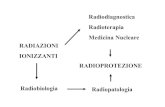

FEDELTA’ NELLA REPLICAZIONE · Radiazioni ultraviolette, ... •Interessare entrambe le eliche...
87
FEDELTA’ NELLA REPLICAZIONE Frequenza di errore = 1 /10 8 TO 10 10 bp replicate • Livelli bilanciati di dNTP • Reazione di polimerizzazione = meccanismo a due step Il dNTP si appaia alla base complementare sullo stampo Il sito attivo della polimerasi si chiude In presenza di un corretto appaiamento avviene la reazione Cambiamento conformazionale nella proteina • Attività 3’ 5’ esonucleasi Correzzione di bozza (proofreading) • Presenza di sistemi enzimatici addizionali • Necessità di un primer per l’attività DNA polimerasica Gli errori di cattivo appaiamento sono più frequenti quando l’elica in sintesi è stabilizzata da pochi legami idrogeno • Riparo degli errori durante e dopo la replicazione
Transcript of FEDELTA’ NELLA REPLICAZIONE · Radiazioni ultraviolette, ... •Interessare entrambe le eliche...

FEDELTA’ NELLA REPLICAZIONE Frequenza di errore = 1 /108 TO 1010 bp replicate
• Livelli bilanciati di dNTP • Reazione di polimerizzazione = meccanismo a due step Il dNTP si appaia alla base complementare sullo stampo Il sito attivo della polimerasi si chiude In presenza di un corretto appaiamento avviene la reazione Cambiamento conformazionale nella proteina • Attività 3’ 5’ esonucleasi Correzzione di bozza (proofreading) • Presenza di sistemi enzimatici addizionali • Necessità di un primer per l’attività DNA polimerasica Gli errori di cattivo appaiamento sono più frequenti quando
l’elica in sintesi è stabilizzata da pochi legami idrogeno • Riparo degli errori durante e dopo la replicazione
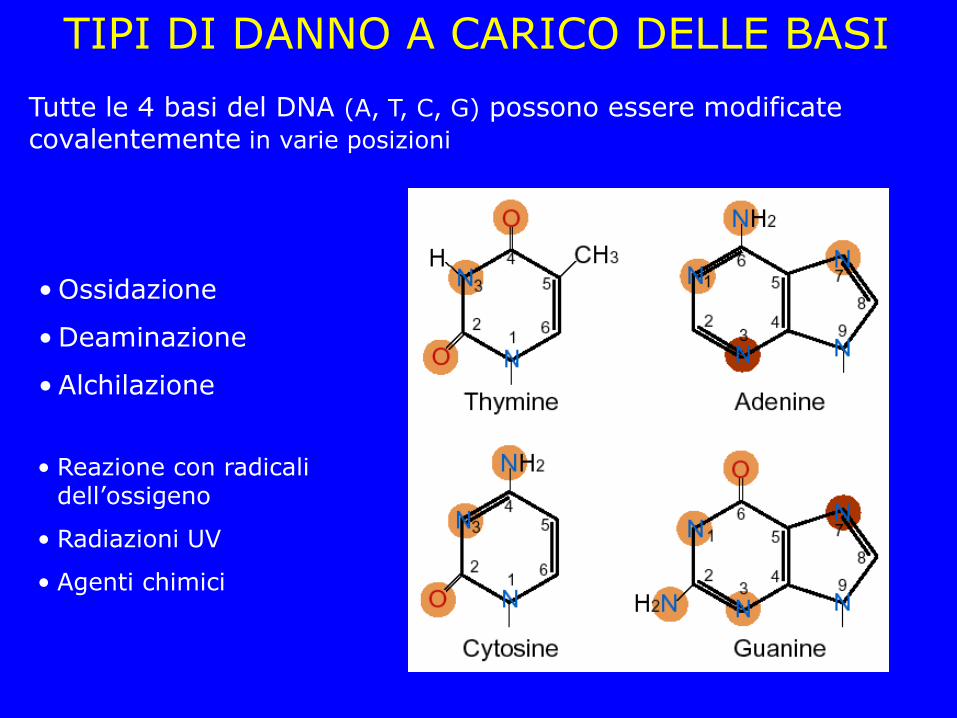
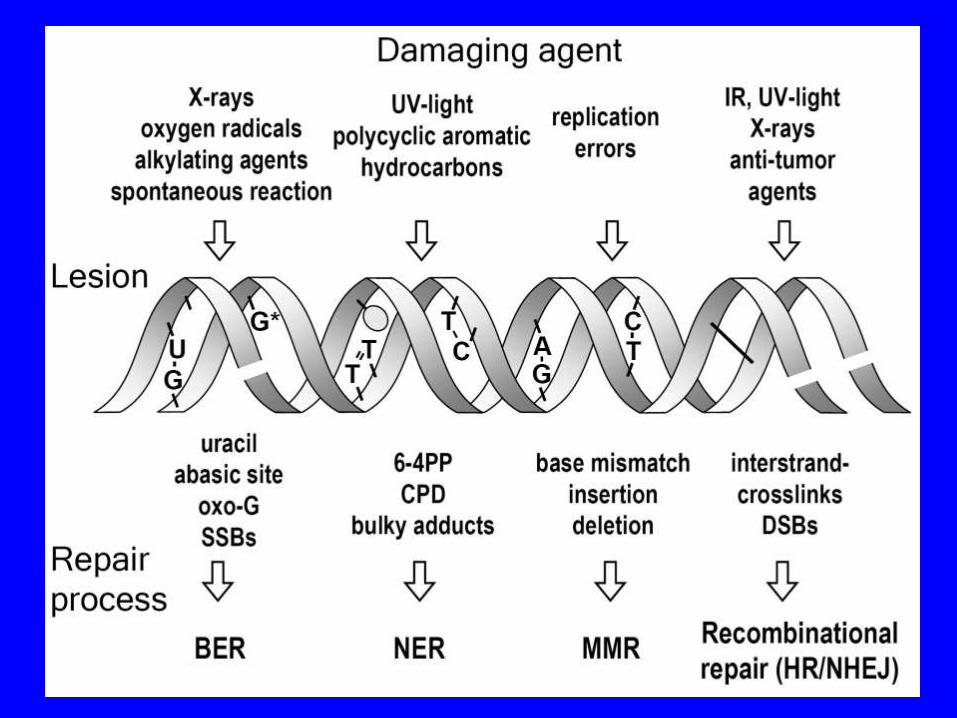
TIPI DI DANNO A CARICO DELLE BASI
Tutte le 4 basi del DNA (A, T, C, G) possono essere modificate covalentemente in varie posizioni
• Ossidazione
• Deaminazione
• Alchilazione
• Reazione con radicali dell’ossigeno
• Radiazioni UV
• Agenti chimici

DANNI SPONTANEI A CARICO DEL DNA
Perdita spontanea di basi Migliaia di purine e centinaia di pirimidine per genoma /giorno
Deaminazione spontanea Centinaia di uracile per genoma /giorno Adenina ipoxantina Gunanina Xantina 5-metil-citosina Timina

Deaminazioni
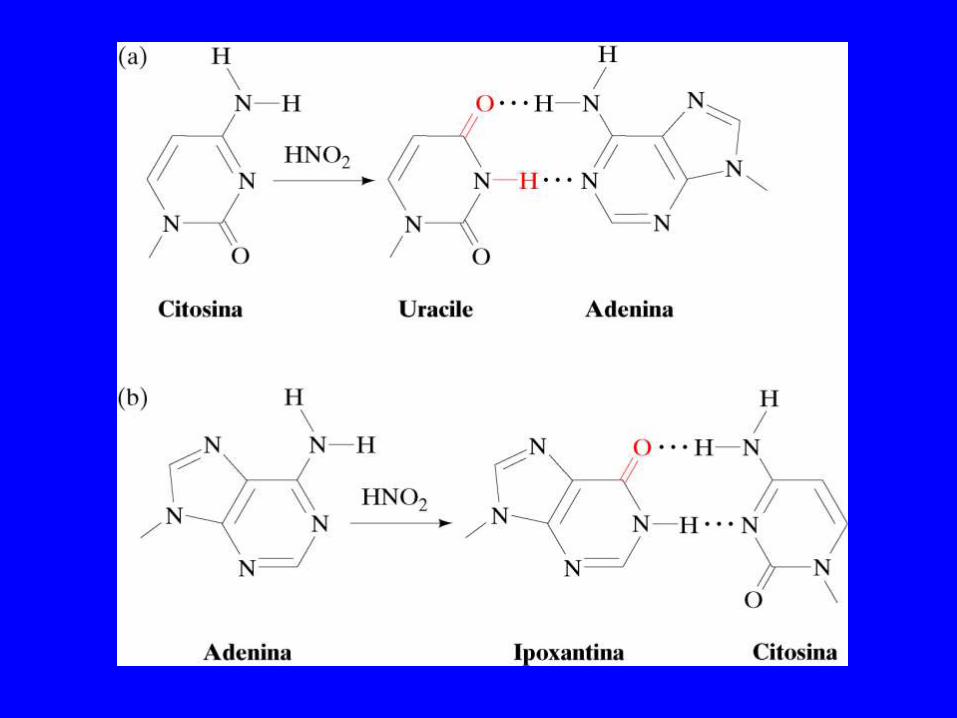
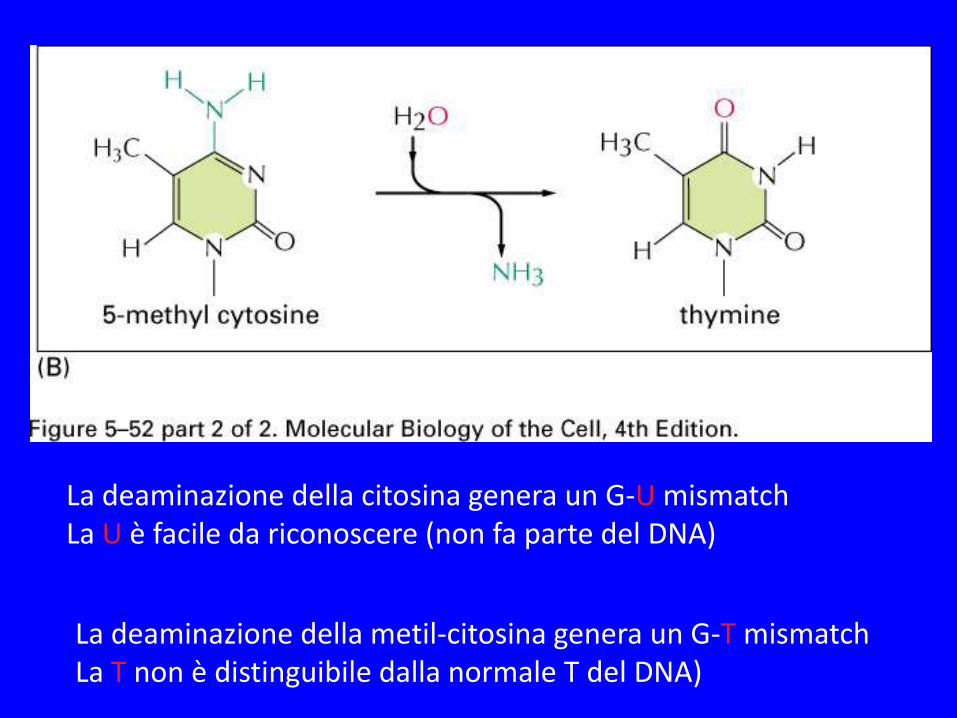
La deaminazione della citosina genera un G-U mismatch La U è facile da riconoscere (non fa parte del DNA) La deaminazione della metil-citosina genera un G-T mismatch La T non è distinguibile dalla normale T del DNA)

AGENTI CHE DANNEGGIANO IL DNA Radiazioni in alcuni intervalli di lunghezza d’onda
Radiazioni ionizzanti come i raggi gamma ed i raggi-X Radiazioni ultraviolette, specialmente i raggi UV-C (~260
nm) che sono fortemente assorbiti dal DNA e le radiazioni a lunghezza d’onda maggiore, i raggi UV-B, che possono attraversare la fascia di ozono atmosferico
I radicali dell’ossigeno, specie chimiche altamente reattive prodotte durante la normale respirazione cellulare o altre vie biochimiche
Agenti chimici ambientali
Molti idrocarburi, compresi quelli presenti nel fumo di sigaretta
Prodotti di microrganismi e piante, come le aflatossine presenti in alimenti ammuffiti
Agenti chimici usati in chemioterapia, in particolare nella lotta
contro i tumori

OSSIDAZIONE DELLE BASI
Guanine modification by hydroxyl radicals. Addition of hydroxyl radical to C-8 of guanine produces an adduct radical, which can be oxidized to 8-hydroxyguanine (8-oxoG) or reduced to give the ring-opened product 2,6-diamino-4-hydroxy-5-formamidopyrimidine (FAPγG).

OSSIDAZIONE DELLE BASI
Modified bases resulting from the attack of reactive oxygen species. Examples of the products that can be formed by hydroxyl radicals addition to position 8 of the purine ring (A) and to positions 5 or 6 of the pyrimidine ring (B).

MODIFICA CHIMICA DELLE BASI
Modified bases resulting from the attack of chemical reactive species. Reactive nitrogen species as N2O3 or HNO2 can promote deamination of DNA bases (guanine to xanthine and adenine to hypoxanthine) and conversion of guanine to oxanine, whereas 8-nitroguanine is formed by reaction of DNA with ONOO- (A). Examples of modified bases formed by the reaction of hypochlorous acid with DNA (B). Some of the modified bases resulting from alkylation (C).
Reactive nitrogen species
hypochlorous acid
alkylation
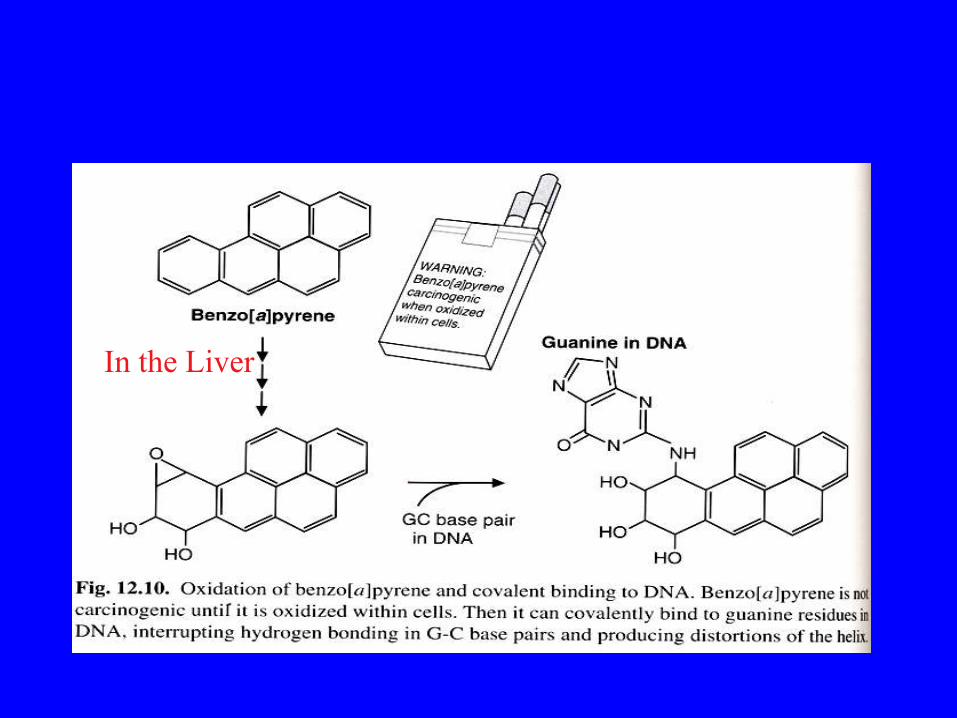
In the Liver

Aflatossine composti prodotti da alcune
specie di funghi.

