
Il virus dell'epatite B -...
5
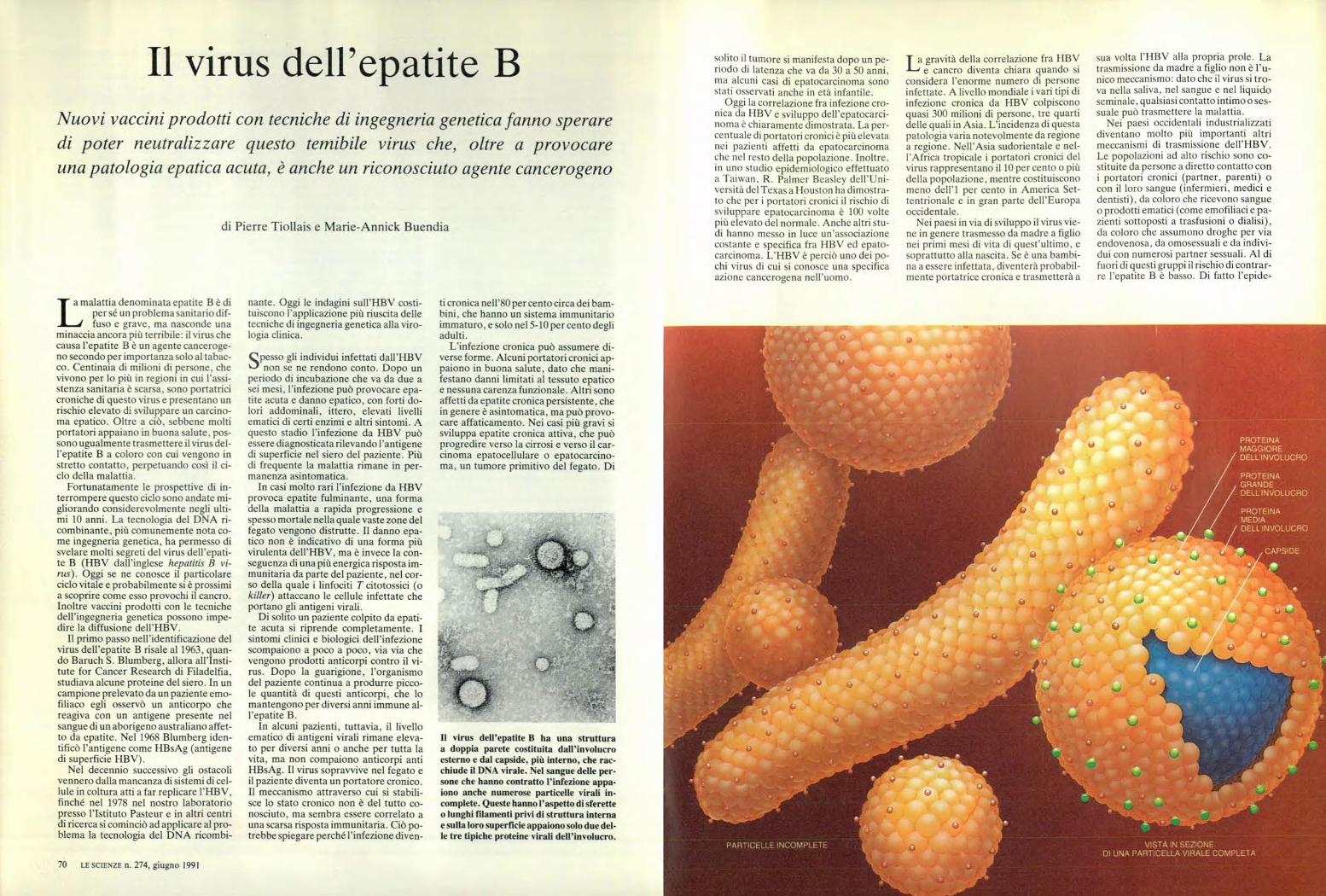
PROTEINA MAGGIORE DELL INVOLUCRO PROTEINA GRANDE DELL INVOLUCRO PRDTE:NA MEDIA DEL L rr,tvoLucRo CAPS DE PARI CELLE INCOMPLETE VISTA IN SEZIONE DI UNA PARTICELLA VIRALE COMPLETA di Pierre Tiollais e Marie-Annick Buendia solito il tumore si manifesta dopo un pe- riodo di latenza che va da 30 a 50 anni, ma alcuni casi di epatocarcinoma sono stati osservati anche in età infantile. Oggi la correlazione fra infezione cro- nica da HBV e sviluppo dell'epatocarci- noma è chiaramente dimostrata. La per- centuale di portatori cronici è più elevata nei pazienti affetti da epatocarcinoma che nel resto della popolazione. Inoltre, in uno studio epidemiologico effettuato a Taiwan, R. Palmer Beasley dell'Uni- versità del Texas a Houston ha dimostra- to che per i portatori cronici il rischio di sviluppare epatocarcinoma è 100 volte più elevato del normale. Anche altri stu- di hanno messo in luce un'associazione costante e specifica fra HBV ed epato- carcinoma. L'HBV è perciò uno dei po- chi virus di cui si conosce una specifica azione cancerogena nell'uomo. T a gravità della correlazione fra HBV I--' e cancro diventa chiara quando si considera l'enorme numero di persone infettate. A livello mondiale i vari tipi di infezione cronica da HBV colpiscono quasi 300 milioni di persone, tre quarti delle quali in Asia. L'incidenza di questa patologia varia notevolmente da regione a regione. Nell'Asia sudorientale e nel- l'Africa tropicale i portatori cronici del virus rappresentano il 10 per cento o più della popolazione, mentre costituiscono meno dell'i per cento in America Set- tentrionale e in gran parte dell'Europa occidentale. Nei paesi in via di sviluppo il virus vie- ne in genere trasmesso da madre a figlio nei primi mesi di vita di quest'ultimo, e soprattutto alla nascita. Se è una bambi- na a essere infettata, diventerà probabil- mente portatrice cronica e trasmetterà a sua volta l'HBV alla propria prole. La trasmissione da madre a figlio non è l'u- nico meccanismo: dato che il virus si tro- va nella saliva, nel sangue e nel liquido seminale, qualsiasi contatto intimo o ses- suale può trasmettere la malattia. Nei paesi occidentali industrializzati diventano molto più importanti altri meccanismi di trasmissione dell'HBV. Le popolazioni ad alto rischio sono co- stituite da persone a diretto contatto con i portatori cronici (partner, parenti) o con il loro sangue (infermieri, medici e dentisti), da coloro che ricevono sangue o prodotti ematici (come emofiliaci e pa- zienti sottoposti a trasfusioni o dialisi), da coloro che assumono droghe per via endovenosa, da omosessuali e da indivi- dui con numerosi partner sessuali. Al di fuori di questi gruppi il rischio di contrar- re l'epatite B è basso. Di fatto l'epide- Il virus dell'epatite B Nuovi vaccini prodotti con tecniche di ingegneria genetica fanno sperare di poter neutralizzare questo temibile virus che, oltre a provocare una patologia epatica acuta, è anche un riconosciuto agente cancerogeno / amalattia denominata epatite B è di per sé un problema sanitario dif- d fuso e grave, ma nasconde una minaccia ancora più terribile: il virus che causa l'epatite B è un agente canceroge- no secondo per importanza solo al tabac- co. Centinaia di milioni di persone, che vivono per lo più in regioni in cui l'assi- stenza sanitaria è scarsa, sono portatrici croniche di questo virus e presentano un rischio elevato di sviluppare un carcino- ma epatico. Oltre a ciò, sebbene molti portatori appaiano in buona salute, pos- sono ugualmente trasmettere il virus del- l'epatite B a coloro con cui vengono in stretto contatto, perpetuando così il ci- clo della malattia. Fortunatamente le prospettive di in- terrompere questo ciclo sono andate mi- gliorando considerevolmente negli ulti- mi 10 anni. La tecnologia del DNA ri- combinante, più comunemente nota co- me ingegneria genetica, ha permesso di svelare molti segreti del virus dell'epati- te B (HBV dall'inglese hepatitis B vi- rus). Oggi se ne conosce il particolare ciclo vitale e probabilmente si è prossimi a scoprire come esso provochi il cancro. Inoltre vaccini prodotti con le tecniche dell'ingegneria genetica possono impe- dire la diffusione dell'HBV. Il primo passo nell'identificazione del virus dell'epatite B risale al 1963, quan- do Baruch S. Blumberg, allora all'Insti- tute for Cancer Research di Filadelfia, studiava alcune proteine del siero. In un campione prelevato da un paziente emo- filiaco egli osservò un anticorpo che reagiva con un antigene presente nel sangue di un aborigeno australiano affet- to da epatite. Nel 1968 Blumberg iden- tificò l'antigene come HBsAg (antigene di superficie HBV). Nel decennio successivo gli ostacoli vennero dalla mancanza di sistemi di cel- lule in coltura atti a far replicare l'HBV, finché nel 1978 nel nostro laboratorio presso l'Istituto Pasteur e in altri centri di ricerca si cominciò ad applicare al pro- blema la tecnologia del DNA ricombi- 70 LE SCIENZE n. 274, giugno 1991 nante. Oggi le indagini