Epitelio respiratorio - libreria universo · lio respiratorio del setto nasale anteriore di...
12
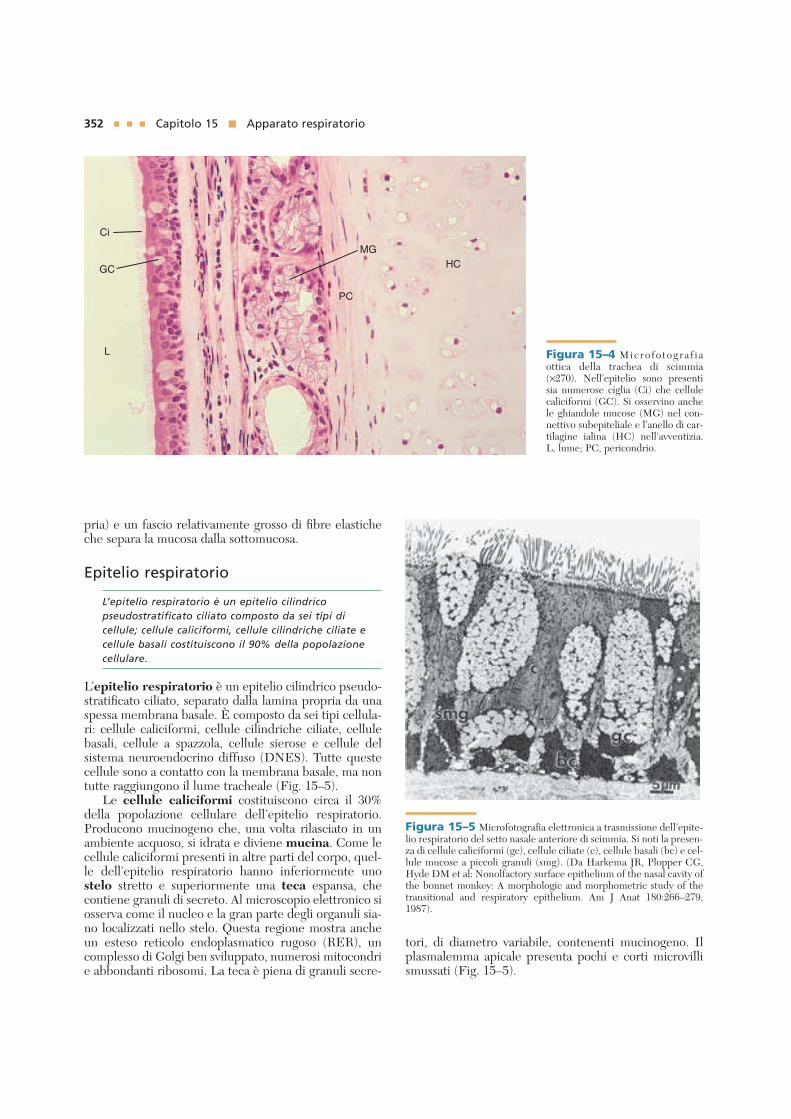
352 Capitolo 15 ■ Apparato respiratorio pria) e un fascio relativamente grosso di fibre elastiche che separa la mucosa dalla sottomucosa. Epitelio respiratorio L’epitelio respiratorio è un epitelio cilindrico pseudostratificato ciliato composto da sei tipi di cellule; cellule caliciformi, cellule cilindriche ciliate e cellule basali costituiscono il 90% della popolazione cellulare. L’epitelio respiratorio è un epitelio cilindrico pseudo- stratificato ciliato, separato dalla lamina propria da una spessa membrana basale. È composto da sei tipi cellula- ri: cellule caliciformi, cellule cilindriche ciliate, cellule basali, cellule a spazzola, cellule sierose e cellule del sistema neuroendocrino diffuso (DNES). Tutte queste cellule sono a contatto con la membrana basale, ma non tutte raggiungono il lume tracheale (Fig. 15–5). Le cellule caliciformi costituiscono circa il 30% della popolazione cellulare dell’epitelio respiratorio. Producono mucinogeno che, una volta rilasciato in un ambiente acquoso, si idrata e diviene mucina. Come le cellule caliciformi presenti in altre parti del corpo, quel- le dell’epitelio respiratorio hanno inferiormente uno stelo stretto e superiormente una teca espansa, che contiene granuli di secreto. Al microscopio elettronico si osserva come il nucleo e la gran parte degli organuli sia- no localizzati nello stelo. Questa regione mostra anche un esteso reticolo endoplasmatico rugoso (RER), un complesso di Golgi ben sviluppato, numerosi mitocondri e abbondanti ribosomi. La teca è piena di granuli secre- tori, di diametro variabile, contenenti mucinogeno. Il plasmalemma apicale presenta pochi e corti microvilli smussati (Fig. 15–5). HC PC MG Ci GC L Figura 15–4 Microfotografia ottica della trachea di scimmia (×270). Nell’epitelio sono presenti sia numerose ciglia (Ci) che cellule caliciformi (GC). Si osservino anche le ghiandole mucose (MG) nel con- nettivo subepiteliale e l’anello di car- tilagine ialina (HC) nell’avventizia. L, lume; PC, pericondrio. Figura 15–5 Microfotografia elettronica a trasmissione dell’epite- lio respiratorio del setto nasale anteriore di scimmia. Si noti la presen- za di cellule caliciformi (gc), cellule ciliate (c), cellule basali (bc) e cel- lule mucose a piccoli granuli (smg). (Da Harkema JR, Plopper CG, Hyde DM et al: Nonolfactory surface epithelium of the nasal cavity of the bonnet monkey: A morphologic and morphometric study of the transitional and respiratory epithelium. Am J Anat 180:266–279, 1987).
Transcript of Epitelio respiratorio - libreria universo · lio respiratorio del setto nasale anteriore di...
352 n n n Capitolo 15 n Apparato respiratorio
pria) e un fascio relativamente grosso di fibre elasticheche separa la mucosa dalla sottomucosa.
Epitelio respiratorio
L’epitelio respiratorio è un epitelio cilindrico
pseudostratificato ciliato composto da sei tipi di
cellule; cellule caliciformi, cellule cilindriche ciliate e
cellule basali costituiscono il 90% della popolazione
cellulare.
L’epitelio respiratorio è un epitelio cilindrico pseudo-stratificato ciliato, separato dalla lamina propria da unaspessa membrana basale. È composto da sei tipi cellula-ri: cellule caliciformi, cellule cilindriche ciliate, cellulebasali, cellule a spazzola, cellule sierose e cellule delsistema neuroendocrino diffuso (DNES). Tutte questecellule sono a contatto con la membrana basale, ma nontutte raggiungono il lume tracheale (Fig. 15–5).
