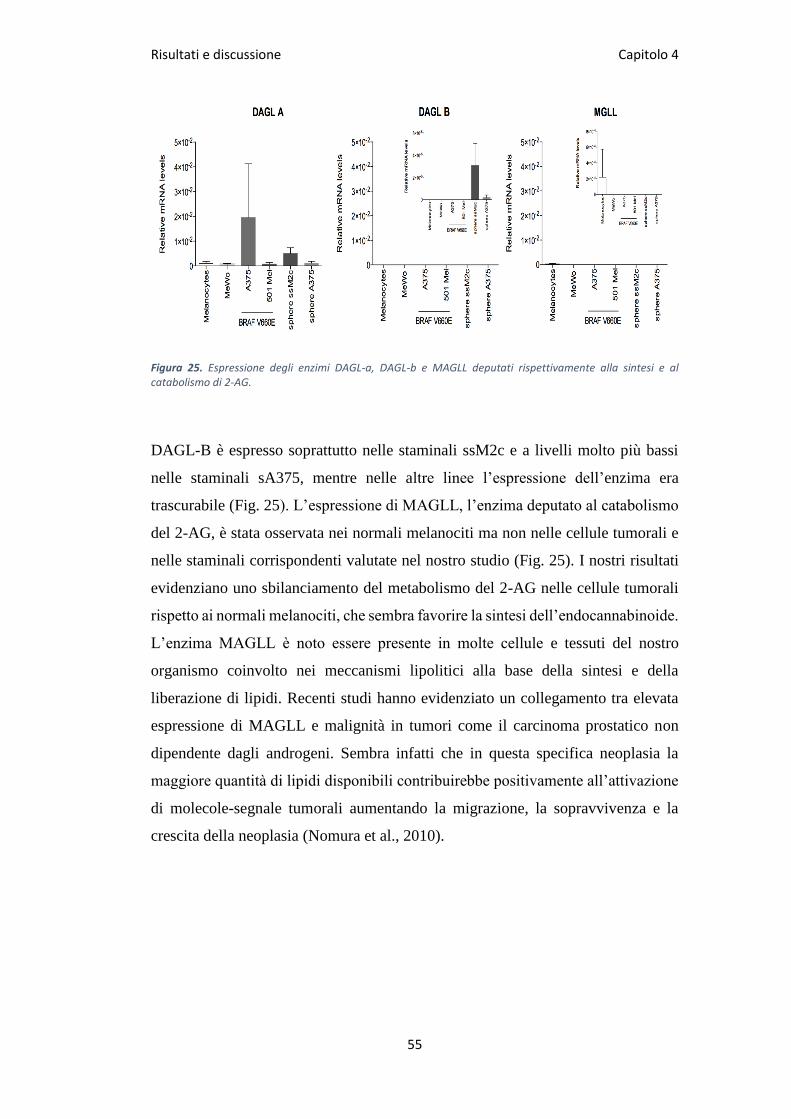
Lo studio di espressione genica ha evidenziato livelli particolarmente elevati di
espressione del gene che codifica per DAGL-A nella linea A375, e in misura minore
nelle altre linee saggiate, comprese le staminali derivate dalla stessa linea A375 e i
normali melanociti (Fig. 25).
Risultati e discussione Capitolo 4
55
Figura 25. Espressione degli enzimi DAGL-a, DAGL-b e MAGLL deputati rispettivamente alla sintesi e al catabolismo di 2-AG.
DAGL-B è espresso soprattutto nelle staminali ssM2c e a livelli molto più bassi
nelle staminali sA375, mentre nelle altre linee l’espressione dell’enzima era
trascurabile (Fig. 25). L’espressione di MAGLL, l’enzima deputato al catabolismo
del 2-AG, è stata osservata nei normali melanociti ma non nelle cellule tumorali e
nelle staminali corrispondenti valutate nel nostro studio (Fig. 25). I nostri risultati
evidenziano uno sbilanciamento del metabolismo del 2-AG nelle cellule tumorali
rispetto ai normali melanociti, che sembra favorire la sintesi dell’endocannabinoide.
L’enzima MAGLL è noto essere presente in molte cellule e tessuti del nostro
organismo coinvolto nei meccanismi lipolitici alla base della sintesi e della
liberazione di lipidi. Recenti studi hanno evidenziato un collegamento tra elevata
espressione di MAGLL e malignità in tumori come il carcinoma prostatico non
dipendente dagli androgeni. Sembra infatti che in questa specifica neoplasia la
maggiore quantità di lipidi disponibili contribuirebbe positivamente all’attivazione
di molecole-segnale tumorali aumentando la migrazione, la sopravvivenza e la
crescita della neoplasia (Nomura et al., 2010).
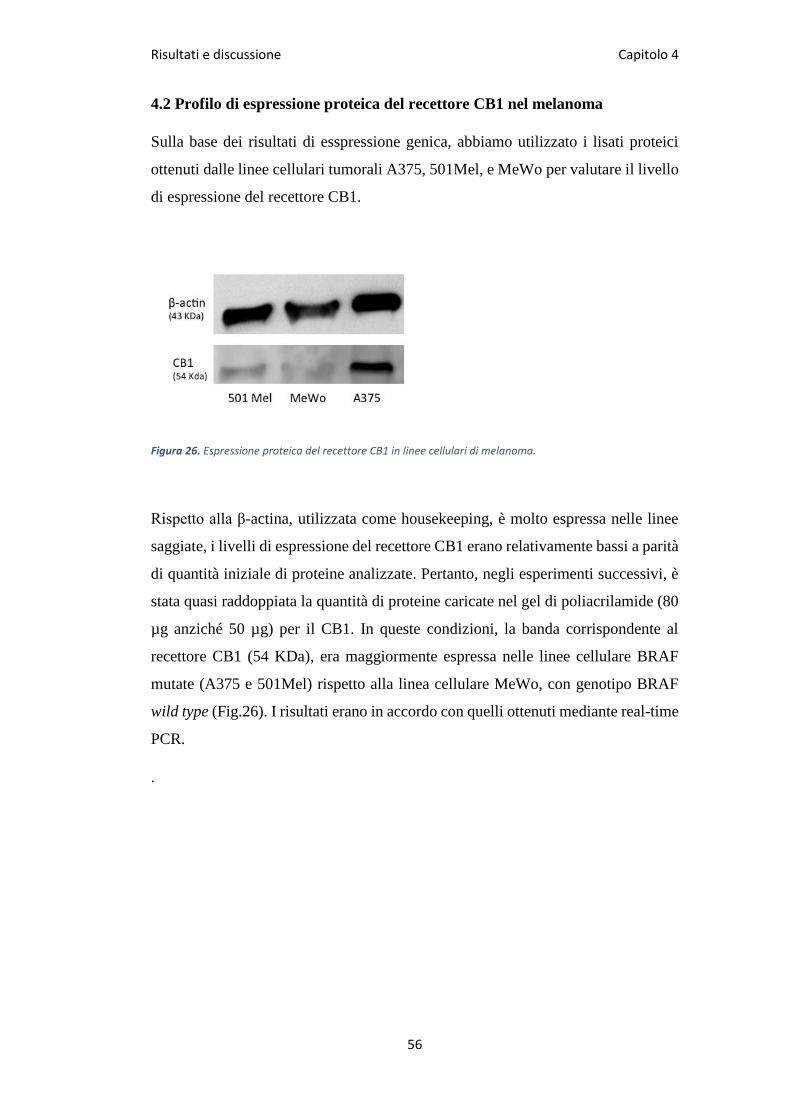
Risultati e discussione Capitolo 4
56
4.2 Profilo di espressione proteica del recettore CB1 nel melanoma
Sulla base dei risultati di esspressione genica, abbiamo utilizzato i lisati proteici
ottenuti dalle linee cellulari tumorali A375, 501Mel, e MeWo per valutare il livello
di espressione del recettore CB1.
Figura 26. Espressione proteica del recettore CB1 in linee cellulari di melanoma.
Rispetto alla β-actina, utilizzata come housekeeping, è molto espressa nelle linee
saggiate, i livelli di espressione del recettore CB1 erano relativamente bassi a parità
di quantità iniziale di proteine analizzate. Pertanto, negli esperimenti successivi, è
stata quasi raddoppiata la quantità di proteine caricate nel gel di poliacrilamide (80
µg anziché 50 µg) per il CB1. In queste condizioni, la banda corrispondente al
recettore CB1 (54 KDa), era maggiormente espressa nelle linee cellulare BRAF
mutate (A375 e 501Mel) rispetto alla linea cellulare MeWo, con genotipo BRAF
wild type (Fig.26). I risultati erano in accordo con quelli ottenuti mediante real-time
PCR.
.

