![h v ] À ] P o ] ^ µ ] ] P o ] ] XZ X v X ó ð ñ o ï ...people.unica.it/orientamento/files/2017/06/2017-18_manifesto_studi.pdf · h v ] À ] P o ] ^ µ ] ] P o ] ] XZ X v X ó](https://static.fdocumenti.com/doc/165x107/5c65c64909d3f2826e8d18e9/h-v-a-p-o-p-o-xz-x-v-x-o-d-n-o-i-h-v-a-p-o.jpg)
Analisi delle correnti di Na secondo Hodgkin e Huxley · Pertanto, la corrente ionica I Na può...
30
Analisi delle correnti di Na secondo Hodgkin e Huxley Prof. Mauro Toselli Dip. Biologia e Biotecnologie Università di Pavia Scuola di Fisiologia e Biofisica 2015 Società Italiana di Fisiologia Pavia, 30/6/2015
Transcript of Analisi delle correnti di Na secondo Hodgkin e Huxley · Pertanto, la corrente ionica I Na può...

Analisi delle correnti di Na
secondo Hodgkin e Huxley Prof. Mauro Toselli
Dip. Biologia e Biotecnologie
Università di Pavia
Scuola di Fisiologia e Biofisica 2015
Società Italiana di Fisiologia
Pavia, 30/6/2015

La corrente di Na
ENa=+52 mV

Lo scopo di misurare le correnti di Na è quello di poter ottenere da esse
informazioni circa la permeabilità della membrana allo ione Na+ (ovvero circa il
comportamento dei canali ionici del Na).
RTVzF
RTVzFCC
RT
VFPzI ei
/exp1
/exp22
DOMANDA:
È lecito assumere che per la corrente di Na valga la legge di Ohm e che quindi INa
possa essere messa in rapporto con la conduttanza gNa secondo la relazione
INa = gNa(Vm-ENa)
anziché con la permeabilità PNa, secondo l’assai più complicata equazione di GHK
per la corrente?
Perché misurare le correnti di Na?
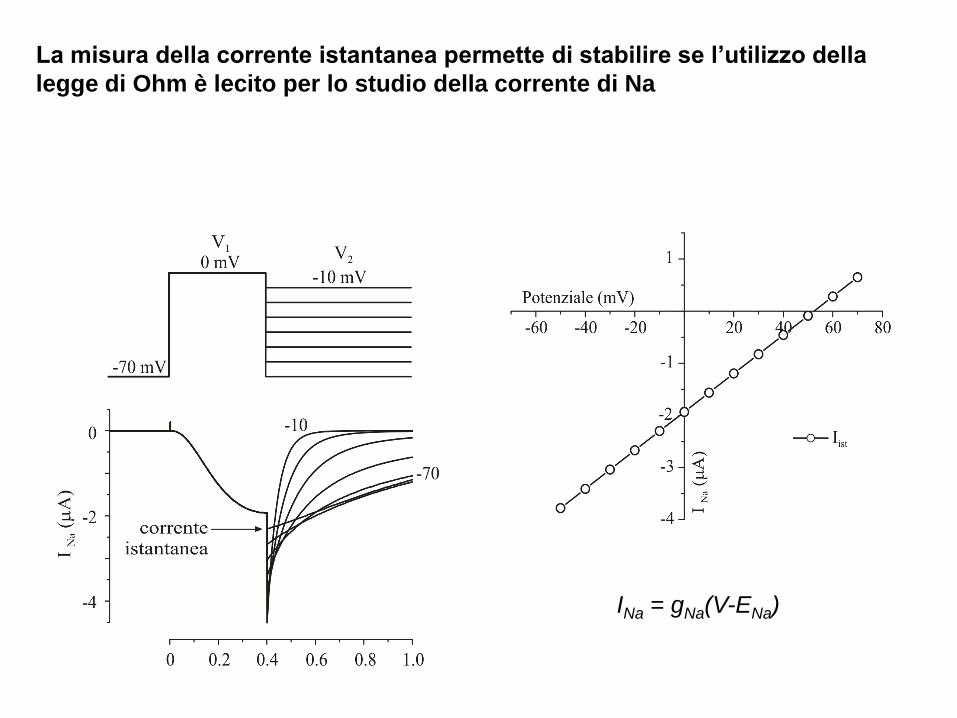
La misura della corrente istantanea permette di stabilire se l’utilizzo della
legge di Ohm è lecito per lo studio della corrente di Na
INa = gNa(V-ENa)

La relazione I/V istantanea per il Na è approssimativamente lineare, come previsto
dalla legge di Ohm
Pertanto gNa può essere utilizzata come misura della permeabilità della membrana
al sodio ovvero della frazione di canali del Na aperti.
INa = gNa(V-ENa) gNa = INa/(V-ENa)

Il modello delle gates secondo Hodgkin e Huxley
Diagramma schematico di un canale voltaggio-dipendente rappresentato come
una proteina transmembranaria con un poro acquoso. Sono indicate le regioni
funzionali essenziali: la gate, il sensore del voltaggio e il filtro di selettività.

