
Insegnamento: BIOCHIMICA E BIOLOGIA Modulo: CHIMICA E ...
64
Alessandro Saba Lezione 9: Biochimica Insegnamento: BIOCHIMICA E BIOLOGIA Modulo: CHIMICA E BIOCHIMICA
Transcript of Insegnamento: BIOCHIMICA E BIOLOGIA Modulo: CHIMICA E ...

Alessandro Saba
Lezione 9: Biochimica
Insegnamento:BIOCHIMICA E BIOLOGIA
Modulo:CHIMICA E BIOCHIMICA

Biochimica

Amminoacidi e
Proteine

Amminoacidi e Proteine
Le proteine rappresentano gli elementi strutturali efunzionali più importanti dei sistemi viventi. Svolgonoun’ampia varietà di funzioni. Sono sintetizzate comesequenza di amminoacidi e hanno struttura polimerica.
In natura esistono circa 300 amminoacidi, ma solo 20 diessi sono codificati dal DNA e sono incorporati nelleproteine. Questi 20 sono degli α-L-amminoacidi.

α-Amminoacidi
Ogni aminoacido possiede:
un carbonio centrale, detto carbonio α (in α rispetto algruppo –COOH) al quale sono legati 4 gruppi:
1. gruppo amminico basico-NH2 (o anche -NH3
+),
2. gruppo carbossilico acido-COOH (o anche -COO-),
3. atomo di H,
4. catena laterale –R diversaper ciascun amminoacido.
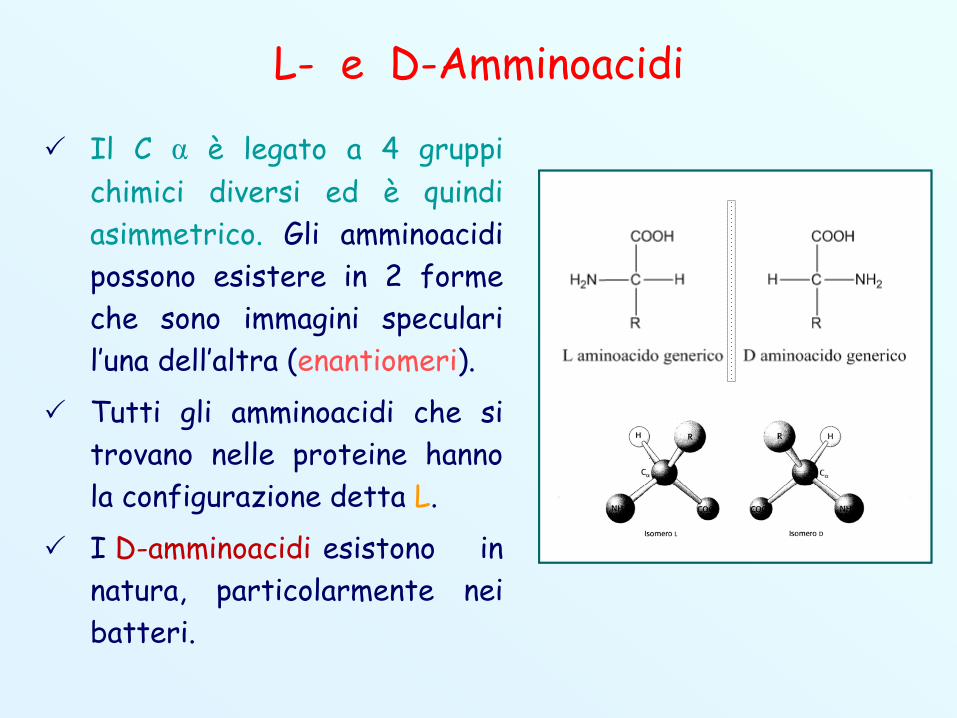
L- e D-Amminoacidi
Il C α è legato a 4 gruppichimici diversi ed è quindiasimmetrico. Gli amminoacidipossono esistere in 2 formeche sono immagini specularil’una dell’altra (enantiomeri).
Tutti gli amminoacidi che sitrovano nelle proteine hannola configurazione detta L.
I D-amminoacidi esistono innatura, particolarmente neibatteri.
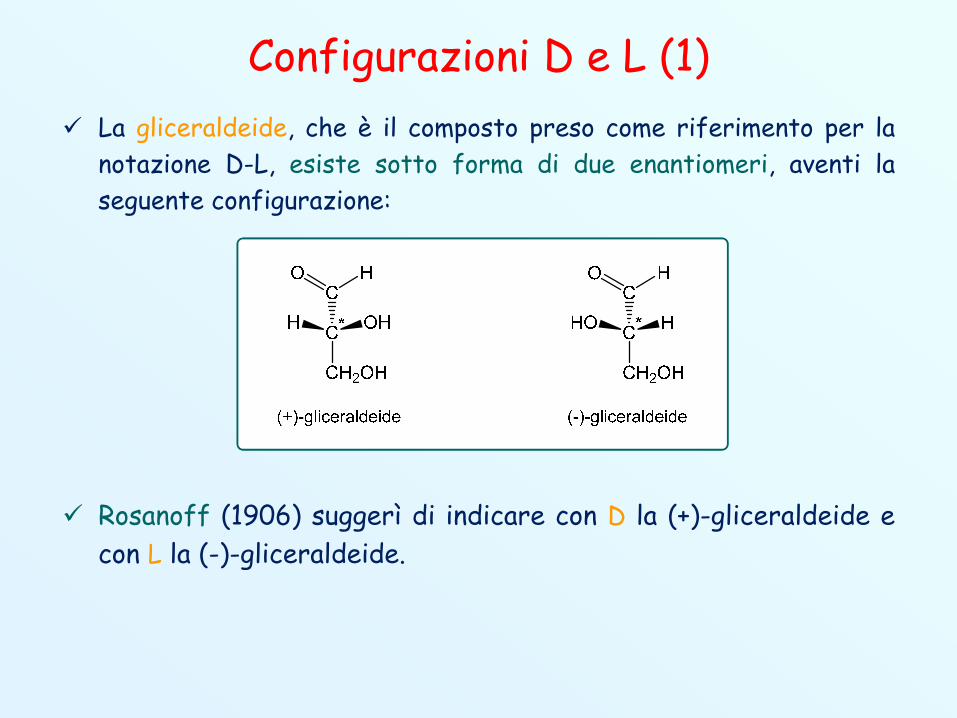
Configurazioni D e L (1) La gliceraldeide, che è il composto preso come riferimento per la
notazione D-L, esiste sotto forma di due enantiomeri, aventi laseguente configurazione:
Rosanoff (1906) suggerì di indicare con D la (+)-gliceraldeide econ L la (-)-gliceraldeide.
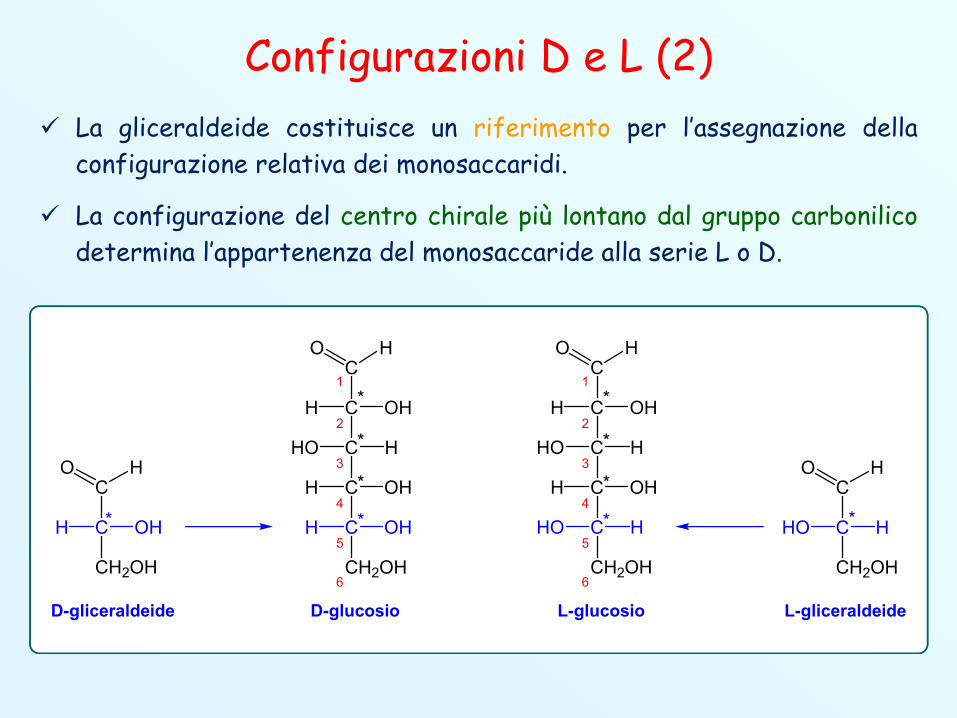
Configurazioni D e L (2) La gliceraldeide costituisce un riferimento per l’assegnazione della
configurazione relativa dei monosaccaridi.
La configurazione del centro chirale più lontano dal gruppo carbonilicodetermina l’appartenenza del monosaccaride alla serie L o D.
C
C
CH2OH
HOC
C
CH2OH
HO
D-gliceraldeide L-gliceraldeide
C
CH2OH
D-glucosio L-glucosio
C
C
C
*
*
*
* *
CO H
*
OH
OH
H
OHH
HO
H
H C
CH2OH
C
C
C
*
*
*
CO H
*
H
OH
H
OHH
HO
H
HOOHH HHO
1
2
3
4
5
6
1
2
3
4
5
6
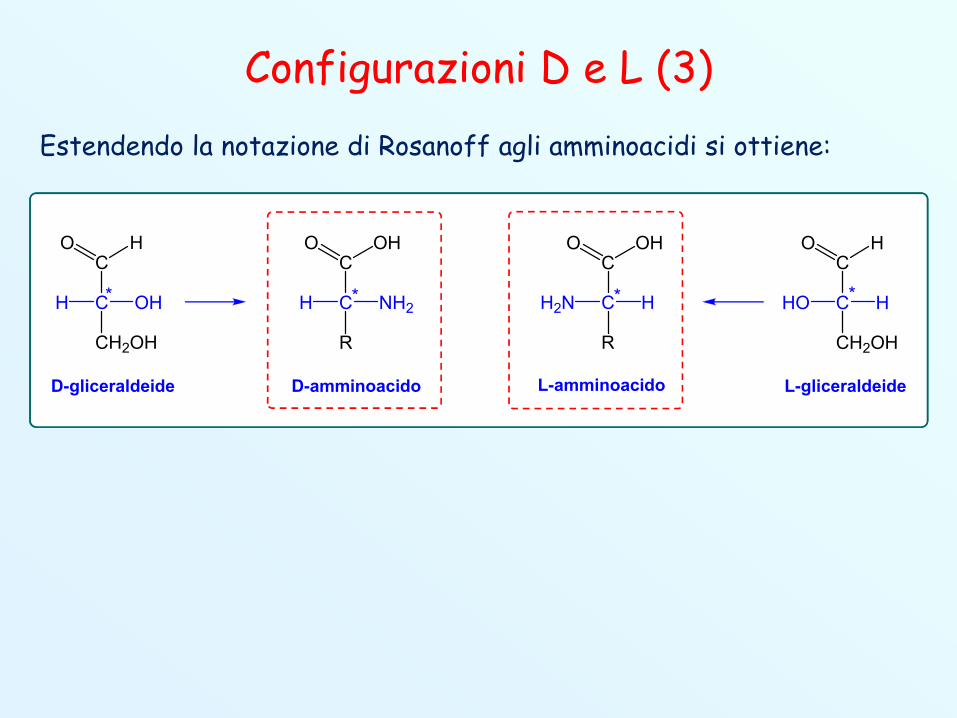
Configurazioni D e L (3)Estendendo la notazione di Rosanoff agli amminoacidi si ottiene:
C
C
CH2OH
HOC
C
CH2OH
HO
D-gliceraldeide L-gliceraldeide
C
R
D-amminoacido L-amminoacido
** *NH2H C
R
* HH2NOHH HHO
C COHO O OH