Conclusioni Capitolo 5
57
CAPITOLO 5
Conclusioni
Il sistema endocannabinoide ha un ruolo funzionale importante nella proliferazione
cellulare e per questo è studiato molto in relazione ai meccanismi di crescita
tumorale. Attraverso gli esperimenti che hanno caratterizzato questo lavoro di tesi
abbiamo iniziato a valutare ciò che nella letteratura scientifica non è ancora ben
chiaro, e cioè il ruolo del sistema endocannabinoide nel melanoma cutaneo
maligno.
I risultati degli esperimenti sulla trascrizione genica dimostrano che il recettore CB1
è più espresso nelle linee cellulari di melanoma con mutazione del gene BRAF (che
codifica per una proteina coinvolta nei processi di proliferazione cellulare) rispetto
alle cellule tumorali con un genotipo BRAF wild type, suggerendo un possibile
collegamento con la maggiore aggressività cellulare associata alla presenza della
mutazione. Lo studio ha inoltre evidenziato l’espressione di tale recettore anche
nelle cellule staminali tumorali derivate da linee BRAF mutate ma non in quelle
derivate da cellule BRAF wild type o nei normali melanociti. Non è stata invece
evidenziata la presenza del recettore CB2, in termini di trascrizione genica, nelle
linee cellulari studiate. Nonostante il ruolo di questo sottotipo recettoriale nel
mantenimento del fenotipo cellulare tumorale sembri non essere determinante, il
suo significato funzionale nella progressione di questa neoplasia necessità di
ulteriori chiarimenti.
Un’altra importante osservazione è stata eseguita sulle cellule tumorali, cresciute in
terreno di coltura con presenza o meno di siero, per valutare un’eventuale
variazione dell’espressione del CB1 e della proteina BRAF. A tal proposito
abbiamo potuto evidenziare il fatto che, nelle cellule private del siero, vengano
messi in atto dei meccanismi compensatori, in risposta alla mancanza di nutrienti,
per i quali si ha un aumento dell’espressione sia del CB1 che di BRAF e quindi
possiamo concludere che, tali cellule, abbiano una maggiore aggressività, rispetto
alle stesse cresciute in terreno di coltura completo.