DANNI DA RADIAZIONE UV
Dimeri di pirimidina ciclobutano CPDs (TT>TC,CT>CC)
Fotoprodotti Pirimidina (6-4) pirimidone 6-4PPs
44° bending
7°-9° bending

MUTAZIONI A CARICO
DELLE BASI
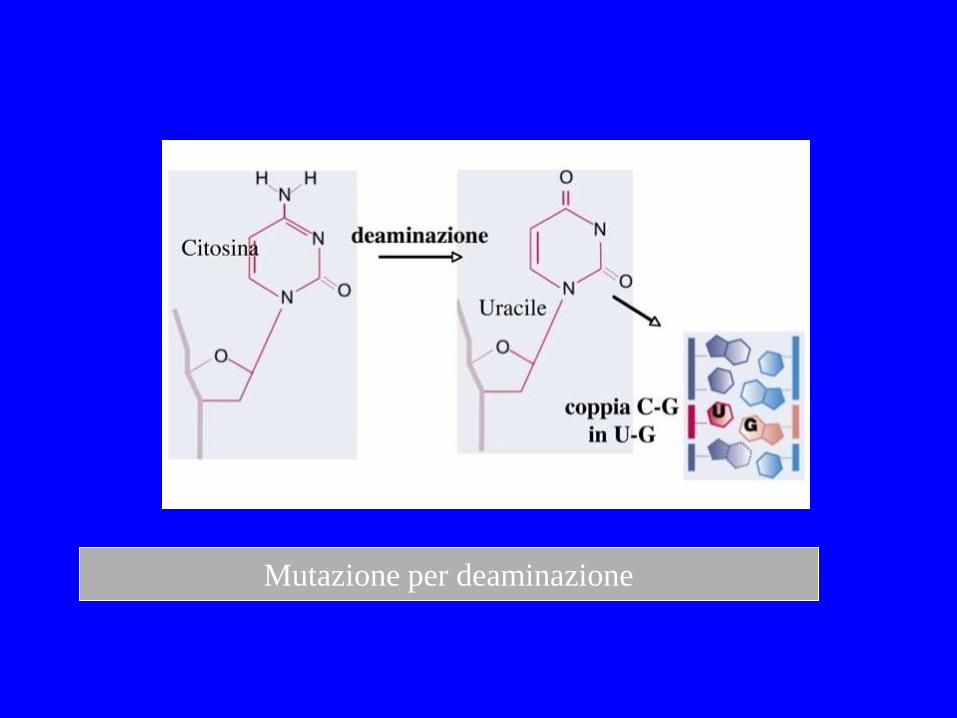
Mutazione per deaminazione

Mutagenesi da alchilazione
Guanosina Metil-Guanosina
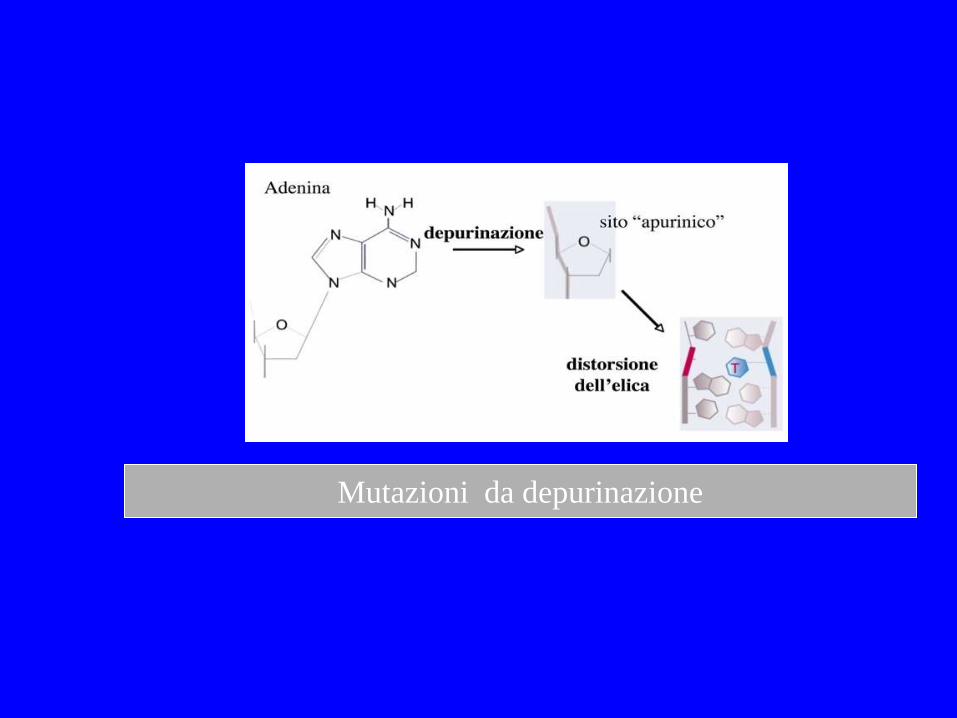
Mutazioni da depurinazione
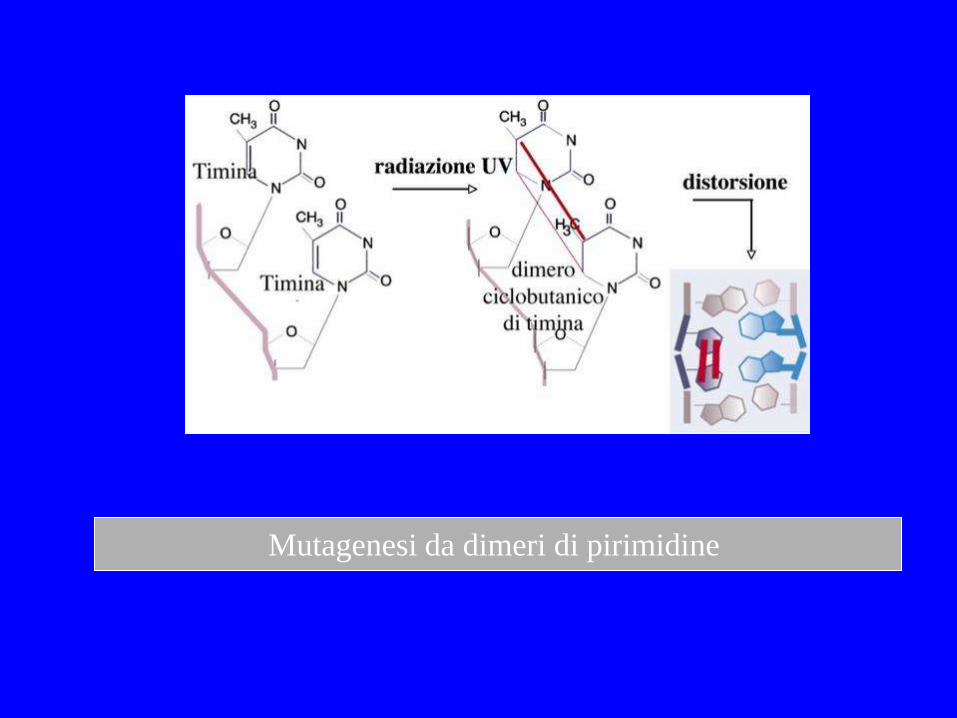
Mutagenesi da dimeri di pirimidine

MUTAZIONI PUNTIFORMI
• Appaiamenti sbagliati (mismatches) tra le basi a causa di errori da parte della DNA-polimerasi durante la replicazione del DNA
• Un esempio è l’incorporazione di U (presente nell’RNA) al posto di T
• Transizioni / Trasversioni
• Inserzioni / delezioni (cambio del frameshift)

EFFETTO DELLE MUTAZIONI
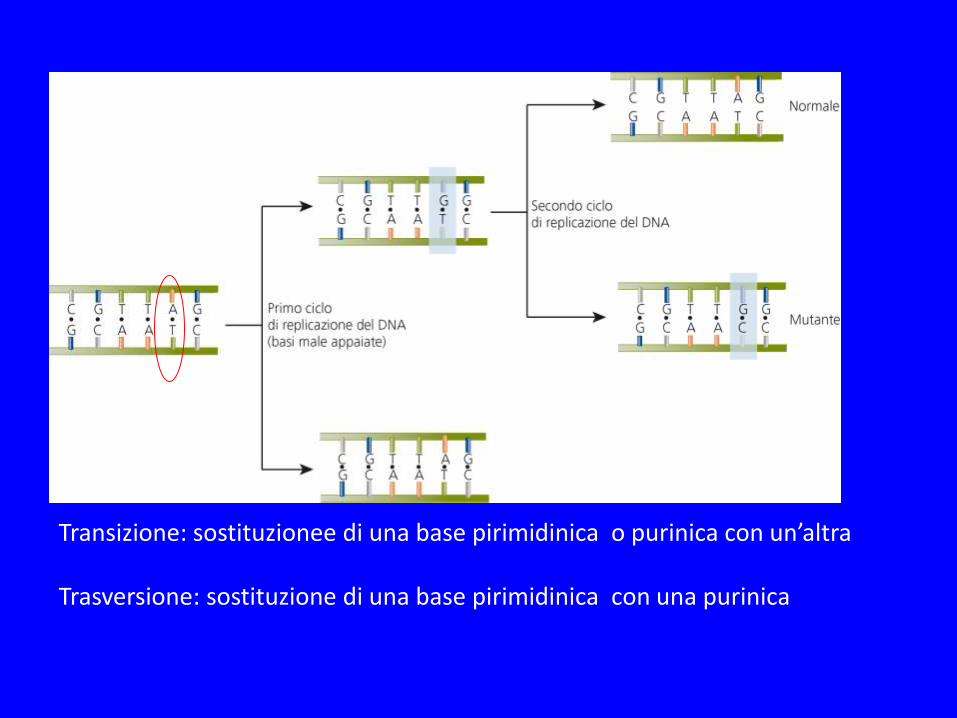
Transizione: sostituzionee di una base pirimidinica o purinica con un’altra Trasversione: sostituzione di una base pirimidinica con una purinica

TIPI DI DANNO A CARICO DEL DNA
• Rotture (breaks) nelle catene riboso-fosfato del DNA
• Possono essere limitate ad una delle due eliche (single-stranded break o SSB) oppure
• Interessare entrambe le eliche (double-stranded break o DSB) • Sono causate frequentemente dalle radiazioni ionizzanti, anche se
possono essere prodotte da alcuni agenti chimici
• Legami crociati (crosslinks) che si possono formare tra le basi
• Sulla stessa elica del DNA ("intrastrand") oppure • Su eliche opposte ("interstrand") • Molti agenti chemioterapici usati nella lotta contro il cancro
provocano legami crociati nel DNA
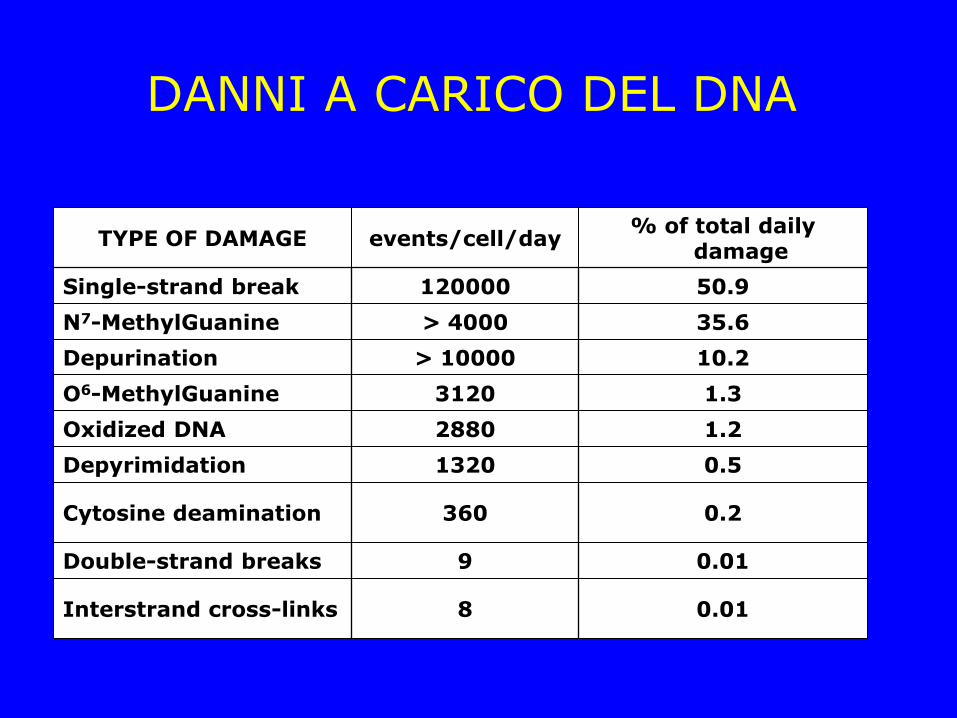
DANNI A CARICO DEL DNA
TYPE OF DAMAGE events/cell/day % of total daily
damage
Single-strand break 120000 50.9
N7-MethylGuanine > 4000 35.6
Depurination > 10000 10.2
O6-MethylGuanine 3120 1.3
Oxidized DNA 2880 1.2
Depyrimidation 1320 0.5
Cytosine deamination 360 0.2
Double-strand breaks 9 0.01
Interstrand cross-links 8 0.01

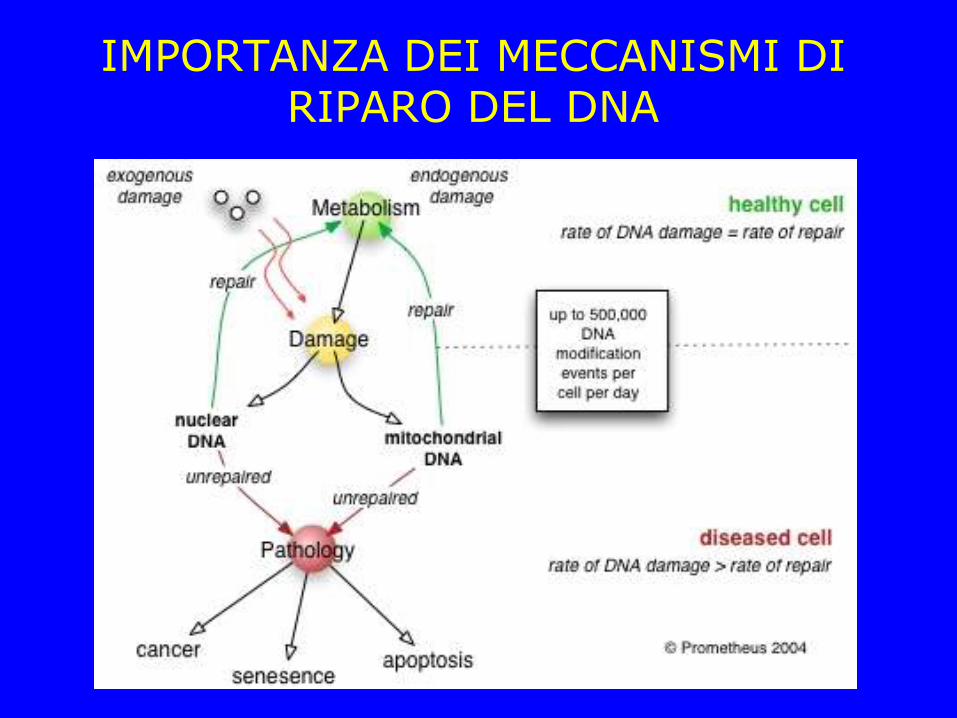
Il DNA cellulare è soggetto a molte alterazioni di natura chimica e fisica
Un fallimento nei sistemi di riparazione del DNA
produce mutazioni (cancerogenesi)
Più di 100 geni (anche nel più semplice organismo) sono coinvolti meccanismi di riparazione del DNA
La recente pubblicazione del genoma umano ha permesso di scoprire più di 130 geni coinvolti
IMPORTANZA DEI MECCANISMI DI RIPARO DEL DNA

MECCANISMI DI RIPARO DELLE BASI DANNEGGIATE
Le basi danneggiate o male appaiate possono essere riparate attraverso diversi meccanismi:
• Ripristino diretto del danno
• Riparo per escissione (Excision Repair), nel
quale la base o le basi danneggiate vengono rimosse e sostituite attraverso la nuova sintesi di DNA.
Esistono diversi meccanismi di riparo per escissione ognuno dei quali utilizza una serie di enzimi specializzati.
1. Base Excision Repair (BER) 2. Nucleotide Excision Repair (NER) 3. Mismatch Repair (MMR)

