
CORSO DI GENETICA - urbigen.altervista.orgurbigen.altervista.org/didattica/lezioni/B15.pdf ·...
43
Roberto Piergentili Universit à di Urbino “Carlo Bo” CORSO DI GENETICA CONTROLLO DELL’ATTIVITÀ GENICA NEI PROCARIOTI
Transcript of CORSO DI GENETICA - urbigen.altervista.orgurbigen.altervista.org/didattica/lezioni/B15.pdf ·...

Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”
CORSO DI GENETICA
CONTROLLO DELL’ATTIVITÀGENICA NEI PROCARIOTI

Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”Trascrizione e traduzione
La trascrizione è il processo con cui l’informazione ereditaria vienetrasferita dal DNA all’RNA. La traduzione è il processo con cuil’informazione viene trasferita dall’RNA alle proteine. Latrascrizione viene effettuata dalla RNA polimerasi. Neiprocarioti esiste solo una RNA polimerasi che trascrive tutti igeni, mentre negli eucarioti ci sono 3 diverse RNA polimerasi:
• RNA polimerasi I per la sintesi dell’RNA ribosomale (rRNA)
• RNA polimerasi II per la sintesi dell’RNA messaggero (mRNA)
• RNA polimerasi III per la sintesi dell’RNA transfer (tRNA)
Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
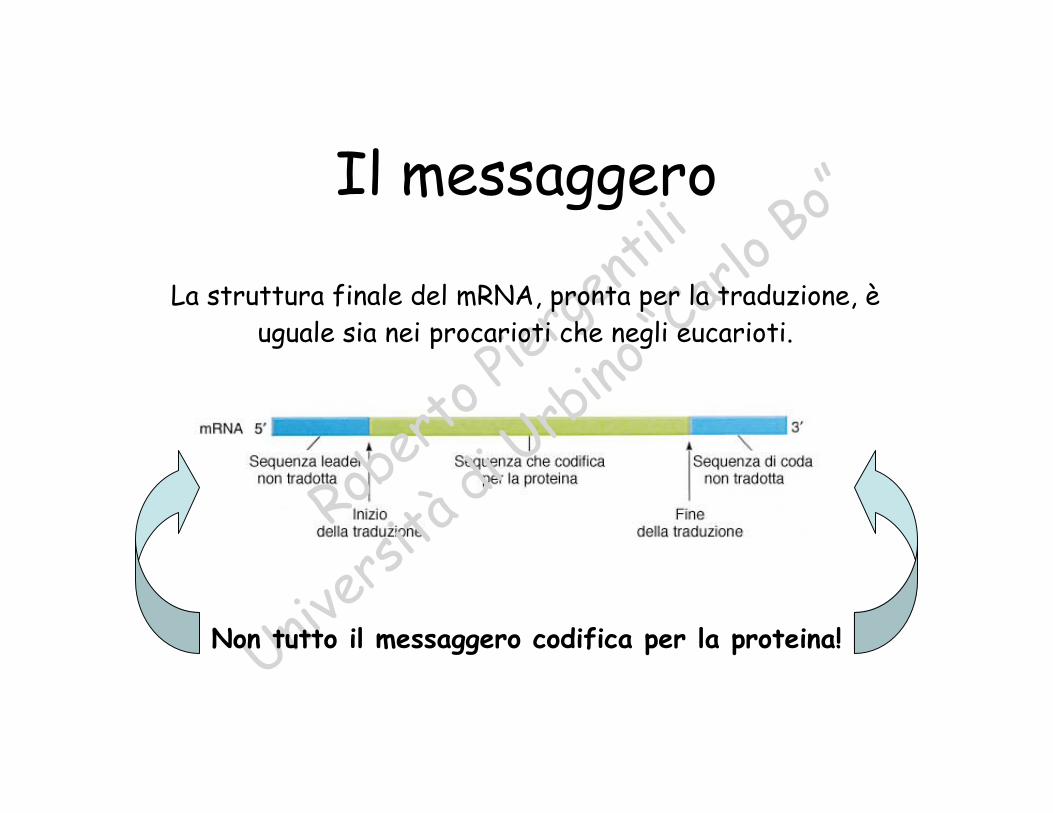
loBo”Il messaggero
La struttura finale del mRNA, pronta per la traduzione, èuguale sia nei procarioti che negli eucarioti.
Non tutto il messaggero codifica per la proteina!

Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”
Riassunto della traduzioneInizio della traduzione
Un ribosoma si attacca alla molecola di mRNA in corrispondenza dellatripletta di inizio AUG. Su questo codone AUG si lega il tRNA conl’anticodone corrispondente che porta la metionina (N-formil metioninanei procarioti).
Sintesi della proteina
Entra il tRNA corrispondente al secondo codone e si forma il legamepeptidico tra la metionina iniziale e il secondo amminoacido. Il ribosomasi sposta sull’mRNA permettendo che una sola tripletta alla volta siadisponibile all’attacco col tRNA specifico. Man mano che il ribosoma sisposta si ha l’allungamento della catena proteica.
Termine della traduzione
Quando il ribosoma raggiunge le triplette di stop (UAG,UAA,UGA) sistacca dall’mRNA.

Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”
Il tRNAEsistono meno di 64tRNA perché in alcunicasi lo stesso tRNA puòleggere più di un codoneche codifica per lo stessoamminoacido.
Ciò è possibile per ilvacillamento (wobbling) ooscillazione della terzabase dell’anticodone.

Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”
Accoppiamento della trascrizionee della traduzione nei procarioti

Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”Regolazione genica nei procarioti
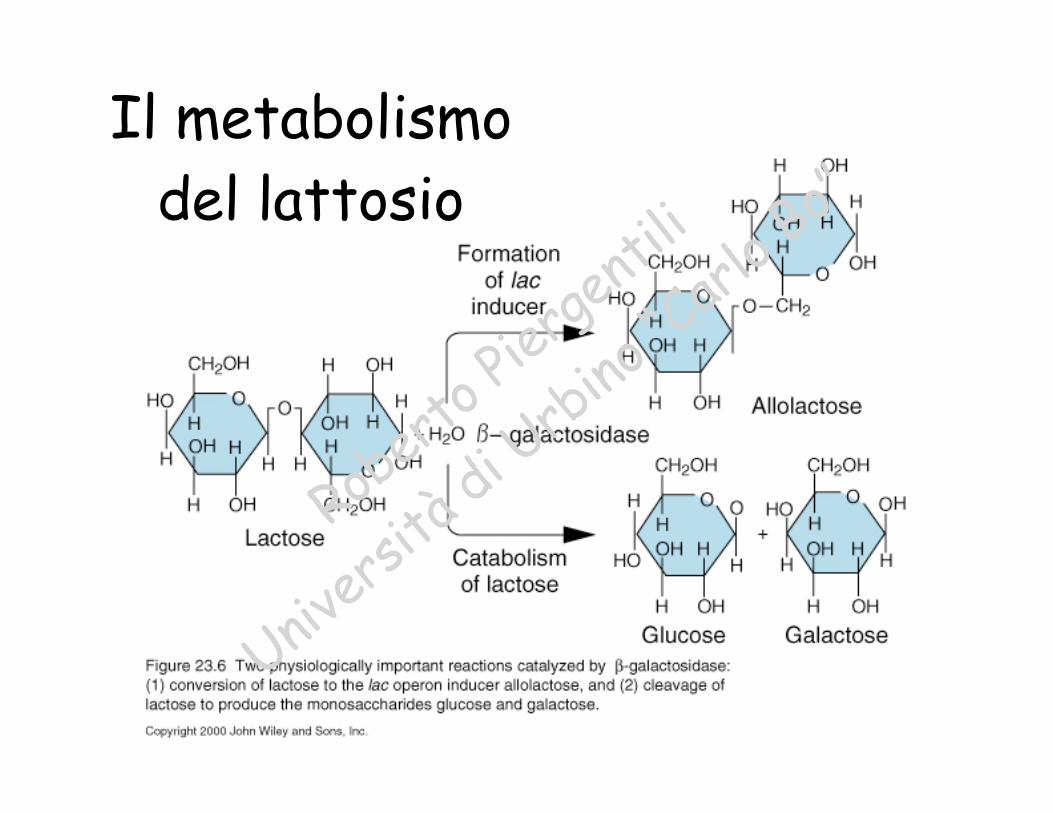
Il metabolismodel lattosio
Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”

Il metabolismodel lattosio èinducibile in
E. coli
Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”
Si parla di induzione se,aggiungendo il substrato,la sintesi enzimatica siinnalza.Si parla di repressione
se, aggiungendo ilsubstrato, la sintesienzimatica si abbassa.

I geni inducibili
Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”