sull'HBV costi- tuiscono l'applicazione più riuscita delle tecniche di ingegneria genetica alla viro- logia clinica. S Spesso gli individui infettati dall'HBV non se ne rendono conto. Dopo un periodo di incubazione che va da due a sei mesi, l'infezione può provocare epa- tite acuta e danno epatico, con forti do- lori addominali, ittero, elevati livelli ematici di certi enzimi e altri sintomi. A questo stadio l'infezione da HBV può essere diagnosticata rilevando l'antigene di superficie nel siero del paziente. Più di frequente la malattia rimane in per- manenza asintomatica. In casi molto rari l'infezione da HBV provoca epatite fulminante, una forma della malattia a rapida progressione e spesso mortale nella quale vaste zone del fegato vengono distrutte. Il danno epa- tico non è indicativo di una forma più virulenta dell'HBV, ma è invece la con- seguenza di una più energica risposta im- munitaria da parte del paziente, nel cor- so della quale i linfociti T citotossici (o killer) attaccano le cellule infettate che portano gli antigeni virali. Di solito un paziente colpito da epati- te acuta si riprende completamente. I sintomi clinici e biologici dell'infezione scompaiono a poco a poco, via via che vengono prodotti anticorpi contro il vi- rus. Dopo la guarigione, l'organismo del paziente continua a produrre picco- le quantità di questi anticorpi, che lo mantengono per diversi anni immune al- l'epatite B. In alcuni pazienti, tuttavia, il livello ematico di antigeni virali rimane eleva- to per diversi anni o anche per tutta la vita, ma non compaiono anticorpi anti HBsAg. Il virus sopravvive nel fegato e il paziente diventa un portatore cronico. Il meccanismo attraverso cui si stabili- sce lo stato cronico non è del tutto co- nosciuto, ma sembra essere correlato a una scarsa risposta immunitaria. Ciò po- trebbe spiegare perché l'infezione diven- ti cronica nell'80 per cento circa dei bam- bini, che hanno un sistema immunitario immaturo, e solo nel 5-10 per cento degli adulti. L'infezione cronica può assumere di- verse forme. Alcuni portatori cronici ap- paiono in buona salute, dato che mani- festano danni limitati al tessuto epatico e nessuna carenza funzionale. Altri sono affetti da epatite cronica persistente, che in genere è asintomatica, ma può provo- care affaticamento. Nei casi più gravi si sviluppa epatite cronica attiva, che può progredire verso la cirrosi e verso il car- cinoma epatocellulare o epatocarcino- ma , un tumore primitivo del fegato. Di Il virus dell'epatite B ha una struttura a doppia parete costituita dall'involucro esterno e dal capside. più interno, che rac- chiude il DNA virale. Nel sangue delle per- sone che hanno contratto l'infezione appa- iono anche numerose particelle virali in- complete. Queste hanno l'aspetto di sferette o lunghi filamenti privi di struttura interna e sulla loro superficie appaiono solo due del- le tre tipiche proteine virali dell'involucro.
Transcript of Il virus dell'epatite B -...
PROTEINAMAGGIOREDELL INVOLUCRO
PROTEINAGRANDEDELL INVOLUCRO
PRDTE:NAMEDIADEL L rr,tvoLucRo
CAPS DE
PARI CELLE INCOMPLETE VISTA IN SEZIONEDI UNA PARTICELLA VIRALE COMPLETA
di Pierre Tiollais e Marie-Annick Buendia
solito il tumore si manifesta dopo un pe-riodo di latenza che va da 30 a 50 anni,ma alcuni casi di epatocarcinoma sonostati osservati anche in età infantile.
Oggi la correlazione fra infezione cro-nica da HBV e sviluppo dell'epatocarci-noma è chiaramente dimostrata. La per-centuale di portatori cronici è più elevatanei pazienti affetti da epatocarcinomache nel resto della popolazione. Inoltre,in uno studio epidemiologico effettuatoa Taiwan, R. Palmer Beasley dell'Uni-versità del Texas a Houston ha dimostra-to che per i portatori cronici il rischio disviluppare epatocarcinoma è 100 voltepiù elevato del normale. Anche altri stu-di hanno messo in luce un'associazionecostante e specifica fra HBV ed epato-carcinoma. L'HBV è perciò uno dei po-chi virus di cui si conosce una specificaazione cancerogena nell'uomo.
T a gravità della correlazione fra HBVI--' e cancro diventa chiara quando siconsidera l'enorme numero di personeinfettate. A livello mondiale i vari tipi diinfezione cronica da HBV colpisconoquasi 300 milioni di persone, tre quartidelle quali in Asia. L'incidenza di questapatologia varia notevolmente da regionea regione. Nell'Asia sudorientale e nel-l'Africa tropicale i portatori cronici delvirus rappresentano il 10 per cento o piùdella popolazione, mentre costituisconomeno dell'i per cento in America Set-tentrionale e in gran parte dell'Europaoccidentale.
Nei paesi in via di sviluppo il virus vie-ne in genere trasmesso da madre a figlionei primi mesi di vita di quest'ultimo, esoprattutto alla nascita. Se è una bambi-na a essere infettata, diventerà probabil-mente portatrice cronica e trasmetterà a
sua volta l'HBV alla propria prole. Latrasmissione da madre a figlio non è l'u-nico meccanismo: dato che il virus si tro-va nella saliva, nel sangue e nel liquidoseminale, qualsiasi contatto intimo o ses-suale può trasmettere la malattia.