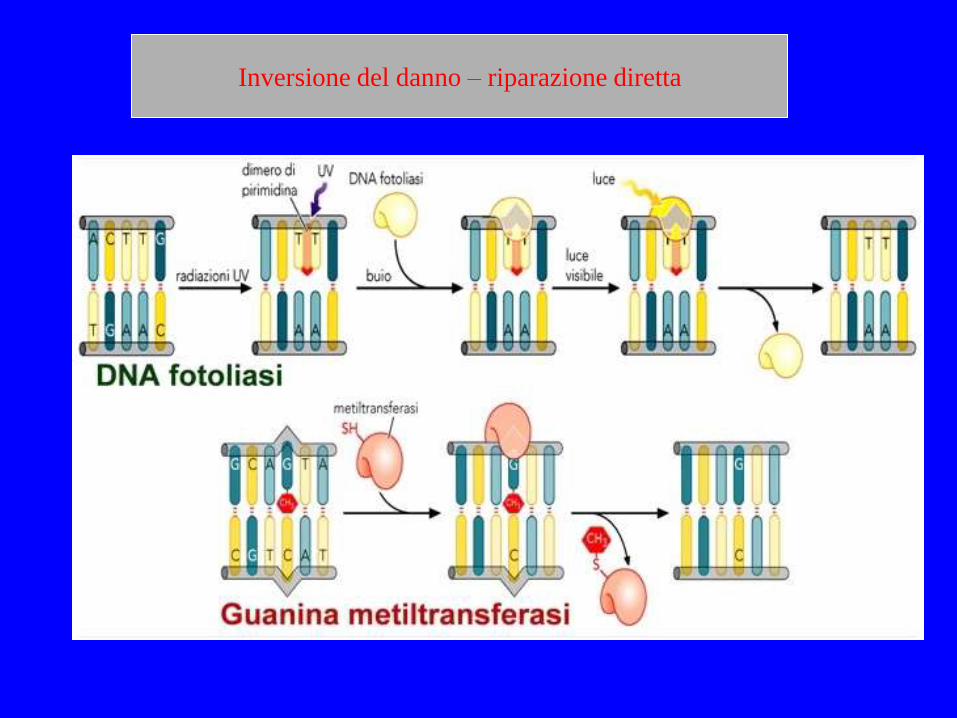
RIPRISTINO DIRETTO DEL DANNO
La causa più frequente di danno chimico a carico delle basi che portano ad una mutazione negli esseri viventi sono la formazione di dimeri di pirimidina l'addizione spontanea di un gruppo metilico alla C, seguita dalla deaminazione con formazione di T Questo tipo di danno può essere riparato da enzimi che agiscono senza il bisogno di rompere la catena riboso-fosfato del DNA. I dimeri di pirimidina possono essere risolti chimicamente attraverso una reazione di fotoriattivazione Alcuni dei gruppi metilici possono essere rimossi da una proteina codificata dal gene MGMT
Inversione del danno – riparazione diretta
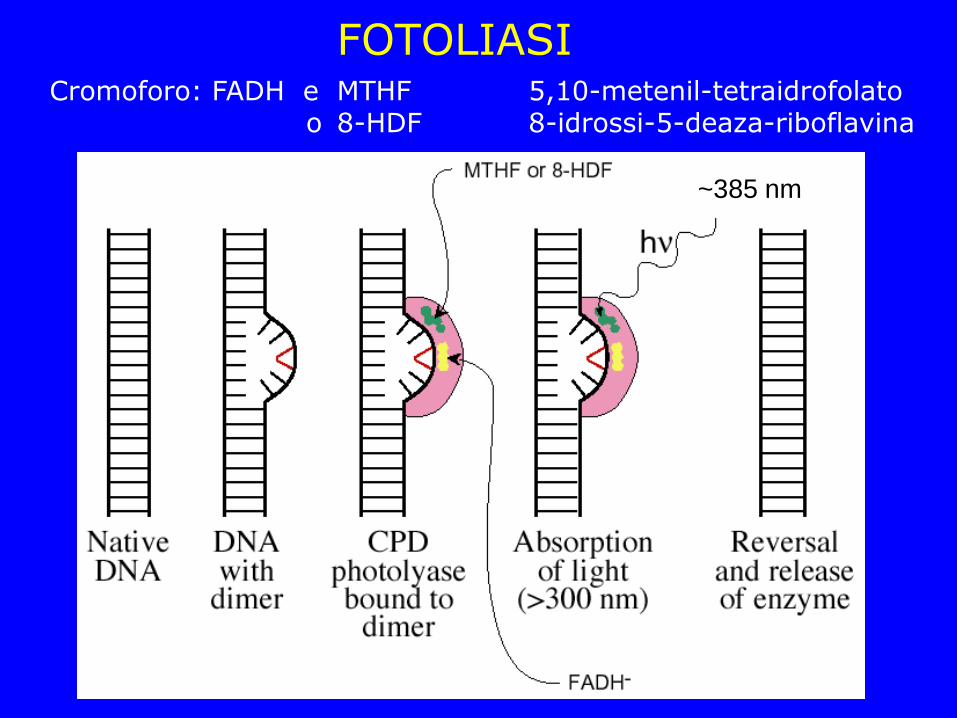
FOTOLIASI Cromoforo: FADH e MTHF 5,10-metenil-tetraidrofolato o 8-HDF 8-idrossi-5-deaza-riboflavina
~385 nm

MGMT O-6-methylguanine-DNA methyltransferase
La proteina possiede una cavità nella quale si inserisce la base alchilata. La metilazione rende inattiva la proteina che quindi può effettuare il riparo solamente una volta.
La O6-methylguanine-DNA methyltransferase o DNA-O6-methylguanine:protein-L-cysteine S-methyltransferase (EC 2.1.1.63) è un enzima di riparo unico presente in molti organismi e codificato dal gene ada. La sua azione rimuove l’addotto mutageno e carcinogenico O6-alchil-guanina presente nel DNA attraverso una reazione chimica in cui il gruppo alchilico viene trasferito dalla guanina ad un residuo di cisteina dell’enzima.

DIOSSIGENASI (ABH2)
Dealchilazione ossidativa di alcune basi attraverso l’azione dell’enzima ABH2 (AlkB)
α-chetoglutarato / O2
1-metil-adenina adenina formaldeide,
3-metil-citosina citosina succinato e CO2
α-chetoglutarato / O2
1-etil-adenina adenina acetaldeide,
succinato e CO2

LIMITI DEI MECCANISMI DI RIPRISTINO DIRETTO
La fotoriattivazione mediante radiazioni UV non rappresenta un valido meccanismo naturale. La MGMT non è un vero enzima in quanto non agisce in maniera catalitica.
Limiti dei meccanismi di ripristino diretto del danno: sono poco efficienti e dispendiosi. Ogni tipi di alterazione chimica a carico delle basi ha bisogno del suo specifico meccanismo di correzione.
Gli organismi viventi hanno sviluppato dei meccanismi più generali, capaci di correggere ogni sorta di danno chimico a carico delle basi attraverso un gruppo limitato di funzioni enzimatiche. Questi sono rappresentati dai meccanismi di riparazione mediante escissione.

BASE EXCISION REPAIR (BER)
Rappresenta uno dei sistemi di riparo più attivi Sono sufficienti 4 attività enzimatiche:
• DNA-Glicosidasi, endonucleasi, DNA-polimerasi e DNA-ligasi e 5 step di reazione • Eliminazione base danneggiata (glicosidasi) • Scissione elica e rimozione residuo abasico (endonucleasi) • Sintesi del DNA (DNA-polimerasi) • Riparo dell’interruzione sull’elica (DNA-ligasi)

SPECIFICITA’ DELLE N-GLICOSIDASI
• uracile
• idrossimetil-uracile
• 5-metil-citosina
• ipoxantina
• T:G timina mismatch
• A:C adenina mismatch
• 3-metil-adenina
• 7-metil-guanina
• 3-metil-guanina
• formamido-pirimidina
• glicol-timina
OGG1 8oxoG
UNG uracile
TDG Timina:G
SMUG1 uracile
NTH1 glicol-timina
MYH A:8oxoG
NEIL1 pirimidine ossidate
NEH2/NEIL2 metil-guanina
MED1 timina, uracile
AAG metilpurine, ipoxantina, 8-
8oxoG, xantina, and oxanina
Varie attività N-glicosidasi Basi modificate

DNA N-GLICOSIDASI UMANE
La porzione C-terminale delle glicosidasi eucarioriche è omologa a quelle procariotiche
Le glicosidasi eucariotiche contengono una porzione N-terminale che controlla la localizzazione intracellulare e le interazioni con altre proteine
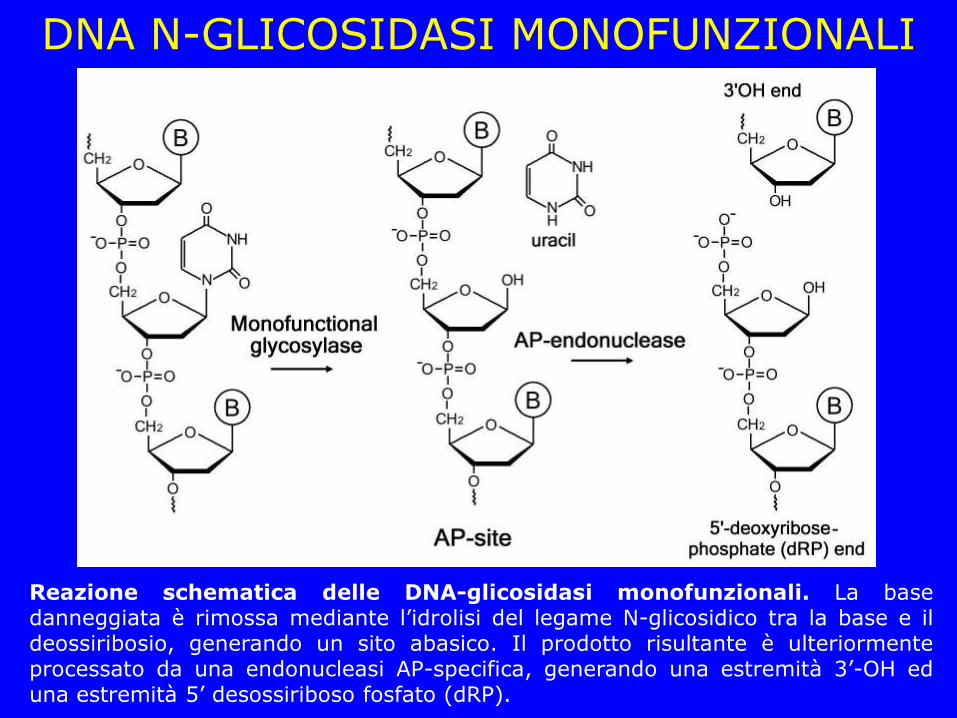
DNA N-GLICOSIDASI MONOFUNZIONALI
Reazione schematica delle DNA-glicosidasi monofunzionali. La base danneggiata è rimossa mediante l’idrolisi del legame N-glicosidico tra la base e il deossiribosio, generando un sito abasico. Il prodotto risultante è ulteriormente processato da una endonucleasi AP-specifica, generando una estremità 3’-OH ed una estremità 5’ desossiriboso fosfato (dRP).
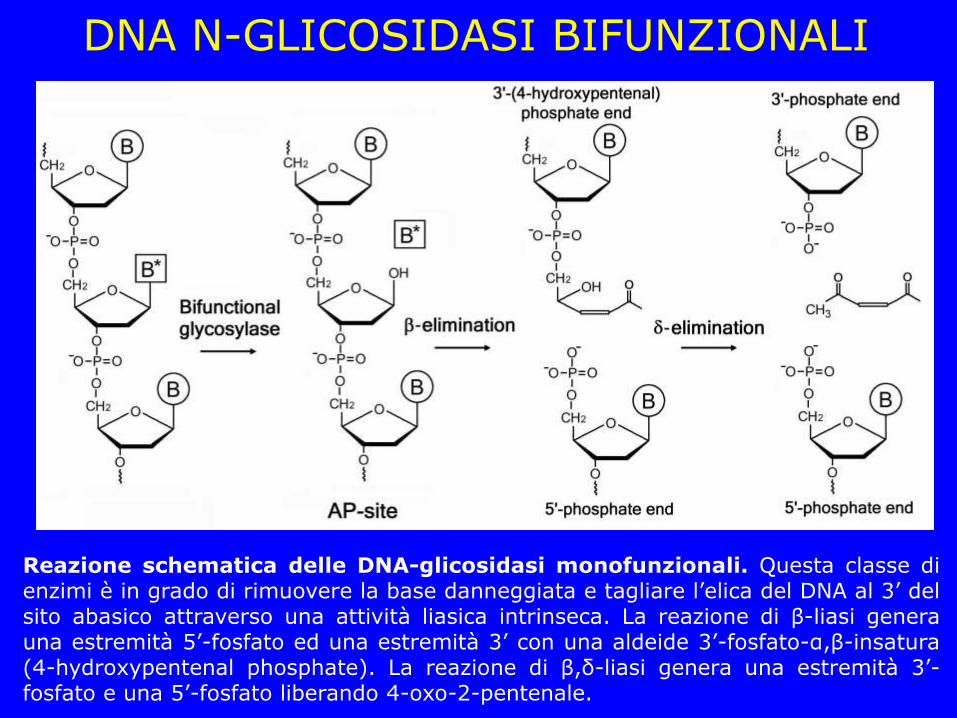
DNA N-GLICOSIDASI BIFUNZIONALI
Reazione schematica delle DNA-glicosidasi monofunzionali. Questa classe di enzimi è in grado di rimuovere la base danneggiata e tagliare l’elica del DNA al 3’ del sito abasico attraverso una attività liasica intrinseca. La reazione di β-liasi genera una estremità 5’-fosfato ed una estremità 3’ con una aldeide 3’-fosfato-α,β-insatura (4-hydroxypentenal phosphate). La reazione di β,δ-liasi genera una estremità 3’-fosfato e una 5’-fosfato liberando 4-oxo-2-pentenale.
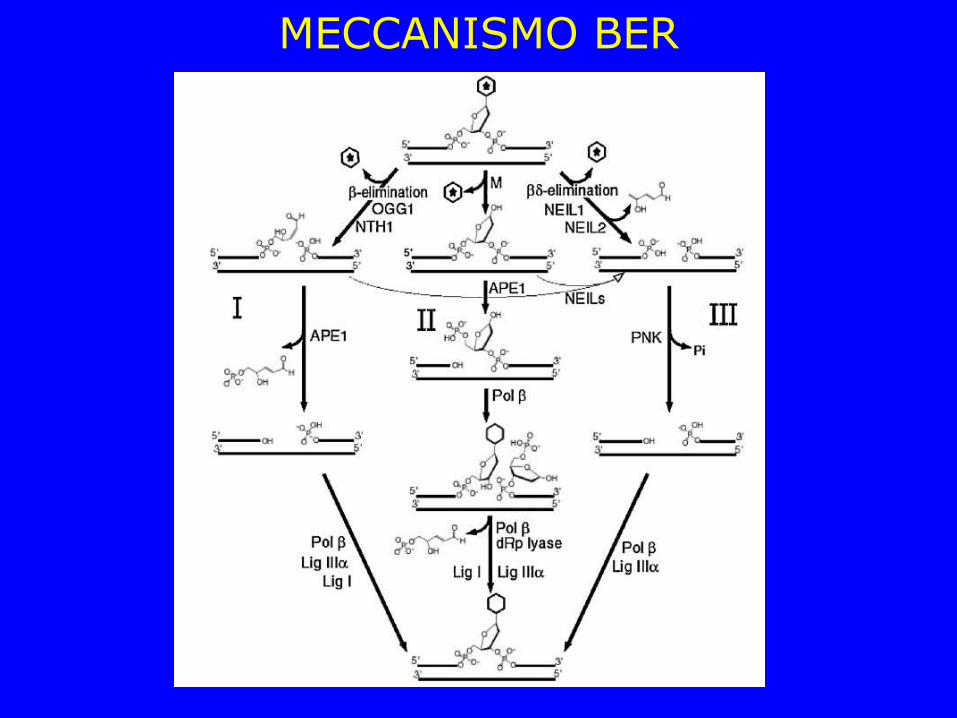
MECCANISMO BER
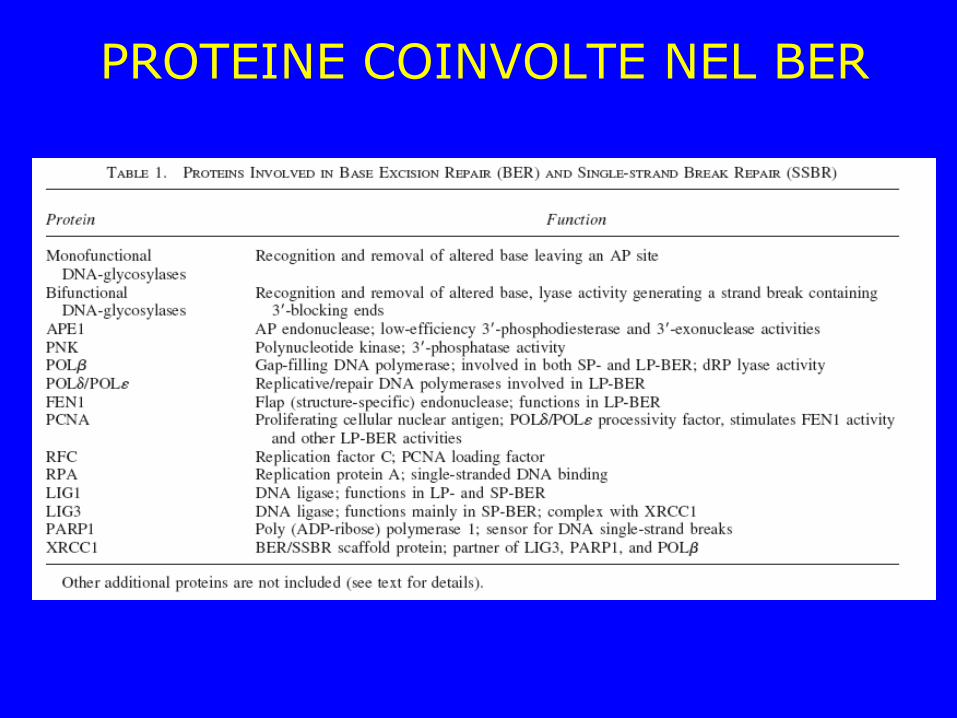
PROTEINE COINVOLTE NEL BER

