2.2 Le cellule procariotiche non contengono organelli ...
42
2.2 Le cellule procariotiche non contengono organelli circondati da membrane. Le cellule procariotiche sono tipicamente più piccole di quelle eucariotiche. In esse, il DNA non è racchiuso all’interno di un nucleo, come invece è nelle cellule eucariotiche, ma è localizzato in una regione limitata della cellula detta nucleoide non delimitata da membrana. In questo senso si spiega anche il termine procariote che appunto significa “prima del nucleo”. Anche altri tipi di organelli delimitati da membrana sono assenti nelle cellule procariotiche. Come le cellule eucariotiche, le cellule procariotiche hanno una membrana plasmatica, che circonda la cellula. Questa membrana delimita il contenuto della cellula, originando un compartimento interno. Molte cellule procariotiche hanno una parete cellulare, struttura che racchiude l’intera cellula, membrana plasmatica compresa. Molto procarioti possiedono flagelli, lunghe fibre che si estendono dalla superficie cellulare, funzionali al movimento dell’organismo. La matrice interna alla cellula procariotica contiene ribosomi, complessi di RNA e proteine, che come abbiamo visto hanno una funzione fondamentale nella sintesi proteica e che nei procarioti sono più piccoli che negli eucarioti. 2.3. Dalla cellula procariotica a quella eucariotica: ipotesi endo-simbiontica L’Ipotesi endo-simbiontica, formulata dalla scienziata Lynn Margulis, è senza dubbio la più convincente tra le teorie addotte per spiegare la comparsa della cellula eucariotica. Essa prende le mosse da un progenitore procariotico che, una volta persa la sua parete cellulare, acquisì la capacità di inglobare materiale esterno introflettendo la membrana plasmatica. Tra le prede di questo progenitore potrebbe esserci stato un procariote eterotrofo dotato di una elevata capacità di ossidare molecole organiche per ottenere energia. Per qualche motivo, il progenitore eucariotico, anziché digerirlo, conservò il batterio come ospite: ciò gli offriva il vantaggio di gestire una fabbrica energetica con capacità ben superiori a quelle che gli erano consentite dalla sua dotazione originaria. Da questo simbionte derivò il mitocondrio della cellula eucariotica. A conferma di questo, è possibile osservare come nelle cellule eucariotiche attuali, i geni ancora presenti nei mitocondri mostrano una organizzazione simile a quella dei geni batterici. Un secondo evento di endosimbiosi ebbe un grandissimo impatto dal momento che, nella popolazione delle cellule eucariotiche già provviste di mitocondri, alcuni organismi fagocitarono un cianobatterio fotosintetico senza digerirlo. Questo nuovo endo-simbionte fotosintetico, una sorta di capostipite dei plastidi, rendeva la cellula completamente autonoma per la nutrizione. Come nel caso dei
Transcript of 2.2 Le cellule procariotiche non contengono organelli ...

2.2 Le cellule procariotiche non contengono organelli circondati da membrane.
Le cellule procariotiche sono tipicamente più piccole di quelle eucariotiche. In esse, il DNA non è
racchiuso all’interno di un nucleo, come invece è nelle cellule eucariotiche, ma è localizzato in una
regione limitata della cellula detta nucleoide non delimitata da membrana. In questo senso si
spiega anche il termine procariote che appunto significa “prima del nucleo”. Anche altri tipi di
organelli delimitati da membrana sono assenti nelle cellule procariotiche. Come le cellule
eucariotiche, le cellule procariotiche hanno una membrana plasmatica, che circonda la cellula.
Questa membrana delimita il contenuto della cellula, originando un compartimento interno. Molte
cellule procariotiche hanno una parete cellulare, struttura che racchiude l’intera cellula,
membrana plasmatica compresa. Molto procarioti possiedono flagelli, lunghe fibre che si
estendono dalla superficie cellulare, funzionali al movimento dell’organismo. La matrice interna
alla cellula procariotica contiene ribosomi, complessi di RNA e proteine, che come abbiamo visto
hanno una funzione fondamentale nella sintesi proteica e che nei procarioti sono più piccoli che
negli eucarioti.
2.3. Dalla cellula procariotica a quella eucariotica: ipotesi endo-simbiontica
L’Ipotesi endo-simbiontica, formulata dalla scienziata Lynn Margulis, è senza dubbio la più
convincente tra le teorie addotte per spiegare la comparsa della cellula eucariotica. Essa prende le
mosse da un progenitore procariotico che, una volta persa la sua parete cellulare, acquisì la
capacità di inglobare materiale esterno introflettendo la membrana plasmatica. Tra le prede di
questo progenitore potrebbe esserci stato un procariote eterotrofo dotato di una elevata capacità
di ossidare molecole organiche per ottenere energia. Per qualche motivo, il progenitore
eucariotico, anziché digerirlo, conservò il batterio come ospite: ciò gli offriva il vantaggio di gestire
una fabbrica energetica con capacità ben superiori a quelle che gli erano consentite dalla sua
dotazione originaria. Da questo simbionte derivò il mitocondrio della cellula eucariotica. A
conferma di questo, è possibile osservare come nelle cellule eucariotiche attuali, i geni ancora
presenti nei mitocondri mostrano una organizzazione simile a quella dei geni batterici. Un secondo
evento di endosimbiosi ebbe un grandissimo impatto dal momento che, nella popolazione delle
cellule eucariotiche già provviste di mitocondri, alcuni organismi fagocitarono un cianobatterio
fotosintetico senza digerirlo. Questo nuovo endo-simbionte fotosintetico, una sorta di capostipite
dei plastidi, rendeva la cellula completamente autonoma per la nutrizione. Come nel caso dei

mitocondri, anche i plastidi hanno un loro genoma, diverso da quello nucleare, e simile in
organizzazione e struttura a quello dei cianobatteri.
2.4 Cellula vegetale e cellula animale
Nei paragrafi precedenti sono state evidenziate le differenze esistenti tra cellula procariotica e
cellula eucariotica. Queste differenze sono maggiori di quanto non lo siano quelle esistenti,
all’interno degli eucarioti, tra cellule animali e vegetali e tra queste ed i funghi o i protisti. Come
vedremo, le cellule vegetali sono diverse da quelle animali perché possiedono: 1. parete cellulare e
plasmodesmi; 2. plastidi; 3. vacuoli; 4. almeno nelle piante vascolari, non hanno centrosomi
(organuli coinvolti nell’organizzazione del fuso durante la fase mitotica). Per il resto sono del tutto
uguali sia nelle componenti, che nella struttura e funzioni.
2.6 Differenze tra vegetali ed animali
La nutrizione eterotrofa per ingestione ha imposto agli animali la necessità del movimento per la
ricerca di cibo ed ha favorito lo sviluppo delle superficie interne del corpo in modo da favorire gli
scambi gassosi, l’assorbimento dei nutrienti, l’eliminazione delle scorie ed assicurare il massimo
risparmio di acqua. Poiché i nutrienti necessari alle piante (anidride carbonica, acqua, sali minerali,
luce) sono diluiti nell’ambiente, esse hanno privilegiato lo sviluppo di superficie esterne (radici,
foglie). Questa grande estensione del corpo impone loro immobilità. Negli animali l’accrescimento
è definito, limitato cioè alla fase giovanile della vita; inoltre, in essi l’organogenesi (cioè la
formazione degli organi) è completa al termine dello sviluppo embrionale e negli stadi successivi lo
sviluppo è limitato ad un aumento in dimensioni degli organi stessi. Le piante, invece, hanno un
accrescimento indefinito grazie alle cellule meristematiche che si mantengono ad uno stadio
giovanile e producono continuamente nuovi organi. Nelle piante, vi è un adattamento della
struttura tale da favorire la crescita verso la luce contro la forza di gravità ed, al contempo,
l’evoluzione di un sistema aperto di circolazione dell’acqua (l’acqua che circola in una pianta non è
mai la stessa diversamente dagli animali dove il sistema è chiuso).
2.7 La vita è caratterizzata da diversi livelli di organizzazione
L’organizzazione biologica si basa su una gerarchia di livelli strutturali ognuno dei quali poggia su
un gradino sottostante. Il livello chimico è quello più semplice e comprende atomi e molecole.

Molte delle molecole di valore biologico sono localizzate in minuscole strutture chiamate organuli
che sono a loro volta i componenti della cellula. Le cellule sono le sub-unità degli organismi. Alcuni
organismi come le amebe corrispondono a singole cellule ma altri sono aggregati multicellulari
cioè sono costituiti da più cellule, coordinate e specializzate. Le attività che un’ameba compie
come singola cellula, una pianta le svolge dividendo il lavoro tra varie linee di cellule specializzate.
Infatti, durante l’evoluzione degli organismi pluricellulari, le cellule si sono associate per formare i
tessuti; ad esempio, i vegetali hanno un tessuto epidermico che serve come copertura di
protezione ed i tessuti vascolari per il movimento dei materiali attraverso il corpo della pianta.
Diversamente dall’ameba nessuna delle cellule di una pianta potrebbe vivere a lungo isolata dalle
altre. I tessuti, a loro volta, si associano in strutture funzionali chiamate organi, come le radici, il
fusto e le foglie; gli organi sono riuniti a formare quella che è l’unità base del mondo organico, cioè
l’organismo. E’ opportuno sottolineare è che ad ogni nuovo passo si manifestano nuove proprietà
che non erano presenti ai livelli più semplici. Tali proprietà, definite come proprietà emergenti,
non sono riscontrabili a livello dei singoli elementi e non sono direttamente derivabili dalle
proprietà di questi ultimi. Tale proprietà dipendono dall’insieme delle interazioni tra i singoli
elementi, cioè dall’organizzazione del sistema. Una molecola, per esempio, una proteina presenta
caratteristiche che non sono riscontrabili in ogni singolo atomo che la compone (né rappresentano
semplicemente la somma delle caratteristiche degli atomi) e non c’è dubbio che una cellula sia
molto di più di un semplice aggregato di molecole.
2.8 Le dimensioni cellulari hanno un limite
La relazione geometrica spiega perché la maggior parte delle cellule sono microscopiche. Nella
diapositiva le cellule sono rappresentate come cubi. Un valore alto del rapporto S/V
(Superficie/Volume) facilita gli scambi di materiali fra la cellula e l’ambiente esterno. La superficie
è rappresentata dalla membrana cellulare, una vera e propria interfaccia tra la cellula e l’ambiente
esterno; si tratta di una specie di barriera selettiva in grado di regolare il passaggio di ossigeno e
nutrienti e l’eliminazione delle scorie. Per fare fronte in maniera adeguata alle esigenze
metaboliche della cellula, la membrana deve essere estesa in maniera da massimizzare la capacità
di assorbire molecole e smaltire prodotti di rifiuto. Per i corpi tridimensionali vale però una regola:
la loro superficie non aumenta in modo proporzionale all’aumento del loro volume, bensì di meno.
Ciò significa che quanto più una cellula è grande tanto meno è estesa, in proporzione, la sua
superficie e quindi la sua membrana plasmatica. Meglio tante ma piccole: se l’organismo è

composto di molte cellule è possibile ripartire i compiti tra tante cellule specializzate. Pur avendo
una struttura comune le cellule possono avere differenti specializzazioni. Il fatto di essere
numerose favorisce la specializzazione. Anche la forma delle cellule può essere funzionale alla
regolazione del S/V: alcune cellule vegetali di grandi dimensioni, ad esempio, sono lunghe e sottili.
2.9 Metodiche per lo studio delle cellule, dei tessuti e degli organi
Prima di fare una piccola panoramica sulle tipologie di microscopi utilizzati per lo studio della
cellula e dei tessuti, vediamo il significato di alcuni termini:
Morfologia = studio di forme e strutture esterne di piante ed animali.
Citologia o biologia cellulare = scienza che studia la cellula dal punto di vista morfologico
(studio strutturale, ad esempio, del nucleo, dei ribosomi o dei vari organuli), e funzionale
(studio dei processi fondamentali come il ciclo cellulare o la meiosi).
Anatomia = studia la forma e la struttura degli organismi a livello di tessuti. Deve il suo
nome al metodo principale d'indagine, la dissezione, rimasta di fondamentale importanza
anche in epoca moderna, per quanto integrata da altri moderni e perfezionati metodi di
indagine.
Fisiologia = scienza integrata che utilizza principi chimico-fisici per spiegare il
funzionamento degli esseri viventi, siano essi vegetali o animali, mono o pluricellulari.
Uno dei più importanti strumenti utilizzabili per lo studio della cellula è senza dubbio il
microscopio. Un microscopio da lui inventato consentì allo scienziato inglese Robert Hooke di
descrivere la prima cellula nel 1665. Hooke esaminò un pezzo di sughero e disegnò e descrisse ciò
che aveva visto scegliendo il termine “cellula” perché la struttura gli ricordava le piccole stanze in
cui vivevano i monaci. In realtà ciò che Hooke vide non erano cellule vive bensì le pareti di cellule
morte. Il primo microscopio ottico moderno fu opera, pochi anni dopo, del naturalista olandese
Antoine van Leeuwenboek che, utilizzando lenti capaci di ingrandire le immagini più di 200 volte,
riuscì a vedere batteri, protisti, cellule del sangue e spermatiche. Ma è solo alla fine del XIX° secolo
che i primi microscopi vennero divulgati e permisero di studiare le cellule in maniera sistematica.
Esistono diverse tipologie di microscopi con differente capacita di ingrandire gli oggetti osservati
(l’ingrandimento corrisponde al rapporto tra le dimensione dell’immagine vista al microscopio e le
dimensioni reali dell’oggetto) e di risoluzione (cioè la capacità di distinguere anche i più piccoli

dettagli di un’immagine) utilizzabili per scopi differenti. Lo stereomicroscopio consente
ingrandimenti fino a 100x (cento volte le sue dimensioni reali) e viene usato spesso per studiare le
superfici di un campione (cioè la sua morfologia) o per eseguire attività come classazione,
dissezione, determinazione di campioni vegetali. I microscopi ottici possono ingrandire il
campione fino ad un massimo di 1000 volte (1000x); ad ingrandimenti superiori l’immagine risulta
progressivamente sfuocata. Per questo motivo, un microscopio ottico non è in grado di risolvere
particolari inferiori a 0,2 micrometri (μm = un millesimo di millimetro), misura che corrisponde a
quella di un piccolo batterio. Poiché l’interno delle cellule è trasparente, è difficile distinguere
specifiche strutture cellulari al microscopio ottico se non attraverso l’uso di sostanze coloranti che
aumentano il contrasto di immagine e spesso possono essere utilizzate per evidenziare particolari
strutture e composti specifici (ad esempio, la presenza di lignina o suberina sulla parete cellulare,
la presenza di amido nella matrice citoplasmatica, ecc.). Tuttavia, ad eccezione degli organuli più
grandi (come il nucleo ed il mitocondrio), la maggior parte delle strutture subcellulari sono troppo
piccole per essere osservate al microscopio ottico. La biologia cellulare ha fatto un rapido passo
avanti negli anni ’50 del secolo scorso con la messa a punto del microscopio elettronico a
trasmissione (TEM) dove il campione viene attraversato o colpito in superficie da un fascio di
elettroni anziché da luce visibile come avviene invece nel microscopio ottico. Il potere risolutivo di
un microscopio è inversamente proporzionale alla lunghezza d’onda delle radiazioni utilizzate ed i
fasci di elettroni hanno lunghezza d’onda assai inferiori a quelli della luce visibile. I moderni
microscopi elettronici possiedono una risoluzione di circa 0,1 nanometri (nm = 1 milionesimo di
millimetro), ma in realtà il limite pratico per l’osservazione delle strutture biologiche è di soli 2 nm,
che tuttavia è circa mille volte quello del microscopio ottico. Si usa il termine ultrastruttura per
indicare l’anatomia cellulare come appare al microscopio elettronico. Il microscopio elettronico a
scansione (SEM) viene utilizzato per studi dettagliati della superficie di un campione in quanto
permette di ottenere un’immagine tridimensionale che mostra la topografia della superficie
dell’oggetto studiato. I microscopi elettronici rivelano la presenza di numerosi organuli non visibili
al microscopio ottico. Il microscopio ottico offre molti vantaggi soprattutto per l’osservazione delle
cellule in vivo.
L’anatomia si studia su sezioni orientate perpendicolarmente o parallelamente rispetto all’asse
maggiore del fusto, della radice o della foglia (sezioni perpendicolari = trasversali e sezioni
parallele = longitudinali). Le sezioni longitudinali a loro volta possono essere:

