1. L’organizzazione generale degli animali · che lo compongono, costituiscono complessivamente...
27
Il progenitore comune di tutti gli animali era un protista coloniale. Il progenitore degli animali era probabilmente un protista flagellato unicellulare che viveva in colonie come quella che vedi nella (▶figura 1). In base all’ipotesi attualmente più accreditata (▶figura 2), all’interno delle colonie di alcuni flagellati certe cellule avrebbero assunto il ruolo di provvedere al movimento, altre alla nutrizione, altre ancora alla riproduzione e così via. Una volta messa in atto questa specializzazione funzionale, le cellule avrebbero potuto continuare a differenziarsi. Gruppi coordinati di cellule specializzate si sarebbero infine evoluti fino a dare origine ai tessuti di organismi più grandi e più complessi, come quelli che oggi chiamiamo animali. Questa differenziazione in gruppi di cellule con funzioni ben precise, che si delinea già dai primi stadi dello sviluppo embrionale, è stata certamente una chiave del successo e della enorme diversità che caratterizzano il regno animale. Figura 1. I protisti flagellati sono imparentati con gli animali. La formazione di colonie da parte di organismi unicellulari ha rappresentato una tappa importante nell’evoluzione degli organismi pluricellulari. Figura 2. Origine degli animali a partire da protozoi. Un’ipotesi di evoluzione di un protoanimale a partire da una colonia di protisti flagellati (le frecce indicano il tempo di evoluzione). La specializzazione dei tessuti è già programmata nello stadio embrionale. Gli animali sono organismi diplobionti nel senso che tutte le cellule somatiche di un animale sono diploidi e le uniche cellule aploidi sono i gameti, prodotti per meiosi all’interno delle gonadi (▶figura 3). Figura 3. Gli animali sono diplobionti. Il ciclo vitale di un animale (stella marina) mostra che l’organismo è diploide e le uniche cellule aploidi sono i gameti. 1. L’organizzazione generale degli animali . A tutt’oggi conosciamo oltre un milione di specie animali, sappiamo però che esistono milioni di altre specie che non sono ancora state descritte. Le somiglianze che accomunano tutti gli animali, e che li distinguono dagli altri organismi, sono sostanzialmente quattro: la pluricellularità, l’eterotrofia, la nutrizione per ingestione e la possibilità di movimento. La struttura generale di un animale, ossia la disposizione dei suoi organi e apparati, insieme alle funzioni delle singole parti che lo compongono, costituiscono complessivamente il suo piano strutturale. Sebbene i piani strutturali animali siano estremamente diversificati, essi possono essere considerati variazioni di quattro caratteristiche fondamentali: la simmetria corporea, la struttura della cavità corporea, la segmentazione del corpo e le appendici esterne che provvedono al movimento del corpo. Tutte queste caratteristiche influiscono sul modo in cui un animale si muove e interagisce con il proprio ambiente.
-
Upload
vuongthuan -
Category
Documents
-
view
214 -
download
0
Transcript of 1. L’organizzazione generale degli animali · che lo compongono, costituiscono complessivamente...

Il progenitore comune di tutti gli animali era un protista coloniale.
Il progenitore degli animali era probabilmente
un protista flagellato unicellulare che viveva in colonie
come quella che vedi nella (▶figura 1). In base
all’ipotesi attualmente più accreditata (▶figura 2),
all’interno delle colonie di alcuni flagellati certe cellule
avrebbero assunto il ruolo di provvedere al movimento,
altre alla nutrizione, altre ancora alla riproduzione e
così via. Una volta messa in atto questa specializzazione
funzionale, le cellule avrebbero potuto continuare a
differenziarsi. Gruppi coordinati di cellule specializzate
si sarebbero infine evoluti fino a dare origine
ai tessuti di organismi più grandi e più complessi, come
quelli che oggi chiamiamo animali. Questa
differenziazione in gruppi di cellule con funzioni ben
precise, che si delinea già dai primi stadi dello sviluppo
embrionale, è stata certamente una chiave del successo
e della enorme diversità che caratterizzano il regno
animale.
Figura 1. I protisti flagellati sono imparentati con gli animali.
La formazione di colonie da parte di organismi unicellulari ha
rappresentato una tappa importante nell’evoluzione degli
organismi pluricellulari.
Figura 2. Origine degli animali a partire da protozoi.
Un’ipotesi di evoluzione di un protoanimale a partire da una
colonia di protisti flagellati (le frecce indicano il tempo di
evoluzione).
La specializzazione dei tessuti è già programmata nello stadio embrionale.
Gli animali sono organismi diplobionti nel senso che
tutte le cellule somatiche di un animale sono diploidi e
le uniche cellule aploidi sono i gameti, prodotti per
meiosi all’interno delle gonadi (▶figura 3).
Figura 3. Gli animali sono diplobionti.
Il ciclo vitale di un animale (stella marina) mostra che
l’organismo è diploide e le uniche cellule aploidi sono i gameti.
1. L’organizzazione generale degli animali.
A tutt’oggi conosciamo oltre un milione di specie animali, sappiamo però che esistono milioni di altre specie che non sono
ancora state descritte. Le somiglianze che accomunano tutti gli animali, e che li distinguono dagli altri organismi, sono
sostanzialmente quattro: la pluricellularità, l’eterotrofia, la nutrizione per ingestione e la possibilità di movimento.
La struttura generale di un animale, ossia la disposizione dei suoi organi e apparati, insieme alle funzioni delle singole parti
che lo compongono, costituiscono complessivamente il suo piano strutturale. Sebbene i piani strutturali animali siano
estremamente diversificati, essi possono essere considerati variazioni di quattro caratteristiche fondamentali: la simmetria
corporea, la struttura della cavità corporea, la segmentazione del corpo e le appendici esterne che provvedono al movimento
del corpo. Tutte queste caratteristiche influiscono sul modo in cui un animale si muove e interagisce con il proprio ambiente.

Nel ciclo vitale degli animali, lo zigote (ovvero il frutto
della fecondazione di una cellula uovo da parte di
uno spermatozoo) va incontro a una serie di divisioni
cellulari molto rapide che originano un embrione
pluricellulare. Questa fase dello sviluppo embrionale
prende il nome di segmentazione e grazie ad essa il
numero delle cellule aumenta ed esse diminuiscono
progressivamente le loro dimensioni. Il primissimo
stadio embrionale, di 12-32 cellule, si chiama morula. Il
secondo stadio è detto blastula e ha l’aspetto di una
piccolissima sfera composta da uno strato di cellule che
racchiude al proprio interno una cavità, il blastocele.
A questo punto comincia anche a cambiare la forma
dell’embrione, prima sostanzialmente sferica: la
blastula si trasforma infatti nella gastrula, un sacco in
cui una parte della parete si ripiega verso l’interno
(▶figura 4); l’apertura che si viene a creare si
chiama blastoporo.
La maturazione dell’embrione prosegue quindi con la
gastrulazione che comporta la formazione di tre strati
di cellule ben differenziate, i foglietti embrionali,
destinati a trasformarsi nei futuri tessuti e organi.
Figura 4. La gastrulazione.
Queste fotografie ottenute al microscopio elettronico a
scansione mostrano la gastrula di riccio di mare a due stadi
diversi, uno più precoce (A) e uno più avanzato (B) in cui le
cellule hanno iniziato a differenziarsi morfologicamente le une
dalle altre.
Per avere un’idea della corrispondenza tra foglietti
embrionali e le strutture dell’adulto, consideriamo
quello che succede nella nostra specie, nella quale, i tre
foglietti embrionali sono:
1. l’ectoderma, lo strato più esterno della gastrula,
che darà origine all’epidermide e al sistema
nervoso;
2. il mesoderma, lo strato intermedio, da cui si
formeranno i muscoli, lo scheletro, il sistema
circolatorio, i reni e l’apparato riproduttore;
3. l’endoderma, lo strato esterno, destinato a formare
il rivestimento del canale digerente e degli organi
annessi, il rivestimento interno dei polmoni e
diverse ghiandole.
In base al numero dei foglietti embrionali, gli animali si
dividono in due grandi gruppi: diblastici e triblastici.
Gli embrioni degli animali diblastici, che comprendono
animali semplici come spugne e meduse, possiedono
soltanto due strati cellulari: l’ectoderma e l’endoderma.
Gli embrioni degli animali triblastici, invece sviluppano,
oltre ad ectoderma ed endoderma, anche il foglietto
intermedio (mesoderma). Gli animali triblastici, che
costituiscono la stragrande maggioranza degli esseri
viventi, si dividono a loro volta in protostomi e
deuterosomi (▶figura 5):
Figura 5. L’albero filogenetico degli animali.
I rapporti evolutivi tra gli animali sono particolarmente utili
nell'individuare diramazioni antiche nelle linee evolutive.
Nei protostomi (termine che deriva dal greco e
significa letteralmente «prima la bocca») il
blastoporo dà origine alla bocca dell’animale,
mentre l'apertura anale si forma in un secondo
momento: i protosomi più noti sono, per esempio,
gli anellidi (lombrichi), i molluschi e gli artropodi
(come insetti e ragni).
Nei deuterostomi, al contrario, dal blastoporo si
origina precocemente l’apertura anale, mentre la
bozza compare in seguito (deuterostoma significa
«la bocca dopo»). Sono esempi di deuterostomi gli
echinodermi (stelle marine e ricci di mare) e tutti i
vertebrati.

Le parole:
Specializzazione è un termine che si usa, per esempio,
nel mondo dello studio o del lavoro. Il senso è molto
simile: anche le cellule si specializzano in base alle loro
capacità naturali (il loro patrimonio genetico) e alla loro
storia individuale («dove» e «quando» si vengono a
trovare).
Morula deriva dal latino «piccola mora», per la sua
forma che ricorda il frutto.
Gastrula in latino significa invece «piccola coppa»
e blastula deriva dal greco blastós, «germe».
Quadri di sviluppo nei protostomi e nei deuterostomi.
Nella linea dei protostomi, la segmentazione dell'uovo
fecondato avviene secondo uno schema determinato.
Anche se nel corso delle divisioni le cellule vengono
artificialmente separate, ognuna da luogo a un preciso
elemento dell'embrione, a seconda della posizione che
in origine occupava nella blastula. La segmentazione
dell'uovo fecondato dei deuterostomi è invece
indeterminata. Cellule embrionali separate dopo un
certo numero di divisioni possono ancora dar luogo a
un embrione completo.
Un'altra differenza sostanziale tra protostomi e
deuterostomi consiste nel fatto che la segmentazione di
questi ultimi è radiale: le cellule si dividono secondo un
piano parallelo oppure perpendicolare rispetto all'asse
maggiore dell'uovo fecondato. Nei protostomi,
generalmente, la segmentazione è invece spirale: il
piano di divisione è obliquo rispetto all'asse
longitudinale dell'uovo e ciò determina una disposizione
spirale delle cellule (▶figura 6):
Figura 6. Quadri di segmentazione dell’uovo.
Le cellule derivate dalle prime due divisioni dello zigote sono
indicate con colori diversi, in modo che se ne possa seguire la
posizione nelle fasi successive dello sviluppo. La segmentazione
radiale produce cellule dimensionalmente uguali, situate
direttamente l'una sull'altra. Nella segmentazione spirale il
piano di divisione (freccia rossa) è invece obliquo, e le cellule
sono disposte a spirale. La segmentazione spirale è ineguale e
produce cellule di dimensioni diverse.
L'apertura boccale dei deuterostomi si origina infine a
una certa distanza dal blastoporo, che nell'adulto
corrisponde all'apertura anale. Nel caso dei protostomi,
la bocca si forma direttamente dal blastoporo, o in una
zona a esso limitrofa.
Tessuti, organi, sistemi.
Utilizzando un microscopio, nel corpo di un animale
complesso come unvertebrato (per esempio un gatto) si
possono riconoscere oltre un centinaio di tipi di cellule
diverse. Questa straordinaria specializzazione serve a
far funzionare in modo più efficiente un organismo
pluricellulare. Nel corpo di un animale si può
riconoscere un ordine gerarchico che, partendo dalla
singola cellula, arriva a quello di organismo nel suo
complesso. Questa organizzazione gerarchica
comprende i seguenti livelli: cellula, tessuto, organo,
sistema di organi e, infine, organismo.
Un tessuto è un insieme di cellule specializzate per
svolgere una determinata attività: per esempio il
tessuto epiteliale è deputato alla funzione di
rivestimento esterno del corpo e interno degli
organi. Nei vertebrati, ad esempio, ci sono quattro
tipi di tessuti fondamentali: epiteliale,
connettivo, muscolare e nervoso. Le spugne,
tuttavia, non hanno tessuti distinti.
Un organo è una struttura composta da due o più
tessuti che nell’insieme svolgono una funzione
comune. Un esempio di organo è il cuore, che è
costituto da tutti e quattro i tipi di tessuto e che
svolge la complessa funzione di «pompa»
dell’intero sistema circolatorio. Le spugne e le
meduse fanno eccezione, dato che sono del tutto
prive di organi.
Un sistema di organi (o apparato) è l’insieme di due
o più organi che svolgono una o più funzioni
complesse e correlate. Parole usate nell’anatomia,
come sistema circolatorio o nervoso, scheletrico o
immunitario, digestivo o respiratorio, si usano
anche nel linguaggio comune, con lo stesso
significato. Per esempio, il sistema tegumentario è
formato da tutti gli organi che proteggono il nostro
corpo esternamente: la pelle e i suoi annessi
(unghie, peli, capelli).
Abbiamo accennato al fatto che alcuni animali, come le
spugne, non hanno strutture definibili come tessuti (e,