NUCLEOTIDE EXCISION REPAIR (NER)
• Il sistema di riparo NER permette la rimozione di danni a carico del DNA che causano una distorsione nella doppia elica
• Il sistema di riparo consiste nella rimozione di un tratto di singola elica del DNA di 12-25 nucleotidi e contenente l’alterazione attraverso la scissione della catena riboso-fosfato alle estremità e la sintesi di un nuovo filamento
• Il sistema di riparo NER è estremamente flessibile in
quanto è in grado di riparare una moltitudine di lesioni, come la formazione di addotti delle basi, che in comune generano una alterazione o distorsione della doppia elica

LESIONI RIMOSSE DAL SISTEMA DI RIPARO NER
Le lesioni più rilevanti rimosse dal sistema di riparo NER sono:
• i dimeri di cis-sin-ciclobutano (CPDs) • i fotoprodotti pirimidina-(6-4)-pirimodone (6-4PPs)
Entrambi si formano tra residui adiacenti di pirimidine e sono le due principali lesione indotte dalla radaizione solare UV. Le lesioni 6-4PPs sebbene siano meno abbondananti distorcono maggiormente l’elica e vengono riparate 5 volte più velocemente.
Il sistema di riparo NER è in grado di rimuovere anche:
• addotti chimici voluminosi a carico delle basi come quelli prodotti dagli idrocarburi policiclici aromatici (fumo di sigaretta)
• cross-linking tra eliche indotto da agenti chemioterapici come il cis-platino o alcuni metaboliti cellulari
Il sistema di riparo NER può anche rimuovere danni minori a carico delle basi come quelli indotti dagli agenti alchilanti ed ossidanti e che non generano distorsioni nell’elica.
• NER come sistema di back-up del BER

MECCANISMO DEL SISTEMA DI RIPARO NER
Il sistema di riparo NER comprende 20-30 proteine e prevede: • Il riconoscimento iniziale del danno che può avvenire
in un punto qualsiasi del genoma (GG-NER)
associato all’evento trascrizionale (TR-NER)
• Il legame di un complesso multiproteico
• Incisioni sui lati 5’ e 3’ dell’elica danneggiata ad una distanza
di diversi nucleotidi dal sito iniziale
• La rimozione dell’elica danneggiata
• La sintesi di nuovo DNA da parte della DNA-polimerasi
• L’unione dei frammenti di DNA da parte della DNA-ligasi

PROTEINE DEL SISTEMA DI RIPARO NER
XPA: Ruolo chiave nell’organizzazione strutturale del compleso NER
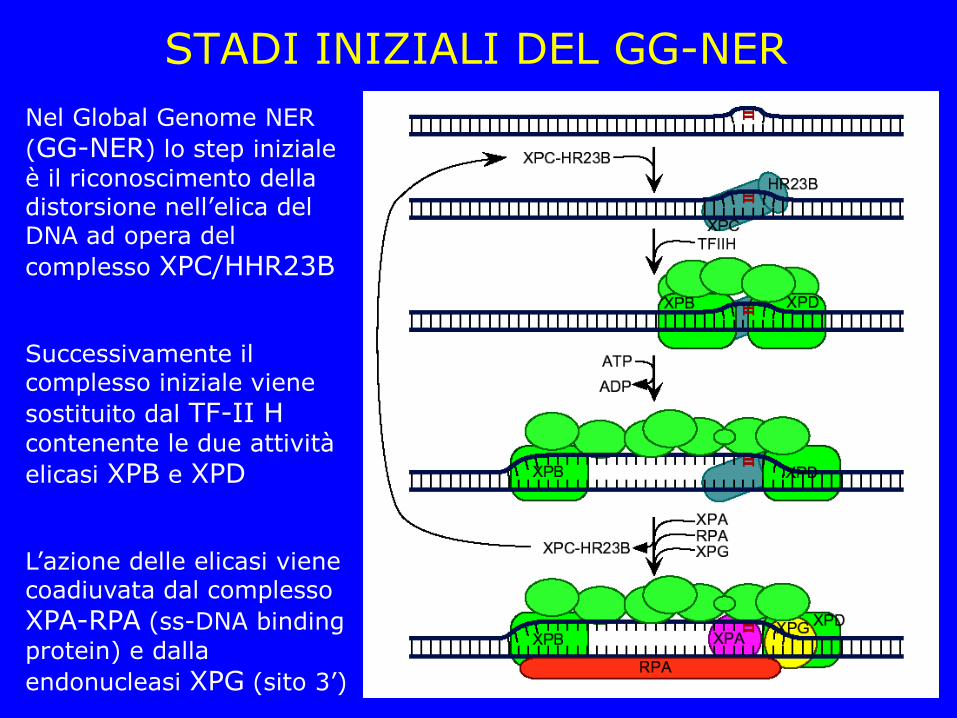
STADI INIZIALI DEL GG-NER
Nel Global Genome NER
(GG-NER) lo step iniziale
è il riconoscimento della distorsione nell’elica del DNA ad opera del
complesso XPC/HHR23B
Successivamente il complesso iniziale viene
sostituito dal TF-II H
contenente le due attività
elicasi XPB e XPD
L’azione delle elicasi viene coadiuvata dal complesso
XPA-RPA (ss-DNA binding
protein) e dalla
endonucleasi XPG (sito 3’)
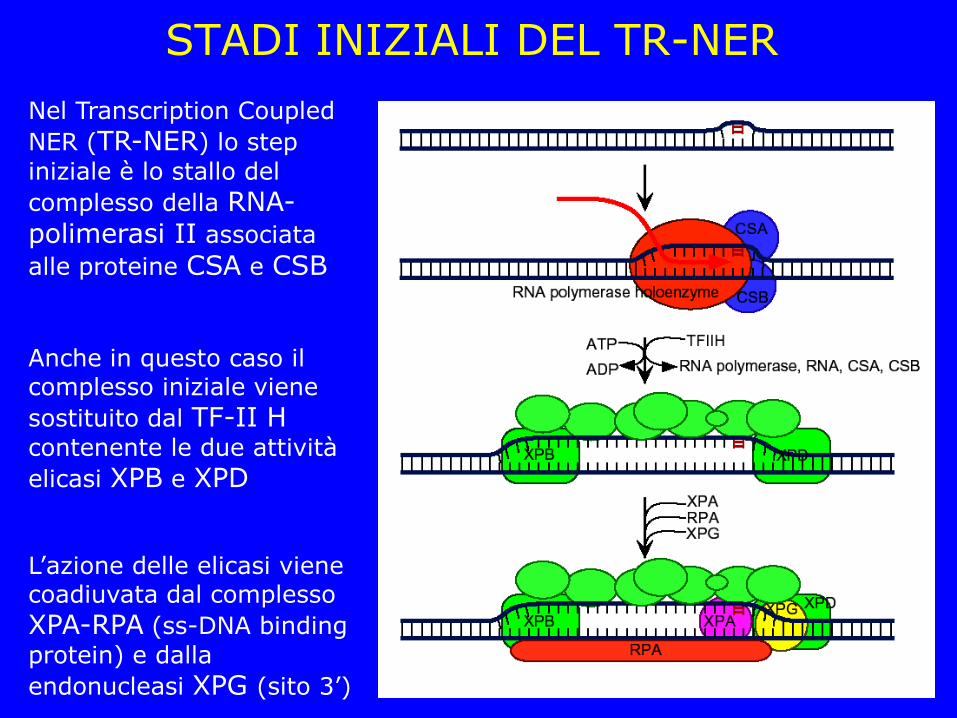
STADI INIZIALI DEL TR-NER
Nel Transcription Coupled
NER (TR-NER) lo step
iniziale è lo stallo del
complesso della RNA-polimerasi II associata
alle proteine CSA e CSB
Anche in questo caso il complesso iniziale viene
sostituito dal TF-II H
contenente le due attività
elicasi XPB e XPD
L’azione delle elicasi viene coadiuvata dal complesso
XPA-RPA (ss-DNA binding
protein) e dalla
endonucleasi XPG (sito 3’)
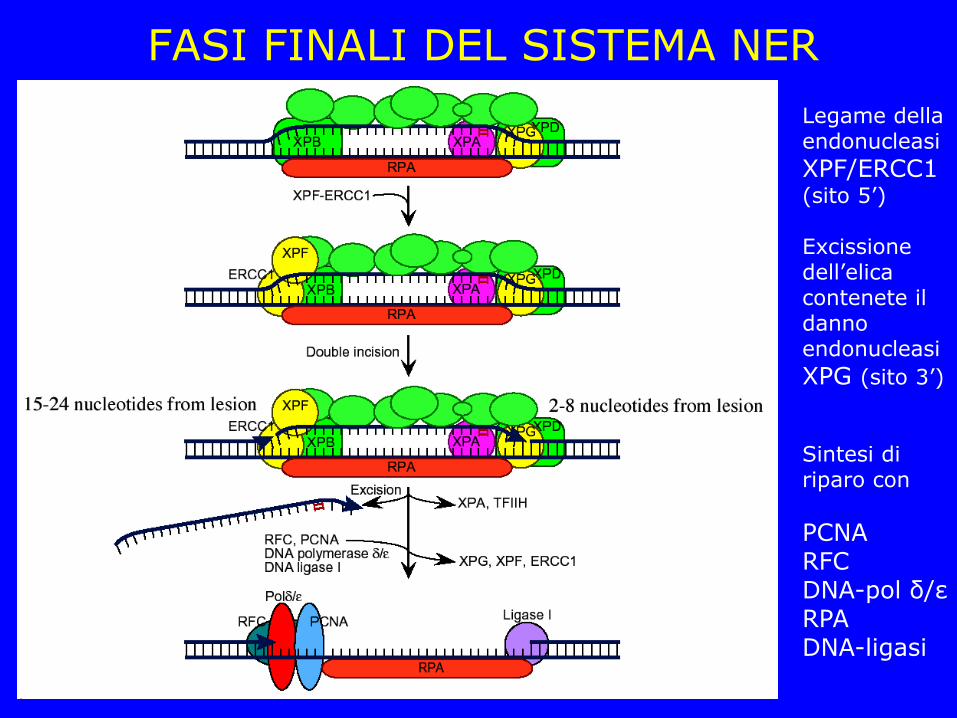
FASI FINALI DEL SISTEMA NER Legame della endonucleasi
XPF/ERCC1 (sito 5’) Excissione dell’elica contenete il danno endonucleasi
XPG (sito 3’)
Sintesi di riparo con
PCNA RFC DNA-pol δ/ε RPA DNA-ligasi

DIFETTI NEL SISTEMA DI RIPARO NER
• riconoscimento iniziale
• DNA-binding
• Elicasi
• Elicasi
• endonucleasi 3’ • endonucleasi 5’
Cockayne’s Syndrome Complementation Groups • CS-A (protein with multiple WD-40 repeats) nizio TC-NER • CS-B (DNA-dependent ATPase of the SNF2 family) inizio TC-NER

MISMATCH REPAIR (MMR)
Meccanismo che riconosce danni a carico del DNA che causano alterazione nella doppia elica del DNA (mismatch tra basi e piccoli loop di ssDNA) Per il riparo viene utilizzata l’elica parentale (si assume che il danno sia avvenuto durante l’ultima replicazione) Costituito da due sistemi enzimatici
• Sistema enzimatico che riconosce il mismatch • Sistema che riconosce l’elica parentale e provoca una
incisione nell’elica neosintetizzata nelle vicinanze del sito danneggiato
Il meccanismo viene completato dall’azione della DNA-polimerasi che sintetizza il nuovo filamento di DNA su stampo del filamento parentale
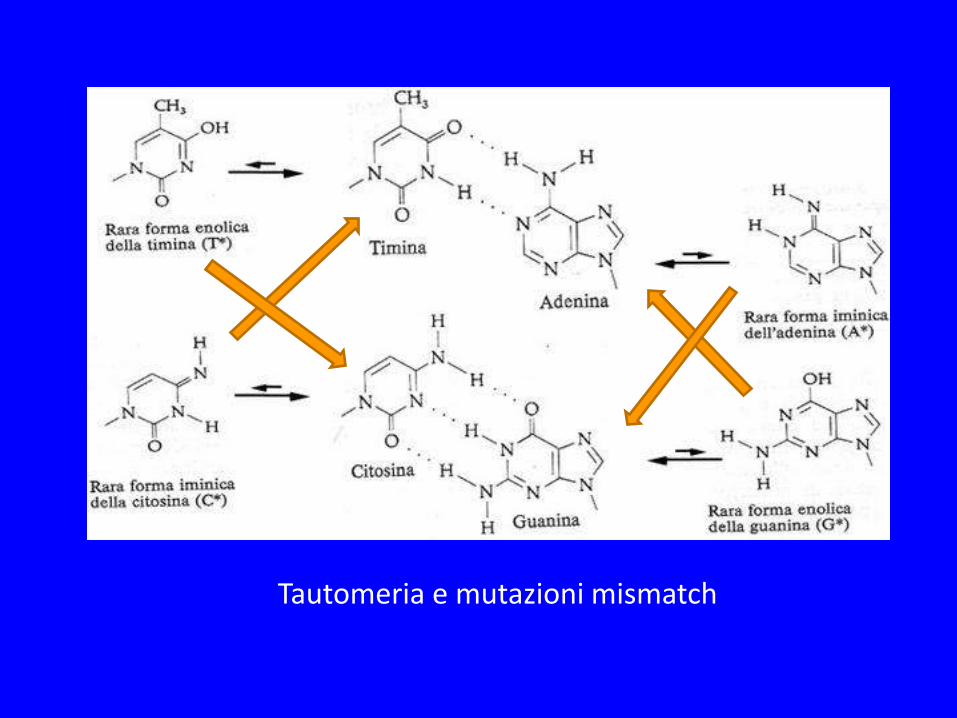
Tautomeria e mutazioni mismatch