Controllo positivo e negativo
Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”
Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”
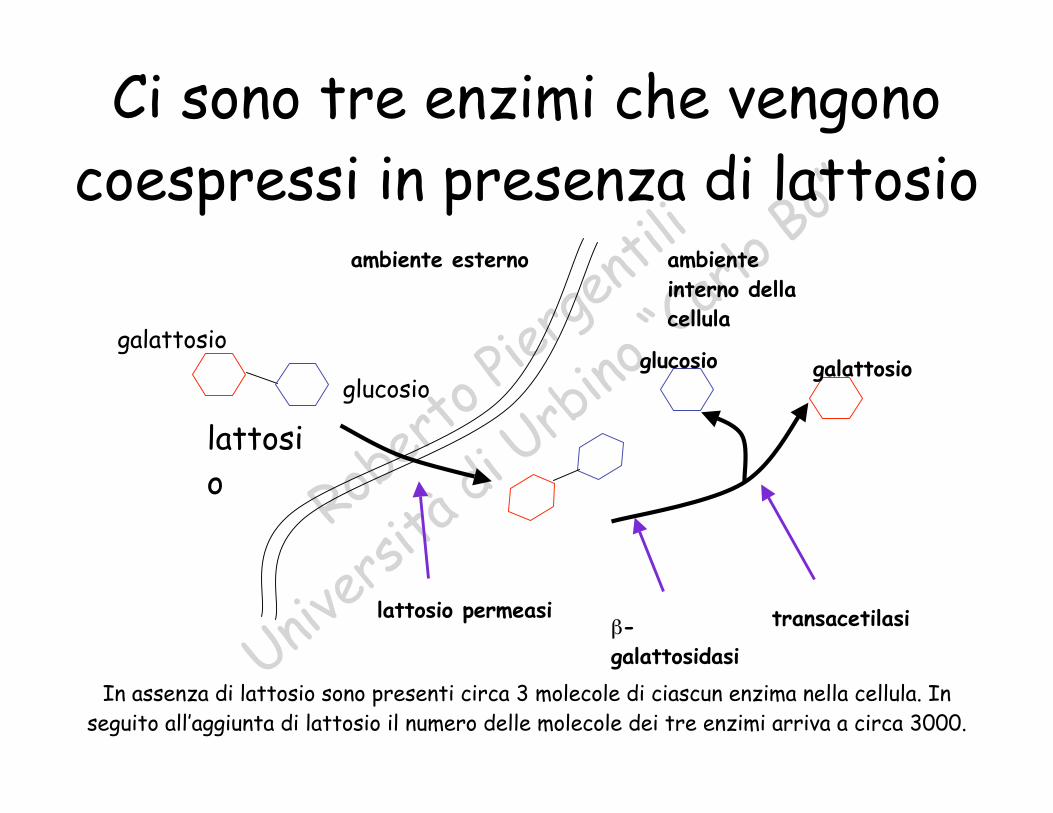
Ci sono tre enzimi che vengonocoespressi in presenza di lattosio
lattosio
galattosio
glucosio
ambiente
interno della
cellula
ambiente esterno
glucosio galattosio
lattosio permeasib-
galattosidasi
transacetilasi
In assenza di lattosio sono presenti circa 3 molecole di ciascun enzima nella cellula. Inseguito all’aggiunta di lattosio il numero delle molecole dei tre enzimi arriva a circa 3000.

Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”
La scopertadell’operone
From left to right: Francois Jacob (1920- ), JacquesMonod (1910-1976) and André Lwoff (1902-1994),awarded the Nobel Prize for Physiology or Medicinein 1965. Jacques Monod was director of the PasteurInstitute in Paris from 1971 to 1976.
Grazie all’uso sistematico di mutantiincapaci di metabolizzare il lattosioe al loro studio tramite creazione didiploidi parziali (sfruttando leproprietà di ceppi F’) tra il 1960 e il1964 F. Jacob, J. Monod e A. Lwoffidentificarono tre categorie dimutanti: o non producevano la b-gal,oppure la permeasi, oppure la
transacetilasi. Esistevano inoltredue tipi di mutazioni: quelle missensoa carico di ciscun gene che nondavano un prodotto funzionale delgene mutato, e quelle nonsenso checompromettevano la sintesi delleproteine a valle (mostravano effetti
polari), suggerendo che il mRNAfosse policistronico.

mRNA policistronici
Una mutazione non-sensoha effetto polare perchégiunto al primo codone distop il ribosoma si stacca enon procede nella tradu-zione dei geni a valle.
Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”

Lemutazioni
polari
Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”
Un individuo chesia eterozigote intrans viene anche
detto trans-eterozigote. Unacellula battericacontenente unesogenote è un
diploide parziale,o merozigote.

Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”
Altre mutazioni delmetabolismo del lattosio
Furono trovate anche mutazioni in altri geni che influenzavano lasintesi delle proteine b-galattosidasi (gene lacZ), permeasi (genelacY) e transacetilasi (gene lacA). Dato che queste mutazionialteravano la modalità di sintesi dei tre enzimi, Jacob Monod eLwoff supposero che fossero mutazioni in geni regolativi (i genilacZ, lacY e lacA sono invece detti geni strutturali).La caratteristica di questi geni regolativi consisteva nel fattoche, quando immessi nel ricevente tramite un F’, alcuni erano ingrado di agire anche in trans, mentre altri potevano esplicare laloro funzione soltanto in cis. Le mutazioni regolative furonomappate in due gruppi di complementazione che definiscono i geniO ed I.

Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”
La genetica dei geni regolatori
--Plac-
+*-ISQ
+*-IQ
++I-d
--Is
++I-
++Oc
induttorepresente
induttoreassente
mutante
* indotti da alte concentrazioni di lattosio
Sintesi di b-galattosidasi,permeasi e transacetilasi

Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”
Organizzazione dell’operone lac
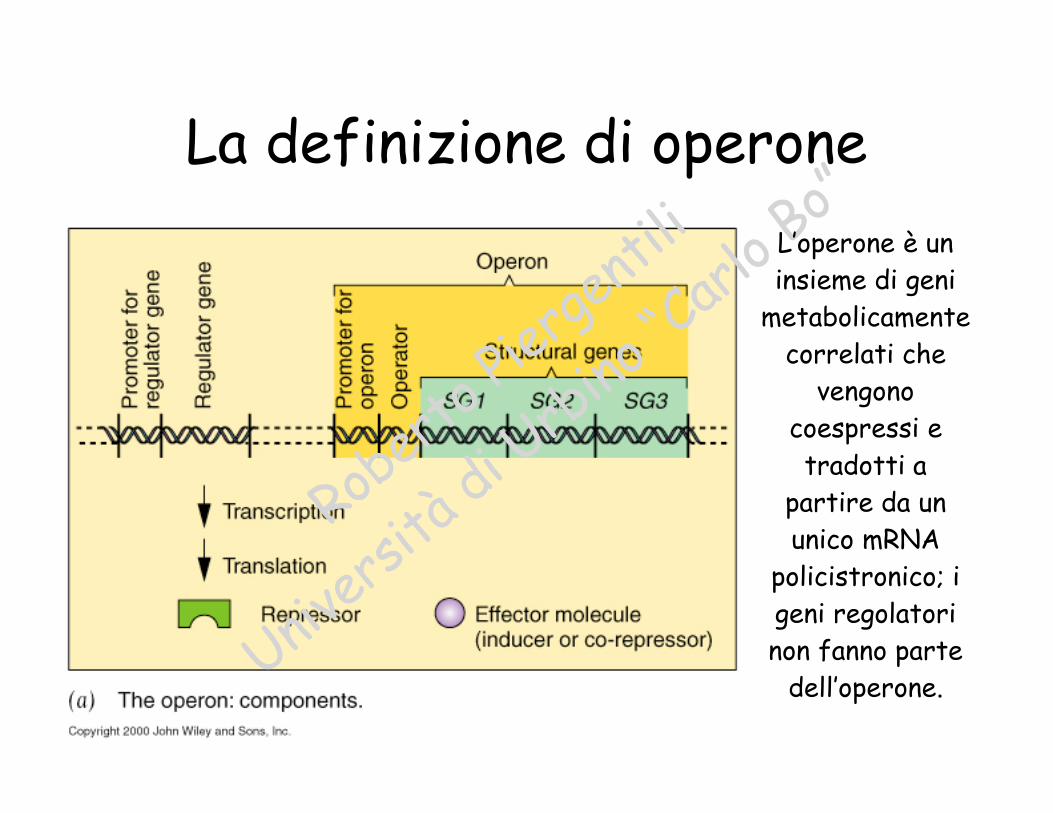
La definizione di operone
Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”
L’operone è uninsieme di geni
metabolicamentecorrelati che
vengonocoespressi etradotti a