di conseguenza, organi). Altri gruppi, come le meduse,
possiedono tessuti; tuttavia essi non sono organizzati
in veri e propri organi. Altri ancora, come le planarie
(vermi piatti) hanno sistemi di organi assai semplici.
Infine, alcuni animali, come i lombrichi e i molluschi,
hanno sistemi di organi complessi e sostanzialmente
simili a quelli dei mammiferi, uomo compreso. Tutto
questo ci indica che la complessità di un organismo
(specie umana compresa) non implica necessariamente
che esso sia più «avanzato» o migliore se confrontato
con animali con meno organi o meno apparati: la
misura del successo biologico, invece, è data
semplicemente da quanto quel particolare animale è
adattato al proprio ambiente. Non si può dire, per
esempio, che una medusa non svolga in modo
eccellente il suo ruolo di animale fluttuante nel mare,
infatti è perfettamente adattato a questo ambiente da
molti milioni di anni.
La maggior parte degli animali è caratterizzata da una simmetria corporea.
La forma complessiva di un animale può essere
descritta in base alla sua simmetria. Un animale viene
definito simmetrico se può essere suddiviso in almeno
due metà simili almeno secondo un piano (sezione). Gli
animali privi di piani di simmetria vengono invece
detti asimmetrici. Sebbene alcuni organismi, come le
spugne, siano asimmetrici, la maggior parte degli
animali manifesta un qualche tipo di simmetria. Durante
il corso dell’evoluzione si è passati da una condizione in
cui vi erano molti piani di simmetria a condizioni in cui
ne sono presenti solo due o più comunemente, uno. Va
sempre ricordato che la simmetria che stiamo
esaminando è spesso solo esteriore in quanto la
disposizione degli organi interni è spesso asimmetrica.
La simmetria radiale è tipica degli animali le cui
parti del corpo sono disposte intorno a un asse
principale; il corpo di questi animali, pertanto, può
essere idealmente suddiviso in metà identiche da
ogni piano passante per l’asse principale (▶figura
7). Alcuni gruppi animali, come per esempio gli
anemoni di mare e le meduse, comprendono
prevalentemente specie caratterizzate da questo
tipo di simmetria. Molti animali caratterizzati da
simmetria radiale sono sessili (cioè sedentari),
mentre altri si muovono lentamente, ma sono in
grado si spostarsi in ogni direzione.
Figura 7. La simmetria raggiata o radiale.
In questo caso il corpo dell’animale può essere idealmente
suddiviso in due parti uguali da infiniti piani di simmetria che
passano per l’asse dell’animale.
La simmetria bilaterale è caratteristica degli animali
che si spostano in una sola direzione. Il loro corpo
può essere suddiviso in due metà speculari (destra
e sinistra) da un singolo piano che passa attraverso
la linea sagittale mediana del corpo (▶figura 8).
Questo piano attraversa il corpo dal capo,
o estremità anteriore, fino alla coda, o estremità
posteriore. La simmetria bilaterale è strettamente
correlata alla cefalizzazione, ossia alla presenza di
un capo voluminoso dove sono localizzati organi di
senso e un cervello più o meno sviluppato, che
vengono quindi a trovarsi in corrispondenza
dell’estremità anteriore dell’animale.
Figura 8. La simmetria corporea.
In questo caso il corpo dell’animale può essere idealmente
suddiviso in due parti simmetriche solo da un piano di
simmetria che viene chiamato piano sagittale mediano.
Le parole:
Cefalizzazione deriva dal greco kephalé, «testa», come
anche in cefalea, mal di testa, o nel pesce
chiamato cefalo perché dotato di un capo
particolarmente grosso.

La struttura della cavità all’interno del corpo influisce sulla mobilità dell’animale.
In base alla presenza e alla struttura di una cavità
corporea piena di liquido, gli animali possono essere
suddivisi in tre tipi: acelomati, pseudocelomati e
celomati (▶figura 9).
Figura 9. La simmetria corporea.
La struttura di tre organismi a confronto. In giallo: i tessuti
derivati dall’endoderma; in rosso: i tessuti derivati dal
mesoderma; in blu: i tessuti derivati dall’ectoderma .
Negli acelomati non esistono altre cavità oltre a quella
digerente (che è aperta), negli pseudocelomati esiste
uno pseudoceloma interposto tra mesoderma ed
endoderma, mentre nei celomati è presente una cavità
all’interno del mesoderma e circondata interamente da
esso: il celoma.
Come vedremo, la struttura della cavità corporea di un
animale influisce fortemente sulle sue modalità di
movimento; in molti animali, infatti, la cavità corporea
svolge le funzioni di scheletro idrostatico. I fluidi sono
relativamente poco comprimibili, cosicché quando i
muscoli che avvolgono la cavità corporea si
contraggono, i liquidi vengono spinti verso un’altra
parte della cavità. Se i tessuti che avvolgono la cavità
corporea sono flessibili, il liquido che fuoriesce da una
parte della cavità può provocare l’espansione di un’altra
parte del corpo. In questo modo i liquidi in movimento
possono indurre lo spostamento di specifiche porzioni
del corpo; puoi comprendere il funzionamento dello
scheletro idrostatico osservando una chiocciola uscire
dalla propria conchiglia (▶figura 10).
Figura 10. Lo scheletro idrostatico.
Questa chiocciola possiede un idroscheletro che le permette di
muoversi grazie alla contrazione dei muscoli e al conseguente
spostamento di liquidi interni.
Il corpo della maggior parte degli animali è suddiviso in segmenti specializzati.
Molti animali possiedono una struttura corporea
suddivisa in segmenti, che possono essere tutti identici
(▶figura 6A) oppure differenti per forma e funzioni
(▶figura 6B).
Questa segmentazione del corpo, chiamata anche
metamerìa, facilita la specializzazione di singole parti e
si è evoluta indipendentemente numerose volte in
diversi gruppi animali (▶figura 11).
Figura 11. In molti animali il corpo è suddiviso in segmenti.
(A) In questo anellide marino tutti i segmenti si assomigliano
(metameria omonoma). Le appendici corrispondono a semplici
setole che contengono una tossina e che forniscono protezione
all’animale. (B) La segmentazione del corpo permette
l’evoluzione di segmenti differenti tra loro. I segmenti di questo
gambero differiscono tra loro per quanto riguarda la forma, la
funzione e le appendici (metameria eteronoma).

Anche nella nostra specie si può riscontrare la
metameria, ad esempio le vertebre, le costole e i
muscoli addominali sono disposti tipicamente in modo
metamerico (▶figura 12).
Figura 12. Anche nell’uomo si riscontra metameria.
Anche nella nostra specie si può ritrovare un esempio di
metameria, nella fotografia si vedono i muscoli addominali (la
cosiddetta “tartaruga”) di un culturista che rispecchiano appunto
la ripetizione di segmenti corporei simili.
Le parole:
Celoma deriva dall’aggettivo greco kôilos, «cavo», con
un evidente richiamo al significato biologico del
termine.
Metameria deriva dal greco metá, qui con il significato
di «insieme», e mèros, «parte». Il significato è quindi
«organizzazione derivante dall’unione di più parti».
Anche nel nostro corpo se ne trovano tracce come ad
esempio le costole, le vertebre e i muscoli addominali.
Le appendici negli animali hanno principalmente funzioni motorie.
Potersi muovere con le proprie forze è importante per
molti animali, poiché permette la ricerca del cibo, la
fuga o la predazione, l’incontro con un compagno per la
riproduzione.
Le appendici che si proiettano all’esterno del corpo
aumentano enormemente le capacità dell’animale di
muoversi. I ricci e le stelle di mare possiedono
numerosi pedicelli che permettono loro di muoversi
lentamente sul substrato. Il movimento rapido, sebbene
rigidamente controllato, è enormemente aumentato
negli animali con appendici modificate a
formare arti specializzati. In due gruppi di animali, gli
artropodi e i vertebrati, la presenza di appendici
articolate costituisce uno dei fattori principali che hanno
contribuito al loro successo evolutivo.
Gli animali: eterotrofi con variazioni sul tema.
Per potersi nutrire, gli animali devono spendere energia
per muoversi fino a raggiungere la fonte di cibo o per
convogliare verso sé stessi il cibo presente nelle
vicinanze.
Gli animali capaci di spostarsi da un luogo all’altro
vengono detti mobili, mentre quelli che rimangono fissi
in un luogo vengono definiti sessili o sedentari.
Molte delle modifiche avvenute a livello del piano
strutturale degli animali influiscono sull’individuazione,
sulla cattura e sulla digestione del cibo. La necessità di
localizzare il cibo ha favorito inoltre l’evoluzione di
strutture sensoriali in grado di fornire agli animali
informazioni sull’ambiente circostante e di un sistema
nervoso capace di ricevere, elaborare e coordinare tali
informazioni.
Secondo la strategia nutritiva utilizzata, gli animali
possono essere suddivisi nelle seguenti categorie
principali: filtratori, erbivori, predatori e detritivori.
Ciascuna di queste modalità nutritive si osserva in
molti gruppi diversi di animali. Inoltre, gli individui di
alcune specie possono utilizzare più di una strategia
nutritiva; infine, alcuni animali impiegano diverse
strategie nutritive nei vari stadi del proprio ciclo vitale.
Gli animali onnivori si alimentano sia di piante sia di
altri animali (la specie umana è tipicamente onnivora).
Molti animali hanno infine una dieta variabile a seconda
del periodo del ciclo vitale, come molti
uccelli granivori (che si nutrono di semi) o frugivori (che
si nutrono di frutta) da adulti, ma che imbeccano i
pulcini con piccoli animali, come gli insetti e le loro
larve.
I filtratori catturano piccole prede
L’aria e l’acqua contengono piccoli organismi e
molecole organiche che costituiscono una potenziale
fonte alimentare per alcuni animali. Lo spostamento
dell’aria e dell’acqua consente di far confluire tali
risorse nutritive verso gli animali che sono posizionati
in un luogo adatto allo scopo.
I filtratori usano poi un dispositivo di filtrazione per
separare le particelle di cibo. Molti animali acquatici
sessili dipendono dalle correnti d’acqua che trasportano
le prede nelle loro vicinanze (▶figura 13A), come il ver-
me albero di Natale.