Gli Amminoacidi sono Zwitterioni Gli amminoacidi possiedono un gruppo acido (-COOH) e uno
basico (-NH2), entrambi deboli. Sono quindi molecole ionizzabili,con grado di ionizzazione dipendente dal pH.
Attorno a pH 7 i gruppi acido e basico sono entrambi in formaionica (zwitterione). La concentrazione dello zwitterione havalore massimo al punto isoelettrico (pI).
Il punto isoelettrico è il valore di pH a cui la carica netta è paria 0 (neutra). Tale valore è dato da: (pK1 + pK2)/2.
Es.: GlicinapI = 5.97
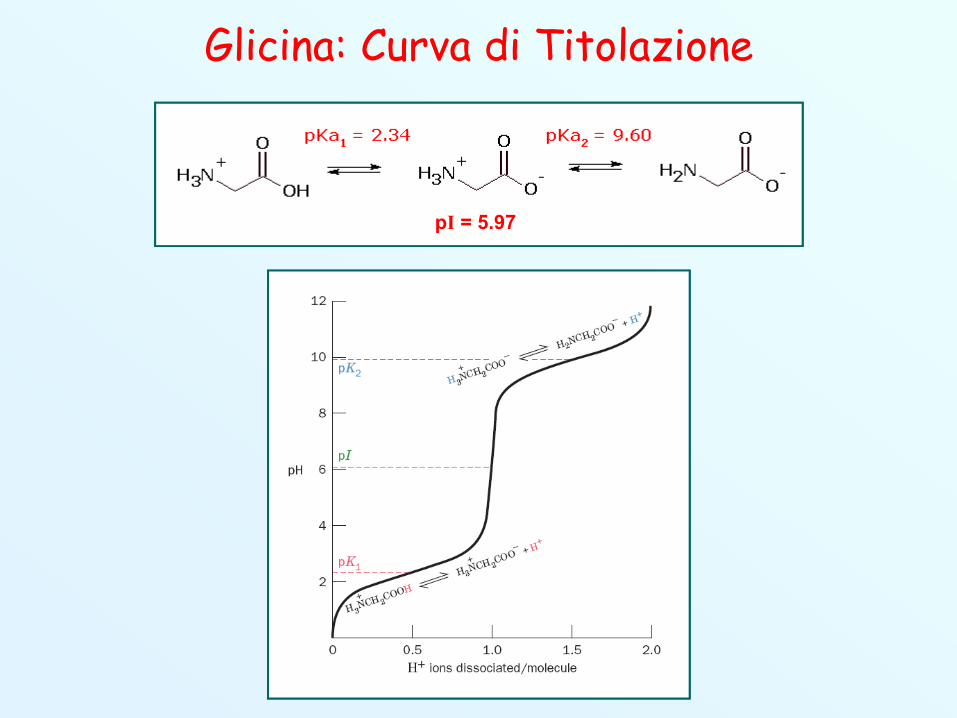
Glicina: Curva di Titolazione
pI = 5.97

Classificazione degli Amminoacidi
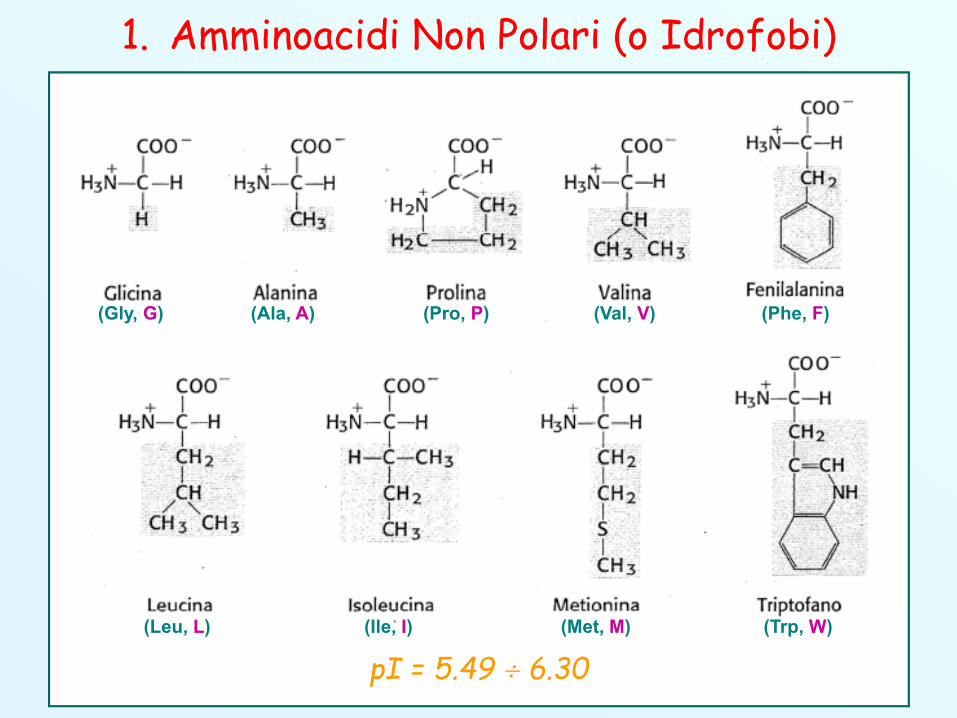
A seconda della natura della loro catena laterale –R, gliamminoacidi possono essere classificati in 4 gruppi:
1. Amminoacidi non polari (idrofobi)
2. Amminoacidi polari non carichi (idrofili)
3. Amminoacidi polari acidi
4. Amminoacidi polari basici
1. Amminoacidi Non Polari (o Idrofobi)
(Ala, A) (Phe, F)(Gly, G)
(Ile, I)(Leu, L) (Met, M)
(Pro, P)
(Trp, W)
(Val, V)
pI = 5.49 6.30
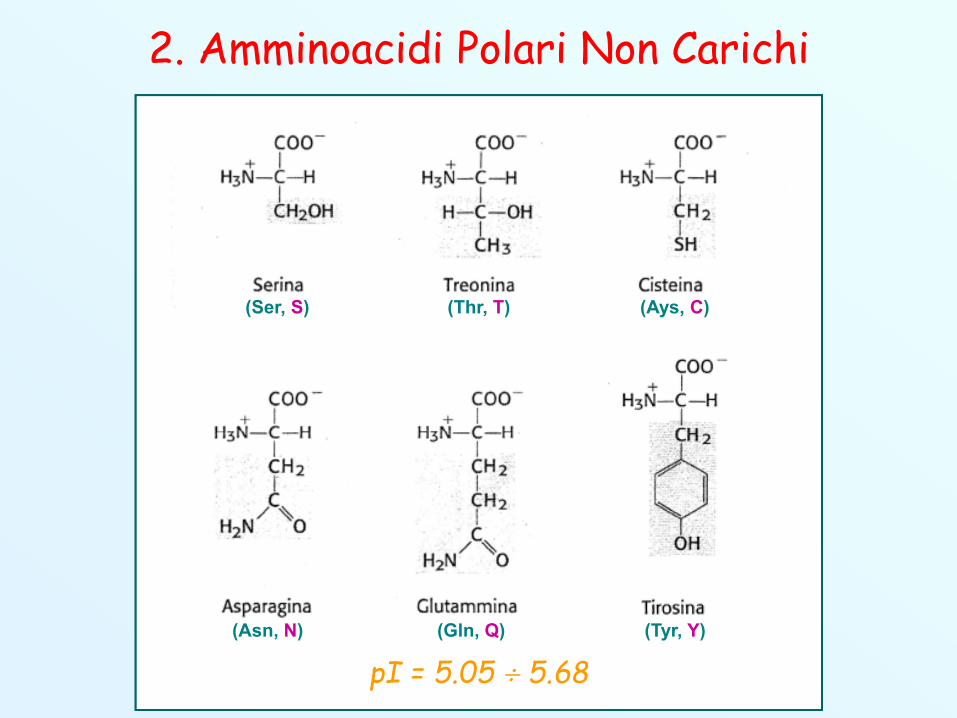
2. Amminoacidi Polari Non Carichi
(Asn, N)
(Ays, C)
(Gln, Q)
(Ser, S)
(Tyr, Y)
(Thr, T)
pI = 5.05 5.68