La proteina-canale va incontro a cambiamenti conformazionali in risposta ad una
variazione del campo elettrico a cavallo della membrana.
In conseguenza di tali cambiamenti conformazionali la proteina-canale passerà da
uno stato chiuso, impervio agli ioni, ad uno stato aperto pervio agli ioni, o vice
versa.
Qualora la transizione tra i due stati chiuso e aperto sia regolata da un’unica gate,
ovvero la transizione avvenga tramite un unico cambiamento conformazionale
della proteina-canale, lo stato di apertura (o chiusura) del canale si identifica
con lo stato in cui si trovano la gate e la particella di gating (sensore del
voltaggio) in quell’istante.
Ciò significa che per un canale che possiede un’unica gate di attivazione la
reazione tra stato chiuso e aperto del canale è esprimibile nel seguente modo:
Assunzioni di base per il modello delle gates
1
p(V)
p1 -(V)
p(V)
p1 -(V)
C O ovvero

Assunzioni di base per il modello delle gates
2
La reazione tra stato non permissivo (chiuso) e permissivo (aperto) della
singola gate segue una cinetica del primo ordine,
ed è quindi descrivibile dalla seguente equazione differenziale:
ppdt
dp)1(
Basato su queste assunzioni, il modello delle gates fornisce una
descrizione sia della voltaggio-dipendenza che della tempo-dipendenza
dei canali ionici.
C O

Quando il potenziale di membrana verrà mantenuto ad un certo valore per un
tempo sufficientemente lungo, la reazione cinetica raggiungerà l’equilibrio (o
stato stazionario). Allo stato stazionario sarà:
0)1( ppdt
dp
)()(
)()(
VV
Vtpe quindi:
Invece, l’andamento temporale con cui lo stato stazionario verrà raggiunto sarà
descritto dalla seguente funzione esponenziale, che è una soluzione
dell’equazione differenziale di partenza t
ppptp o exp)()(
Dove p∞ è il valore di p allo stato stazionario e po è il valore di p a t=0 e in cui la
costante di tempo è in relazione con le due costanti di velocità e nel
seguente modo:
)()(
1
VV
LA REAZIONE ALL’EQUILIBRIO
CINETICA DELLA REAZIONE
po
p∞

Pertanto, ricavando sperimentalmente i valori di p∞(V) e di (V), è possibile ottenere le
costanti di velocità (V) e (V), che ci permettono di descrivere quantitativamente le
caratteristiche di tempo-dipendenza e voltaggio-dipendenza del canale.
RIASSUMENDO:
)()(
)(
VV
Vp
)()(
1
VV
/p /)1( p
OVVERO:
C O