SISTEMA DI RIPARO MMR IN E.Coli
MutS riconosce i punti di mismatch MutL stabilizza il complesso MutH riconosce il DNA emimetilato DNA metilato nei siti GATC La metilazione avviene in ritardo rispetto alla sintesi del DNA per cui è possibile riconoscere l’elica di nuova sintesi
MutS/MutL attiva MutH che taglia il DNA sull’elica non metilata
Una attività elicasi (UvrD o Helicase II) svolge l’elica dal sito di taglio sino al punto di mismatch mentre una attività esonucleasi (5’ o 3’ in base al punto di taglio) rimuove il singolo filamento
La DNA pol-III sintetizza il nuovo filamento e la DNA-ligasi unisce l’interruzione
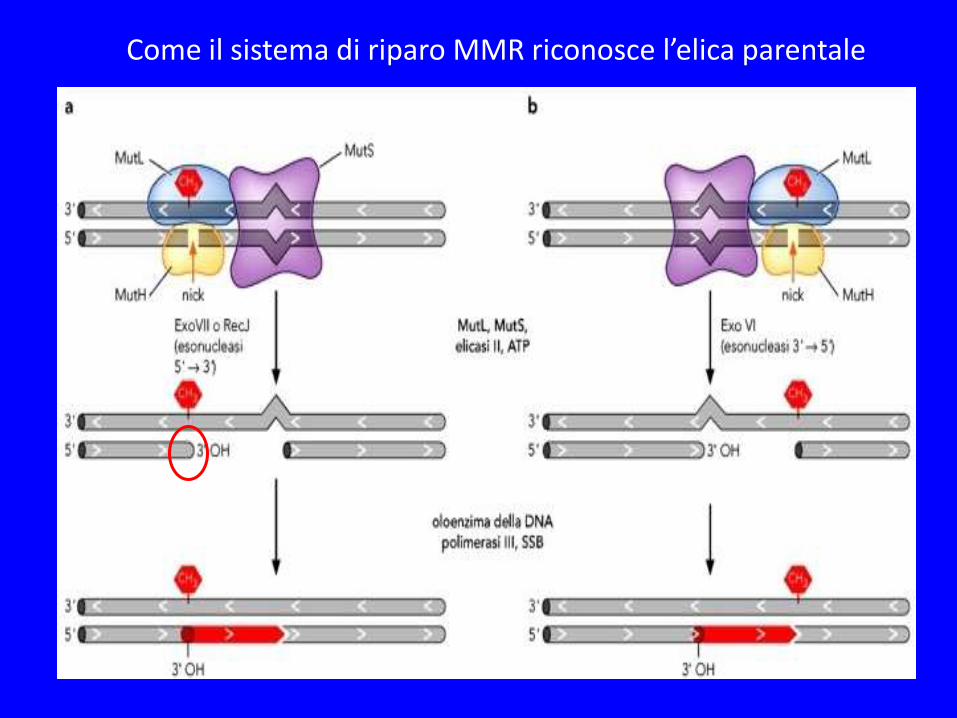
Come il sistema di riparo MMR riconosce l’elica parentale

GENI COINVOLTI NEL SISTEMA MMR
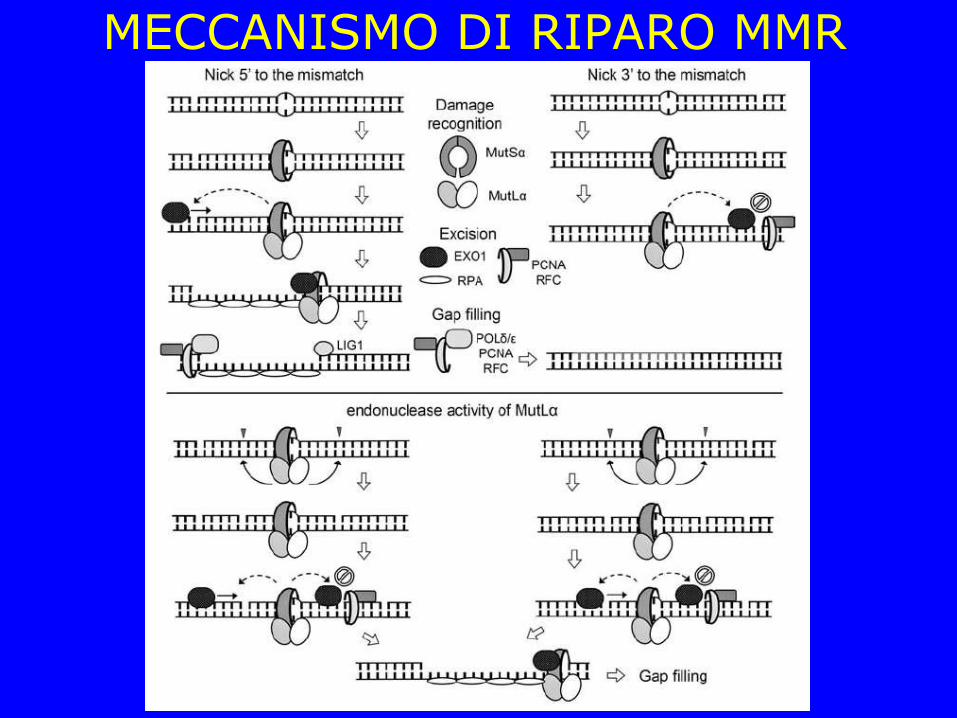
MECCANISMO DI RIPARO MMR

Correzione di scivolamenti nella replicazione di regioni con ripetizioni in tandem (DNA microsatellite) Tendenza della polimerasi a scivolare sullo stampo copiando più volte la stessa regione (causa di polimorfismo e di patologie: distrofia muscolare, X fragile, malattia di Huntington estensione della tripletta CAG che codifica per la glutamina)

DIFETTI NEL SISTEMA DI RIPARO MMR
Hereditary non-polyposis colon cancer (HNPCC) • forma di cancro al colon caratterizzata dalla insorgenza in età precoce
e dalla ereditarietà autosomica dominante • frequentemente associata con difetti nei geni codificanti MSH2 (35%
dei casi) e MLH1 (circa 60% dei casi) • MSH2 e MLH1 sono essenziali per la formazione di eterodimeri
funzionali di MutS e MutL • saltuariamente associata con difetti in altri geni del sistema di riparo
MMR (MSH6, PMS2 e PMS1), conseguenza della ridondanza di questi ultimi geni che li rende singolarmente non essenziali per il MMR
La HNPCC è quasi sempre associata con difetti nella riparazione del DNA evidenziate nella "instabilità del DNA microsatellite" (microsatellite instability o MIN) rappresentata da variazioni nel numero delle unità di sequenza ripetute [An, (GGC)n, (CA)n] In molti casi di cancro al colon associati con MIN ma senza alterazioni a carico dei geni del sistema MMR, è stata osservata una metilazione estesa del promotore di MLH1 che lo silenzia
• probabilmente tutti i casi di cancro al colon associati a MIN sono dovuti a difetti nel sistema di riparo MMR
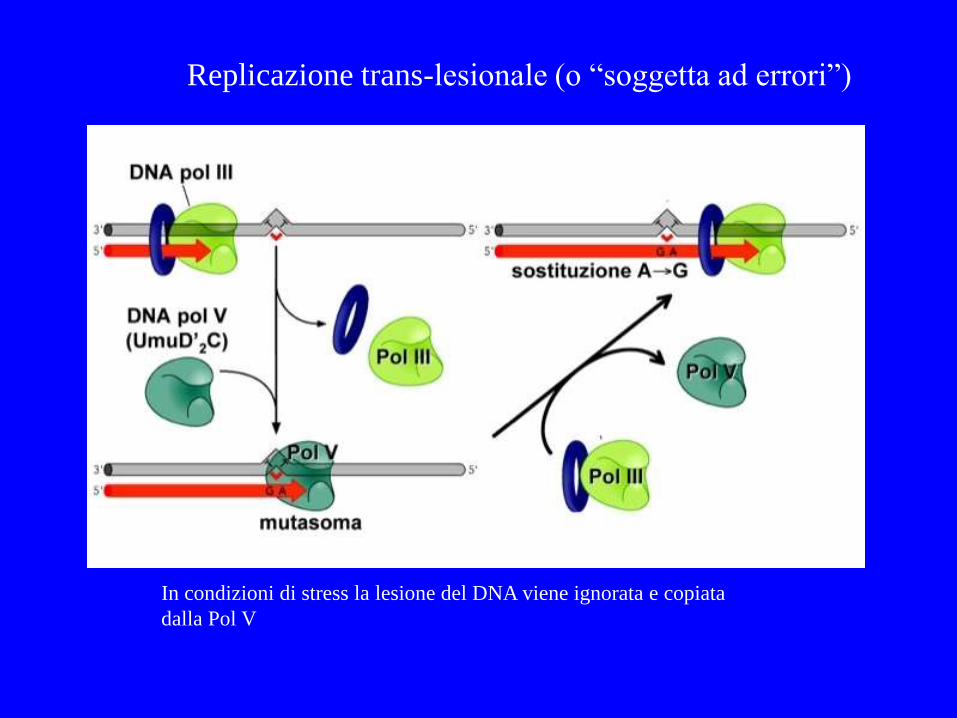
In condizioni di stress la lesione del DNA viene ignorata e copiata
dalla Pol V
Replicazione trans-lesionale (o “soggetta ad errori”)
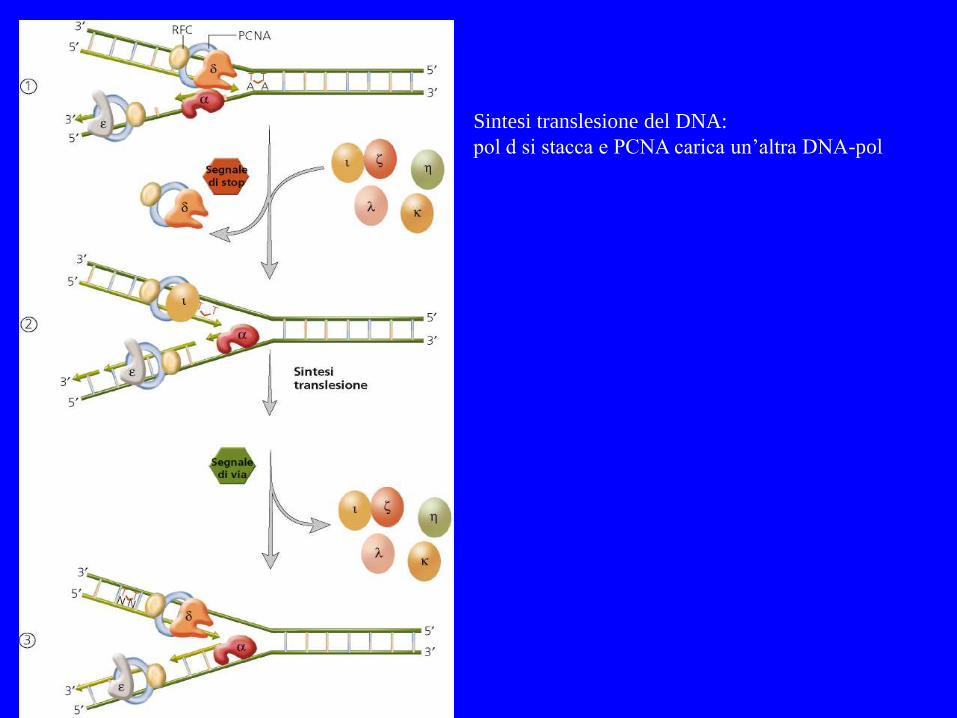
Sintesi translesione del DNA:
pol d si stacca e PCNA carica un’altra DNA-pol

TRANSLESIONAL SYNTHESIS
Translesional o bypass synthesis: DNA polimerasi alternative a bassa processività Possono essere sia error free che error prone Ad esempio Pol incorpora correttamente dA in presenza di CPD (dimeri timina)
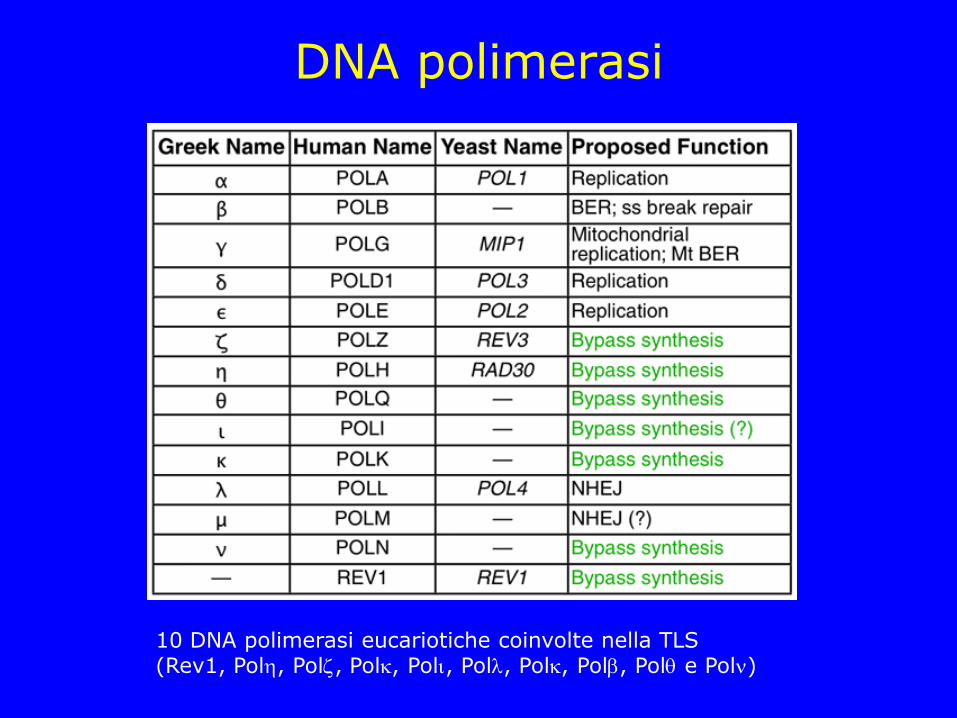
DNA polimerasi
10 DNA polimerasi eucariotiche coinvolte nella TLS (Rev1, Pol, Pol, Pol, Pol, Pol, Pol, Pol, Pol e Pol)

STRAND-BREAK REPAIR Le radiazioni ionizzanti ed alcuni agenti chimici possono produrre rotture che riguardano
• la singola elica (single-strand breaks o SSBs)
• la doppia elica (double-strand breaks o DSBs)
I meccanismi di riparo per gli strand-breaks sono essenziali per la sopravvivenza della cellula Le rotture nella singola elica vengono riparate attraverso un solo tipo di meccanismo
• Single-strand breaks repair (SSBR)
Le rotture nella doppia elica possono essere riparate attraverso due tipi di meccanismo
• Double-strand breaks repair by non-homologous end joining (NHEJ) • Double-strand breaks repair by homologous recombination (HR)