Conclusioni Capitolo 5
58
Infine, abbiamo dimostrato l’espressione genica di enzimi deputati alla sintesi e al
metabolismo degli endocannabinoidi endogeni, AEA e 2-AG. I risultati indicano
che, rispetto ai normali melanociti, le cellule di melanoma cutaneo maligno
sembrano metabolicamente orientate a conservare la sintesi di endocannabinoidi.
Questi studi rappresentano un punto di partenza importante indicando un possibile
ruolo del sistema endocannabinoide nel melanoma cutaneo maligno. Sono in corso
d’opera esperimenti con tecniche di silenziamento genico, finalizzati a valutare il
significato funzionale del recettore CB1 nella linea A375. Dati preliminari indicano
che il silenziamento del recettore induce un rallentamento della crescita tumorale
in vitro e una minore attività clonogenica delle cellule tumorali, nella direzione
degli studi di trascrizione genica oggetto di questa tesi di laurea. Il nostro obiettivo
è quello di capire se la modulazione del sistema endocannabinoide possa
rappresentare una nuova e concreta strategia farmacologica in grado di arrestare
questa devastante malattia.

Nomenclatura
59
Nomenclatura
2-AG 2-arachidonoilglicerolo
501 Mel Linea cellulare di melanoma metastatico
A375t Linea cellulare di melanoma metastatico
A375s Sfere di melanoma metastatico della linea A375
AbI e AbII Anticorpo primario e secondario
AEA Anandamide
APS e TEMED Catalizzatori per gel di Western Blotting
BRAF V600E Mutazione a carico della proteina BRaf
CB1 e CB2 Recettori cannabinoidi
CDK Ciclina chinasi-dipendente che inattiva Rb
CMC Cellule staminali tumorali
COX-2 Enzima coinvolto nel metabolismo di 2-AG
CT Ciclo Threshold
DAGL-A e DAGL-B Enzimi che sintetizzano il 2-AG
DMEM Dulbecco’s Modified Medium, mezzo di coltura
DMSO Dimetilsolfossido, parte del mezzo di congelamento
ECS Sistema endocannabinoide
ERK Extracellular signal-regulated kinases
FAAH Enzima che metabolizza l’AEA
FBS Siero fetale bovino
GAPDH Gene housekepping
M2 Melanocytes Growth Medium, mezzo di coltura

Nomenclatura
60
MAPK Mitogen-actvated protein kinase
Me5 t Linea tumorale isolata da paziente
Me5 s Sfere della linea corrispondente Me5
Me26c t Linea tumorale isolata da paziente
Me26c s Sfere della linea corrispondente Me26c
MGLL Enzima che metabolizza IL 2-AG
MeWo Linea cellulare di melanoma metastatico
NAPE-PLD Enzima che sintetizza l’AEA
P/S Penicillina/ Streptomicina
PBS Phosfate saline buffer
Rb retinoblastoma, soppressore tumorale
RPMI Rosewell Park Memorial Institute, mezzo di coltura
SDS Sodio dodecil solfato
SSM2c t Linea tumorale isolata da paziente
SSM2c s Sfere della linea corrispondente SSM2c
TA Temperatura di annealing
TM Temperatura di melting
Try Tripsina

61
Bibliografia
Aguado T1, Carracedo A, Julien B, Velasco G, Milman G, Mechoulam R, Alvarez
L, Guzmán M, Galve-Roperh I. Cannabinoids induce glioma stem-like cell
differentiation and inhibit gliomagenesis. J Biol Chem. 2007 Mar 2;282(9):6854-
62. Epub 2007 Jan 2.
Aiom-airtum, I numeri del cancro in Italia 2014, Available: [http://www.registri-
tumori.it/cms/it/numeri]
Aitken J1, Welch J, Duffy D, Milligan A, Green A, Martin N, Hayward N.
CDKN2A variants in a population-based sample of Queensland families with
melanoma. J Natl Cancer Inst. 1999 Mar 3;91(5):446-52.
Alger BE1, Kim J. Supply and demand for endocannabinoids. Trends Neurosci.
2011 Jun;34(6):304-15. doi: 10.1016/j.tins.2011.03.003. Epub 2011 Apr 18.
Amiri KI1, Richmond A. Role of nuclear factor-kappa B in melanoma. Cancer
Metastasis Rev. 2005 Jun;24(2):301-13.
Börner C1, Höllt V, Sebald W, Kraus J. Transcriptional regulation of the
cannabinoid receptor type 1 gene in T cells by cannabinoids. J Leukoc Biol. 2007
Jan;81(1):336-43. Epub 2006 Oct 13.
Carnero A1. Targeting the cell cycle for cancer therapy. Br J Cancer. 2002 Jul
15;87(2):129-33.