radiali: superfici longitudinali che decorrono lungo un raggio dell’asse.
tangenziali: superfici longitudinali poste ad angolo retto rispetto all’asse del raggio.
2.10 La cellula vegetale
Una cellula vegetale è costituita dal protoplasto e dalla parete cellulare. Il protoplasto è tutto il
contenuto del lume cellulare (cioè l’insieme di citoplasma, nucleo e vacuoli) e vi si comprende
anche la membrana citoplasmatica. Il citoplasma è quella porzione del protoplasto contenuta
all’interno della membrana plasmatica che include organelli circondati da membrana (ad esempio
cloroplasti e mitocondri), sistemi di membrane (quali il reticolo endoplasmatico ed l’apparato di
Golgi) e corpi sprovvisti di membrana (ad esempio ribosomi, filamenti di actina e microtubuli). Il
resto del citoplasma, cioè la matrice in cui sono sospesi il nucleo, i vari organelli ed i sistemi di
membrana, si chiama citosol. Si tratta di una soluzione colloidale costituita da acqua, ioni, lipidi,
proteine enzimatiche e strutturali, zuccheri, nucleotidi, ormoni.
2.10.1 Parete cellulare
Quando Robert Hooke, osservando al microscopio una sezione di sughero, descrisse una struttura
regolare e propose per essa il nome di cellula, in realtà stava osservando quello che in un tessuto
morto rimane della cellula, ovvero la sua parete. La parete è il compartimento esterno alla
membrana plasmatica presente in tutte le cellule vegetali (ma anche nei funghi ed in molti
batteri); la presenza di questa parete, come ho detto precedentemente, costituisce uno degli
elementi distintivi tra cellula vegetale e cellula animale. Si tratta di una struttura molto complessa,
con accrescimento centripeto (cioè gli strati che compongono la parete vengono deposti
dall’esterno verso il plasmalemma), che presenta molti “canali” che mettono in comunicazione le
cellule fra di loro e la cellula con l’ambiente esterno. La composizione e l’organizzazione delle
componenti di parete rispondono alle necessità di un compromesso tra rigidità e plasticità. La
rigidità permette di controllare l’espansione del protoplasto ed impedire la rottura della
membrana plasmatica quando il protoplasto aumenta di volume, in seguito all’assorbimento di
acqua da parte del vacuolo. Inoltre, è importante nell’imporre le dimensioni e la forma alle cellule,
nel modulare la consistenza del tessuto ed, in ultimo, nel conferire la forma definitiva agli organi
della pianta. Al contempo, una certa plasticità è necessaria per consentire la distensione della
cellula e, quindi, il suo accrescimento. La parete ha però anche altre funzioni: protegge dai fattori
di stress sia di natura abiotica che biotica provenienti dall’ esterno (aridità, gelo, patogeni, ecc.),

contribuisce al trasporto dell’acqua (come vedremo il “continuum” di pareti cellulari va a costituire
la cosiddetta via apoplastica), produce molecole segnale specialmente in relazione ad attacchi di
agenti patogeni, accumula sostanze di riserva come le emicellulose (polisaccaridi scarsamente
solubili associati alla cellulosa). Nella maggior parte dei tessuti, essa svolge la sua funzione in
cellule vive; in alcune cellule, però, la parete cellulare è il solo compartimento che rimane dopo la
morte del citoplasma. Il sughero, ad esempio, è un tessuto tegumentale secondario, privo di spazi
intercellulari, formato da cellule morte, la cui parete è ispessita e suberificata (e proprio queste
modificazioni di parete determinano la morte delle cellule) e il lume cellulare è ripieno d'aria.
Grazie a questa struttura il sughero ha proprietà di isolamento proteggendo i tessuti sottostanti
dagli scambi termici e dagli scambi di sostanze chimiche liquide o gassose. Lo xilema è costituito da
cellule con parete ispessita che subiscono un processo di lignificazione delle pareti secondarie, così
da risultare morte a maturità. L'assenza di citoplasma e organelli al loro interno è funzionale in
quanto riduce al minimo la resistenza offerta dal tessuto alla risalita contro gravità della linfa
grezza.
La parete riveste un ruolo assai importante nelle attività economiche umane. Da essa si ricavano
carta, fibre tessili (cotone, canapa, lino ed altre), polimeri industriali (pellicole, colle, addensanti
alimentari, plastiche biodegradabili), legno da industria e legna da ardere, ed indirettamente
petrolio e carbone. Inoltre, essendo la più grande riserva di carbonio organico presente in natura,
ha un ruolo ecologico determinante nel ciclo di questo elemento.
Biogenesi della parete. Una prima fase della sua formazione è connessa alla divisione cellulare: la
divisione della cellula in due cellule figlie si realizza mediante la formazione di un setto di
separazione di cui la porzione centrale costituisce la lamella mediana. Si tratta di un processo
relativamente semplice che consente di raddoppiare il numero di cellule ad ogni ciclo di divisone
cellulare; è un po’ come se si volesse dividere una stanza in due stanze costruendo una parete
divisoria. Come si forma la lamella mediana? Ne riparleremo successivamente a proposito della
mitosi, ma già da ora conviene sottolineare come il primo atto della citodieresi (cioè della divisione
del citoplasma) è rappresentato dalla comparsa del fragmoplasto che è l’apparato con cui prende
avvio la formazione del setto di separazione e quindi della parete cellulare. Dopo che i cromosomi
si sono allontanati gli uni dagli altri, nel piano equatoriale del fuso mitotico che si estende tra i due
nuclei figli, si forma il fragmoplasto (al microscopio ottico appare come una zona citoplasmatica
più densa solcata da tante fibrille) dove si vanno addensando, trasportati dalle vescicole del Golgi,

materiali quali glicoproteine, pectine, emicellulose che vanno a costruire il setto di separazione. La
formazione di questo setto di separazione non avviene contemporaneamente su tutto il piano di
divisione della cellula, ma è limitata inizialmente al centro e poi gradualmente si estende verso la
periferia fino a raggiungere le preesistenti pareti laterali della cellula. Mano a mano che il setto si
forma, il fragmoplasto scompare. In una seconda fase successiva alla costruzione di questo setto di
separazione, ogni cellula “figlia” costruisce per proprio conto, a ridosso della lamella mediana, una
sottile parete pectocellulosica alla quale viene dato il nome di parete primaria. Successivamente,
quando la cellula differenziandosi (cioè specializzandosi) si accresce, la parete effettua una intensa
crescita (accrescimento per distensione) durante la quale aumenta la propria superficie
mantenendo generalmente costante il suo spessore. Raggiunte le dimensioni finali, alcune cellule,
in relazione alla loro posizione ed alla funzione che svolgono, provvedono ad effettuare un
adeguato rinforzamento della parete incrementandone lo spessore: si forma così un robusto
strato di ispessimento che si appone all’interno della parete primaria e che costituisce la parete
secondaria. Nelle cellule giovanili la parete è costituita sempre dalla lamella mediana e da un’esile
parete primaria; in quelle adulte, come detto, spesso è presente una parete secondaria
pluristratificata.
Lamella mediana. La lamella mediana è la porzione più esterna della parete, e quindi della cellula.
E’ un sottile strato dello spessore non maggiore di un decimo di μm interposto tra le due pareti
primarie di due cellule attigue. La principale componente della lamella mediana sono le pectine:
macromolecole che derivano dalla polimerizzazione dell’acido galatturonico cui possono essere
attaccati anche altri zuccheri come l’arabinosio. Si tratta di composti fortemente idrofili in grado di
formare colloidi gelatinosi con azione cementante che consente loro di saldare insieme le due
cellule figlie; ciò conferisce, fra l’altro, croccantezza a frutta o verdura. Con la maturazione della
cellula, le pectine vengono idrolizzati da enzimi quali la pectasi e la pectinasi producendo gli spazi
intercellulari. La maturazione e, successiva marcescenza di un frutto, è accompagnata proprio da
questi processi di idrolisi. La pectina trova impiego nell'industria alimentare come gelificante,
soprattutto nella realizzazione di marmellate e confetture, assumendo la denominazione di E440.
A livello di lamella mediana sono presenti anche proteine sia enzimatiche che strutturali.
Parete primaria. La parete primaria è lo strato che si trova a ridosso della lamella mediana: di
solito è più grosso della lamella mediana ed ha uno spessore che va da un decimo di μm ad 1 μm.
La parete primaria è formata da una matrice di acqua (60%), sostanze pectiche (10-35%) ed

emicellulose (25-50%) prodotte dal Golgi, in cui è dispersa una componente fibrillare di cellulosa
(9-25%) sintetizzata da complessi enzimatici localizzati sulla superficie esterna della membrana
plasmatica.
Ma vediamo ora nel dettaglio quali sono le componenti che concorrono a formare la parete
primaria partendo dalla più importante e cioè dalla cellulosa. La cellulosa è un polisaccaride a
catena lineare costituito da unità di ß-glucosio legate tra loro da legami 1-4. Per realizzare questo
legame ß-1,4 ciascuna unità di ß-glucosio è ruotata di 180° rispetto all’unità precedente. Ne
consegue che l’unità strutturale ripetentesi nella catena cellulosica è il disaccaride cellobiosio. La
presenza del legame ß-1,4 e l'assenza, nei mammiferi, di enzimi specifici per la sua degradazione,
rendono non assimilabile il glucosio proveniente dalla cellulosa. Solamente i ruminanti ed altri tipi
di mammiferi, sono in grado di utilizzare il glucosio della cellulosa, grazie alla presenza, nel loro
sistema digerente, di batteri che rompono tale legame. Anche molti microrganismi del suolo
possiedono enzimi cellulosolitici che consentono di degradare la cellulosa e riutilizzare il carbonio
in essa contenuto. Le catene di cellulosa, che come ho già evidenziato sono prodotte da enzimi
localizzati sulla membrana plasmatica, sono riunite in microfibre del diametro di circa 10-25
nanometri. Le microfibre a loro volta si aggregano a formare le fibre, filamenti di 0,5 μm di
diametro ad arrangiamento cilindrico o piatto il che conferisce una resistenza alla trazione pari o
maggiore a quella di una fune di acciaio di uguale spessore. Le fibrille sono disperse (cioè sono
orientate secondo direzioni diverse) nella matrice tanto è vero che, nel caso della parete primaria,
si parla di tessitura dispersa.
Emicellulose: si tratta di un gruppo di polisaccaridi ramificati fortemente legati alla componente
fibrillare della parete grazie alla loro capacità di formare legami idrogeno. Grazie a questi legami,
le emicellulose stabiliscono la connessione fra componente fibrillare e matrice e limitano
l’estensibilità della parete, regolando le dimensioni delle cellule.
Proteine. Le pareti possono contenere anche glicoproteine (sono proteine strutturali che
conferiscono rigidità e svolgono un ruolo importante nella comunicazione cellula-cellula) ed
enzimi. Tra proteine di parete più conosciute vi sono le estensine, cosiddette perché si riteneva
fossero coinvolte nella distensione della parete cellulare. Secondo alcuni studiosi, tuttavia, il
deposito di estensina conferirebbe addirittura maggiore rigidità alla parete, rendendola meno
estensibile. Le espansine sono invece coinvolte nel “rilassamento” della componente fibrillare
della parte, un requisito, questo, fondamentale per il suo accrescimento.

Parete secondaria. Per la maggior parte delle cellule la parete primaria è la parete definitiva. Le
cellule che devono svolgere funzioni di supporto e meccaniche, nonché quelle di trasporto dei vasi
xilematici, dovranno dotarsi di parete secondaria. La parete secondaria si appone a ridosso ed
internamente alla parete primaria una volta che quest’ultima ha terminato la crescita in spessore.
La parete secondaria ha uno spessore di 3-5 μm ed è formata da una matrice (prevalentemente
emicellulose mentre le pectine possono mancare) in quantità più scarsa rispetto alla parete
primaria e da un sistema fibrillare, costituito dalla cellulosa, che può raggiungere valori tra l’80 ed
il 95%. Per questo motivo, la parete secondaria è rigida e non facilmente estensibile. Le proteine
strutturali e gli enzimi, che sono relativamente abbondanti nella parete primaria, sembrano
mancare in quella secondaria. In una parete secondaria, solitamente, si possono distinguere tre
strati (soprattutto nei tessuti meccanici ed in quelli vascolari) che differiscono gli uni dagli altri per
l’orientamento delle microfibrille di cellulosa. Mentre la parete primaria è caratterizzata da una
tessitura dispersa, nella parete secondaria le fibrille sono disposte parallelamente e le fibrille di
uno strato sono generalmente disposte perpendicolarmente allo strato precedente.
Punteggiature. La composizione e la struttura della parete cellulare potrebbero renderne difficile
l’attraversamento da parte dei vari composti necessari all’attività metabolica del protoplasto. Ho
detto difficile ma non impossibile. La cellulosa, infatti, è una molecola che, come è possibile
osservare versando un po’ di acqua colorata su un foglio di carta, può essere attraversata da acqua
ed altre sostanze. Tuttavia, per aumentare l’efficienza degli scambi che avvengono tra cellula e
cellula la parete cellulare si è dotata di perforazioni, chiamate punteggiature, nelle quali si
insinuano i plasmodesmi, che assicurano la continuità citoplasmatica tra cellule adiacenti. I
plasmodesmi sono strutture delimitate dal plasmalemma e formati da un tubulo di reticolo
endoplasmatico, detto desmotubulo, rimasto intrappolato durante la formazione del setto di
separazione della cellula madre. L’insieme del volume totale citoplasmatico di tutte le cellule
collegate tra loro dai plasmodesmi è detto simplasto. In parete primaria, le punteggiature sono
riunite in zone dette campi di punteggiature primarie mentre in parete secondaria, che non viene
deposta in corrispondenza dei gruppi di plasmodesmi, sono riunite in zone dette semplicemente
punteggiature o porocanali. Normalmente una punteggiatura di una cellula corrisponde ad
un’altra della cellula adiacente (punteggiatura appaiata).
Il trasporto avviene sia nel simpasto che nell’apoplasto. Come evidenziato sopra, il movimento
dell’acqua e dei soluti in essa disciolti da una cellula all’altra può avvenire nello spazio interno