I filtratori mobili fanno in modo che il mezzo
contenente i nutrienti li raggiunga. Il becco serrato del
fenicottero, per esempio, filtra piccoli organismi
contenuti nell’acqua fangosa che vi viene introdotta
(▶figura 13B).
Figura 13. La nutrizione per filtrazione.
(A) Questi filtratori marini sessili si nutrono del plancton che
viene trasportato dalle correnti oceaniche. (B) I fenicotteri sono
filtratori mobili: nelle lagune e nei laghi salati, filtrano con il
becco microrganismi dall’acqua.
Gli erbivori si nutrono di piante
Gli animali che si nutrono di vegetali vengono
detti erbivori, come un bruco. Essi possiedono
tipicamente un canale digerente molto lungo e
complesso, che permette loro di digerire le varie parti
delle piante (contenenti sostanze difficili da demolire,
come per esempio la cellulosa; (▶figura 14).
Figura 14. Una singola pianta può nutrire molti erbivori diversi.
Molte specie di insetti si nutrono di una singola specie di salice,
ognuna consumando tessuti diversi.
I predatori catturano e uccidono le prede
I predatori sono dotati di caratteristiche che permettono
loro di catturare e sopraffare altri animali (detti prede).
Molti predatori vertebrati possiedono organi sensoriali
per la localizzazione delle vittime e denti o artigli affilati
per catturare e sopraffare prede anche di elevate
dimensioni (▶figura 15). Un’altra arma dei predatori è
costituita dalle tossine, come quelle contenute nel
veleno di alcuni serpenti.
Figura 15. Denti e artigli.
I denti e gli artigli di questo leone sono perfettamente adattati
alla vita da predatore.
I detritivori si cibano di materia organica
decomposta
Gli animali definiti detritivori, o saprofagi, seguono un
regime alimentare basato sul consumo di materia
organica di origine animale o vegetale in avanzato stato
di decomposizione (▶figura 16). Di solito sono
organismi di piccole dimensioni come pesci, insetti o
altri invertebrati, ma anche alcuni uccelli o mammiferi
più grandi (come gli avvoltoi o le iene) che si nutrono
prevalentemente di cadaveri.
Figura 16. Lo scarabeo stercorario.
Questi insetti detritivori si nutrono di sterco, che raccolgono per
cibarsene e per deporvi le uova, facendone palline che poi fanno
rotolare spingendole con le zampe.
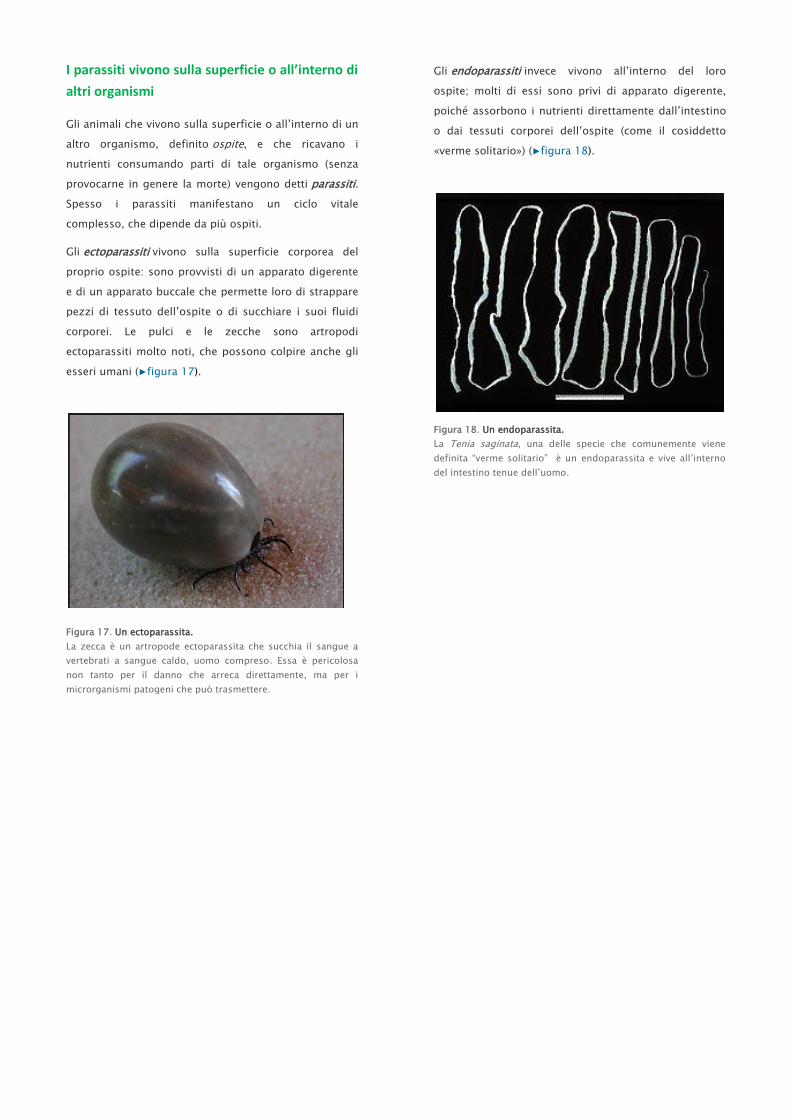
I parassiti vivono sulla superficie o all’interno di
altri organismi
Gli animali che vivono sulla superficie o all’interno di un
altro organismo, definito ospite, e che ricavano i
nutrienti consumando parti di tale organismo (senza
provocarne in genere la morte) vengono detti parassiti.
Spesso i parassiti manifestano un ciclo vitale
complesso, che dipende da più ospiti.
Gli ectoparassiti vivono sulla superficie corporea del
proprio ospite: sono provvisti di un apparato digerente
e di un apparato buccale che permette loro di strappare
pezzi di tessuto dell’ospite o di succhiare i suoi fluidi
corporei. Le pulci e le zecche sono artropodi
ectoparassiti molto noti, che possono colpire anche gli
esseri umani (▶figura 17).
Figura 17. Un ectoparassita.
La zecca è un artropode ectoparassita che succhia il sangue a
vertebrati a sangue caldo, uomo compreso. Essa è pericolosa
non tanto per il danno che arreca direttamente, ma per i
microrganismi patogeni che può trasmettere.
Gli endoparassiti invece vivono all’interno del loro
ospite; molti di essi sono privi di apparato digerente,
poiché assorbono i nutrienti direttamente dall’intestino
o dai tessuti corporei dell’ospite (come il cosiddetto
«verme solitario») (▶figura 18).
Figura 18. Un endoparassita.
La Tenia saginata, una delle specie che comunemente viene
definita “verme solitario” è un endoparassita e vive all’interno
del intestino tenue dell’uomo.

I Poriferi, privi di simmetria corporea, sono gli animali più semplici.
Le spugne (phylum Porifera, dal latino «portatori di pori»)
rappresentano uno dei primi gruppi animali evolutisi dai
protisti. Le spugne sono animali sessili, che filtrano il cibo
direttamente dall'acqua che attraversa il loro corpo. Il
flusso d'acqua viene promosso dal battito dei flagelli di
cui sono provviste le cellule che tappezzano la cavità
corporea. Queste cellule, i coanociti uniche responsabili
del processo di alimentazione, sono fornite di un
collaretto, formato da ciglia modificate, che accoglie al
suo interno il flagello. Cellule di questo tipo caratterizzano
anche i protisti flagellati appartenenti all'ordine dei
coanoflagellati, e ciò lascia ipotizzare un rapporto
filogenetico tra i due gruppi. In seguito, cambiamenti del
piano organizzativo delle spugne consentirono una più
efficiente raccolta del cibo presente nell'acqua. Le spugne
sono del resto così esclusive nella loro organizzazione
interna, che molti specialisti, nell’ambito del regno
animale, le considerano un sottoregno a parte, quello dei
parazoi.
I Poriferi, anche quelli di grandi dimensioni, hanno un
piano organizzativo particolarmente semplice. Il corpo di
una spugna consiste infatti di una libera aggregazione di
cellule, organizzate attorno a un sistema di canali
acquiferi. Una spugna non dispone né di una bocca, né di
una vera e propria cavità digerente, né di muscoli, né di
sistema nervoso; non esistono cioè organi intesi nel senso
tradizionale. Una spugna è del resto un organismo così
mediocremente organizzato che se le sue cellule vengono
dissociate e filtrate, esse possono associarsi di nuovo a
formare una nuova spugna. I Poriferi sono organismi
sessili, e restano aderenti al substrato per la maggior parte
del loro ciclo vitale.
Essi si alimentano filtrando l'acqua e trattenendo al loro
interno piccoli organismi e particelle alimentari trasportati
dalla corrente (▶figura 19).
Figura 19. Le spugne sono filtratori.
Un porifero provvede a far circolare l’acqua contenente il cibo
attraverso il proprio corpo grazie al battito di flagelli collocati
sui coanociti, cellule specializzate per la nutrizione. L’acqua
penetra nella spugna attraverso piccoli pori e passa entro canali
acquiferi, dove i coanociti catturano le particelle alimentari.
Il flusso d'acqua giunge nella cavità corporea sia attraverso
pori inalanti che attraversano singole cellule epidermiche
(nelle spugne più semplici) sia attraverso pori intercellulari
(nelle spugne a organizzazione più complessa), e le
particelle alimentari vengono trattenute dai coanociti.
L'uscita dell'acqua avviene infine attraverso una o più
aperture di grandi dimensioni denominate osculi.
Tra il sottile piano epidermico costituito da cellule parietali
dette pinacociti e quello interno dei coanociti è situato uno
strato contenente amebociti, cellule capaci di movimenti
ameboidi responsabili della sia pur modesta
comunicazione fra cellule.
2. Semplici aggregazioni di individui: i Poriferi.
In questo paragrafo descriveremo il phylum dei Poriferi che comprende animali molto semplici: le spugne. Esse sono prive di
veri e propri tessuti e la maggior parte delle loro cellule mantengono un alto grado di indipendenza e sono in grado di
cambiare forma e funzione. Anche se le spugne sono spesso grandi animali pluricellulari, sono organismi con un grado di
complessità paragonabile a quello di organismi unicellulari. Le strategie di nutrizione, l’organizzazione cellulare, gli scambi
gassosi, la riproduzione e la risposta agli stimoli ambientali avvengono con modalità simili a quelle dei protozoi.
Il fatto che siano animali sessili (ancorati al substrato), la completa mancanza di movimenti e la loro crescita generalmente
asimmetrica, avevano convinto gli antichi naturalisti che le spugne fossero piante. La loro natura animale venne riconosciuta
solo nel 1765, quando venne descritta la natura della circolazione dell’acqua all’interno del loro corpo.
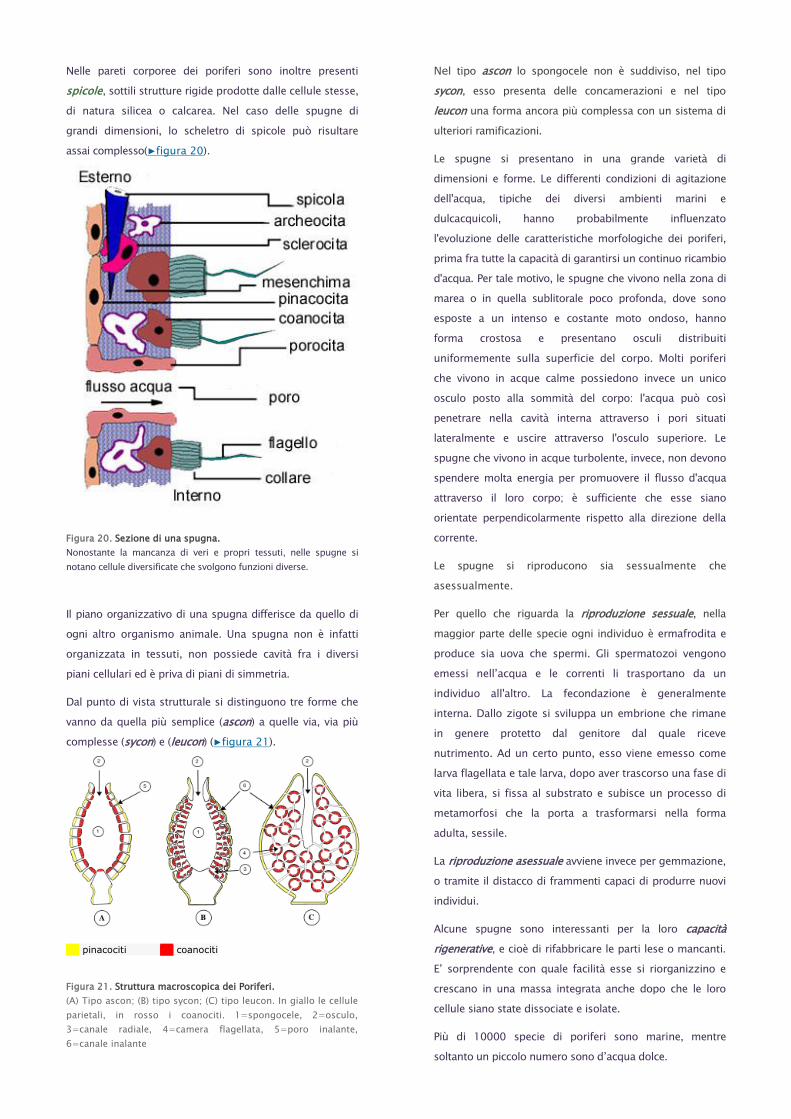
Nelle pareti corporee dei poriferi sono inoltre presenti
spicole, sottili strutture rigide prodotte dalle cellule stesse,
di natura silicea o calcarea. Nel caso delle spugne di
grandi dimensioni, lo scheletro di spicole può risultare
assai complesso(▶figura 20).
Figura 20. Sezione di una spugna.
Nonostante la mancanza di veri e propri tessuti, nelle spugne si
notano cellule diversificate che svolgono funzioni diverse.
Il piano organizzativo di una spugna differisce da quello di
ogni altro organismo animale. Una spugna non è infatti
organizzata in tessuti, non possiede cavità fra i diversi
piani cellulari ed è priva di piani di simmetria.
Dal punto di vista strutturale si distinguono tre forme che
vanno da quella più semplice (ascon) a quelle via, via più
complesse (sycon) e (leucon) (▶figura 21).
██ pinacociti ██ coanociti
Figura 21. Struttura macroscopica dei Poriferi.
(A) Tipo ascon; (B) tipo sycon; (C) tipo leucon. In giallo le cellule
parietali, in rosso i coanociti. 1=spongocele, 2=osculo,
3=canale radiale, 4=camera flagellata, 5=poro inalante,
6=canale inalante
Nel tipo ascon lo spongocele non è suddiviso, nel tipo
sycon, esso presenta delle concamerazioni e nel tipo
leucon una forma ancora più complessa con un sistema di
ulteriori ramificazioni.
Le spugne si presentano in una grande varietà di
dimensioni e forme. Le differenti condizioni di agitazione
dell'acqua, tipiche dei diversi ambienti marini e
dulcacquicoli, hanno probabilmente influenzato
l'evoluzione delle caratteristiche morfologiche dei poriferi,
prima fra tutte la capacità di garantirsi un continuo ricambio
d'acqua. Per tale motivo, le spugne che vivono nella zona di
marea o in quella sublitorale poco profonda, dove sono
esposte a un intenso e costante moto ondoso, hanno
forma crostosa e presentano osculi distribuiti
uniformemente sulla superficie del corpo. Molti poriferi
che vivono in acque calme possiedono invece un unico
osculo posto alla sommità del corpo: l'acqua può così
penetrare nella cavità interna attraverso i pori situati
lateralmente e uscire attraverso l'osculo superiore. Le
spugne che vivono in acque turbolente, invece, non devono
spendere molta energia per promuovere il flusso d'acqua
attraverso il loro corpo; è sufficiente che esse siano
orientate perpendicolarmente rispetto alla direzione della
corrente.
Le spugne si riproducono sia sessualmente che
asessualmente.
Per quello che riguarda la riproduzione sessuale, nella
maggior parte delle specie ogni individuo è ermafrodita e
produce sia uova che spermi. Gli spermatozoi vengono
emessi nell’acqua e le correnti li trasportano da un
individuo all'altro. La fecondazione è generalmente
interna. Dallo zigote si sviluppa un embrione che rimane
in genere protetto dal genitore dal quale riceve
nutrimento. Ad un certo punto, esso viene emesso come
larva flagellata e tale larva, dopo aver trascorso una fase di
vita libera, si fissa al substrato e subisce un processo di
metamorfosi che la porta a trasformarsi nella forma
adulta, sessile.
La riproduzione asessuale avviene invece per gemmazione,
o tramite il distacco di frammenti capaci di produrre nuovi
individui.
Alcune spugne sono interessanti per la loro capacità
rigenerative, e cioè di rifabbricare le parti lese o mancanti.
E’ sorprendente con quale facilità esse si riorganizzino e
crescano in una massa integrata anche dopo che le loro
cellule siano state dissociate e isolate.
Più di 10000 specie di poriferi sono marine, mentre
soltanto un piccolo numero sono d’acqua dolce.