Nei paesi occidentali industrializzatidiventano molto più importanti altrimeccanismi di trasmissione dell'HBV.Le popolazioni ad alto rischio sono co-stituite da persone a diretto contatto coni portatori cronici (partner, parenti) ocon il loro sangue (infermieri, medici edentisti), da coloro che ricevono sangueo prodotti ematici (come emofiliaci e pa-zienti sottoposti a trasfusioni o dialisi),da coloro che assumono droghe per viaendovenosa, da omosessuali e da indivi-dui con numerosi partner sessuali. Al difuori di questi gruppi il rischio di contrar-re l'epatite B è basso. Di fatto l'epide-
Il virus dell'epatite BNuovi vaccini prodotti con tecniche di ingegneria genetica fanno speraredi poter neutralizzare questo temibile virus che, oltre a provocareuna patologia epatica acuta, è anche un riconosciuto agente cancerogeno
/ amalattia denominata epatite B è diper sé un problema sanitario dif-
d fuso e grave, ma nasconde unaminaccia ancora più terribile: il virus checausa l'epatite B è un agente canceroge-no secondo per importanza solo al tabac-co. Centinaia di milioni di persone, chevivono per lo più in regioni in cui l'assi-stenza sanitaria è scarsa, sono portatricicroniche di questo virus e presentano unrischio elevato di sviluppare un carcino-ma epatico. Oltre a ciò, sebbene moltiportatori appaiano in buona salute, pos-sono ugualmente trasmettere il virus del-l'epatite B a coloro con cui vengono instretto contatto, perpetuando così il ci-clo della malattia.
Fortunatamente le prospettive di in-terrompere questo ciclo sono andate mi-gliorando considerevolmente negli ulti-mi 10 anni. La tecnologia del DNA ri-combinante, più comunemente nota co-me ingegneria genetica, ha permesso disvelare molti segreti del virus dell'epati-te B (HBV dall'inglese hepatitis B vi-rus). Oggi se ne conosce il particolareciclo vitale e probabilmente si è prossimia scoprire come esso provochi il cancro.Inoltre vaccini prodotti con le tecnichedell'ingegneria genetica possono impe-dire la diffusione dell'HBV.
Il primo passo nell'identificazione delvirus dell'epatite B risale al 1963, quan-do Baruch S. Blumberg, allora all'Insti-tute for Cancer Research di Filadelfia,studiava alcune proteine del siero. In uncampione prelevato da un paziente emo-filiaco egli osservò un anticorpo chereagiva con un antigene presente nelsangue di un aborigeno australiano affet-to da epatite. Nel 1968 Blumberg iden-tificò l'antigene come HBsAg (antigenedi superficie HBV).
Nel decennio successivo gli ostacolivennero dalla mancanza di sistemi di cel-lule in coltura atti a far replicare l'HBV,finché nel 1978 nel nostro laboratoriopresso l'Istituto Pasteur e in altri centridi ricerca si cominciò ad applicare al pro-blema la tecnologia del DNA ricombi-
70 LE SCIENZE n. 274, giugno 1991
nante. Oggi le indagini sull'HBV costi-tuiscono l'applicazione più riuscita delletecniche di ingegneria genetica alla viro-logia clinica.
SSpesso gli individui infettati dall'HBV
non se ne rendono conto. Dopo unperiodo di incubazione che va da due asei mesi, l'infezione può provocare epa-tite acuta e danno epatico, con forti do-lori addominali, ittero, elevati livelliematici di certi enzimi e altri sintomi. Aquesto stadio l'infezione da HBV puòessere diagnosticata rilevando l'antigenedi superficie nel siero del paziente. Piùdi frequente la malattia rimane in per-manenza asintomatica.
In casi molto rari l'infezione da HBVprovoca epatite fulminante, una formadella malattia a rapida progressione espesso mortale nella quale vaste zone delfegato vengono distrutte. Il danno epa-tico non è indicativo di una forma piùvirulenta dell'HBV, ma è invece la con-seguenza di una più energica risposta im-munitaria da parte del paziente, nel cor-so della quale i linfociti T citotossici (okiller) attaccano le cellule infettate cheportano gli antigeni virali.
Di solito un paziente colpito da epati-te acuta si riprende completamente. Isintomi clinici e biologici dell'infezionescompaiono a poco a poco, via via chevengono prodotti anticorpi contro il vi-rus. Dopo la guarigione, l'organismodel paziente continua a produrre picco-le quantità di questi anticorpi, che lomantengono per diversi anni immune al-l'epatite B.
In alcuni pazienti, tuttavia, il livelloematico di antigeni virali rimane eleva-to per diversi anni o anche per tutta lavita, ma non compaiono anticorpi antiHBsAg. Il virus sopravvive nel fegato eil paziente diventa un portatore cronico.Il meccanismo attraverso cui si stabili-sce lo stato cronico non è del tutto co-nosciuto, ma sembra essere correlato auna scarsa risposta immunitaria. Ciò po-trebbe spiegare perché l'infezione diven-
ti cronica nell'80 per cento circa dei bam-bini, che hanno un sistema immunitarioimmaturo, e solo nel 5-10 per cento degliadulti.
L'infezione cronica può assumere di-verse forme. Alcuni portatori cronici ap-paiono in buona salute, dato che mani-festano danni limitati al tessuto epaticoe nessuna carenza funzionale. Altri sonoaffetti da epatite cronica persistente, chein genere è asintomatica, ma può provo-care affaticamento. Nei casi più gravi sisviluppa epatite cronica attiva, che puòprogredire verso la cirrosi e verso il car-cinoma epatocellulare o epatocarcino-ma , un tumore primitivo del fegato. Di
Il virus dell'epatite B ha una strutturaa doppia parete costituita dall'involucroesterno e dal capside. più interno, che rac-chiude il DNA virale. Nel sangue delle per-sone che hanno contratto l'infezione appa-iono anche numerose particelle virali in-complete. Queste hanno l'aspetto di sferetteo lunghi filamenti privi di struttura internae sulla loro superficie appaiono solo due del-le tre tipiche proteine virali dell'involucro.

Portatori cronici dell'epatite B
aZg;Rh.
I portatori cronici del virus dell'epatite B sono numerosi soprattutto nei paesi in via disviluppo, dove questa infezione è endemica. Nella sola Asia ve ne sono circa 225 milioni.
miologia di questa malattia è molto simi-le a quella della sindrome da immuno-deficienza acquisita (AIDS), il che spie-ga perché l'infezione da HBV sia comu-ne nei pazienti affetti da AIDS o dal co-siddetto complesso correlato all'AIDS.L'epatite, tuttavia, è molto più contagio-sa dell'AIDS.
Pertanto l'epatite B è soprattutto unamalattia infantile nelle nazioni in via disviluppo, mentre nei paesi occidentalicolpisce per lo più gli adulti. Questa di-stinzione ha importanza pratica per lefuture strategie di vaccinazione. Neipaesi in via di sviluppo sarà necessaria lavaccinazione di massa, mentre nei paesioccidentali basterà proteggere solo lapopolazione ad alto rischio.
I vaccini antivirali funzionano sensibi-lizzando il sistema immunitario di un
individuo alle molecole virali. Le primericerche sull'HBV si sono quindi con-centrate per lo più sulla struttura e sulciclo vitale del virus al fine di identificarei bersagli adatti per il vaccino.
Tutti i virus sono parassiti cellulari for-mati da acido nucleico, che ne costitui-sce il genoma, racchiuso da un rivesti-mento proteico o capside. Per replicarsiun virus deve penetrare nella cellula eutilizzare i meccanismi cellulari per sin-tetizzare le proteine del rivestimento e ilgenoma virale. Una volta che gli acidinucleici sono completi del loro rivesti-mento, le nuove particelle virali lascianola cellula e vanno a infettarne altre.
Il virus dell'epatite B ha una strutturaa doppia parete costituita da due rivesti-menti proteici concentrici. L'involucroesterno contiene tre proteine denomina-te proteina maggiore, media e grande;l'antigene di superficie si trova su tutte etre le proteine. Il rivestimento interno,che è il capside vero e proprio, è costi-
tuito da una singola proteina che circon-da il DNA virale e interagisce con esso.