attraverso i plasmodesmi. In quanto strettamente connesso dai plasmodesmi, l’insieme di tutti i
protoplasti del corpo della pianta e dei loro plasmodesmi costituisce un continuum detto
simplasto. Il movimento delle sostanze da cellula a cellula tramite i plasmodesmi è detto
simplastico e necessita l’attraversamento della membrana plasmatica o plasmalemma. Questa, al
pari di tutte le membrane lipoproteiche, come vedremo più avanti, è altamente impermeabile agli
ioni; inoltre, sovente il trasporto all’interno della cellula avviene contro il gradiente
concentrazione, e quindi è evidente che l’attraversamento della membrana plasmatica non può
avvenire secondo le modalità della semplice diffusione. Il movimento di ioni o grandi molecole
attraverso la membrana dipende innanzitutto dal metabolismo cellulare e dalla presenza nelle
membrane di specifici trasportatori che assicurano la necessaria selettività. Il movimento invece
nel continuum delle pareti e del sistema di conduzione xilematico, dove avviene il trasporto
apoplastico, prende il nome di apoplasto o spazio libero (gli spazi intercellulari, essendo pieni
d’aria, non fanno parte dello spazio libero). Per le proprietà idrofile dei suoi componenti (cellulosa,
emicellulose, pectine) la parete cellulare è permeabile all'acqua: l'acqua può agevolmente fluire
attraverso le pareti cellulari secondo un gradiente di diffusione senza che debba necessariamente
attraversare membrane ed entrare quindi nel protoplasto. Si tratta di un sistema efficiente di
trasporto che non prevede investimento energetico da parte della cellula. Non tutte le pareti,
però, sono idrofile e quindi non tutte sono in grado di assicurare un trasporto di tipo apoplastico:
come vedremo nel paragrafo successivo alcune modificazioni di parete (o meglio delle
incrostazioni che vengono depositate sulle pareti della cellula) rendono la parete cellulare
altamente impermeabile in modo tale da interrompere la diffusione dell’acqua e dei soluti.
Modificazioni della parete cellulare. Lo scambio di materiali ed informazioni tra cellula e cellula è
di fondamentale importanza nel funzionamento metabolico cellulare tanto che, come abbiamo
visto, le cellule si sono dotate di vie di comunicazione interne quali i plasmodesmi. Tuttavia, in
alcuni casi, la cellula può “ritenere” conveniente dotarsi di strutture isolanti in grado di limitare ed,
in alcuni casi, addirittura interrompere le relazioni con l’ambiente esterno. E’ un sistema di
protezione analogo a quanto messo in atto nella pratica edile quando si vuole isolare le pareti
della casa dal freddo (cappotto termico) o dall’umidità (mediante l’inserimento di una guaina che
blocca la penetrazione dell’acqua). In questo senso, le pareti di alcune tipologie di cellule possono
essere modificate per svolgere particolari funzioni. Queste modifiche possono consistere sia nella
deposizione di particolari molecole strutturali, generalmente non presenti nella maggior parte
delle cellule, che nell’alterata proporzione con cui i normali costituenti vengono deposti. Alcune di

queste modifiche determinano anche la morte della cellula: ne sono un esempio la suberificazione
del periderma e la lignificazione dei vasi xilematici. D’altra parte, la morte della cellula è in questi
casi un “male” necessario al raggiungimento della specifica funzionalità e cioè l’isolamento
dall’ambiente esterno (il sughero) e l’efficienza di trasporto (i vasi xilematici). La suberificazione è
una modificazione della parete cellulare che consiste nella deposizione di lamelle di suberina, un
polimero piuttosto complesso altamente idrofobico. In seguito al processo di suberificazione, le
pareti cellulari diventano barriere impermeabili con conseguente morte della cellula che, a quel
punto, si riempie d’aria. Un esempio della efficienza del sughero nel proteggere i tessuti
sottostanti dalla perdita d’acqua per evaporazione è fornito dai giovani tuberi di patata sui quali è
presente come tessuto di rivestimento una pellicola di sughero che consente ai tuberi di
mantenersi turgidi per lungo tempo. Basta asportare questa pellicola perché il tubero in poco
tempo si dissecchi. Il sughero è anche tessuto in grado di limitare le escursioni termiche e per
questo motivo viene anche utilizzato nelle pratiche di bioedilizia come isolante termico. La
lignificazione è un processo, invece, che consiste nell’ incrostazione della lignina (polimero molto
complesso costituito principalmente da composti fenolici) tra le maglie del reticolo cellulosico
della parete. La lignificazione ha il significato funzionale di conferire idrofobicità e compattezza alla
parete, rendendola impermeabile e resistente alla compressione. L’unione di lignina e cellulosa è
simile all’accoppiata cemento-ferro del cemento armato, dove il primo conferisce resistenza alla
compressione e il secondo alla tensione. In seguito alla lignificazione, inoltre, la parete cellulare
viene anche preservata dalla demolizione dei polisaccaridi da parte di enzimi idrolizzanti; è
appunto alla lignificazione che si deve la grande resistenza del legno all’attacco dei microrganismi;
solo alcuni funghi presenti nel tratto gastrointestinale degli erbivori (ruminanti e non) e di molti
uccelli possono idrolizzare la lignina. La lignificazione è tipica delle cellule dei tessuti meccanici e
dei vasi, elementi ai quali, per lo svolgimento delle loro funzioni, è richiesta, come già evidenziato,
una elevata rigidità della parete cellulare. Dopo la cellulosa, la lignina è il polimero più abbondante
di una pianta (fino al 20-30% in peso) e quindi del nostro pianeta. Essendo legata covalentemente
alla cellulosa e ad altri polimeri della pianta è molto difficile estrarla. Per questo motivo le cartiere
sono molto inquinanti: l’estrazione di pasta di cellulosa dal legno richiede, infatti, trattamenti
chimici molto forti. Le cellule dei tessuti tegumentali hanno, invece, pareti modificate per essere
impermeabili all’acqua. L’epidermide delle foglie ed dei germogli è ricoperta da uno strato di
spessore variabile denominato cuticola. Si tratta di una struttura non cellulare, stratificata,
costituita da cutina, cere ed altri polimeri. La cutina è una sostanza lipofila la cui molecola

presenta una struttura tridimensionale assimilabile nella forma ad un graticcio. Di per sé non
sarebbe del tutto impermeabile perché le maglie del graticcio sono lasse se non fosse che in
queste maglie alloggiano le cere, sostanze queste ultime altamente idrofobiche. Tali strutture
sono importanti da un punto di vista ecologico in quanto, soprattutto in ambienti aridi o
potenzialmente tali, limitano la perdita di acqua che può avvenire attraverso i tessuti. Nelle aree
caratterizzate, viceversa, da un regime pluviale, la presenza della cuticola facilità il deflusso
dell’acqua lungo la foglia evitando ristagni che in condizioni di temperature elevate potrebbero
facilitare la proliferazione di parassiti e di colonie algali.
2.10.2 Le membrane
Se la parete cellulare è una peculiarità dei procarioti e tra gli eucarioti delle piante, delle alghe e
dei funghi, le membrane biologiche si trovano in tutte le cellule. Si parla di membrane in quanto si
fa riferimento ad un sistema composto da membrane interne (quelle che circondano gli organuli
ed in alcuni casi, come nei plastidi e nei mitocondri, si estendono anche al loro interno) e da
membrana esterna (plasmalemma).
Funzioni delle membrane biologiche. Oltre a delimitare la cellula ed i compartimenti della cellula
dall’ambiente esterno le membrane regolano selettivamente il passaggio in entrata ed in uscita
delle sostanze. Sono una sorta di “posto di dogana” dove si effettuano controlli sia in entrata che
in uscita e si lasciano entrare o si bloccano le merci o se ne regola il passaggio mediante l’utilizzo di
specifici trasportatori. In questo modo la membrana contribuisce a mantenere un ambiente
interno compatibile con la vita. Ma è anche una superficie di comunicazione a livello della quale si
assiste ad uno scambio continuo di informazioni tra l’ambiente intra- ed extra-cellulare. Vista
l’abbondante presenza di proteine enzimatiche è anche una superficie dove si svolge un’intensa
attività catalitica. Inoltre, le membrane sono una parte essenziale dei sistemi di trasferimento e di
immagazzinamento dell’energia in quanto svolgono una funzione fondamentale nel processo di
chemiosmosi sia della catena respiratoria che della fase luminosa della fotosintesi.
Composizione e struttura delle membrane biologiche. Le membrane biologiche sono strutture
altamente conservate; infatti tutte le membrane di qualsiasi cellula di vivente hanno struttura e
composizione comparabili. Al microscopio elettronico è possibile evidenziarne la caratteristica
struttura “a sandwich” proteine-lipidi-proteine: i due strati esterni elettrondensi (si definisce
elettrondensa una zona della sezione che al microscopio elettronico si manifesta opaca o scura in
quanto si comporta da isolante e non si fa attraversare dal fascio di elettroni) sono composti da

proteine; un terzo strato, che appare invece come luminoso ed è collocato tra i due strati proteici,
è costituito da fosfolipidi. I fosfolipidi sono i principali responsabili delle proprietà fisiche delle
membrane biologiche. Ciò è dovuto al fatto che alcuni fosfolipidi hanno caratteristiche uniche,
come quella di formare strutture a doppio strato. Le molecole di questa classe di composti organici
presentano una testa polare idrosolubile e una coda apolare non idrosolubile. La testa polare è
costituita da glicerolo (un alcol a tre atomi di carbonio), un gruppo fosfato e la colina (un’ammina).
Al glicerolo sono attaccati due catene di acidi grassi che formano la porzione apolare e idrofobica
del fosfolipide. Le molecole di questo tipo che hanno regioni idrofiliche ed idrofobiche distinte
vengono chiamate anfipatiche. La disposizione a doppio strato permette alle teste idrofiliche dei
fosfolipidi di associarsi liberamente con l’ambiente acquoso, mentre le catene idrofobiche degli
acidi grassi sono relegate all’interno della struttura, lontane dalle molecole d’acqua. Un acido
grasso è insaturo (presenta uno o più doppi legami) mentre l’altro è saturo (non presenta alcun
doppio legame). La presenza del doppio legame influisce sulle caratteristiche chimico-fisiche dei
grassi: in linea generale, gli acidi grassi insaturi, come la maggior parte dei grassi vegetali, non si
solidificano se non a temperature molto basse; gli acidi grassi saturi, come quelli animali ma anche
l’olio di palma e di cocco ed il burro di cacao, si solidificano a temperature relativamente basse.
Nella componente lipidica sono immerse proteine integrali o transmembrana che possiedono
regioni idrofobiche (a contatto con il doppio strato fosfolipidico) e regioni idrofiliche esposte sui
due lati della membrana. Anche queste proteine, quindi, possono essere considerate come
molecole anfipatiche. Secondo il modello a mosaico fluido, il doppio strato di molecole
fosfolipidiche ha una consistenza fluida ed in esso sono immerse, come tante tessere di un
mosaico, le proteine. Questo modello a mosaico non è statico, poiché le proteine cambiano
continuamente posizione e si muovono come degli iceberg che galleggiano in un mare
rappresentato dal doppio strato fosfolipidico. Oltre alle proteine di membrana vi sono anche
proteine periferiche che non sono completamente immerse nello strato fosfolipidico, sono invece
delle appendici legate debolmente alla superficie della membrana, spesso ancorate a porzioni
esposte delle proteine integrali. La membrana è costituita anche da carboidrati, localizzati
esclusivamente sulla superficie esterna, che sembrano avere un ruolo importante nel
riconoscimento tra cellula e cellula tramite il quale una cellula è in grado di distinguere i diversi tipi
di cellule adiacenti. Nel concetto di membrana a mosaico fluido è insito, come detto sopra, il
concetto di fluidità. Quando la membrana solidifica (ad esempio a temperature basse) cambia la
sua permeabilità e le proteine enzimatiche presenti sulla membrana possono inattivarsi. Per

questo motivo, le cellule sono in grado di modificare la composizione lipidica delle proprie
membrane in risposta a variazioni di temperatura. In molte varietà di piante resistenti a
temperature basse (ad esempio, i cereali vernini) la percentuale di fosfolipidi insaturi aumenta in
autunno, un adeguamento, questo, che impedisce la solidificazione delle membrane cellulari
durante l’inverno. Ognuno di voi avrà sicuramente notato che lasciando il burro fuso a
temperatura ambiente, esso si solidifica. Gli oli vegetali, invece, a temperatura ambiente
rimangono liquidi. Infatti, come detto precedentemente, il burro è un grasso saturo che non ha
doppi legami a livello degli acidi grassi che lo compongono. Al contrario, un olio vegetale è
polinsaturo ed ha due o più doppi legami nelle sue catene di acidi grassi. I doppi legami fanno sì
che nelle molecole si vengano a formare ripiegamenti che impediscono alle catene idrocarburiche
di avvicinarsi fra loro ed interagire: non riuscendo a disporsi in modo ordinato, i legami tra le
molecole sono inferiori e quindi la temperatura di fusione è più bassa (cioè, ci vuole meno energia
per rompere i legami). In questo modo, i grassi insaturi abbassano la temperatura alla quale
solidificano sia l’olio sia i lipidi di membrana.
La peculiare struttura influenza la permeabilità della membrana cellulare. La struttura a mosaico
fluido consente alle membrane di funzionare come barriere selettivamente permeabili, cioè di
permettere il passaggio di alcune sostanze ma non di tutte. Più una molecola è piccola e solubile
nei lipidi (idrofobica) e più facilmente attraversa la membrana. I gas come l’ossigeno e l’anidride
carbonica sono piccole molecole apolari che attraversano rapidamente il doppio strato lipidico.
Sebbene siano polari, le molecole d’acqua attraversano, anche se lentamente, la membrana,
essendo sufficientemente piccole da poter passare attraverso interruzioni che si vengono a creare
temporaneamente tra le catene di acidi grassi. Molecole grosse e polari non diffondono attraverso
la membrana senza l’azione di specifiche proteine di membrana. Come vedremo queste proteine
ricadono in 3 categorie: canali che permettono il movimento passivo di ioni; pompe primarie, che
usano l’energia dell’ATP per trasportare attivamente ioni; trasportatori secondari che accoppiano
il movimento di uno ione con quello di un altro.
L’acqua e le cellule vegetali
Diffusione. Una immediata conseguenza del movimento continuo delle molecole nei gas e nelle
soluzioni è che le molecole si diffondono secondo un gradiente di concentrazione: da una zona di
maggior concentrazione ad una zona a minor concentrazione finché la concentrazione delle
molecole non diviene uniforme nelle due zone. A questo movimento di molecole si dà il nome di