Le classi dei poriferi.
Il phylum dei Poriferi viene suddiviso in tre classi:
1) Calcispongie
2) Ialospongie
3) Demospongie
Le Calcispongie vivono a bassa profondità, sono in
genere di grandezza ridotta, sono caratterizzate dalla
presenza di spicole calcaree e le spicole sono sempre
isolate e di varia forma: aghiformi, a tre assi o a quattro
assi (▶figura 22).
Figura 22. Calcispongie.
Sono dette spugne calcaree e possiedono uno scheletro formato
da carbonato di calcio (CaCO3). Nella foto: Leucilla nuttingi.
Le Ialospongie sono spugne tipiche delle acque
profonde. Sono spugne distinguibili dalle altre per la
presenza di un'impalcatura scheletrica esclusivamente
di spicole silicee a simmetria triassiale. Le spicole silicee
possono essere isolate o formare un reticolato. La
forma più comune è cilindrica con ampia cavità atriale,
munita di apertura con opercolo (▶figura 23).
Figura 23. Ialosponge.
Sono dette spugne vitree e possiedono strutture di sostegno
costituite da spicole silicee SiO2). Nella foto: Euplectella
aspergillum.
Le Demospongie (▶figura 24) comprendono le spugne
la cui impalcatura scheletrica è costituita da materiale
corneo (spongina), di spicole o di queste ultime miste a
spongina. L'organizzazione strutturale è complessa, con
molte e minute camere flagellate comunicanti con i pori
esalanti e inalanti tramite una fitta rete di canali. Il 95%
delle specie di Poriferi appartiene a questa classe che
comprende anche le ben note spugne da bagno.
Figura 24. Demospongie.
Possiedono strutture di sostegno costituite da spicole silicee
e/o fibre di una proteina detta spongina. Nella foto:
Xestospongia testudinaria.
Le parole:
Spugna è un termine di uso comune, ma la maggior
parte degli oggetti domestici che chiamiamo spugne
sono formati da materiali artificiali o da cellulosa. Solo
le «spugne naturali» hanno a che fare con gli omonimi
animali, sono infatti gli scheletri di un gruppo di
spugne.
Coanocita deriva dal greco choáne, «imbuto», e kýtos,
«cavità», inteso nel senso di cellula. I coanociti sono,
pertanto, «cellule a forma di imbuto».
Osculo deriva dal latino os, oris, «bocca», e quindi è
inteso come il diminutivo, nel senso di «boccuccia».
Questo nome deriva da un’errata interpretazione,
perché inizialmente si pensò che fosse un’apertura dalla
quale entra qualcosa, quindi una bocca.

Gli Cnidari o Celenterati sono animali a simmetria radiale dotati di una cavità gastrovascolare.
L'innovazione sostanziale del piano organizzativo degli
Cnidari è rappresentata da particolari cellule, definite
cnidociti, che contengono al loro interno strutture urticanti
definite nematocisti (▶figura 25). Queste ultime possono
essere espulse per la cattura di prede oppure a scopo
difensivo.
Figura 25. Le nematocisti costituiscono armi molto potenti.
I tentacoli di questa caravella portoghese sono provvisti di
numerose cellule specializzate contenenti strutture urticanti, le
nematocisti, che iniettano tossine nella preda.
Alcuni studiosi ipotizzano che gli cnidociti derivino dalla
simbiosi di Cnidari e Protisti. Tali cellule avrebbero
permesso ai primi cnidari di catturare prede di grandi
dimensioni e di ottenere così un rapido successo evolutivo. I
primitivi Cnidari si trasformarono infatti in un numero assai
elevato di specie, che durante il primo periodo Cambriano
costituivano probabilmente più di metà delle specie animali
esistenti. Attualmente il phylum degli Cnidari comprende
circa 10000 specie, per la maggior parte marine.
Il corpo degli Cnidari è caratterizzato dalla presenza di
tentacoli, sui quali di regola sono distribuite gli cnidociti. Vi
sono inoltre cellule epiteliali provviste di fibre muscolari
che consentono a questi animali di muoversi, ed è presente
un primitivo sistema nervoso a rete diffusa costituito da due
reti nervose che provvedono alla coordinazione delle varie
attività vitali. In alcuni casi, sono presenti fotorecettori
(per la percezione visiva) e meccanorecettori statici (per
l’equilibrio).
Gli Cnidari sono provvisti di un'apertura boccale che
immette in una cavità digerente a fondo cieco, definita
cavità gastrovascolare (o celenteron). Esiste dunque
un'unica via di comunicazione con l'esterno, che funge sia
da cavità boccale che anale. Tale apparato digerente,
unitamente alla presenza di tentacoli armati di cnidociti
attorno alla bocca, consente agli Cnidari di catturare e
inghiottire una gamma di prede assai più ampia di quella
disponibile per le spugne. Dopo che una preda è stata
catturata, essa viene infatti avviata alla bocca tramite i
tentacoli; gli cnidociti servono per paralizzare le prede e
trattenerle. Tali cellule sono fra l'altro responsabili
dell'azione urticante, talora intensa, che certe meduse o altri
cnidari possono causare anche all'uomo. In casi limite,
rappresentati dalla cosiddetta «vespa di mare» (genere
Chironex) o dalla «caravella portoghese» (Physalia), gli effetti
possono essere addirittura letali.
Variazioni di forma negli Cnidari.
La maggior parte degli Cnidari subisce una variazione della
forma del corpo e del tipo di riproduzione nel corso del
proprio ciclo vitale (▶figura 26).
3. L’evoluzione degli animali diblastici: Cnidari e Ctenofori.
In questo paragrafo prenderemo in considerazione due phyla di animali diblastici: gli Cnidari e gli Ctenofori.
Questi sono i primi animali in cui compare per la prima volta un’organizzazione di tipo tessutale, e quindi esistono veri e
propri tessuti. Si tratta di animali diblastici e quindi nell’embrione si differenziano due soli strati cellulari: l’ectoderma
(esterno) e l’endoderma (interno). Manca il mesoderma ma è presente uno strato gelatinoso, amorfo e non cellularizzato
interposto tra ectoderma ed endoderma: la mesoglea. Fanno la loro comparsa anche le cellule nervose e quelle muscolari e
una cavità digerente. La simmetria è radiale (Cnidari) oppure è bilaterale doppia (Ctenofori).
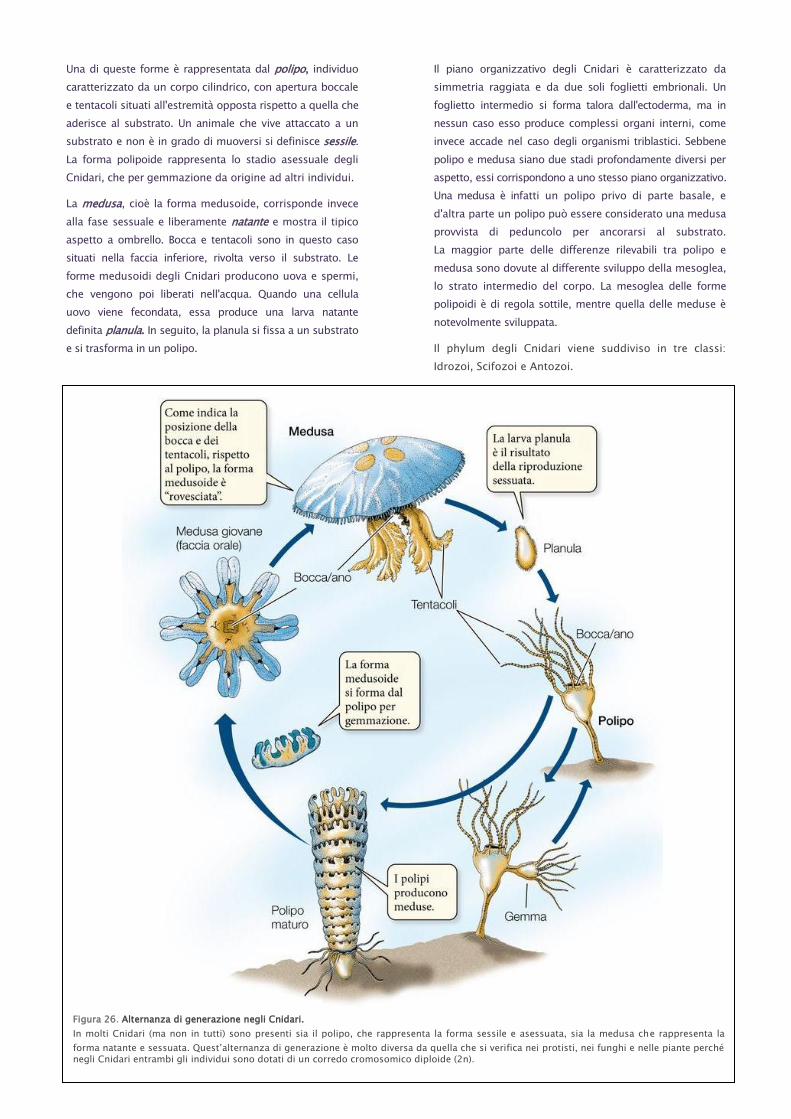
Una di queste forme è rappresentata dal polipo, individuo
caratterizzato da un corpo cilindrico, con apertura boccale
e tentacoli situati all'estremità opposta rispetto a quella che
aderisce al substrato. Un animale che vive attaccato a un
substrato e non è in grado di muoversi si definisce sessile.
La forma polipoide rappresenta lo stadio asessuale degli
Cnidari, che per gemmazione da origine ad altri individui.
La medusa, cioè la forma medusoide, corrisponde invece
alla fase sessuale e liberamente natante e mostra il tipico
aspetto a ombrello. Bocca e tentacoli sono in questo caso
situati nella faccia inferiore, rivolta verso il substrato. Le
forme medusoidi degli Cnidari producono uova e spermi,
che vengono poi liberati nell'acqua. Quando una cellula
uovo viene fecondata, essa produce una larva natante
definita planula. In seguito, la planula si fissa a un substrato
e si trasforma in un polipo.
Il piano organizzativo degli Cnidari è caratterizzato da
simmetria raggiata e da due soli foglietti embrionali. Un
foglietto intermedio si forma talora dall'ectoderma, ma in
nessun caso esso produce complessi organi interni, come
invece accade nel caso degli organismi triblastici. Sebbene
polipo e medusa siano due stadi profondamente diversi per
aspetto, essi corrispondono a uno stesso piano organizzativo.
Una medusa è infatti un polipo privo di parte basale, e
d'altra parte un polipo può essere considerato una medusa
provvista di peduncolo per ancorarsi al substrato.
La maggior parte delle differenze rilevabili tra polipo e
medusa sono dovute al differente sviluppo della mesoglea,
lo strato intermedio del corpo. La mesoglea delle forme
polipoidi è di regola sottile, mentre quella delle meduse è
notevolmente sviluppata.
Il phylum degli Cnidari viene suddiviso in tre classi:
Idrozoi, Scifozoi e Antozoi.
Figura 26. Alternanza di generazione negli Cnidari.
In molti Cnidari (ma non in tutti) sono presenti sia il polipo, che rappresenta la forma sessile e asessuata, sia la medusa che rappresenta la
forma natante e sessuata. Quest’alternanza di generazione è molto diversa da quella che si verifica nei protisti, nei funghi e nelle piante perché negli Cnidari entrambi gli individui sono dotati di un corredo cromosomico diploide (2n).
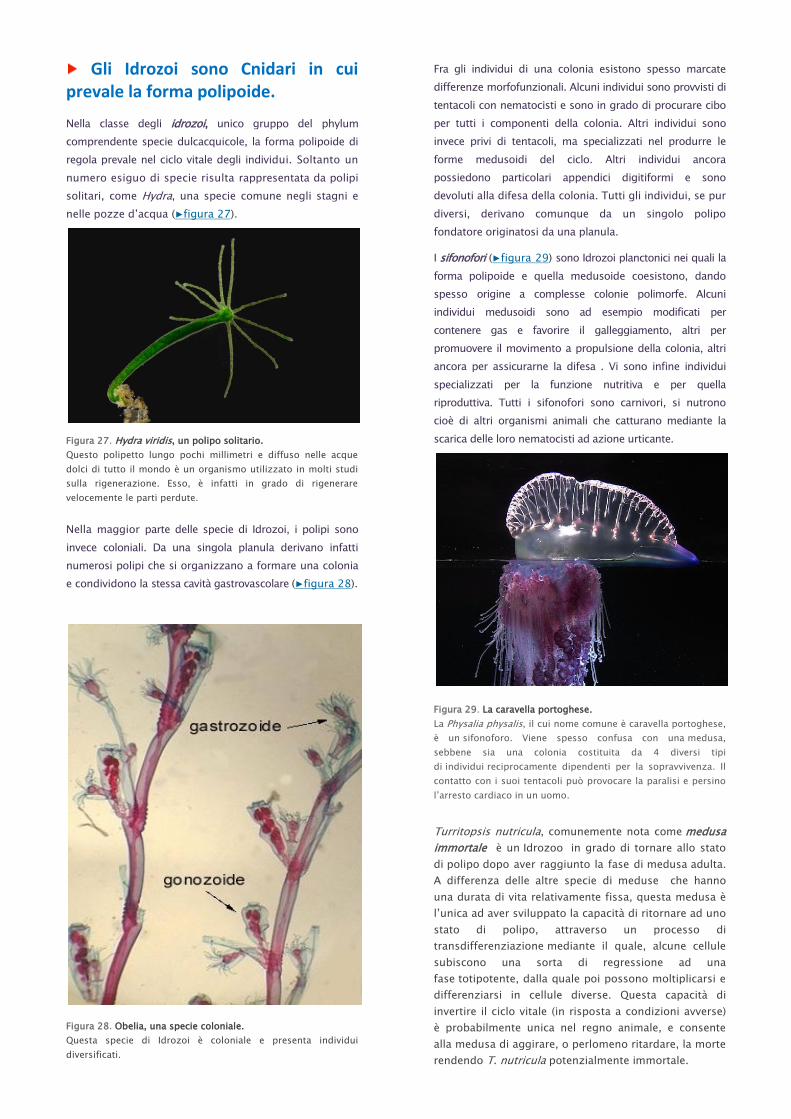
Gli Idrozoi sono Cnidari in cui prevale la forma polipoide.
Nella classe degli idrozoi, unico gruppo del phylum
comprendente specie dulcacquicole, la forma polipoide di
regola prevale nel ciclo vitale degli individui. Soltanto un
numero esiguo di specie risulta rappresentata da polipi
solitari, come Hydra, una specie comune negli stagni e
nelle pozze d’acqua (▶figura 27).
Figura 27. Hydra viridis, un polipo solitario.
Questo polipetto lungo pochi millimetri e diffuso nelle acque
dolci di tutto il mondo è un organismo utilizzato in molti studi
sulla rigenerazione. Esso, è infatti in grado di rigenerare
velocemente le parti perdute.
Nella maggior parte delle specie di Idrozoi, i polipi sono
invece coloniali. Da una singola planula derivano infatti
numerosi polipi che si organizzano a formare una colonia
e condividono la stessa cavità gastrovascolare (▶figura 28).
Figura 28. Obelia, una specie coloniale.
Questa specie di Idrozoi è coloniale e presenta individui
diversificati.
Fra gli individui di una colonia esistono spesso marcate
differenze morfofunzionali. Alcuni individui sono provvisti di
tentacoli con nematocisti e sono in grado di procurare cibo
per tutti i componenti della colonia. Altri individui sono
invece privi di tentacoli, ma specializzati nel produrre le
forme medusoidi del ciclo. Altri individui ancora
possiedono particolari appendici digitiformi e sono
devoluti alla difesa della colonia. Tutti gli individui, se pur
diversi, derivano comunque da un singolo polipo
fondatore originatosi da una planula.
I sifonofori (▶figura 29) sono Idrozoi planctonici nei quali la
forma polipoide e quella medusoide coesistono, dando
spesso origine a complesse colonie polimorfe. Alcuni
individui medusoidi sono ad esempio modificati per
contenere gas e favorire il galleggiamento, altri per
promuovere il movimento a propulsione della colonia, altri
ancora per assicurarne la difesa . Vi sono infine individui
specializzati per la funzione nutritiva e per quella
riproduttiva. Tutti i sifonofori sono carnivori, si nutrono
cioè di altri organismi animali che catturano mediante la
scarica delle loro nematocisti ad azione urticante.
Figura 29. La caravella portoghese.
La Physalia physalis, il cui nome comune è caravella portoghese,
è un sifonoforo. Viene spesso confusa con una medusa,
sebbene sia una colonia costituita da 4 diversi tipi
di individui reciprocamente dipendenti per la sopravvivenza. Il
contatto con i suoi tentacoli può provocare la paralisi e persino
l’arresto cardiaco in un uomo.
Turritopsis nutricula, comunemente nota come medusa
immortale è un Idrozoo in grado di tornare allo stato
di polipo dopo aver raggiunto la fase di medusa adulta.
A differenza delle altre specie di meduse che hanno
una durata di vita relativamente fissa, questa medusa è
l’unica ad aver sviluppato la capacità di ritornare ad uno
stato di polipo, attraverso un processo di
transdifferenziazione mediante il quale, alcune cellule
subiscono una sorta di regressione ad una
fase totipotente, dalla quale poi possono moltiplicarsi e
differenziarsi in cellule diverse. Questa capacità di
invertire il ciclo vitale (in risposta a condizioni avverse)
è probabilmente unica nel regno animale, e consente
alla medusa di aggirare, o perlomeno ritardare, la morte
rendendo T. nutricula potenzialmente immortale.