62
Chin L1. The genetics of malignant melanoma: lessons from mouse and man. Nat
Rev Cancer. 2003 Aug;3(8):559-70.
Compagnucci C1, Di Siena S, Bustamante MB, Di Giacomo D, Di Tommaso M,
Maccarrone M, Grimaldi P, Sette C. Type-1 (CB1) cannabinoid receptor promotes
neuronal differentiation and maturation of neural stem cells. PLoS One.
2013;8(1):e54271. doi: 10.1371/journal.pone.0054271. Epub 2013 Jan 23.
Currais A1, Hortobágyi T, Soriano S. The neuronal cell cycle as a mechanism of
pathogenesis in Alzheimer's disease. Aging (Albany NY). 2009 Apr 28;1(4):363-
71.
Daniel K. Nomura, Jonathan Z. Long, Sherry Niessen, Heather S. Hoover, Shu-
Wing Ng, and Benjamin F. Cravatt. Monoacylglycerol lipase regulates a fatty acid
network that promotes cancer pathogenesis. Cell.2010 Jan 8: 140(1): 49-61. doi:
10.1016/j.cell.2009.11.027.
Daniel J. Hermanson and Lawrence J. Marnett* Cannabinoids, Endocannabinoids
and Cancer . Cancer Metastasis Rev. Author manuscript; available in PMC 2012
Jun 3. Published in final edited form as: Cancer Metastasis Rev. 2011 Dec; 30(3-
4): 599–612. doi: 10.1007/s10555-011-9318-8.
Dankort D1, Curley DP, Cartlidge RA, Nelson B, Karnezis AN, Damsky WE Jr,
You MJ, DePinho RA, McMahon M, Bosenberg M. Braf(V600E) cooperates with
Pten loss to induce metastatic melanoma. Nat Genet. 2009 May;41(5):544-52. doi:
10.1038/ng.356. Epub 2009 Mar 12.
Datta SR1, Brunet A, Greenberg ME. Cellular survival: a play in three Akts. Genes
Dev. 1999 Nov 15;13(22):2905-27.

63
Davis MI, Ronesi J, Lovinger DM. (2003) A predominant role for inhibition of the
adenylate cyclase/ protein kinase A pathway in ERK activation by cannabinoid
receptor 1 in N1E-115 neuroblastoma cells. J Biol Chem. 278:48973-80.
Di Marzo V1. The endocannabinoid system: its general strategy of action, tools for
its pharmacological manipulation and potential therapeutic exploitation. Pharmacol
Res. 2009 Aug;60(2):77-84. doi: 10.1016/j.phrs.2009.02.010. Epub 2009 Mar 4.
Fang D1, Nguyen TK, Leishear K, Finko R, Kulp AN, Hotz S, Van Belle PA, Xu
X, Elder DE, Herlyn M. A tumorigenic subpopulation with stem cell properties in
melanomas. Cancer Res. 2005 Oct 15;65(20):9328-37.
Eisenmann KM1, VanBrocklin MW, Staffend NA, Kitchen SM, Koo HM.
Mitogen-activated protein kinase pathway-dependent tumor-specific survival
signaling in melanoma cells through inactivation of the proapoptotic protein bad.
Cancer Res. 2003 Dec 1;63(23):8330-7.
Emma L Curry, Mohammad Moad, Craig N Robson, and Rakesh Heer Using
induced pluripotent stem cells as a tool for modelling carcinogenesis World J Stem
Cells. 2015 Mar 26; 7(2): 461–469. Published online 2015 Mar 26. doi:
10.4252/wjsc.v7.i2.461
Fargnoli MC1, Argenziano G, Zalaudek I, Peris K. High- and low-penetrance
cutaneous melanoma susceptibility genes. Expert Rev Anticancer Ther. 2006
May;6(5):657-70.
Galve-Roperh I1, Chiurchiù V, Díaz-Alonso J, Bari M, Guzmán M, Maccarrone M.
Cannabinoid receptor signaling in progenitor/stem cell proliferation and

64
differentiation. Prog Lipid Res. 2013 Oct;52(4):633-50. doi:
10.1016/j.plipres.2013.05.004. Epub 2013 Sep 25.
Globacan 2012, (2014) Available:
[http://globocan.iarc.fr/old/pie_site.asp?selection=16120&title=Melanoma+of+ski
n&sex=0&type=0&populations=0&window=1&join=1&submit=%C2%A0Execu
te%C2%A0]
Gandini S1, Sera F, Cattaruzza MS, Pasquini P, Abeni D, Boyle P, Melchi CF.
Meta-analysis of risk factors for cutaneous melanoma: I. Common and atypical
naevi. Eur J Cancer. 2005 Jan;41(1):28-44.
Gandini S1, Sera F, Cattaruzza MS, Pasquini P, Picconi O, Boyle P, Melchi CF.
Meta-analysis of risk factors for cutaneous melanoma: II. Sun exposure. Eur J
Cancer. 2005 Jan;41(1):45-60.
Gandini S1, Sera F, Cattaruzza MS, Pasquini P, Zanetti R, Masini C, Boyle P,
Melchi CF. Meta-analysis of risk factors for cutaneous melanoma: III. Family
history, actinic damage and phenotypic factors. Eur J Cancer. 2005
Sep;41(14):2040-59.
Giles N1, Pavey S, Pinder A, Gabrielli B. Multiple melanoma susceptibility factors
function in an ultraviolet radiation response pathway in skin. Br J Dermatol. 2012
Feb;166(2):362-71. doi: 10.1111/j.1365-2133.2011.10635.x. Epub 2011 Dec 5.
Girouard SD1, Murphy GF. Melanoma stem cells: not rare, but well done. Lab
Invest. 2011 May;91(5):647-64. doi: 10.1038/labinvest.2011.50. Epub 2011 Mar
28.