Il genoma dell'HBV, che venne isola-to per la prima volta nel 1974 da WilliamS. Robinson della Stanford UniversitySchool of Medicine, è una molecola cir-colare di DNA costituita da sole 3200subunità nucleotidiche. È il genoma piùpiccolo che si conosca fra tutti quelli deivirus animali: il genoma del comune vi-rus dell'herpes simplex, per esempio, è50 volte più grande. Come le molecoledi DNA in gran parte degli organismi, ilgenoma dell'HBV è formato da due fi-lamenti uniti tramite le basi azotate, mapossiede una caratteristica insolita: unfilamento è più lungo dell'altro. Il brevefilamento «più», di lunghezza variabile,è lungo solo il 50-80 per cento del corri-spondente filamento «meno». (Comespiegheremo più avanti, questa insolitastruttura è una conseguenza del partico-larissimo meccanismo di replicazione delvirus.) La struttura circolare del genomaè assicurata dall'accoppiamento dellebasi alle estremità dei filamenti.
Usando le tecniche del DNA ricombi-nante il nostro gruppo di ricerca riuscì,alla fine degli anni settanta, a clonare ilgenoma dell'HBV nel batterio Escheri-chia coli. Questo fondamentale passoavanti rese possibile la produzione digrandi quantità del virus e delle sue com-ponenti per ulteriori studi. Patrick Char-nay del nostro laboratorio, in collabora-zione con Francis Galibert dell'Ospeda-le Saint-Louis di Parigi, determinò in se-guito la sequenza completa di nucleotidinel genoma dell'HBV, fornendo così leprime informazioni disponibili sull'orga-nizzazione genetica del virus.
Il genoma dell'HBV è un prodigio dicompattezza. E costituito da quattro soligeni potenziali, denominati S, C, P e X,che si sovrappongono ampiamente. Le
sequenze regolatrici che controllano laproduzione delle proteine virali e il ciclodi replicazione sono anch'esse inserite inqueste sequenze codificanti.
Il gene S codifica per la proteina mag-giore dell'involucro e specifica in modocompleto l'HBsAg. Una sequenza di cir-ca 500 nucleotidi che precede il gene Spuò venire a sua volta trascritta insiemecon esso. Come ha dimostrato WolframH. Gerlich dell'Università di Gottinga,questa sequenza «a monte» può esseredivisa in due regioni, dette pre-S/ e pre--S2, che sono coinvolte nella sintesi dellealtre proteine dell'involucro: la proteinamedia è codificata dalla regione pre-S2e dal gene S, mentre la proteina grandeè codificata dalle regioni pre-S/ e pre-S2e dal gene S. La regione pre-S/ svolgeanche un ruolo importante al momentodell'ingresso del virus in una cellula epa-tica, come ha dimostrato A. RobertNeurath del New York Blood Center.
Il gene C codifica per la proteina delcapside. Come il gene S, anche il C èpreceduto da una breve regione pre-Cche codifica per un peptide idrofobo at-tivo nell'assemblaggio della particella vi-rale . Il gene P, che è così lungo da inclu-dere parti di tutti gli altri geni, codificaper enzimi essenziali al ciclo di replica-zione virale. Il gene X si trova a cavallodelle estremità unite dei filamenti diDNA virale. Il suo prodotto proteico fa-vorisce l'espressione di tutti i geni viraliinteragendo con una specifica sequenzadi DNA presente nel genoma dell'HBV.
Nel ciclo vitale dell'HBV la sintesi del-le proteine virali è strettamente regolataa livello sia della trascrizione sia dellatraduzione. Si conoscono due tipi diRNA messaggero (m-RNA) copiato dalgenoma virale. Il più piccolo, della lun-ghezza di circa 2100 nucleotidi, codificaper le proteine maggiore e media dell'in-volucro. L'm-RNA più grande, dellalunghezza di circa 3500 nucleotidi, è sor-prendente: è addirittura più lungo delgenoma completo e contiene una se-quenza ripetitiva terminale di circa 100nucleotidi. Questo m-RNA codifica perla proteina del capside e per i prodot-ti del gene P, come hanno dimostratoHarold E. Varmus dell'Università del-la California a San Francisco e HeinzSchaller dell'Università di Heidelberg;rappresenta inoltre un intermedio dellareplicazione del DNA virale, come ve-dremo fra poco.
L'espressione di tutti i geni virali è at-tivata da elementi presenti nel genomadell'HBV che favoriscono la trascrizionee operano di preferenza nelle celluleepatiche. Altri elementi regolatori mo-dulano i livelli di singole proteine. Unbuon esempio di questo complesso mec-canismo è dato dalle variazioni che siosservano nelle quantità delle tre protei-ne dell'involucro sintetizzate in celluleepatiche infettate. La proteina grande èprodotta in piccole quantità e si trovasolo alla superficie di particelle viralicomplete, in grado di propagare l'infe-
72 LE SCIENZE n. 274, giugno 1991
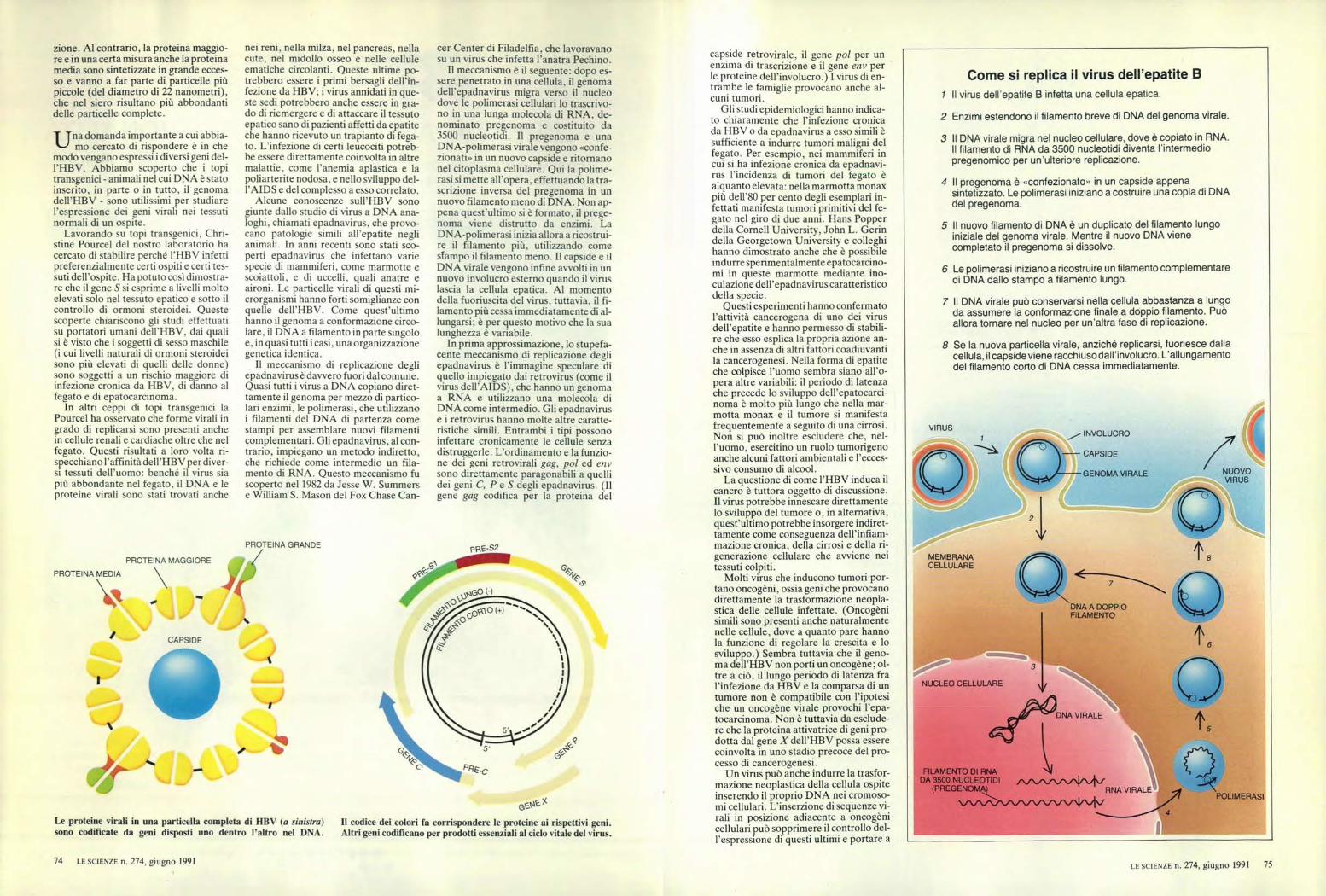
Come si replica il virus dell'epatite B
i Il virus dell'epatite B infetta una cellula epatica.