SINGLE-STRAND BREAKS REPAIR (SSBR) Le rotture nella singola elica vengono riparate attraverso un meccanismo comune al sistema di riparo BER (short-patch BER)
PARP riconosce le interruzioni XRCC1 ha le funzioni di proteina scaffold La PNK Polinucleotide kinasi/3’-fosfatasi prepara le estremità La DNA polimerasi β riempie l’interruzione La DNA ligasi 3 unisce le estremità
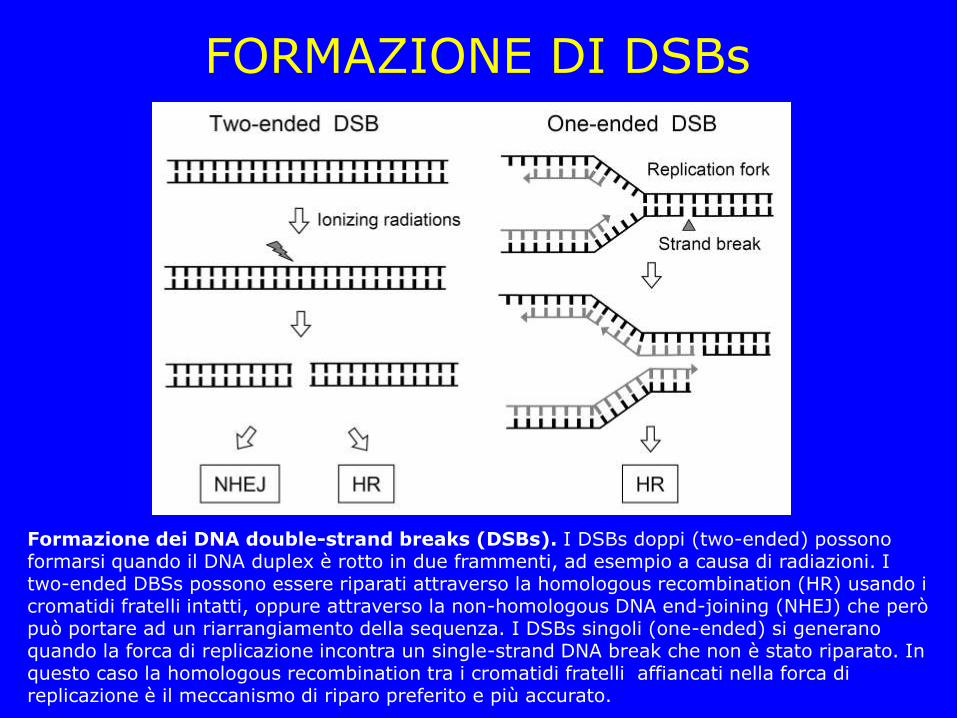
FORMAZIONE DI DSBs
Formazione dei DNA double-strand breaks (DSBs). I DSBs doppi (two-ended) possono formarsi quando il DNA duplex è rotto in due frammenti, ad esempio a causa di radiazioni. I two-ended DBSs possono essere riparati attraverso la homologous recombination (HR) usando i cromatidi fratelli intatti, oppure attraverso la non-homologous DNA end-joining (NHEJ) che però può portare ad un riarrangiamento della sequenza. I DSBs singoli (one-ended) si generano quando la forca di replicazione incontra un single-strand DNA break che non è stato riparato. In questo caso la homologous recombination tra i cromatidi fratelli affiancati nella forca di replicazione è il meccanismo di riparo preferito e più accurato.
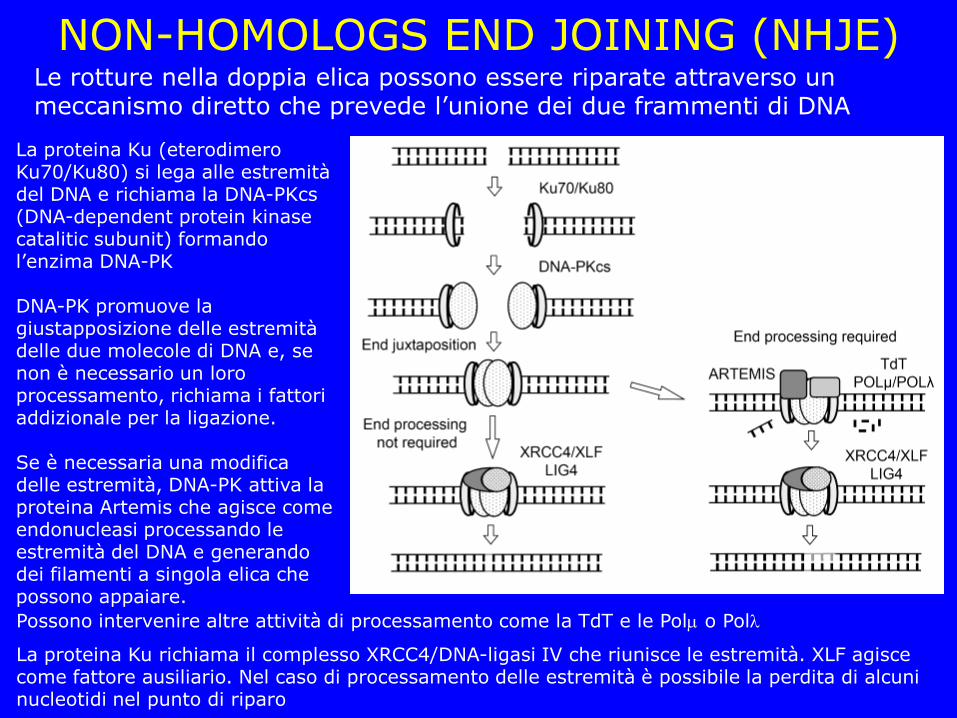
NON-HOMOLOGS END JOINING (NHJE) Le rotture nella doppia elica possono essere riparate attraverso un meccanismo diretto che prevede l’unione dei due frammenti di DNA
La proteina Ku (eterodimero Ku70/Ku80) si lega alle estremità del DNA e richiama la DNA-PKcs (DNA-dependent protein kinase catalitic subunit) formando l’enzima DNA-PK DNA-PK promuove la giustapposizione delle estremità delle due molecole di DNA e, se non è necessario un loro processamento, richiama i fattori addizionale per la ligazione. Se è necessaria una modifica delle estremità, DNA-PK attiva la proteina Artemis che agisce come endonucleasi processando le estremità del DNA e generando dei filamenti a singola elica che possono appaiare.
Possono intervenire altre attività di processamento come la TdT e le Pol o Pol
La proteina Ku richiama il complesso XRCC4/DNA-ligasi IV che riunisce le estremità. XLF agisce come fattore ausiliario. Nel caso di processamento delle estremità è possibile la perdita di alcuni nucleotidi nel punto di riparo
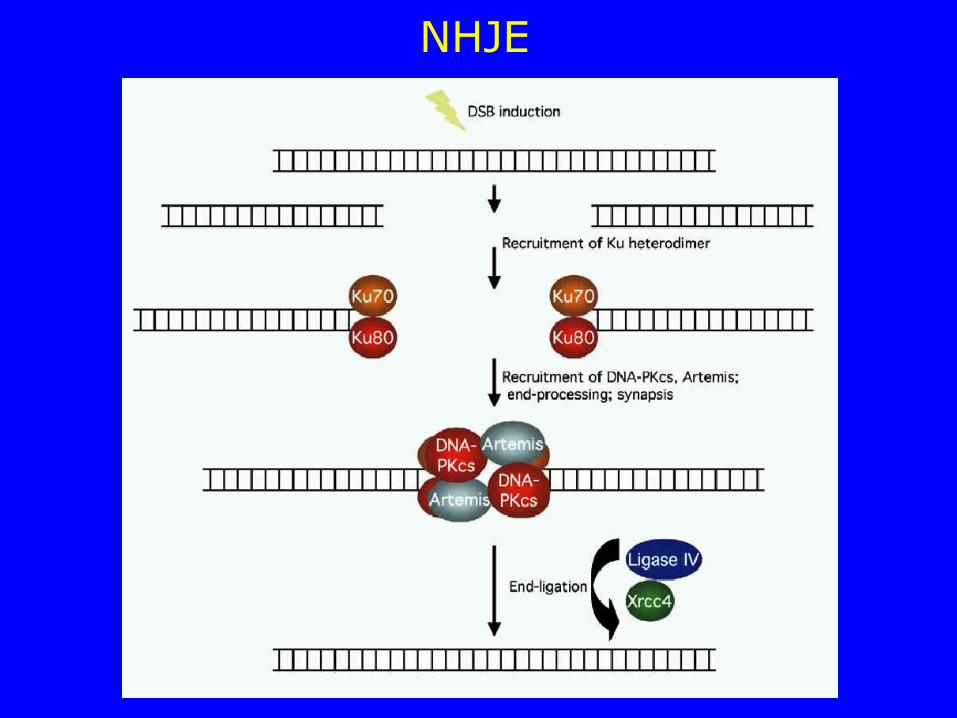
NHJE

LINFOMA DI BURKITT Il linfoma di Burkitt è un tumore di solido costituito da linfociti B. Nella parte maggior parte dei casi di linfoma di Burkitt (>90%) si osserva una traslocazione del proto-oncogene c-myc dalla sua posizione normale sul cromosoma 8 ad una posizione adiacente all’enhancer del gene della catena pesante delle immunoglobuline G situato sul cromosoma 14
C-myc è un fattore essenziale nella regolazione della mitosi cellulare e la sua sovraespressione rende la cellula tumorale
Il rischio di traslocazioni che riguardano le regioni codificanti i geni degli anticorpi è elevato in quanto durante la maturazione e la sintesi delle catene degli anticorpi avvengono riarrangiamenti a carico della sequenza del DNA. Cromosoma 14 (catene pesanti), cromosoma 2 (catene leggere kappa) e cromosoma 22 (catene leggere lambda).

LEUCEMIA MIELOIDE CRONICA (CML) Le leucemia sono proliferazioni incontrollate dei leucociti e la leucemia mieloide cronica (CML) colpisce cellule staminali emopoietiche precursori dei granulociti e dei macrofagi. Molti casi di CML sono associati ad una traslocazione tra la regione terminale del cromosoma 9 ed il cromosoma 22 con la formazione di un cromosoma 22 piccolo detto cromosoma Philadephia
Il proto-oncogene c-abl si fonde con il gene bcr. Il prodotto BCR-ABL è una tirosina kinasi costitutivamente attivata che stimola risposte cellulari normalmente sotto il controllo del PDGF (platelet-derived growth factor) provocando una mitosi incontrollata ed una inibizione dell’apoptosi
L’imatinib mesilate (Gleevec®) è un inibitore dell’attività kinasi di ABL

LEUCEMIA DELLE CELLULE-B Le cellule B hanno un tempo di vita limitato e vanno spontaneamente incontro ad apoptosi dopo che è terminata la risposta immunitaria
Una traslocazione tra il cromosoma 18 ed il cromosoma 14 porta il gene bcl-2 adiacente all’enhancer del gene della catena pesante delle immunoglobuline presente sul cromosoma 14
BCL-2 è una proteina ad attività anti-apoptotica.
La sovraespressione del gene bcl-2 protegge la cellula nei confronti dell’apotosi prevenendo l’azione delle caspasi e rendondo la cellula tumorale
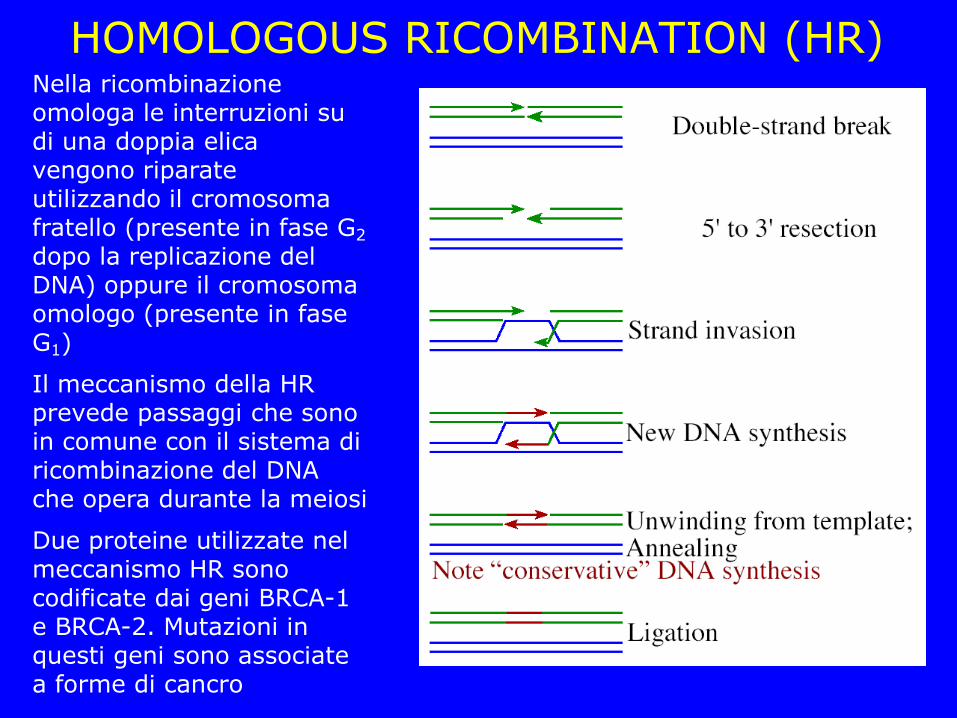
HOMOLOGOUS RICOMBINATION (HR) Nella ricombinazione omologa le interruzioni su di una doppia elica vengono riparate utilizzando il cromosoma fratello (presente in fase G2 dopo la replicazione del DNA) oppure il cromosoma omologo (presente in fase G1)
Il meccanismo della HR prevede passaggi che sono in comune con il sistema di ricombinazione del DNA che opera durante la meiosi
Due proteine utilizzate nel meccanismo HR sono codificate dai geni BRCA-1 e BRCA-2. Mutazioni in questi geni sono associate a forme di cancro
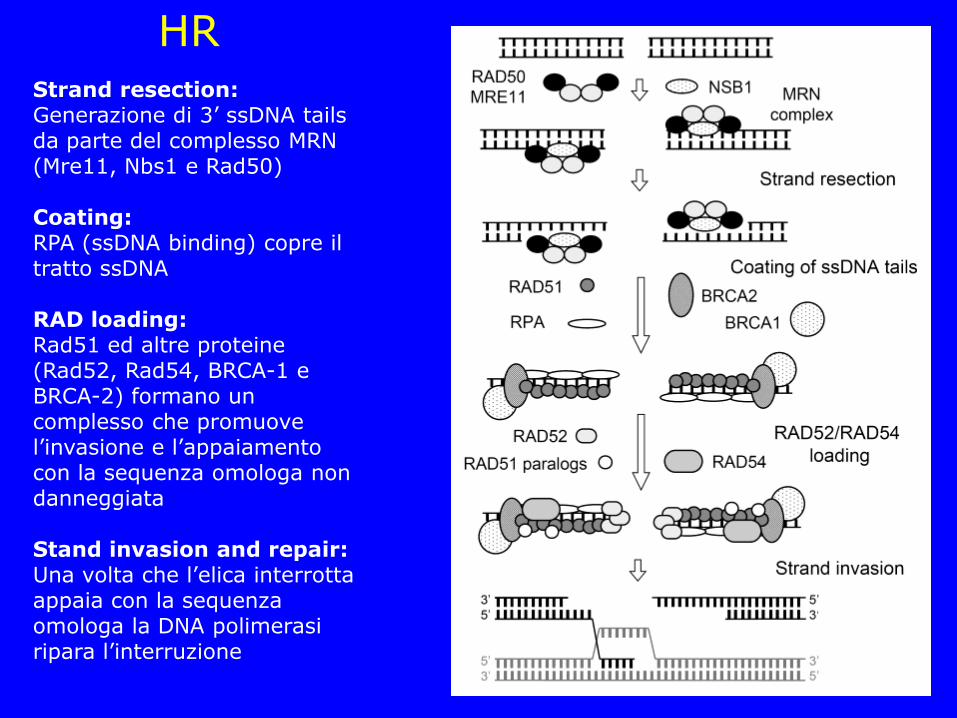
HR Strand resection: Generazione di 3’ ssDNA tails da parte del complesso MRN (Mre11, Nbs1 e Rad50) Coating: RPA (ssDNA binding) copre il tratto ssDNA RAD loading: Rad51 ed altre proteine (Rad52, Rad54, BRCA-1 e BRCA-2) formano un complesso che promuove l’invasione e l’appaiamento con la sequenza omologa non danneggiata Stand invasion and repair: Una volta che l’elica interrotta appaia con la sequenza omologa la DNA polimerasi ripara l’interruzione
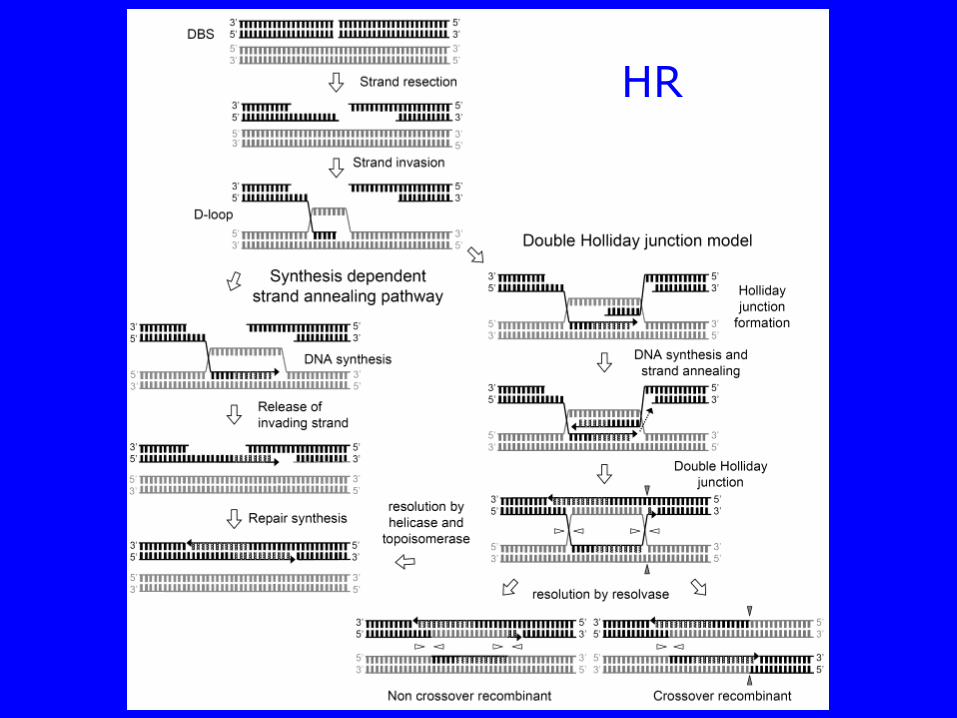
HR
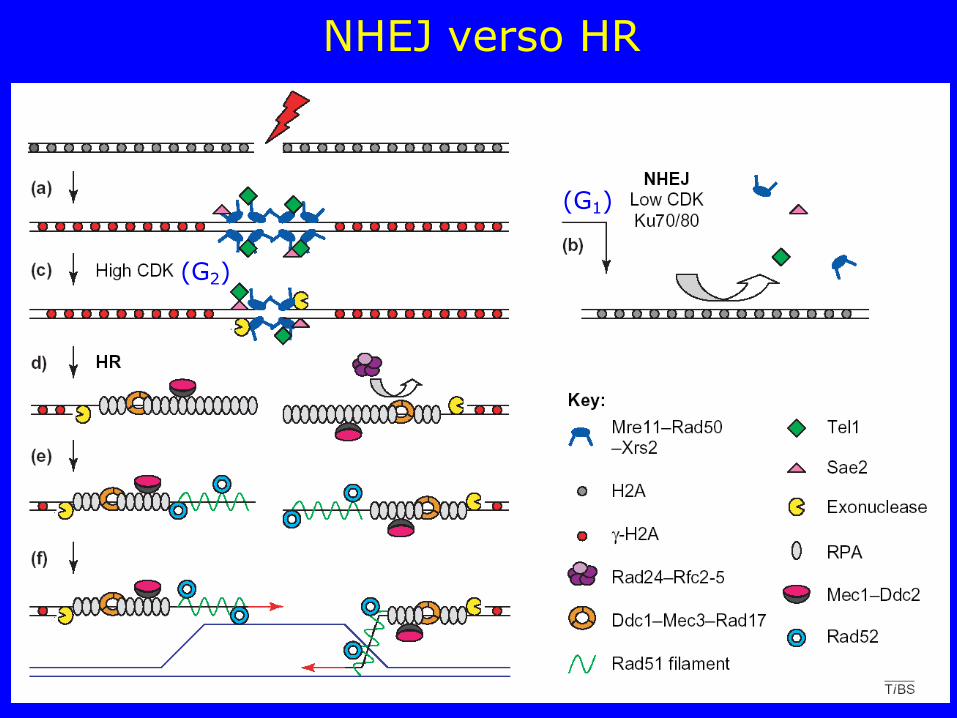
NHEJ verso HR
(G1)
(G2)