3. Amminoacidi Polari Acidi
(Asp, D)pI=2.85
(Glu, E)pI=3.15
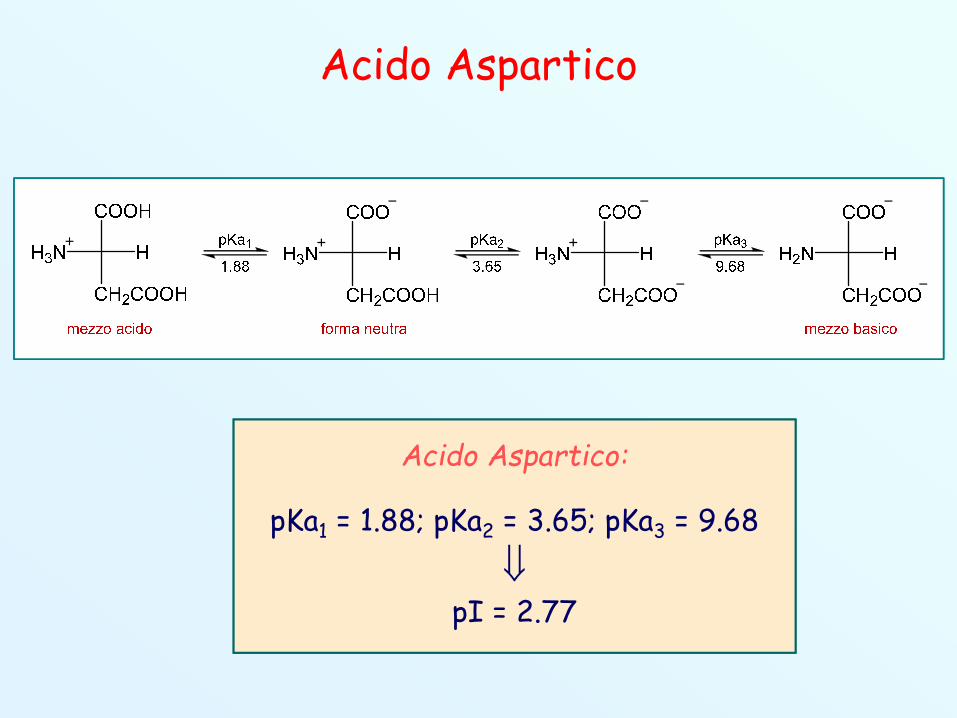
Acido Aspartico
Acido Aspartico:
pKa1 = 1.88; pKa2 = 3.65; pKa3 = 9.68
pI = 2.77

4. Amminoacidi Polari Basici
(Arg, R)pI=10.76
(His, H)pI=7.60
(Lys, K)pI=9.60
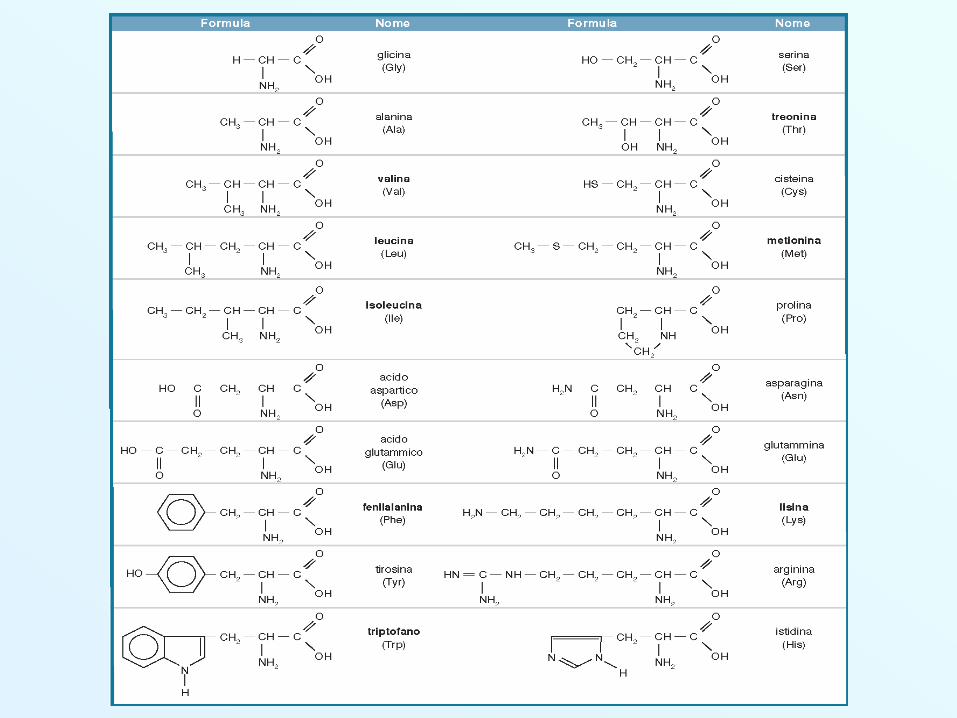

Amminoacidi Essenziali e Non Gli amminoacidi essenziali sono amminoacidi che l’organismo non può
sintetizzare. Devono quindi essere somministarti con la dieta.
Sono 9: Istidina (His) Isoleucina (Ile) Leucina (Leu)Lisina (Lys) Metionina (Met) Fenilalanina (Phe)Treonina (Thr) Triptofano (Trp) Valina (Val)
Altri amminoacidi possono essere essenziali in alcune fasi della vita,quali l’infanzia e l’adolescenza e/o talvolta durante la malattia.
Sono 6: Arginina (Arg) Cisteina (Cys) Glutammina (Gln)Glicina (Gly) Prolina (Pro) Tirosina (Tyr)
Gli amminoacidi NON essenziali
Sono 5: Alanina (Ala) Asparagina (Asp) Aspartato (Asp)Serina (Ser) Glutammato (Glu)

Il Legame Peptidico
Le proteine si formano per unione di molecole diamminoacidi, che si legano covalentemente mediantelegame peptidico.
Il legame peptidico si realizza mediante condensazionedel gruppo carbossilico -COOH del primo amminoacidocon il gruppo amminico -NH2 del secondo amminoacido,con eliminazione di una molecola d’ acqua.

Il Legame Peptidico
PEPTIDI: gli oligopeptidi, o semplicemente peptidi, sonocostituiti da meno di 20 amminoacidi.Es: alcuni ormoni/neurotrasmettitori come la vasopressina,l’ossitocina, il TRH (ormone di rilascio della tireotropina).
POLIPEPTIDI: costituiti da 20-49 di amminoacidi.Es: alcuni ormoni/ neurotrasmettitori come la calcitonina eil VIP (peptide intestinale vasoattivo).
PROTEINE: costituite da 50 o più amminoacidi.Es: alcuni ormoni/ neurotrasmettitori come l’ insulina (laproteina più piccola, con 50 amminoacidi), il GH (ormonedella crescita), la prolattina, la tireotropina.

Caratteristiche del Legame Peptidico
Ha il carattere di un doppio legame parziale (è più corto diun legame singolo).
E’ rigido e planare (non è possibile la rotazione attorno allegame tra il carbonio carbonilico e l’azoto del legamepeptidico).
In genere è un legame con configurazione trans, a causa diinterferenze steriche tra i gruppi -R (i legami tra un Cα eun gruppo α-amminico o α-carbossilico possono ruotare!)
I gruppi -C=O ed -NH del legame peptidico non hanno unacarica elettrica (a differenza del gruppo α-amminicoall’estremità N-terminale ed α-carbossilico al C-terminale)ma sono polari e partecipano alla formazione di legami aidrogeno.

Funzioni Principali delle Proteine (1)
Legame: il riconoscimento specifico di altre molecole(ligandi) è centrale per la funzione delle proteine. Leproteine sono in grado di riconoscere e legare molecolepiccole come la molecola dell’ossigeno o grandi come unsegmento specifico di DNA. Il legame specifico è regolatodalla complementarità della forma e da interazioni polaricome il legame idrogeno.
Catalisi: negli organismi viventi ogni reazioni chimica habisogno di un catalizzatore la maggior parte dei quali ècostituita da catalizzatori biologici. Le reazioni possonoessere accelerate fino a 17 ordini di grandezza.

Funzioni Principali delle Proteine (2)
Interruttore: le proteine possono variare la loroconformazione in base al pH o al legame con ligandi. Talicambiamenti possono essere utilizzati come interruttorimolecolari per controllare processi cellulari.
Proteine strutturali: la funzione strutturale dipendedalla loro capacità di associarsi con se stesse o con altreproteine o molecole per formare sistemi complessi eresistenti.Es.: cheratina, che costituisce la struttura dei capelli,
delle unghie e dello strato corneo della cute;collagene e l'elastina, che appartengono al tessutoconnettivo sottocutaneo (cartilagine, osso, fasce,tendini, legamenti, …).

Funzioni Principali delle Proteine (3)
Riserva: alcune proteine vegetali e animali svolgonofunzione di riserva di nutrienti essenziali (caseina…).
Risposta allo stress: molte proteine del nostroorganismo sono in grado di convertire contaminantiorganici tossici in derivati meno tossici o di ripararedanni provocati dagli stessi contaminanti (es.: heatshock proteins*...).
* proteine indotte da ‘’shock termico’’ o altri stress, comeinfezioni infiammazioni, esposizioni a tossine. Agiscono da‘’chaperones’’ molecolari, accoppiandosi a proteine denaturateper ripristinarne il ripiegamento tridimensionale e quindi lecondizioni funzionali.