65
Goggins WB1, Tsao H. A population-based analysis of risk factors for a second
primary cutaneous melanoma among melanoma survivors. Cancer. 2003 Feb
1;97(3):639-43.
Gudbjartsson DF1, Sulem P, Stacey SN, Goldstein AM, Rafnar T, Sigurgeirsson B,
Benediktsdottir KR, Thorisdottir K, Ragnarsson R, Sveinsdottir SG, Magnusson V,
Lindblom A, Kostulas K, Botella-Estrada R, Soriano V, Juberías P, Grasa M, Saez
B, Andres R, Scherer D, Rudnai P, Gurzau E, Koppova K, Kiemeney LA,
Jakobsdottir M, Steinberg S, Helgason A, Gretarsdottir S, Tucker MA, Mayordomo
JI, Nagore E, Kumar R, Hansson J, Olafsson JH, Gulcher J, Kong A,
Thorsteinsdottir U, Stefansson K. ASIP and TYR pigmentation variants associate
with cutaneous melanoma and basal cell carcinoma. Nat Genet. 2008 Jul;40(7):886-
91. doi: 10.1038/ng.161. Epub 2008 May 18.
Gustafsson SB1, Wallenius A, Zackrisson H, Popova D, Plym Forshell L,
Jacobsson SO. Effects of cannabinoids and related fatty acids upon the viability of
P19 embryonal carcinoma cells. Arch Toxicol. 2013 Nov;87(11):1939-51. doi:
10.1007/s00204-013-1051-3. Epub 2013 Apr 4.
Halaban R1. Rb/E2F: a two-edged sword in the melanocytic system. Cancer
Metastasis Rev. 2005 Jun;24(2):339-56.
Hardin, Bertoni, Kleinsmith (2012) Principles of Cell Biology Available:
[http://www.mun.ca/biology/desmid/brian/BIOL2060/BIOL2060-19/CB19.html].
Harland M1, Cust AE2, Badenas C3, Chang YM1, Holland EA4, Aguilera P3,
Aitken JF5, Armstrong BK2, Barrett JH1, Carrera C3, Chan M1, Gascoyne J1,
Giles GG6, Agha-Hamilton C4, Hopper JL7, Jenkins MA7, Kanetsky PA8, Kefford
RF4, Kolm I4, Lowery J9, Malvehy J3, Ogbah Z10, Puig-Butille JA3, Orihuela-
Segalés J11, Randerson-Moor JA1, Schmid H4, Taylor CF9, Whitaker L1, Bishop

66
DT1, Mann GJ4, Newton-Bishop JA1, Puig S3. Prevalence and predictors of
germline CDKN2A mutations for melanoma cases from Australia, Spain and the
United Kingdom. Hered Cancer Clin Pract. 2014 Nov 20;12(1):20. doi:
10.1186/1897-4287-12-20. eCollection 2014.
Haskó J1, Fazakas C2, Molnár J3, Nyúl-Tóth Á4, Herman H5, Hermenean A6,
Wilhelm I7, Persidsky Y8, Krizbai IA9. CB2 receptor activation inhibits melanoma
cell transmigration through the blood-brain barrier. Int J Mol Sci. 2014 May
8;15(5):8063-74. doi: 10.3390/ijms15058063.
Hennessy BT1, Smith DL, Ram PT, Lu Y, Mills GB. Exploiting the PI3K/AKT
pathway for cancer drug discovery. Nat Rev Drug Discov. 2005 Dec;4(12):988-
1004.
Hocker T1, Tsao H. Ultraviolet radiation and melanoma: a systematic review and
analysis of reported sequence variants. Hum Mutat. 2007 Jun;28(6):578-88.
Ibrahim N1, Haluska FG. Molecular pathogenesis of cutaneous melanocytic
neoplasms. Annu Rev Pathol. 2009;4:551-79. doi:
10.1146/annurev.pathol.3.121806.151541.
Jemal A1, Saraiya M, Patel P, Cherala SS, Barnholtz-Sloan J, Kim J, Wiggins CL,
Wingo PA. Recent trends in cutaneous melanoma incidence and death rates in the
United States, 1992-2006. J Am Acad Dermatol. 2011 Nov;65(5 Suppl 1):S17-
25.e1-3. doi: 10.1016/j.jaad.2011.04.032.
Jiang S1, Fu Y, Williams J, Wood J, Pandarinathan L, Avraham S, Makriyannis A,
Avraham S, Avraham HK. Expression and function of cannabinoid receptors CB1