I concetti appena espressi verranno ora utilizzati per
una analisi quantitativa del canale del Na da un punto
di vista biofisico
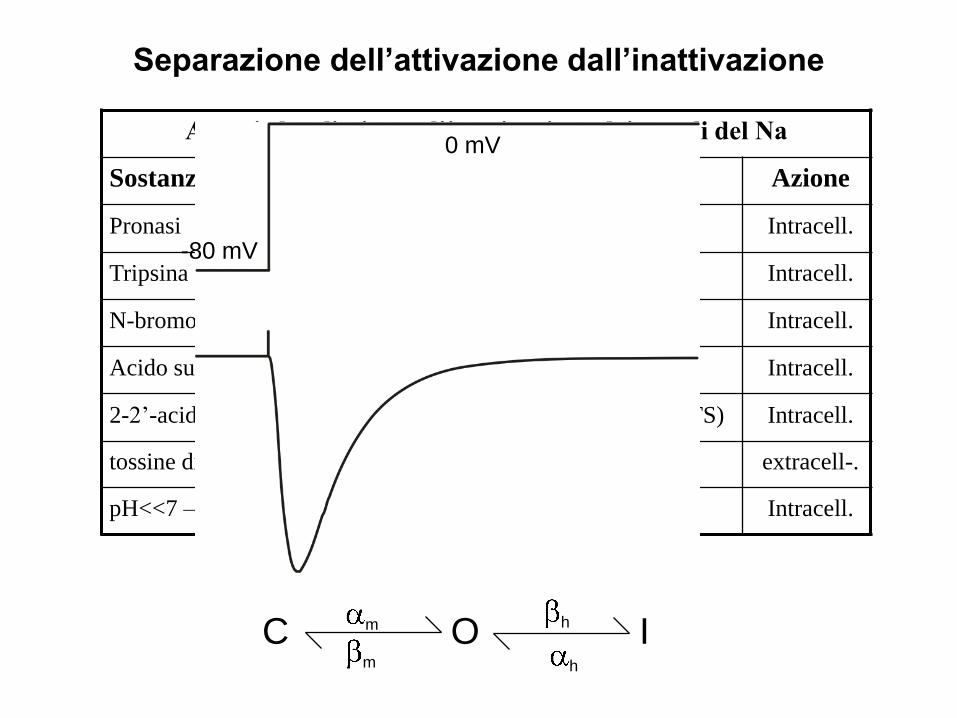
Separazione dell’attivazione dall’inattivazione
Agenti che eliminano l’inattivazione dei canali del Na
Sostanza Azione
Pronasi Intracell.
Tripsina Intracell.
N-bromoacetamide (NBA) Intracell.
Acido sulfonico 2,4,6-trinitrobenzene (TNBS) Intracell.
2-2’-acido disulfonico 4-acetamide-4-isotiocianostilbene (SITS) Intracell.
tossine di scorpione e celenterato extracell-.
pH<<7 – pH>>7 Intracell.
h
h
m
m
C O I
-80 mV
0 mV
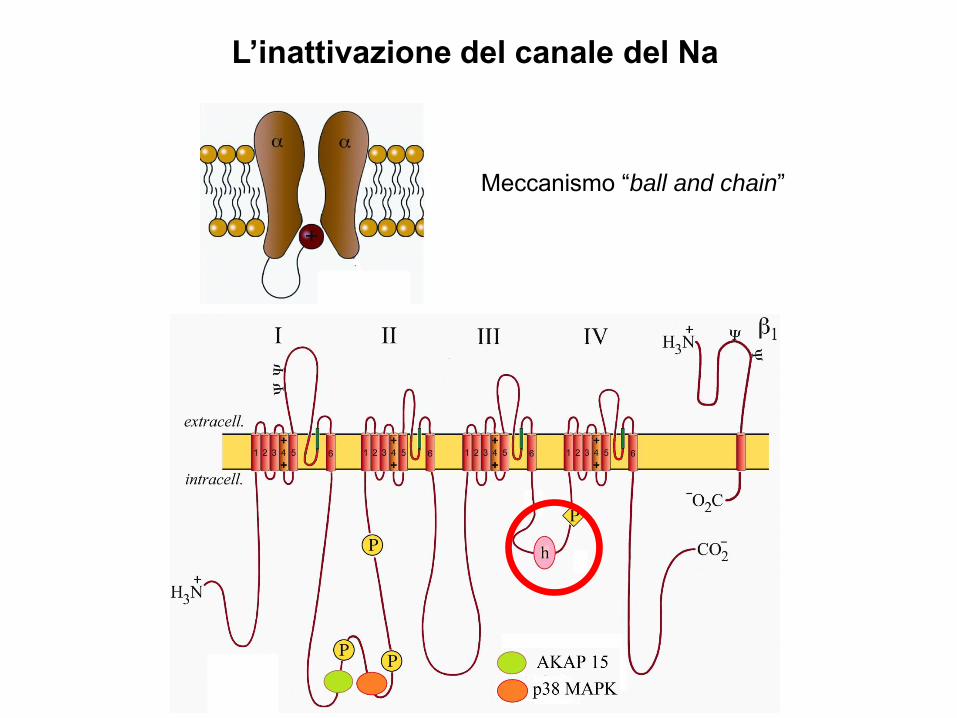
L’inattivazione del canale del Na
Meccanismo “ball and chain”
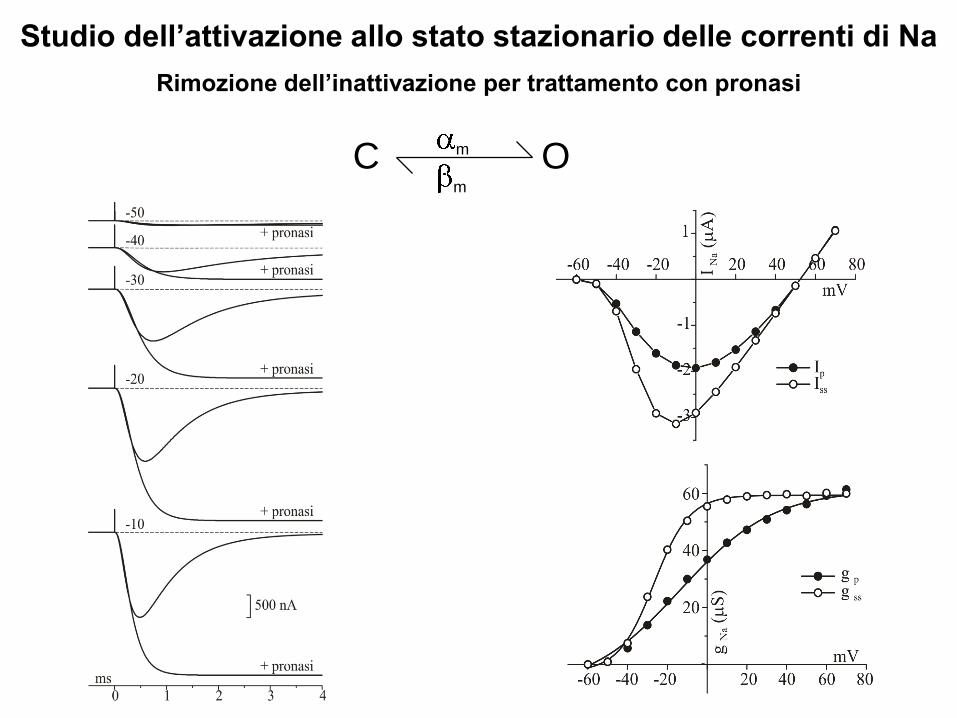
Studio dell’attivazione allo stato stazionario delle correnti di Na
Rimozione dell’inattivazione per trattamento con pronasi
m
m
C O
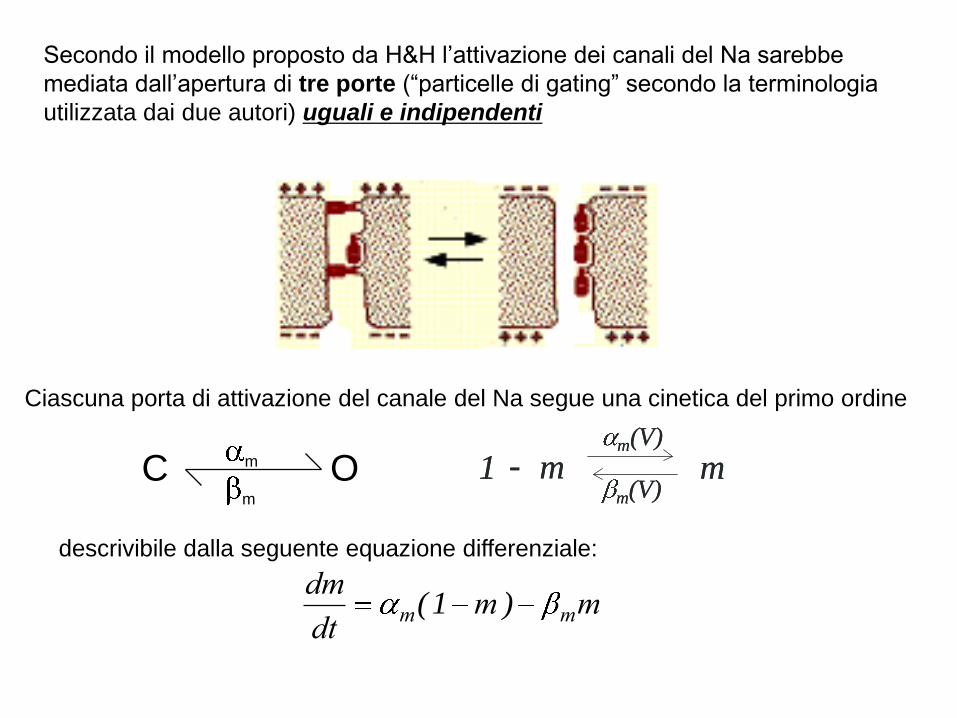
Ciascuna porta di attivazione del canale del Na segue una cinetica del primo ordine
mm(V)
m1 -m(V)
mm(V)
m1 -m(V)
descrivibile dalla seguente equazione differenziale:
mmdt
dmmm )1(
Secondo il modello proposto da H&H l’attivazione dei canali del Na sarebbe
mediata dall’apertura di tre porte (“particelle di gating” secondo la terminologia
utilizzata dai due autori) uguali e indipendenti
m
m
C O