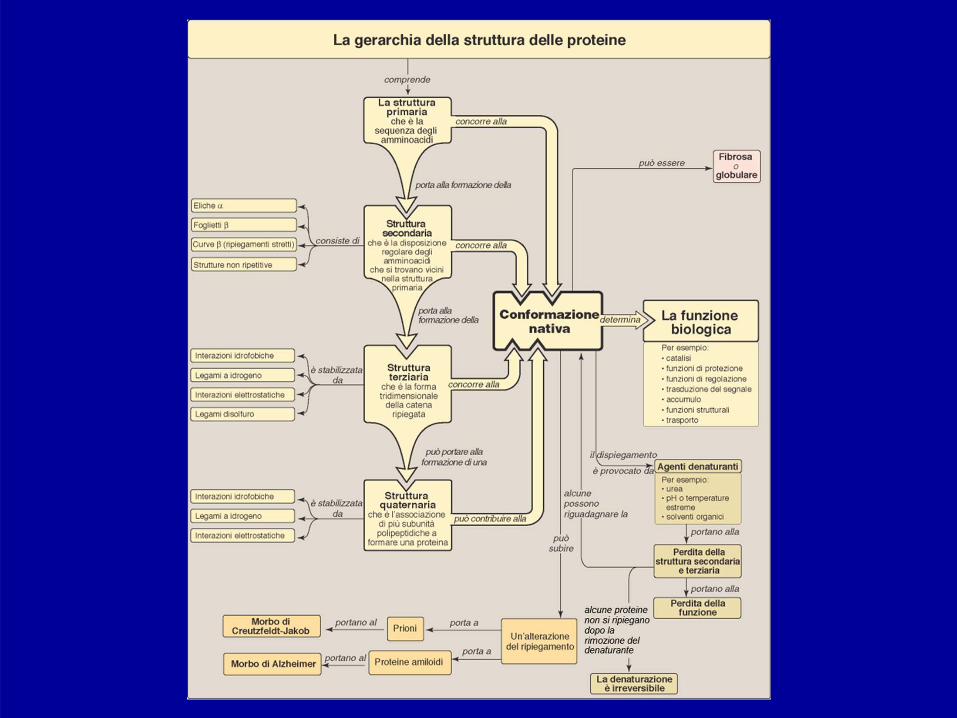
Struttura delle Proteine (1)
Struttura primaria: è la sequenza degli amminoacidi checostituiscono la catena polipeptidica.
Struttura secondaria: è la capacità di un polipeptide diassumere una struttura spaziale regolare e ripetitiva.
Struttura terziaria: descrive gli aspetti tridimensionalidell’intero polipeptide.
Struttura quaternaria: quando una proteina è costituitada un’associazione di più “sub-unità” polipeptidichetenute insieme da interazioni non covalenti,l’arrangiamento spaziale complessivo è detto strutturaquaternaria.
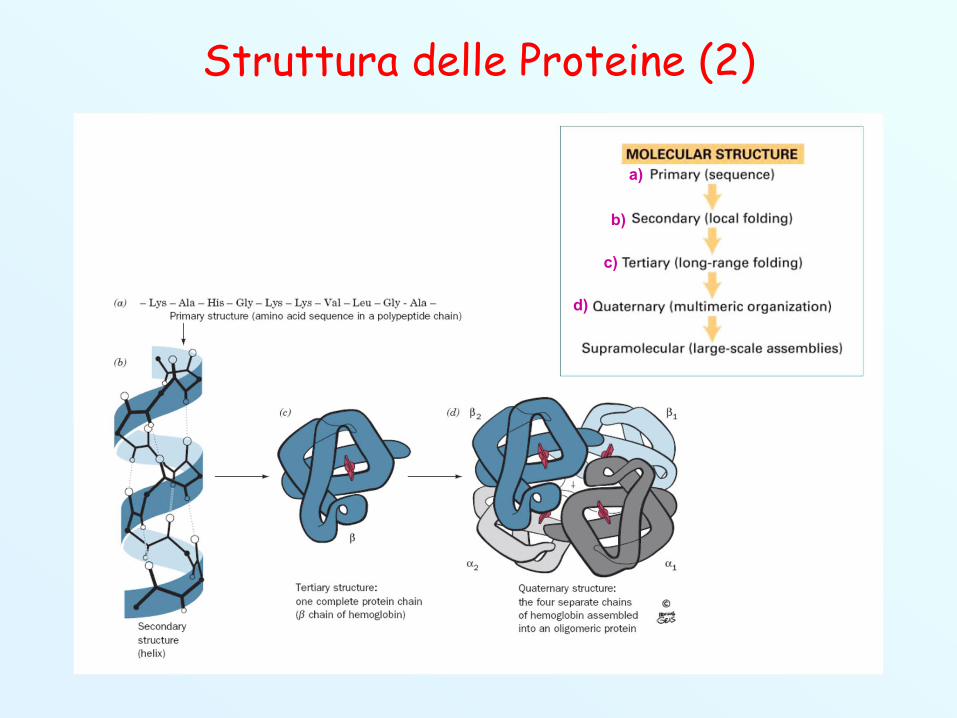
Struttura delle Proteine (2)
a)
b)
c)
d)

Struttura Primaria La peculiare sequenza amminoacidica di una catena
polipeptidica rappresenta la struttura primaria.
In alcune proteine la catena peptidica presenta legamitrasversali, il più comune dei quali è il ponte disolfuro, che siforma per ossidazione laterale di due residui di cisteina.
I ponti disolfuro possono formarsi tra residui di cisteinaall’interno della stessa catena polipeptidica, oppure possonounire due catene diverse.

Struttura Secondaria
La struttura secondaria di una proteina si riferisce aorganizzazioni regolari e ricorrenti nello spazio deiresidui aminoacidici adiacenti di una catenapolipeptidica.
Strutture secondarie più rappresentate: α-elica struttura a foglietto ripiegato
(β-struttura o β-foglietto)
Le strutture secondarie sono stabili in quantominimizzano le repulsioni steriche e allo stesso temporendono massima la potenzialità di formazione deilegami a idrogeno.
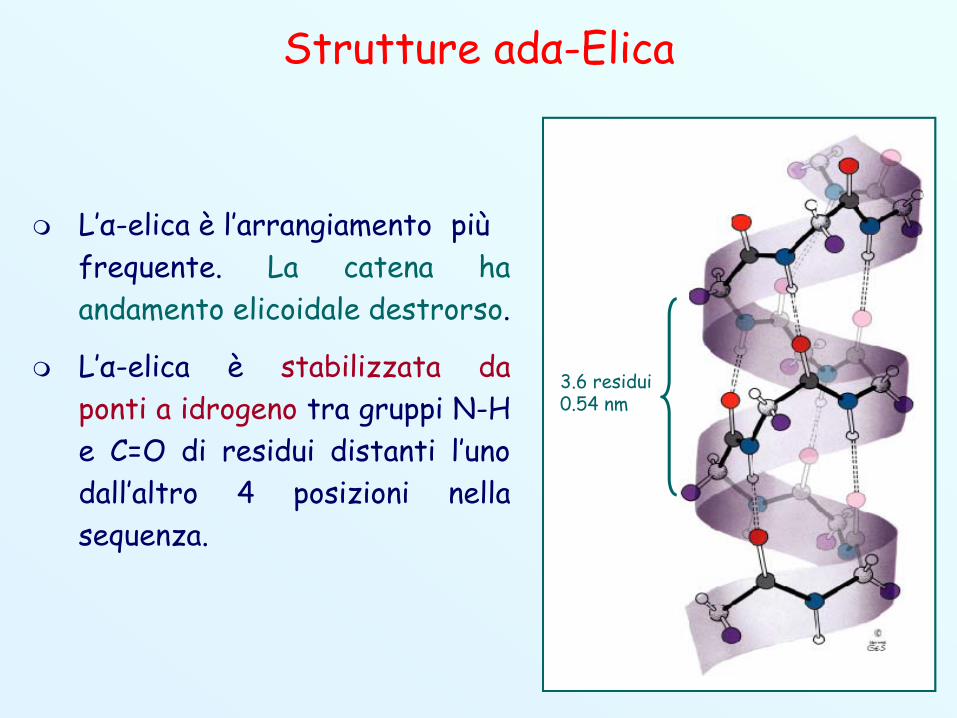
Strutture adα-Elica
L’α-elica è l’arrangiamento piùfrequente. La catena haandamento elicoidale destrorso.
L’α-elica è stabilizzata daponti a idrogeno tra gruppi N-He C=O di residui distanti l’unodall’altro 4 posizioni nellasequenza.
3.6 residui0.54 nm
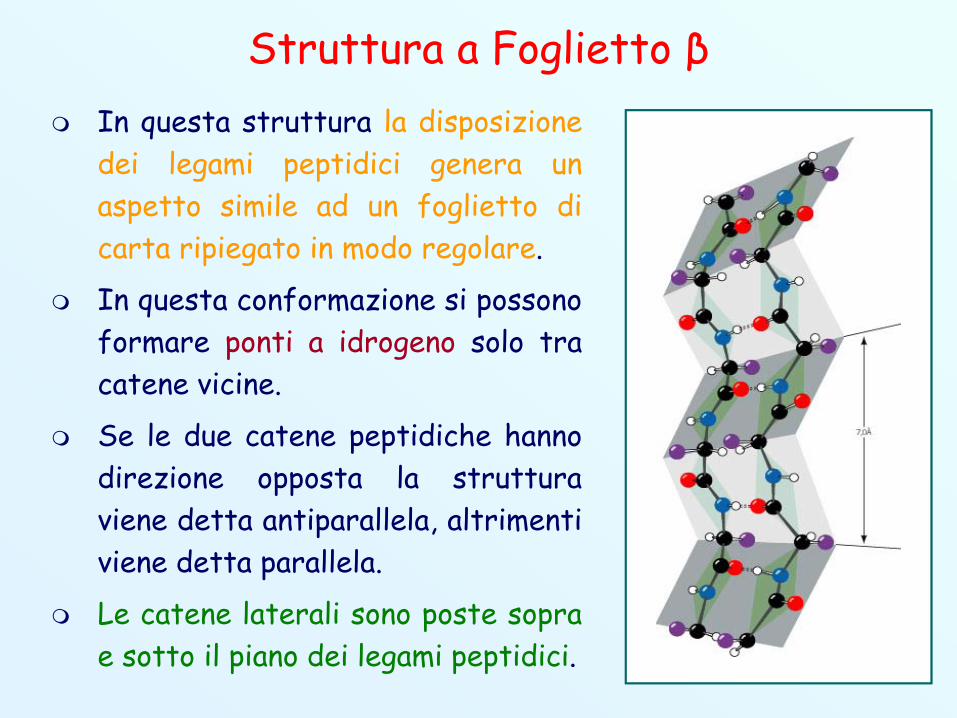
Struttura a Foglietto β In questa struttura la disposizione
dei legami peptidici genera unaspetto simile ad un foglietto dicarta ripiegato in modo regolare.
In questa conformazione si possonoformare ponti a idrogeno solo tracatene vicine.
Se le due catene peptidiche hannodirezione opposta la strutturaviene detta antiparallela, altrimentiviene detta parallela.
Le catene laterali sono poste soprae sotto il piano dei legami peptidici.