67
and CB2 and their cognate cannabinoid ligands in murine embryonic stem cells.
PLoS One. 2007 Jul 25;2(7):e641.
Karbowniczek M1, Spittle CS, Morrison T, Wu H, Henske EP. mTOR is activated
in the majority of malignant melanomas. J Invest Dermatol. 2008 Apr;128(4):980-
7. Epub 2007 Oct 4.
Kauffmann RM1, Chen SL2. Workup and staging of malignant melanoma. Surg
Clin North Am. 2014 Oct;94(5):963-72, vii. doi: 10.1016/j.suc.2014.07.001. Epub
2014 Aug 6.
Kenessey I1, Bánki B, Márk A, Varga N, Tóvári J, Ladányi A, Rásó E, Tímár J.
Revisiting CB1 receptor as drug target in human melanoma. Pathol Oncol Res.
2012 Oct;18(4):857-66. Epub 2012 Mar 24.
Laderoute KR1, Amin K, Calaoagan JM, Knapp M, Le T, Orduna J, Foretz M,
Viollet B. 5'-AMP-activated protein kinase (AMPK) is induced by low-oxygen and
glucose deprivation conditions found in solid-tumor microenvironments. Mol Cell
Biol. 2006 Jul;26(14):5336-47.
Lasithiotakis K1, Leiter U, Meier F, Eigentler T, Metzler G, Moehrle M,
Breuninger H, Garbe C. Age and gender are significant independent predictors of
survival in primary cutaneous melanoma. Cancer. 2008 Apr 15;112(8):1795-804.
doi: 10.1002/cncr.23359.
Lee N1, Barthel SR1, Schatton T2. Melanoma stem cells and metastasis: mimicking
hematopoietic cell trafficking? Lab Invest. 2014 Jan;94(1):13-30. doi:
10.1038/labinvest.2013.116. Epub 2013 Oct 14.

68
Madonna G1, Ullman CD, Gentilcore G, Palmieri G, Ascierto PA. NF-κB as
potential target in the treatment of melanoma. J Transl Med. 2012 Mar 20;10:53.
doi: 10.1186/1479-5876-10-53.
Markovic SN1, Erickson LA, Rao RD, Weenig RH, Pockaj BA, Bardia A, Vachon
CM, Schild SE, McWilliams RR, Hand JL, Laman SD, Kottschade LA, Maples WJ,
Pittelkow MR, Pulido JS, Cameron JD, Creagan ET; Melanoma Study Group of the
Mayo Clinic Cancer Center. Malignant melanoma in the 21st century, part 1:
epidemiology, risk factors, screening, prevention, and diagnosis. Mayo Clin Proc.
2007 Mar;82(3):364-80.
Merriam-Webster. Melanoma. Available [http://www.merriam-
webster.com/medical/melanoma].
Meyle KD1, Guldberg P. Genetic risk factors for melanoma. Hum Genet. 2009
Oct;126(4):499-510. doi: 10.1007/s00439-009-0715-9. Epub 2009 Jul 8.
Mihić LL1, Bulat V, Situm M, Krolo I, Seserko A. The role of apoptosis in the
pathogenesis of malignant melanoma. Coll Antropol. 2010 Apr;34 Suppl 2:303-6.
Na YR1, Seok SH, Kim DJ, Han JH, Kim TH, Jung H, Lee BH, Park JH. Isolation
and characterization of spheroid cells from human malignant melanoma cell line
WM-266-4. Tumour Biol. 2009;30(5-6):300-9. doi: 10.1159/000261073. Epub
2009 Nov 23.
National Cancer Institute. Melanoma. Available:
[http://www.cancer.gov/cancertopics/types/melanoma].

69
Nones J1, Spohr TC, Furtado DR, Sartore RC, Paulsen BS, Guimarães MZ, Rehen
SK. Cannabinoids modulate cell survival in embryoid bodies. Cell Biol Int. 2010
Mar 12;34(4):399-408. doi: 10.1042/CBI20090036.
Oh HA1, Kwon S, Choi S, Shin H, Yoon KH, Kim WJ, Lim HJ. Uncovering a role
for endocannabinoid signaling in autophagy in preimplantation mouse embryos.
Mol Hum Reprod. 2013 Feb;19(2):93-101. doi: 10.1093/molehr/gas049. Epub 2012
Oct 30.
Palmieri G1, Capone M, Ascierto ML, Gentilcore G, Stroncek DF, Casula M, Sini
MC, Palla M, Mozzillo N, Ascierto PA. Main roads to melanoma. J Transl Med.
2009 Oct 14;7:86. doi: 10.1186/1479-5876-7-86.
Paria BC1, Ma W, Andrenyak DM, Schmid PC, Schmid HH, Moody DE, Deng H,
Makriyannis A, Dey SK. Effects of cannabinoids on preimplantation mouse embryo
development and implantation are mediated by brain-type cannabinoid receptors.
Biol Reprod. 1998 Jun;58(6):1490-5.
Platz A1, Egyhazi S, Ringborg U, Hansson J. Human cutaneous melanoma; a
review of NRAS and BRAF mutation frequencies in relation to histogenetic
subclass and body site. Mol Oncol. 2008 Apr;1(4):395-405. doi:
10.1016/j.molonc.2007.12.003. Epub 2007 Dec 28.
Pisanti S1, Picardi P, D'Alessandro A, Laezza C, Bifulco M. The endocannabinoid
signaling system in cancer. Trends Pharmacol Sci. 2013 May;34(5):273-82. doi:
10.1016/j.tips.2013.03.003. Epub 2013 Apr 17.
Pópulo H, Soares P, Faustino A, Rocha AS, Silva P, Azevedo F, Lopes JM. mTOR
pathway activation in cutaneous melanoma is associated with poorer prognosis