Dove m∞ è il valore di m allo stato stazionario e mo è il valore di m a t=0 e in cui
la costante di tempo m sono in relazione con le due costanti di velocità m e m
nel seguente modo:
)()(
1
VV mm
m
Mentre, l’andamento temporale con cui lo stato stazionario verrà raggiunto
sarà descritto dalla seguente funzione esponenziale, che rappresenta una
soluzione dell’equazione differenziale
m
o
tmmmtm exp)()(
La soluzione di tale equazione allo stato stazionario (t→∞) è:
)()(
)(
VV
Vm
mm
m
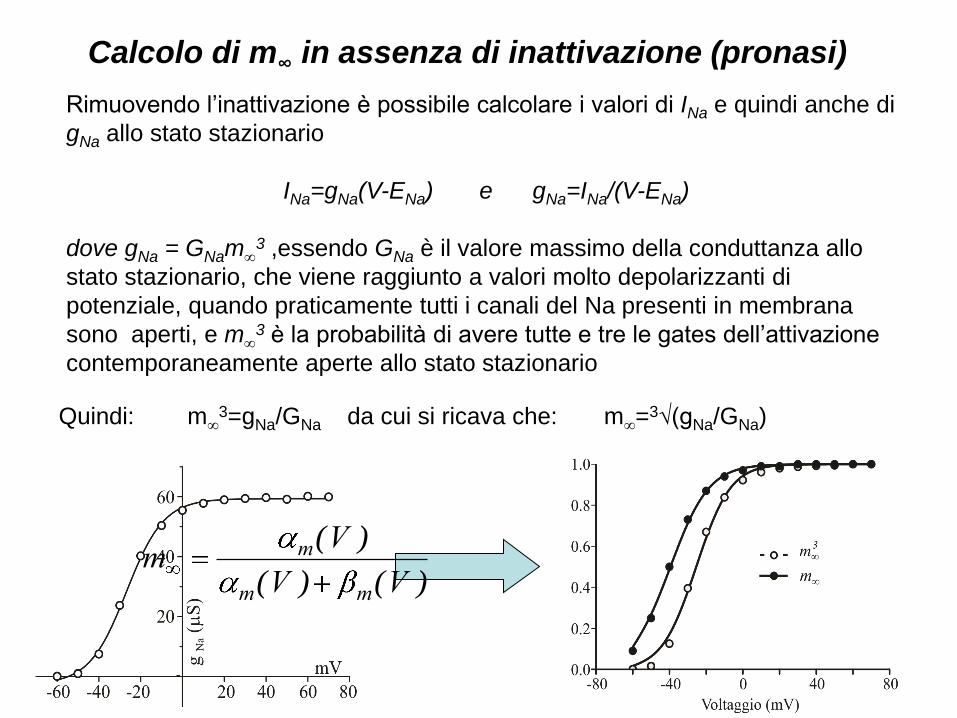
Calcolo di m∞ in assenza di inattivazione (pronasi)
Rimuovendo l’inattivazione è possibile calcolare i valori di INa e quindi anche di
gNa allo stato stazionario
INa=gNa(V-ENa) e gNa=INa/(V-ENa)
dove gNa = GNam∞3 ,essendo GNa è il valore massimo della conduttanza allo
stato stazionario, che viene raggiunto a valori molto depolarizzanti di
potenziale, quando praticamente tutti i canali del Na presenti in membrana
sono aperti, e m∞3 è la probabilità di avere tutte e tre le gates dell’attivazione
contemporaneamente aperte allo stato stazionario
Quindi: m∞3=gNa/GNa da cui si ricava che: m∞=3√(gNa/GNa)
)()(
)(
VV
Vm
mm
m

