COMPOSTI AD ALTA ENERGIA - … · 1,3-bifosfoglicerato fosfoenolpiruvato fosfocreatina ATP. Title:...
23
COMPOSTI AD ALTA ENERGIA
Transcript of COMPOSTI AD ALTA ENERGIA - … · 1,3-bifosfoglicerato fosfoenolpiruvato fosfocreatina ATP. Title:...

COMPOSTI AD ALTA ENERGIA

ENTALPIA: misura della variazione di energia di una reazione condotta a pressione costante
∆H = ∆E + P∆V
ENTROPIA: misura del grado di disordine
S = k lnW (k=cos di Boltzmann, cioè la cost dei gas R/n. di Avogadro; W numero di sottostati di uguale energia)

L’ENERGIA LIBERA DI GIBBS
G = H-TS
Questa funzione di stato tiene conto sia del termine entalpico(H), che misura la variazione di energia a pressione costante, sia di quello entropico (S), che valuta l’importanza del grado di disordine.
Per una variazione di ENERGIA LIBERA (∆G) in un sistema a temperatura e pressione costante possiamo scrivere:
∆G = ∆H - T∆S
Una diminuzione di energia (∆H negativo) e/o un aumento di entropia (∆S positivo) sono tipici delle trasformazioni favorite, e entrambe queste condizioni tenderanno a rendere il ∆G negativo.
Un processo in un sistema isolato a pressione e temperatura costanti è favorito se il ∆G è NEGATIVO!!!

Le trasformazioni accompagnate da variazioni negative dell’energia libera si dicono ESOERGONICHE e sono termodinamicamente favorite, mentre le trasformazioni accompagnate da variazioni positive dell’energia libera si dicono ENDOERGONICHE e sono termodinamicamentesfavorite (è favorita la reazione inversa).
Se ∆H = T∆S allora ∆G = 0
Allora la trasformazione non è favorita né in un senso nénell’altro: il sistema è all’equilibrio. In questa condizione la reazione è reversibile, cioè, può essere spostata in qualsiasi direzione da una spinta infinitesimale in un senso o nell’altro.
Il segno del ∆G di un processo indica se questo processo, oppure il suo contrario, sia termodinamicamente favorito.
Il valore assoluto del ∆G è indicazione di quanto il processo sia lontano dall’equilibrio.