70
characteristics. Pigment Cell Melanoma Res. 2011 Feb;24(1):254-7. doi:
10.1111/j.1755-148X.2010.00796.x. Epub 2010 Nov 19.
Porta C1, Paglino C1, Mosca A2. Targeting PI3K/Akt/mTOR Signaling in Cancer.
Front Oncol. 2014 Apr 14;4:64. doi: 10.3389/fonc.2014.00064. eCollection 2014.
R. G. Pertwee,corresponding author A. C. Howlett, M. E. Abood, S. P. H.
Alexander, V. Di Marzo, M. R. Elphick, P. J. Greasley, H. S. Hansen, G. Kunos, K.
Mackie, R. Mechoulam, and R. A. Ross International Union of Basic and Clinical
Pharmacology. LXXIX. Cannabinoid Receptors and Their Ligands: Beyond CB1
and CB2 Pharmacol Rev. 2010 Dec; 62(4): 588–631. doi: 10.1124/pr.110.003004
PMCID: PMC2993256
Rigel DS1. Trends in dermatology: melanoma incidence. Arch Dermatol. 2010
Mar;146(3):318. doi: 10.1001/archdermatol.2009.379.
Roger S Day What Tumor Dynamics Modeling Can Teach us About Exploiting the
Stem-Cell View for Better Cancer Treatment Cancer Inform. 2015; 14(Suppl 2):
25–36. Published online 2015 Feb 24. doi: 10.4137/CIN.S17294 PMCID:
PMC4345852
Sánchez MG1, Ruiz-Llorente L, Sánchez AM, Díaz-Laviada I. Activation of
phosphoinositide 3-kinase/PKB pathway by CB(1) and CB(2) cannabinoid
receptors expressed in prostate PC-3 cells. Involvement in Raf-1 stimulation and
NGF induction. Cell Signal. 2003 Sep;15(9):851-9.
Sarbassov DD1, Ali SM, Kim DH, Guertin DA, Latek RR, Erdjument-Bromage H,
Tempst P, Sabatini DM. Rictor, a novel binding partner of mTOR, defines a

71
rapamycin-insensitive and raptor-independent pathway that regulates the
cytoskeleton. Curr Biol. 2004 Jul 27;14(14):1296-302.
Schatton T1, Frank MH. Cancer stem cells and human malignant melanoma.
Pigment Cell Melanoma Res. 2008 Feb;21(1):39-55. doi: 10.1111/j.1755-
148X.2007.00427.x.
Sekulic A1, Haluska P Jr, Miller AJ, Genebriera De Lamo J, Ejadi S, Pulido JS,
Salomao DR, Thorland EC, Vile RG, Swanson DL, Pockaj BA, Laman SD,
Pittelkow MR, Markovic SN; Melanoma Study Group of Mayo Clinic Cancer
Center. Malignant melanoma in the 21st century: the emerging molecular
landscape. Mayo Clin Proc. 2008 Jul;83(7):825-46.
Siegel R1, Ma J, Zou Z, Jemal A. Cancer statistics, 2014. CA Cancer J Clin. 2014
Jan-Feb;64(1):9-29. doi: 10.3322/caac.21208. Epub 2014 Jan 7.
Smalley KS1. Understanding melanoma signaling networks as the basis for
molecular targeted therapy. J Invest Dermatol. 2010 Jan;130(1):28-37. doi:
10.1038/jid.2009.177.
Stahl JM1, Cheung M, Sharma A, Trivedi NR, Shanmugam S, Robertson GP. Loss
of PTEN promotes tumor development in malignant melanoma. Cancer Res. 2003
Jun 1;63(11):2881-90.
Stahl JM1, Sharma A, Cheung M, Zimmerman M, Cheng JQ, Bosenberg MW,
Kester M, Sandirasegarane L, Robertson GP. Deregulated Akt3 activity promotes
development of malignant melanoma. Cancer Res. 2004 Oct 1;64(19):7002-10.