NHEJ verso HR
Modello di riparazione delle double-strand breaks (DSB): NHEJ verso HR
a) La formazione di una DSB richiama le componenti di controllo iniziali: il complesso MRN (MRX nel lievito), Tel1 e Sae2. La fosforilazione dell’istone H2a (γ-H2a) si estende intorno al sito di rottura
b) Il sistema di riparo NHEJ, che agisce tramite Ku70 o Ku80, è la via di riparazione preferita quando l'attività delle CDK è bassa (fase G1)
c) Quando l'attività delle CDK è alta (fase G2), vengono generate le estremità 3’ ssDNA ed il complesso MRN/MRX, Tel1 e Sae2 cominciano a dissociare
d) RPA lega le ssDNA e recluta Ddc2-Mec1 mentre i complessi Rad24-Rfc2-5 e Ddc1-Mec3-Rad17 (9-9-1 complesso) si associano al sito della rottura
e) Rad52 e Rad51 sono reclutati nel sito di rottura, spostano RPA, e avviano l'invasione e l’appaiamento con la sequenza di DNA omologa

FARMACI Antimetaboliti
• 5-fluorouracil • 6-tioguanine
Alchilanti • N-methyl-N-nitrosourea (MNU) • N-methyl-N’-nitro-N-nitrosoguanidine (MNNG) • 1-(2-chloroethyl)-3-cyclohexyl-1-nitrosourea (CCNU) • Mitomycin C
Platino • Cis-diamminedichloro-platinum • Oxaliplatin
Inibitori topoisomerasi • Camptotecin (CPT) • Etoposide (ETP)
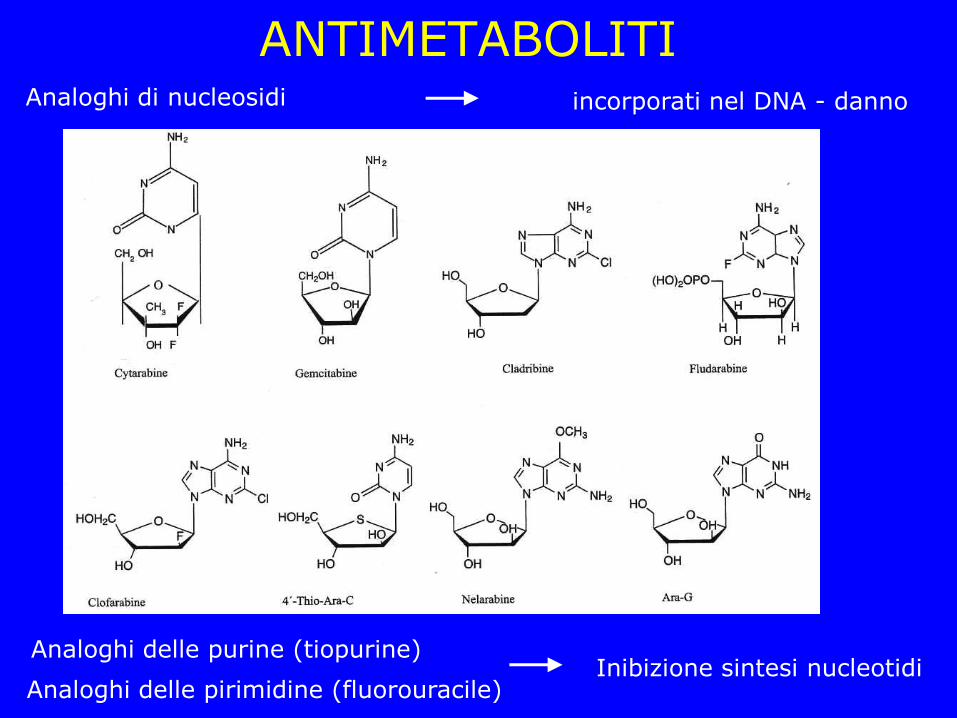
ANTIMETABOLITI
Analoghi delle purine (tiopurine)
Analoghi delle pirimidine (fluorouracile)
Analoghi di nucleosidi incorporati nel DNA - danno
Inibizione sintesi nucleotidi
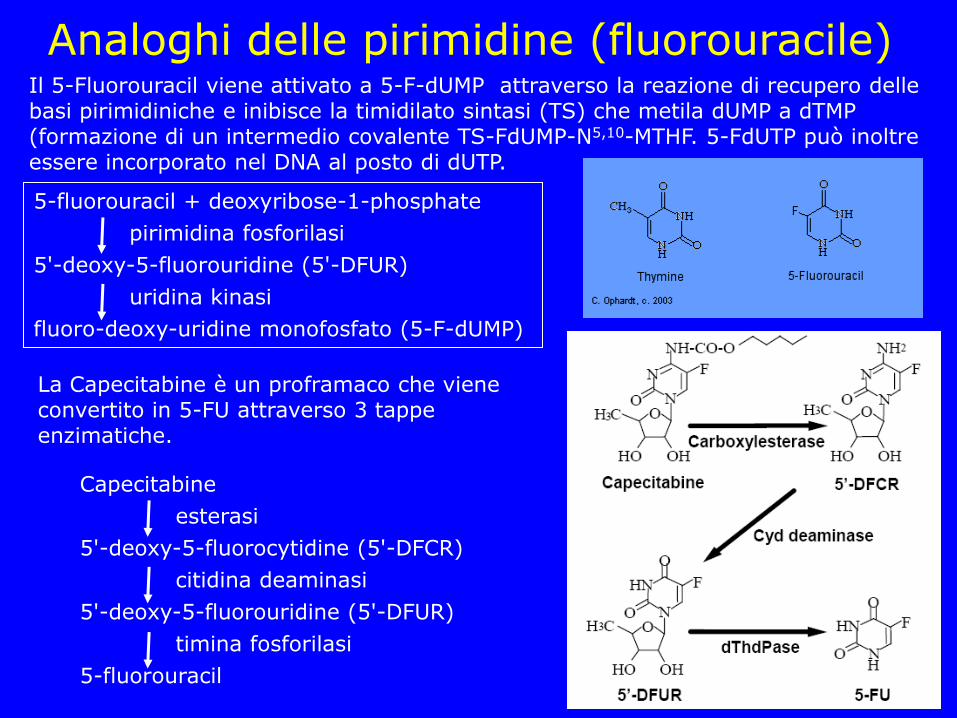
Analoghi delle pirimidine (fluorouracile) Il 5-Fluorouracil viene attivato a 5-F-dUMP attraverso la reazione di recupero delle basi pirimidiniche e inibisce la timidilato sintasi (TS) che metila dUMP a dTMP (formazione di un intermedio covalente TS-FdUMP-N5,10-MTHF. 5-FdUTP può inoltre essere incorporato nel DNA al posto di dUTP.
Capecitabine
esterasi
5'-deoxy-5-fluorocytidine (5'-DFCR)
citidina deaminasi
5'-deoxy-5-fluorouridine (5'-DFUR)
timina fosforilasi
5-fluorouracil
5-fluorouracil + deoxyribose-1-phosphate
pirimidina fosforilasi
5'-deoxy-5-fluorouridine (5'-DFUR)
uridina kinasi
fluoro-deoxy-uridine monofosfato (5-F-dUMP)
La Capecitabine è un proframaco che viene convertito in 5-FU attraverso 3 tappe enzimatiche.
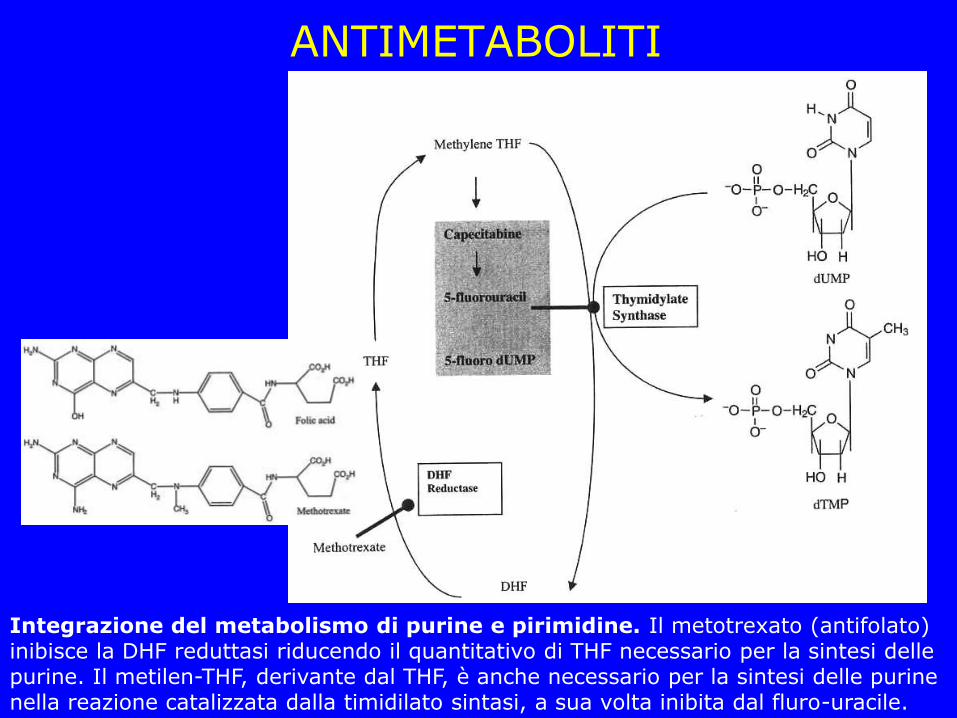
ANTIMETABOLITI
Integrazione del metabolismo di purine e pirimidine. Il metotrexato (antifolato) inibisce la DHF reduttasi riducendo il quantitativo di THF necessario per la sintesi delle purine. Il metilen-THF, derivante dal THF, è anche necessario per la sintesi delle purine nella reazione catalizzata dalla timidilato sintasi, a sua volta inibita dal fluro-uracile.