Calcolo di m in assenza di inattivazione (pronasi)
Si parte da un potenziale di holding piuttosto negativo, ad es. -70 mV, e si
depolarizza la membrana a vari potenziali per diversi ms, come nell’esempio
in figura;
Per la corrente di Na, m è calcolabile interpolando l’andamento temporale di gNa,
ottenuto ai vari potenziali in depolarizzazione, con una funzione esponenziale
elevata alla terza potenza del tipo
3/exp tBAACy /exp tBAACy
L’andamento della
conduttanza
rispecchia
l’andamento della
probabilità di
apertura del canale
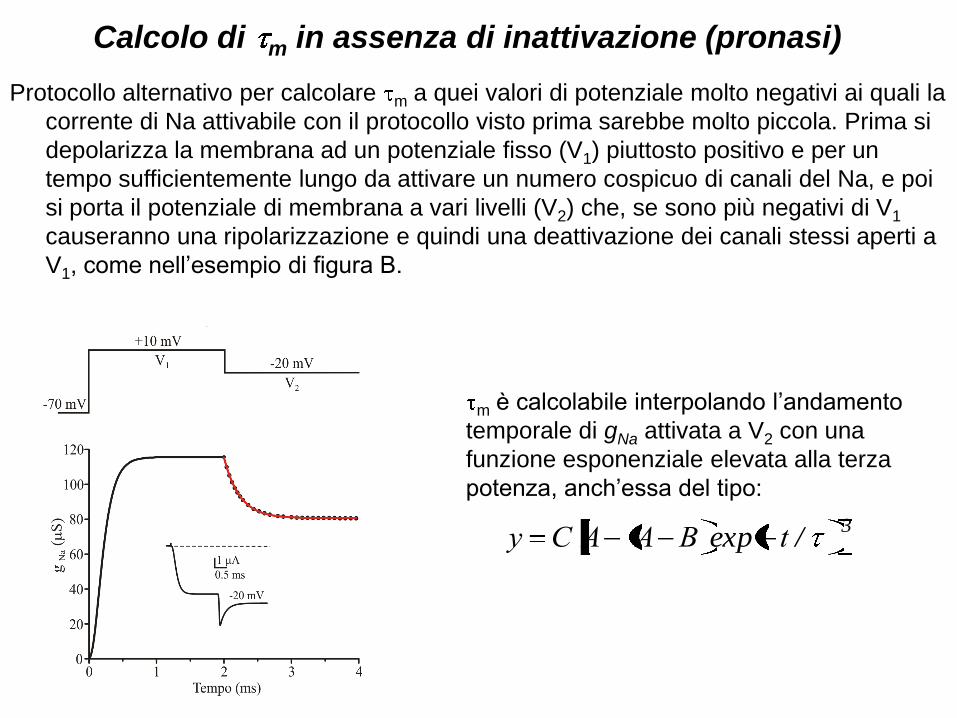
Calcolo di m in assenza di inattivazione (pronasi)
Protocollo alternativo per calcolare m a quei valori di potenziale molto negativi ai quali la
corrente di Na attivabile con il protocollo visto prima sarebbe molto piccola. Prima si
depolarizza la membrana ad un potenziale fisso (V1) piuttosto positivo e per un
tempo sufficientemente lungo da attivare un numero cospicuo di canali del Na, e poi
si porta il potenziale di membrana a vari livelli (V2) che, se sono più negativi di V1
causeranno una ripolarizzazione e quindi una deattivazione dei canali stessi aperti a
V1, come nell’esempio di figura B.
m è calcolabile interpolando l’andamento
temporale di gNa attivata a V2 con una
funzione esponenziale elevata alla terza
potenza, anch’essa del tipo:
3/exp tBAACy