diffusione. La diffusione è un processo spontaneo: l’energia è immagazzinata sotto forma di
energia potenziale e viene rilasciata quando gli ioni o le molecole si spostano da una regione ad
alta concentrazione verso una regione a bassa concentrazione. Molecole che vanno nella direzione
opposta (da bassa verso alta concentrazione), si muovono contro gradiente di concentrazione, ma
questo non può avvenire spontaneamente ma solo in seguito ad un input di energia. La legge di
Fick ci dice anche che la diffusione è rapida per distanze brevi (ad esempio, nel caso di trasporto
che avviene nell’ordine delle dimensioni cellulari), ma molto lenta per le lunghe distanze (ad
esempio, nel caso di risalita delle soluzioni nelle piante). Se per diffondere su una distanza di 50
μm (le dimensioni di una cellula) una molecola di glucosio in soluzione acquosa impiega 2,5
secondi, per essere trasportata ad un metro di distanza impiega 32 anni. Questo è dovuto al fatto
che man mano che la sostanza diffonde lontano dal punto di partenza la forza guida, cioè il
gradiente di concentrazione, diventa meno forte e quindi il movimento netto diventa più lento.
Come vedremo, il trasporto dell’acqua e dei soluti su lunghe distanze può essere spiegato con il
flusso di massa; il flusso di massa è un movimento globale delle molecole di un liquido (pensate ad
una cascata d’acqua oppure ad un gomma per l’irrigazione), che si spostano tutte insieme in una
direzione e serve in genere per spostare l’acqua ed i soluti da una parte all’altra di un sistema
pluricellulare (ad esempio xilema e floema). Se nella diffusione la forza guida è il gradiente di
concentrazione, nel flusso di massa è invece un gradiente di pressione idraulica fra regioni diverse
della soluzione.
L’osmosi è la diffusione di acqua attraverso una membrana selettivamente permeabile.
I fenomeni osmotici sono essenziali per tutte le cellule in quanto è attraverso di essi che si realizza
in gran parte l’assorbimento dell’acqua da parte di una cellula. Nella cellula vegetale questi
fenomeni svolgono ancor più un ruolo fondamentale poiché sono alla base di quel processo tipico
della cellula vegetale che è la crescita per distensione. L’osmosi è un tipo particolare di diffusione
che comporta il movimento netto di acqua attraverso una membrana selettivamente permeabile,
da una regione a concentrazione maggiore ad una a concentrazione minore. Le molecole d’acqua
passano liberamente in entrambe le direzioni ma come in tutti i tipi di diffusione il movimento
netto avviene dalla regione in cui le molecole d’acqua sono più concentrate a quella dove lo sono
meno. La maggior parte delle molecole di soluto (ad esempio, zucchero e sale) non può diffondere
liberamente attraverso le membrane cellulari selettivamente permeabili. L’osmosi è facilmente
osservabile utilizzando uno strumento chiamato osmometro costituito da un tubo ad U nel quale i

due bracci sono separati da una membrana semipermeabile. Da un lato viene posta una soluzione
acqua/soluto, mentre dall’altro lato viene messa acqua distillata. Si verrà quindi a creare una
differenza di potenziale dell’acqua, dato che i soluti abbassano tale potenziale (molte delle
molecole d’acqua si legano al soluto e pertanto non sono più libere di diffondere attraverso la
membrana), che genera un movimento netto di molecole d’acqua dalla parte in cui è contenuta
acqua distillata verso quella contenente acqua/soluto. Ne risulta che il livello del fluido cala dalla
parte dell’acqua pura e aumenta dalla parte dell’acqua/soluto. Inizialmente, l’acqua entra
velocemente, perché è elevata la differenza di potenziale dell’acqua tra un braccio e l’altro
dell’osmometro. Mano a mano che le molecole d’acqua si diffondono secondo il gradiente di
potenziale il livello di fluido continua ad aumentare dal lato contenente il soluto e,
proporzionalmente all’altezza raggiunta dalla colonna di acqua, si ha un aumento di pressione
all’interno del tubo. L’aumento della pressione innalza il potenziale chimico dell’acqua e così il
potenziale idrico nel tubo comincia a crescere verso lo zero. La pressione aumenta fino a che il
potenziale dell’acqua della soluzione all’interno del tubo diventa uguale a zero, cioè eguaglia il
potenziale dell’acqua distillata che si trova nell’altro tubo. Questo si verifica in coincidenza del
raggiungimento del livello massimo del liquido nel tubo. A questo punto il sistema ha raggiunto
uno stato di equilibrio per cui la differenza di potenziale fra i due lati della membrana è zero (la
variazione di energia libera è zero) e cessa la diffusione. Nel raggiungimento dell’equilibrio,
bisogna tener conto anche dell’effetto della diluizione. Come l’acqua si diffonde attraverso la
membrana, non soltanto questa provoca un aumento di pressione ma tende anche a diluire la
soluzione all’interno del tubo. Ciò aumenta il potenziale osmotico dell’acqua della soluzione (lo
rende meno negativo), così che la pressione richiesta per raggiungere l’equilibrio è minore di
quella che sarebbe stata necessaria. Si definisce pressione osmotica la pressione che deve essere
esercitata sulla soluzione più concentrata per impedire la diffusione dell’acqua dalla parte meno
concentrata a quella più concentrata. Una soluzione con un’alta concentrazione di soluto ha una
bassa concentrazione di acqua ed un’alta pressione osmotica (si parla di soluzione ipertonica); al
contrario, una soluzione con un basso quantitativo di soluto ha un’alta concentrazione di acqua ed
una pressione osmotica bassa (si parla di soluzione ipotonica). Due soluzioni di una sostanza che
hanno la stessa pressione osmotica sono dette isotoniche.
Le cellule sono osmometri artificiali. Nella cellula esistono tutte le condizioni per la realizzazione
di processi osmotici. Esistono membrane semipermeabili e soluzioni di vari soluti racchiusi in
compartimenti delimitati da membrane semipermeabili. Molti sono i compartimenti della cellula

che sono sede di fenomeni osmotici: i mitocondri, i plastidi, le cisterne del Golgi, il reticolo
endoplasmatico, ecc. tuttavia, nelle cellule vegetali, è il vacuolo, come vedremo, a rivestire un
ruolo di primaria importanza.
La tabella sottostante riassume il movimento dell’acqua verso l’interno o l’esterno di una soluzione
(o di una cellula) in rapporto alle concentrazioni relative di soluto
Concentrazione
del soluto nella
soluzione A
Concentrazione
del soluto nella
soluzione B
Tonicità Direzione del movimento
netto d’acqua
Maggiore Minore A ipertonica rispetto a B; B
ipotonica rispetto ad A
Da B verso A
Minore Maggiore B ipertonica rispetto ad A;
A ipotonica rispetto a B
Da A verso B
Uguale Uguale A e B sono isotoniche Nessun movimento netto
Se la soluzione è isotonica, ovvero ha la stessa concentrazione del citoplasma non avviene alcun
flusso di acqua attraverso la membrana. Se la soluzione circostante è più concentrata (ipertonica)
rispetto alla soluzione cellulare (ipotonica), una cellula posta in tali condizioni perde acqua e tende
a raggrinzirsi, cioè si ha un restringimento del circostante citoplasma ed un progressivo distacco
dalla parete cellulare. A tale fenomeno si dà il nome di plasmolisi. Se la soluzione cellulare è più
concentrata (ipertonica) di quella esterna (ipotonica), l’acqua attraverserà la membrana
dall’esterno verso l’interno della cellula riempiendo il vacuolo centrale e facendo distendere la
cellula. Mano a mano che l’acqua entra nella cellula, il volume della cellula aumenta e la parete
cellulare, che è elastica, viene progressivamente più tesa proprio come si tende la parete di un
palloncino che viene gonfiato. Il ruolo della parete cellulare è assimilabile a quello esercitato dalla
colonna d’acqua nel tubo di un osmometro; la progressiva tensione della parete cellulare,
generata dall’ingresso dell’acqua, si traduce in una pressione diretta verso l’interno che tende a
cacciare fuori l’acqua dalla cellula. Quanto più acqua penetra nella cellula tanto più forte diventa
questa contropressione fino al raggiungimento di un equilibrio stabile in cui la tendenza dell’acqua
ad entrare è esattamente bilanciata dalla contropressione della parete. A questo punto, la cellula
non introduce più acqua ed è detta turgida. In tali condizioni la pressione esercitata dalla parete
cellulare, detta pressione di turgore, ha raggiunto un valore uguale a quello della pressione

osmotica. Il turgore cellulare è un fattore di grande importanza per la vita dell’intera pianta. Le
cellule si accrescono solo quando sono turgide ed anche una piccola diminuzione del contenuto di
acqua al di sotto della piena saturazione determina un calo nella velocità di crescita. Gli apparati
stomatici basano il loro meccanismo di apertura/chiusura sulla variazione del turgore cellulare. Le
cellule se perdono troppa acqua si afflosciano e di conseguenza le foglie, i fiori, i frutti, piantine
intere perdono la loro consistenza. Inoltre, la pressione di turgore fornisce pressoché l’unico
sostegno alle piante non legnose.
La diffusione facilitata. Abbiamo visto come le molecole apolari ma anche alcune molecole polari
di piccole dimensioni (ad esempio l’acqua) possono attraversare la membrana per semplice
diffusione, cioè secondo un gradiente di concentrazione. Vi è tuttavia un diverso tipo di diffusione,
detto diffusione facilitata in cui una specifica proteina di trasporto rende la membrana
permeabile anche ad uno ione o ad una molecola polare. Attenzione: la diffusione facilitata
consente l’attraversamento della membrana a sostanze che, per lo più, non potrebbero
diffondersi liberamente ma il loro movimento netto avviene sempre da una regione a
concentrazione maggiore verso una regione a concentrazione minore e, quindi, secondo
gradiente. Ciò significa che la diffusione facilitata, al pari della diffusione semplice, non prevede il
dispendio di energia metabolica e per questo motivo si parla di trasporto passivo. Tra le proteine
trasportatrici possiamo distinguere: proteine canale e proteine carrier. Le proteine canale
formano dei “tunnel” che si lasciano attraversare da ioni e molecole d’acqua secondo i loro
gradienti. Per quanto riguarda gli ioni, come vedremo dopo, trattandosi di particelle cariche,
questi gradienti sono di tipo elettrochimico. Si tratta di canali che vengono aperti o chiusi
attraverso l’azione di “cancelletti chimici” e che possono essere attraversati anche da 100 milioni
di ioni al secondo. Le acquaporine sono proteine trans-membrana che favoriscono elevati flussi
d’acqua attraverso la membrana, consentendo, in questo modo, rapide contrazioni in risposta a
stimoli ambientali. Ad esempio, la chiusura a scatto tipo trappola della pianta carnivora Dionaea
muscipula o venere acchiappa mosche, il chiudersi e ritrarsi al contatto di Mimosa pudica. Il
trasporto dei soluti attraverso le proteine carrier è più lento di quello che avviene attraverso le
proteine canale. La proteina carrier lega una o più molecole di soluto su un lato della membrana,
dopodichè subisce un cambiamento della sua forma che determina lo spostamento del soluto
sull’altro lato della membrana. Una volta scaricata la molecola di soluto, la proteina riprende la
sua conformazione originaria.

Trasporto attivo. Sebbene alcune sostanze possano essere trasportate attraverso la membrana
cellulare per diffusione, una cellula spesso ha bisogno di trasportare i soluti anche contro
gradiente di concentrazione. La ragione è che molte sostanze sono richieste dalla cellula a
concentrazioni più alte rispetto a quelle esterne. Se l’energia per la diffusione è fornita dal
gradiente di concentrazione della sostanza che deve essere trasportata, nel caso del trasporto
attivo, cioè contro gradiente, è necessario un dispendio di energia metabolica (ATP) che alimenti il
processo. Tuttavia, il trasporto attivo può essere accoppiato all’ATP in maniera indiretta: l’ATP
viene usato per “pompare”, ad esempio, protoni contro il gradiente; in una seconda fase, il
gradiente di concentrazione fornisce esso stesso l’energia per il cotrasporto di un’altra sostanza.
Occorre qui evidenziare che il movimento dei soluti entro e fuori della cellula, o di un
compartimento cellulare, è guidato, nel caso di molecole cariche, da una combinazione di
gradiente di concentrazione e gradiente elettrico (per cui si parla di gradiente elettrochimico). In
tutte le cellule c’è un voltaggio tra i due lati della membrana (ad esempio, il citoplasma è carico
negativamente rispetto al fluido extracellulare in quanto vi è una distribuzione diseguale di anioni
e cationi sui lati opposti della membrana). Questo potenziale, che varia tra i -50 e -200 millivolt (il
segno meno indica che l’interno è negativo rispetto all’esterno), agisce come una batteria, una
fonte energetica che influenza il movimento di tutte le sostanze cariche attraverso la membrana. A
causa della differenza di carica tra interno ed esterno, il trasporto di membrana favorisce il
trasporto passivo di cationi verso l’interno ed anioni verso l’esterno. Quindi, a livello di membrana,
sono due le forze implicate: una chimica (il gradiente di concentrazione dello ione) ed una elettrica
(l’effetto del potenziale di membrana sul movimento dello ione).
Le piante hanno particolari pompe idrogeno che espellono ioni idrogeno. Alcune pompe, dette
pompe primarie od elettrogeniche, in grado di trasportare attivamente ioni, possono creare uno
squilibrio di cariche e quindi aumentare la componente elettrica del gradiente elettrochimico. La
pompa primaria delle membrane delle cellule vegetali è una pompa protonica (H+) localizzata sulla
membrana plasmatica e sul tonoplato (la membrana che delimita il vacuolo). L’energia liberata
dall’idrolisi dell’ATP viene utilizzata per trasportare gli ioni H+ dall’interno verso l’esterno della
cellula. In questo modo, si alza il pH del citoplasma (che si mantiene costante a circa 7,4) e, cosa
più importante, si genera un potenziale elettrico di membrana. Questo accade perché l’interno
della membrana si trova ad avere più cariche negative dell’esterno, dopo che gli ioni carichi
positivamente sono stati trasportati all’esterno. I gradienti di H+ possono essere immaginati come
serbatoi di energia potenziale usabile per scopi diversi. Mano a mano che la sua energia potenziale

viene usata, il gradiente protonico viene ricostituito dalle pompe. Si tratta quindi di una delle tante
forme di accoppiamento fra un processo esoergonico ed uno endoergonico che si trovano negli
esseri viventi. Se vogliamo ricorrere nuovamente all’analogia sistema metabolico – sistema
idraulico, il riflusso dei protoni è equivalente all’acqua che, scendendo da un bacino di montagna,
compie lavoro; il livello del bacino non si abbassa indefinitamente perché l’acqua viene
nuovamente pompata su man mano, con spesa di energia.
Il cotrasporto. Nel paragrafo precedente, si è visto che la dissipazione del gradiente protonico,
generato dall’attività della pompa, avviene grazie al riflusso passivo, cioè mediante il ritorno,
secondo il gradiente di concentrazione, degli H+ nel compartimento da cui erano stati espulsi.
Poiché il doppio strato fosfolipidico è impermeabile ai protoni, questo riflusso può avvenire
unicamente attraverso dei “varchi” controllati, ovvero attraverso dei canali. Se così non fosse i
protoni appena espulsi attraverso la membrana ritornerebbero indietro e quindi non si potrebbe
avere un gradiente. Un sistema di cotrasporto si configura, in un certo senso, come un trasporto di
tipo passivo che muove soluti (zuccheri, amminoacidi, anioni) attraverso una membrana contro
gradiente elettrochimico sfruttando l’energia fornita dal flusso di ritorno dei protoni. Ad esempio
gli ioni potassio, il fosfato ed il nitrato si muovono tutti verso l’interno della cellula con il flusso
protonico di ritorno, un movimento catalizzato da trasportatori specifici chiamati simporti. Gli ioni
sodio invece si muovono verso l’esterno della cellula e cioè in senso opposto alla direzione del
flusso protonico su un trasportatore specifico definito antiporto. Anche una macromolecola come
il saccarosio “accompagna” il rientro dei protoni seguendo il gradiente di concentrazione degli ioni
idrogeno mantenuto dalla pompa; evento questo di fondamentale importanza, come vedremo, nel
trasporto dei fotosintati.
Esocitosi ed endocitosi. Le molecole più grandi sono trasportate per endocitosi ed esocitosi. Si
tratta di processi di ingestione (endocitosi) o secrezione (esocitosi ) di sostanze mediante la
formazione di vescicole. L’ endocitosi può avvenire secondo due modalità: fagocitosi (“la cellula
mangia" ad esempio amebe e muffe) e pinocitosi (“la cellula beve" nel caso di sostanze liquide).
Procediamo il nostro viaggio nella cellula vegetale analizzando le altre componenti a partire dal
citoscheletro.
2.10.3 Citoscheletro. Il citoscheletro è una complessa rete di filamenti proteici che si estende nel
citosol della cellula eucariotica. Questi filamenti, oltre a fornire un supporto meccanico alla cellula,
aiutano gli organelli a mantenere una posizione, aiutano nel trasporto di materiali all’interno della