Mercaptopurine (6-MP) e thioguanine (6-TG) inibiscono la sinteisi dei nucleotidi purinici
Analoghi delle purine (tiopurine)
6-MP viene convertito in ribonucleotide e inibisce la reazione che converte inosina in adenina

ANTIMETABOLITI
Agisce anche come free radical scavenger
Utile in chemioterapia
idrossiurea Idrossiurea
L’idrossiurea è un inibitore della ribonucleotide reduttasi (RR)
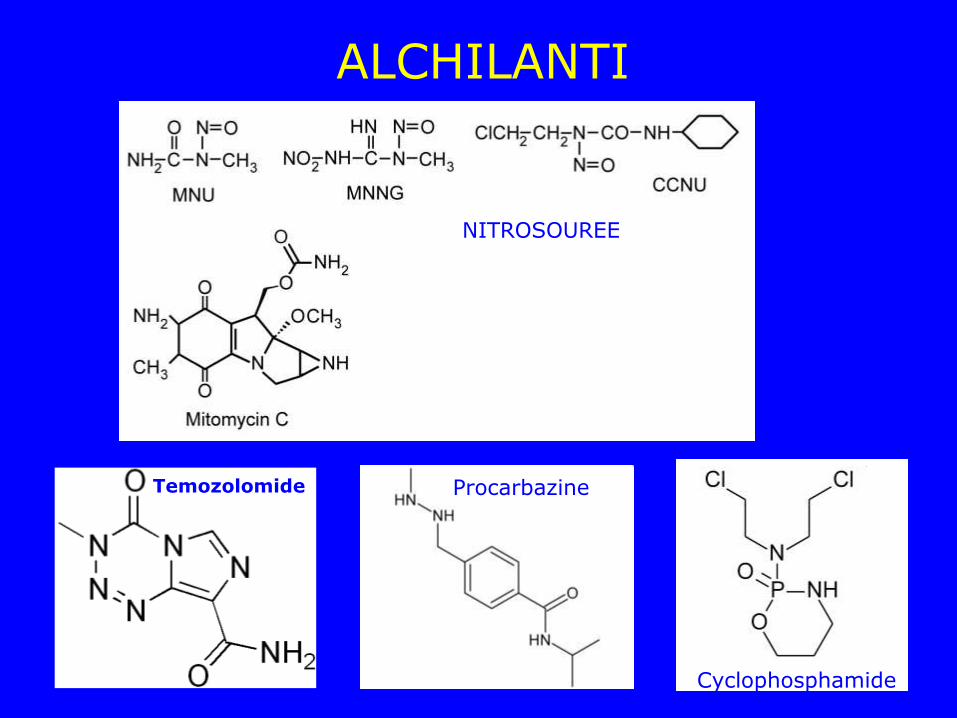
ALCHILANTI
Temozolomide Procarbazine
NITROSOUREE
Cyclophosphamide
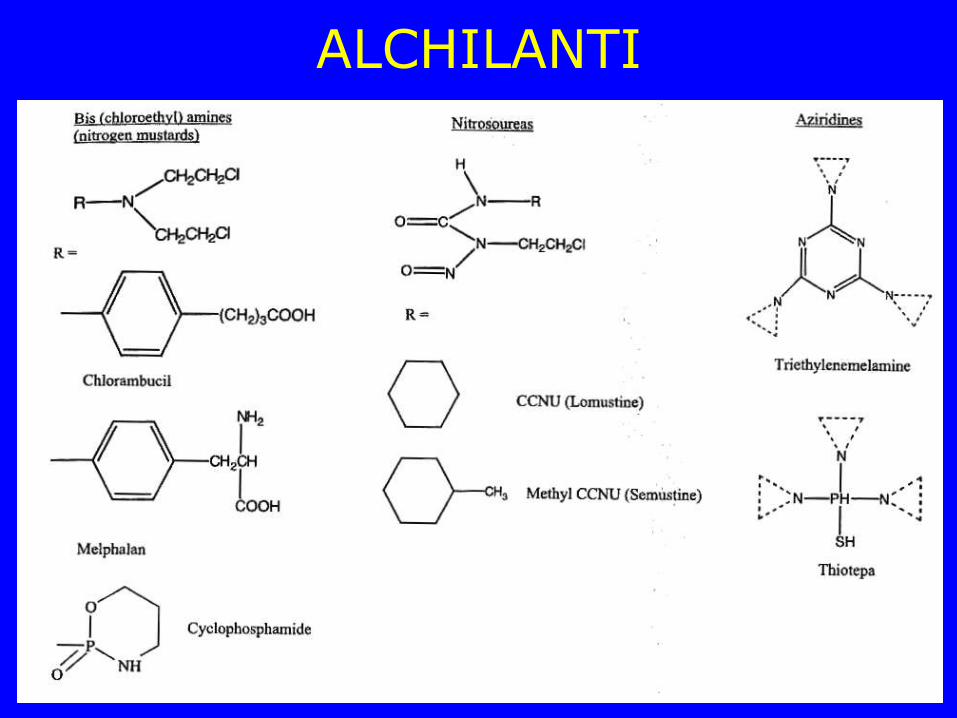
ALCHILANTI
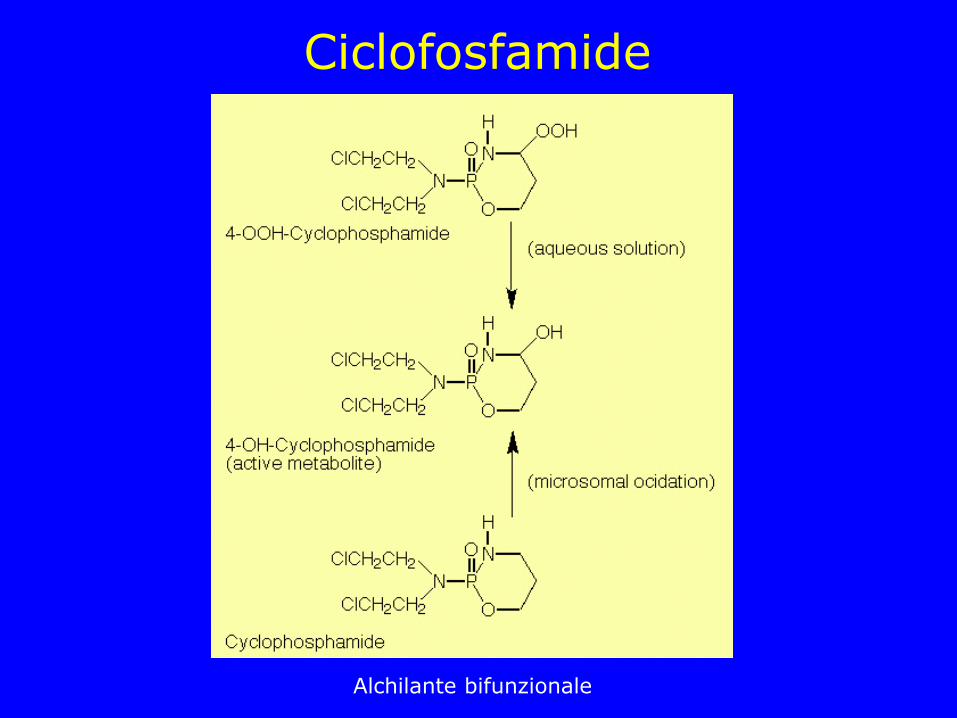
Ciclofosfamide
Alchilante bifunzionale
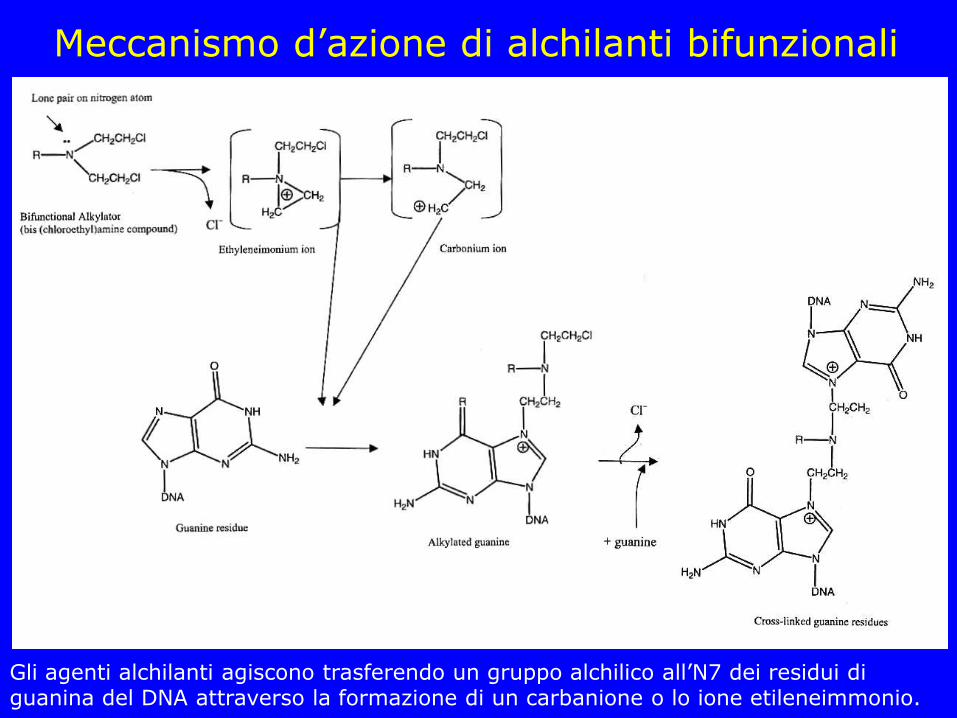
Meccanismo d’azione di alchilanti bifunzionali
Gli agenti alchilanti agiscono trasferendo un gruppo alchilico all’N7 dei residui di guanina del DNA attraverso la formazione di un carbanione o lo ione etileneimmonio.
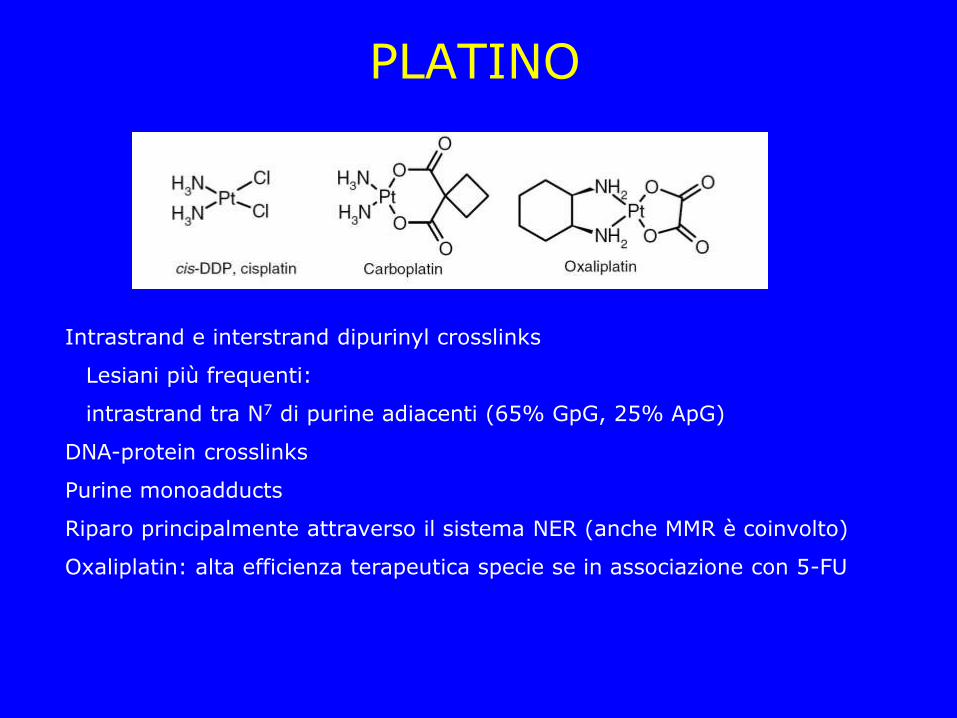
PLATINO
Intrastrand e interstrand dipurinyl crosslinks
Lesiani più frequenti:
intrastrand tra N7 di purine adiacenti (65% GpG, 25% ApG)
DNA-protein crosslinks
Purine monoadducts
Riparo principalmente attraverso il sistema NER (anche MMR è coinvolto)
Oxaliplatin: alta efficienza terapeutica specie se in associazione con 5-FU
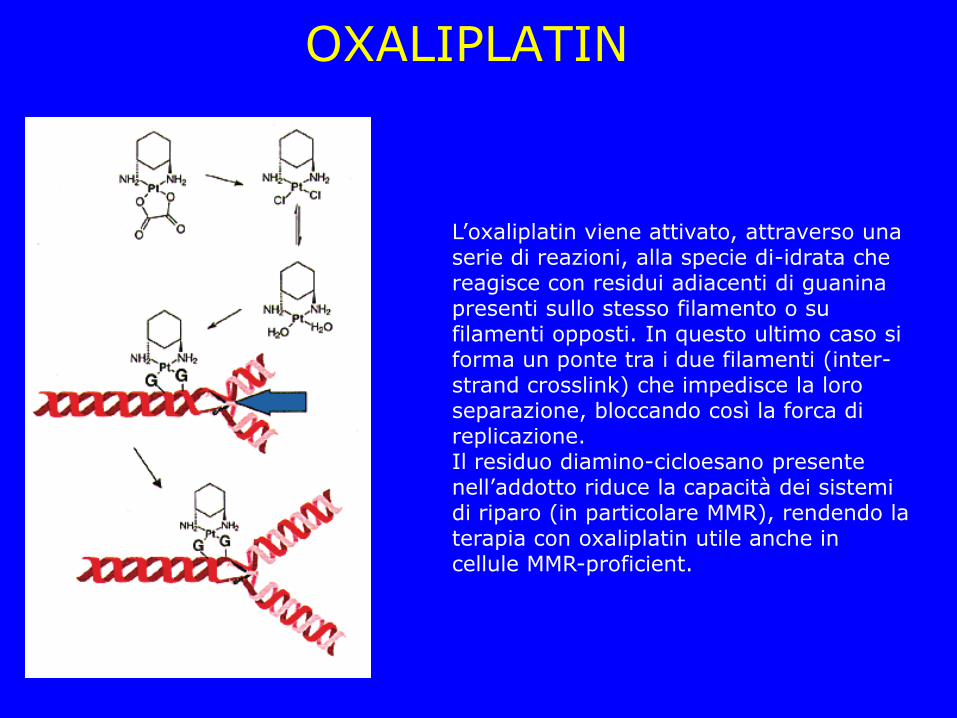
OXALIPLATIN
L’oxaliplatin viene attivato, attraverso una serie di reazioni, alla specie di-idrata che reagisce con residui adiacenti di guanina presenti sullo stesso filamento o su filamenti opposti. In questo ultimo caso si forma un ponte tra i due filamenti (inter-strand crosslink) che impedisce la loro separazione, bloccando così la forca di replicazione. Il residuo diamino-cicloesano presente nell’addotto riduce la capacità dei sistemi di riparo (in particolare MMR), rendendo la terapia con oxaliplatin utile anche in cellule MMR-proficient.
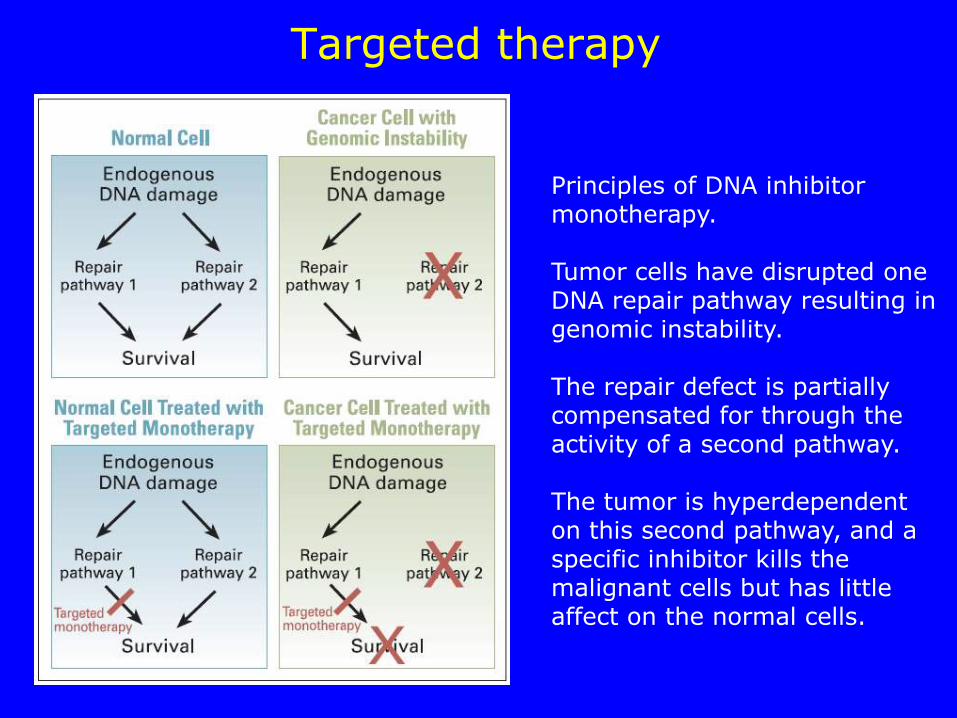
Targeted therapy
Principles of DNA inhibitor monotherapy. Tumor cells have disrupted one DNA repair pathway resulting in genomic instability. The repair defect is partially compensated for through the activity of a second pathway. The tumor is hyperdependent on this second pathway, and a specific inhibitor kills the malignant cells but has little affect on the normal cells.