Gli Scifozoi sono cnidari in cui prevale la forma medusoide.
Tutte le varie centinaia di specie appartenenti alla classe
degli Scifozoi sono marine e alcune annoverano individui
di dimensioni superiori ai cinquanta centimetri. Lo strato
di mesoglea risulta in questo caso particolarmente spesso
e compatto e conferisce alle meduse il loro tipico aspetto.
Esse hanno complessivamente la forma di una coppa
capovolta (ombrella), al di sotto della quale sono situati i
tentacoli provvisti di nematocisti. Il movimento è reso
possibile dalla contrazione delle fibre muscolari disposte ai
bordi dell'ombrella, le quali favoriscono l'espulsione
dell'acqua dalla cavità gastrovascolare. Quando i muscoli
si rilasciano, l'ombrella torna a espandersi e a riempirsi di
acqua. Il cibo catturato mediante i tentacoli viene avviato
alla bocca, e da qui nella cavità digerente. Quest'ultima è
suddivisa in quattro camere gastriche, al cui interno
avviene la digestione a opera di particolari enzimi. Alcune
meduse appartenenti alla classe degli Scifozoi sono molto
comuni nei nostri mari (▶figure 30, 31, 32, 33).
Figura 30. La medusa quadrifoglio.
Aurelia aurita è una delle meduse più note e diffuse. L’ ombrello
è diafano e trasparente e presenta 4 strutture circolari,
le gonadi, che formano una struttura a forma di quadrifoglio.
Possiede corti e sottili tentacoli urticanti e 4 braccia più spesse
che dipartono dal centro dell'ombrello.
Figura 31. La medusa luminosa.
Pelagia noctiluca presenta il fenomeno della bioluminescenza ed
è comune nel Mar Mediterraneo e nell'Oceano Atlantico. Se
viene sfiorata, provoca dolorose irritazioni. È pelagica, ma nel
periodo autunnale e primaverile si avvicina alla costa.
Figura 32. Polmone di mare.
Rhizostoma pulmo, il polmone di mare.
Figura 33. Cassiopea.
Cotylorhiza tubercolata detta Cassiopea.
Il ciclo biologico degli Scifozoi è caratterizzato dalla forma
medusoide anziché da quella polipoide. Gli organi sessuali
(gonadi) si sviluppano nei tessuti gastrodermali in
prossimità delle camere gastriche e ogni individuo
medusoide produce sia uova che spermi che vengono poi
liberati in mare aperto. Dalle uova fecondate si forma una
planula ciliata, che si fissa a un substrato e si trasforma in un
piccolo polipo. L'individuo polipoide inizia ad alimentarsi e ad
accrescersi e può formare altri polipi per gemmazione. Dopo
un certo periodo di crescita, il polipo comincia a produrre
piccoli individui medusoidi mediante un processo ripetuto
di divisione trasversa del corpo di forma colonnare, definito
strobilazione. I nuovi individui così formatisi si accrescono e
si trasformano in meduse adulte. In definitiva, il polipo che
si origina da un singolo uovo fecondato è in grado di
produrre numerosi individui medusoidi identici
geneticamente, che in seguito effettueranno il processo di
riproduzione sessuale.
Tra gli Scifozoi vengono spesso incluse anche le cubomeduse
le cui meduse sono dotate di ombrello cuboidale e 4
tentacoli. Per la maggior parte sono meduse tropicali di
piccole dimensioni, ma molto velenose come le cosiddette
“vespe di mare”.

Gli Antozoi sono cnidari in cui manca la forma medusoide.
Le circa 6000 specie di anemoni di mare e coralli
costituiscono la classe degli Antozoi. Gli appartenenti a
questa classe differiscono da tutti gli altri Cnidari per la
totale assenza dello stadio medusoide dal loro ciclo
biologico, e per il fatto che i gameti (uova e spermi)
vengono prodotti da individui polipoidi. Dalle uova
fecondate si sviluppa una planula che si trasforma
direttamente in un polipo. Molte specie di Antozoi possono
inoltre riprodursi asessualmente per gemmazione o per
scissione. Al pari di tutti gli altri Cnidari, anche gli Antozoi
sono carnivori, e catturano le prede mediante la scarica
delle nematocisti situate sui tentacoli. La cavità digerente
degli Antozoi è invece più complessa di quella degli altri
Cnidari e risulta suddivisa da numerosi setti (mesenteri) in
varie concamerazioni che ne aumentano la superficie. La
maggiore presenza di enzimi digestivi aumenta così
l'assorbimento delle sostanze nutritive.
Gli anemoni di mare (▶figure 34, 35) sono Antozoi
solitari privi di strutture protettive rigide, diffusi sia nei
mari freddi che caldi. La maggior parte delle specie sono
sessili, vivono cioè aderenti a un substrato, ma alcune sono
capaci di piccoli spostamenti dopo che la loro porzione
basale (disco pedale) si è staccata dal substrato. Altre
specie, infine, possono compiere brevi spostamenti
nuotando.
Figura 34. Antozoi solitari.
Actinia equina, (pomodoro di mare).
Figura 35. Antozoi solitari.
Anemonia sulcata (capelli di venere).
I coralli (▶figure 36, 37) sono di regola organismi sessili e
coloniali. Ogni polipo della colonia secerne una matrice
organica sulla quale si deposita in molti casi uno scheletro di
carbonato di calcio. La struttura calcarea delle madrepore
ha una forma peculiare per ogni specie, e conferisce alle
colonie forme diversissime, come del resto indicano le
denominazioni attribuite alle varie specie: madrepore a
corna d'alce, a cervello, a ventaglio, a canne d'organo e
altre ancora. Man mano che una colonia cresce, gli individui
più vecchi muoiono e sui loro scheletri calcarei crescono i
nuovi individui. Le barriere coralline formate dallo scheletro
calcareo degli Antozoi coloniali si sviluppano soprattutto in
condizioni di acque limpide e calde. Esse sono
particolarmente abbondanti nell'oceano Indo-Pacifico, dove
formano strutture talora imponenti a forma di anello (atolli)
o nastro (barriere) (▶figure 38 e 39).
Figura 36 Antozoi coloniali.
Corallium rubrum (corallo rosso), una specie coloniale.
Figura 37. Antozoi coloniali.
Molte specie di Antozoi costituiscono le barriere coralline.
La Grande Barriera Corallina che fiancheggia le coste nord-
orientali dell'Australia è lunga oltre 2000 km e larga più di
150 km. Una barriera corallina lunga centinaia di miglia è
inoltre presente nel Mar Rosso ed è stato stimato che essa
contenga più materiale calcareo di quello che forma gli
edifici delle maggiori città del Nord America.