partire da ununico mRNA
policistronico; igeni regolatorinon fanno parte
dell’operone.
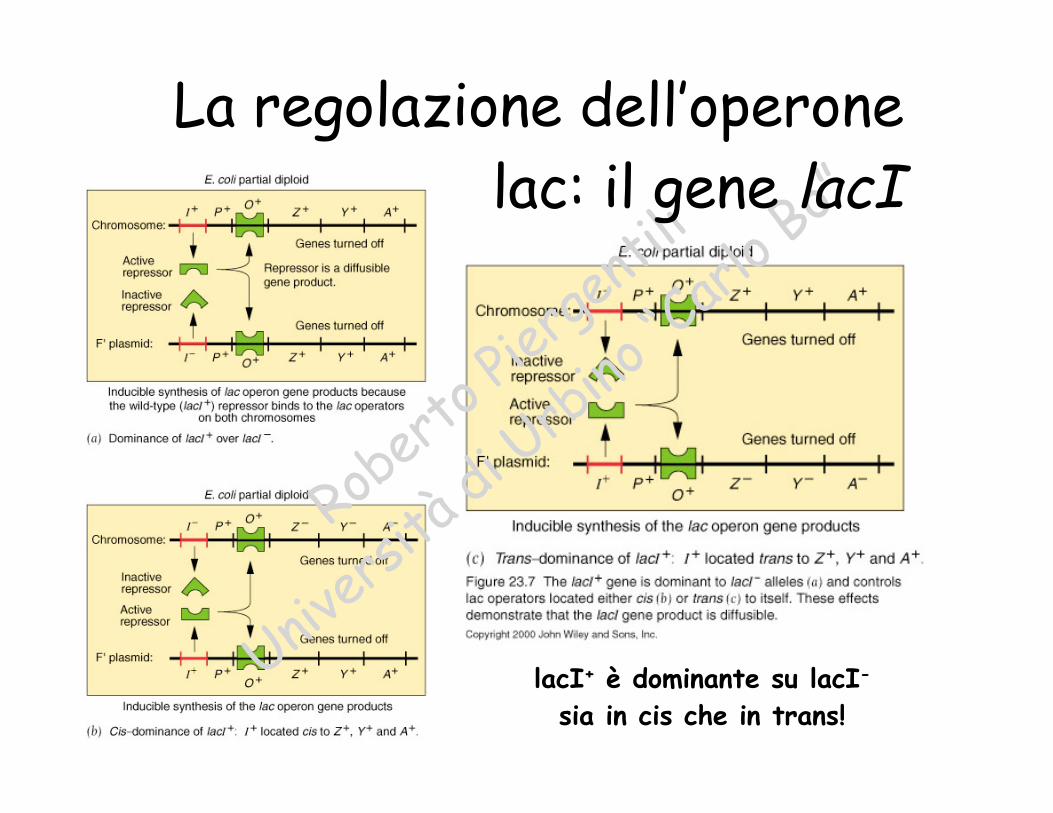
lacI+ è dominante su lacI-
sia in cis che in trans!
Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”
La regolazione dell’operonelac: il gene lacI
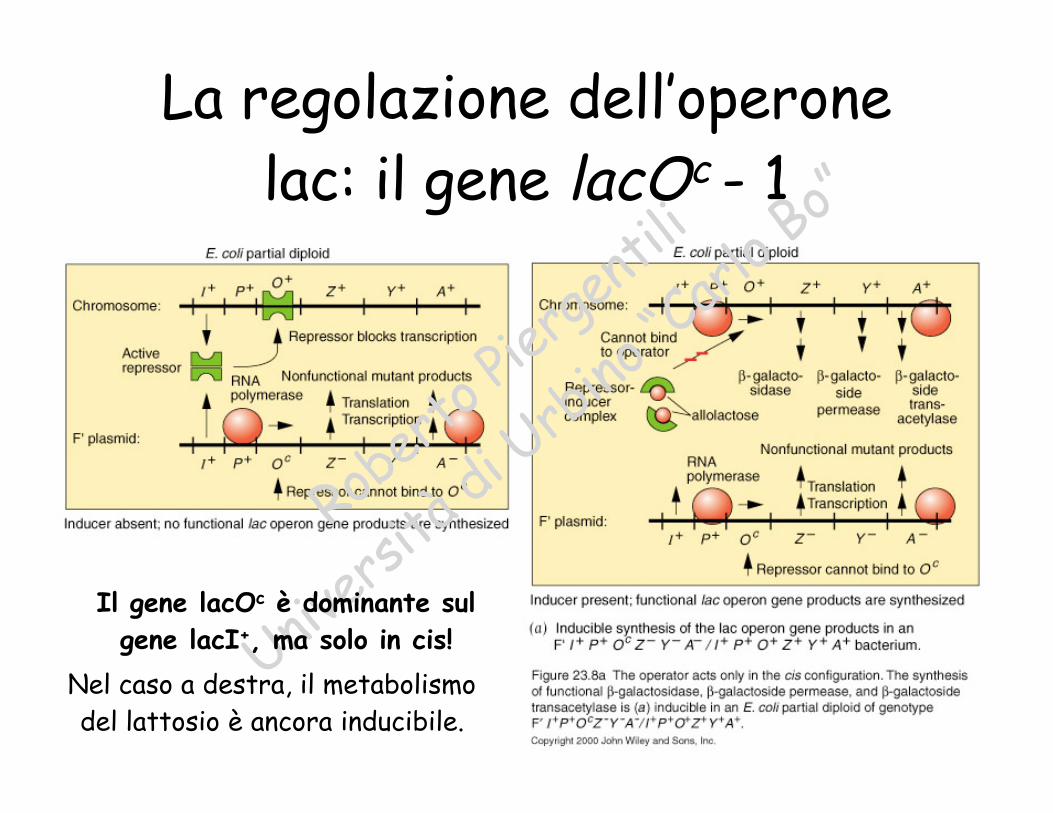
La regolazione dell’operonelac: il gene lacOc - 1
Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”
Il gene lacOc è dominante sul
gene lacI+, ma solo in cis!
Nel caso a destra, il metabolismodel lattosio è ancora inducibile.