Le cellule caliciformi costituiscono circa il 30%della popolazione cellulare dell’epitelio respiratorio.Producono mucinogeno che, una volta rilasciato in unambiente acquoso, si idrata e diviene mucina. Come lecellule caliciformi presenti in altre parti del corpo, quel-le dell’epitelio respiratorio hanno inferiormente unostelo stretto e superiormente una teca espansa, checontiene granuli di secreto. Al microscopio elettronico siosserva come il nucleo e la gran parte degli organuli sia-no localizzati nello stelo. Questa regione mostra ancheun esteso reticolo endoplasmatico rugoso (RER), uncomplesso di Golgi ben sviluppato, numerosi mitocondrie abbondanti ribosomi. La teca è piena di granuli secre-
tori, di diametro variabile, contenenti mucinogeno. Ilplasmalemma apicale presenta pochi e corti microvillismussati (Fig. 15–5).
HC
PC
MG
Ci
GC
L Figura 15–4 Micro fo togra f i aottica della trachea di scimmia(×270). Nell’epitelio sono presentisia numerose ciglia (Ci) che cellulecaliciformi (GC). Si osservino anchele ghiandole mucose (MG) nel con-nettivo subepiteliale e l’anello di car-tilagine ialina (HC) nell’avventizia.L, lume; PC, pericondrio.
Figura 15–5 Microfotografia elettronica a trasmissione dell’epite-lio respiratorio del setto nasale anteriore di scimmia. Si noti la presen-za di cellule caliciformi (gc), cellule ciliate (c), cellule basali (bc) e cel-lule mucose a piccoli granuli (smg). (Da Harkema JR, Plopper CG,Hyde DM et al: Nonolfactory surface epithelium of the nasal cavity ofthe bonnet monkey: A morphologic and morphometric study of thetransitional and respiratory epithelium. Am J Anat 180:266–279,1987).

Capitolo 15 n Apparato respiratorio n n n 353
Le cellule cilindriche ciliate rappresentano il 30%circa delle cellule; sono alte e sottili, con un nucleo basa-le, e presentano ciglia e microvilli sulla porzione apicaledella membrana plasmatica (Fig. 15–6). Il citoplasmaapicale è ricco di mitocondri e contiene il complesso delGolgi. Il resto del citoplasma presenta poco RER epochi ribosomi. Queste cellule spostano il muco e ilmateriale corpuscolato inglobato in esso, mediante ilmovimento delle ciglia, verso il nasofaringe per la loroeliminazione.
Le cellule basali costituiscono il 30% della popola-zione cellulare; poggiano sulla membrana basale e laloro superficie apicale non raggiunge il lume dell’organo(vedi Fig. 15–5). Sono poco differenziate e considerateperciò cellule staminali, capaci di proliferare per sosti-tuire le cellule cilindriche ciliate, a spazzola e calicifor-mi morte.
Le cellule a spazzola (piccole cellule mucose) rap-presentano il 3% della popolazione cellulare totale; sono
cellule strette, di tipo cilindrico, con alti microvilli. Laloro funzione è sconosciuta ma, dato che sono in relazio-ne con le terminazioni nervose, alcuni autori hanno sug-gerito un loro ruolo nell’attività sensoriale; altri ricerca-tori ipotizzano che siano semplicemente cellule cali-ciformi che hanno già rilasciato il loro mucinogeno.
Le cellule sierose sono cellule cilindriche che rap-presentano il 3% circa delle cellule totali dell’epiteliorespiratorio. All’apice presentano microvilli e granulicontenenti un prodotto di secrezione sieroso elettron-denso.
Le cellule DNES, conosciute anche come cellule apiccoli granuli o cellule di Kulchitsky, rappresentano il3–4% della popolazione cellulare. Molte di queste cellu-le possiedono dei lunghi e sottili prolungamenti che siestendono fino al lume, e si crede che possiedano lacapacità di monitorare i livelli di ossigeno e anidride car-bonica nel lume delle vie aeree. Queste cellule sonostrettamente associate con terminazioni nervose nude,con le quali stabiliscono contatti sinaptici, e insieme aqueste fibre nervose costituiscono i corpi neuroepite-liali polmonari. Le cellule DNES contengono nume-rosi granuli nel loro citoplasma basale, in cui è possibiletrovare sostanze farmacologicamente attive, comeammine, peptidi, acetilcolina e adenosina trifosfato. Incondizioni di ipossia queste sostanze sono rilasciate nonsolo nella fessura sinaptica, ma anche negli spazi del tes-suto connettivo della lamina propria, ove agiscono comeormoni paracrini oppure passano in circolo agendocome veri e propri ormoni. Perciò, è stato suggerito chequesti corpi neuroepiteliali possano avere effetti localiper limitare condizioni di ipossia localizzata regolando laperfusione e la ventilazione nelle zone limitrofe, oppureche possano avere effetti sistemici attraverso l’innerva-zione efferente che ritrasmette l’informazione di condi-zioni ipossiche ai centri regolatori della respirazionesituati a livello del midollo allungato.
Lamina propria e fibre elastiche
La lamina propria della trachea è composta da tessutoconnettivo lasso di tipo fibroelastico. Contiene elementilinfoidi (noduli linfatici, linfociti, neutrofili) e ghiandolesieromucose e mucose, i cui dotti si aprono sulla super-ficie epiteliale. Un denso strato di fibre elastiche, lalamina elastica, separa la lamina propria dalla sottostan-te sottomucosa.
Sottomucosa
La sottomucosa tracheale è costituita da tessuto connet-tivo denso, irregolare, di tipo fibroelastico, contenentenumerose ghiandole mucose e sieromucose. I corti dot-ti di queste ghiandole perforano la lamina elastica e lalamina propria per aprirsi sulla superficie epiteliale. Nel-la sottomucosa sono presenti elementi linfoidi; questaregione è inoltre ricca di vasi sanguigni e linfatici, i cuirami più piccoli raggiungono la lamina propria.
Figura 15–6 Microfotografia elettronica a scansione di tracheafetale umana in cui sono visibili cellule ciliate e non ciliate (×5500).(Da Montgomery PQ, Stafford ND, Stolinski C: Ultrastructure of thehuman fetal trachea. A morphologic study of the luminal and glandu-lar epithelia at the mid–trimester. J Anat 173:43–59, 1990).

354 n n n Capitolo 15 n Apparato respiratorio
Avventizia
L’avventizia della trachea è composta da tessuto connet-tivo fibroelastico (vedi Fig. 15–4). Le caratteristicheprincipali di quest’area sono gli anelli a C di cartilagineialina e il tessuto connettivo fibroso. L’avventizia è ancheresponsabile dell’ancoraggio della trachea alle struttureadiacenti (esofago e tessuto connettivo del collo).
no nel polmone a livello dell’ilo. Il bronco di destra è piùdritto di quello di sinistra e si divide in tre bronchi, chepenetrano nei tre lobi del polmone destro; il bronco disinistra si biforca in due rami, che entrano nei due lobidel polmone sinistro. Queste ramificazioni lobari rap-presentano i bronchi intrapolmonari.