Abbiamo detto che le barriere coralline si sviluppano nelle
acque calde e limpide dei mari tropicali, notoriamente
povere di sostanze nutritive. Per lungo tempo gli scienziati
si sono dunque chiesti come i polipi di tali colonie
riuscissero a catturare zooplancton sufficiente per crescere
con tanta rapidità. La risposta è venuta dalla scoperta che
nei tessuti dei coralli sono presenti organismi simbionti
rappresentati da dinoflagellati altamente specializzati. Si
tratta di organismi fotosintetici, che forniscono ai loro
ospiti carboidrati e favoriscono la deposizione del
carbonato di calcio per lo scheletro delle colonie.
L'esistenza di questa simbiosi tra dinoflagellati e
madreporari spiega fra l'altro perché le formazioni coralline
siano presenti unicamente a una profondità limitata, dove
arriva abbondante la radiazione solare. In queste
condizioni i dinoflagellati possono infatti effettuare la
fotosintesi rimanendo protetti dai predatori all'interno
dello scheletro dei coralli.
Figura 38. Barriere coralline.
Distribuzione geografica delle barriere coralline.
Figura 39. Un atollo.
Un atollo (isola corallina).
Le ragioni del successo evolutivo degli Cnidari.
Gli Cnidari divennero gli organismi marini predominanti
durante il Cambriano, 600 milioni di anni fa, e tutt'oggi essi
rappresentano un'importante componente delle comunità
marine. La ragione del loro successo evolutivo va
essenzialmente attribuita al peculiare piano organizzativo,
che associa un tasso metabolico poco elevato con la
capacità di catturare prede relativamente grandi. La massa
corporea delle meduse e di molti polipi è essenzialmente
dovuta al grande sviluppo della mesoglea. Da ciò deriva
che anche Cnidari di grandi dimensioni come gli anemoni
di mare necessitano di modeste quantità di sostanze
nutritive, e possono digiunare per settimane o addirittura
mesi. La presenza delle nematocisti consente inoltre agli
cnidari di catturare anche prede notevolmente più attive e
strutturalmente più complesse rispetto a loro stessi. Altri
Cnidari, come ad esempio le madrepore e i coralli, possono
inoltre nutrirsi di microrganismi e sopravvivere anche in
acque povere di plancton grazie alla simbiosi con i
dinoflagellati. Tutte queste caratteristiche hanno in
definitiva consentito agli cnidari di colonizzare anche
habitat dove le prede non sono sufficienti ad assicurare la
sopravvivenza di organismi con maggiori esigenze
metaboliche.
Le parole:
Cnidàrio deriva dal greco kníde, «ortica», il che rende
bene l’idea dell’azione urticante delle nematocisti.
Il suffisso -zoo (pl. zoi) deriva dal greco zôion, «essere
vivente», inteso come «animale».
Per gli cnidari si trova associato a:
hýdor, «acqua», con allusione al fatto che
gli idrozoi sono fatti per oltre il 90% di acqua;
skýphos, «tazza», per la forma degli scifozoi;
ánthos, «fiore», perché gli antozoi sembrano più piante
fiorite che animali.

Gli Ctenofori sono animali diblastici dal corpo gelatinoso.
Gli Ctenofori sono un phylum di organismi diblastici,
marini, caratterizzati da un corpo gelatinoso e
trasparente provvisto di file di ciglia disposte a pettine
(ctenofori = portatori di pettini). Il piano organizzativo di
ctenofori e cnidari può risultare in apparenza simile, ma
tra i due gruppi esistono in realtà sostanziali differenze.
Come gli Cnidari, essi possiedono due strati cellulari
separati da una spessa mesoglea di consistenza
gelatinosa e spesso tentacoli per la cattura delle prede.
Mentre gli Cnidari hanno però molti piani di simmetria che
passano per l’asse dell’animale (simmetria radiale), gli
Ctenofori ne hanno solo due (simmetria biradiale).
Mentre negli Cnidari esiste sia la forma sessile (polipo)
che quella natante (medusa), negli Ctenofori esiste solo la
forma natante.
Gli Ctenofori hanno inoltre una cavità digerente che si
apre all’esterno mediante due pori anali situati dalla
parte opposta alla bocca; l'alimento si muove dunque in
una precisa direzione attraverso l'intestino.
Gli Ctenofori possiedono otto file di ciglia, definite pettini,
responsabili del movimento degli animali.
I tentacoli degli Ctenofori sono solidi e privi di nematocisti,
ma sono coperti da colloblasti, cellule dotate di una
superficie collosa che aderisce a piccole particelle che
servono da nutrimento. I tentacoli possono inoltre essere
retratti e portati alla bocca. In alcune specie, l'intera
superficie del corpo è ricoperta di muco con proprietà
adesive per la cattura delle prede. Tutte le oltre 100
specie di Ctenofori sono infatti organismi carnivori.
A differenza degli Cnidari, gli Ctenofori non possono
catturare prede di dimensioni uguali o superiori a loro
stessi. I loro tentacoli vischiosi possono tuttavia raccogliere
grandi quantità di piccoli organismi planctonici. Gli
Ctenofori hanno un tasso metabolico relativamente basso,
dal momento che, al pari degli cnidari, sono
essenzialmente formati da mesoglea inerte. Come gli
Cnidari, anche gli Ctenofori possono dunque svolgere con
successo il ruolo di predatori in mare aperto, dove la
presenza di prede è spesso assai scarsa.
Il ciclo vitale degli Ctenofori è semplice. Dalle gonadi
localizzate nella parete della cavità gastrovascolare i gameti
vengono liberati all'interno della cavità stessa e in seguito
emessi all'esterno attraverso la bocca o i pori anali. Il
processo di fecondazione avviene dunque in mare aperto.
In quasi tutte le specie, dalle uova fecondate si sviluppa
direttamente un organismo simile per aspetto all'adulto
(sviluppo diretto); stadio che tuttavia verrà raggiunto
gradualmente in seguito al progressivo accrescimento
dell'individuo.
Alcuni Ctenofori sono rappresentati nelle (▶figure 40, 41
e 42).
Figura 40. Ctenofori.
Pleurobrachia, uno ctenoforo dotato di tentacoli.
Figura 41. Ctenofori.
Beroe, uno ctenoforo privo di tentacoli e dal corpo globoso.
Figura 42. Ctenofori.
Cestum veneris, uno ctenoforo, privo di tentacoli e dal corpo
schiacciato che gli fa assumere l’aspetto di una cintura.
Le parole:
Ctenoforo deriva dal greco ktenís, «pettine», e foro,
«porto», con il significato di portatori di pettini.

La maggior parte degli animali possiede una cavità corporea.
Dopo la comparsa degli animali diblastici a simmetria
radiale, si è evoluto un terzo foglietto embrionale: il
mesoderma. Questo carattere si trova nei due grandi
gruppi di animali triblastici, i protosomi e i
deuterostomi. Come abbiamo già detto, il termine
protostomi significa letteralmente «prima la bocca» e fa
riferimento al fatto che nella maggior parte dei
protostomi la bocca dell’animale si sviluppa
dal blastoporo embrionale, mentre nei deuterostomi
ilblastoporo dà origine all’apertura anale del canale
gastrointestinale.
I protostomi sono estremamente diversificati, ma hanno
tutti simmetria bilaterale e un corpo caratterizzato da
un cervello (o encefalo) anteriore, che avvolge l’apertura
del canale digerente, e un sistema nervoso ventrale,
formato da cordoni nervosi longitudinali appaiati o fusi.
La comparsa di una cavità corporea interna piena di
liquido è stata una tappa significativa nella storia
evolutiva degli animali; in base a tale caratteristica, essi
possono essere suddivisi in acelomati, pseudocelomati
e celomati (▶figura 43):
gli acelomati, come i platelminti (vermi piatti), sono
privi di una cavità corporea ben delimitata. In
questi animali lo spazio tra l’intestino e la parete
corporea muscolare è occupato da un ammasso di
cellule complessivamente note come mesenchima .
Questi animali si muovono tipicamente per mezzo
del battito di ciglia.
gli pseudocelomati, come i nematodi (vermi
cilindrici), possiedono una cavità corporea
detta pseudoceloma contenente un liquido nel
quale sono sospesi molti organi interni.
i celomati, tra cui i lombrichi (vermi segmentati),
sono provvisti di celoma, ossia di una cavità
corporea rivestita da una membrana
chiamata peritoneo che avvolge anche gli organi
interni.
Gli animali celomati hanno un migliore controllo sul
movimento dei liquidi all’interno della cavità corporea
rispetto agli pseudocelomati.
Sebbene la funzione idrostatica del celoma sia
importante, la maggior parte degli animali ha sviluppato
anche uno scheletro rigido che fornisce protezione e
facilita i movimenti: come vedremo, i muscoli si
inseriscono su queste strutture, che possono essere
collocate all’interno del corpo dell’animale oppure sulla
sua superficie esterna (sotto forma di guscio o di
cuticola).
Figura 43. Le cavità corporee degli animali.
(A) Gli animali acelomati non presentano cavità corporee chiuse.
(B) Gli animali pseudocelomati sono caratterizzati da una cavità
corporea chiamata pseudoceloma. (C) Gli animali celomati
possiedono una cavità corporea (celoma) rivestita dal peritoneo,
che avvolge gli organi interni.
Le parole:
Elminta deriva da hélmins, hèlminthos, che in greco
significa «verme». Il termine ricorre nel nome scientifico
di vari gruppi, come i platelminti (da platýs, «piatto»).
Verme in biologia non è una categoria sistematica ma è
solo un termine che viene usato per indicare in modo
generico un organismo dal corpo allungato, privo di
scheletro rigido e di arti, come ad esempio platelminti,
nemertini, nematodi e anellidi. Nel linguaggio comune, i
«vermi» sono di solito i lombrichi e le larve di alcuni tipi
di insetti, il cui aspetto è senz’altro vermiforme ma la
cui biologia è del tutto diversa.
I turbellari, provvisti di ciglia, nell’acqua
generano turbella, dal latino «moti disordinati».
I trematodi possiedono ventose per aderire ai loro
ospiti (trematódes, dal greco «dotati di fori, ventose»).
I cestodi infine sono nastriformi (cestus in latino
significa cinto.
4. La comparsa del mesoderma e della simmetria bilaterale: Platelminti e Nemertini.
Questo paragrafo si occupa di due phyla di animali triblastici a simmetria bilaterale che manifestano un’ampia varietà di
dimensioni e di forme. I Platelminti (vermi piatti) sono animali relativamente semplici, provvisti di un tubo digerente a fondo
cieco e privi di sistema di trasporto interno. I Nemertini (vermi a nastro) presentano caratteristiche simili ai platelminti, ma
anche due grosse novità evolutive: la presenza di un tubo digerente completo e di un sistema di trasporto interno.

Platelminti: gli animali a simmetria bilaterale più semplici e privi di celoma.
I Platelminti, definiti anche vermi piatti, rappresentano un
phylum di organismi evolutisi da masse appiattite di
cellule. I Platelminti sono animali triblastici, caratterizzati da
simmetria bilaterale e dal corpo appiattito in senso
dorsoventrale. La loro organizzazione interna è assai più
complessa di quella di Cnidari e Ctenofori (▶figura 44).
Essi, tuttavia, non possiedono cavità del corpo, e sono privi
sia di apparato circolatorio che di strutture per il trasporto
di ossigeno. Un simile piano organizzativo comporta che ogni
cellula sia localizzata vicino alla superficie del corpo, in
modo da ricevere direttamente dall'esterno sufficiente
ossigeno e alimento; un'esigenza che viene adeguatamente
soddisfatta da una forma appiattita del corpo.
Figura 44. I platelminti possono condurre vita libera o essere
parassiti.
(A) Questo animale vive nell’apparato digerente dei ricci di
mare. Il disegno mostra la struttura anatomica dei platelminti
parassiti: poiché l’ospite fornisce loro tutti i nutrienti necessari,
non necessitano di organi digestivi, e gran parte del corpo è
occupato dagli organi riproduttivi. (B) Alcune specie di
platelminti, come questo verme marino del Pacifico, conducono
vita libera.
Le forme a vita libera strisciano sul substrato grazie
all’azione della muscolatura e alla presenza di un’ampia
superficie marginale provvista di ciglia.
Quando esiste, il canale digerente dei vermi piatti consiste
di una bocca che comunica con una cavità a fondo cieco e
si tratta pertanto di una cavità gastrovascolare. In molti
casi, tuttavia, tale sacco (intestino) è notevolmente
ramificato e ciò determina un aumento di superficie utile
per l'assorbimento delle sostanze alimentari.
Il sistema nervoso è più centralizzato rispetto a quello
degli Cnidari ed è di tipo cordonale: sono presenti due
cordoni nervosi longitudinali, uno a destra e uno a sinistra
connessi tra loro da delle commessure e nell’estremità
cefalica sono presenti dei gangli nervosi che contengono i
corpi cellulari dei neuroni (▶figura 45).
Figura 45. I platelminti presentano un sistema nervoso di tipo
cordonale e una cavità gastrovascolare.
In questa figura si evidenziano il sistema nervoso cordonale con
due cordoni nervosi e i gangli cefalici e la cavità digerente molto
ramificata.
Sono presenti i protonefridi (in greco, piccoli reni
primordiali) che hanno soprattutto una funzione
osmoregolatrice importante per le specie che vivono in
acque dolci e che devono eliminare l’eccesso di acqua che
tende a d entrare nel corpo per osmosi. I protonefridi sono
tubuli che dalla superficie corporea si addentrano
all’interno del corpo ramificandosi e terminando con delle
cellule dotate di un ciuffo di ciglia, dette cellule a fiamma. I
protonefridi sembra che oltre a eliminare l’acqua in
eccesso, abbiano anche la capacità di eliminare le scorie
metaboliche azotate e quindi possono essere considerati
organi escretori (▶figura 46).
Figura 46. I platelminti presentano un sistema escretore.
In questa figura si evidenzia il sistema escretore costituito dai
protonefridi.