Struttura Terziaria (1) La struttura terziaria è la conformazione 3D, avvolta, di una
proteina. La struttura primaria di una catena polipeptidica determina la
sua struttura terziaria. Quando una proteina si avvolge su se stessa, gli amminoacidi
che si trovano in regioni lontane della sequenza polipeptidicapossono ugualmente interagire tra loro.

Struttura Terziaria (2) Anche la struttura terziaria è stabilizzata da legami non
covalenti come ponti idrogeno, interazioni idrofobiche (i gruppilipofili si dispongono all’interno della molecola) tra amminoacidinon polari e legami ionici.
Una ulteriore contributo alla stabilizzazione viene dato dalegami covalenti, sotto forma di ponti disolfuro fra duecisteine.
Ogni proteina possiede una struttura terziaria caratteristica,composta di elementi di struttura secondaria (α-eliche,foglietti β, regioni non regolari dette ripiegamenti o loop)ripiegati in modo specifico, per adattarsi al particolare ruolofunzionale che la proteina riveste.
La struttura terziaria ha un’influenza fondamentale sull’attivitàbiologica delle proteine.

Proteine Fibrose e Globulari
Proteine globulari- solubili in acqua- usate a livello cellulare- hanno un struttura 3D complessa
Proteine Fibrose- insolubili in acqua- utilizzate per tessuti connettivi- collagene, cheratina
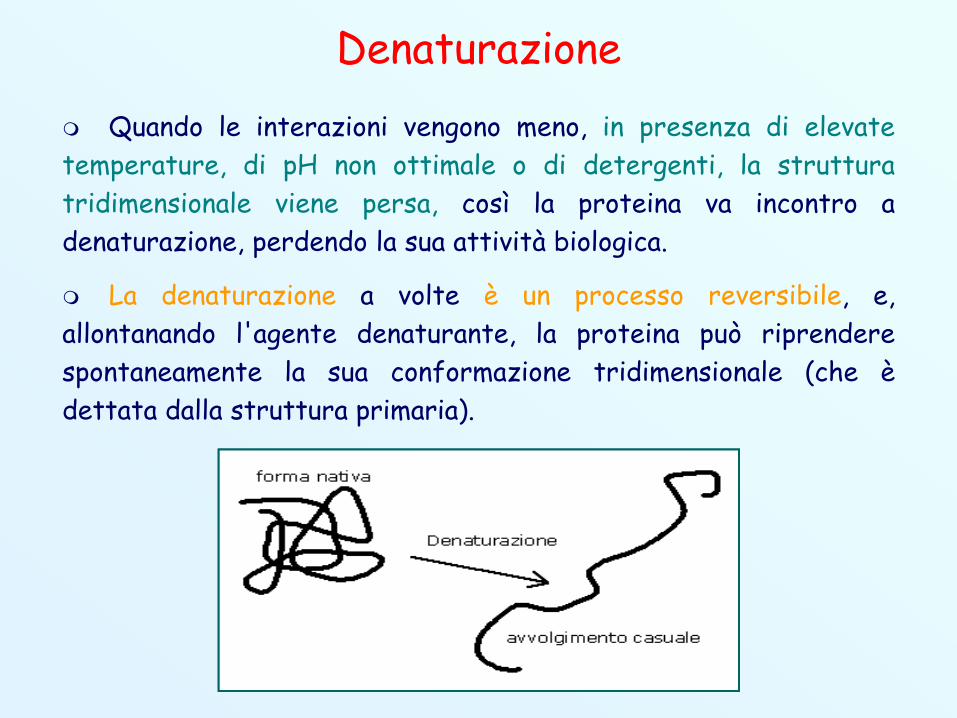
Denaturazione Quando le interazioni vengono meno, in presenza di elevatetemperature, di pH non ottimale o di detergenti, la strutturatridimensionale viene persa, così la proteina va incontro adenaturazione, perdendo la sua attività biologica.
La denaturazione a volte è un processo reversibile, e,allontanando l'agente denaturante, la proteina può riprenderespontaneamente la sua conformazione tridimensionale (che èdettata dalla struttura primaria).
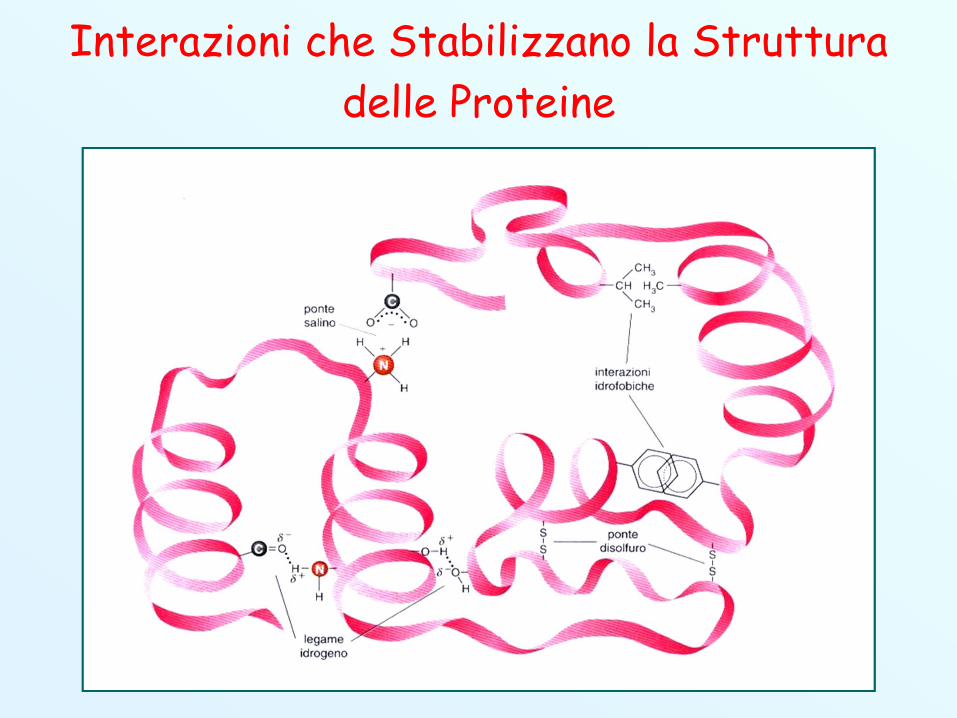
Interazioni che Stabilizzano la Struttura delle Proteine

Struttura Quaternaria
Molte proteine sono costituite da una sola catenapolipeptidica (proteine monomeriche).
Alcune proteine sono costituite da 2 o più catenepolipeptidiche (sub-unità) strutturalmente identiche odiverse (proteine multimeriche).
L’associazione delle sub-unità polipeptidiche costituiscela struttura quaternaria.
Le sub-unità sono tenute insieme da interazioni noncovalenti.
Questa organizzazione quaternaria può essere di due tipi:associazione tra sub-unità identiche o quasi (omotipica) odi struttura molto diversa (eterotipica).

Proteine con Gruppi Diversi dagli Amminoacidi
Molti enzimi contengono solo amminoacidi e nessun altro gruppochimico PROTEINE SEMPLICI.
Altre proteine contengono anche gruppi chimici funzionalipermanentemente associati PROTEINE CONIUGATE. Laparte non amminoacidica è il GRUPPO PROSTETICO.

Proteine Globulari:
Enzimi

Gli Enzimi
Ad eccezione di un piccolo gruppo di molecole, glienzimi sono proteine la cui attività catalitica dipendedall’integrità della loro conformazione nativa.
Praticamente tutte le reazioni biochimiche sonocatalizzate da enzimi. Nelle condizioni biologiche lereazioni non catalizzate sarebbero troppo lente.
Sono catalizzatori molto efficaci, che aumentano lavelocità delle reazioni di vari ordini di grandezza(107 – 1014).

Cofattori
Alcuni enzimi, per la loro attività si avvalgonounicamente delle catene laterali dei loro residuiamminoacidici. Altri invece hanno bisogno dicomponenti chimici addizionali chiamati COFATTORI.
Il cofattore può essere:- uno o più ioni inorganici (Fe2+, Mg2+, Mn2+, Zn2+) metalloproteasi.
- molecole organiche o metallorganiche complesselegate non covalentemente dette coenzimi.
- molecole organiche complesse o ioni metallicilegati covalentemente alla proteina enzimaticadette gruppi prostetici.

Un pò di terminologia…
Oloenzima: è l’enzima cataliticamente attivo con tuttii suoi cofattori.
Apoenzima o Apoproteina: è la parte proteica di unenzima.
Isoenzimi o Isozimi: sono enzimi diversi (differentestruttura chimica diverse proprietà chimicofisiche, quali il pH e il punto isoelettrico) checatalizzano la stessa reazione. Sono generalmentepresenti in tessuti differenti.

Classificazione degli enzimiGli enzimi vengono classificati in base al tipo di reazione checatalizzano, utilizzando una nomenclatura internazionale.
Ad ogni enzima viene associato un codice costituito dalle lettere "EC“(Enzyme Commission) seguite da 4 set di numeri separati da punti.
Esempio: EC.3.4.11.4
EC 3: enzimi della famiglia delle idrolasi (il primo numeroidentifica la reazione catalizzata dall'enzima).
EC 3.4: idrolasi che agiscono su un legame peptidico (il secondonumero identifica il tipo di substrato).
EC 3.4.11: idrolasi che agiscono solo sull'amminoacido N-terminaledi un peptide (il terzo numero fornisce ulterioridettagli sul tipo di substrato).
EC 3.4.11.4: idrolasi che agiscono solo sull'amminoacido N-terminaledi un tripeptide (il quarto è un numero d'ordine e segnal'ordine di scoperta degli enzimi).