Bronchi secondari e terziari(intrapolmonari)
Ogni bronco intrapolmonare serve un lobo dei polmoni;
i bronchi terziari servono i segmenti broncopolmonari.
Ogni bronco intrapolmonare rappresenta la via aerea diun lobo del polmone. Tali vie sono simili ai bronchi pri-mari, con alcune eccezioni: gli anelli a C cartilagineisono sostituiti da placche irregolari di cartilagine ialinache circondano completamente il lume del bronco intra-polmonare; in questo modo queste vie aeree non pre-sentano una parte schiacciata, ma sono del tutto roton-deggianti. Lo strato muscolare liscio è localizzato all’in-terfaccia tra la lamina propria fibroelastica e la sottomu-cosa, ed è costituito da due strati ad andamento spira-liforme orientati in direzioni opposte. Le fibre elastiche,che si irradiano dall’avventizia, si connettono con le fibreelastiche provenienti dalla avventizia di una regionediversa dell’albero bronchiale.
Come nei bronchi primari e nella trachea, le ghian-dole sieromucose e gli elementi linfoidi sono presentinella lamina propria e nella sottomucosa dei bronchiintrapolmonari. I dotti di queste ghiandole trasportano iprodotti di secrezione alla superficie dell’epitelio ciliatopseudostratificato che delimita il lume. I noduli linfaticisono particolarmente evidenti nei punti in cui le vieaeree si ramificano per formare bronchi di diametrosempre più piccolo. I bronchi intrapolmonari più picco-li hanno pareti sempre più sottili, con una quantità infe-riore di placche di tessuto cartilagineo, e presentano cel-lule epiteliali più basse.
I bronchi secondari, derivanti da quelli primari,entrano nei lobi del polmone e sono conosciuti anchecome bronchi lobari. Il polmone di sinistra è suddivisoin due lobi e quindi possiede due bronchi secondari,mentre quello di destra ha tre lobi e contiene perciò trebronchi secondari.
Appena penetrati nei lobi polmonari, i bronchisecondari si suddividono in rami più piccoli, i bronchiterziari (o segmentali). Ciascun bronco terziario si rami-fica, ma si dirige verso una sezione distinta del tessutopolmonare conosciuta come segmento broncopolmo-nare. Ogni polmone ha 10 segmenti broncopolmonari,che sono completamente separati gli uni dagli altri datessuto connettivo e sono importanti da un punto di vistaclinico negli interventi chirurgici che interessano i pol-moni.
Poiché le ramificazioni dei bronchi intrapolmonaridiminuiscono di calibro, alla fine portano ai bronchioli.
CORRELAZIONI CLINICHE
L’epitelio respiratorio delle persone esposte cro-nicamente ad agenti irritanti, come il fumo disigaretta e la polvere di carbone, subisce altera-zioni reversibili conosciute come metaplasia,associate ad un aumento del numero delle cellu-le caliciformi rispetto a quello delle cellule cilia-te. Tale aumento provoca uno strato di muco piùspesso per la rimozione degli agenti irritanti, mail ridotto numero di ciglia ritarda l’eliminazionedel muco provocando congestione. Inoltre, leghiandole sieromucose della lamina propria edella sottomucosa aumentano di dimensioni e,di conseguenza, è maggiore la secrezione. Dopopochi mesi dalla cessata esposizione agli agentiinquinanti, la percentuale cellulare ritorna allanormalità (1 : 1) e le ghiandole sieromucoseritornano alle loro normali dimensioni.
Albero bronchiale
L’albero bronchiale inizia alla biforcazione della trachea,con i bronchi primari destro e sinistro, che si arborizza-no (formano rami che diminuiscono progressivamentedi dimensioni). L’albero bronchiale è costituito dalle vieaeree esterne ai polmoni (bronchi primari, bronchiextrapolmonari) e dalle vie aeree interne ai polmoni:bronchi intrapolmonari (bronchi secondari e terziari),bronchioli, bronchioli terminali e bronchioli respiratori(Fig. 15–7). L’albero bronchiale si divide 15-20 volte pri-ma di raggiungere il livello dei bronchioli terminali.Mano a mano che le vie aeree diminuiscono di calibro,si possono osservare una diminuzione della quantità dicartilagine, del numero di ghiandole e cellule caliciformie dell’altezza delle cellule epiteliali, ed un aumento deltessuto muscolare liscio e del tessuto elastico (rispettoallo spessore della parete).
Bronchi primari (extrapolmonari)
La struttura dei bronchi primari è identica a quella del-la trachea, ma questi hanno un diametro più piccolo epareti più sottili. Ogni bronco primario si accompagnaalle arterie polmonari, vene e vasi linfatici che penetra-
Capitolo 15 n Apparato respiratorio n n n 355
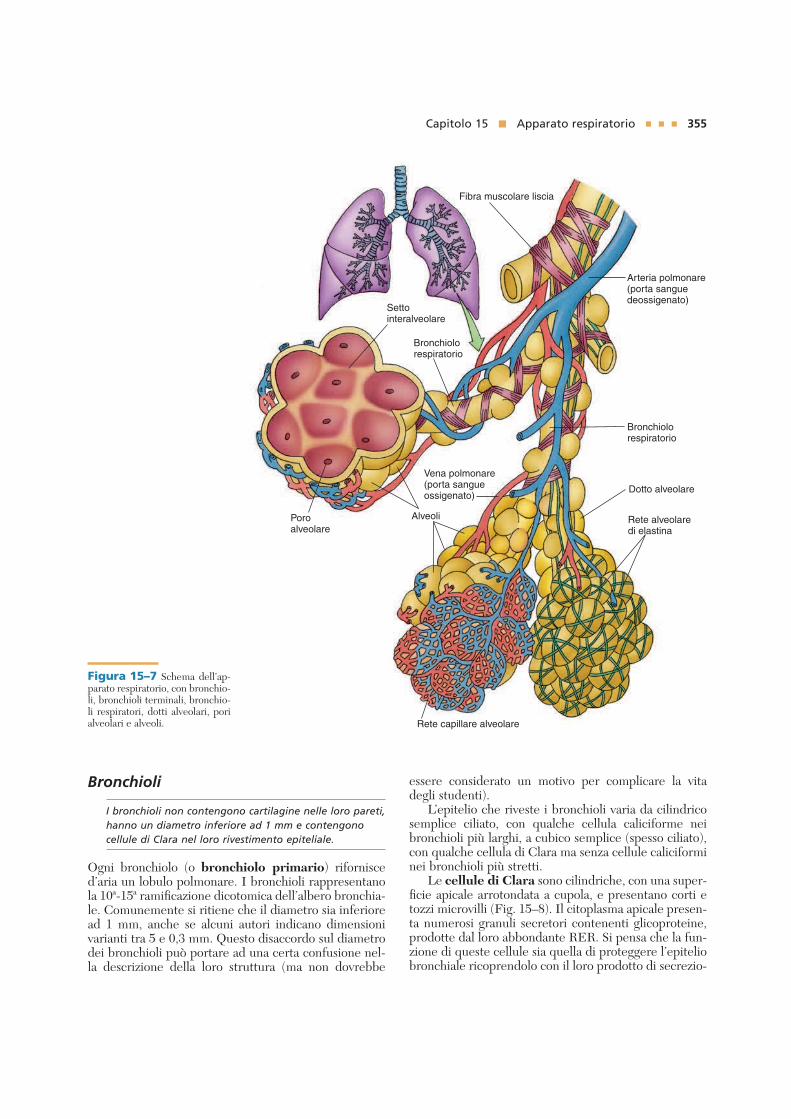
Bronchioli
I bronchioli non contengono cartilagine nelle loro pareti,
hanno un diametro inferiore ad 1 mm e contengono
cellule di Clara nel loro rivestimento epiteliale.