cellula, controllano i movimenti dei cromosomi durante la divisione del nucleo. E’ facile
immaginare il citoscheletro come qualcosa di statico, un specie di lattice di filamenti che tengono
le cellule in forme specifiche e i componenti cellulari in posti specifici. Tuttavia, le cellule si
dividono spesso, cambiano di dimensione e forma e gli organelli ed altri componenti interne di
tutte le cellule viventi sono per lo più in movimento costante; la qual cosa significa che il
citoscheletro non può essere rigido. Ci sono almeno tre tipi di filamenti:
1. microtubuli (i più grandi): tubi cavi di 18-25 nm di diametro composti da due subunità
proteiche, α e β tubulina. Sono abbastanza rigidi, pur potendo allungarsi aggiungendo
molecole di tubulina ad una estremità o accorciarsi perdendo unità di tubulina ad una
estremità della stessa parte. Oltre a svolgere un ruolo strutturale nella formazione del
citoscheletro, i microtubuli sono coinvolti nel movimento dei cromosomi durante la
divisione cellulare. Inoltre, fungono da “binari” per alcuni movimenti intracellulari. Infatti,
alcune proteine motrici (chinesine e dineine) sfruttano l’energia fornita dall’ATP per
muoversi lungo i filamenti citoscheletrici, trasportando come carico organuli e vescicole.
2. Filamenti di actina. I filamenti più piccoli del citoscheletro sono quelli di actina (4-7 nm)
costituiti da subunità di una proteina, l’actina, che si avvolgono a spirale. La proteina
motrice che si muove lungo il filamento è la miosina e l’energia deriva sempre dall’idrolisi
dell’ATP.
3. Filamenti intermedi. I filamenti intermedi sono chiamati così perché hanno una
dimensione media tra quella dei microtubuli ed i filamenti di actina. Sono fatti di proteine
avvolte in fasci a forma di corda ed hanno diametro tra 8 e 12 nm. Al contrario della altre
due categorie di filamenti del citoscheletro, non sono strutture dinamiche ma piuttosto
stabili e durature. La loro funzione è quella di fornire alla cellula resistenza meccanica
contro lo stiramento.
2.10.4 Organuli coinvolti nella sintesi proteica e nel trasporto. Sono numerosi gli organuli che
svolgono un ruolo nella sintesi delle proteine e in loro trasporto da un compartimento all’altro
della cellula. Il nucleo contiene ed esprime le informazioni genetiche. I ribosomi ed i componenti a
loro associati sintetizzano le proteine. Il reticolo endoplasmatico svolge un ruolo importante nella
sintesi e nell’assemblaggio delle proteine. L’ apparato di Golgi guida il movimento delle proteine
verso specifici compartimenti.

Nucleo. Il nucleo è l’organulo di maggiori dimensioni (fino 10 µm di diametro); è sede
dell’informazione genetica (genoma), provvede alla duplicazione e alla trascrizione del DNA. Si
chiama cromatina la forma “allentata”, meno strutturata, in cui gli acidi nucleici si trovano nella
cellula durante l’interfase del ciclo cellulare. La cromatina è costituita da DNA, RNA e proteine sia
acide che basiche (dette istoni). Durante il processo di divisione cellulare la cromatina diventa
progressivamente più condensata fino a diventare visibile al microscopio sotto forma di
cromosomi distinti. Ogni cromosoma, dopo il processo di duplicazione, presenta una forma ad X
poiché costituito da una coppia di unità identiche, chiamate cromatidi fratelli, associate tra loro
grazie in corrispondenza del loro centromero. I cromosomi sono solitamente presenti in coppie
nelle cellule somatiche degli organismi. I membri di ogni coppia, chiamati cromosomi omologhi,
sono simili per forma, dimensione, e posizione dei loro centromeri. Uno dei cromosomi della
coppia è di origine materna e l’altro di origine paterna: entrambi portano nella stessa posizione i
medesimi geni, cioè l’informazione per il controllo degli stessi caratteri genetici, sebbene non
necessariamente la medesima informazione. Ad esempio, un gene che codifica per il colore di un
fiore può presentarsi sui cromosomi omologhi sotto forma di due diverse varianti alleliche: una
variante che codifica per il colore giallo sul cromosoma di origine materna e per il colore rosso
quello sul cromosoma paterno o vice versa. Nel caso siano presenti entrambe le varianti alleliche si
parla di eterozigosi e l’individuo si dice eterozigote per quel carattere; se invece i due cromosomi
omologhi portano la medesima variante allelica si parla di omozigosi e l’individuo si dice
omozigote per quel carattere. Il numero di cromosomi varia notevolmente tra le specie differenti
e non è comunque correlato alla complessità degli organismi: ad esempio, alcune felci hanno
anche 1220 cromosomi mentre l’Arabidopsis thaliana, una piccola erbacea infestante ampiamente
usata per ricerche genetiche, ne ha solo 10. Le cellule riproduttive (cioè le cellule che partecipano
alla riproduzione sessuale come le cellule spermatiche e la cellula uovo), hanno solo la metà del
numero di cromosomi caratteristico delle cellule somatiche di ciascun organismo (cioè portano
solo un membro della coppia di omologhi); tale numero è indicato come corredo aploide (n), in
contrapposizione a quello diploide (2n) delle cellule somatiche (cioè quelle che formano il corpo
della pianta) dove sono portati entrambi i membri di ciascuna coppia di omologhi. Ad esempio, nel
Mais il numero dei cromosomi nelle cellule somatiche è 2n = 20 (10 coppie di cromosomi
omologhi: 10 cromosomi di origine materna e 10 di origine paterna); le cellule della linea
germinale hanno un corredo cromosomico n= 10 (per ciascuna coppia di omologhi è presente solo
un cromosoma di origine materna o paterna). Le cellule che hanno più di un corredo di cromosomi

sono dette poliploidi (3n, 4n, 5n, o più). La poliploidia è piuttosto frequente negli organismi
vegetali (quasi l’80% delle angiosperme è poliploide) ed è stato un fattore importante
nell’evoluzione delle piante. Nel nucleo si trovano sempre uno o più nucleoli, piccoli corpi
subsferici sede della sintesi dei ribosomi e dell’RNA ribosomiale (rRNA). Il nucleo è racchiuso da
una doppia membrana nucleare interrotta da pori attraverso i quali passano, oltre alle
macromolecole, le due sub-unità dei ribosomi che si assemblano solo nel citoplasma. La
membrana nucleare è connessa con il reticolo endoplasmico da cui si origina.
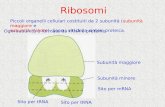
Ribosomi. Sono particelle presenti sia nelle cellule procariotiche che eucariotiche, con una
dimensione di 17-30 nm, costituite da molecole di RNA ribosomiale e proteine che si associano a
formare due subunità di dimensioni differenti. Ne esistono due classi: quelli presenti nel citosol e
quelli associati al reticolo endoplasmatico ruvido. Si presentano come particelle individuali o
riunite in aggregati detti polisomi, tenuti insieme da un filamento di mRNA. Sono sede della sintesi
proteica: quelli liberi sintetizzano le proteine per il metabolismo cellulare; quelli associati al RER
codificano le proteine destinate ad essere escrete dalla cellula. Si ricorda che i ribosomi non sono
presenti nel nucleo ma si trovano nel citoplasma; sono presenti anche nello stroma dei mitocondri
e plastidi.
Reticolo endoplasmatico (RE). E’ costituito da una rete di membrane che formano sacche
appiattite (cisterne) o strutture tubulari (tubuli). Lo spazio interno che si viene a formare è
chiamato lume e nella maggior parte delle cellule dà origine ad un unico compartimento interno
che è in comunicazione con lo spazio presente fra la membrana nucleare interna e la membrana
nucleare esterna. Il reticolo endoplasmatico funziona come un sistema di comunicazione
all’interno della cellula per il trasporto di materiali, quali proteine e lipidi. E’ connesso con i
plasmodesmi permettendo, in questo modo, di favorire l comunicazione tra cellula e cellula. E’
tuttavia anche il luogo dove avvengono la sintesi e l’assemblaggio di alcune proteine e la sintesi di
alcuni lipidi ed alcuni carboidrati. Il reticolo endoplasmatico viene classificato in due tipologie: 1.
RE rugoso costituito da cisterne con numerosi polisomi aderenti alla loro superficie esterna che ha
un ruolo fondamentale nella sintesi e nell’assemblaggio delle proteine. Molte delle proteine che
sono esportate fuori della cellula e quelle destinate ad altri organuli sono sintetizzate sui ribosomi
attaccati alla membrana del reticolo endoplasmatico. In alcuni casi queste proteine vengono
modificate mediante l’aggiunta di carboidrati (glicoproteine) o lipidi (fosfolipidi); RE liscio
costituito prevalentemente da tubuli, privi di ribosomi, che svolge un ruolo importante nella

sintesi di numerosi lipidi (l’altro organulo coinvolto nella sintesi dei lipidi è il plastidio) e
carboidrati. I corpi oleosi sono gocciole lipidiche che si originano nel reticolo endoplasmatico e da
qui vengono rilasciati nel citosol; sono particolarmente abbondanti nei frutti e nei semi. Circa il
45% del peso di semi di girasole, arachide, lino e sesamo è costituito di olio utilizzato dalla pianta
per sostenere il suo sviluppo.
Apparato del Golgi. Il termine apparato del Golgi viene usato quando si vuole indicare il
complesso dei corpi di Golgi di una data cellula. I corpi di Golgi (o dittiosomi) sono formati da
numerosi gruppi di dischi appiattiti o cisterne impilati e che spesso si presentano ramificati ai
margini in una complessa rete di tubuli. Ciascuna di queste cisterne ha uno spazio interno, detto
lume. L’apparato del Golgi possiede due facce distinte: una di formazione, o cis, che è
strutturalmente associata con la porzione liscia del reticolo endoplasmatico; e una di maturazione,
o trans, che è rivolta verso il plasmalemma e dalla quale gemmano le vescicole. I corpi di Golgi
sono coinvolti nei processi di secrezione che, nelle piante, consistono principalmente nella sintesi
e nella secrezione dei polisaccaridi non cellulosici della parete cellulare (emicellulose e pectine). Il
complesso di Golgi, inoltre, processa, smista e modifica le glicoproteine e le proteine destinate al
vacuolo. Il percorso generale è dai ribosomi al lume del RE rugoso, da qui al complesso del Golgi e
quindi alla destinazione finale. Specificatamente, i polipeptidi, sintetizzati a livello dei ribosomi,
vengono trasportati al RE rugoso dove avviene la produzione della glicoproteina mediante
assemblaggio della proteina ed successiva aggiunta del carboidrato alla proteina. La glicoproteina
viene trasportata mediante vescicole di trasporto, che si formano dalla membrana del RE e si
muovono su microtubuli, alla superficie cis del complesso del Golgi dove può subire alcune
modificazioni (ad esempio, i carboidrati della glicoproteina possono essere modificati). La
glicoproteina, nella regione trans, viene impacchettata in vescicole secretorie che si staccano dalla
membrana del Golgi trasportando il loro contenuto a specifiche destinazioni. Le glicoproteine
svolgono un ruolo di grande importanza nel riconoscimento cellulare. Per esempio, le
glicoproteine presenti nelle cellule che formano i peli radicali del trifoglio bianco (Trifolium repens)
sono in grado di riconoscere, quindi legarsi alla parete cellulare del batterio Rhizobium trifoli. Una
volta legatisi ai peli radicali, questi batteri infettano le radici e causano la formazione dei noduli
all’interno dei quali avviene la fissazione dell’azoto atmosferico. Nel sangue umano, la diversa
natura della glicoproteina presente sulla superficie dei globuli rossi, invece, consente di classificarli
in uno dei quattro gruppi sanguigni A, B, Ab e 0. Qualora, un individuo riceva sangue non

compatibile con il proprio, vengono riconosciuti come «estranei» i globuli rossi con una diversa
glicoproteina di superficie e si attiva di conseguenza un meccanismo che li distrugge.
2.10.5. Organuli coinvolti nel metabolismo energetico. Gli organuli coinvolti nel metabolismo
energetico sono i plastidi ed i mitocondri. I plastidi si trovano solo nelle alghe e nelle piante (i
cianobatteri, procarioti fotosintetici, non contengono plastidi ma solo un sistema di
endomembrane, dette tilacoidi, su cui sono presenti i pigmenti fotosintetici). I mitocondri sono
invece presenti in tutte le cellule eucariotiche.
Plastidi. Dal momento che non sono presenti in tutte le cellule degli organismi viventi, i plastidi
sono da ritenersi organuli non indispensabili al funzionamento della cellula (al contrario del
nucleo, dei ribosomi e dei mitocondri). Tuttavia, la loro presenza, ed in particolare quella dei
cloroplasti, rende possibile la vita autotrofa. Esistono diversi tipi di plastidi che si differenziano in
base alla loro struttura ed alla funzione che essi svolgono nella pianta: i cloroplasti contengono
pigmenti fotosintetici e sono responsabili della trasformazione dell’energia luminosa in energia
chimica; i cromoplasti contengono pigmenti non fotosintetici e conferiscono una colorazione
giallo-arancio-rosso ad alcuni organi della pianta; i leucoplasti sono plastidi che accumulano
sostanze di riserva ed in particolare l’amido (gli amiloplasti) ed i lipidi (gli elaioplasti). Infine, gli
ezioplasti sono cloroplasti o proplastidi che non sono stati esposti alla luce (ad esempio, nel caso
di piante cresciute al buio) e, quindi, oltre a possedere una struttura ed una organizzazione poco
definite, non sono in grado di fare fotosintesi. Nonostante le differenze sopra accennate, tutti
plastidi hanno alcune caratteristiche comuni quali: la forma relativamente lenticolare; la presenza
di una doppia membrana lipoproteica (una membrana esterna ed una membrana interna) che
delimita una matrice interna chiamata stroma; un complesso sistema di endomembrane
(endomembrane = membrane che si sviluppano internamente all’organulo) dette tilacoidi;
presenza di DNA, RNA e ribosomi; totipotenza, cioè capacità di differenziarsi, sdifferenziarsi e
ridifferenziarsi.
Proplastidi. I proplastidi rappresentano lo stadio giovanile, ancora non differenziato dei plastidi e
si trovano, nell’embrione e nelle cellule meristematiche (cellule giovanili) degli apici radicali e
caulinari. Sono generalmente piccoli (1-3 µm), sferici o elissoidali, incolori (sono sprovvisti di
clorofilla), con un sistema di endomembrane poco sviluppato (si parla, infatti, di protilacoidi
anziché di tilacoidi). Nello stroma si possono evidenziare plastoglobuli (goccioline di lipidi con la
funzione di mantenere la conformazione delle membrane) e riserve d’amido. Dai proplastidi, col