2 Enzimi estendono il filamento breve di DNA del genoma virale.
3 Il DNA virale migra nel nucleo cellulare, dove è copiato in RNA.Il filamento di RNA da 3500 nucleotidi diventa l'intermediopregenomico per un'ulteriore replicazione.
4 Il pregenoma è «confezionato» in un capside appenasintetizzato. Le polimerasi iniziano a costruire una copia di DNAdel pregenoma.
5 II nuovo filamento di DNA è un duplicato del filamento lungoiniziale del genoma virale. Mentre il nuovo DNA vienecompletato il pregenoma si dissolve.
6 Le polimerasi iniziano a ricostruire un filamento complementaredi DNA dallo stampo a filamento lungo.
7 Il DNA virale può conservarsi nella cellula abbastanza a lungoda assumere la conformazione finale a doppio filamento. Puòallora tornare nel nucleo per un'altra fase di replicazione.
8 Se la nuova particella virale, anziché replicarsi, fuoriesce dallacellula, il capside viene racchiuso dall'involucro. L'allungamentodel filamento corto di DNA cessa immediatamente.
INVOLUCRO
CAPSIDE
VIRUS
NUOVOVIRUS
MEMBRANACELLULARE
DNA A DOPPIOFILAMENTO
NUCLEO CELLULARE
FILAMENTO DI RNADA 3500 NUCLEOTIDI
(PREGENOMA) RNA VIRALEPOLIMERASI
3
Le proteine virali in una particella completa di HBV (a sinistra)sono codificate da geni disposti uno dentro l'altro nel DNA.
codice dei colori fa corrispondere le proteine ai rispettivi geni.Altri geni codificano per prodotti essenziali al ciclo vitale del virus.
PROTEINA GRANDE
41.• —#
PROTEINA MAGGIORE
PROTEINA MEDIA
CAPSIDE
zione. Al contrario, la proteina maggio-re e in una certa misura anche la proteinamedia sono sintetizzate in grande ecces-so e vanno a far parte di particelle piùpiccole (del diametro di 22 nanometri),che nel siero risultano più abbondantidelle particelle complete.
Una domanda importante a cui abbia-mo cercato di rispondere è in che
modo vengano espressi i diversi geni del-l'HBV. Abbiamo scoperto che i topitransgenici - animali nel cui DNA è statoinserito, in parte o in tutto, il genomadell'HBV - sono utilissimi per studiarel'espressione dei geni virali nei tessutinormali di un ospite.
Lavorando su topi transgenici, Chri-siine Pourcel del nostro laboratorio hacercato di stabilire perché l'HBV infettipreferenzialmente certi ospiti e certi tes-suti dell'ospite. Ha potuto così dimostra-re che il gene S si esprime a livelli moltoelevati solo nel tessuto epatico e sotto ilcontrollo di ormoni steroidei. Questescoperte chiariscono gli studi effettuatisu portatori umani dell'HBV, dai qualisi è visto che i soggetti di sesso maschile(i cui livelli naturali di ormoni steroideisono più elevati di quelli delle donne)sono soggetti a un rischio maggiore diinfezione cronica da HBV, di danno alfegato e di epatocarcinoma.
In altri ceppi di topi transgenici laPourcel ha osservato che forme virali ingrado di replicarsi sono presenti anchein cellule renali e cardiache oltre che nelfegato. Questi risultati a loro volta ri-specchiano l'affinità dell'HBV per diver-si tessuti dell'uomo: benché il virus siapiù abbondante nel fegato, il DNA e leproteine virali sono stati trovati anche
nei reni, nella milza, nel pancreas, nellacute, nel midollo osseo e nelle celluleematiche circolanti. Queste ultime po-trebbero essere i primi bersagli dell'in-fezione da HBV; i virus annidati in que-ste sedi potrebbero anche essere in gra-do di riemergere e di attaccare il tessutoepatico sano di pazienti affetti da epatiteche hanno ricevuto un trapianto di fega-to. L'infezione di certi leucociti potreb-be essere direttamente coinvolta in altremalattie, come l'anemia aplastica e lapoliarterite nodosa, e nello sviluppo del-l'AIDS e del complesso a esso correlato.
Alcune conoscenze sull'HBV sonogiunte dallo studio di virus a DNA ana-loghi, chiamati epadnavirus, che provo-cano patologie simili all'epatite neglianimali. In anni recenti sono stati sco-perti epadnavirus che infettano variespecie di mammiferi, come marmotte escoiattoli, e di uccelli, quali anatre eaironi. Le particelle virali di questi mi-crorganismi hanno forti somiglianze conquelle dell'HBV. Come quest'ultimohanno il genoma a conformazione circo-lare, il DNA a filamento in parte singoloe, in quasi tutti i casi, una organizzazionegenetica identica.
Il meccanismo di replicazione degliepadnavirus è davvero fuori dal comune.Quasi tutti i virus a DNA copiano diret-tamente il genoma per mezzo di partico-lari enzimi, le polimerasi, che utilizzanoi filamenti del DNA di partenza comestampi per assemblare nuovi filamenticomplementari. Gli epadnavirus, al con-trario, impiegano un metodo indiretto,che richiede come intermedio un fila-mento di RNA. Questo meccanismo fuscoperto nel 1982 da Jesse W. Summerse William S. Mason del Fox Chase Can-
cer Center di Filadelfia, che lavoravanosu un virus che infetta l'anatra Pechino.
Il meccanismo è il seguente: dopo es-sere penetrato in una cellula, il genomadell'epadnavirus migra verso il nucleodove le polimerasi cellulari lo trascrivo-no in una lunga molecola di RNA, de-nominato pregenoma e costituito da3500 nucleotidi. Il pregenoma e unaDNA-polimerasi virale vengono «confe-zionati» in un nuovo capside e ritornanonel citoplasma cellulare. Qui la polime-rasi si mette all'opera, effettuando la tra-scrizione inversa del pregenoma in unnuovo filamento meno di DNA. Non ap-pena quest'ultimo si è formato, il prege-noma viene distrutto da enzimi. LaDNA-polimerasi inizia allora a ricostrui-re il filamento più, utilizzando comestampo il filamento meno. Il capside e ilDNA virale vengono infine avvolti in unnuovo involucro esterno quando il viruslascia la cellula epatica. Al momentodella fuoriuscita del virus, tuttavia, il fi-lamento più cessa immediatamente di al-lungarsi; è per questo motivo che la sualunghezza è variabile.
In prima approssimazione, lo stupefa-cente meccanismo di replicazione degliepadnavirus è l'immagine speculare diquello impiegato dai retrovirus (come ilvirus dell'AIDS), che hanno un genomaa RNA e utilizzano una molecola diDNA come intermedio. Gli epadnaviruse i retrovirus hanno molte altre caratte-ristiche simili. Entrambi i tipi possonoinfettare cronicamente le cellule senzadistruggerle. L'ordinamento e la funzio-ne dei geni retrovirali gag, poi ed envsono direttamente paragonabili a quellidei geni C, P e S degli epadnavirus. (Ilgene gag codifica per la proteina del
capside retrovirale, il gene poi per unenzima di trascrizione e il gene env perle proteine dell'involucro.) I virus di en-trambe le famiglie provocano anche al-cuni tumori.