Classificazione degli enzimi
Le principali classi di enzimi sono le seguenti:
EC1 Ossidoreduttasi: trasferimento di elettroni (deidrogenasi,ossidasi).
EC2 Transferasi: reazioni di trasferimento di gruppifunzionali (transaminasi, chinasi).
EC3 Idrolasi: reazione di idrolisi (lipasi, amilasi, proteasi).EC4 Liasi: addizione di gruppi a legami doppi o formazione di
doppi legami mediante rimozioni di gruppi (decarbossilasi).EC5 Isomerasi: trasferimento di gruppi all’interno di molecole
formando isomeri (isomerasi, mutasi).ù
EC6 Ligasi: formazione di legami C-C, C-S, C-O, C-N (sintetasi,polimerasi).

Proteine Globulari:
Emoglobina e Mioglobina

Il Trasporto di O2
O2 è poco solubile in H2O disciolto semplicemente nel sanguenon potrebbe essere trasportato in quantità adeguata allenecessità dell’organismo.
Nessuna catena laterale degli amminoacidi è idonea a legarereversibilmente O2.
Questo ruolo viene svolto da alcuni metalli di transizione tra cui ilFe.
Se il Fe fosse libero, interagendo con O2 genererebbe speciealtamente reattive (Reactive Oxygen Specie, ROS). I ROSpossono reagire con quasi tutte le molecole organiche(proteine, lipidi, glucidi, DNA), generando stress ossidativo.
Per evitare ciò, il Fe è inglobato in un gruppo prostetico legatoalla proteina di trasporto, detto EME, che caratterizza lamioglobina e l’emoglobina.
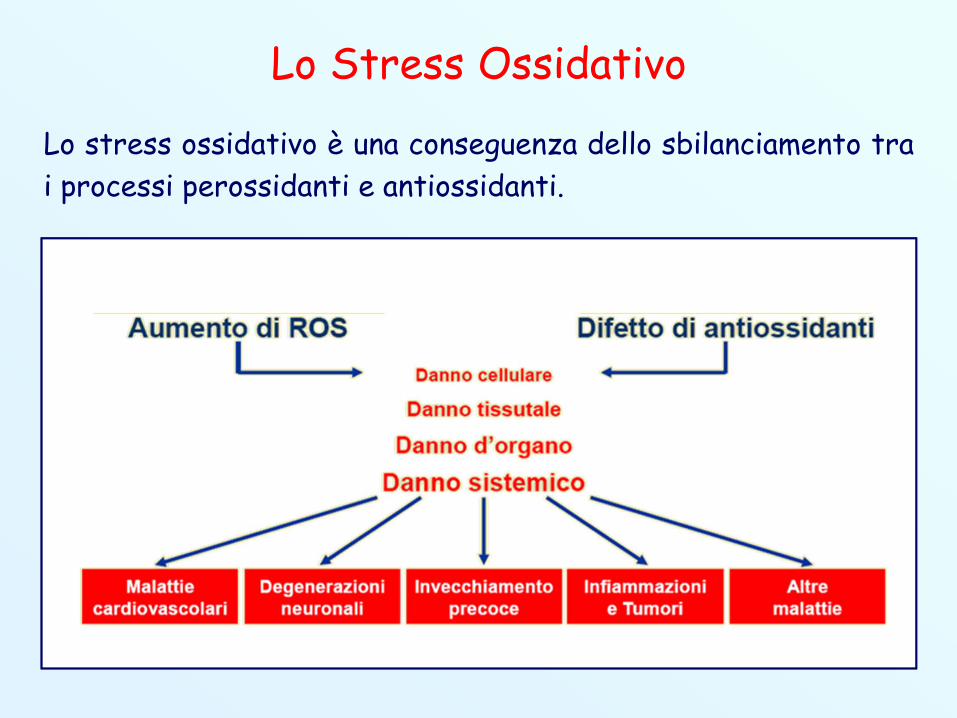
Lo Stress Ossidativo
Lo stress ossidativo è una conseguenza dello sbilanciamento trai processi perossidanti e antiossidanti.
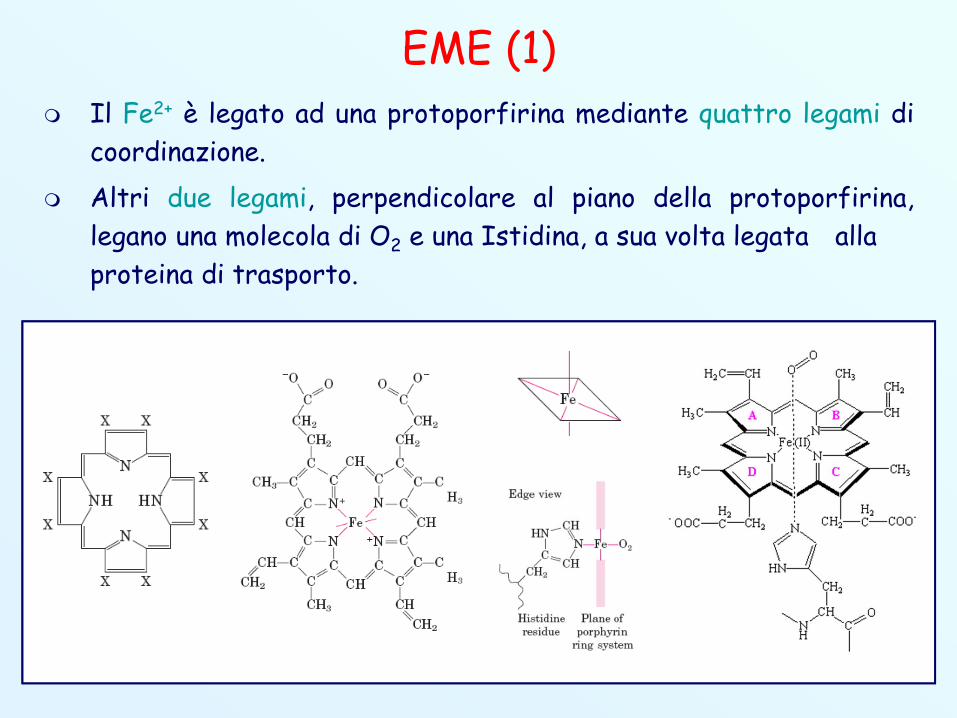
EME (1) Il Fe2+ è legato ad una protoporfirina mediante quattro legami di
coordinazione. Altri due legami, perpendicolare al piano della protoporfirina,
legano una molecola di O2 e una Istidina, a sua volta legata allaproteina di trasporto.

EME (2) Se l’EME non fosse legato alla proteina attraverso l’Istidina, in
presenza di O2 il Fe2+ potrebbe ossidarsi a Fe3+ (reazioneirreversibile). La proteina, quindi, protegge il Fe2+ dall’ossidazione.
L’EME lega O2 in maniera reversibile.
Alcune molecole di piccole dimensioni (es. NO o CO) si possonofacilmente coordinare al Fe al posto di O2, con un’affinitàmaggiore. Questo limiterebbe il trasporto di ossigeno nel sangue.(es. carbossiemoglobina).

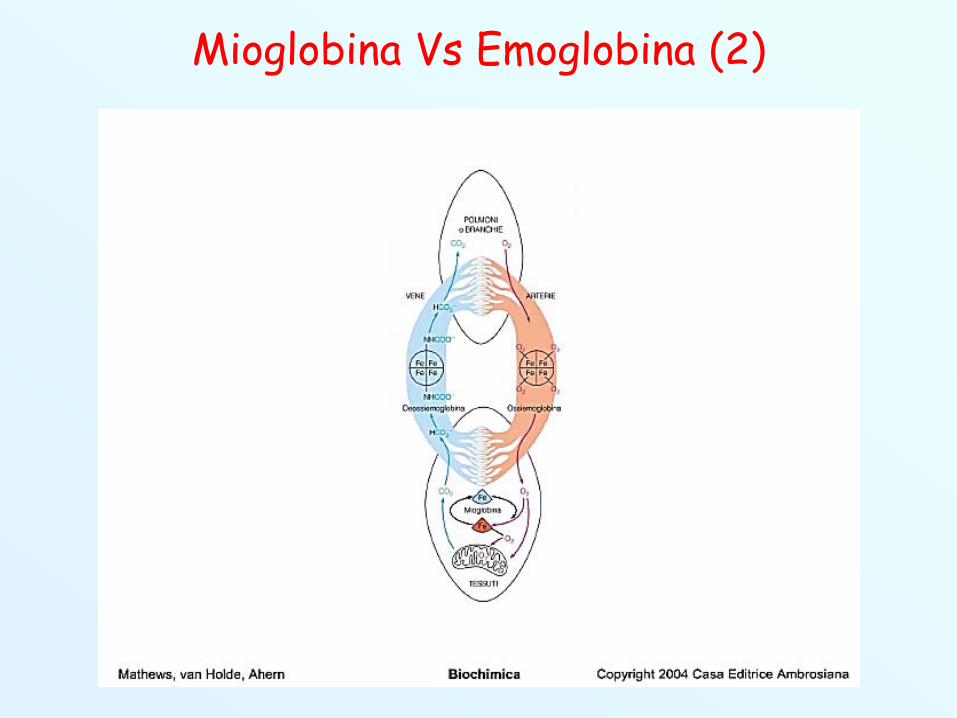
Mioglobina Vs Emoglobina (1) Mioglobina e Emoglobina sono i più importanti membri della
famiglia delle Globine. L’Emoglobina è una proteina intracellulare presente negli
eritrociti (globuli rossi, cellule del sangue). Trasporta O2
dai polmoni ai tessuti. La Mioglobina è una proteina intracellulare, presente nei
muscoli dei vertebrati. E’ in grado di legare l’O2 rilasciatodall’emoglobina ai tessuti e trasportarlo ai vari organellicellulari che lo utilizzano (es. mitocondri).La mioglobina funziona da deposito di O2 a livello muscolare.Quando il muscolo è a riposo la mioglobina rimane legataall’O2, quando il muscolo è attivo viene consumato O2
diminuisce la pressione a livello dei tessuti la mioglobinarilascia l’O2.
Mioglobina Vs Emoglobina (2)

La Mioglobina La Mioglobina (Mb) è una proteina globulare costituita da 150
amminoacidi, avente un peso molecolare di 16,7 KDa. E’ costituita da 8 segmenti di
α-elica (indicati con A ÷ H)uniti da ripiegmenti (indicaticon AB, BC, CD, …).
Il legame dell’O2 all’EMEdipende dalla “respirazione”della proteina che ne rendepossibile l’ingresso e l’uscitadalla tasca dell’ EME, che èimmersa in profondità nellastruttura della proteina.