differenziamento cellulare, derivano i diversi plastidi della cellula adulta ed, in questo senso, si
tratta di organuli totipotenti. La possibilità di trasformarsi in uno specifico plastidio piuttosto che
in un altro dipende sia da fattori ambientali, quali luce e temperatura, che dalla particolare
funzionalità dell’organo: se una plantula viene fatta crescere al buio, i proplastidi non diventano
amiloplasti, come accade nella radice, ma ezioplasti. Differentemente, i proplastidi della radice,
esposta ad illuminazione continua, si trasformano in cloroplasti.
Ezioplasti. La trasformazione dei proplastidi in cloroplasti è strettamente determinata dalla luce.
Se sperimentalmente semi di angiosperme vengono fatti germinare al buio, le piantine che si
sviluppano in queste condizioni (si parla di piantine eziolate) appaiono di coloro bianco o verde
pallido. Al buio non si sintetizza clorofilla; la sequenza di reazioni che portano alla formazione della
clorofilla si arresta allo stadio di protoclorofilla, un precursore della clorofilla. Quando posti al
buio, i proplastidi pur cominciando a differenziarsi, non completano il loro sviluppo e, al
microscopio elettronico, sono visibili plastidi (detti ezioplasti) dotati di una struttura anormale. La
struttura di un ezioplasto è molto più semplice di quella di un cloroplasto ma più complessa di un
proplastidio. Oltre a non contenere clorofilla, gli ezioplasti sono privi di sistema lamellare (si parla
di corpo prolammelare); anche la sintesi delle proteine di membrana è molto ridotta, mentre i
lipidi sono sintetizzati normalmente. Se le piante cresciute al buio vengono illuminate, gli ezioplasti
si differenziano in cloroplasti: la luce catalizza la sintesi della clorofilla e delle proteine, il corpo
prolamellare si disgrega e si trasforma gradualmente nel sistema di membrane tilacoidali tipiche
del cloroplasto.
Cloroplasti. Come già detto, i cloroplasti sono gli organuli in cui ha luogo la fotosintesi e sono
presenti in tutti gli organismi eucariotici autotrofi (alghe e piante). In un millimetro quadrato di
foglia possono essere presenti alcune centinaia di migliaia di cloroplasti e le cellule di un
parenchima clorofilliano possono contenere da 40 a 50 cloroplasti. I cloroplasti hanno una
dimensione intorno ai 5-10 µm e sono di forma ellissoidale con una faccia piana ed una convessa.
La composizione e struttura della membrana è simile alle altre membrane biologiche fatta
eccezione per una maggiore presenza di galattolipidi ed un minore contenuto di fosfolipidi. La
membrana esterna, dotata di particolari proteine, le porine, che formano dei canali, è permeabile
a molecole di piccole dimensioni sia di natura idrofila che lipofila; la membrana interna, invece, è
altamente selettiva. Internamente, i cloroplasti presentano una matrice detta stroma contenente
gli enzimi utilizzati nella fase oscura della fotosintesi (organicazione e riduzione della CO2), oltre ad

un complesso sviluppo di endomembrane, dette membrane tilacoidali o tilacoidi, che si ripiegano
su se stesse a formare delle regioni più dense e scure, chiamate grana, connesse da porzioni
lineari, i tilacoidi intergranali. Tutti i tilacoidi sono in continuità tra loro costituendo un sistema
chiuso di membrane che racchiude una singola camera interconnessa definita lume. Il microscopio
elettronico fornisce solo immagini bidimensionali del complesso sistema di membrane interne,
partendo da queste immagini si è tentato di ricostruirne la struttura tridimensionale. Secondo uno
schema tra i più accreditati, l’insieme delle connessioni segue un andamento ad elica, simile a
quello dei gradini di una scala a chiocciola. Poiché nello spessore della membrana dei tilacoidi è
localizzato l’apparato fotochimico della fotosintesi (pigmenti fotosintetici, sistemi enzimatici per il
trasferimento di elettroni e fattori di accoppiamento per la sintesi di ATP), questo tipo di
organizzazione permette di concentrare e disporre ordinatamente in ciascun cloroplasto un gran
numero di queste molecole. Inoltre, il lume offre un continuum di spazi interni interconnessi fra
loro che favorisce la creazione del gradiente protonico necessario al funzionamento dell’ATP-asi. Il
sistema dei tilacoidi è immerso nello stroma del cloroplasto in cui, oltre agli enzimi necessari alla
fase oscura della fotosintesi, si trovano anche molecole di DNA e RNA, ribosomi, plastoglobuli
(corpuscoli contenenti una grande varietà di lipidi e composti lipofili) e amido come surplus dei
materiali glucidi. Infatti, durante l’attività fotosintetica, nelle ore diurne, la quantità di zuccheri
prodotti può essere superiore rispetto ai fabbisogni metabolici della cellula; questi zuccheri non
possono essere accumulati all’interno al cloroplasto in quanto, abbassando il potenziale osmotico,
richiamerebbero acqua dal citoplasma e farebbero scoppiare l’organulo. Per questo motivo,
l’eccesso viene polimerizzato ad amido (sostanza osmoticamente inattiva) ed accumulato
temporaneamente nello stroma del cloroplasto. Durate le ore notturne, questo amido, detto
amido primario, è idrolizzato in zuccheri solubili per essere in parte consumato per le attività
metaboliche della cellula ed in parte trasportato negli amiloplasti dove viene riformato un amido
che, per semplicemente distinguerlo dal precedente, viene detto secondario.
Leucoplasti: con il termine leucoplasto si indica qualsiasi plastidio privo di colore. I leucoplasti
sono classificati in base alle sostanze prodotte e/o accumulate in: amiloplasti (accumulano
amido), elaioplasti (accumulano lipidi) e proteinoplasti (accumulano proteine). Gli amiloplasti
sono plastidi non pigmentati (incolori), per lo più privi di tilacoidi o con un sistema di
endomembrane disorganizzato, contenenti granuli di amido secondario. Si trovano in tutte le
cellule adulte del corpo della pianta che non sono esposte alla luce e quindi tipicamente in tutte le
cellule della radice e comunque nei tessuti di riserva di semi, frutti, fusti e radici. L’amido

rappresenta la riserva glucidica più importante per i vegetali e dal punto di vista chimico è
composto da due diversi polisaccaridi di α-glucosio: l’amilopectina (componente principale)
solitamente a catena ramificata e l’amilosio a catena lineare. Attenzione: l’unica differenza
chimica tra amido e cellulosa è che nel primo i legami sono principalmente α (1→4) glucosidici
mentre nel secondo sono β (1→4) glucosidici. Questa piccola differenza, tuttavia, comporta
l’impossibilità da parte dell’organismo umano di digerire la cellulosa poiché manca degli enzimi
specifici necessari per la sua degradazione. L’amido può essere accumulato negli amiloplasti per
ore, giorni, mesi, anni fino al momento del suo utilizzo: ad esempio, durante il periodo di fioritura
e fruttificazione, momento in cui la pianta necessita di notevoli risorse energetiche, oppure
all’inizio del periodo vegetativo quando il “risveglio” metabolico della pianta non è supportato da
un’adeguata attività fotosintetica. Nei semi, l’amido costituisce la fonte di zuccheri che consente
l’avvio dei processi metabolici che sono alla base della germinazione del seme e dello sviluppo
della plantula. Sovente l’accumulo di amido è così imponente che alla fine l’amiloplasto è
costituito da una grossa massa interna di amido delimitata perifericamente dalla membrana
esterna dell’amiloplasto. Gli amiloplasti presenti nelle cellule della cuffia della radice (i granuli
d’amido in questo caso prendono il nome di statoliti) hanno, invece, la funzione di percepire la
direzione della forza di gravità e innescare la risposta gravitropica positiva della radice, cioè la sua
crescita verso il basso. La forma dei granuli di amido è diversa da specie a specie ed è tipicamente
costante nella stessa specie. Quasi tutti i granuli di amido presentano un centro di formazione
(detto ilo) anche questo di forma, posizione e grandezza specie-specifica. Possono essere semplici
(la deposizione inizia in un solo punto) o composti (l’accumulo avviene contemporaneamente in
diversi punti dello stesso amiloplasto e quindi si osservano più ili). Una specifica analisi
microscopica può quindi consentire di individuare eventuali frodi perpetrate mediante l’aggiunta
alla farina di frumento di farine di altri cereali, di leguminose o di patate.
Gli elaioplasti sono coinvolti nella sintesi e nell’accumulo dei monoterpeni (lipidi volatili che
conferiscono odori, sapori, rappresentano importanti agenti farmacologici). Si trovano nella polpa
dei frutti e nelle cellule delle ghiandole di secrezione associate ai tricomi e delle cavità di
secrezione (ad esempio, della buccia dell’arancia).
Cromoplasti. I cromoplasti sono plastidi colorati (ma non fotosintetici), mancanti di un sistema
tilacoidale organizzato, che contengono caroteni e xantofille (sostanze appartenenti alla famiglia
dei carotenoidi), pigmenti responsabili del colore giallo, arancio, rosso di molti fiori (ranuncolo,

calendula), di alcuni frutti (pomodoro, peperone) e di radici (carota, barbabietola). Alcuni di questi
carotenoidi sono presenti anche nei cloroplasti (ad esempio, il ß-carotene), altri invece sono
esclusivi dei cromoplasti (ad esempio, il licopene ed il d-carotene). Come vedremo, il colore dei
fiori e dei frutti, di cui sono responsabili i carotenoidi dei cromoplasti e gli antociani disciolti nel
succo vacuolare, riveste un ruolo importante nell’attrazione dei pronubi (animali impollinatori) e
dei disseminatori (animali coinvolti nella dispersione dei semi). I cromoplasti generalmente si
differenziano a partire dai cloroplasti, come avviene nel pomodoro e nel peperone, ma possono
derivare anche da plastidi non fotosintetici quali i proplastidi ed i leucoplasti (barbabietola e
carota). La conversione dei cloroplasti in cromoplasti è facilmente osservabile: molti frutti
(peperone, pomodoro, arancia) che da principio sono verdi per la presenza di cloroplasti, in
seguito, con la maturazione, diventano di colore giallo, rosso, arancione per la conversione dei
cloroplasti in cromoplasti. Durante la maturazione la clorofilla, e più in generale l’intero apparato
fotosintetico dei cloroplasti, viene distrutta e nei plastidi compaiono i carotenoidi che possono
essere accumulati in goccioline lipidiche giallo-arancio (plastoglobuli) o sotto forma di cristalli.
Questo processo, legato a fattori endogeni alla pianta (ormoni e nutrienti) ed ambientali
(fotoperiodo e temperatura), è generalmente irreversibile. Tuttavia, in opportune condizioni può
essere reversibile: per esempio, per effetto del freddo o di intensa illuminazione i cloroplasti
possono arrossarsi per poi tornare verdi quando si ristabiliscono le condizioni iniziali. Per molto
tempo si è i ritenuto che i cromoplasti non fossero altro che cloroplasti invecchiati e
metabolicamente inattivi. Tuttavia, studi recenti condotti sul pomodoro hanno evidenziato che i
cromoplasti sono dotati di tutti gli enzimi necessari a portare avanti le reazioni tipiche della catena
respiratoria; sono cioè in grado di produrre autonomamente ATP tanto è vero che è stato coniato
il termine respirazione cromoplastica. Nelle colture agrarie, la variazione di colore del frutto è
spesso una delle manifestazioni più evidenti della sindrome di domesticazione operata dall’uomo.
Ad esempio, la variazione della bacca da verde a pigmentata nel pomodoro è avvenuta quando un
ancestrale a bacca verde ha verosimilmente accumulato le mutazioni che accompagnano la
maturazione del frutto con la perdita di clorofilla e la sintesi di pigmenti carotenoidi (β-carotene
e/o licopene) ed antociani. La carota selvatica ha una radice incolore di sapore amaro ed
internamente di consistenza legnosa; nonostante questi caratteri non particolarmente appetibili,
veniva coltivata probabilmente per le foglie ed i semi aromatici. Le giovani radici delle carote
coltivate hanno un colore pallido; dopo il primo mese di crescita cominciano a produrre pigmenti
fino ad accumularne ad alti livelli prima che la crescita secondaria sia conclusa. In base alla

pigmentazione della radici, le carote coltivate possono essere distinte in due gruppi: la carote del
tipo orientale (selezionate in Afghanistan e di colore giallo o porpora) la cui colorazione è dovuta
all’accumulo di antociani nel vacuolo delle cellule e le carote del tipo occidentale (probabilmente
selezionate in Anatolia e di colore giallo, arancione o rosso) la cui colorazione è invece dovuta ai
carotenoidi. La varietà oggi più diffusa accumula una grande quantità di carotenoidi ed è stata
probabilmente selezionata, nel sedicesimo secolo, in Olanda.
Gerontoplasti. Nelle foglie senescenti è possibile osservare plastidi, che in seguito a processi
degradativi (degradazione delle clorofille, distruzione del sistema tilacoidale, accumulo di
carotenoidi), assumono un aspetto simile a quello dei cromoplasti. In questo caso, però, si parla di
gerontoplasti, cioè di plastidi che rappresentano uno stadio degenerativo dei cloroplasti e non
vanno confusi con i veri cromoplasti (i gerontoplasti non hanno la capacità né di dividersi né di
sintetizzare ex novo i carotenoidi). La trasformazione da cloroplasto a gerontoplasto, tuttavia, non
sembra essere un processo irreversibile: alcuni studi hanno infatti dimostrato che il gerontoplasto
non è un plastidio morto bensì può essere riconvertito in cloroplasto. La funzione di questi plastidi,
probabilmente in associazione con altri organelli, è quella di attuare una sorta di controllo sulla
demolizione dell’apparato fotosintetico che avviene durante il processo di senescenza. Questo
controllo è necessario per due motivi: 1. i cloroplasti contengono circa il 75% delle proteine della
foglia; ne consegue che il catabolismo e la mobilizzazione di questo contenuto proteico fornisce
una importante sorgente di aminoacidi e di azoto per altri organi della pianta; 2. I prodotti che
derivano dalla degradazione delle clorofille possono essere, in determinate condizioni, molto
tossici per la cellula e devono essere rimossi.
Ciclo di sviluppo dei plastidi. Un plastidio deriva sempre da un altro plastidio per scissione binaria,
la forma di riproduzione tipica dei procarioti, e successivo differenziamento. Dunque, questi
organuli non sono in grado di formarsi ex novo in una cellula vegetale. In effetti, tutti i plastidi
derivano dai proplastidi dello zigote che a sua volta li ha ereditati da quelli delle cellule gamiche.
Nella maggior parte delle piante il gamete maschile concorre con il solo nucleo alla formazione
dello zigote, gli altri organuli vengono trasmessi dal citoplasma del gamete femminile, cioè per via
materna; le conifere, che hanno una trasmissione per via paterna, fanno eccezione. Il
differenziamento dei proplastidi nelle varie forme di plastidi dipende sia da fattori ambientali,
come luce e temperatura, che da meccanismi di regolazione interni relativi all’organo. Come si è
visto, se una plantula viene fatta crescere al buio, i proplastidi della foglia diventano ezioplasti,