Gli studi epidemiologici hanno indica-to chiaramente che l'infezione cronicada HBV o da epadnavirus a esso simili èsufficiente a indurre tumori maligni delfegato. Per esempio, nei mammiferi incui si ha infezione cronica da epadnavi-rus l'incidenza di tumori del fegato èalquanto elevata: nella marmotta monaxpiù dell'80 per cento degli esemplari in-fettati manifesta tumori primitivi del fe-gato nel giro di due anni. Hans Popperdella Cornell University, John L. Gerindella Georgetown University e colleghihanno dimostrato anche che è possibileindurre sperimentalmente epatocarcino-mi in queste marmotte mediante ino-culazione dell'epadnavirus caratteristicodella specie.
Questi esperimenti hanno confermatol'attività cancerogena di uno dei virusdell'epatite e hanno permesso di stabili-re che esso esplica la propria azione an-che in assenza di altri fattori coadiuvantila cancerogenesi. Nella forma di epatiteche colpisce l'uomo sembra siano all'o-pera altre variabili: il periodo di latenzache precede lo sviluppo dell'epatocarci-noma è molto più lungo che nella mar-motta monax e il tumore si manifestafrequentemente a seguito di una cirrosi.Non si può inoltre escludere che, nel-l'uomo, esercitino un ruolo tumorigenoanche alcuni fattori ambientali e l'ecces-sivo consumo di alcool.
La questione di come l'HBV induca ilcancro è tuttora oggetto di discussione.Il virus potrebbe innescare direttamentelo sviluppo del tumore o, in alternativa,quest'ultimo potrebbe insorgere indiret-tamente come conseguenza dell'infiam-mazione cronica, della cirrosi e della ri-generazione cellulare che avviene neitessuti colpiti.
Molti virus che inducono tumori por-tano oncogèni, ossia geni che provocanodirettamente la trasformazione neopla-stica delle cellule infettate. (Oncogènisimili sono presenti anche naturalmentenelle cellule, dove a quanto pare hannola funzione di regolare la crescita e losviluppo.) Sembra tuttavia che il geno-ma dell'HBV non porti un oncogène; ol-tre a ciò, il lungo periodo di latenza fral'infezione da HBV e la comparsa di untumore non è compatibile con l'ipotesiche un oncogène virale provochi l'epa-tocarcinoma. Non è tuttavia da esclude-re che la proteina attivatrice di geni pro-dotta dal gene X dell'HBV possa esserecoinvolta in uno stadio precoce del pro-cesso di cancerogenesi.
Un virus può anche indurre la trasfor-mazione neoplastica della cellula ospiteinserendo il proprio DNA nei cromoso-mi cellulari. L'inserzione di sequenze vi-rali in posizione adiacente a oncogènicellulari può sopprimere il controllo del-l'espressione di questi ultimi e portare a
74 LE SCIENZE n. 274, giugno 1991LE SCIENZE n. 274, giugno 1991 75
GENE DELLA CRESCITACELLULARE ATTIVATO
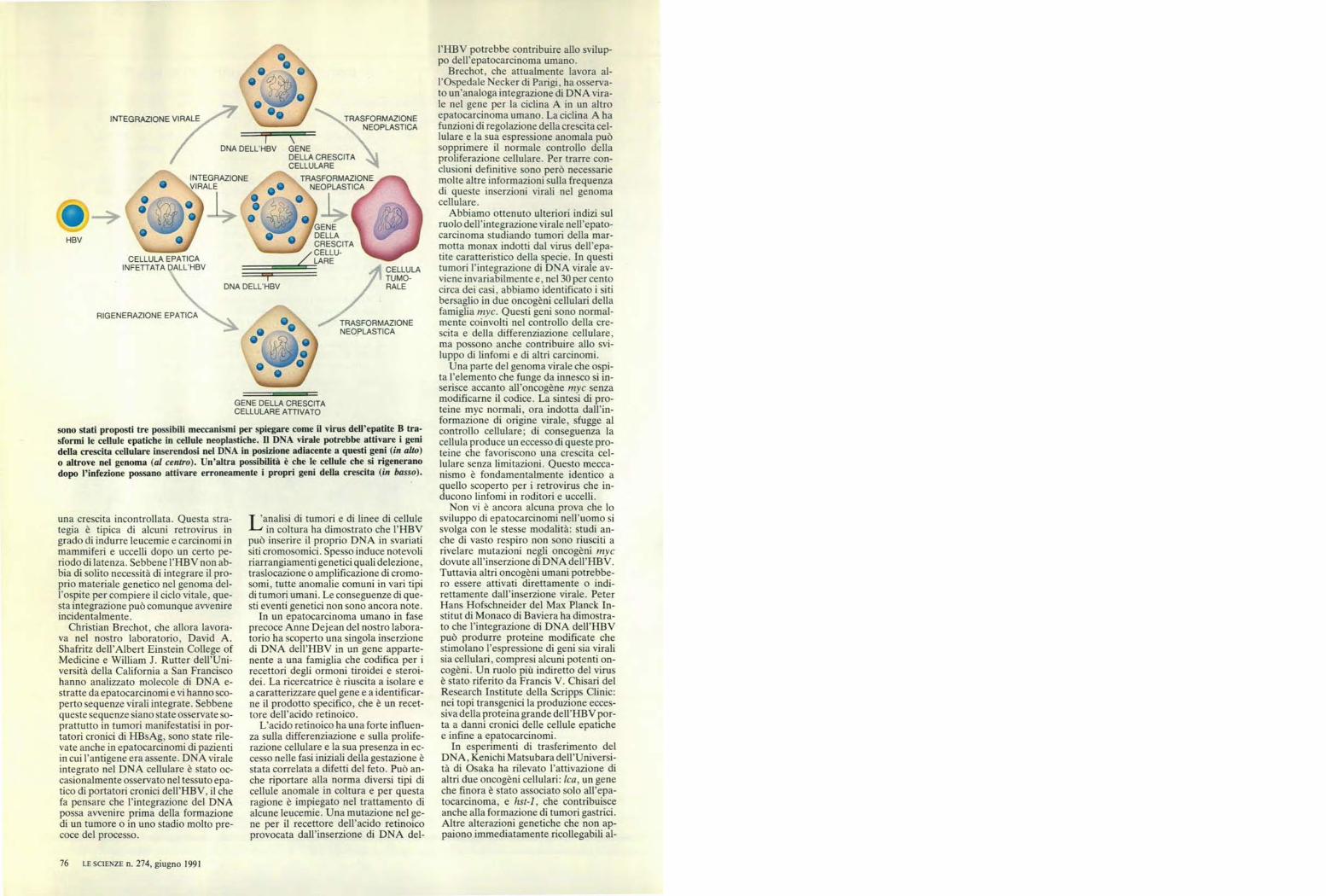
sono stati proposti tre possibili meccanismi per spiegare come il virus dell'epatite B tra-sformi le cellule epatiche in cellule neoplastiche. Il DNA virale potrebbe attivare i genidella crescita cellulare inserendosi nel DNA in posizione adiacente a questi geni (in alto)o altrove nel genoma (al centro). Un'altra possibilità è che le cellule che si rigeneranodopo l'infezione possano attivare erroneamente i propri geni della crescita (in basso).