Il phylum dei Platelminti viene suddiviso in tre classi:
Turbellari, Trematodi e Cestodi.
Turbellari: Platelminti a vita libera.
Sono considerati tra gli attuali vermi piatti, quelli più simili
alle forme primitive. Si tratta per lo più di piccoli organismi
marini o di acqua dolce, oltre a poche specie che vivono
sulla terraferma, in ambienti umidi. I più semplici
turbellari marini sono privi di cavità digerente, di organi
escretori e di veri e propri organi riproduttori. Le più
comuni specie di vermi piatti sono comunque
rappresentate dalle forme dulcacquicole appartenenti al
genere Dugesia, meglio conosciute con il nome di planarie
(▶figura 47 e 48).
Figura 47. I turbellari comprendono le planarie.
Dugesia, una planaria d’acqua dolce, molto diffusa.
Esse possiedono un'estremità cefalica provvista di un
organo chemiorecettore, di due occhi molto semplici
definiti ocelli, e di un rudimentale cervello formato da un
ispessimento del cordone nervoso longitudinale. Tutti i
vermi piatti attualmente viventi si nutrono di sostanze
animali; alcuni come predatori, altri come parassiti, altri
ancora come detritivori. I primitivi vermi piatti potrebbero
invece essere stati erbivori, adattatisi secondariamente a
una dieta formata da piccoli animali raccolti sullo strato
algale. Alcuni, infine, si sarebbero specializzati per vivere
parassiti sopra o all'interno di animali più grandi.
Figura 48. La faringe delle planarie è estroflettibile.
In questa figura si evidenzia la faringe estroflettibile con
l’apertura boccale alla sua estremità.
Le planarie sono dotate di capacità rigenerative veramente
straordinarie: se il corpo viene tagliato in due parti con
una sezione trasversale, ogni parte rigenera quella
mancante; se vengono effettuati molti tagli trasversali,
ogni parte ottenuta rigenera il capo dalla parte anteriore e
una coda dalla parte opposta; se viene effettuato un taglio
longitudinale che passa per il piano sagittale mediano a
livello del capo, si ottiene una planaria a due teste!
(▶figura 49)
Figura 49. La rigenerazione nelle planarie.
In questa figura si mostra l’estesa capacità rigenerativa nella
planaria.
Queste estese capacità rigenerative vengono utilizzate per
compiere la riproduzione asessuata che avviene per
frammentazione del corpo.
Le planarie si riproducono anche mediante riproduzione
sessuata e presentano ermafroditismo insufficiente, nel
senso che non possono praticare l’autofecondazione.
Durante la stagione riproduttiva, gran parte del loro corpo
è occupato dai testicoli, dalle vie spermatiche, dalle ovaie
e dagli ovidotti che trasportano i gameti all’esterno del
corpo.
Sebbene i primitivi vermi piatti fossero organismi a vita
libera, in seguito il loro piano organizzativo si è
specializzato anche per la vita parassita, che garantisce
l'assunzione di sostanze nutritive a spese di un ospite
con minimo dispendio di energia. Un'ipotesi plausibile circa
l'evoluzione delle strategie alimentari dei vermi piatti è che
essi siano gradualmente passati da una dieta a base di
organismi in decomposizione a una composta da
organismi morenti e infine da organismi viventi.
Attualmente, le più diffuse forme parassite sono
rappresentate dai Trematodi e dai Cestodi. Si tratta di
organismi che vivono nel corpo dei vertebrati, e che nel
caso dell'uomo possono causare gravi danni.

Trematodi: Platelminti parassiti.
I Trematodi sono tutti parassiti e da adulti sono quasi
tutti endoparassiti di vertebrati. Hanno comunemente
forma ovale più o meno allungata e dimensioni in
genere piccole: la loro lunghezza può infatti variare da
una frazione di millimetro fino a 10 mm. Alcune specie
possono però giungere anche a 2 cm di lunghezza.
Sono muniti di ventose e presentano vari stadi larvali. Il
termine Trematodi deriva dal greco "trema", che vuol
dire "foro", in quanto le ventose furono erroneamente
interpretate quali fori.
I Trematodi annoverano le fascìole (▶figura 50),
parassiti che infestano il fegato. Le fasciole conducono
esistenze complesse con molti stadi asessuati e con due
o più ospiti (tipicamente tre, talvolta due o quattro).
Due ospiti intermedi, che ospitano stadi larvali o
giovanili, possono essere invertebrati o vertebrati;
l’ospite definitivo, che ospita lo stadio sessualmente
maturo, è quasi sempre un vertebrato (▶figura 51).
Figura 50. La fasciola epatica.
Fasciola hepatica, un parassita che infesta il fegato di molti
animali, uomo compreso.
Figura 51. Ciclo vitale della fasciola epatica.
Il ciclo vitale della Fasciola hepatica è molto complesso. Il
parassita viene trasmesso da una chiocciola d’acqua dolce, la
Lymnea.
Tra i Trematodi sono compresi anche gli schistosomi,
ossia le fasciole del sangue, che causano nell’uomo una
grave parassitosi, la schistosomiasi (▶figura 52).
Figura 52. Schistosoma.
Schistosoma japonicum un parassita che causa la
schistosomiasi. La parassitosi che può colpire l’uomo ha forme
acute e croniche diverse a seconda delle specie in causa e degli
organi o tessuti coinvolti.
Esse affliggono circa 250 milioni di persone, per la
maggior parte in Asia e in Africa tropicali e subtropicali,
e nell’America meridionale nord-orientale. Tra le
malattie di origine parassitaria che affliggono l’uomo, la
schistosomiasi è oggi un problema sanitario mondiale,
secondo solo alla malaria.. le vittime raramente
vengono uccise subito, più spesso vengono debilitate
da anni di disagi e perdita di energia, e le loro vite
finiscono con l’essere abbreviate da malattie
secondarie. Il controllo della parassitosi, trattando le
vittime con farmaci risulta soddisfacente ma non per le
persone che sono regolarmente esposte alla re
infestazione. Vaccini efficaci non sono stati ancora
sviluppati. L’eliminazione delle feci e urine umane e
l’educazione sanitaria alla popolazione sono molto
importanti. Inoltre, è importante controllare le
chiocciole non usando veleni ma rendendo loro
l’ambiente inospitale (▶figura 53).
Figura 53. Ciclo vitale di Schistosoma.
Il ciclo di Schistosoma è più semplice di quello della Fasciola.

Cestodi: Platelminti parassiti.
I Cestodi, infine, anch’essi parassiti soprattutto di
erbivori, includono le tenie (comunemente definite
«vermi solitari» che infestano vari mammiferi e gli esseri
umani. I cestodi sono endoparassiti specializzati privi di
apparato digerente e possono raggiungere lunghezze
fino a 10-15 metri. Il loro corpo allungato e nastriforme
consta di 3 parti caratteristiche (▶figura 54):
Scolice,(erroneamente chiamato “testa”) dotato di
ventose e/o uncini per l'adesione alla parete intestinale
dell'ospite;
Collo, la zona di accrescimento dell'animale;
Strobila, il tratto più o meno lungo costituito da
proglottidi prodotte dal collo e spinte lontano da esso
tramite la formazione di nuove proglottidi. Quest'ultime
contengono l'apparato riproduttore e consentono la
riproduzione sessuale che, in questi parassiti, è
imponente per il numero di uova fecondate prodotte.
Figura 54. La tenia (verme solitario).
In alto: scolice di Tenia solium, definita tenia armata perché
dotata di uncini con le quali si attacca alla mucosa intestinale.
Essa ha come ospite intermedio i suini e come ospite definitivo
l’uomo. In basso: Tenia saginata definita tenia inerme perché
priva di uncini. Essa ha come ospite intermedio i bovini e come
ospite definitivo l’uomo.
I parassiti vivono in un ambiente ricco di sostanze
nutritive e non hanno perciò alcun bisogno di andare alla
ricerca di cibo. Essi devono comunque soddisfare altre
fondamentali esigenze: per completare il loro ciclo vitale i
parassiti devono vincere le difese dei loro ospiti e lasciarli
al momento opportuno per trasferirsi su altri. La maggior
parte delle specie parassite mostra pertanto un complesso
ciclo vitale, che prevede il passaggio in due o più ospiti
attraverso parecchie fasi larvali (▶figura 55).
Un ciclo vitale di tale complessità si è probabilmente
affermato perché aumenta le probabilità dei parassiti di
incontrare nuovi ospiti.
Figura 55. Il ciclo vitale di Diphyllobothrium latum.
Questo verme parassita dei pesci deve infestare un copepode
(un crostaceo) e un pesce prima di poter reinfestare l’ospite
primario, un mammifero.
Le principali specie di tenia che colpiscono l’uomo in
europa sono la Tenia solium e la Tenia saginata. L’uomo
è infestato mangiando carne cruda o mal cotta di
animali infestati, che contengono nei muscoli larve di
tenia (chiamate cisticerchi). Nell’intestino la larva si
sviluppa in una sorta di testa, detta scolice , munita di
ventose o uncini, con la quale si ancora alla parete
dell’intestino. Dallo scolice si producono poi un gran
numero di segmenti corporei, chiamati proglottidi, che
si allungano per anche più di un metro. La tenia è in
pratica priva di sistema nervoso e apparato digerente
(regressione parassitaria). Le proglottidi contengono i
due tipi di gonadi (ermafroditismo) da cui avviene
l’autofecondazione. Quando un segmento è maturo, si
stacca, e le uova fecondate sono espulse con le feci. Le
larve che ne derivano possono essere ingerite
dall’ospite intermedio: se ciò avviene esse raggiungono
i muscoli dove si incistano, ricominciando il ciclo. Un
essere umano infettato può eliminare tramite le feci
milioni di uova al giorno, sia allo stato "libero" sia
racchiuse nelle proglottidi. Queste uova possono
sopravvivere in natura per diversi mesi, durante i quali
possono infettare un nuovo ospite intermedio.

La comparsa dell’apertura anale e del sistema circolatorio: i Nemertini.
Il phylum dei Nemertini, o «vermi a nastro» è costituito
da organismi triblastici con corpo appiattito
dorsoventralmente e con sistema nervoso ed escretore
simili a quelli dei platelminti (▶figura 56).
A differenza di questi ultimi, tuttavia, i nemertini sono
provvisti di un apparato digerente completo, con
bocca e ano situati ai poli opposti. Dal momento che i
nemertini possiedono un apparato digerente
completo, con una bocca all'estremo anteriore e un
ano all'estremo opposto, il cibo transita in una precisa
direzione e durante il percorso viene attaccato da vari
enzimi digestivi; un sistema assai più efficiente di
quello che prevede l'eliminazione delle scorie
attraverso la stessa via di accesso.
Figura 56. Per i nemertini la proboscide rappresenta l’organo
per la cattura del cibo.
La proboscide può essere mossa rapidamente, mentre l’animale
si muove soltanto lentamente. Si osservi che la proboscide e la
bocca si aprono separatamente all’esterno.
Il corpo è di regola compatto per quasi tutta la sua
lunghezza, ma in alcune specie esiste una cavità
ripiena di liquido, detta rincocele, nella quale si trova
una struttura a forma di proboscide, muscolare e cava.
Le contrazioni dei muscoli che circondano il rincocele
provocano l'estroflessione improvvisa della proboscide
attraverso l'apertura anteriore del corpo. Il rincocele
trasmette dunque forze che muovono rapidamente la
proboscide senza che vengano mosse le altre parti del
corpo. Alcuni Nemertini di piccole dimensioni possono
muoversi mediante il battito di ciglia. Quelli di
dimensioni maggiori utilizzano invece le onde di
contrazione della muscolatura per muoversi alla
ricerca di cibo o per nascondersi. I movimenti
risultano comunque lenti. La proboscide della maggior
parte dei nemertini è armata da uno stiletto per
trafiggere le prede. Tossine a effetto paralizzante
vengono inoltre prodotte dalla proboscide e immesse
nella ferita provocata alla preda dallo stiletto. Alcune
specie sono prive di stiletto e catturano le prede
avvolgendole e bloccandole con la proboscide.
Quest'ultima viene poi retratta nel rincocele a opera di
muscoli retrattori e la preda viene avviata alla bocca.
Oltre che per la cattura delle prede, il rincocele serve
ai Nemertini per il movimento: la proboscide può
essere infissa nel substrato e la sua successiva
contrazione spinge in avanti l'animale. Un meccanismo
di questo tipo si ritrova anche in altri phyla.
L’altra grande novità evolutiva è rappresentata
dall’apparato circolatorio. Esso è privo di cuore ed è
semplicemente costituito da vasi sanguigni principali e
vasi secondari che trasportano nutrimento, ossigeno,
scorie metaboliche da una parte all’altra del corpo.
Quando i Nemertini vengono disturbati, possono
andare incontro ad autotomia (autoframmentazione
del corpo) seguita da rigenerazione. La riproduzione
può essere asessuata o sessuata ed essi,
contrariamente ai platelminti, sono in genere a sessi
separati. Delle 900 specie di Nemertini, la maggior
parte sono marine; solo poche vivono in acqua dolce o
nel substrato umido delle foreste tropicali. Tutti i
Nemertini sono carnivori, e si nutrono principalmente
di artropodi e piccoli vermi (▶figura 57).
Figura 57. I Nemertini possono essere anche intensamente
colorati.
Alcuni Nemertini sono di colore tenue come quello che è
raffigurato sopra, altri presentano intense colorazioni come la
specie marina che è raffigurata sotto.
Le parole:
Nemertini, deriva dal greco nema, nematos, «filo».