Molte reazioni necessarie per la vita sono ENDOERGONICHE (ad es.trasporto contro gradiente di concentrazione), tali reazioni intrinsecamente sfavorite possono essere rese termodinamicamentefavorite accoppiandole a reazioni fortemente esoergoniche.
A B ∆G = + 10 kJ/mole
C D ∆G = -30 kJ/mole
se la cellula riuscirà ad accoppiare queste due reazioni il ∆G per il processo complessivo sarà dato dalla somma algebrica dei singoli valori :
A B ∆G = + 10 kJ/mole
C D ∆G = -30 kJ/mole
A + C B + D ∆G = -20 kJ/mole
L’equilibrio del processo complessivo risulta ora molto spostato a destra.
B risulta quindi efficientemente prodotto a partire da A.
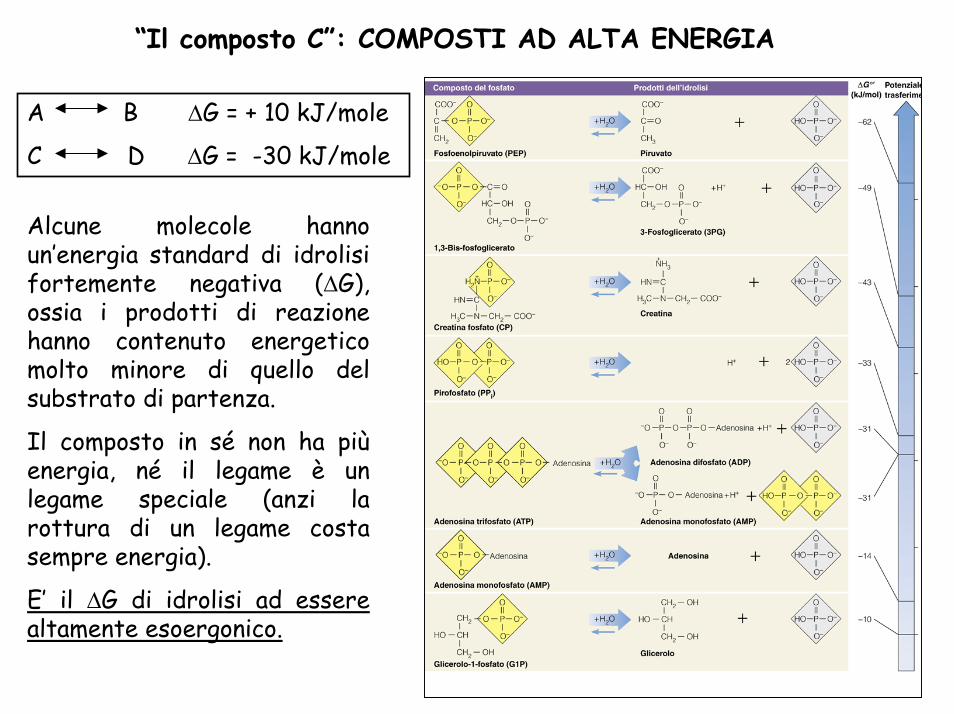
“Il composto C”: COMPOSTI AD ALTA ENERGIA
Alcune molecole hanno un’energia standard di idrolisi fortemente negativa (∆G), ossia i prodotti di reazione hanno contenuto energetico molto minore di quello del substrato di partenza.
Il composto in sé non ha piùenergia, né il legame è un legame speciale (anzi la rottura di un legame costa sempre energia).
E’ il ∆G di idrolisi ad essere altamente esoergonico.
A B ∆G = + 10 kJ/mole
C D ∆G = -30 kJ/mole

FATTORI CHE CONTRIBUISCONO A RENDERE negative LE ∆G DI IDROLISI DEI COMPOSTI A ALTA ENERGIA
1. Delocalizzazione degli elettroni del legame O-P:
la stabilizzazione per risonanza del Pi comporta che i prodotti siano + stabili dei reagenti, cioèa uno stato energetico inferiore. Le diverse forme di risonanza (W) fanno aumentare l’entropia del sistema (S = k lnW) che influisce sul ∆G (∆G = ∆H - T∆S) facendolo aumentare.
2. Ulteriore idratazione dei prodotti di idrolisi: il rilascio del Pi aumenta le possibilità di idratazione, specialmente quando i prodotti sono carichi. Tale idratazione è uno stato energeticamente favorito.
3. Repulsione elettrostatica tra prodotti carichi: nella maggior parte dei casi l’idrolisi di tali molecole a alta energia genera prodotti carichi, la repulsione tra questi prodotti ionici favorisce fortemente la reazione di idrolisi.
4. Aumento di stabilizzazione per risonanza delle molecole prodotte: in alcuni casi non èsolo il Pi che si stabilizza per risonanza ma anche la restante parte della molecola (PEP→ piruvato).

Le categorie di composti ad alta energia più importanti sono due:
TIOESTERI
COMPOSTI FOSFORILATI

TIOESTERE
C H 3 CO
S R
C H 3 CO
O RC H 3 C
O
O -
-ESTERE NORMALE
Stabilizzazione per risonanza
∆G di idrolisi del tioestere
Stabilizzazione dell’estere per risonanza
∆G di idrolisi dell’estere normale
I tioesteri rispetto agli esteri normali hanno un livello energetico più alto.
Negli esteri normali vi è una delocalizzazione degli elettroni π, che conferisce carattere di parziale doppio legame al C-O. Nei tioesteri le maggiori dimensioni dell’S riducono il grado di delocalizzazione, risulta quindi ridotto il grado di stabilizzazione per risonanza→ ∆G di idrolisi maggiore.