Ogni bronchiolo (o bronchiolo primario) rifornisced’aria un lobulo polmonare. I bronchioli rappresentanola 10ª-15ª ramificazione dicotomica dell’albero bronchia-le. Comunemente si ritiene che il diametro sia inferioread 1 mm, anche se alcuni autori indicano dimensionivarianti tra 5 e 0,3 mm. Questo disaccordo sul diametrodei bronchioli può portare ad una certa confusione nel-la descrizione della loro struttura (ma non dovrebbe
essere considerato un motivo per complicare la vitadegli studenti).
L’epitelio che riveste i bronchioli varia da cilindricosemplice ciliato, con qualche cellula caliciforme neibronchioli più larghi, a cubico semplice (spesso ciliato),con qualche cellula di Clara ma senza cellule caliciforminei bronchioli più stretti.
Le cellule di Clara sono cilindriche, con una super-ficie apicale arrotondata a cupola, e presentano corti etozzi microvilli (Fig. 15–8). Il citoplasma apicale presen-ta numerosi granuli secretori contenenti glicoproteine,prodotte dal loro abbondante RER. Si pensa che la fun-zione di queste cellule sia quella di proteggere l’epiteliobronchiale ricoprendolo con il loro prodotto di secrezio-
Fibra muscolare liscia
Settointeralveolare
Bronchiolo respiratorio
Poro alveolare
Rete capillare alveolare
Vena polmonare(porta sangueossigenato)
Dotto alveolare
Bronchiolo respiratorio
Alveoli
Arteria polmonare(porta sangue deossigenato)
Rete alveolaredi elastina
Figura 15–7 Schema dell’ap-parato respiratorio, con bronchio-li, bronchioli terminali, bronchio-li respiratori, dotti alveolari, porialveolari e alveoli.

356 n n n Capitolo 15 n Apparato respiratorio
ne. Queste cellule provvedono inoltre a degradare letossine contenute nell’aria inalata per mezzo degli enzi-mi del citocromo P–450 presenti nel reticolo endopla-smatico liscio. Alcuni autori hanno suggerito che le cel-lule di Clara producano un materiale, simile al surfattan-te, che riduce la tensione superficiale dei bronchioli efacilita quindi il mantenimento della loro pervietà. Pareinoltre che queste cellule, proliferando, rigenerino l’epi-telio dei bronchioli.
La lamina propria dei bronchioli è sprovvista dighiandole ed è circondata da una lassa rete di fascimuscolari lisci ad andamento elicoidale (Fig. 15–9). Lepareti dei bronchioli e delle loro diramazioni sono privedi cartilagine. Fibre elastiche si irradiano dal tessutoconnettivo fibroelastico che circonda il rivestimentomuscolare dei bronchioli. Durante l’inspirazione, quan-do i polmoni si espandono, tali fibre esercitano una ten-sione sulle pareti bronchiolari; tirando in manierauniforme in tutte le direzioni, le fibre elastiche contri-buiscono a mantenere pervii i bronchioli.
Bronchioli terminali
I bronchioli terminali formano le parti più piccole e più
lontane della porzione conduttrice dell’apparato
respiratorio.
Ogni bronchiolo si suddivide a formare parecchi bron-chioli terminali più piccoli, di diametro inferiore a 0,5mm, che costituiscono la parte terminale del sistema di
Figura 15–8 Microfotogra f iaelettronica a scansione delle celluledi Clara e delle cubiche ciliate deibronchioli terminali di ratto (×1817).(Da Peao MND, Aguas AP, De SaCM, Grande NR: Anatomy of Claracell secretion: Surface changesobserved by scanning electron micro-scopy. J Anat 183:377–388, 1993).
Figura 15–9 Microfotografia ottica di un bronchiolo (×117). Sinoti la presenza di muscolatura liscia (SM) e l’assenza di cartilaginenella parete. Si osservi che l’intera struttura è intrapolmonare ed è cir-condata da tessuto polmonare. A, alveolo; E, epitelio; L, lume.
CORRELAZIONI CLINICHE
La muscolatura liscia dei bronchioli è controllatadal sistema nervoso parasimpatico. In condizioninormali la muscolatura liscia si contrae al terminedell’espirazione e si rilascia durante l’inspirazio-ne. Nelle persone sofferenti di asma, comunque,gli strati muscolari vanno incontro a contrazioniprolungate durante l’espirazione, con conseguen-te difficoltà nell’espulsione dell’aria dai polmoni.Steroidi e β2–agonisti rilasciano la muscolaturaliscia bronchiale e sono di solito utilizzati per alle-viare gli attacchi asmatici.
A
SM
L
E

Capitolo 15 n Apparato respiratorio n n n 357
conduzione dell’apparato respiratorio. Queste struttureriforniscono d’aria gli acini polmonari, le ultime porzio-ni del lobulo polmonare. L’epitelio dei bronchioli termi-nali è costituito da cellule di Clara e cellule cubiche,alcune delle quali provviste di ciglia. La sottile laminapropria è costituita da tessuto connettivo fibroelastico edè circondata da uno o due strati di muscolatura liscia. Lefibre elastiche partono dall’avventizia e, come nei bron-chioli, si legano alle fibre elastiche che si irradiano daglialtri componenti dell’albero bronchiale. I bronchioli ter-minali si ramificano per formare i bronchioli respiratori.
LA PORZIONE RESPIRATORIADELL’APPARATO RESPIRATORIO
La porzione respiratoria dell’apparato respiratorio ècostituita dai bronchioli respiratori, dai dotti alveolari,dai sacchi alveolari e dagli alveoli.
Bronchioli respiratori
I bronchioli respiratori sono la prima parte dell’apparato
respiratorio in cui avvengono gli scambi gassosi.