72
Tsao H1, Atkins MB, Sober AJ. Management of cutaneous melanoma. N Engl J
Med. 2004 Sep 2;351(10):998-1012.
Uzdensky AB1, Demyanenko SV, Bibov MY. Signal transduction in human
cutaneous melanoma and target drugs. Curr Cancer Drug Targets. 2013
Oct;13(8):843-66.
Vainio H1, Miller AB, Bianchini F. An international evaluation of the cancer-
preventive potential of sunscreens. Int J Cancer. 2000 Dec 1;88(5):838-42.
Van Dross R1, Soliman E, Jha S, Johnson T, Mukhopadhyay S. Receptor-
dependent and receptor-independent endocannabinoid signaling: a therapeutic
target for regulation of cancer growth. Life Sci. 2013 Mar 19;92(8-9):463-6. doi:
10.1016/j.lfs.2012.09.025. Epub 2012 Oct 12.
Van Sickle MD1, Duncan M, Kingsley PJ, Mouihate A, Urbani P, Mackie K, Stella
N, Makriyannis A, Piomelli D, Davison JS, Marnett LJ, Di Marzo V, Pittman QJ,
Patel KD, Sharkey KA. Identification and functional characterization of brainstem
cannabinoid CB2 receptors. Science. 2005 Oct 14;310(5746):329-32.
Wang H1, Guo Y, Wang D, Kingsley PJ, Marnett LJ, Das SK, DuBois RN, Dey
SK. Aberrant cannabinoid signaling impairs oviductal transport of embryos. Nat
Med. 2004 Oct;10(10):1074-80. Epub 2004 Sep 19.
Weinstock MA. Reducing death from melanoma and standards of evidence. J Invest
Dermatol. 2012 May;132(5):1311-2. doi: 10.1038/jid.2012.57.

73
Wolchok JD (2014) Melanoma Available: [http://www.cancerresearch.org/cancer-
immunotherapy/impacting-all-cancers/melanoma].
Woods D1, Cherwinski H, Venetsanakos E, Bhat A, Gysin S, Humbert M, Bray PF,
Saylor VL, McMahon M. Induction of beta3-integrin gene expression by sustained
activation of the Ras-regulated Raf-MEK-extracellular signal-regulated kinase
signaling pathway. Mol Cell Biol. 2001 May;21(9):3192-205.

74
Indice delle Tabelle
Tabella 1.1 Fattori di rischio di melanoma cutaneo 16
Tabella 3.2 Cellule isolate da singolo paziente 30
Tabella 3.3 Sequenza dei primer utilizzati in Real Time PCR 31
Tabella 3.4 Primer utilizzati in sonde TaqMan 32
Tabella 3.5 Volume di tampone RLT da utilizzare in base al numero di cellule 36
Tabella 3.6 Protocollo di real-time PCR con sonda SYBERGreen 40
Tabella 3.7 Protocollo di real-time PCR con sonda TaqMan 42
Tabella 3.8 Valori per taratura con BSA, Metodo Bradford 44
Tabella 3.9 Sostanze per preparazione gel in Western Blotting 45
Tabella 4.10 Espressione del recettore CB2 53

75
Indice delle Figure
Figura 1.1 Struttura chimica dei principali composti cannabinoidi 5
Figura 1.2 Struttura dei recettori cannabinoidi CB1 e CB2 5
Figura 1.3 Struttura del recettore CB1 6
Figura 1.4 Catabolismo degli endocannabinoidi 8
Figura 1.5 Biosintesi e metabolismo degli endocannabinoidi 9
Figura 1.6 Rappresentazione schematica di cute sana 13
Figura 1.7 Percentuali dei casi di melanoma 14
Figura 1.8 Incidenza e mortalità di melanoma nel mondo 15
Figura 1.9 Prevalenza di mutazioni a carico del gene CDKN2A 17
Figura 1.10 Ciclo cellulare in cellule eucariote 19
Figura 1.11 Rappresentazione della via di segnale MAPK/ERK 20
Figura 1.12 Cellule iniziatrici di melanoma 22
Figura 3.13 Rappresentazione linea cellulare A375 25
Figura 3.14 Rappresentazione linea cellulare MeWo 26
Figura 3.15 Rappresentazione melanociti NHEM 27
Figura 3.16 Schema di estrazione dell’RNA 38
Figura 3.17 Meccanismo d’azione della sonda TaqMan 41
Figura 3.18 Schema dell’SDS page 46
Figura 3.19 Componenti del sandwich e camera di trasferimento 47
Figura 3.20 Trattamento della nitrocellulosa con i rispettivi anticorpi 49
Figura 3.21 Strumento ImageQuant LAS 4000 49
Figura 4.22 Espressione del recettore CB1 51
Figura 4.23 Espressione del recettore CB1 in cellule staminali di melanoma 52
Figura 4.24 Espressione NAPE-PLD e FAAH 54
Figura 4.25 Espressione DAGL-a, DAGL-b e MAGL 55
Figura 4.26 Espressione proteica del recettore CB1 in linee di melanoma 56
Figura 4.27 Espressione proteica del recettore CB1 e BRAF in linee starvate e non
starvate 57

76
Ringraziamenti
Desidero ringraziare il Professor S. Fogli, mio relatore, per la sua disponibilità e
pazienza, ma soprattutto professionalità, mostrata nei mie confronti durante tutto
il periodo di tesi.
Un sentito ringraziamento va a tutto il laboratorio di farmacologia, Dott.ssa Carpi,
nonché mia correlatrice, Dott.ssa Polini e Prof.ssa Nieri, che mi hanno supportato
ed aiutato nello sviluppo di questo lavoro fornendomi suggerimenti preziosi.
Infine desidero precisare e ringraziare l’importante collaborazione con l’ITT
(Istituto Toscano Tumori), in particolare la Dott.ssa Barbara Stecca, grazie ai
quali ci sono stati forniti elementi necessari per eseguire i nostri studi.