Calcolo di m in assenza di inattivazione (pronasi)
Utilizzando entrambi i metodi è possibile ottenere sperimentalmente i valori di
m per un ampio range di potenziali
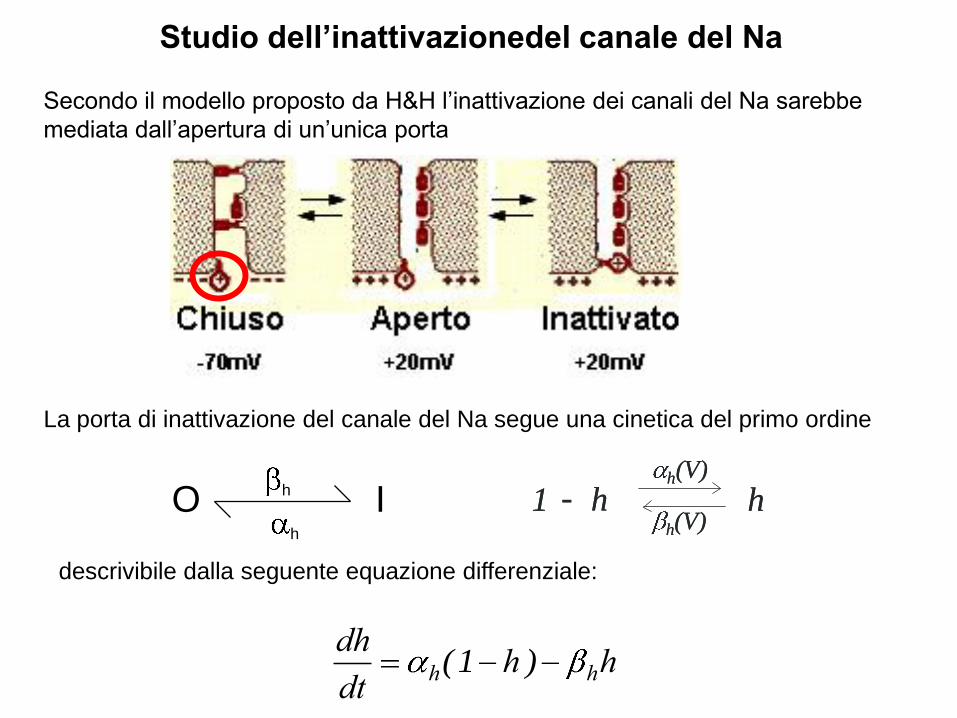
Studio dell’inattivazionedel canale del Na
La porta di inattivazione del canale del Na segue una cinetica del primo ordine
descrivibile dalla seguente equazione differenziale:
hh(V)
h1 -h(V)
hh(V)
h1 -h(V)
hhdt
dhhh )1(
Secondo il modello proposto da H&H l’inattivazione dei canali del Na sarebbe
mediata dall’apertura di un’unica porta
h
h
O I

Dove h∞ è il valore di h allo stato stazionario e ho è il valore di h a t=0 e in cui
la costante di tempo h è in relazione con le due costanti di velocità h e h nel
seguente modo:
Mentre, l’andamento temporale con cui lo stato stazionario verrà raggiunto
sarà descritto dalla seguente funzione esponenziale, che rappresenta una
soluzione dell’equazione differenziale
h
o
thhhth exp)()(
)()(
1
VV hh
h
La soluzione di tale equazione allo stato stazionario (t→∞) è:
)()(
)(
VV
Vh
hh
h

Studio dell’inattivazione allo stato stazionario
delle correnti di Na: calcolo di h∞
h
h
O I
)()(
)(
VV
Vh
hh
h

/exp tBAACy
Calcolo di h
Nel range di potenziali depolarizzanti a partire da -50 mV, dove il fenomeno
dell’inattivazione è chiaramente manifesto, come in figura, una misura della
cinetica dell’inattivazione la si può ottenere interpolando la componente
inattivante della corrente di Na con una funzione monoesponenziale decrescente,
dalla quale è calcolabile direttamente la costante di tempo dell’inattivazione ( h
secondo la terminologia di Hodgkin e Huxley).