I bronchioli respiratori hanno una struttura simile aibronchioli terminali, ma la loro parete è interrotta dastrutture a parete sottile a forma di sacco, conosciutecome alveoli, all’interno delle quali avvengono gli scam-bi respiratori (O2 per CO2). Mano a mano che i bron-chioli respiratori si ramificano, il loro diametro si restrin-ge e il numero di alveoli aumenta. Dopo alcune biforca-zioni, ogni bronchiolo respiratorio termina in un dottoalveolare (Fig. 15–10).
Dotti alveolari, atri e sacchi
alveolari
I dotti alveolari, gli atri e gli alveoli sono riforniti da una
ricca rete di capillari.
I dotti alveolari sono privi di pareti proprie e sonosemplici arrangiamenti lineari di alveoli (Figg. 15–11 e15–12). Un dotto alveolare, che origina da un bronchio-lo respiratorio, si ramifica, e ciascun dotto alveolare chesi foma normalmente termina con un rigonfiamento afondo cieco costituito da due o più piccoli gruppi dialveoli, detti sacchi alveolari. Ogni sacco si apre così inuno spazio comune, che alcuni ricercatori indicano conil nome di atrio.
Elementi sottili di tessuto connettivo tra gli alveoli, isetti interalveolari, rinforzano i dotti alveolari e inqualche modo li stabilizzano. Inoltre, l’imbocco di ognialveolo nel dotto alveolare presenta fibre collagene ditipo III ed è controllato da una singola cellula muscolareliscia (“manopola” di muscolo liscio) che forma un deli-cato sfintere capace di regolare il diametro dell’apertura.
Sottili fibre elastiche si ramificano dalla periferia deidotti alveolari e dei sacchi e si mescolano alle fibre ela-stiche provenienti da altri elementi intrapolmonari.Questa rete di fibre elastiche non solo mantiene la per-vietà di queste delicate strutture durante l’inspirazione,ma le protegge anche dal danneggiamento che si puòavere con la distensione ed è responsabile dell’espirazio-ne spontanea.
Alveoli
Gli alveoli sono piccoli sacchi aerei composti da
pneumociti di tipo I, molto appiattiti, e da numerosi
pneumociti di tipo II.
Gli alveoli sono piccoli diverticoli di circa 200 µm didiametro derivanti dai bronchioli respiratori, dai dotti edai sacchi alveolari (Fig. 15–13; vedi anche Figg. 15–11Ae B e 15–12). Gli alveoli formano la struttura funzionaleprimaria dell’apparato respiratorio, in quanto le loro sot-tili pareti permettono lo scambio di CO2 per O2, tra l’a-
Figura 15–10 Microfotografia ottica di un bronchiolo respiratorioumano (R) da cui prende origine un dotto alveolare (A). I bronchiolirespiratori hanno pareti definite con gli alveoli inframmezzati. I dottialveolari non hanno parete propria; i dotti sono creati da alveoli adia-centi.
A
R
358 n n n Capitolo 15 n Apparato respiratorio
CO2
Alveolo
Diffusione di CO2 dal sangue agli alveoli
Alveolo del polmone
Diffusione di CO2 nel sangue e conversione in HCO3–
Tessuto corporeo
Hb
CO2+H2O
Anidrasi carbonica H2CO3
HCO3–+H+
HCO3–
Cl–
CO2
CO2
Prodottadallecelluledei tessuti
Capillare
Cl–
CO2+H2O H2CO3
HCO3–+H+
Hb
Anidrasi carbonica
CO2
Capillare
Poro alveolare Bronchiolo respiratorio
Dotto alveolare
Alveolo
Sangue ricco di ossigeno al cuore
Plasma
Interno dell’alveolo
Sangue povero di ossigeno dal cuore
Eritrocita
Cellula alveolare O2
CO2
Capillare alveolare
C D
A
B
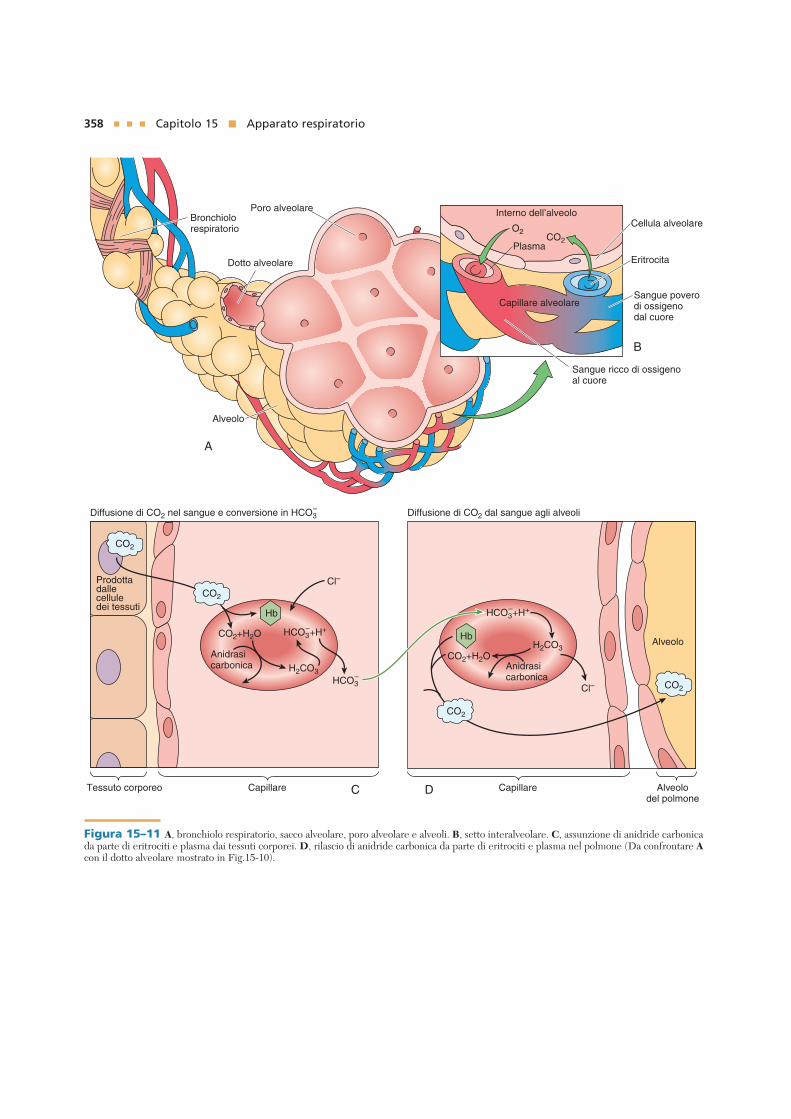
Figura 15–11 A, bronchiolo respiratorio, sacco alveolare, poro alveolare e alveoli. B, setto interalveolare. C, assunzione di anidride carbonicada parte di eritrociti e plasma dai tessuti corporei. D, rilascio di anidride carbonica da parte di eritrociti e plasma nel polmone (Da confrontare Acon il dotto alveolare mostrato in Fig.15-10).