L’Emoglobina L’Emoglobina (Hb) è una proteina globulare avente un peso
molecolare di 64,5 KDa. E’ costituita da 4 sub-unità
(ha quindi una strutturaquaternaria) ognuno delle qualipossiede un gruppo prostetico.
2 catene sono α (Hbα) e 2catene sono β (Hbβ). Essedifferiscono per il numero diresidui (141 vs. 146).
Le catene α e β sono moltosimili tra loro, ma nonidentiche e presentano unastruttura simile alla catenapolipetidica della mioglobina.
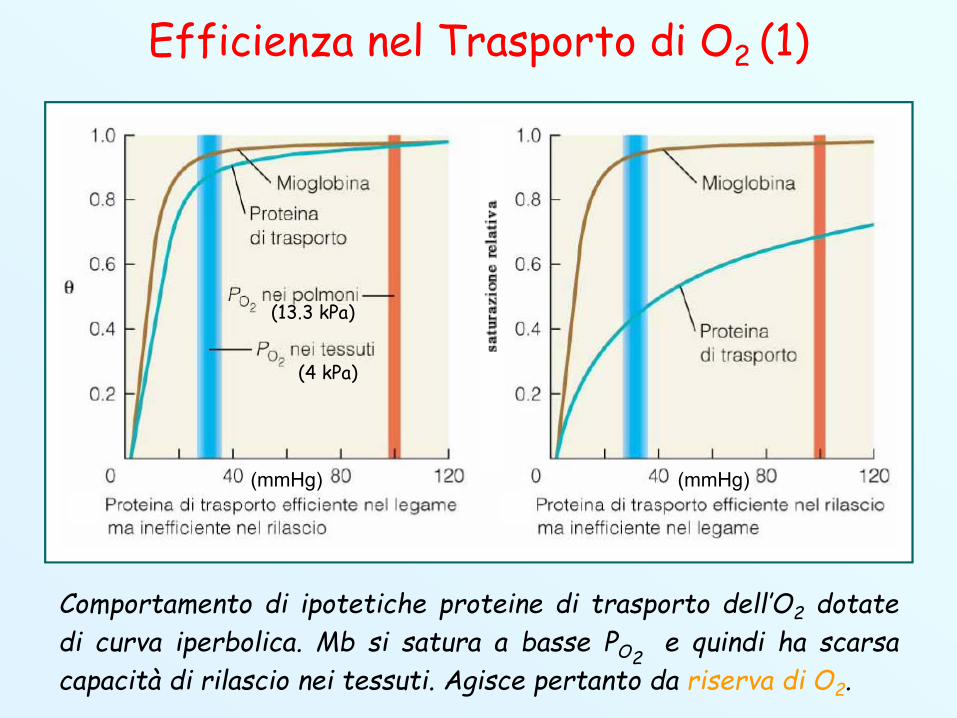
Efficienza nel Trasporto di O2 (1)
Comportamento di ipotetiche proteine di trasporto dell’O2 dotatedi curva iperbolica. Mb si satura a basse PO2
e quindi ha scarsacapacità di rilascio nei tessuti. Agisce pertanto da riserva di O2.
(4 kPa)
(13.3 kPa)
(mmHg) (mmHg)
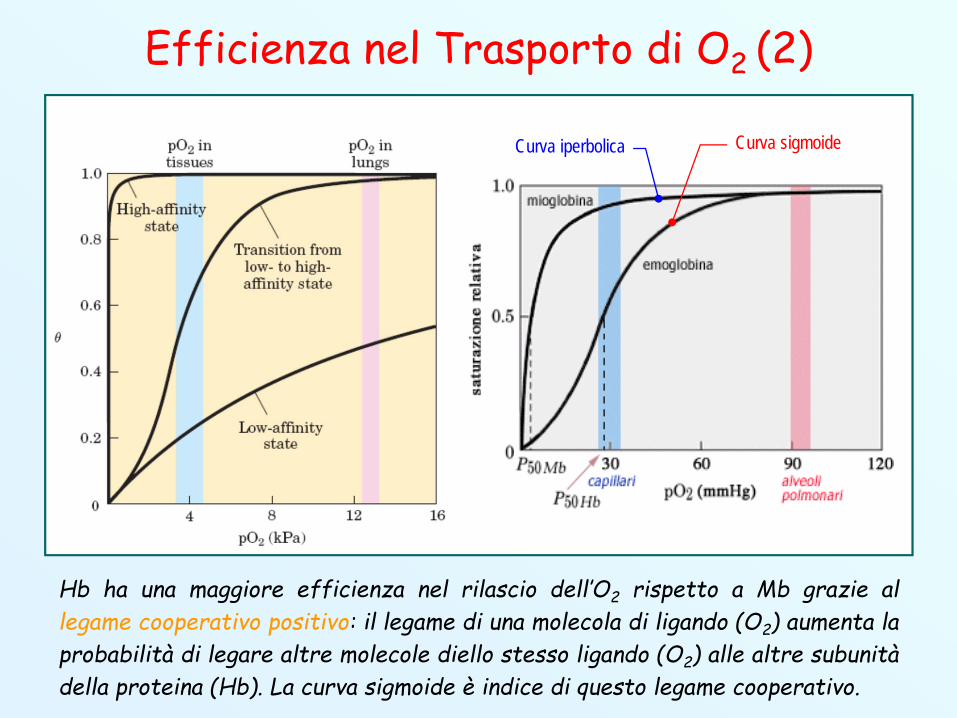
Efficienza nel Trasporto di O2 (2)
Hb ha una maggiore efficienza nel rilascio dell’O2 rispetto a Mb grazie allegame cooperativo positivo: il legame di una molecola di ligando (O2) aumenta laprobabilità di legare altre molecole diello stesso ligando (O2) alle altre subunitàdella proteina (Hb). La curva sigmoide è indice di questo legame cooperativo.
Curva sigmoideCurva iperbolica
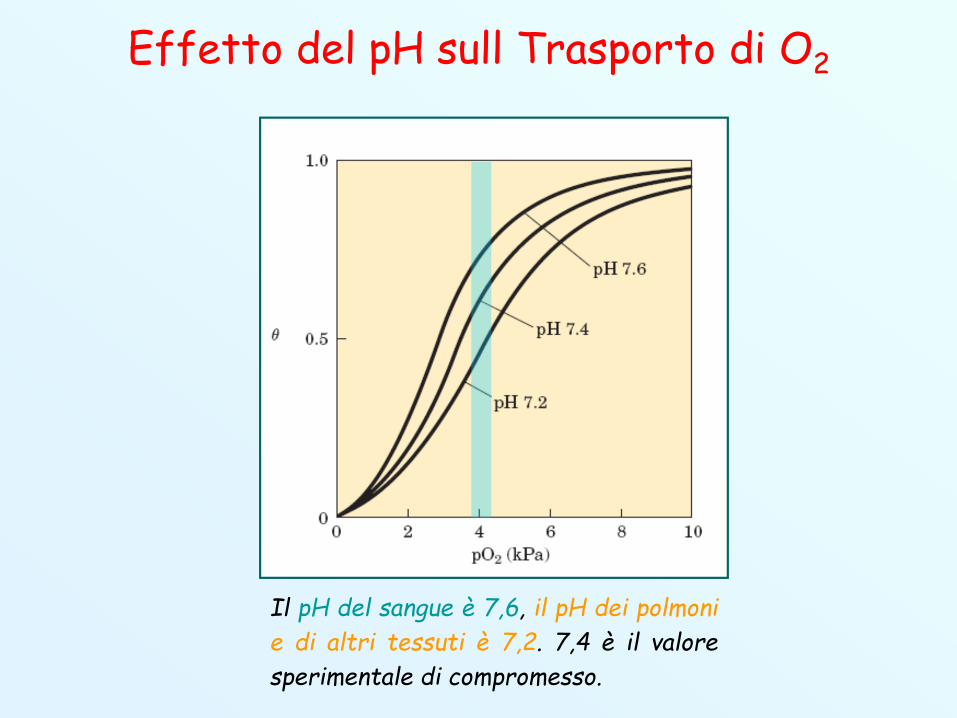
Effetto del pH sull Trasporto di O2
Il pH del sangue è 7,6, il pH dei polmonie di altri tessuti è 7,2. 7,4 è il valoresperimentale di compromesso.
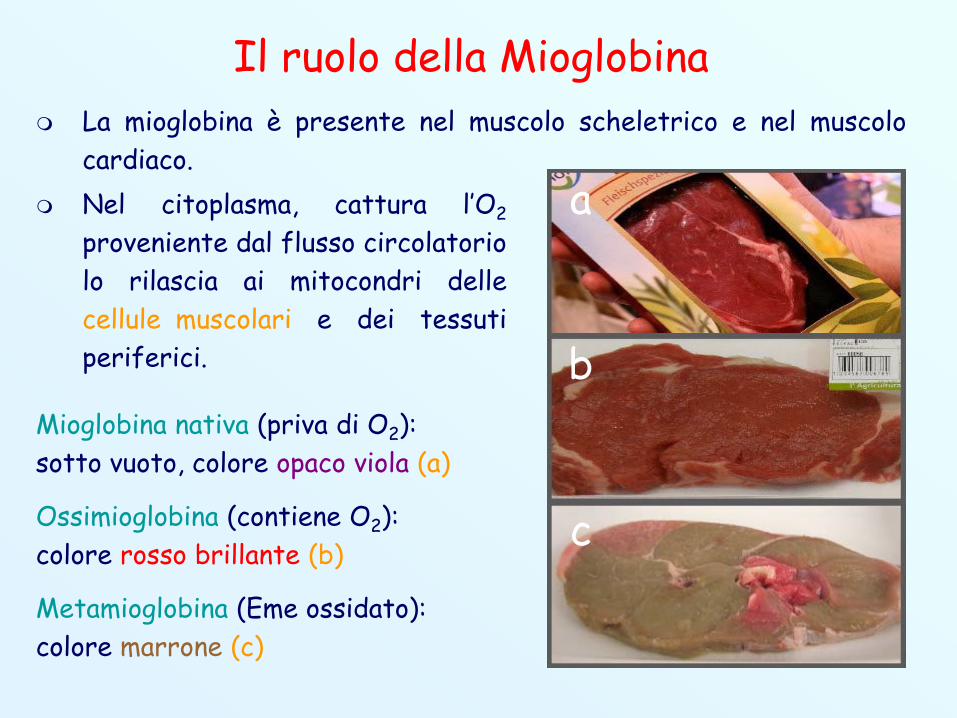
Il ruolo della Mioglobina La mioglobina è presente nel muscolo scheletrico e nel muscolo
cardiaco. Nel citoplasma, cattura l’O2
proveniente dal flusso circolatoriolo rilascia ai mitocondri dellecellule muscolari e dei tessutiperiferici.
Mioglobina nativa (priva di O2):sotto vuoto, colore opaco viola (a)
Ossimioglobina (contiene O2):colore rosso brillante (b)
Metamioglobina (Eme ossidato):colore marrone (c)
a
b
c