quelli della radice amiloplasti; ma, differentemente, i proplastidi della radice esposti alla luce si
trasformeranno in cloroplasti. Non solo i proplastidi ma anche i plastidi adulti sono in grado di
dividersi e trasformarsi l’uno nell’altro. L’esposizione alla luce è accompagnata dalla conversione
degli amiloplasti in cloroplasti. I cromoplasti si differenziano dai cloroplasti, e in particolari
condizioni, come nel caso della carota esposta alla luce, è possibile anche il processo inverso.
Mitocondri. Sono gli organuli in cui avviene la respirazione aerobica, un processo che richiede
ossigeno e che consiste in una serie di reazioni che trasformano l’energia chimica, sotto forma di
carboidrati, in ATP. La cellula vegetale può contenere centinaia o migliaia di mitocondri in funzione
del tipo di cellula e dello stadio di sviluppo fino ad occupare il 20% del volume del citoplasma nelle
cellule con metabolismo molto attivo. Sono circondati da una doppia membrana che delimita una
matrice interna : la membrana esterna è liscia e permette il passaggio di molte molecole di
piccole dimensioni; al contrario, la membrana mitocondriale interna è selettivamente permeabile,
può essere cioè attraversata solo da alcuni tipi di molecole. Tale membrana è ripiegata per
formare estroflessioni, chiamate creste mitocondriali, che si estendono all’interno della matrice.
Le creste aumentano enormemente la superficie della membrana mitocondriale interna, fornendo
un’area in cui avvengono gran parte delle reazioni che trasformano l’energia chimica in ATP. Nella
matrice, che comprende anche molecole di DNA, RNA e ribosomi, sono presenti enzimi coinvolti
nel ciclo di Krebs. I mitocondri derivano tutti dai mitocondri dello zigote e quindi, come nel caso
dei plastidi, dal gamete materno. Come già evidenziato per i plastidi, si dividono per scissione
analogamente ad un batterio.
2.10.6. Vacuolo. I vacuoli sono delle cisterne all’interno della cellula, di forma tondeggiante,
ripiene di un succo (succo vacuolare) ed esternamente delimitate da una singola membrana
lipoproteica chiamata tonoplasto. Nelle cellule vegetali meristematiche e quindi giovanili si
trovano numerosi e piccoli vacuoli, che durante il differenziamento confluiranno in un unico
vacuolo centrale (può occupare anche più del 90% del volume delle cellule adulte), il che relega il
citoplasma ad un sottile strato addossato al plasmalemma. La membrana del vacuolo (tonoplasto)
è asimmetrica nel senso che la superficie esterna è più ricca di proteine integrali (pompe
protoniche, carriers, proteine canale, enzimi) rispetto a quella interna. All’interno del vacuolo è
presente una soluzione acquosa chiamata succo vacuolare contenente numerose sostanze, di
natura diversa, che possono essere o disciolte, o presenti allo stato cristallino o variamente
condensate. I vacuoli possono accumulare cationi (K+, Ca++, Mg++) ed anioni (Cl-, SO42-, NO3-),

metaboliti secondari (fenoli, alcaloidi, glucosidi), sostanze di riserva (come glucosio, fruttosio,
saccarosio, fruttani), acidi organici come acido succinico (nei germogli di erba medica), acido
malico e acido citrico (nei frutti immaturi), acido ossalico. I vacuoli possono contenere pigmenti
idrosolubili come i flavonoidi (antociani e flavoni). Il pH del succo vacuolare ha un valore medio
compreso tra 5,0 e 5,5 ma l’intervallo di tale valore può estendersi da circa 2,5 (i vacuoli del frutto
di limone) a valori maggiori di 7 (nei vacuoli di riserva delle proteine).
Funzioni del vacuolo. I vacuoli delle piante sono compartimenti che svolgono diverse funzioni
riguardanti, a seconda dei casi, la distensione cellulare, il turgore cellulare, i processi di
detossificazione, le funzioni di riserva.
Distensione cellulare ed osmoregolazione. A differenza delle cellule animali in cui per la crescita è
necessaria la sintesi dell’intero protoplasma, nelle cellule vegetali la crescita avviene soprattutto
grazie all’aumento del contenuto d’acqua del vacuolo; anche le cellule vegetali devono produrre
nuove proteine, membrane ed organelli, ma a parità di volume, in minore quantità rispetto alle
cellule animali. La presenza del vacuolo centrale fa sì che il citoplasma venga confinato ad un
sottile strato periferico aumentando, così, la superficie di scambio tra la cellula e l’ambiente
esterno. Come si è già avuto modo di evidenziare a proposito dei processi osmotici che avvengono
nella cellula, il vacuolo svolge anche un ruolo fondamentale nella regolazione della pressione di
turgore grazie al trasporto attivo di ioni (ioni K+ per la maggior parte delle piante e Na+ per le
alofite, specie vegetali adattate a crescere su suoli salini) mediato dalle pompe protoniche
presenti sul tonoplasto e loro accumulo all’interno del vacuolo. Il movimento dell’acqua viene
invece mediato da aquaporine specifiche del tonoplasto (aquaporine -TIP) che rappresentano
probabilmente le proteine del tonoplasto presenti in maggior abbondanza.
Attività litica. In analogia a quanto avviene nei lisosomi delle cellule animali, i vacuoli possono
svolgere anche un’attività litica grazie alla presenza di idrolasi acide (proteasi, nucleasi, lipasi,
glicosidasi) che contribuiscono alla rottura ed al riciclo di quasi tutti i componenti cellulari. Questo
riciclo si rende necessario non solo per il ricambio delle strutture cellulari, ma anche per il
recupero di nutrienti durante la morte cellulare programmata (apoptosi) connessa con lo sviluppo
e la senescenza.
Funzione di riserva. I vacuoli possono accumulare ed immagazzinare saccarosio, fruttosio,
sorbitolo e fruttani, oltre a grandi quantità di proteine. Quest’ultima funzione caratterizza i vacuoli

di riserva proteica PSV (Proteine Storage Vacuole) tipici dei semi (dove si trovano nei cotiledoni e
nell’endosperma) ma talvolta abbondanti anche nei tuberi e nelle foglie. Un esempio è fornito dai
granuli di aleurone dell'endosperma (cioè il tessuto nutritivo) dei semi di ricino. Questi sono
piccoli vacuoli nei quali le proteine precipitano in seguito alla disidratazione del seme. Durante la
germinazione e le prime fasi di crescita della plantula, il contenuto dei corpi proteici viene
idrolizzato per fornire substrati e energia per la crescita della plantula. Nei semi delle Graminacee
la parte esterna dell’endosperma è formata da uno o più strati di cellule (strato aleuronico) ricche
di granuli di aleurone o glutine. Il glutine è una sostanza lipoproteica che si origina dall'unione, in
presenza di acqua, di due tipi di proteine: la gliadina e la glutenina presenti principalmente
nell'endosperma delle cariosside dei cereali quali frumento, farro, segale, kamut e orzo.
Funzione di accumulo di composti tossici e metaboliti secondari. I vacuoli svolgono un ruolo
molto importante nei processi di detossificazione della pianta accumulando composti
potenzialmente tossici. La presenza di elevate concentrazioni di metalli pesanti induce, ad
esempio, l’accumulo nel vacuolo di fitochelatine, molecole in grado di chelare (cioè di legare) i
metalli pesanti, che vengono rimossi dal citoplasma e trasportati nel vacuolo dove formano il
complesso metallo-fitochelatine. All’interno del vacuolo possono essere depositati anche
precipitati di ossalati di calcio in forma di rafidi (cristalli prismatici aghiformi disposti a fascetti),
druse (cristalli prismatici a bipiramidi tetragonali), stiloidi (cristalli prismatici massicci). La
formazione di questi cristalli potrebbe essere connessa a processi di detossificazione, ad esempio
dall’eccesso di calcio assorbito dalle radici (ne sono ricche ad esempio le specie che vivono in zone
aride e su suoli ricchi di calcio), ma anche ad una funzione di difesa nei confronti degli insetti
fitofagi. Gli ossalati sono particolarmente abbondanti negli spinaci, nelle barbabietole, nelle
carote, nei piselli, nei fagioli, pomodori, nel cacao, nei kiwi, nelle foglie di thè e se consumati in
eccesso possono favorire l’insorgenza, nell’uomo, di gravi calcolosi renali. Nel vacuolo si
accumulano glucosidi ed alcaloidi. Uno dei principali glucosidi cianogenetici è l’amigdalina dei semi
e delle foglie delle Rosaceae (mandorle amare, albicocche, pesche, ciliegie, prugne) che per idrolisi
libera acido cianidrico. Gli alcaloidi, composti contenenti azoto, possono essere molto tossici per
l’uomo ma rivestono al tempo stesso un ruolo di grande importanza sia da un punto di vista
economico che medicinale. Infatti le proprietà che caratterizzano alcune piante di importanza
fondamentale per l'economia di diversi popoli (per esempio il caffè, il tè, il tabacco, ecc.) sono
dovute alla presenza di determinati alcaloidi; d'altro canto sono alcaloidi i principi attivi di varie
droghe, quali la marijuana, la cocaina, l'oppio, l'hashish. In medicina, gli alcaloidi rivestono

un'importanza del tutto particolare in quanto la loro azione farmacologica si esplica
prevalentemente sul sistema nervoso influenzando quindi non solo le facoltà volitive e intellettive,
ma anche tutte quelle funzioni, quali il metabolismo, la pressione sanguigna, la temperatura
corporea, la respirazione, la diuresi, che sono regolate da impulsi nervosi.
I metaboliti secondari sono sostanze che, sebbene non coinvolte direttamente nei processi vitali
della cellula, svolgono una funzione estremamente importante nei meccanismi di interazione
pianta-ambiente. Questi metaboliti (fenoli, tannini, alcaloidi, resine, oli essenziali) possono essere
secreti all’esterno della cellula o venire accumulati all’interno di essa. Nel primo caso essi possono
localizzarsi a livello delle pareti mentre nel secondo caso il vacuolo rappresenta la sede
intracellulare in cui questi composti vengono segregati in modo da evitarne l’interazione con i
diversi componenti cellulari visto che molte di queste sostanze sono citotossiche. Come detto
possono svolgere importanti funzioni ecologiche, in quanto proteggono la pianta da organismi
animali predatori, e sono coinvolti nei meccanismi di resistenza ai patogeni. Alcuni composti
fitotossici possono essere vantaggiosi nella competizione fra specie diverse che condividono lo
stesso habitat, un fenomeno chiamato allelopatia. Molte di queste sostanze vengono utilizzate
anche dall’uomo in campo farmacologico. Tra i composti fenolici, i flavonoidi comprendono gli
antociani, pigmenti di colore variabile dal rosso al viola (dipende dal pH del mezzo) che danno il
colore ai petali dei fiori (ad esempio geranio e calendula), a diversi frutti (ad esempio, uva e
ciliegia) e a radici come il ravanello e la barbabietola, e le antoxantine che danno colore giallo alle
uve bianche, ai cedri ed ai limoni. Gli antociani svolgono un ruolo importante anche in piante
giovani o con getti nuovi, proteggendole dai raggi ultravioletti quando la produzione di clorofilla e
di cere non è ancora iniziata. L'intera pianta può assumere una colorazione rosso-brunastra (come
per esempio i nuovi getti di rose in primavera), che si riduce man mano che la produzione di
clorofilla inizia. Il crescente interesse dell’uomo per queste sostanze deriva dalle loro proprietà
antiossidanti che le rendono efficaci protettori dell’integrità vascolare, antinfiammatori, ed agenti
antitumorali.
2.10.7. Altri organuli.
Gliossisomi: organuli di piccole dimensioni delimitati da una singola membrana, molto ricchi di
proteine enzimatiche. Sono coinvolti nel metabolismo lipidico ed in particolare nella
trasformazione dei grassi in carboidrati

Perossisomi: sono organuli coinvolti nel processo di fotorespirazione nei quali l’acido glicolico
(prodotto nei cloroplasti) è ossidato ad acido gliossilico con formazione di acqua ossigenata, che è
demolita in acqua e ossigeno molecolare.
Mitosi e meiosi
Il ciclo cellulare. La cellula si riproduce mediante un processo noto come divisione cellulare
durante il quale il contenuto di una cellula è diviso tra due nuove cellule figlie. Molto importante è
il fatto che ciascuna nuova cellula eredita una replica esatta dell’informazione genetica della cellula
madre: per questo motivo le due cellule figlie, a meno che non intervengano mutazioni, sono
geneticamente uguali sia alla cellula madre che tra di loro. Negli organismi pluricellulari come le
piante una complessa successione di divisioni cellulari determina lo sviluppo completo
dell’organismo. La prima cellula che si divide, nell’ambito del ciclo vitale della pianta, è lo zigote
che si forma in seguito alla fusione di un gamete maschile con il gamete femminile. Lo zigote si
accresce in una struttura embrionale mediante divisione cellulare ed espansione delle cellule.
L’embrione è dotato di regioni specializzate, chiamate meristemi, che vanno incontro a divisioni
cellulari. Quando il seme germina, le cellule presenti all’apice meristematico della radichetta e
all’apice meristematico del germoglio continuano a dividersi producendo sempre nuove cellule
che mano a mano si differenziano e si aggregano a formare tessuti con funzioni specializzate come
ad esempio, i tessuti fotosintetici, i tessuti di riserva, i tessuti di trasporto. Le divisioni cellulari
continuano anche in un organismo che ha completato il suo sviluppo, per compensare le perdite
dovute alla morte cellulare o per garantire la crescita degli organi (ad esempio la crescita del fusto,
dei rami e della radice) oppure per consentire il ricambio di alcuni organi (si pensi alla formazione
di nuove foglie in primavera). E’ corretto dire, quindi, che le cellule meristematiche non smettono
mai di dividersi per tutta la vita della pianta. Non solo, anche le cellule già differenziate, purché
vive, possono tornare ad uno stadio meristematico e cominciare a dividersi nuovamente. Questo
avviene in quanto tutte le cellule vegetali, al contrario di quelle animali, sono totipotenti, ovvero, in
opportune condizioni, possono riacquisire la capacità di dividersi e produrre qualsiasi cellula
differenziata di un organismo. Gli stadi attraverso i quali una cellula passa da una divisione
cellulare alla successiva costituiscono il ciclo cellulare. La durata del ciclo cellulare varia ma nelle
cellule vegetali in attiva crescita è compresa fra le 8 e le 20 ore. Il ciclo cellulare consiste di due fasi
principali, l’interfase e la mitosi (o fase M), ciascuna delle quali può essere riconosciuta mediante
un microscopio ottico. L’interfase che precede la mitosi è un periodo di intensa attività cellulare,
durante il quale vengono fatto complessi preparativi per la divisione cellulare, inclusa la
duplicazione dei cromosomi e la produzione di gran parte delle proteine, dei lipidi e degli altri