Capitolo 15 n Apparato respiratorio n n n 359
Figura 15–12 Microfotografia elettronica a scan-sione di un polmone di ratto che mostra un bronchio-lo (b), una piccola arteria (v) e alveoli (d), alcuni deiquali presentano pori. (Da Leeson TS, Leeson CR,Paparo AA: Text/Atlas of Histology. Philadelphia, WBSaunders, 1988).
Figura 15–13 Microfotografia elettronica atrasmissione di un setto interalveolare di scimmia.Si noti la presenza di alveoli (a), eritrociti (e) den-tro i capillari (c), e macrofagi alveolari (m). Si noti-no filopodi (frecce) e pori alveolari (asterischi).(Da Maina JN: Morphology and morphometry ofthe normal lung of the adult vervet monkey (Cer-copithecus aethiops). Am J Anat 183:258–267,1988).

360 n n n Capitolo 15 n Apparato respiratorio
ria nel loro lume e il sangue nei capillari adiacenti. Seb-bene ogni alveolo sia una piccola struttura di circa 0,002mm3, il loro numero totale è di circa 300 milioni e que-sto conferisce al polmone una struttura spugnosa. Si cal-cola che l’area totale degli alveoli utile per gli scambigassosi superi i 140 m2 (circa lo spazio di un apparta-mento con due stanze da letto oppure le dimensioni diun campo da tennis).
A causa del loro elevato numero, gli alveoli sonospesso addossati gli uni agli altri, con la scomparsadell’interstizio di tessuto connettivo tra loro. In taliaree di contatto gli spazi aerei di due alveoli possonocomunicare tra loro per mezzo di un poro alveolare(poro di Kohn), il cui diametro varia dagli 8 ai 60 µm(vedi Fig. 15–12). Questi pori probabilmente hanno loscopo di equilibrare la pressione dell’aria all’internodei segmenti polmonari. La regione tra due alveoliadiacenti è conosciuta come setto interalveolare.Essa è occupata da un vasto letto capillare costituitoda capillari continui che ricevono il sangue dall’arte-ria polmonare e sono drenati dalla vena polmonare. Iltessuto connettivo del setto interalveolare è ricco difibre elastiche e di fibre collagene di tipo III (fibrereticolari).
Poiché sia gli alveoli che i capillari sono costituiti datessuto epiteliale, entrambi presentano una discretamembrana basale. Le aperture degli alveoli associati coni sacchi alveolari, a differenza di quelli dei bronchiolirespiratori e dei dotti alveolari, sono prive di cellulemuscolari lisce. Il loro orifizio è circondato da fibre ela-stiche e soprattutto da fibre reticolari. Le pareti alveola-ri sono costituite da due tipi cellulari: pneumociti di tipoI e di tipo II.
Pneumociti di tipo I
Il 95% circa della superficie alveolare è costituito da epi-telio squamoso semplice, le cui cellule sono note comepneumociti di tipo I (detti anche cellule alveolari ditipo I o cellule alveolari squamose). Le cellule diquesto epitelio sono fortemente appiattite e così il lorocitoplasma può raggiungere lo spessore di soli 80 nm
(Fig. 15–14; vedi anche Fig. 15–12). La regione delnucleo è più ampia e ospita la maggior parte degli orga-nelli cellulari, costituita da pochi mitocondri, un RER edun apparato del Golgi scarsi.
Gli pneumociti di tipo I sono uniti fra loro da giun-zioni occludenti che prevengono l’infiltrazione del fluidoextracellulare (fluido tissutale) nel lume dell’alveolo.Alla base di queste cellule vi è una membrana basaleben sviluppata, che si estende quasi al bordo dei porialveolari. Il bordo di ogni poro alveolare è formato dallafusione delle membrane cellulari di due pneumociti ditipo I a stretto contatto di due alveoli diversi. La facciaprospiciente il lume degli pneumociti di tipo I è rivesti-ta di surfattante, come descritto più avanti.
Pneumociti di tipo II
Sebbene gli pneumociti di tipo II (conosciuti anchecome grandi cellule alveolari, cellule settali e cellu-le alveolari di tipo II) siano più numerosi degli pneu-mociti di tipo I, di fatto occupano solo il 5% della super-ficie alveolare. Sono cellule cubiche sparse tra gli pneu-mociti di tipo I, con i quali formano giunzioni occluden-ti. La loro superficie apicale è a forma di cupola e spor-ge nel lume degli alveoli (Figg. 15–15 e 15–16). Si trova-no spesso in regioni dove alveoli adiacenti sono separatigli uni dagli altri da un setto (da cui il nome di cellulesettali) e sotto la superficie opposta al lume vi è unamembrana basale.
Le fotografie al microscopio elettronico degli pneu-mociti di tipo II mostrano corti microvilli apicali, unnucleo centrale, abbondante RER, un apparato del Gol-gi ben sviluppato e numerosi mitocondri. La caratteristi-ca principale di queste cellule è la presenza di corpilamellari, delimitati da membrana, che contengono ilsurfattante polmonare, un prodotto di secrezione diqueste cellule.
Questo surfattante è sintetizzato a livello del RERdegli pneumociti di tipo II ed è costituito principalmenteda due fosfolipidi, la dipalmitoil fosfatidilcolina e ilfosfatidilglicerolo, da lipidi neutri e da quattro tipicheproteine, le apoproteine surfattanti SP-A, SP-B, SP-C
aep
en
p
b
r
Figura 15–14 Microfotografia elettronica a trasmissione della barriera sangue–gas (×71.250). Si noti la presenza dell’alveolo (a), di pneumo-citi di tipo I assottigliati (ep), membrane basali fuse (b), cellule endoteliali appiattite dei capillari (en) con vescicole pinocitotiche (frecce), plasma(p) ed un eritrocita (r) nel lume capillare. (Da Maina JN: Morphology and morphometry of the normal lung of the adult vervet monkey (Cerco-pithecus aethiops). Am. J Anat 183:258–267, 1988).

Capitolo 15 n Apparato respiratorio n n n 361
e SP-D. Il surfattante è modificato a livello dell’ap-parato del Golgi ed è rilasciato dalla rete trans delGolgi in vescicole secretorie conosciute come corpicompositi, che rappresentano i precursori dei corpilamellari.
Il surfattante viene rilasciato mediante esocitosi nellume alveolare. Qui forma una rete larga simile al latti-ce, nota come mielina tubulare, che successivamentesi separa nelle sue componenti lipidica e proteica. I lipi-di sono inseriti in un film monomolecolare di fosfolipidi,costituendo l’interfaccia con l’aria, mentre le proteineentrano nello strato acquoso tra gli pneumociti e il filmdi fosfolipidi. Il surfattante riduce la tensione superficia-le, prevenendo così il collasso degli alveoli, l’atelettasia.Esso viene continuamente prodotto dagli pneumociti ditipo II ed è poi fagocitato dagli pneumociti di tipo II epiù raramente dai macrofagi alveolari.