Velocità del recupero dall’inattivazione (de-inattivazione) della corrente
di Na
La corrente viene evocata in successione a due potenziali test (V1 e V2) aventi la
stessa ampiezza ma separati da un ritorno al potenziale di holding di durata variabile.
V1 evoca una corrente sufficientemente ampia, utilizzata come controllo, la cui durata
è tale da permettere l’inattivazione pressoché completa dei canali. V2 partirà dopo che
la membrana verrà ripolarizzata al potenziale di holding per un numero crescente di
ms, permettendo ad una frazione di canali sempre più ampia di rimuovere
l’inattivazione. Il confronto tra le correnti evocate durante V2 e V1 permetterà di
stimare la frazione di canali che hanno recuperato dall’inattivazione.
Il recupero
dall’inattivazione sarà
approssimativamente
descrivibile da una
funzione
monoesponenziale del
tipo y=1-exp(-t/ h), dove
h è la costante di tempo
dell’inattivazione.
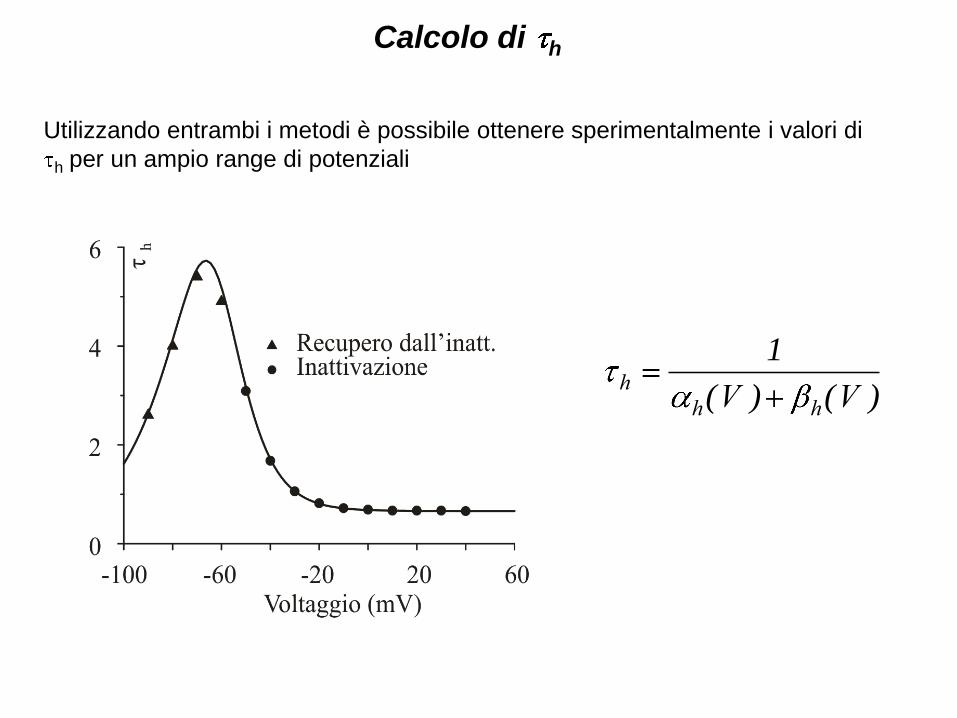
Utilizzando entrambi i metodi è possibile ottenere sperimentalmente i valori di
h per un ampio range di potenziali
Calcolo di h
)()(
1
VV hh
h
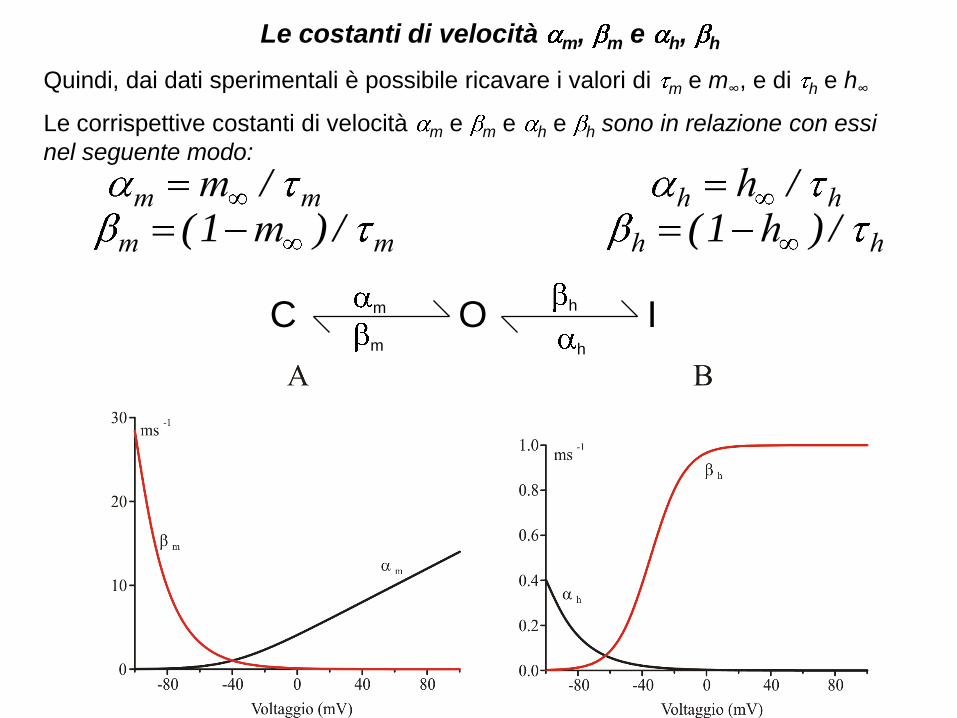
Le costanti di velocità m, m e h, h
Quindi, dai dati sperimentali è possibile ricavare i valori di m e m∞, e di h e h∞
Le corrispettive costanti di velocità m e m e h e h sono in relazione con essi
nel seguente modo:
mm m /
mm m /)1(hh h /
hh h /)1(
h
h
m
m
C O I
O C C C
3 1
2
2
2
3
3 I
h
h
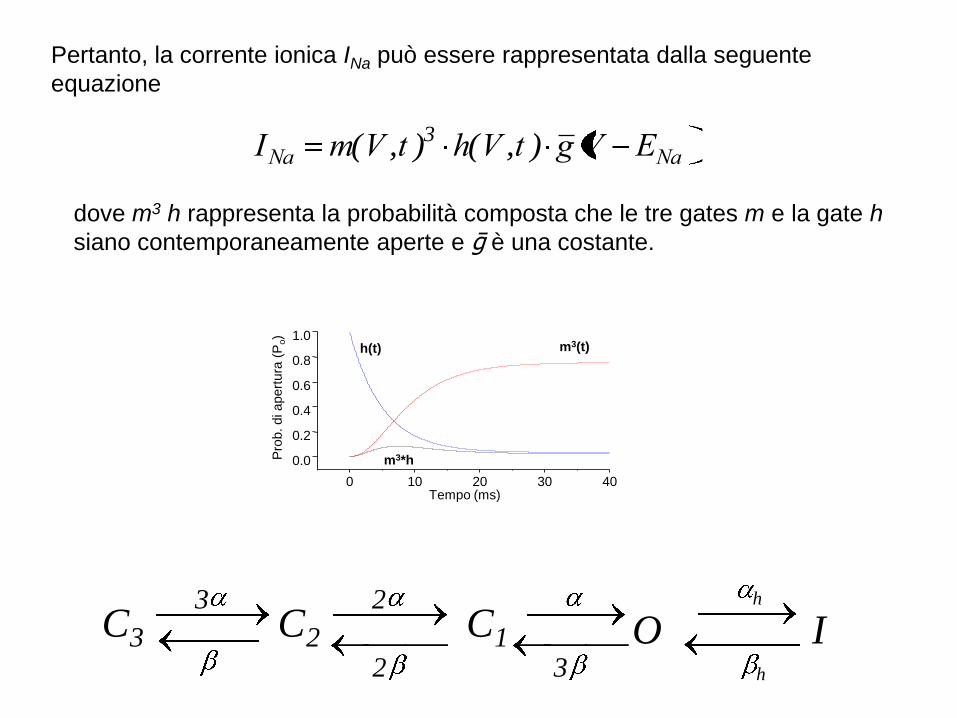
Pertanto, la corrente ionica INa può essere rappresentata dalla seguente
equazione
NaNa EVgtVhtVmI ),(),( 3
dove m3·h rappresenta la probabilità composta che le tre gates m e la gate h
siano contemporaneamente aperte e ḡ è una costante.
0 10 20 30 40
0.0
0.2
0.4
0.6
0.8
1.0
Pro
b. di apert
ura
(P
o)
Tempo (ms)
m3(t) h(t)
m3*h