DNA DELL'HBV GENEDELLA CRESCITACELLULARE
TRASFORMAZIONEO NEOPLASTICA
CELLULA EPATICAINFETTATA DALL'HBV
O—
GENEDELLACRESCITACELLU-
/ LARECELLULATUMO-RALE
RIGENERAZIONE EPATICATRASFORMAZIONENEOPLASTICA
TRASFORMAZIONENEOPLASTICA
INTEGRAZIONEVIRALE
DNA DELL HBV
INTEGRAZIONE VIRALE
HBV
una crescita incontrollata. Questa stra-tegia è tipica di alcuni retrovirus ingrado di indurre leucemie e carcinomi inmammiferi e uccelli dopo un certo pe-riodo di latenza. Sebbene l'HBV non ab-bia di solito necessità di integrare il pro-prio materiale genetico nel genoma del-l'ospite per compiere il ciclo vitale, que-sta integrazione può comunque avvenireincidentalmente.
Christian Brechot, che allora lavora-va nel nostro laboratorio, David A.Shafritz dell'Albert Einstein College ofMedicine e William J. Rutter dell'Uni-versità della California a San Franciscohanno analizzato molecole di DNA e-stratte da epatocarcinomi e vi hanno sco-perto sequenze virali integrate. Sebbenequeste sequenze siano state osservate so-prattutto in tumori manifestatisi in por-tatori cronici di HBsAg, sono state rile-vate anche in epatocarcinomi di pazientiin cui l'antigene era assente. DNA viraleintegrato nel DNA cellulare è stato oc-casionalmente osservato nel tessuto epa-tico di portatori cronici dell'HBV, il chefa pensare che l'integrazione del DNApossa avvenire prima della formazionedi un tumore o in uno stadio molto pre-coce del processo.
J:analisi di tumori e di linee di cellulein coltura ha dimostrato che l'HBV
può inserire il proprio DNA in svariatisiti cromosomici. Spesso induce notevoliriarrangiamenti genetici quali delezione,traslocazione o amplificazione di cromo-somi, tutte anomalie comuni in vari tipidi tumori umani. Le conseguenze di que-sti eventi genetici non sono ancora note.
In un epatocarcinoma umano in faseprecoce Anne Dejean del nostro labora-torio ha scoperto una singola inserzionedi DNA dell'HBV in un gene apparte-nente a una famiglia che codifica per irecettori degli ormoni tiroidei e steroi-dei. La ricercatrice è riuscita a isolare ea caratterizzare quel gene e a identificar-ne il prodotto specifico, che è un recet-tore dell'acido retinoico.
L'acido retinoico ha una forte influen-za sulla differenziazione e sulla prolife-razione cellulare e la sua presenza in ec-cesso nelle fasi iniziali della gestazione èstata correlata a difetti del feto. Può an-che riportare alla norma diversi tipi dicellule anomale in coltura e per questaragione è impiegato nel trattamento dialcune leucemie. Una mutazione nel ge-ne per il recettore dell'acido retinoicoprovocata dall'inserzione di DNA del-
l'HBV potrebbe contribuire allo svilup-po dell'epatocarcinoma umano.
Brechot, che attualmente lavora al-l'Ospedale Necker di Parigi, ha osserva-to un'analoga integrazione di DNA vira-le nel gene per la ciclina A in un altroepatocarcinoma umano. La ciclina A hafunzioni di regolazione della crescita cel-lulare e la sua espressione anomala puòsopprimere il normale controllo dellaproliferazione cellulare. Per trarre con-clusioni definitive sono però necessariemolte altre informazioni sulla frequenzadi queste inserzioni virali nel genomacellulare.
Abbiamo ottenuto ulteriori indizi sulruolo dell'integrazione virale nell'epato-carcinoma studiando tumori della mar-motta monax indotti dal virus dell'epa-tite caratteristico della specie. In questitumori l'integrazione di DNA virale av-viene invariabilmente e, nel 30 per centocirca dei casi, abbiamo identificato i sitibersaglio in due oncogèni cellulari dellafamiglia myc. Questi geni sono normal-mente coinvolti nel controllo della cre-scita e della differenziazione cellulare,ma possono anche contribuire allo svi-luppo di linfomi e di altri carcinomi.
Una parte del genoma virale che ospi-ta l'elemento che funge da innesco si in-serisce accanto all'oncogène myc senzamodificarne il codice. La sintesi di pro-teine myc normali, ora indotta dall'in-formazione di origine virale, sfugge alcontrollo cellulare; di conseguenza lacellula produce un eccesso di queste pro-teine che favoriscono una crescita cel-lulare senza limitazioni. Questo mecca-nismo è fondamentalmente identico aquello scoperto per i retrovirus che in-ducono linfomi in roditori e uccelli.
Non vi è ancora alcuna prova che losviluppo di epatocarcinomi nell'uomo sisvolga con le stesse modalità: studi an-che di vasto respiro non sono riusciti arivelare mutazioni negli oncogèni mycdovute all'inserzione di DNA dell'HBV.Tuttavia altri oncogèni umani potrebbe-ro essere attivati direttamente o indi-rettamente dall'inserzione virale. PeterHans Hofschneider del Max Planck In-stitut di Monaco di Baviera ha dimostra-to che l'integrazione di DNA dell'HBVpuò produrre proteine modificate chestimolano l'espressione di geni sia viralisia cellulari, compresi alcuni potenti on-cogèni. Un ruolo più indiretto del virusè stato riferito da Francis V. Chisari delResearch Institute della Scripps Clinic:nei topi transgenici la produzione ecces-siva della proteina grande dell'HBV por-ta a danni cronici delle cellule epatichee infine a epatocarcinomi.
In esperimenti di trasferimento delDNA, Kenichi Matsubara dell'Universi-tà di Osaka ha rilevato l'attivazione dialtri due oncogèni cellulari: lca, un geneche finora è stato associato solo all'epa-tocarcinoma, e hst-I , che contribuisceanche alla formazione di tumori gastrici.Altre alterazioni genetiche che non ap-paiono immediatamente ricollegabili al-
76 LE SCIENZE n. 274, giugno 1991

l'infezione da HBV sono state anch'esseosservate in epatocarcinomi dell'uomo.Saranno necessari ulteriori studi percomprendere a fondo il ruolo dei virusnella cancerogenesi a livello molecolare.
Gli esperimenti descritti dimostrano
quanto sia stata fondamentale latecnologia del DNA ricombinante per laconoscenza della biologia di base del-l'HBV. Questa tecnologia ha anche ap-plicazioni pratiche nello sviluppo di nuo-ve tecniche diagnostiche e di vaccini.
I test diagnostici per l'epatite B sonostati migliorati grazie alla tecnica dellaibridazione molecolare, che sfrutta l'ele-vata affinità di brevi filamenti di DNAdonato nei confronti del relativo DNAcomplementare. Il DNA virale clonatoe marcato con isotopi radioattivi puòfungere da sonda sensibile e affidabileper rilevare nel siero la presenza di par-ticelle virali in grado di diffondere l'in-fezione. Attualmente la tecnica dell'ibri-dazione molecolare che utilizza DNAdell'HBV viene impiegata abitualmenteper individuare un'infezione recente daquesto virus. Poiché l'identificazione delDNA virale consente anche una valuta-zione accurata del livello di replicazionevirale in corso, l'ibridazione molecolareè una tecnica adatta per seguire i pro-gressi di un trattamento antivirale.
La sensibilità del metodo può essereulteriormente migliorata mediante latecnica della reazione a catena della po-limerasi, un semplice procedimento en-zimatico per duplicare anche tracce dispecifiche sequenze di DNA a velocitàesponenziale (si veda l'articolo La sco-perta della reazione a catena della poli-merasi di Kary B. Mullis in «Le Scienze»n. 262, giugno 1990). Con questa tecnicasi può aumentare di più di 1000 volte lasensibilità dei test diagnostici basati sul-l'ibridazione: si riescono a individuareanche solo 100 particelle virali in un mil-lilitro di siero, che è considerata la con-centrazione minima perché si possa dif-fondere l'infezione.