Oltre a produrre e fagocitare surfattante, gli pneu-mociti di tipo II vanno incontro a mitosi per rigenerarsi,come gli pneumociti di tipo I.
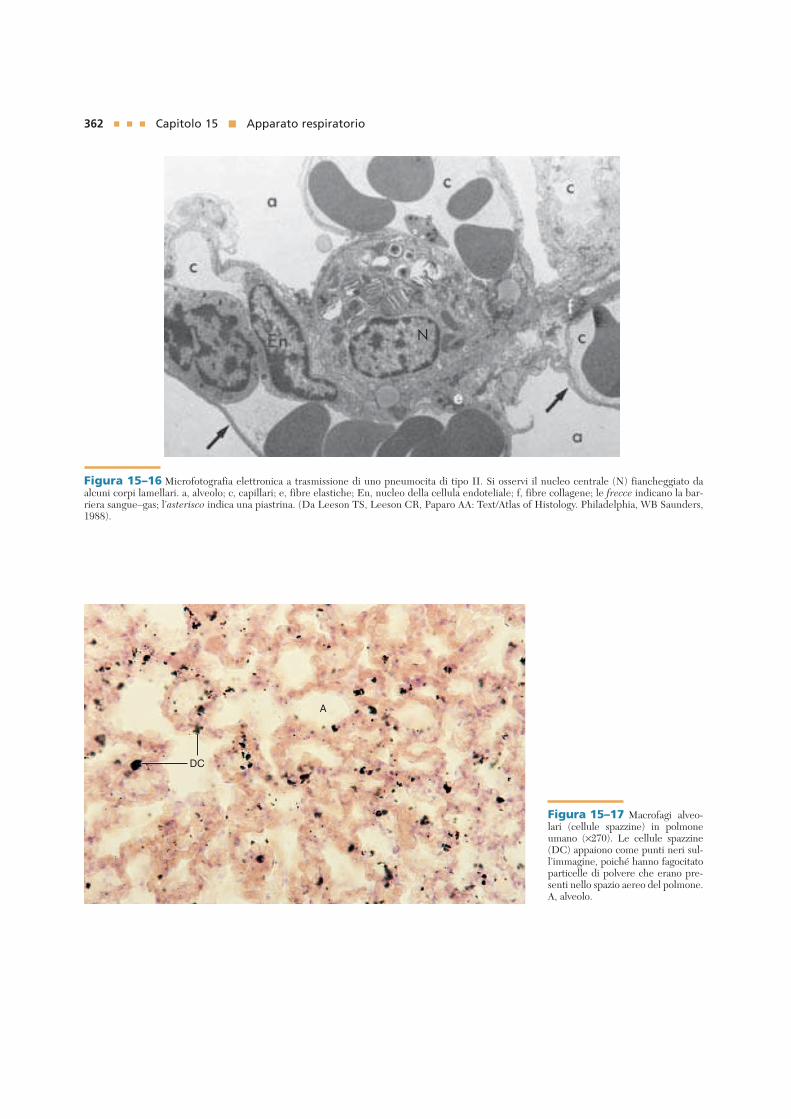
Macrofagi alveolari (cellule pazzine)
I macrofagi alveolari fagocitano materiale corpuscolato
nel lume degli alveoli e negli spazi interalveolari.
I monociti raggiungono l’interstizio polmonare, diven-gono macrofagi alveolari (o cellule spazzine), migranotra gli pneumociti di tipo I e quindi entrano nel lumealveolare. Queste cellule fagocitano il materiale corpu-scolato, come la polvere e i batteri, ed in questo modomantengono un ambiente intrapolmonare sterile (Fig.15–17; vedi anche Fig. 15–13). Le cellule spazzine, inol-tre, cooperano con gli pneumociti di tipo II nel recupe-ro del surfattante. Circa 100 milioni di macrofagi migra-no nei bronchi ogni giorno e da qui vengono trasportati,per azione delle ciglia, alla faringe per essere eliminatimediante la deglutizione o l’espettorazione. Alcunimacrofagi alveolari, comunque, rientrano nell’interstiziopolmonare e passano nei vasi linfatici per abbandonare ipolmoni.
Surfattante liberato dallavescicola lipoproteica
Strato acquoso sottostante
Monostrato lipidicoSurfattante
Piccolo corpolamellare chesi fonde con lavescicolalipoproteica
Corpomultivescicolare
Piccolo corpo lamellare (fosfolipidi)
Sintesi proteica
Golgi
Sintesi di fosfatidilcolina
Aminoacidi
Colina
Figura 15–15 Schema di pneumocita di tipo II. (Da confrontarecon lo pneumocita di tipo II mostrato in Fig. 15-16).
CORRELAZIONI CLINICHE
Alla nascita, i polmoni del neonato si espandonofin dal primo atto respiratorio e la presenza delsurfattante permette agli alveoli di rimanere per-vii. I neonati immaturi (nati prima del settimomese di gestazione), che non hanno ancora pro-dotto il surfattante (o che ne hanno prodotto unaquantità inadeguata), possono subire lo stressrespiratorio del neonato, che può essere fatale.Questi neonati sono trattati con una terapia com-binata di surfattante sintetico e glucocorticoidi. Ilprimo agisce immediatamente per ridurre la ten-sione superficiale, i secondi stimolano la sua pro-duzione da parte degli pneumociti di tipo II.
CORRELAZIONI CLINICHE
I macrofagi alveolari in soggetti con congestionepolmonare e insufficienza cardiaca congestiziafagocitano globuli rossi travasati. Questi macrofa-gi sono generalmente denominati cellule del-l’insufficienza cardiaca.
L’enfisema è una patologia associata di solitoad una sequela di esposizioni a lungo termine alfumo di sigaretta o ad altri inibitori della proteinaα1–antitripsina. Questa proteina salvaguarda ipolmoni dall’azione dell’elastasi sintetizzata dallecellule spazzine, che distrugge le fibre elastiche.In tali pazienti l’elasticità del tessuto polmonare èridotta e sono presenti grosse sacche piene difluido, che fanno diminuire la capacità di scambiogassoso della porzione respiratoria dell’apparatorespiratorio.
362 n n n Capitolo 15 n Apparato respiratorio
DC
A
Figura 15–16 Microfotografia elettronica a trasmissione di uno pneumocita di tipo II. Si osservi il nucleo centrale (N) fiancheggiato daalcuni corpi lamellari. a, alveolo; c, capillari; e, fibre elastiche; En, nucleo della cellula endoteliale; f, fibre collagene; le frecce indicano la bar-riera sangue–gas; l’asterisco indica una piastrina. (Da Leeson TS, Leeson CR, Paparo AA: Text/Atlas of Histology. Philadelphia, WB Saunders,1988).