Il metabolismo del lattosio questa volta NON è inducibile!
Laregolazione
dell’operone lac:
il genelacOc - 2
Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”

Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”
ConclusioniDa tutti questi dati si dedusse che I+ è trans dominante e siipotizzò che codificasse per una proteina diffusibile nel citoplasmain grado di influenzare anche i geni in trans, mentre per Oc sidedusse che era una mutazione cis dominante (in grado diinfluenzare solo i geni in cis) e che il gene O non producesse unaproteina diffusibile ma che fosse una sequenza di regolazione, chevenne chiamata operatore. Oc è un operatore costitutivo. Jacob,Monod e Lwoff ipotizzarono anche che il gene O fosse una sequenzadi DNA alla quale si lega il prodotto del gene I (la proteinarepressore). Il lattosio, legandosi al repressore, favorisce il suodistacco dal gene O consentendo alla polimerasi di legarsi alpromotore per iniziare la trascrizione dei geni a valle. L’operone lacè un modello di controllo negativo.

Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”
Ilmodello

lacI è una proteina allosterica
Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”

Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”La mutazione lacIS
È anche detta super
repressore. In questocaso il repressore èmutato nel sito di legamecon il lattosio, ma non nelsito di legame conl’operatore: il repressoreè quindi insensibile allattosio, per cui l’operoneè sempre “spento”.Is è dominante su I+.
* indotti da alteconcentrazioni di
lattosio
Sintesi di b-galattosidasi,permeasi e transacetilasi
--Plac-
+*-ISQ
+*-IQ
++I-d
--Is
++I-
++Oc
induttorepresente
induttoreassente
mutante

La mutazione lacI-d
--Plac-
+*-ISQ
+*-IQ
++I-d
--Is
++I-
++Oc
induttorepresente
induttoreassente
mutante
* indotti da alteconcentrazioni di
lattosio
Sintesi di b-galattosidasi,permeasi e transacetilasi
Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”
Nelle cellule aploidi le mutazioni I-d hanno fenotipo costitutivo. Neidiploidi parziali I+/I-d le mutazioni I-d sono trans dominanti su I+. Ilrepressore selvatico è un tetramero. La mutazione I-d produce unaproteina alterata che nei diploidi parziali I+/I-d si lega alle molecole“buone” impedendone il legame all’operatore.

Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”Le mutazioni IQ e ISQ
--Plac-
+*-ISQ
+*-IQ
++I-d
--Is
++I-
++Oc
induttorepresente
induttoreassente
mutante
* indotti da alteconcentrazioni di
lattosio
Sintesi di b-galattosidasi,permeasi e transacetilasi
Sono mutazioni a carico delpromotore del gene I edeterminano la produzionedi un gran numero dimolecole di repressore.Riducono l’efficienza diinduzione e sono indotti soloda alte concentrazioni dilattosio.

Le mutazioni P
--Plac-
+*-ISQ
+*-IQ
++I-d
--Is
++I-
++Oc
induttorepresente
induttoreassente
mutante
* indotti da alteconcentrazioni di
lattosio
Sintesi di b-galattosidasi,permeasi e transacetilasi
Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”Le mutazioni in P, sia nell’operone che nell’induttore, regolano il legame
della RNA polimerasi. Quindi se un operone è P- comunque non funziona,anche se tutto il resto è a posto.

Controllo positivo dell’operone lac
Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”
I geni lacZ, lacY e lacA sono espressi solo se nel terreno non è presenteil glucosio. Altrimenti questo zucchero viene usato preferenzialmenteanche in presenza di lattosio. In presenza di glucosio si ha larepressione da catabolita: il glucosio mantiene basso il livello di cAMP(AMP ciclico) impedendo la formazione del complesso CAP-cAMP.

Zuccheri nelterreno di
coltura
glucosio
glucosio+lattosio
lattosio
Quantitàrelativa di b-galattosidasi
1
50
2500
Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”
Il ruolo delcAMP
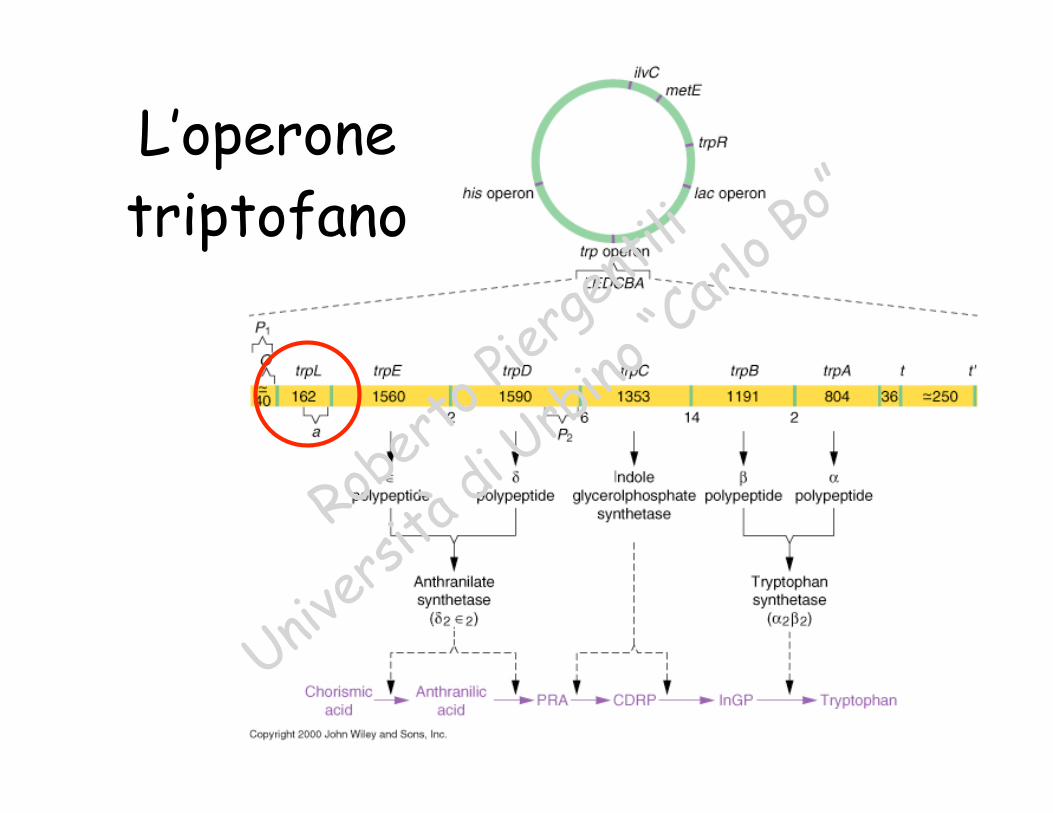
L’operonetriptofano
Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”