I Nematodi possiedono una cuticola di rivestimento che mutano periodicamente. II vantaggio di movimenti più efficienti, resi possibili dalla
presenza di cavità piene di liquido, ha favorito lo sviluppo
di tali cavità per tutta l'estensione del corpo. Un gruppo
che testimonia i risultati di questo cambiamento è
rappresentato dai vermi tondi (phylum Nematodi).
Gli organi interni dei Nematodi sono contenuti nello
pseudocele e il corpo è provvisto di una spessa cuticola
prodotta dal piano epidermico (▶figura 58).
Figura 58. I Nematodi.
(A) Questo disegno mostra la struttura del corpo di Trichinella
spiralis, un nematode parassita che causa la trichinosi. (B) Una
cisti di Trichinella spiralis nel tessuto muscolare di un ospite.
(C) Questo nematode invece conduce vita libera, muovendosi
attraverso i sedimenti marini.
Quando un Nematode cresce, esso abbandona la vecchia
cuticola e successivamente ne forma una nuova: questo
processo è definito muta ed è comune in molti gruppi di
animali.
Dorsalmente, ventralmente e lateralmente, l’epidermide
si ingrossa a formare 4 creste longitudinali, dette
cordoni. Sotto all’epidermide e tra le creste epidermiche
si trovano 4 bande di muscoli longitudinali formati da
cellule muscolari allungate che decorrono parallelamente
all’asse del verme. Mancano completamente i muscoli
circolari e quindi la locomozione avviene grazie al gioco
di forze opposte esercitate dai muscoli longitudinali sulla
cuticola e questo provoca dei tipici movimenti ondulatori
serpentiformi.
Il tubo digerente è diritto e possiede un’apertura
all’estremità anteriore, la bocca e una, vicino all’estremità
posteriore, l’ano. Dopo la bocca è presente una faringe
muscolare e in seguito l’intestino. Poiché la sezione
trasversale del corpo dei nematodi è rotonda, essi
possiedono una superficie corporea relativamente limitata
per gli scambi con l'ambiente. Gli scambi avvengono
soprattutto attraverso la cuticola, ma si realizzano anche a
livello dell'intestino, che è formato da un solo strato di
cellule. Sebbene la pressione idrostatica all'interno dello
pseudocele sia piuttosto elevata, il cibo riesce a transitare
attraverso l'intestino grazie alle contrazioni ritmiche della
faringe muscolosa situata all'estremità anteriore del tratto
digerente.
Il tratto digerente sembra avere anche un ruolo
nell’escrezione delle scorie azotate mentre la principale
funzione del sistema escretore è molto probabilmente in
prevalenza osmoregolatrice.
Mancano completamente sia strutture adibite alla
respirazione, che alla circolazione dei liquidi corporei.
Il sistema nervoso consiste in un anello nervoso e di
gangli posto attorno alla faringe e di 4 cordoni nervosi
principali che decorrono all’interno delle creste
epidermiche. Il cordone nervoso dorsale e ventrale hanno
una funzione prevalentemente motore, quelli laterali,
sensoriale.
Sono presenti organi sensoriali concentrati soprattutto
alle estremità anteriore e posteriore del corpo.
5. La comparsa dello pseudoceloma: Nematodi e Rotiferi.
Come è già stato descritto, fin dalle prime fasi di sviluppo, nella blastula di molti gruppi animali si forma un blastocele. In alcune
specie il blastocele persiste poi anche nell'adulto sotto forma di pseudocele, una cavità piena di liquido nella quale sono
contenuti gli organi. Lo pseudocele può inoltre funzionare come scheletro idrostatico, ma in alcuni pseudocelomati esso è
parzialmente occupato da masse cellulari. Fra i molti phyla di organismi pseudocelomati, verranno qui descritti i due più ricchi di
specie e cioè i Nematodi e i Rotiferi.

L’apparato riproduttore si trova nella cavità del corpo.
Generalmente i sessi sono separati e la fecondazione è
interna. In alcune specie è stato osservato
ermafroditismo mentre in altre partenogenesi (sviluppo
a partire da cellule uovo non fecondate).
I vermi tondi sono uno dei gruppi animali più ricchi di
specie e universalmente diffusi. Noi stessi ingeriamo
inconsapevolmente, con i cibi e con le bevande, grandi
quantità di Nematodi. Una mela marcia può contenere fino
a 90000 Nematodi, mentre in un metro quadrato di
sedimento fangoso se ne contano circa 4420000 (▶figura
58). Molte specie di vermi tondi vivono come detritivori nel
suolo, sul fondo di fiumi e laghi, o come parassiti nel
corpo di organismi vegetali o animali. Attualmente si
conoscono circa 20000 specie di Nematodi, ma è probabile
che ne esistano più di un milione.
Un Nematode del suolo, Caenorhabitis elegans, è un
«organismo modello» ampiamente studiato dai biologi
che si occupano di genetica e di sviluppo (▶figura 59).
Questo organismo si presta particolarmente bene a tali
ricerche poiché è facile da allevare, raggiunge la
maturità in soli tre giorni e possiede un numero fisso di
cellule corporee (tra l’altro, il genoma di C. elegans è
stato completamente sequenziato).
Figura 59. Un verme modello.
Caenorhabditis elegans è un nematode lungo circa 1 mm, che
vive nel suolo. Si tratta di un organismo modello molto usato in
biologia, soprattutto in genetica e biologia dello sviluppo (le
strutture tondeggianti sono uova).
La dieta dei nematodi è varia quanto i loro possibili
habitat. Molte specie vivono infatti come parassiti, altri
sono predatori di protisti o di altri piccoli invertebrati
(compresi altri vermi tondi). Molti studi su questo phylum
sono stati motivati proprio dalla necessità di comprendere
la biologia delle specie parassite al fine di poterne
controllare la trasmissione. I vermi tondi sono infatti
parassiti dell'uomo, di molti animali domestici e anche di
specie vegetali economicamente importanti. Il nematode di
maggiori dimensioni, che misura circa nove metri, vive
nella placenta dei capodogli. Tra le tante specie di
Nematodi, solo la minoranza è rappresentata da parassiti.
L’uomo è ospite di circa 50 specie di Nematodi e una
dozzina di queste causa seri problemi sanitari dato che
interessa milioni, a volte centinaia di milioni di persone.
La struttura dei Nematodi parassiti non differisce molto da
quella delle specie che conducono vita libera, ma
sicuramente il ciclo vitale dei primi è modificato per
consentire il passaggio degli individui da un ospite
all'altro. Un metodo relativamente semplice per effettuare
tale passaggio viene attuato da Trichinella spiralis, l'agente
responsabile di una grave malattia nota come trichinosi.
Le forme larvali di Trichinella formano cisti calcificate nello
spessore dei muscoli dei mammiferi loro ospiti (▶figura
58) e queste, se presenti in gran numero, possono causare
gravi danni o addirittura la morte. La Trichinella viene
trasmessa a un nuovo ospite quando un mammifero
mangia le carni di un individuo infetto. Le larve attraversano
infatti la parete intestinale del nuovo ospite e tramite il
circolo sanguigno giungono ai muscoli dove si localizzano
e formano nuove cisti. L'infezione si trasmette dunque da
mammifero a mammifero, e spesso sono le carni di maiale
che trasmettono l'infezione all'uomo. Nel caso di altri
Nematodi, invece, il ciclo biologico del parassita è assai più
complesso e prevede il passaggio in diversi ospiti
appartenenti a classi diverse.
Alcuni importanti parassiti nell’uomo sono gli strongili, gli
anchilostomi, gli ossiuri, gli ascaridi e infine le filarie,
parassiti con un artropode come ospite intermedio e che
causano la filariosi, una parassitosi che colpisce l’uomo e
diversi animali domestici. A volte essa può provocare
l’elefantiasi dovuta al blocco del sistema linfatico e alla
crescita incontrollata di tessuti fibrosi che portano a
rigonfiamenti grotteschi degli arti affetti (▶figura 60).
Figura 60 Elefantiasi, causata da Wuchereria bancrofti.
Se i parassiti si accumulano nei vasi sanguigni, possono limitare
la circolazione e causare accumulo di fluido nei tessuti
circostanti. I più comuni, segni visibili di infezione sono braccia,
gambe, seni e genitali spropositatamente enormi.
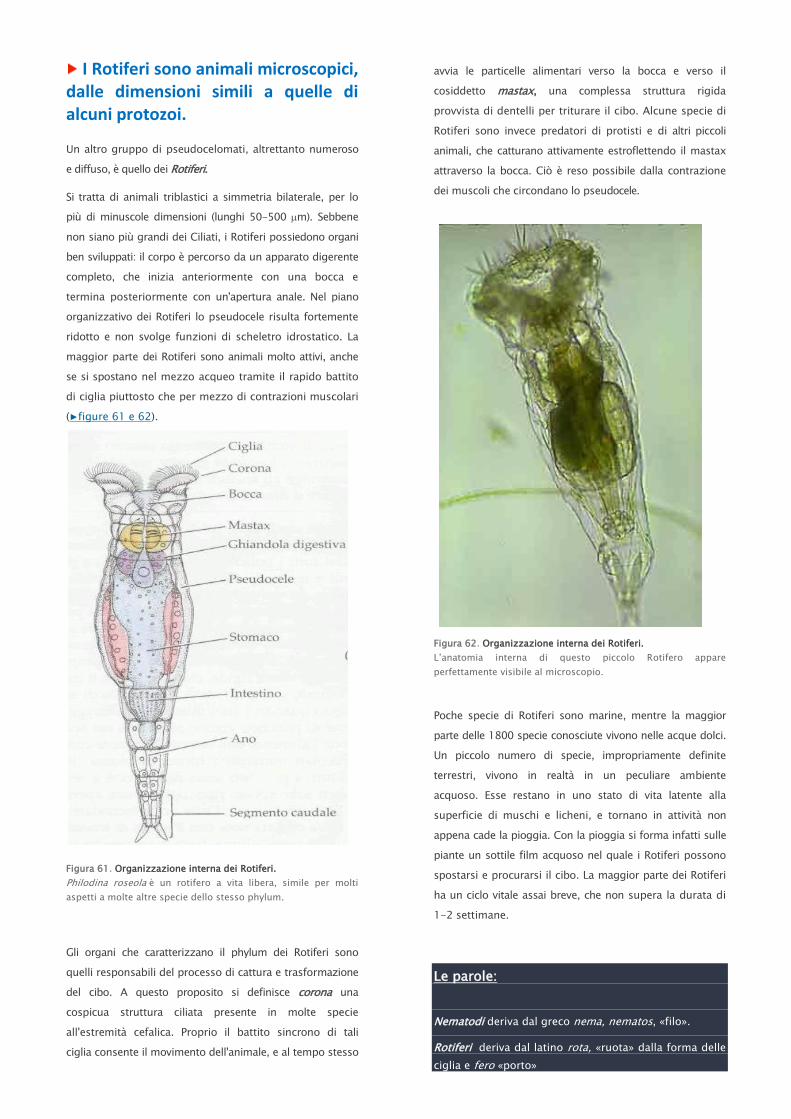
I Rotiferi sono animali microscopici, dalle dimensioni simili a quelle di alcuni protozoi.
Un altro gruppo di pseudocelomati, altrettanto numeroso
e diffuso, è quello dei Rotiferi.
Si tratta di animali triblastici a simmetria bilaterale, per lo
più di minuscole dimensioni (lunghi 50-500 m). Sebbene
non siano più grandi dei Ciliati, i Rotiferi possiedono organi
ben sviluppati: il corpo è percorso da un apparato digerente
completo, che inizia anteriormente con una bocca e
termina posteriormente con un'apertura anale. Nel piano
organizzativo dei Rotiferi lo pseudocele risulta fortemente
ridotto e non svolge funzioni di scheletro idrostatico. La
maggior parte dei Rotiferi sono animali molto attivi, anche
se si spostano nel mezzo acqueo tramite il rapido battito
di ciglia piuttosto che per mezzo di contrazioni muscolari
(▶figure 61 e 62).
Figura 61. Organizzazione interna dei Rotiferi.
Philodina roseola è un rotifero a vita libera, simile per molti
aspetti a molte altre specie dello stesso phylum.
Gli organi che caratterizzano il phylum dei Rotiferi sono
quelli responsabili del processo di cattura e trasformazione
del cibo. A questo proposito si definisce corona una
cospicua struttura ciliata presente in molte specie
all'estremità cefalica. Proprio il battito sincrono di tali
ciglia consente il movimento dell'animale, e al tempo stesso
avvia le particelle alimentari verso la bocca e verso il
cosiddetto mastax, una complessa struttura rigida
provvista di dentelli per triturare il cibo. Alcune specie di
Rotiferi sono invece predatori di protisti e di altri piccoli
animali, che catturano attivamente estroflettendo il mastax
attraverso la bocca. Ciò è reso possibile dalla contrazione
dei muscoli che circondano lo pseudocele.
Figura 62. Organizzazione interna dei Rotiferi.
L’anatomia interna di questo piccolo Rotifero appare
perfettamente visibile al microscopio.
Poche specie di Rotiferi sono marine, mentre la maggior
parte delle 1800 specie conosciute vivono nelle acque dolci.
Un piccolo numero di specie, impropriamente definite
terrestri, vivono in realtà in un peculiare ambiente
acquoso. Esse restano in uno stato di vita latente alla
superficie di muschi e licheni, e tornano in attività non
appena cade la pioggia. Con la pioggia si forma infatti sulle
piante un sottile film acquoso nel quale i Rotiferi possono
spostarsi e procurarsi il cibo. La maggior parte dei Rotiferi
ha un ciclo vitale assai breve, che non supera la durata di
1-2 settimane.
Le parole:
Nematodi deriva dal greco nema, nematos, «filo».
Rotiferi deriva dal latino rota, «ruota» dalla forma delle
ciglia e fero «porto»