Acetil-CoA + H2O acetato + CoA + H+ (∆G°= -32,2 kJ/mole)
CH3 CSCoA
O
H2OCoASH
CCH3
O
OH
H+
O-
O
CH3 C
Stabilizzazione per risonanza
Acetil-CoA
IDROLISI
Acido Acetico
IONIZZAZIONE
Acetato

I composti fosforilati presenti nelle cellule possono essere distinti in due gruppi in base alla loro energia libera di idrolisi:
A BASSA ENERGIA (∆G < -25 kj/mole)
A ALTA ENERGIA (∆G > -30 kj/mole)

MOLECOLE AD ALTA ENERGIA E RELATIVI ∆G
-31.4Acetil-CoA-9.2Glicerolo-1-P-13.8Glucosio-6-P-15.9Fruttosio-6-P-20.9Glucosio-1-P-33.4PPi(→2Pi)-14.2AMP (→adenosina+ Pi)-32.2ATP(→AMP+ PPi)
ATP(→ADP+ Pi) -30.5ADP (→AMP+ Pi)
-43.0fosfocreatina-49.31,3bifosfoglicerato (→3fosfoglicerato +Pi)-61.9Fosfoenolpiruvato
kJ/moleCOMPOSTO FOSFORILATO

Il PEP, quando viene idrolizzato, genera piruvato che esiste in due forme tautomeriche (enolica e chetonica) molto più stabili del substrato iniziale.
PEP3- + H2O piruvato + Pi (∆G°= -61,9 kJ/mole)
PEPPIRUVATO
forma enolicaPIRUVATO
forma chetonica
idrolisitautomerizzazione
-
O- C
O
CO
CH2
PO
OO H2O
P I
CH2
OHC
OO- C O- C
O
C
CH3
O
- -
-

ATP: adenosina trifosfato
Un discorso speciale merita la molecola di ATP che pur avendo un ∆G di idrolisi meno negativo è il composto ad alta energia universalmente utilizzato nei sistemi biologici per le reazioni accoppiate, le biosintesi, il lavoro meccanico, osmotico, etc…

ATP 4- + H2O ADP3- + Pi2- + H+ (∆G° = -30,5 kJ/mole)
ADP 3-
PO-
O
O Rib Ade ninaPO-
OO-
P
O
O
O O
H2O
OO P
O
OO- P
O-
O
Ade ninaPO-
O
O RibPO-
OO-
OH
O-
- 3-
H+ O-P
O
O-
PO-
O
O Rib Ade ninaO- P
O
OOH+
ATP 4-
ADP 2-
Stabilizzazione per risonanza
Idrolisi con rimozione della repulsione tra le cariche
ionizzazione
H
La reazione di idrolisi del legame fosfoanidridico genera due prodotti: ADP e Pi. Il primo si ionizza rilasciando un protone, il secondo si stabilizza per risonanza.


∆G TEORICO E REALE
I ∆G di idrolisi delle molecole considerate differiscono dai ∆G reali perché all’interno della cellula la concentrazione di ATP, ADP e Pi sono molto inferiori alla concentrazione standard 1M rispetto alla quale il ∆G° è calcolato.
Anche il pH può non essere 7 e la temperatura può non essere 25°C, tali valori dipendono in genere dalla specie considerata.
Equazione che descrive il ∆G reale di idrolisi dell’ATP, considerando il pH=7:
∆G = ∆G° + RT ln [ADP] [Pi ] / [ATP]
Anche ammettendo pH 7 e T=25°C la ∆G reale in base alle concentrazioni passa da -30.5 kj/mole a -51.8 kj/mole nel globulo rosso.
Pertanto il ∆Greale varia da una cellula all’altra e nel tempo in base alle condizioni metaboliche.