materiali biologicamente importanti. L’interfase occupa la parte più lunga dell’intero ciclo cellulare
e può essere divisa in tre fasi che vengono indicate come G1, S e G2 (dove G sta per gap, un
intervallo durante il quale non c’è sintesi di DNA e S sta per sintesi, ovvero replicazione del DNA ).
Il periodo che intercorre tra la fine della mitosi e la fase S è definito fase G1: si tratta di un periodo
di intensa attività biochimica durante il quale la giovane cellula sintetizza i costituenti cellulari
(organelli, ribosomi, sistemi di membrane, enzimi, ecc.), i nucleotidi che le serviranno nella
successiva fase di sintesi del DNA ed aumenta le proprie dimensioni. Le cellule che non sono
destinate a dividersi ulteriormente si fermano in questa fase del ciclo (in tal caso detta G0). Dopo la
duplicazione del contenuto di DNA del nucleo la fase S si conclude e la cellula entra in fase G2. Il
principale ruolo è quello di preparare la cellula alla divisione mitotica sintetizzando le tubuline del
fuso mitotico ed altre proteine che interagiscono con i cromosomi. L’avanzamento del ciclo
cellulare è controllato in due punti di passaggio, chiamati punti di controllo, uno alla fine della fase
G1 e uno alla fine della fase G2. E’ al punto di controllo G1 che il sistema provvede ad arrestare il
ciclo o avviare il processo che darà inizio alla fase S. Al punto di controllo G2 il sistema provvede
ad arrestare il ciclo oppure ad avviare la mitosi. Ad esempio, le cellule dei tessuti meristematici
sono caratterizzate da un particolare tipo di divisone cellulare in cui una delle due cellule figlie
continua a dividersi mentre l’altra inizia un processo di differenziamento (fermandosi alla G0): i
processi di controllo del ciclo sono fondamentali per stabilire che una cellula continui il ciclo
cellulare mentre l’altra inizi a differenziarsi.
Prima che inizi la mitosi, i microtubuli ed i filamenti di actina si organizzano a formare una
piccola fascia intorno al nucleo, detta banda pre-profasica. Questa banda, che scompare poco
prima della divisione mitotica, costituisce una specie di “cicatrice” che definisce in anticipo il punto
in cui avverrà la divisione del citoplasma.
Le fasi in cui si esplica la divisione mitotica possono essere così riassunte:
1. Profase. La cromatina si condensa rapidamente (cioè i filamenti di DNA si accorciano ed
ispessiscono) e cominciano ad apparire i singoli cromosomi; l’involucro nucleare ed i
nucleoli scompaiano. Ogni cromosoma è stato duplicato nella precedente fase S ed ora
consiste in una coppia di unità identiche, chiamate cromatidi fratelli. Vi è una regione
chiamata centromero dove i cromatidi sono strettamente associati.
2. Metafase. Dopo che l’involucro nucleare si è dissolto, cominciano a comparire numerose
fibre parallele, chiamate fibre del fuso, costituite da microtubuli. A maturazione, il fuso
assume una forma allungata, con le fibre che si estendono dagli opposti poli alla regione

equatoriale. Queste fibre si attaccano a ciascun cromosoma ad un lato del centromero, in
una struttura proteica, a forma di disco, chiamata cinetocore. Com’è possibile osservare al
microscopio ottico, verso la fine della metafase, i cromosomi si allineano,
indipendentemente l’uno dall’altro, su un piano circolare perpendicolare all‘asse del fuso.
Questo piano viene detto piastra metafasica. I cromosomi ora sono pronti a subire la
rottura dei loro centromeri e la separazione dei loro cromatidi.
3. Anafase. I cromatidi fratelli, associati allo stesso centromero, si separano e migrano verso i
poli opposti del fuso trasportati dai microtubuli. Nel momento in cui i cromatidi si separano
e non sono più attaccati alle loro copie ogni cromatidio è considerato un cromosoma in
quanto ognuno di essi è dotato del proprio centromero. Alla fine dell’anafase, i cromosomi
risultano separati in due nuclei geneticamente uguali.
4. Telofase. I nuclei figli ripetono gli eventi che hanno caratterizzato l’anafase ma al contrario.
I microtubuli del fuso scompaiono, i cromosomi si decondensano, attorno a ciascuna serie di
cromosomi si forma l’involucro nucleare e nucleoli compaiono in ciascuno dei nuovi nuclei.
I misteri del fuso mitotico: come avviene il movimento dei cromosomi? Il movimento dei
cromosomi in anafase è il risultato di due forze: una forza è quella operata dai microtubuli
cinetocorici (cioè quelli attaccati al cinetocoro) che, accorciandosi, “tirano” i cromosomi verso i
poli opposti; l’altra è esercitata dai microtubuli polari (cioè quelli che pur convergendo ai poli del
fuso non sono attaccati al cinetocoro) che, allungandosi, “spingono” i poli del fuso in direzione
opposta. Quando i cromatidi fratelli si allontanano, i microtubuli del cinetocoro si accorciano, a
cominciare dalle estremità. Ma il movimento dei cromosomi non è solo il risultato
dell’accorciamento dei microtubuli, sembra infatti che le proteine motrici (quali la dineina)
forniscano la vera e propria forza motrice per spingere i cromosomi lungo i microtubuli cui sono
legati.
La citodieresi. Alla mitosi segue la citodieresi, cioè la divisione del citoplasma. In anafase o
telofase già si assiste ad un evento che in qualche modo prepara alla successiva citodieresi e che
consiste nella formazione, tra i due nuclei figli, di un sistema di corti microtubuli, a forma di botte.
Questo sistema, chiamato fragmoplasto, è diviso dal piano stabilito, immediatamente prima
dell’inizio della profase, dalla banda pre-profasica, e corrisponde anche alla posizione occupata
dalla piastra metafasica. Questo piano costituisce il piano di divisione a livello del quale si forma
la piastra cellulare che deriva dalla fusione delle vescicole del Golgi, trasportate in questa regione
dai microtubuli del fragmoplasto con l’aiuto delle proteine motrici e contenenti molecole della

matrice (pectine e/o emicellulose). Quando le vescicole si fondono le loro membrane
contribuiscono alla formazione della membrana plasmatica, da una parte e l’altra della piastra
cellulare. In questo momento si formano anche i plasmodesmi come segmenti del reticolo
endoplasmatico liscio, intrappolati fra le vescicole che si fondono. La piastra cellulare si accresce
con un meccanismo centrifugo (dal centro verso i lati) fino a fondersi con la parete della cellula
madre. Dopo la formazione della lamella mediana, come già si è detto a proposito della biogenesi
della parete, ciascun protoplasto produce una parete primaria, ad essa contigua. Inoltre, ciascuna
cellula figlia deposita un nuovo strato di parete primaria tutt’intorno al protoplasto in continuazione
con quella che si è formata in corrispondenza della piastra cellulare. L’aumento in volume delle
cellule figlie determina, infatti, lo stiramento e la rottura della parete della cellula madre.
Meiosi. In tutti gli organismi, la riproduzione sessuale contempla due processi fondamentali: la
meiosi, un processo di divisione cellulare che produce cellule aploidi a partire da cellule diploidi, e
la gamia, un processo che ristabilisce la diploidia attraverso la fusione di due cellule aploidi
(gameti) in una cellula diploide (zigote).
Per capire la meiosi, è utile ricordare alcuni concetti già espressi quando ho parlato del nucleo:
aploide: numero dei cromosomi nei gameti (n)
diploide: numero dei cromosomi nelle cellule somatiche (2n)
I cromosomi sono omologhi quando portano gli stessi geni (ma non necessariamente gli stessi
alleli); un omologo deriva dal gamete di uno dei genitori, il suo compagno dal gamete dell’altro
genitore). Dopo la fecondazione entrambi gli omologhi saranno presenti nello zigote. Durante la
meiosi, il corredo diploide di cromosomi, che contiene i due omologhi di ogni coppia, si riduce a un
corredo aploide, che contiene solo un omologo per ogni coppia. In questo modo, la meiosi
controbilancia gli effetti della fecondazione, assicurando che il numero dei cromosomi rimanga
costante da una generazione all’altra.
Come si è visto precedentemente, la mitosi consiste di una singola divisione e dà come risultato la
formazione 2 di cellule figlie, ciascuna delle quali è una copia esatta dei cromosomi della cellula
genitrice. La divisione meiotica, invece, consiste di due successive divisioni (meiosi I e meiosi II)
che formano in totale 4 cellule figlie contenenti ciascuna la metà del corredo cromosomico presente
nella cellula madre. Inoltre, ogni cellula figlia differisce geneticamente dalle altre cellule “sorelle”
tra loro e dalla cellula madre. Gli eventi chiave della meiosi avvengono durante l’interfase che

precede la meiosi e durante la profase della meiosi I. Durante questa interfase, al pari di quanto
accade nella mitosi, i cromosomi si duplicano; tuttavia, nonostante le due divisioni nucleari, si ha
una sola duplicazione e quindi una sola interfase che precede la prima divisione meiotica.
All’inizio della meiosi, quindi, ciascun cromosoma è composto da due cromatidi fratelli identici,
tenuti insieme dal centromero. All’inizio della profase I, dopo la duplicazione dei cromosomi, i
cromosomi omologhi si uniscono in coppie grazie all’azione di alcune proteine che, nell’insieme,
vanno sotto il nome di complesso sinaptinemale. Va sottolineato che il processo può avvenire solo
nelle cellule diploidi: nelle cellule aploidi, infatti, i cromosmi omologhi non sono presenti. Una
volta che è avvenuto il contatto tra alcuni punti dei cromosomi omologhi, l’appaiamento si estende
per l’intera lunghezza dei cromatidi, così che ogni porzione di un cromosoma omologo viene a
trovarsi vicino a quella corrispondente dell’altro omologo. Poiché ogni cromosoma si compone di
due cromatidi identici, nell’appaiamento dei cromosomi omologhi sono coinvolti 4 cromatidi. I
cromosomi omologhi appaiati sono detti bivalenti o tetradi. L’uso del termine tetradi pone l’accento
sul fatto che ci sono 4 cromatidi per bivalente. In una cellula con numero diploide 4, ci sono due
tetradi (ed un totale di 8 cromatidi); nelle cellule di mais, che ha 20 cromosomi, ci sono 10 tetradi
(ed un totale di 40 cromatidi) a questo stadio. E’ a questo punto che avviene un processo cruciale
che altera la composizione genetica dei cromosomi e che può essere considerato uno dei motori del
processo di evoluzione biologica. Durante la fase di appaiamento, i cromatidi non fratelli (ad
esempio, uno del cromosoma omologo paterno e l’altro di quello materno) si intersecano, formando
i chiasmi, e scambiano tra loro pezzi di cromosoma attraverso l’azione di enzimi che tagliano e
riuniscono le molecole di DNA. Tale fenomeno, noto come crossing-over o ricombinazione, fa sì
che alla fine dell’appaiamento un cromatidio di un cromosoma omologo contenga pezzi di DNA
dell’altro omologo e viceversa (cromosomi ricombinante). La ricombinazione genetica che ne
risulta aumenta notevolmente la variabilità genetica, cioè le nuove combinazioni di caratteri, tra i
discendenti della riproduzione sessuata. A differenza della mitosi, durante la quale i singoli
cromosomi raggiungono il fuso, nella metafase I della meiosi i cromosomi omologhi appaiati e
ricombinanti si dispongono in coppia lungo la piastra equatoriale tra i due poli della cellula, tirati
dai microtubuli del fuso. I cinetocori fratelli di un cromosoma sono connessi mediante le fibre del
fuso solo ad uno dei due poli, mentre quelli del cromosoma omologo sono collegati all’altro polo.
Durante l’anafase I, i cromosomi omologhi di ogni coppia si separano e migrano verso i poli
opposti; è questa un’altra importante caratteristica della meiosi. Mentre, infatti, durante la mitosi,
nell’anafase ogni cromosoma duplicato si scinde nei due cromatidi (a questo punto diventati a tutti
gli effetti cromosomi) fratelli che migrano ciascuno ai due poli della cellula, durante la meiosi i
cromatidi non si separano ed i cromatidi fratelli rimangono uniti; è la coppia di cromosomi

omologhi che si separa ed ognuno dei cromosomi, ancora formato da due cromatidi fratelli, migra
verso uno dei due poli. Quindi, alla fine della telofase I, ognuno dei due nuclei figli che deriva dalla
meiosi I è dunque aploide, vale a dire contiene soltanto un assetto cromosomico , rispetto ai due
presenti nel nucleo diploide di partenza. Precisamente, per ogni coppia di cromosomi omologhi ne
avrà ereditato, a caso, o soltanto quello paterno o soltanto quello materno, fenomeno noto come
assortimento indipendente dei cromosomi. Se ad esempio in un dato nucleo diploide esistono due
coppie di cromosomi omologhi, uno dei nuclei figli potrebbe ricevere il cromosoma 1 dalla madre e
quello 2 dal padre, o viceversa, oppure potrebbe ricevere entrambi i cromosomi di derivazione
paterna o ambedue di derivazione materna. Tutto dipende dall’allineamento casuale delle coppie
omologhe in corrispondenza della piastra equatoriale durante la metafase I. Naturalmente, più è
elevato il numero dei cromosomi che costituiscono un corredo genetico, minore è la probabilità che
si ristabiliscano le combinazioni parentali. Ad esempio, nel mais che ha 20 coppie di omologhi, si
possono prevedere fino a 220 (1.048.576) possibili combinazioni. Quasi contemporaneamente alla
fine della prima divisione meiotica può iniziare la citodieresi con la divisione del citoplasma in due
cellule figlie. La seconda divisione meiotica (meiosi II) è molto simile ad una normale mitosi, con i
cromosomi, ciascuno formato ancora da due cromatidi, che durante la metafase II si dispongono
lungo la piastra equatoriale. Come avviene nella mitosi, durante l’anafase II, la separazione dei
cinetocori consente che questa volta siano i singoli cromatidi, ora cromosomi figli, a migrare verso i
rispettivi poli. Con la seconda citodieresi si formano, in totale, 4 cellule figlie, ciascuna contenente
l’esatta metà del contenuto genetico della cellula madre che si è divisa per meiosi. Le 4 cellule,
seppure equivalenti nel contenuto di DNA, saranno tutte differenti fra loro per ricombinazione
dovuto a crossing-over durante la profase I e all’assortimento indipendente paterni e materni
durante l’anafase I. Questi due processi, associati all’unione casuale dei gameti maschili e
femminili nella fecondazione, fanno sì che le possibili combinazioni genetiche risultanti nella
riproduzione sessuale siano praticamente infinite.

Mitosi e meiosi portano a risultati diversi. La mitosi è una singola divisione nucleare durante la
quale i cromatidi fratelli si separano l’uno dall’altro e vengono distribuiti alle due cellule figlie
dando luogo alla fine a 2 cellule figlie diploidi identiche fra loro ed alla cellula madre. Una diploide
che entra in mitosi produce due cellule diploidi. Allo stesso modo, una cellula aploide che entra in
mitosi produce due cellule aploidi. Da notare che durante tutta la mitosi i cromosomi omologhi non
si associano mai fisicamente. La meiosi consiste di 2 divisioni nuclerai successive, meiosi I e
meiosi II. Durante la meiosi I si separano i cromosomi omologhi, mentre nella meiosi II si separano
i cromatidi fratelli. La meiosi si conclude con la formazione di 4 cellule aploidi geneticamente
differenti. I destini di queste cellule dipendono dal tipo di ciclo vitale; come vedremo negli animali
si differenziano come gameti, nelle piante diventano spore.