Con la tecnologia del DNA ricombi-nante si sta procedendo anche allo svi-luppo di nuovi vaccini. Queste ricercheebbero inizio nel 1970, quando SaulKrugman e i suoi collaboratori dellaNew York University dimostrarono cheil siero, inattivato termicamente , di unportatore cronico poteva avere un effet-to protettivo in persone suscettibili al-l'infezione da HBV. In seguito diversigruppi di ricerca hanno riscontrato uneffetto protettivo negli scimpanzé di par-ticelle virali purificate che portavanol'antigene di superficie.
Nel 1976 Philippe Maupas e collabo-ratori dell'Università di Tours hanno ri-ferito i primi risultati di vaccinazioni ese-guite su esseri umani. Il vaccino consi-steva in particelle virali purificate incom-plete - essenzialmente involucri vuoti re-canti l'HBsAg - estratte dal sangue diportatori cronici e trattate in modo daeliminare tutte le particelle che poteva-
no diffondere l'infezione. È stato l'unicovaccino antivirale mai sviluppato a par-tire dal sangue di pazienti. Maupas e col-leghi hanno poi dimostrato chiaramentela sicurezza e l'efficacia del vaccino in unampio gruppo di soggetti, che compren-dono bambini di regioni dove l'HBV èendemico e membri di popolazioni adalto rischio di paesi in cui il virus non èendemico. (Il vaccino si è dimostratomeno efficace in pazienti dializzati, pro-babilmente a causa dell'indebolimentodel loro sistema immunitario.)
Sebbene il vaccino derivato dal sierosia valido, la sua produzione pone diver-si problemi: la disponibilità di siero uma-no prelevato a portatori cronici è limita-ta e il procedimento di purificazione èlungo e costoso. Inoltre ciascuna partitadi vaccino preparato da una fonte diver-sa deve essere provata su scimpanzé pergarantirne la sicurezza. Per tutte questeragioni la produzione di un vaccino perl'epatite B con le tecniche dell'ingegne-ria genetica è una prospettiva vantaggio-sa da vari punti di vista.
Q ono diverse le tecniche di ingegneriagenetica che potrebbero essere uti-
lizzate per lo sviluppo di vaccini, ma inpratica le scelte sono più limitate perchél'HBsAg è pienamente immunogeno so-lo se conserva in modo completo la pro-pria struttura naturale. Batteri modifica-ti per mezzo dell'ingegneria geneticapossono produrre grandi quantità di an-tigene, che però non ha la conformazio-ne chimica e la struttura corretta; invecei lieviti e le cellule di mammifero sono ingrado di sintetizzare le proteine specifi-che dell'antigene.
Attualmente sono già sul mercato vac-cini per l'HBV ottenuti in cellule di lie-viti o di mammiferi. Rutter ha prodottoun vaccino impiegando il comune lievitodi birra, Saccharomyces cerevisiae. Nel1984 Marie Louise Michel del nostro la-boratorio ha costruito geneticamenteuna linea di cellule ovariche di cricetocinese che sono in grado di produrre an-tigeni dell'HBV. Il DNA virale ricombi-nante da lei inserito conteneva sia il geneS sia la regione pre-S2. Le particelle vi-rali risultanti recano perciò tanto la pro-teina maggiore quanto la proteina mediadell'involucro e presentano i due deter-minanti antigenici HBsAg e pre-S2.
Questo fatto potrebbe essere signifi-cativo, perché David R. Milich dellaScripps Clinic ha dimostrato che nei topiil determinante pre-S2 scatena una ri-sposta immunitaria più intensa che nonl'HBsAg. Inoltre in ceppi di topo chenon producono anticorpi per l'HBsAgl'immunizzazione con le particelle ri-combinanti ottenute dalla Michel con-sente di aggirare l'ostacolo della manca-ta risposta: questa proprietà potrebbe ri-velarsi importante nella vaccinazione dipersone che non rispondono all'antigenedi superficie.
Un altro valido impiego della tecnicadel DNA ricombinante sarebbe quello
di costruire un vaccino contenente par-ticelle vive impiegando come vettori ilvirus del vaiolo bovino o l'adenovirus.Per mezzo dell'ingegneria genetica sipuò fare in modo che questi virus inno-cui esprimano antigeni dell'HBV, maquesta tecnica non è stata finora appli-cata a vaccini destinati all'uomo. Do-vrebbe essere possibile anche sintetizza-re chimicamente sequenze di amminoa-cidi che duplichino i determinanti anti-genici dell'HBV e ottenere in questomodo un vaccino poco costoso. Oggipurtroppo questo metodo non è pratica-bile perché l'immunogenicità dei peptidiartificiali, come quelli prodotti dai bat-teri, è estremamente bassa.
Oltre ai vaccini che possono impedirel'infezione da HBV, la tecnologia delDNA ricombinante potrebbe essere sulpunto di fornire una cura per i portatoricronici. Nell'agosto 1990 Robert P. Pe-rillo della Washington University e col-leghi hanno annunciato che il trattamen-to quotidiano con interferone alfa, unaproteina ottenuta con tecniche di inge-gneria genetica che potenzia la rispostaimmunitaria, ha eliminato o inibito l'in-fezione in più di un terzo di 85 portatoricronici dell'epatite B. Ulteriori ricerchesull'uso dell'interferone contro l'HBVpotranno perfezionare questo tipo ditrattamento.
Negli ultimi 10 anni la conoscenzasempre crescente della biologia moleco-lare dell'HBV ha trovato diverse appli-cazioni cliniche, soprattutto per preve-nire l'infezione, ma molti dei possibilibenefici sono stati appena intravisti. Neipaesi in via di sviluppo la vaccinazionedi massa con il vaccino ricombinante - ilprimo vaccino destinato all'uomo svi-luppato con le tecnologie dell'ingegneriagenetica - costituirà un grande progressonel controllo dell'epatite B. (In Italia lavaccinazione obbligatoria verrà attuatacon questo tipo di vaccino.) Una campa-gna di questo genere darà un duplice be-neficio: la vaccinazione potrà prevenirenon solo l'epatite acuta, ma anche il tu-more che le è spesso associato.
BIBLIOGRAFIA
CHISARI FRANCIS V. (a cura), Advan-ces in Hepatitis Research, Masson Publi-shing USA, 1984.
GERETY ROBERT L (a cura), Hepati-tis B, Academic Press, 1985.
TIOLLAIS PIERRE, POURCEL CHRISTINEe DEJEAN ANNE, The Hepatitis B Virus in<Nature», 317, n. 6037, 10 ottobre 1985.
GANEM DON e VARMUS H. E., The Mo-lecular Biology of the Hepatitis B Virusesin «Annual Review of Biochemistry»,56, pagg. 651-693, 1987.
ZUCKERMAN ARIE J. (a cura), Vira!Hepatitis and Liver Disease, Alan R.Liss, Inc., 1988.
78 LE SCIENZE n. 274, giugno 1991








![Tales of Childhood [Op.46] · dim. poco a tempo cou tenere corda din. e calando PPP Tempo I. > dim. rall. dim. cresc. dim. cresc. e Tall. poco a poco cresc. poco e rall. c tempo din.](https://static.fdocumenti.com/doc/165x107/610be0f99e7a740cb429fe89/tales-of-childhood-op46-dim-poco-a-tempo-cou-tenere-corda-din-e-calando-ppp.jpg)