Se poi si corregono T, pH e presenza di ioni si hanno ulteriori fluttuazioni: nel caso dell’ATP, ad es., il Mg++ può mascherare cariche negative, modificando dei gruppi fosforici.
∆Greale per ATP: -50 kj/mole -65 kj/mole
P
O
O
O O R ib A d e n in aP
O
O
O O P
O
O
O O
M g + +
P
O
O
O O P
O
O
O O R ib
M g + +
A d e n in a
Mg ATP2-
Mg ADP-

Generalmente l’idrolisi dell’ATP nelle reazioni accoppiatecomprende 2 fasi:
1.a) trasferimento del gruppo Pi sul substrato o dell’AMPsul substrato o enzima.
ATP ADP + Pi (caso I)
ATP AMP + PPi (caso II)
1.b) conseguente aumento di E libera (∆G) di substrato o enzima.
2. Rilascio Pi o AMP.
Esistono tuttavia casi in cui la scissione dell’ATP in sèfornisce l’energia atta a far cambiare conformazione ad es. ad una proteina carrier.
ATP
ADP
NH3
ATP ADP P ICOO-
CHNH3+
CH2
CH2
C
O NH2
COO-
CHNH3+
CH2
CH2
CO-
O
COO-
CHNH3+
CH2
CH2
C
O
PI
NH3
O OP
O-O-
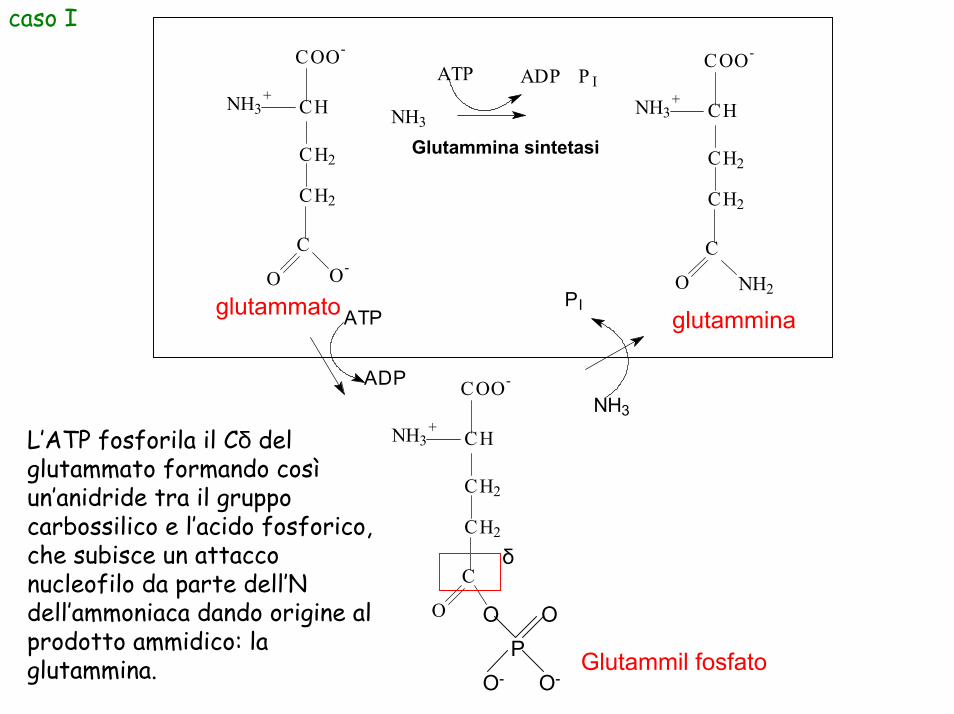
glutammato glutammina
Glutammil fosfato
Glutammina sintetasi
δ
L’ATP fosforila il Cδ del glutammato formando cosìun’anidride tra il gruppo carbossilico e l’acido fosforico, che subisce un attacco nucleofilo da parte dell’N dell’ammoniaca dando origine al prodotto ammidico: la glutammina.