THAT’S ALL FOLKS!
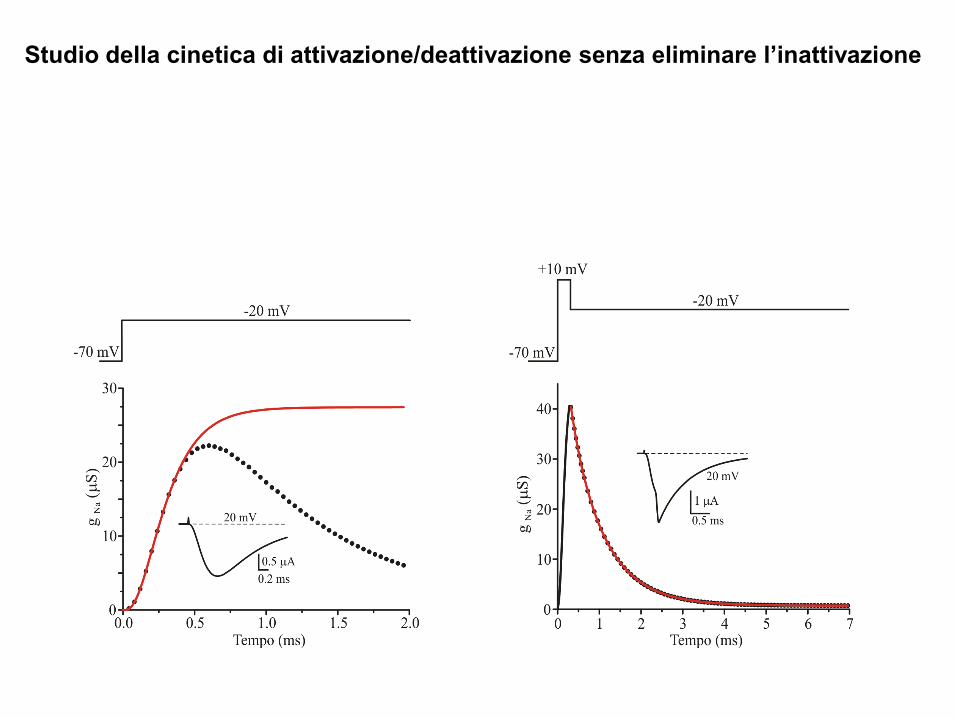
Studio della cinetica di attivazione/deattivazione senza eliminare l’inattivazione
![P ] ] h o } o } ] v ] ] ] o o o } o ] o } Y i & o } v ] ] - UniPa · 2019-02-04 · > ] } v } o } p ] w } ] ] } o } p ] ] ] v } o ] } o } p ] o ] v ] h o } o } ] v ] ] ] o o o } o](https://static.fdocumenti.com/doc/165x107/5f34e7b7a9690435fc2e2132/p-h-o-o-v-o-o-o-o-o-y-i-o-v-unipa-2019-02-04.jpg)






![µ Ì ] } v ] v } u ] o · txdqwlwj vlpphwulfd n vxffhvvl 2 : g l x j l s r á l l r ä w ; l s r h {h z h y h x h w h v h u h t x h w h v h u h t h v h u h t ® r ä w 5 4 l r ä](https://static.fdocumenti.com/doc/165x107/5f143fd7e52ca628f67db95b/-oe-v-v-u-o-txdqwlwj-vlpphwulfd-n-vxffhvvl-2-g-l-x-j-l-s-r-l-l.jpg)





![Cominardi - Musicoterapia · Dh^/ D DKZ/ h v [ ] À ] u µ ] o } u v v Ì } v ] U Dh^/ Kd Z W/ /E Z^ > ] u } o Ì ] } v } v } µ v ] u } v } ] }](https://static.fdocumenti.com/doc/165x107/5ff7e0218391042aab768891/cominardi-dh-d-dkz-h-v-u-o-u-v-v-oe-v-u-dh-kd-z-w-e.jpg)



![I MISTERI DELLE PIRAMIDI EGIZIE · 2021. 1. 14. · } ] ' ] u ^ À } v o ] u d À ] } ] & v ï w p ] v ï ] í ð = o o p } ~ ð d & h fkh oh whfqlfkh h l pdwhuldol lpslhjdwl shu](https://static.fdocumenti.com/doc/165x107/61325d69dfd10f4dd73a6708/i-misteri-delle-piramidi-2021-1-14-u-v-o-u-d-.jpg)