Il peptide leader
Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”
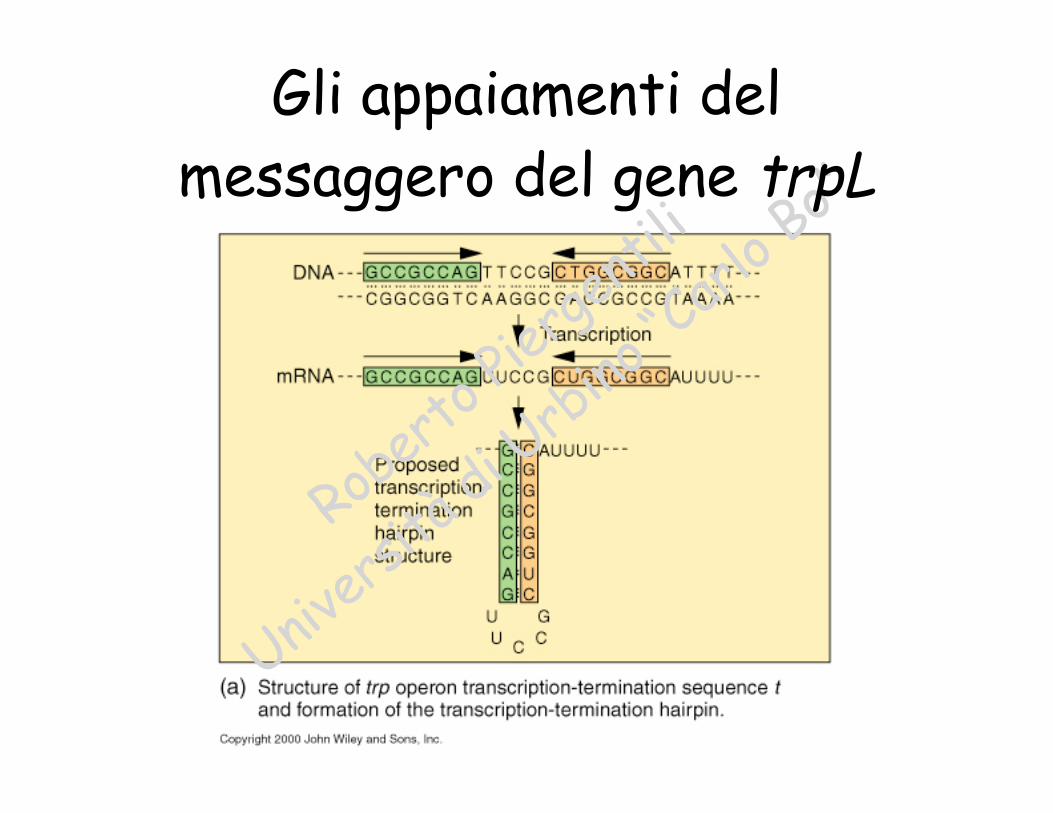
Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”
Gli appaiamenti delmessaggero del gene trpL