caso I
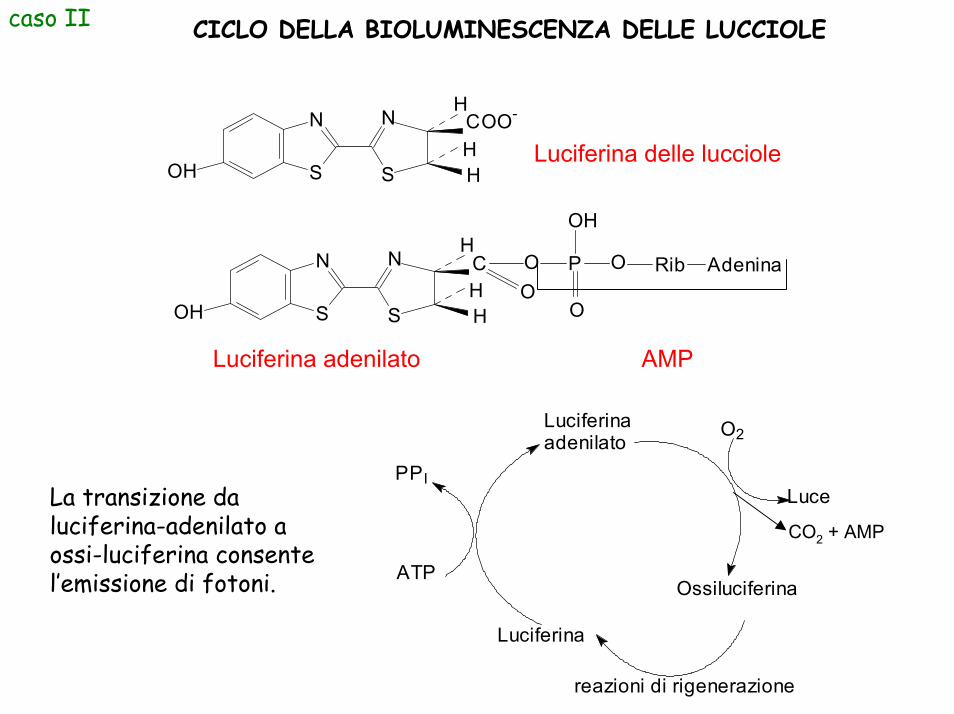
caso II CICLO DELLA BIOLUMINESCENZA DELLE LUCCIOLE
OH
N
S
N
S
HCOO-
HH
HHC
H
OH
N
S
N
SOO P
OH
O
O Rib Adenina
PPI
ATP
Luciferina adenilato
Ossiluciferina
Luciferina
O2
Luce
reazioni di rigenerazione
Luciferina delle lucciole
Luciferina adenilato AMP
CO2 + AMP
La transizione da luciferina-adenilato a ossi-luciferina consente l’emissione di fotoni.

COMPOSTI FOSFORILATI A BASSA ENERGIA
I composti fosforilati a potenziale più negativo possono trasferire il gruppo fosforico all’ADP per dare ATP il quale può trasferirlo ai prodotti di idrolisi (es.fruttosio) dei composti a ∆G meno negativo (es. a dare F6P).
I composti fosforilati a “bassa energia” sono tali perché i loro prodotti di idrolisi non possono avere stabilizzazioni per risonanza (sono alcooli).
-CH2OPi CH2
OH
In questo contesto l’ATP si comporta come una moneta di scambio universale.

CCHOH
CH2 O P
O O PCCH2
COO-
O P
P CREATINA
ADENINA Rib P P P
Glucosio Glicerolo
-70
-60
-50
-40
-30
-20
-10
∆G
6P P
Pi
Composti ad alta energia
Composti ad bassa energia
1,3-bifosfoglicerato fosfoenolpiruvato
fosfocreatina
ATP