Il ruolo dell’Emoglobina Trasporta l’O2 dai polmoni ai globuli rossi (sistema vascolare),
dove una parte di esso può essere direttamente utilizzata per ilmetabolismo nei mitocondri. Trasporta anche CO2 e ioni H+,rilasciati dai processi ossidativi, dai tessuti ai polmoni.
Anemia: riduzione della quantità totale di emoglobina circolantenel sangue e all'interno dei globuli rossi.

Anemia Falciforme (1) L’emoglobina presenta 500 varianti genetiche (sostituzione di uno
o più residui aminoacidi) che provocano solo piccoli effettistrutturali e funzionali.
Un’eccezione è la sostituzione del glutammato in posizione 6 dellacatena β con una valina che producendo una zona idrofobica“appiccicosa” determina un’associazione anormale dell’Hb.
Quando la concentrazione di ossigeno cade al di sotto di un certolivello critico, le subunità polimerizzano in aggregati fibrosilineari, che modificano la forma della cellula.
Il risultato di questi eventi è l’assunzione da parte degli eritrocitidi una forma a falce, molto fragile, che intasa i capillari,impedendo l’ossigenazione dei tessuti.
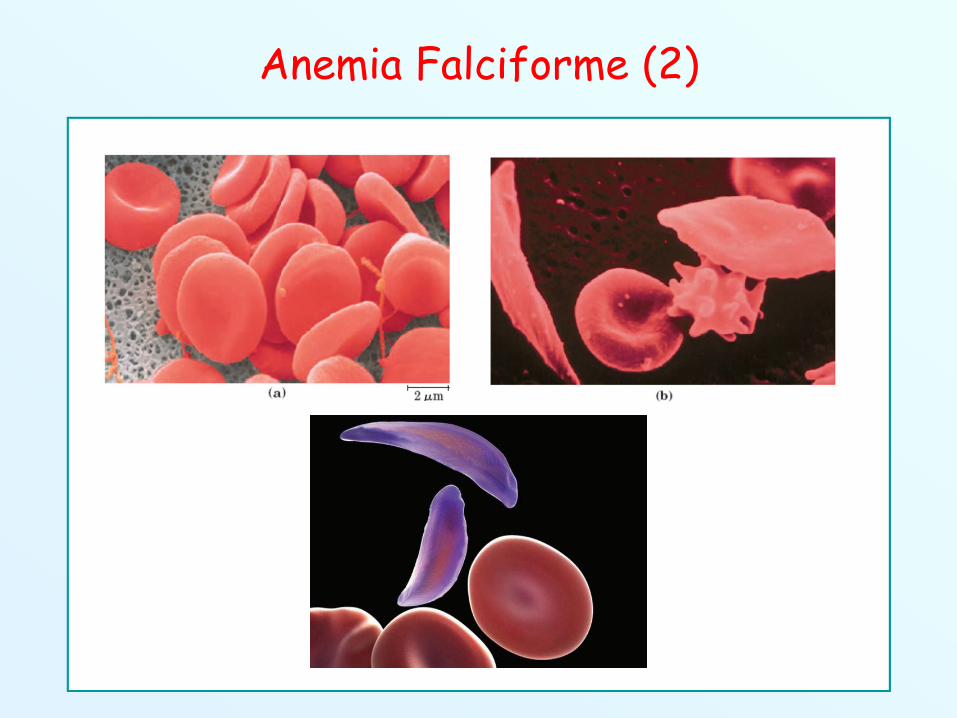
Anemia Falciforme (2)

Valutazione della Glicosilazione delle Proteine: Parametri Usuali
Concentrazione glucosio nel sangue: esprime laglicemia al momento dell’analisi.
Livello di Hb Glicosilata: riflette il controllometabolico mediato su due mesi precedenti l’analisi.
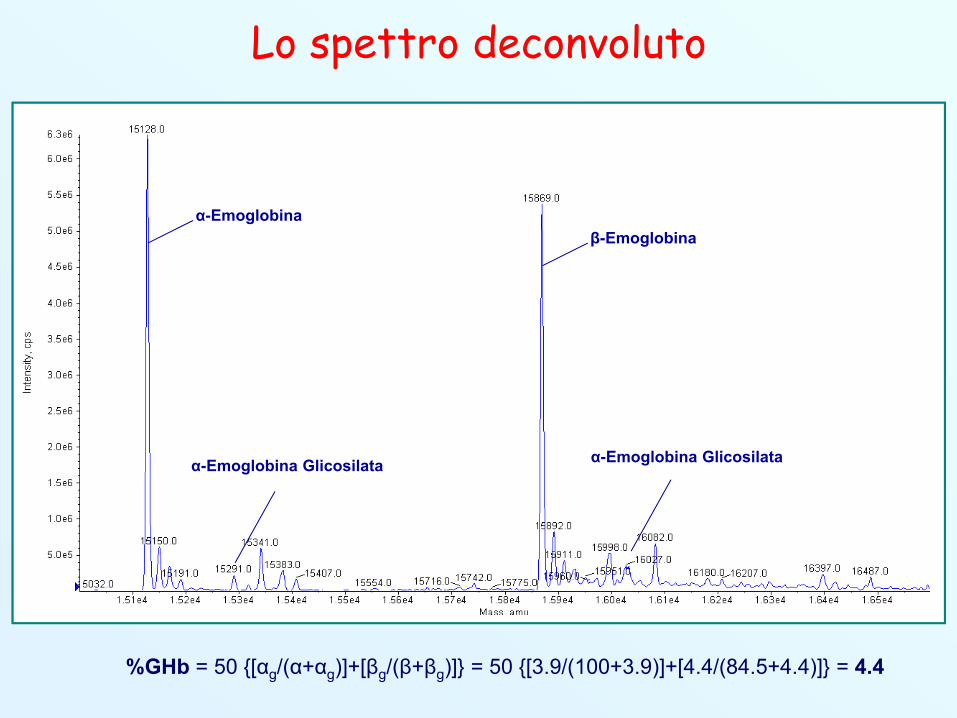
Lo spettro deconvoluto
α-Emoglobinaβ-Emoglobina
α-Emoglobina Glicosilata α-Emoglobina Glicosilata
%GHb = 50 {[αg/(α+αg)]+[βg/(β+βg)]} = 50 {[3.9/(100+3.9)]+[4.4/(84.5+4.4)]} = 4.4
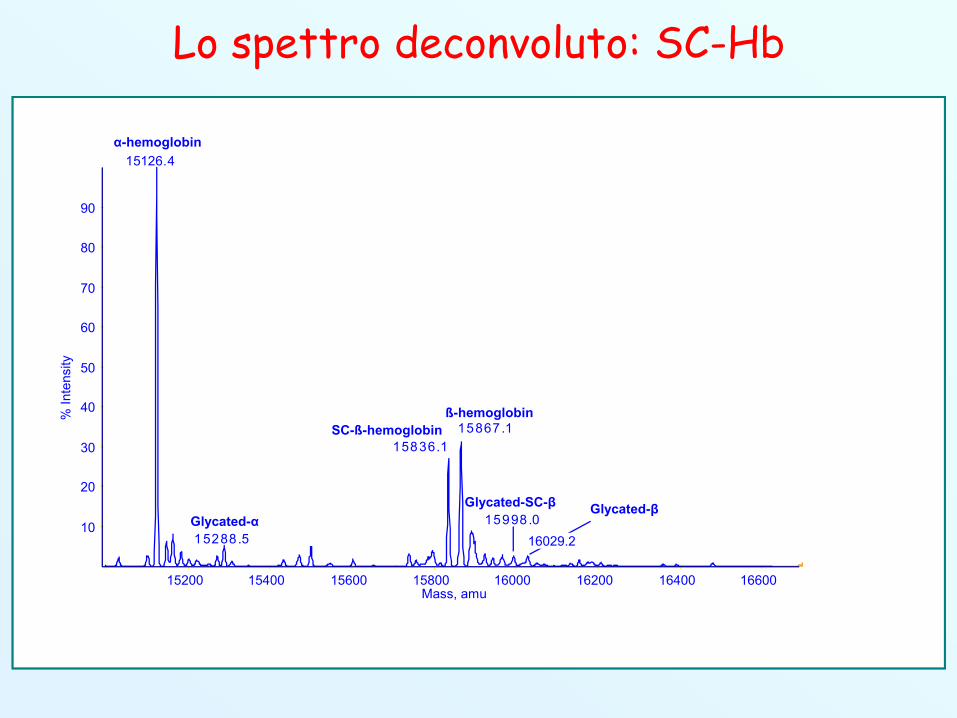
Lo spettro deconvoluto: SC-Hb
15288.5
15836.115867.1
15998.0
15200 15400 15600 15800 16000 16200 16400 16600Mass, amu
10
20
30
40
50
60
70
80
90
% In
tens
ity
15126.4
Glycated-αGlycated-β
α-hemoglobin
ß-hemoglobinSC-ß-hemoglobin
Glycated-SC-β
16029.2


