Figura 15–17 Macrofagi alveo-lari (cellule spazzine) in polmoneumano (×270). Le cellule spazzine(DC) appaiono come punti neri sul-l’immagine, poiché hanno fagocitatoparticelle di polvere che erano pre-senti nello spazio aereo del polmone.A, alveolo.

Capitolo 15 n Apparato respiratorio n n n 363
Setto interalveolare
La regione tra due alveoli adiacenti, conosciuta comesetto interalveolare, è rivestita su entrambi i lati da epi-telio alveolare (vedi Fig. 15–13). Il setto interalveolarepuò essere estremamente stretto e ospitare solo uncapillare continuo e la sua membrana basale, oppuretalvolta può essere più ampio e includere elementi ditessuto connettivo, come fibre collagene di tipo III efibre elastiche, macrofagi, fibroblasti (e miofibroblasti),mastociti ed elementi linfoidi.
Barriera sangue–gas
La barriera sangue–gas è la regione del setto
interalveolare che è attraversata da O2 e da CO2 quando
questi gas vanno dal vaso sanguigno al lume degli
alveoli, e viceversa.
Le regioni più sottili del setto interalveolare, in cui i gaspossono essere scambiati, sono definite come barrierasangue–gas (vedi Fig. 15–14). La barriera sangue-gas piùstretta, dove gli pneumociti di tipi I sono in stretto con-tatto con l’endotelio dei capillari e dove le membranebasali dei due epiteli si fondono, è la più efficiente per loscambio di O2 (nel lume alveolare) e CO2 (nel sangue).Queste regioni sono costituite dalle seguenti strutture:
n surfattante e pneumociti di tipo In membrane basali fuse di pneumociti di tipo I e di cel-
lule endoteliali dei capillarin cellule endoteliali dei capillari continui
Scambi gassosi tra tessuti e polmoni
Nei polmoni, l’O2 viene scambiato con la CO2 trasportata
dal sangue; nei tessuti del corpo, la CO2 viene scambiata
con l’O2 trasportato dal sangue.
Durante l’inspirazione, l’ossigeno dell’aria entra neglispazi alveolari dei polmoni. Poiché l’area totale deglialveoli supera i 140 m2 ed il volume totale del sangue intutti i capillari polmonari in qualsiasi momento nonsupera i 140 ml, lo spazio disponibile per la diffusionedei gas è enorme. Inoltre, il diametro dei capillari è suf-ficientemente stretto da far passare i globuli rossi in sin-gola fila; in questa maniera l’ossigeno può raggiungereciascun eritrocita da ogni parte, utilizzando l’interasuperficie del globulo rosso disponibile allo scambio digas. L’ossigeno diffonde attraverso la barrierasangue–gas, entra nel lume dei capillari e si lega all’emedell’emoglobina degli eritrociti formando l’ossiemoglo-bina. La CO2 lascia il sangue e diffonde, attraverso labarriera sangue–gas, nel lume degli alveoli e lascia glispazi alveolari quando l’aria ricca di CO2 è espirata. Ilpassaggio di O2 e CO2 attraverso la barriera sangue-gas èdovuto alla diffusione passiva in risposta alla pressioneparziale di questi gas nel sangue e negli alveoli.
Le cellule del corpo formano ogni minuto circa 200ml di CO2; questa entra in circolo ed è trasportata in tremodi: (1) come gas disciolto nel plasma (20 ml), (2) lega-ta all’emoglobina (40 ml) oppure (3) sotto forma di ionibicarbonato nel plasma (140 ml). La sequenza di eventiche si susseguono negli scambi gassosi è qui brevemen-te riassunta (vedi Fig. 15–11C):
1 Una gran parte della CO2 sciolta nel plasma diffondenel citosol degli eritrociti.
2 Una parte della CO2 si lega alla porzione globularedell’emoglobina. Sebbene la CO2 sia trasportata inpunti diversi della molecola di emoglobina, la suacapacità di legame è maggiore in assenza di O2 nell’e-me.
3 La gran parte della CO2 che si trova all’interno delcitosol degli eritrociti, si combina con l’acqua in unareazione catalizzata dalla anidrasi carbonica e for-ma l’acido carbonico, che si dissocia in ione idrogeno(H+) e ione bicarbonato (HCO3
–). Gli ioni idrogeno silegano all’emoglobina e quelli bicarbonato lascianogli eritrociti per entrare nel plasma. Per il manteni-mento dell’equilibrio ionico, lo ione cloruro (Cl–)passa dal plasma agli eritrociti; questo scambio dibicarbonato contro cloruro è detto “shift” del clo-ruro.
Il sangue ricco di bicarbonato raggiunge i polmoniattraverso le arterie polmonari. Poiché il livello di CO2 èmaggiore nel sangue rispetto a quello del lume alveola-re, la CO2 è liberata (secondo il gradiente di concentra-zione). Il meccanismo di rilascio è l’inverso della prece-dente reazione. Gli eventi che si verificano sono iseguenti (vedi Fig. 15–11D):
1 Gli ioni bicarbonato entrano negli eritrociti (con con-seguente rilascio di Cl– dai globuli rossi nel plasma,shift del cloruro).
2 Gli ioni bicarbonato e idrogeno nel citosol degli eri-trociti si combinano, con conseguente formazione diacido carbonico.
3 Nel polmone, il legame dell’O2 con l’emoglobina ren-de quest’ultima più acida e riduce la sua capacità dilegare CO2. Inoltre, l’eccesso di ioni idrogeno libera-ti a causa dell’alta acidità dell’emoglobina si lega agliioni bicarbonato, formando acido carbonico.
4 L’anidrasi carbonica catalizza la rottura del legamedell’acido carbonico, con formazione di acqua e CO2.
5 La CO2 sciolta nel plasma, legata all’emoglobina erilasciata dall’acido carbonico, segue il gradiente diconcentrazione, per diffondere attraverso la barrierasangue–gas ed entrare quindi nel lume alveolare.
L’emoglobina possiede anche due siti di legame perl’ossido nitrico (NO), un neurotrasmettitore che,quando viene liberato dalle cellule endoteliali dei vasisanguigni, induce un rilasciamento della muscolaturaliscia vascolare, con conseguente dilatazione dei vasi








![L'APPARATO RESPIRATORIO [modalit compatibilit ])€¦ · La RESPIRAZIONE è lo scambio di O2 e CO2 tra l’atmosfera e cellule corporee. Prevede: • RESPIRAZIONE ESTERNA avviene](https://static.fdocumenti.com/doc/165x107/5f0b3d367e708231d42f882e/lapparato-respiratorio-modalit-compatibilit-la-respirazione-lo-scambio-di.jpg)