Le strutture a forcina
Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”
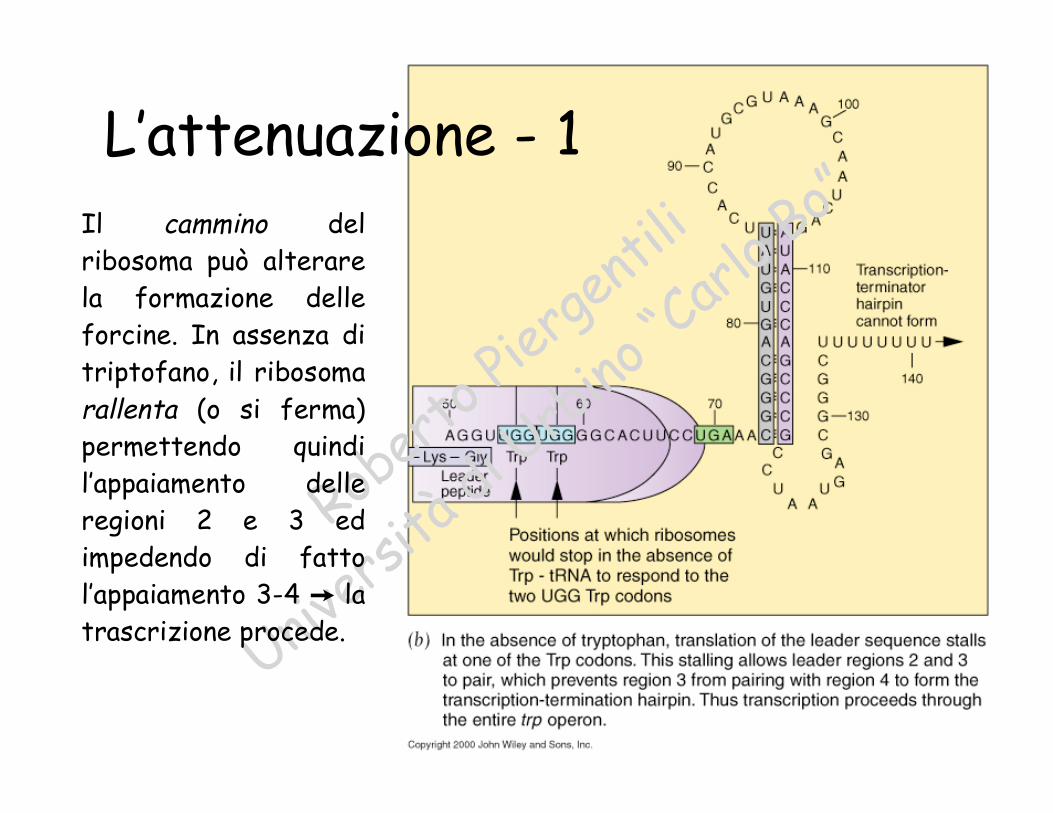
L’attenuazione - 1
Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”Il cammino del
ribosoma può alterarela formazione delleforcine. In assenza ditriptofano, il ribosomarallenta (o si ferma)permettendo quindil’appaiamento delleregioni 2 e 3 edimpedendo di fattol’appaiamento 3-4 Ÿ latrascrizione procede.

L’attenuazione - 2
Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”
In presenza ditriptofano, il ribosomanon si ferma e procedesulla zona 2 (da cui poisi stacca per lapresenza di un codonedi stop); in questomodo permette quindil’appaiamento 3-4:questa forcinadestabilizza la RNApolimerasi (ricordareche nei procarioti latrascrizione e latraduzione sonoaccoppiate!) e la
trascrizione si ferma.

Il fago l
Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”Il cromosoma di l esiste sia
nella forma lineare checircolare. La forma lineare(quella inattiva nel capside)favorisce l’iniezione del DNAnell’ospite. Una voltainiettato, il DNA passa allaforma trascrizionalmenteattiva, quella circolare.

profago quiescente
Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”
Ciclo liticoe lisogenicoIl fago l può riprodursi
sia attraverso ilclassico ciclo litico, sia
passare al ciclolisogenico. In questo
caso l’anello di DNA delfago ricombina con ilcromosoma batterico
(un crossing over)sempre nello stessopunto (ricordare la
trasduzionespecializzata!)

La repressione del ciclo litico
operatore destrooperatore sinistro
Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”

Il mantenimentodel ciclo
lisogenico
Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”
Il repressore agisce intrans e impedisce sia alfago di innescare ilproprio ciclo litico, siaad altri eventuali fagi ldi farlo (effettodominante). Inoltre,come profago, un solo l
è presente nella cellulaperché c’è un solo sitod’inserzione.

Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”
L’innesco del ciclo litico
Il repressore automantiene il sistema teoricamente all’infinito.L’unico modo per uscire dal ciclo lisogenico è l’eliminazione fisica delrepressore. Questo può avvenire tramite taglio proteolitico.La proteina RecA è una proteina batterica che viene innescata dalsistema SOS: questo sistema monitorizza continuamente l’integritàdel DNA batterico e se questo viene danneggiato promuove lasintesi di geni e l’attivazione di altri tramite tagli proteolitici. I geniattivati bloccano il ciclo cellulare e promuovono la riparazione delDNA tramite (anche) ricombinazione (da cui il nome di RecA).
Tra i bersagli degli enzimi proteolitici del sistema SOS c’è anche ilrepressore di l: quando la nave affonda, i topi scappano…

L’innesco del ciclo litico
Rober
to Pi
erge
ntili
Univer
sità di U
rbino
“Car
loBo”


















